
On the largest Ichthyosaurus: A new specimen of Ichthyosaurus somersetensis containing an embryo
DEAN R. LOMAX and SVEN SACHS
Lomax, D.R. and Sachs, S. 2017. On the largest Ichthyosaurus: A new specimen of Ichthyosaurus somersetensis containing an embryo. Acta Palaeontologica Polonica 62 (3): 575–584.
A formerly undescribed Ichthyosaurus specimen from the collection of the Niedersächsisches Landesmuseum (Lower Saxony State Museum) in Hannover, Germany, provides valuable new information. The skeleton was collected from the Lower Jurassic strata (lower Hettangian, Blue Lias Formation) of Doniford Bay, Somerset, UK. However, the specimen is a composite as almost the entire tail has been added and other parts are reconstructed. Regardless of the incomplete preservation, the estimated total length of this individual, based on the skull and precaudal length, is between 300 and 330 cm and it is thus the largest unequivocal example of the genus Ichthyosaurus. Cranial and postcranial characters, specifically from the maxilla, lacrimal, jugal, the humerus, and the ilium justify a referral to I. somersetensis. A fork-like shape of the proximal end of the ilium is unusual and has not been reported for any species of Ichthyosaurus. Likewise the presence of four elements in the third row of the hindfin, indicated by the presence of a bifurcation is novel for the species and has wider implications for the taxonomic utility of hindfins within the genus. The specimen also bears an embryo, which is only the third embryo known for Ichthyosaurus and the first to be positively identified to species level.
Key words: Ichthyosauria, Ichthyosaurus somersetensis, embryo, Jurassic, Hettangian, UK, Somerset.
Dean R. Lomax [dean.lomax@manchester.ac.uk], School of Earth and Environmental Sciences, The University of Manchester, Oxford Road, Manchester, M13 9PL, UK.
Sven Sachs [Sachs.Pal@gmail.com], Naturkundemuseum Bielefeld, Abteilung Geowissenschaften, Adenauerplatz 2, 33602 Bielefeld, Germany; Im Hof 9, 51766 Engelskirchen, Germany.
Received 26 April 2017, accepted 9 June 2017, available online 28 August 2017.
Copyright © 2017 D.R. Lomax and S. Sachs. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
During the mid-1990s, professional fossil collector Peter Langham found a large, well-preserved and mostly articulated ichthyosaur skeleton at Doniford Bay, Watchet, Somerset, UK (Fig. 1). The specimen was recovered from the Early Jurassic (lower Hettangian) Blue Lias Formation, specifically the Caloceras johnstoni Ammonite Subzone of the Psiloceras planorbis Ammonite Zone (Bed 36) and is now curated in the Niedersächsisches Landesmuseum (NLMH) in Hannover, Germany.
Numerous ichthyosaurs have been collected in Somerset. Most are historical specimens discovered in quarries around Street and surrounding areas during the early and middle 19th century, although several have been found on the Somerset coast more recently (Delair 1969; Deeming et al. 1993; McGowan 2003; Massare and Lomax 2016a). The majority of this material is referable to the genus Ichthyosaurus, which has been recorded from various locations in the UK, most notably the Lyme Regis-Charmouth location on the Dorset coast and from quarries in Street and surrounding areas in Somerset, but also outside the UK (Massare and Lomax 2017a).
There are six valid species of Ichthyosaurus: I. communis De la Beche and Conybeare, 1821; I. breviceps Owen, 1881; I. conybeari Lydekker, 1888; I. anningae Lomax and Massare, 2015; I. larkini Lomax and Massare, 2017; and I. somersetensis Lomax and Massare, 2017. The studied specimen (NLMH 106234) can be assigned to Ichthyosaurus because it possesses diagnostic traits for the genus, which are: a forefin with an anterior digital bifurcation, a humerus that is almost as wide proximally as distally, a hindfin with a bifurcation, and a tripartite pelvis (Massare and Lomax 2017a). A tripartite pelvis has been reported in the coeval Temnodontosaurus and Leptonectes specimens, but the morphology of those elements are distinctly different to those in Ichthyosaurus (McGowan and Motani 2003; Delsett et al. 2017). NLMH 106234 can further be assigned to Ichthyosaurus somersetensis based on the morphology of the skull, humerus, and ilium. I. somersetensis is known from multiple specimens, including complete to fairly complete skeletons (Lomax and Massare 2017). In fact most Ichthyosaurus specimens found in Somerset belong to this species (Lomax and Massare 2017; DRL personal observation). The studied specimen provides new information on the morphology of the pelvis and hindfin of the species, along with the size range of the genus.
An embryo is also preserved with NLMH 106234, positioned between the ribs. The embryo comprises a portion of articulated vertebral column, fragments of ribs, a section of an articulated forefin, and probably a scapula. This is only the third embryo reported for Ichthyosaurus and the first to be positively identified to species level (Pearce 1846; Deeming et al. 1993).
Fig. 1. Map of Somerset including the location of Doniford Bay (indicated by a star), Watchet, UK.
Institutional abbreviations.—ANSP, Academy of Natural Sciences, Philadelphia, USA; BRSMG, Bristol City Museum and Art Gallery, Bristol, UK; IRSNB, Royal Belgian Institute of Natural Sciences, Brussels, Belgium; BU, Lapworth Museum of Geology, University of Birmingham, Birmingham, UK; NHMUK (formerly BMNH), The Natural History Museum, London, UK; NLMH, Niedersächsisches Landesmuseum (Lower Saxony State Museum), Hannover, Germany; PETMG, Peterborough Museum and Art Gallery, Peterborough, UK; SMF, Sauriermuseum, Frick, Switzerland; UWGM, University of Wisconsin Geological Museum, Madison, USA.
Material and methods
History and reconstruction of the specimen.—The studied specimen, NLMH 106234, appears to be a complete skeleton, but is actually a composite (Fig. 2). When found, most of the caudal vertebrae could not be collected and the tail associated with NLMH 106234 comprises vertebrae from another individual as well as replicated ones (DRL and SS personal observation; personal communication Peter Langham, 2017). Composites of Ichthyosaurus have been reported, but most are from historical collections, although recently collected specimens are also known for composites or have modifications (Massare and Lomax 2014, 2016b), including specimens restored by Peter Langham (Deeming et al. 1993; Massare and Lomax 2016a).
NLMH 106234 was initially prepared by P. Langham who added a tail from a smaller ichthyosaur and various ammonites to create a better specimen for display. A large crack runs across the mid-dorsal region of the specimen where two adjacent blocks meet (Fig. 2). The vertebrae are disarticulated at this point and several centra are probably missing, as are several ribs. The neural spines appear consistent across the two blocks, although they are set in plaster. Parts of the skull, some neural spines and centra, and the forefin have also been reconstructed (Figs. 2B, 3, 4).
The forefin, however, requires some explanation. A portion of the radius, most of the radiale and distal carpal 3, all of distal carpal 2, metacarpal 2 and 3, and up to four phalanges of digits II and III, and several posterior phalanges of digit IV, are reconstructed from plaster (Fig. 4). The element that contacts the radiale anteriorly, along with the next four elements, are entirely reconstructed from plaster. Although, the sixth element is genuine and indicates the presence of an anterior bifurcation of digit II. Aside from the additions and reconstructions, the rest of the specimen, including several of the ammonites (Caloceras johnstoni) and a plant frond (probably an example of Otozamites or Ptilophyllum) are genuine (our observation; personal communication Peter Langham, 2017; Fig. 2).
The specimen was later sold to Siber + Siber, a Swiss company specializing in minerals, rocks and fossils. It was purchased from Siber + Siber by the Hannover-based art collector Ernst Schwitters who used it as a wall decoration in his living room. When Schwitters died the Kurt & Ernst Schwitters Foundation became the owner. In 2005, the specimen was placed on loan to the NLMH, but was officially purchased by the museum in 2013. Initially, a wooden frame surrounded the skeleton, that was later removed when it was conserved for display in 2006 and 2007. Since December 2007 the specimen has been on display at NLMH, where one of us (SS) first became aware of it.
Methods for size estimation.—Another issue that requires discussion is the skull, jaw, and precaudal lengths of NLMH 106234 (Table 1). The skull length is 57.5 cm and the jaw length has a maximum of 67 cm and a minimum of 64 cm, accounting for some reconstruction (3 cm) in the posterior portion of the mandible. The precaudal vertebral column length, measured along the line of the vertebral column from the first vertebra at the back of the skull to the first definite single-headed rib, is approximately 107 cm. However, where the two adjacent blocks meet at the mid-dorsal region at least three vertebrae, and possibly a fragment of a fourth, are disarticulated and positioned adjacent to the rest of the vertebral column. At this position, the bones are all set in plaster. It is possible that the two large blocks were placed too close together and that some vertebrae are missing. Furthermore, the precaudal count for the studied specimen is 40, which is considerably less than for other examples of Ichthyosaurus somersetensis. The count should be greater than 46 (Lomax and Massare 2017), which suggests at least six vertebrae are missing in the mid-dorsal region. A mid-to-posterior dorsal centrum on this specimen measures approximately 2.5 cm long in lateral view, which, accounting for the missing centra, would add roughly 15 cm to the precaudal length, giving a total of 122 cm. Buchholtz (2001: fig. 5) showed that the tail stock (pelvis to bend in the tail) for Ichthyosaurus is about 60% of the precaudal length, which would suggest a tail stock length of approximately 73 cm for NLMH 106234 (i.e., a preflexural total length of 195 cm, not including the skull). For comparison, the tail stock is 56% of the precaudal length in the holotype of I. somersetensis (ANSP 15766). NHMUK OR2013*, a referred specimen of I. somersetensis, has a total length from tip of snout to tip of fluke of 268 cm, including a fluke length of ~ 40 cm. Considering the size of the studied specimen, it is probable that the fluke length may have been as much as 50 cm. Adding this fluke length and the skull length to the estimated preflexural length of NLMH 106234, suggests an animal with a total length of just over 300 cm (Table 1).
Furthermore, using the equation between skull length and vertebral column length from Deeming et al. (1993) suggests that NLMH 106234 should have a vertebral column total length of 272 cm. Adding the skull length of 57.5 cm equates to a total body length of 329.5 cm, about 30 cm longer than the length suggested above, but that difference could probably be accounted for in the tail.
Table 1. Measurements (in cm) of Ichthyosaurus somersetensis specimens, including the holotype (ANSP 15766) and a referred specimen, compared with NLMH 106234. (L/R), left/right skeletal elements; e, estimated measurement because the bone was damaged, rotated or partially buried; 1, see Materials and methods for details. The precaudal and preflexural lengths do not include the skull length. Note: NHMUK OR2013* is behind glass and not accessible for measurement, so a cast was measured instead. Some of the measurements are estimates because the cast does not display good enough detail.
|
Length |
ANSP |
NHMUK OR2013* |
NLMH |
|
skull |
43.8 |
47.0 |
57.5 |
|
preorbital |
28.0 |
30.5 |
40.0 |
|
prenarial |
21.0 |
31.1 |
33.3 |
|
maxilla |
11.8 |
|
14.5 |
|
jaw |
47.0 |
53.0 |
64.0–67.01 |
|
scapula |
|
12 (R) |
15.5 (R) |
|
humerus |
8.8 (L), 8.2e (R) |
9–10e (R) |
10.4 (R) |
|
femur |
6.0 (L) 6.3 (R) |
7 (R) |
7.7 (L), 7.8 (R) |
|
ischium |
|
|
8.6 (L), 8.4 (R) |
|
pubis |
|
|
8.3e (L) |
|
ilium |
|
|
8.4 (L) |
|
precaudal |
102 |
118e |
122e |
|
preflexural (includes apical centra) |
157 |
181e |
195e |
|
total length (from tip of snout to tip of tail) |
230 (some posterior caudal vertebrae may be missing) |
268 |
300–330e |
Ichthyosaurus size.—In the following discussion, the total length includes the skull length and the vertebral column length from the back of the skull to the tip of the tail. As currently defined, the smallest species of Ichthyosaurus is I. conybeari. The holotype of I. conybeari is the largest reported specimen, but it only comprises an anterior skeleton, including a damaged skull. This specimen probably has a total body length estimate of < 150 cm (Massare and Lomax 2016a; McGowan and Motani 2003). However, Massare and Lomax (2016a) noted that the species may be larger, in light of a fragmentary skeleton that may or may not belong to the species. Next in size is I. anningae which, based on the holotype, has a total length estimate of < 180 cm (Lomax and Massare 2015). Of similar size is I. breviceps. The largest specimen of I. breviceps has a total length estimate of < 190 cm (Massare and Lomax 2014). In comparison, I. communis is slightly larger, with a probable total length of around 200 cm (Massare and Lomax 2017a; DRL personal observation). I. larkini is the next largest species, with the holotype having a total length of 220 cm, although Lomax and Massare (2017) indicated the species probably reached a total length of < 250 cm. I. somersetensis is the largest species, with the largest complete referred specimen (NHMUK OR2013*) having a total length of 270 cm. Lomax and Massare (2017), however, estimated the total length of the species as no greater than 300 cm.
McGowan and Motani (2003) gave the maximum length of the largest Ichthyosaurus as 250 cm. However, Massare et al. (2015) estimated a body length, from tip of the snout to the tail bend, of just under 300 cm for the genus based on an isolated forefin with a humerus length of 11.7 cm. The specimen was not assigned to a species of Ichthyosaurus because it was too incomplete. The total length of this individual was re-estimated as just over 300 cm from the tip of the snout to the end of the tail (Massare and Lomax 2017b), which still is the largest estimate for the genus until now.
However, some very large examples of Ichthyosaurus have been reported from Europe, including an incomplete ichthyosaur skull (SMF 46) from the lower Sinemurian of Frick, Switzerland (Maisch et al. 2008) and a fragmentary skull (IRSNB R145) from the upper Sinemurian of Bonnert, Belgium (Godefroit 1996). Massare et al. (2015) suggested that the Frick specimen could not be assigned to Ichthyosaurus, whereas the Bonnert specimen was identified as cf. Ichthyosaurus. This is contrary to Massare and Lomax (2017a), who erroneously referred both specimens to the genus. We agree with Massare et al. (2015), pending detailed examination of the European specimens.
NLMH 106234 has a skull length of 57.5 cm, which is the largest skull of an Ichthyosaurus on record (compare McGowan 1974; Massare et al. 2015) and with an estimated total length of 300–330 cm, the specimen is the largest, unequivocal example of Ichthyosaurus known. Furthermore, it extends the total body length estimate for I. somersetensis to < 350 cm.
Systematic palaeontology
Order Ichthyosauria de Blainville, 1835
Family Ichthyosauridae Bonaparte, 1841
Genus Ichthyosaurus De la Beche and Conybeare, 1821
Type species: Ichthyosaurus communis De la Beche and Conybeare, 1821; upper Hettangian–lower Sinemurian, Lower Jurassic of England, UK.
Ichthyosaurus somersetensis Lomax and Massare, 2017
Referred material.—NLMH 106234, a virtually complete, articulated skeleton lying on its left side, and missing almost the entire tail (see Material; Fig. 2). From Doniford Bay, Watchet, Somerset, UK. It was recovered from the Lower Jurassic (lower Hettangian) Blue Lias Formation, specifically the Caloceras johnstoni Ammonite Subzone of the Psiloceras planorbis Ammonite Zone (Bed 36).
Emended diagnosis.—As in Lomax and Massare (2017) with the following changes: total length > 300 cm but < 350 cm; hindfin with three or four elements in third row, with one element (tarsal 3) in broad contact with astragalus; notching usually present in some elements of the leading edge of the hindfin, but absent in the tibia.
Description.—The specimen can be assigned to I. somersetensis, because it possesses the following autapomorphies: a broad, asymmetric maxilla with a fairly long anterior process, and delicate, very long, and slender posterior process in lateral view; a triradiate lacrimal with a posterior shelf at the base of the dorsal process; a jugal dorsal ramus that is only slightly curved, lacking a right angle dorsal bend; and an ilium that is wide relative to its length and more oblong than rib-like (Lomax and Massare 2017). The specimen also possesses the unique combination of characters found in I. somersetensis, including: a prefrontal that excludes the dorsal process of the lacrimal from the orbit margin; the anterior process of the jugal extends slightly beyond the anterior inner edge of the orbit; the premaxilla supranarial and subnarial processes are about equal in length, extending about half way across the dorsal and ventral margins of the external naris, with the nasal making up about half of the dorsal margin; and a humerus that is long relative to its width, with a small dorsal process that does not extend far down the shaft (Lomax and Massare 2017).
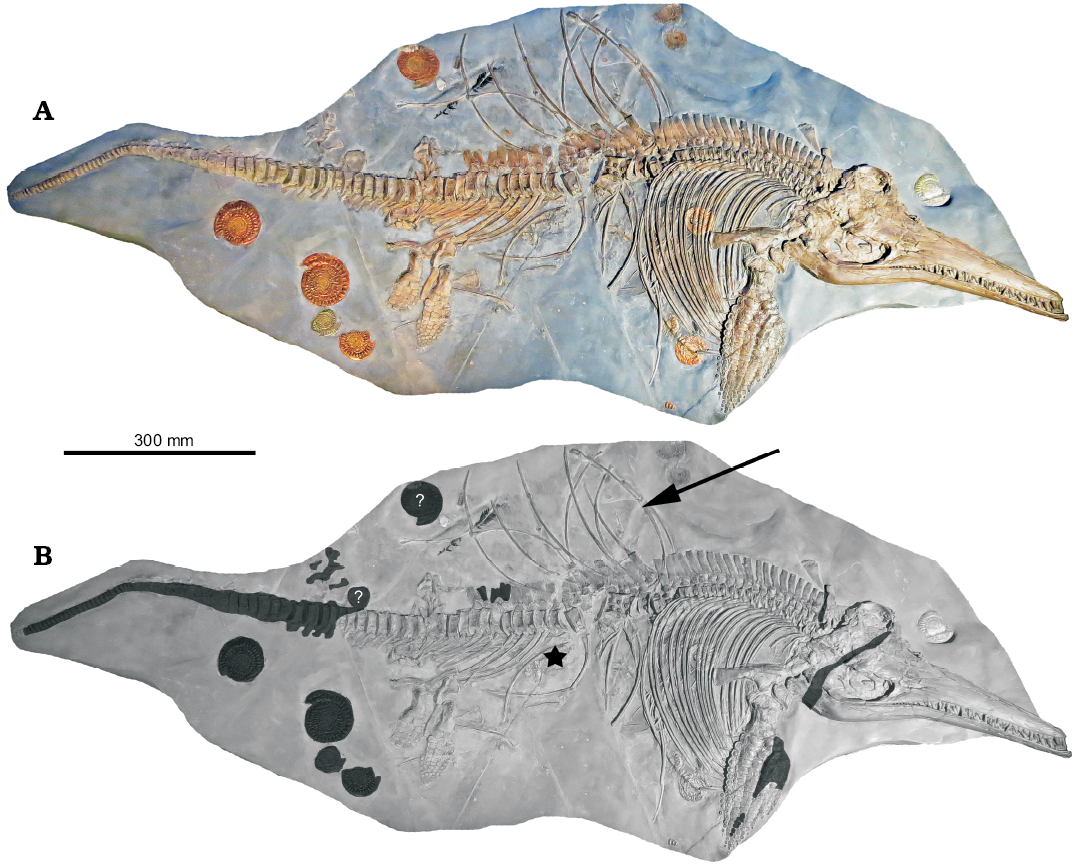
Fig. 2. A. Skeleton of Ichthyosaurus somersetensis Lomax and Massare, 2017 (NLMH 106234) from the Lower Jurassic (lower Hettangian) of Doniford Bay, Watchet, Somerset, UK. B. Explantory photograph with the composite/reconstructed sections shown in black; the question marks indicate that the element may have been added to the specimen; arrow points to a large crack that runs across the specimen; star indicates the position of the embryo.
Skull, mandible, and dentition: The skull is 57.5 cm long, and is preserved in dorsolateral view (Fig. 3). It is complete, but damaged posteriorly and dorsoventrally crushed, with some bones missing and others difficult to identify. The skull has also been restored in the posterior region, where a crack runs across the skull and mandible (Figs. 2B, 3). The orbit is crushed and the shape is not original, although a complete sclerotic ring is preserved and does not fill the orbital margin. Unless otherwise stated, the skull morphology concurs with other specimens of the species.
The skull roof is damaged and crushed, but some information can be gleaned. A low medial ridge is formed by the parietals and the lateral border of the temporal opening is formed largely by the supratemporal. The contact of the latter with the parietal and postfrontal is indiscernible.
An element that is probably the squamosal is displaced posterior to the postorbital and separated by a large crack. This element is roughly rectangular and positioned ventral to the supratemporal. Irrespective of crushing, the prefrontal forms a small portion of the dorsal and at least half of the anterior margin of the orbit. The prefrontal extends ventrally to the level of the external naris, excluding the dorsal process of the lacrimal from the anterior margin of the orbit.

Fig. 3. Skull of Ichthyosaurus somersetensis Lomax and Massare, 2017 (NLMH 106234) from the Lower Jurassic (lower Hettangian) of Doniford Bay, Watchet, Somerset, UK; in dorsolateral view. Arrow points to the reconstruction that extends across the skull.
The lacrimal is triradiate, similar to the condition in the holotype of the species (ANSP 15766), but the dorsal process is larger and more robust in NLMH 106234. This difference may be due to the crushing or represent individual variation. There appears to be a shelf at the base of the lacrimal, which is found in all examples of the species (Lomax and Massare 2017), although this is somewhat obscured due to the crushing. The dorsal process of the lacrimal makes up the posterior margin of the external naris and the anterior process of the lacrimal, although broken, forms about half of the ventral border of the external naris. The posteroventral process of the lacrimal makes up less than half of the anterior margin of the orbit. A thickened anterior process of the jugal extends slightly beyond the anterior margin of the orbit (although some of it may be buried by the maxilla) and alongside the posteroventral process of the lacrimal. The dorsal ramus of the jugal is damaged, but has a robust shape and clearly lacks a dorsal bend, as is characteristic for I. somersetensis (Lomax and Massare 2017).
In dorsal view, the nasal is wide posteriorly, with the anterior-most portion extending as far forward as the anterior process of the maxilla. In lateral view, the nasal forms about half of the dorsal border of the external naris. The supranarial and subnarial processes of the premaxilla make up at least half of the external naris borders. The maxilla is very large, as in all specimens of the species, and its maximum (dorsoventral) height is about even with the mid-posterior edge of the external naris. The maxilla is asymmetric in lateral view, with a fairly long and abruptly narrowing anterior process that extends beyond the external naris. The posterior process of the maxilla is delicate, very long, slender, and extends well under the orbit.
In lateral view, the angular forms a small portion of the mandible and the anterior end extends forward to about even with the posterior edge of the orbit. The surangular, however, extends farther forward than the angular, as far anteriorly as the highest point of the maxilla. The posterior end of the dentary ends abruptly, about half-way across the orbit, where it overlaps the surangular.
The teeth have largely slender crowns with longitudinal striations. Most of the roots are continuous with the crown, although some of the roots are wider than the crown. The roots have coarse, longitudinal grooves. The posterior maxillary tooth crowns are much smaller than the premaxillary teeth.
Axial skeleton: There are at least 40 precaudal centra, including the atlas-axis, although six are probably missing (see Material and methods). At least one of the posterior dorsals or anterior caudals, probably the latter based on shape, is disarticulated and positioned beneath some of the caudal ribs. There are eight caudal vertebrae preserved in articulation, and an isolated caudal centrum in the matrix. The isolated caudal centrum may not belong with the specimen as it appears smaller than the preserved caudal vertebrae, although it could conceivably be from a more posterior section. In addition, the last two caudal vertebrae in articulation are partially reconstructed and all of the vertebrae beyond that point have been added to the specimen and are not discussed further (see Material and methods; Fig. 2).
The neural spine apices of most of the cervical through mid-dorsal neural spines appear to have been “carved” during preparation, as the tips are squared and differ in colour from the rest of the neural spine. However, it is difficult to confirm whether this is an artefact of preparation or genuine. Regardless, the portions of the neural spines that are definitely genuine are at least twice as high as the centrum height. The posterior dorsal and anterior caudal neural spines are unusual in having a V-shape notch at the tip (Fig. 5), which is uncommon in Ichthyosaurus. This morphology is genuine, although some of the neural spines have been reconstructed to mirror the morphology (Fig. 5). The neural spines of the posterior dorsals and anterior caudals are also wider and about the same height as the centra.
The longest rib measures 49 cm along its curvature, but the distal end is buried.
Pectoral girdle and forefin: Most of the pectoral girdle is obscured by matrix. However, the complete right scapula is preserved and visible in lateral view. It is a long, slender element, measuring 15.5 cm along the long-axis. The anterior end is wide and the shaft is narrow, whereas the posterior end is slightly flared, as is typical of all species of the genus (Massare and Lomax 2017a).
The right forefin is preserved in dorsal view, but has been partially reconstructed (see Material and methods; Fig. 4). The humerus is crushed, in that a portion of the proximal region has been displaced posteriorly and some of the shaft surface is missing. The proximodistal length of the humerus measures 10.4 cm, its proximal and distal widths are about equal and the dorsal process is small. There is an irregular depression on the head, ventral and slightly anterior to the dorsal process. The dorsal process does not form a prominent ridge. The depression appears deeper than is typical of Ichthyosaurus somersetensis (Lomax and Massare 2017: fig. 7B), but this could be due to crushing, which gives the appearance of a tall proximal region (Fig. 4). Alternatively it might be a pathological deformation (Rothschild and Storrs 2003). About midway down the shaft is a slight expansion of the anterior side. This may be an artefact of crushing. The anterodistal end of the humerus is expanded slightly, but there is no anterior facet.
There are at least five primary digits, identified by the presence of an anterior digital bifurcation (Fig. 4). Irrespective of the reconstruction, the bifurcation is present in digit II, at the third phalangeal row. The elements of the bifurcation are much smaller, more circular than the other digits, as is often seen in some specimens of Ichthyosaurus, including in the holotype of I. somersetensis (DRL personal observation of ANSP 15766). A posterior accessory digit contacts the ulna and extends to almost the distal end of the fin. A second posterior accessory digit is present at the fifth phalangeal row. Digit V is prominent, as in all specimens of the genus. Although a large portion of the forefin is reconstructed, it is clear that the intermedium contacts both distal carpals 3 and 4. This contact rules out any suggestion of the specimen being an example of Protoichthyosaurus (Appleby 1979; Lomax and Massare in press).

Fig. 4. Right forefin of Ichthyosaurus somersetensis Lomax and Massare, 2017 (NLMH 106234) from the Lower Jurassic (lower Hettangian) of Doniford Bay, Watchet, Somerset, UK; in dorsal view (anterior to the right). Grey indicates plaster filler (elements are not genuine); black indicates crushed and displaced portion of humerus. Arrow points to bifurcation. Photograph (A), explanatory drawing (B).
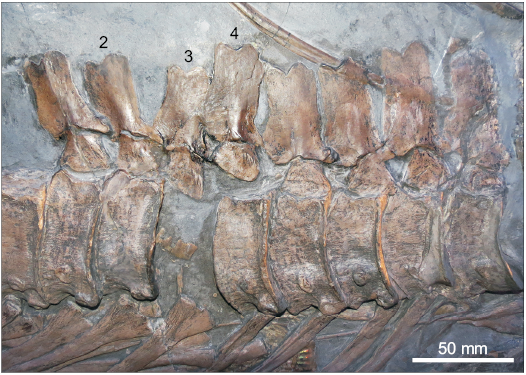
Fig. 5. Close-up of some mid-posterior dorsal vertebrae of Ichthyosaurus somersetensis Lomax and Massare, 2017 (NLMH 106234) from the Lower Jurassic (lower Hettangian) of Doniford Bay, Watchet, Somerset, UK; showing the unusual V-shape morphology of the neural spine apices. Note, the second, third, and fourth neural spine from the left are mostly reconstructed and the morphology is not genuine.
Pelvic girdle and hindfin: The pelvis is tripartite, as in all species of Ichthyosaurus (Lomax and Massare 2017; Fig. 6). Both ilia are present and probably exposed in lateral view. The right ilium, identified as that closest to the skull, is damaged proximally, thus the description is based upon the left ilium (Fig. 6). The ilium is more oblong, than rib-like, as in other species of the genus (Lomax and Massare 2017). It is slightly curved, with the concave side presumably facing posteriorly, although the orientation of the element makes this difficult to confirm. It is marginally shorter than the ischium, but about the same length as the pubis. The ilium has a fork-like proximal end, which is an unusual trait that, to our knowledge, has not been reported in any other Lower Jurassic ichthyosaur. A lateral ridge runs from the middle of the ilium to the proximal end, where it is flared. This ridge is also present on the damaged right ilium. The central portion of the fork-like proximal end is separated from both lateral ridges and extends further proximally. This morphology has not previously been reported in any species of Ichthyosaurus, although the lateral ridge is reminiscent of the ilium in BRSMG Ce 16611, another Ichthyosaurus specimen with an embryo. It is possible that this morphology may be due to pathology. The ischium is the longest of the pelvic elements. It is elongate and robust, compared to either the ilium or pubis, and is only slightly flared proximally. The pubis has a very narrow shaft with a widely expanded, “fan” shaped, distal end. This pubis morphology is present in several examples of Ichthyosaurus spp., including BRSMG Ce16611 and NHMUK R3372, which both contain embryos.
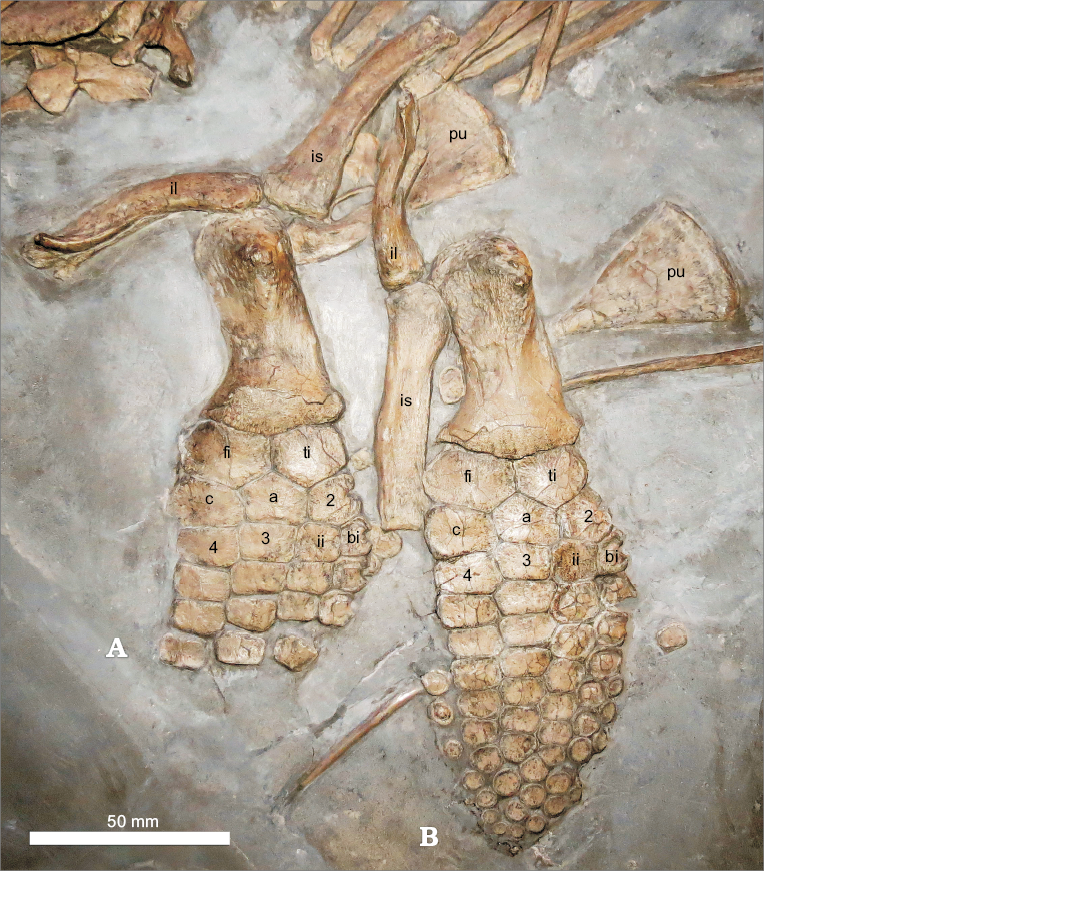
Fig. 6. Hindfins and pelvis of Ichthyosaurus somersetensis Lomax and Massare, 2017 (NLMH 106234) from the Lower Jurassic (lower Hettangian) of Doniford Bay, Watchet, Somerset, UK. The right hindfin (B) is the more complete of the two, exposed in dorsal view. The left hindfin (A) is in ventral view. Abbreviations: 2, tarsal two; 3, tarsal three; 4, tarsal four; a, astragalus; bi, bifurcation; c, calcaneum; fi, fibula; ii, metatarsal 2; il, ilium; is, ischium; pu, pubis; ti, tibia.
Both hindfins are preserved. The more complete, preserved closest to the skull and identified as the right, is exposed in dorsal view, whereas the left hindfin is exposed in ventral view. The femur is long relative to its distal width. Its anterodistal end has an anterior facet and the posterodistal end is expanded posteriorly. The dorsal process is offset anteriorly. The ventral process is more centrally located, but is slightly offset anteriorly. The fibula is anteroposteriorly wider than the tibia but is proximodistally about the same length. Tarsal 2, the first element of the bifurcation, and the first two phalanges are notched, although the shape of the notch differs, as has been reported in some specimens of Ichthyosaurus (Massare and Lomax 2016a: fig 10). There is one element (tarsal 3) in broad contact with the astragalus. However, a bifurcation of tarsal 2 is present, which results in four elements in the third row. We identify the anterior branch of digit II as the bifurcation. A small portion of metatarsal 2 contacts the astragalus. This differs from the condition described by Lomax and Massare (2017), who found that I. somersetensis had only one element (tarsal 3) directly in contact with the astragalus, with three elements in that row, and a bifurcation in a more distal row. In NLMH 106234, a distal bifurcation is present at the fourth phalangeal row, similar to that seen in NHMUK OR2013*. A posterior accessory digit is also present in the right hindfin, but is incomplete.
Embryo: The embryo is positioned between the ribs, near the mid-posterior dorsal vertebrae, on the block posterior to the crack (Figs. 2B, 7). There are 23 centra preserved in the embryo, although only 16 are articulated or associated. Of the articulated section, the 11th vertebra, counted from the left, is poorly preserved and/or partly missing; it may have been restored (Fig 7A). The articulated vertebral column is 6.37 cm long. It is not possible to identify the exact portion within the column. However, some loose and isolated centra are very round and could belong to dorsal vertebrae. This may suggest that the articulated section also comprises dorsal centra, which would be consistent with the interpretation of the fin as a forefin. Numerous delicate ribs are preserved; some fragments are in articulation with the vertebral column whereas others are lying in the matrix. A very small partial fin is preserved, which is probably the forefin. The total length is 1.84 cm, but the humerus is missing or buried. Four primary digits are evident, although a fifth primary digit may be present, indicated by a possible distal bifurcation (Fig. 7B). It appears that the ?radiale, ?distal carpal, and ?metacarpal are notched. This may, however, be an artefact of preservation (Fig. 7). All of the forefin elements are highly cancellous, displaying a spongious texture. The rim of many of the phalanges possesses a “bottle-cap” like morphology, suggestive of poorly ossified bone, or perhaps calcified cartilage. Similar preservation is also present in very small examples of Ichthyosaurus (e.g., BU 5289). A large element, proximal to the fin, is too long to be the humerus and is probably the scapula, which is expanded proximally.
Stratigraphic and geographic range.—Hettangian (lowermost Jurassic), Blue Lias Formation of Somerset and possibly Dorset, UK (see Lomax and Massare 2017, for more details).

Fig. 7. Embryo of Ichthyosaurus somersetensis Lomax and Massare, 2017 (NLMH 106234) from the Lower Jurassic (lower Hettangian) of Doniford Bay, Watchet, Somerset, UK. A. Articulated vertebral column, isolated forefin, probable scapula (white arrow), ribs, and isolated centra. B. Close-up of the forefin. Arrows point to probable notching of the ?radiale, ?distal carpal, and ?metacarpal.
Discussion and conclusions
Despite NLMH 106234 being a composite, the specimen provides important new information on Ichthyosaurus somersetensis. In the diagnosis for I. somersetensis, Lomax and Massare (2017) stated that the species has one element (tarsal 3) in broad contact with the astragalus, with a total of three elements in that row (third row). In NLMH 106234, tarsal 3 is in broad contact with the astragalus but a bifurcation of tarsal 2 results in four elements in the third row (Fig. 6). Metatarsal 2 also contacts the astragalus. In addition, both the proximal element of the bifurcation and metatarsal 2 are smaller than tarsals 3 and 4. This has not been reported for the species so far, and shows that the hindfin of I. somersetensis is more variable than previously thought. This hindfin morphology, however, is present in other species of the genus as well (Lomax and Massare 2017; DRL personal observation) and so hindfin morphology must be used in conjunction with other features to assign a specimen to I. somersetensis.
The ilium morphology of NLMH 106234 is unusual and may reflect individual variation, pathology, or could be size related. It is possible that this morphology is present in other specimens, but has gone unnoticed because the ilium must be well-preserved and exposed in a particular orientation in order to see the morphology. For example, in the holotype of I. somersetensis (ANSP 15766) the ilium is overlain by ribs and the proximal end cannot be examined in detail. Regardless, the morphology of the ilium in NLMH 106234 provides new information.
Only two other Lower Jurassic genera have been reported with embryos. Stenopterygius, which is known from multiple specimens (McGowan 1979) and Leptonectes, which is known from a single specimen (Lomax and Massare 2012). NLMH 106234 is the third Ichthyosaurus specimen from Somerset to be found with an embryo and the first to be positively identified to species level. Although, the other two specimens with embryos (NHMUK R3372 and BRSMG Ce16611) may also be I. somersetensis. Each of the three gravid females have just one embryo. This may be because remains of other embryos have not been preserved or were unknowingly removed during preparation. Alternatively, the presence of a single embryo may suggest that Ichthyosaurus carried just one embryo. But without more specimens, it is not possible to confirm this observation.
Lomax and Massare (2015) discussed possible sexual dimorphism in the humerus morphology of Ichthyosaurus anningae. One feature that they recorded was the presence of an anterior facet on the humerus of some specimens, which they hypothesized could be because male I. anningae required additional musculature in the forefin to position themselves during mating. Lomax and Massare (2015) also noted that the anterior facet is seen in other species, but not on all specimens. Clearly, NLMH 106234, BRSMG Ce 16611 and NHMUK R3372 are females. All three specimens lack an anterior facet on the humerus. In addition, Lomax and Massare (2017) noted that some, but not all specimens of Ichthyosaurus, have a pubis with a very broad and asymmetric, almost “fan” shaped, distal end. They suggested that this may also be a sexually dimorphic character. All three gravid females of Ichthyosaurus have this pubis morphology. Thus, the lack of an anterior facet in the humerus and an identically shaped, distally broad pubis in all three specimens lends support for these characters being sexually dimorphic.
The length of the skull, jaw and estimated total body length of NLMH 106234 are the largest published records of an unequivocal example of Ichthyosaurus. It is also the hitherto largest example of I. somersetensis known.
Acknowledgements
We would like to thank Annette Richter (NLMH) for access to and discussion of NLMH 106234, Elijah Widman (NLMH) for discussing the conservation of the specimen, and Kerstin Schmidt (NLMH) for taking photos of the fossil plant. Thanks also to Peter Langham (Dorset, UK) for providing DRL with additional information about the collection and history of the studied specimen. Thanks also to Paul Kenrick (NHMUK) for confirming the identification of the fossil plant and Kevin Page (Plymouth University, UK) for confirming the identification of the ammonites and discussing the stratigraphic position of the specimen. Thanks to Edward Daeschler and Edward Gilmore (both ANSP), Deborah Hutchinson (BRSMG), Jon Clatworthy (BU), Sandra Chapman and Paul Barrett (both NHMUK), and Carrie Eaton and David Lovelace (both UWGM) for access to and help with specimens in their collections. Katrin Sachs (Bielefeld Museum, Germany) is also acknowledged for her kind assistance with examining NLMH 106234 and co-creating some of the figures. DRL is grateful to Judy Massare (SUNY College at Brockport, NY, USA) for discussing the specimen and for assisting with the total length estimate, along with providing an early review of the manuscript. Furthermore, DRL wishes to acknowledge Judy Massare, Michael Maisch (University of Tübingen, Germany), and Valentin Fischer (University of Liège, France) for discussion of the ilium morphology. DRL also thanks William Wahl (Wyoming Dinosaur Center, Thermopolis, USA) for discussing the taphonomy and conservation of the specimen. DRL’s travel was covered in part by a Dean’s Doctoral Scholarship Award from The University of Manchester, and funding from Research Councils United Kingdom. Finally, we would also like to acknowledge Aubrey Roberts (University of Southampton, UK) and Benjamin Moon (University of Bristol, UK) for their helpful reviews and suggestions.
References
Appleby, R.M. 1979. The affinities of Liassic and later ichthyosaurs. Palaeontology 22: 921–946.
Bonaparte, C.L. 1841. A new systematic arrangement of vertebrated animals. Transactions of the Linnean Society of London 18: 247–304. Crossref
Buchholtz, E.A. 2001. Swimming styles in Jurassic ichthyosaurs. Journal of Vertebrate Paleontology 21: 61–73. Crossref
Blainville, H.M.D. de 1835. Description de quelques espèces de reptiles de la Californie, précédée de l’analyse d’un systéme general d’herpetologie et d’amphibiologie. Nouvelles Annales du Muséum d’Histore Naturelle 4: 236–296.
Deeming, D.C., Halstead, L.B., Manabe, M., and Unwin, D.M. 1993. An ichthyosaur embryo from the Lower Lias (Jurassic: Hettangian) of Somerset, England, with comments on the reproductive biology of ichthyosaurs. Modern Geology 18: 423–442.
De la Beche, H.T. and Conybeare, W.D. 1821. Notice of the discovery of a new fossil animal, forming a link between the Ichthyosaurus and crocodile, together with general remarks on the osteology of the Ichthyosaurus. Transactions of the Geological Society of London 5: 559–594. Crossref
Delair, J.B. 1969. A history of the early discoveries of Liassic ichthyosaurs in Dorset and Somerset (1779–1835). Proceedings of the Dorset Natural History & Archaeological Society 90: 1–9.
Delsett, L.L., Roberts, A.J, Druckenmiller, P.S., and Hurum, J.H. 2017. A new ophthalmosaurid (Ichthyosauria) from Svalbard, Norway, and evolution of the ichthyopterygian pelvic girdle. PLoS ONE 12: e0169971. Crossref
Godefroit, P. 1996. Un crâne d’Ichthyosaurus communis (Reptilia, Ichthyosauria) du Sinémurian supérieur de Lorrain belge. Bulletin de la Société belge de géologie 104: 77–90.
Lomax, D.R. and Massare, J.A. 2012. The first Leptonectes (Reptilia: Ichthyosauria) with associated embryos, from Somerset, England. Paludicola 8: 263–276.
Lomax, D.R. and Massare, J.A. 2015. A new species of Ichthyosaurus from the Lower Jurassic of West Dorset, England. Journal of Vertebrate Paleontology 35: e903260. Crossref
Lomax, D.R. and Massare, J.A. 2017. Two new species of Ichthyosaurus from the lowermost Jurassic (Hettangian) of Somerset, England. Papers in Palaeontology 3: 1–20. Crossref
Lomax, D.R. and Massare, J.A. (in press). The taxonomic utility of forefin morphology in Lower Jurassic ichthyosaurs: Protoichthyosaurus and Ichthyosaurus. Journal of Vertebrate Paleontology. Crossref
Lydekker, R. 1888. Note on the classification of the Ichthyopterygia (with a notice of two new species). Geological Magazine 5: 309–314. Crossref
Maisch, M.W., Reisdorf, A., Schlatter, R., and Wetzel, A. 2008. A large skull of Ichthyosaurus (Reptilia: Ichthyosauria) from the Lower Sinemurian (Lower Jurassic) of Frick (NW Switzerland). Swiss Journal of Geosciences 101: 617–627. Crossref
Massare, J.A. and Lomax, D.R. 2014. An Ichthyosaurus breviceps collected by Mary Anning: new information on the species. Geological Magazine 151: 21–28. Crossref
Massare, J.A. and Lomax, D.R. 2016a. A new specimen of Ichthyosaurus conybeari (Reptilia, Ichthyosauria) from Watchet, Somerset, England, U.K., and a re-examination of the species. Journal of Vertebrate Paleontology 36: e1163264. Crossref
Massare, J.A. and Lomax, D.R. 2016b. Composite skeletons of Ichthyosaurus in historic collections. Paludicola 10: 207–250.
Massare, J.A. and Lomax, D.R. 2017a. A taxonomic reassessment of Ichthyosaurus communis and I. intermedius and a revised diagnosis for the genus. Journal of Systematic Palaeontology [published online]. Crossref
Massare, J.A. and Lomax, D.R. 2017b. Correction to: A large forefin of Ichthyosaurus from the U.K., and estimates of the size range of the genus. Paludicola 11: 73–75.
Massare, J.A., Lomax, D.R., and Klein, A. 2015. A large forefin of Ichthyosaurus from the U.K., and estimates of the size range of the genus. Paludicola 10: 119–135.
McGowan, C. 1974. A revision of the latipinnate ichthyosaurs of the Lower Jurassic of England (Reptilia: Ichthyosauria). Life Science Contributions of the Royal Ontario Museum 100: 1–30. Crossref
McGowan, C. 1979. A revision of the Lower Jurassic ichthyosaurs of Germany with descriptions of two new species. Palaeontographica Abteilung A 166: 93–135.
McGowan, C. 2003. A new specimen of Excalibosaurus from the English Lower Jurassic. Journal of Vertebrate Paleontology 23: 950–956. Crossref
McGowan, C. and Motani, R. 2003. Handbook of Paleoherpetology, Ichthyopterygia. 175 pp. Verlag Dr Friedrich Pfeil, Munich.
Owen, R. 1881. Part 3. Ichthyopterygia. In: A Monograph of the Fossil Reptilia of the Liassic Formations, 83–134. Palaeontographical Society (Great Britain), London.
Pearce, J.C. 1846. Notice of what appears to be the embryo of an Ichthyosaurus in the pelvic cavity of Ichthyosaurus (communis?). Annals & Magazine of Natural History (First Series) 17: 44–46. Crossref
Rothschild, B.M. and Storrs, G.W. 2003. Decrompression syndrome in plesiosaurs (Sauropterygia: Reptilia). Journal of Vertebrate Paleontology 23: 324–328. Crossref
Acta Palaeontol. Pol. 62 (3): 575–584, 2017
https://doi.org/10.4202/app.00376.2017