
The oldest post-Paleozoic (Ladinian, Triassic) brachiopods from the Betic Range, SE Spain
JOSÉ FRANCISCO BAEZA-CARRATALÁ, FERNANDO PÉREZ-VALERA, and JUAN ALBERTO PÉREZ-VALERA
Baeza-Carratalá, J.F., Pérez-Valera, F., and Pérez-Valera, J.A. 2018. The oldest post-Paleozoic (Ladinian, Triassic) brachiopods from the Betic Range, SE Spain. Acta Palaeontologica Polonica 63 (1): 71–85.
Triassic brachiopods from the Betic Range were unknown hitherto. Herein we describe the first brachiopod occurrences in the early Ladinian of this domain referable to a new genus and species Misunithyris goyi derived from three localities of the south-Iberian Triassic outcrops. The analysis of internal and external characters of this brachiopod allowed to characterize systematically and biogeographically this fauna in a chronostratigraphic interval when the paucity of brachiopod records is attributable to the entire peri-Iberian epicontinental platform system established in the westernmost Tethyan margin. The new record is endemic to the Betic Range and represents a new faunal constituent of the multicostate zeillerids stock. This fauna inhabited the epicontinental seas of the Sephardic bioprovince since a closer affinity with the low-latitude Tethyan assemblages is revealed. The possible linkage of the Triassic stock with the Early–Middle Jurassic multicostate zeillerid representatives suggests feasible phylogenetic relationships between both groups.
Key words: Brachiopoda, Zeillerioidea, palaeobiogeography, Triassic, Ladinian, Spain.
José F. Baeza-Carratalá [jf.baeza@ua.es] and Fernando Pérez-Valera [fperez@ua.es], Departamento de Ciencias de la Tierra y Medio Ambiente, Universidad Alicante, Apdo. 99, San Vicente del Raspeig, 03080 Alicante, Spain.
Juan A. Pérez-Valera [jalbertoperez@geo.ucm.es], Departamento de Paleontología, Facultad de Ciencias Geológicas, Universidad Complutense, C/ José Antonio Novais 2, 28040 Madrid, Spain.
Received 31 July 2017, accepted 27 October 2017, available online 29 January 2018.
Copyright © 2018 J.F. Baeza-Carratalá et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Triassic Period represented one of the most significant episodes in the evolutionary history of brachiopods, inasmuch as severe biotic turnovers took place between the Permian–Triassic extinction event and the end-Triassic extinction boundary, two of the big five extinctions during Phanerozoic (Raup and Sepkoski 1982). Most of the diverse and abundant Paleozoic brachiopod faunas became extinct in the end-Permian extinction (Gould and Calloway 1980; Sepkoski 1996). After this biotic crisis, marine benthic biota underwent a new radiation in the late Early–Middle Triassic (cf. Hallam and Wignall 1997; Payne et al. 2004), contributing to the definitive replacement by the new evolutionary Mesozoic fauna. Brachiopods recovery started in the Olenekian–early Anisian (Chen et al. 2005, 2015; Shen et al. 2006; Zakharov and Popov 2014; Ke et al. 2016; Gaetani 2016), and experienced a new peak at community-level for the brachiopod clades during the Mesozoic (cf. Ager 1965; Sandy 1995; Dulai 2001; Tomašových 2006, and references therein).
In this timespan, brachiopod communities proliferated in the shallow epicontinental seas in several paleogeographical domains of the Paleo- and Neo-Tethys Ocean (e.g., Bittner 1890; Siblík 1972, 1998; Dagys 1974; Vörös and Pálfy 1989; Pálfy 1991, 2003; Pálfy and Török 1992; Senkowiczowa and Popiel-Barczyk 1993; Kaim 1997; Torti and Angiolini 1997; Hagdorn and Sandy 1998; Benatov 2001; Feldman 2005, 2013), but in the westernmost Tethyan basins the record is very scarce. Prior to the widely accredited Early Jurassic bloom in brachiopod diversity (e.g., Jiménez de Cisneros 1923; Baeza-Carratalá 2011, 2013; Baeza-Carratalá and García Joral 2012), no other Mesozoic brachiopods have been reported from elsewhere in the Betic Range hitherto. The only pre-Jurassic brachiopod evidence in this area is restricted to the indefinite exceptional record of a few cross-sections in the Early Devonian of the Nevado-Filábride complex (Laborda-López et al. 2015).
In the South-Iberian Paleomargin, collecting effort in numerous Triassic localities over the last decades resulted in extensive collections of Ladinian nautiloids and ammonoids (Pérez-Valera 2015; Pérez-Valera et al. 2017). In contrast, only three of these localities yielded brachiopod specimens suitable for taxonomy and having appropriate stratigraphic framework based on ammonoids (Pérez-Valera 2015; Pérez-Valera et al. 2017). It has also substantiated the record of brachiopods in the Triassic from the Betic Range for the first time, resulting in a new genus and species, allowing detailed descriptions of its external and internal structure.
This record in the Betic Range is also remarkable because it makes available new data among the sparse brachiopod occurrences in this chronostratigraphic interval. In fact, this paucity of brachiopod records is not confined to the Betic Range, but is attributable to the entire peri-Iberian epicontinental platform system established in the westernmost Neo-Tethys margin. Brachiopod occurrences into these Triassic epicontinental seas around Iberia have only been reported in the Iberian Range through the monospecific Lingularia bursts (Márquez-Aliaga et al. 1999, 2007; Escudero-Mozo et al. 2015; Giannetti et al. 2017), and four specimens attributed to Coenothyris sp. (Márquez-Aliaga et al. 1994; Escudero-Mozo et al. 2015) and C. vulgaris (Schmidt 1935), all of them mainly Ladinian in age, plus the sporadic Anisian Mentzelia (M. mentzeli, M. sp.), and Koeveskallina koeveskalyensis, mainly from Menorca and the Catalonian Coastal Range (Schmidt 1935; Escudero-Mozo et al. 2015).
Finally, this study enables the establishment of palaeobiogeographical affinities, by analyzing the similarity of the recorded fauna with those assemblages from several well-documented basins from the Germanic, Tethyan, and Sephardic bioprovinces.
Institutional abbreviations.—DCTMA, Earth and Environmental Science Department collections, University of Alicante, Spain.
Other abbreviations.—L, length; W, width.
Material and methods
Amidst the profuse Triassic outcrops prospected in the Betic Range by the authors, only a total number of six brachiopods were collected and studied from three localities (Fig. 1). Taxonomic supra-generic assignments follow the published determinations: mainly by Kaesler and Selden (1997–2007). Recent supplementary data (Manceñido 1993a; Baeza-Carratalá and García Joral 2014) are considered for the systematic arrangement into the zeilleriids. The main biometric parameters (in mm) and indices were measured for the taxonomical analysis when it was possible. Internal structure was studied using the conventional method of taking acetate peels after making transverse serial sections. The distance between serial sections was 0.1 mm. High resolution microphotographs of acetate peels were taken using an optical microscope (Nikon CFI60 E600POL). All specimens were coated with magnesium oxide prior to photographing. All specimens figured and acetate peels are deposited in the collections of Earth and Environmental Sciences Department (University of Alicante, Spain).
The ammonoid zonal scheme used is based on the standard zones proposed by Balini et al. (2010) modified by recent chronostratigraphic data from Pérez-Valera (2015) for the Betic domain. Paleobiogeographical analysis performed is plotted on a paleomap slightly modified after Pérez-López and Pérez-Valera (2007). It is worth noting that the paleobiogeographic relationships are inferred on the basis of the unique record in the Triassic Betic Domain hitherto; consequently the accuracy of the analysis would be improved if further brachiopod faunas were to be found in the future.
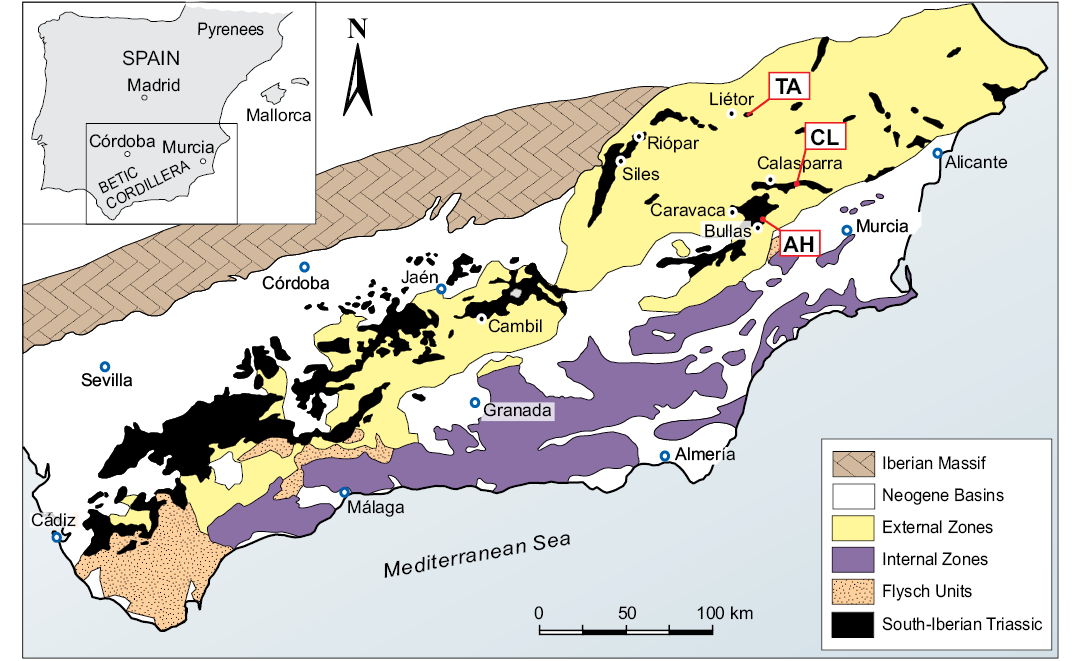
Fig. 1. Geographical and geological setting of the localities studied yielding Ladinian brachiopods in the Betic Range context emphasizing the South-Iberian Triassic outcrops. AH, Arroyo Hurtado section; CL, Calasparra section; TA, Talave section.
Geographical and geological setting
The Betic-Rif orogen is an extremely arcuate orogen that comprises the south of Spain and the north of Morocco, both connected by the so-called Gibraltar Arc, a structure formed in the Miocene (Platt et al. 1995; Balanyá et al. 2007). The north branch of this orogen is the Betic Range (Fig. 1), traditionally divided in two different domains: the Internal and External zones. Three tectonic complexes constitute the Internal Zones: Nevado-Filábride Complex, Alpujárride Complex, and Maláguide Complex, structurally stacked from bottom to top. These complexes are limited by major tectonic contacts and show varied metamorphism and structural features that indicate long-term and multifaceted trajectories during their emplacement as the internal part of the orogen (García-Dueñas et al. 1992; Lonergan and White 1997). These units come from the Alboran Domain, and are largely allochthonous.
On the other hand, the External Zones are composed of non metamorphic units that are actually forming the external fold-and-thrust belt of the Betic Range (Martín-Algarra and Vera 2004). The External Zones are formed by Triassic to the lower Miocene sediments deposited in the South-Iberian Palaeomargin. Conventionally, the External Zones are divided in two domains (Prebetic and Subbetic), through stratigraphic and paleogeographic significant changes after Lower Jurassic in the South-Iberian Paleomargin (García-Hernández et al. 1980), depending on their position with respect to the foreland: the Prebetic, more proximal and the Subbetic, more distal.
In the External Zones of the Betic Range, the classical tripartite Triassic crops out extensively and encompasses siliciclastic and evaporitic units of Buntsandstein and Keuper facies and epicontinental carbonates of Muschelkalk facies (Pérez-López 1998). All Triassic units have been included in the so-called “South-Iberian Triassic” (Fig. 1; Pérez-López and Pérez-Valera 2007). The carbonates of Muschelkalk facies consist of two coeval units, formally named the Siles Formation and the Cehegín Formation (Pérez-Valera and Pérez-López 2008), respectively representing the proximal and distal regions of the South-Iberian Palaeomargin in the Middle Triassic (Fig. 2).
Brachiopods studied in the present work derive from three localities (Talave, Calasparra, and Arroyo Hurtado sections) integrated in these two aforementioned units of the easternmost External Betic zones (Figs. 1, 2). These outcrops are located in the Murcia and Albacete provinces (SE-Spain, Fig. 1).
The stratigraphic arrangement and facies present in the Siles and Cehegín formations show the evolution of an epicontinental platform that was emplaced over the siliciclastic deposit of Buntsandstein facies (Arroyo Molinos Formation; Pérez-Valera F. 2005). Both formations are divided into two members (Fig. 2). The lower member is developed in a ramp during a transgressive stage, and the most characteristic feature is the presence of three laminated limestone beds (Pérez-Valera and Pérez-López 2008). The upper member is located over a regional unconformity that represents the maximum flooding surface, with presence of abundant cephalopods (ammonoids and nautiloids) and other representative fauna (Pérez-López et al. 2005). This member is constituted by a succession of marly limestone beds, lutites, thin bedded limestone and bioclastic limestone beds (tempestites) showing a progradational pattern in a high-stand stage (Pérez-Valera and Pérez-López 2008).
The brachiopod-bearing deposits are situated in the middle part of the upper member of Siles Formation (Talave section, Fig. 2), representing an equivalent stratigraphic position in the Cehegín Formation (Calasparra and Arroyo Hurtado sections, Fig. 2). Brachiopods have been found ocassionally together with bivalves, ammonoids, and nautiloids in a cm-thick alternation of marls and thin-bedded marly limestone levels (Fig. 2). In Talave and Calasparra sections, this part of the succession is situated over a bioclastic key-level that probably mark the end of a regressive parasequence and therefore, the alternation of marly limestone beds and marls with pelagic fossils (e.g., ammonoids) containing brachiopods, can represent the deeper part of the next parasequence, as proposed by Pérez-Valera and Pérez-López (2007). In the Arroyo Hurtado section, bioclastic levels with borings at the top mark the end of the regressive parasequences, and brachiopods have been found in levels together with ammonoids, developed over the bioclastic beds.
The recent finding of a large number of ammonoids has allowed a new biostratigraphical framework in the Muschelkalk carbonates of the Betic Range in which three ammonite biochronozones have been distinguished (Fig. 2), all belonging to the Ladinian stage (Pérez-Valera J.A. 2005; Pérez-Valera et al. 2011; Pérez-Valera 2015): (i) Eoprotrachyceras curionii Zone (lower part of Fassanian, lower Ladinian), which is subdivided into the Eoprotrachyceras curionii and Gevanites awadi subchronozones; (ii) Gevanites epigonus Zone (upper part of Fassanian, lower Ladinian); and (iii) Protrachyceras hispanicum Zone (lower part of Longobardian, upper Ladinian). This is consistent with the previous data provided by bivalves, conodonts, and nautiloids of Ladinian age (Márquez-Aliaga et al. 2001; Plasencia et al. 2007; Pérez-Valera et al. 2017). All the levels containing brachiopods in the three studied outcrops are situated in the Gevanites epigonus Zone (Fig. 2) and they can be therefore attributed to the uppermost Fassanian (lower Ladinian).
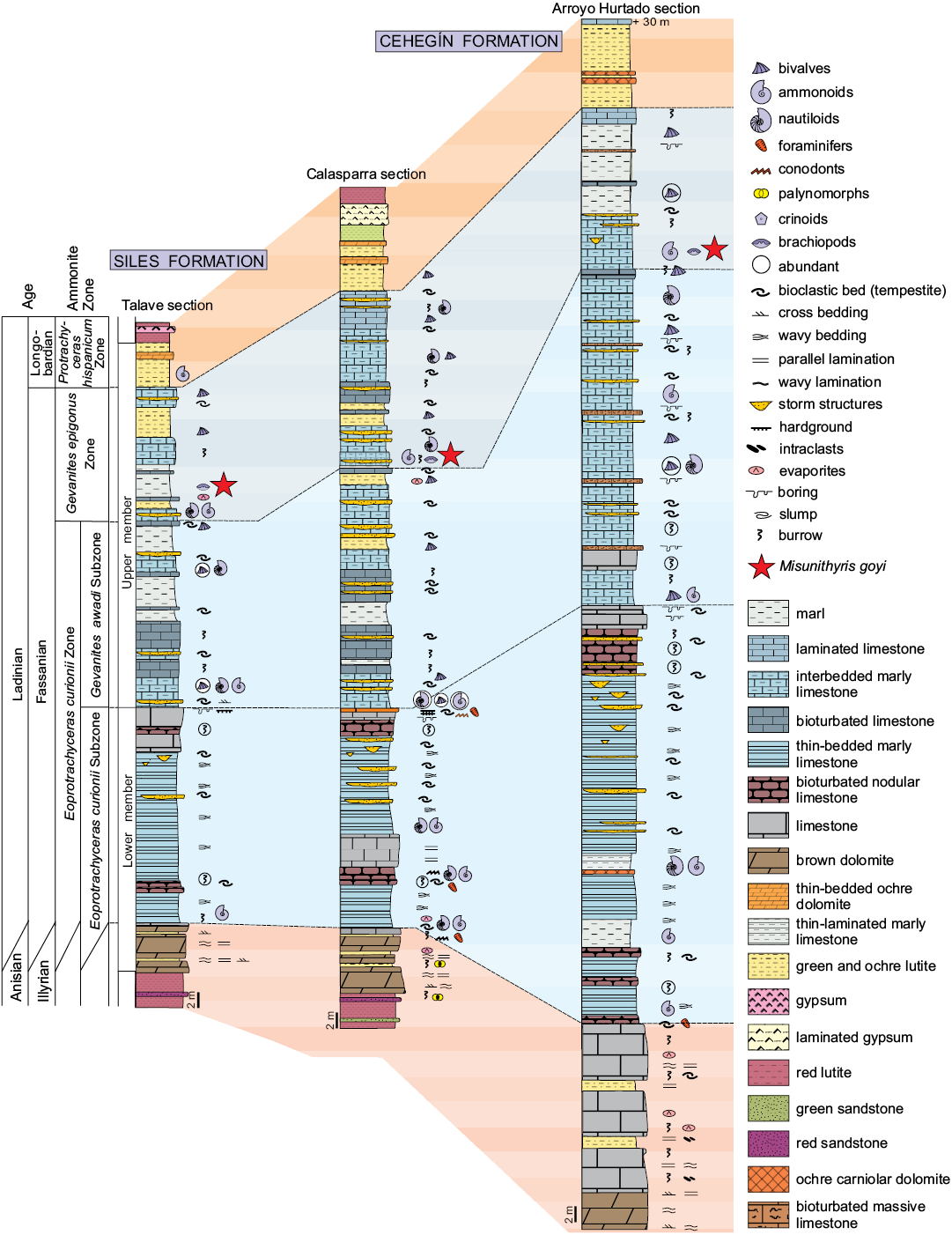
Fig. 2. Ladinian lithostratigraphical sections showing the occurrences of Misunithyris goyi in Talave, Calasparra, and Arroyo Hurtado sections. Ammonite zones and subzones after Pérez-Valera (2015).
Systematic palaeontology
Phylum Brachiopoda Duméril, 1805
Subphylum Rhynchonelliformea Williams, Carlson, and Brunton, 1996
Class Rhynchonellata Williams, Carlson, and Brunton, 1996
Order Terebratulida Waagen, 1883
Suborder Terebratellidina Muir-Wood, 1955
Superfamily Zeillerioidea Allan, 1940
?Family Zeilleriidae Schuchert in Schuchert and Le Vene, 1929
Genus Misunithyris nov.
Etymology: From Mīšūnīš, ancient name of the current Mundo River; in the Mundo River valley, the most significant outcrops of specimens from which this genus is erected are found.
Type species: Misunithyris goyi sp. nov. (by monotypy); see below.
Diagnosis.—As for the type species by monotypy.
Remarks.—The supra-generic systematic arrangement of Misunithyris is debatable, depending on the diagnostic criteria selected. Exceptional concurrence of several internal and external features can make attributable this genus to different groups within Terebratulida. On the one hand, the new genus herein erected shows common features with the superfamily Dielasmatoidea Schuchert, 1913, which includes some genera with enveloped dental plates and cardinal process. However, the assignment to Dielasmatoidea can be problematic because Misunithyris does not evidence a dielasmoid-type brachidium (sensu Dagys 1974 or Smirnova 2008). In this sense, supplementary elements such as median ridges or vertical plates (even not forming part in the development of brachidium, as stated by Dagys 1974 and Smirnova 2008) have not been observed in the internal structure of Misunithyris. Furthermore, some Dielasmatoidea representatives show septum-supported architectures and often short-looped developments (e.g., Adygella Dagys, 1959; Dielasmina Waagen, 1882; Tunethyris Calzada, Peybernes, Kamoun, and Youssef, 1994). In addition, crural bases are given off dorsally instead the distinctive crural progress revealed in Misunithyris.
Within Dielasmatoidea, higher similarity was expected with the anteriorly multicostate stock attributed to the Permian Dielasmina Waagen, 1882 and Hemiptychina Waagen, 1882 and the Permian–Triassic Costoconcha Jin, Sun, and Ye, 1979, since besides the anterior ribbed pattern, all genera share quite a few beak features and the presence of cardinal process, but the rest of the internal structure is totally different, mainly referred to the dental plates and the crural development, which is clearly a brachidium-supported structure in the first stages.
Another dielasmatoid morphotype, widely distributed and to some extent contemporary with Misunithyris is represented by Coenothyris Douvillé, 1879. The species of this Triassic genus display comparable dental plates, undeveloped or fused with the thickened shell wall, evident cardinal process, and long loop. Conversely, it evidences an initial septum-supported structure, notable septalium and, especially, the external features (such as smooth shell, often with strong uniplication) are entirely different (e.g., Popiel-Barczyk and Senkowiczowa 1989; Török 1993; Senkowiczowa and Popiel-Barczyk 1996; Kaim 1997; Pálfy 2003; Feldman 2005).
Arrangement within Zeillerioidea Allan, 1940 is the most plausible option, mainly because of the presence of dental lamellae, the well-developed and large dorsal median septum, and a clear zeilleroid-type brachidium (sensu Smirnova 2008) with a long-looped development, not connected to the median septum. However, some characters do not fully agree with the various families up to now determined in Zeillerioidea.
Misunithyris shares with the family Eudessidae Muir-Wood, 1965 the envelopment of dental plates, the presence of a cardinal process and most of the beak features. The most remarkable difference to Eudessidae are in a short dorsal median septum and the crural bases given off dorsally, as well as the usual growth of a median cardinal plate of the later group and not perceived in Misunithyris. Some external diagnostic criteria are very different as well, since even showing Eudessidae multicostate shells, Misunithyris shows a marginal ribbing pattern instead the entire multicostate shell-length of Eudessidae. Folding pattern is also unrelated as the conspicuous dorsal sulcus developed by Misunithyris is not shared with any Eudessidae representatives.
The affinities with the family Zeilleridae are found in several subfamilies. Some representatives of the subfamily Vectellinae Baker, 2006 exhibit a long-looped development and cardinal process as a knob or poorly developed callus. However, Middle–Late Triassic representatives of this subfamily are very different in both external and internal features to the new established genus. It is the case of Fletcherithyroides Dagys, 1977, Aulacothyroides Dagys, 1965, and Parantiptychia Xu and Liu, 1983, consisting on smooth morphotypes, also showing a long stage of crura supported by septal pillars. Probably the clearest affinity in this subfamily is found in the Upper Triassic?–lowermost Jurassic Tauromenia Seguenza, 1885, due to comparable anterior ribbing pattern and the beak features, as well as the well-developed dorsal median septum (Alméras et al. 2007; Baeza-Carratalá and García Joral 2012) and long-looped brachidium (Alméras et al. 2007).
Finally, there are several arguments for and against assigning Misunithyris to the subfamily Zeilleriinae Schuchert, 1929 as was defined in Kaesler and Selden (1997–2007). The septum is clearly well-developed and the brachidium evidences a long-looped progression. Conversely, the presence of cardinal process is atypical in this subfamily, although some genera show primitive lobes as massive callus or small knobs (e.g., Antiptychina Zittel, 1880; Kolymithyris Dagys, 1965). On the other hand, some of them also reveal enveloped dental lamellae instead the typical strong and unenveloped dental plates characteristic of this subfamily. As for Tauromenia, the closer external affinities are found in the anteriorly ribbed representatives of this subfamily, i.e., Calpella Owen and Rose, 1997 and Parathyridina Schuchert and Le Vene, 1929, the internal structure of which remains poorly known except for the presence of a prominent median septum (e.g., Cooper 1983; Baker 2006; Alméras et al. 2010b). In this sense, very close internal and external affinities have been recognized with the recent erected multicostate genus Menathyris Feldman, 2013, except for the cardinal process and the anterior folding pattern.
It must be kept in mind that the subfamily Zeilleriinae Schuchert, 1929 was recently split into several subfamilies by Baeza-Carratalá and García Joral (2014) on the basis of the hinge plates-crural bases relationship, thus resulting three subfamilies (Aulacothyrinae Babanova, 1964, Zeilleriinae Schuchert, 1929, and Securininae Baeza-Carratalá and García Joral, 2014). Attending to this criterion, Misunithyris clearly shows a Bakonyithyris-type pattern and might be arranged into Aulacothyrinae. However, the representatives of Aulacothyrinae (sensu Baeza-Carratalá and García Joral 2014) have not evidenced cardinal process so far, and the progress of dental plates is rather different.
Summarizing, the arrangement of Misunithyris into family Zeilleridae is the best plausible determination, but does not fully agree with the currently defined subfamilies. Combining external and internal features, it seems to be closer to the multicostate zeilleriid stock, such as Menathyris Feldman, 2013, Calpella Owen and Rose, 1997, or Tauromenia Seguenza, 1885. A new subfamily might be erected, either with Misunithyris as monotypic taxon, or together with the aforementioned genera, but further studies of internal structures in their representatives will be required to establish the validity of this approach, emphasizing examination of cardinal process and brachidium architecture in addition to the already known shared features.
Stratigraphic and geographic range.—Misunithyris is so far a monospecific genus recorded in the Gevanites epigonus Chronozone of the Fassanian (lower Ladinian, Middle Triassic) from the Betic Range (Fig. 2).
Misunithyris goyi sp. nov.
Fig. 3.
Etymology: In a tribute to Antonio Goy (Complutense University of Madrid), prominent Triassic and Jurassic palaeontologist, to whom the authors are very indebted for long term teaching and collaboration.
Type material: Holotype: DCTMA-BQ-TA1.2 (Fig. 3F; Table 1) from the Talave section. Paratype: DCTMA-BQ-AH2.2 (Figs. 3B, 4, 5). Intraspecific variability is shown through the paratypes (Fig. 3) and measured in Table 1.
Type locality: Talave section (External Betic Zone, SE Spain).
Type horizon: Upper member of Siles Formation, Fassanian (early Ladinian); Gevanites epigonus Biochronozone.
Material.—Six specimens (BQ-CL1.1, BQ-CL1.2, BQ-TA1.1, BQ-TA1.2, BQ-AH2.1, and BQ-AH2.2), including type material, with diverse state of preservation. All individuals show articulated but generally fragmented and encrusted valves. Internal recrystallization is visible in two specimens.
Diagnosis.—Medium-sized and ventribiconvex zeilleriid, pyriform in outline with strong sub-labiate beak and epithyrid/permesothyrid foramen. The lateral commissure is straight and the anterior one is clearly sulcate. Sharp ribs (8–13) appearing only near the anterior margin (3–5 in the median sulcus), often bifurcate and occasionally intercalate. Deltidial plates present and well-developed striate cardinal process. Crural bases are given off dorsally and hinge plates initially fused and subparallel. Dorsal septum long, exceeding the length of the brachidium. Long descending branches, distally with ventral progression.
Description.—External features (Fig. 3): Medium-sized ventribiconvex shells (Table 1), pyriform to subovoidal in dorsal outline. W/L ratio is rather variable, but all specimens show L > W. Thickness is about 3/5 of the length. Maximum convexity lies in the posterior third while maximum width lies in the anterior third of the shell. Maximum thickness is observed near the mid-length. The beak is massive, sub-labiate, and strong, erect to slightly incurved, with a medium-sized epithyrid to permesothyrid foramen; the beak ridges are well-perceived and blunt, developing short and narrow interareas. The lateral commissure is straight and the anterior one is clearly sulcate, with a wide and arcuate dorsal median sinus. Sulcation is generally shallow, being more pronounced in the larger specimens.
Ribbing pattern is distinctive of this species. Ribs are absent in the posterior third of the shell; costation consists of 8 to 13 ribs on each valve, 3–5 of which can be present in the median sulcus. Ribs are wide, strong, and square in cross-section, becoming stronger and sharper near the anterior margin. They are often bifurcate; the thicker specimens possess a set of weaker, intercalated ribs. Costation is more evident in the ventral valve and on the flanks of the dorsal one. In the median dorsal sulcus, ribs are only noticed from the mid-length onwards. Concentric, dense and strong growth lines are present on the entire surface giving a reticulate aspect to the shell.

Fig. 3. Intraspecific variability of zeillerid brachiopod Misunithyris goyi gen. et sp. nov. of the lower Ladinian from the South-Iberian Triassic platform, the Arroyo Hurtado (A, B), Calasparra (C, E), Talave (D, F) sections; in dorsal (A1–F1), anterior (A2–F2), and lateral (A3–F3) views. A. BQ-AH2.1. B. BQ-AH2.2, sectioned in the present work. C. BQ-CL1.1. D. BQ-TA1.1. E. BQ-CL1.2. F. BQ-TA1.2, holotype. All specimens were coated with magnesium oxide.
Internal structure (Figs. 4, 5): This species shows a sub-octagonal to ovoidal delthyrial cavity in cross-section where the remains of a short pedicle collar and well-developed deltidial plates are visible. Dental plates are difficult to distinguish as they are enveloped in a thickened-shell wall (Fig. 5). They are short and subparallel. Hinge teeth are massive with crenulations, inserted in broad, shallow, and also crenulated sockets; small denticula are also discernible. It has a noticeable cardinal process, striated and raised by a high cardinal platform where a central cavity is located under the myophore (Fig. 5). Hinge plates are initially horizontal, fused and subparallel. The dorsal median septum is long and well-developed up to 2/3 of the shell. A short elliptical septalium is discernible. Crural bases are located in the ventral area of the hinge plates but an incipient dorsal thickening emerge clearly showing an early dorsal development (Fig. 4), suggesting a Bakonyithyris-type hinge plate/crural base inter-relation (sensu Baeza-Carratalá and García Joral 2014). The descending branches of the brachidium show subparallell and vertical plates and they are developed in the commissural plane in the posterior third of the shell, acquiring a slight ventral development anteriorly. The transverse band is not visible.
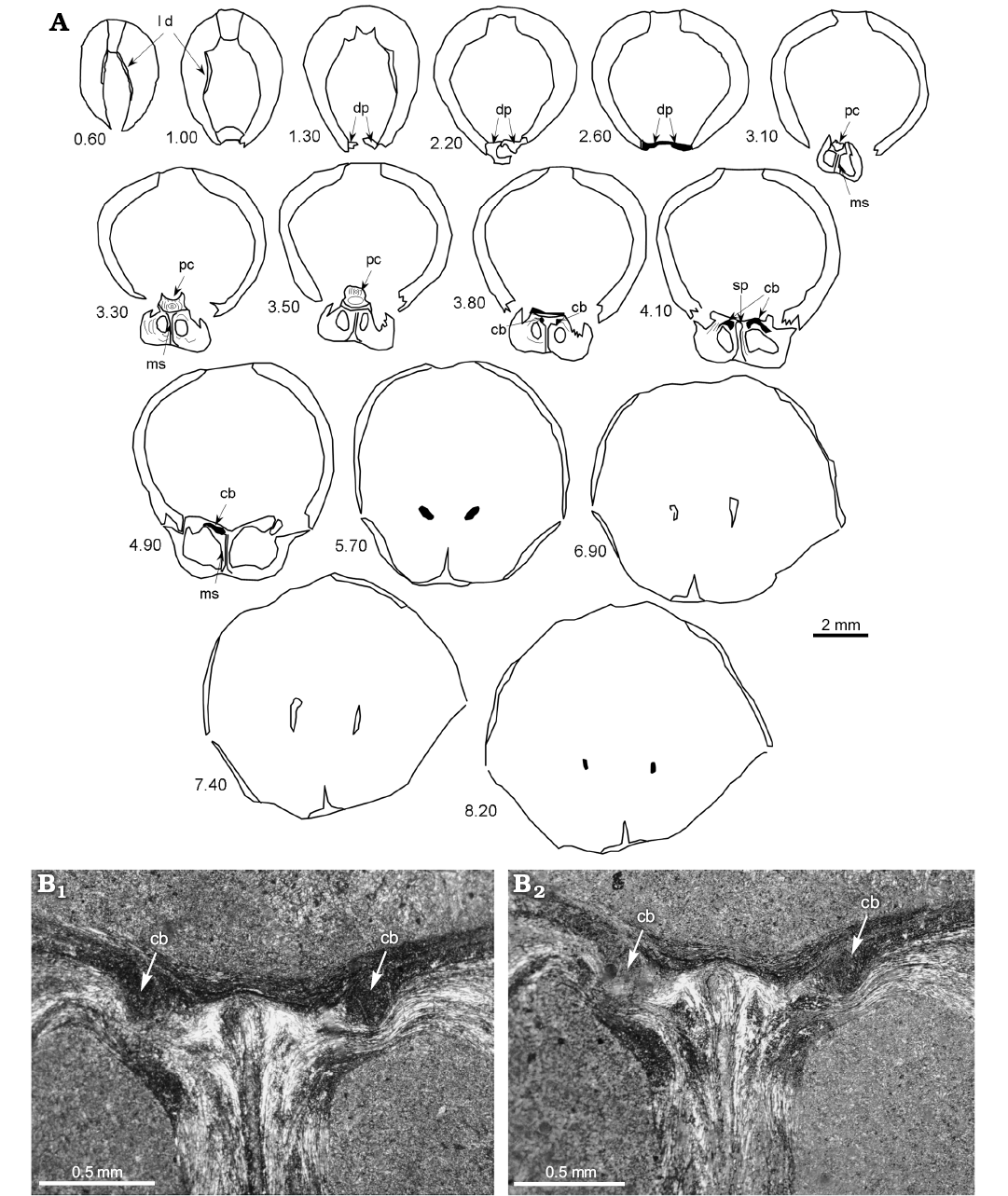
Fig. 4. Internal structure of zeillerid brachiopod Misunithyris goyi gen. et sp. nov. (BQ-AH2.2) from the lower Ladinian of the South-Iberian Triassic platform, the Arroyo Hurtado section. A. Transverse serial sections through the specimen (distance from the beak in mm). B. Microphotographs of acetate peels showing the hinge plates-crural bases relationship: crural bases are located in the ventral part of the hinge plates but a dorsal thickening emerges towards an early dorsal development. Section at 4.10 mm (B1) and 4.30 mm (B2) from the apex. Abbreviations: cb, crural bases; dp, deltidial plates; ld, dental lamellae; ms, median septum; pc, cardinal process; sp, septalium.
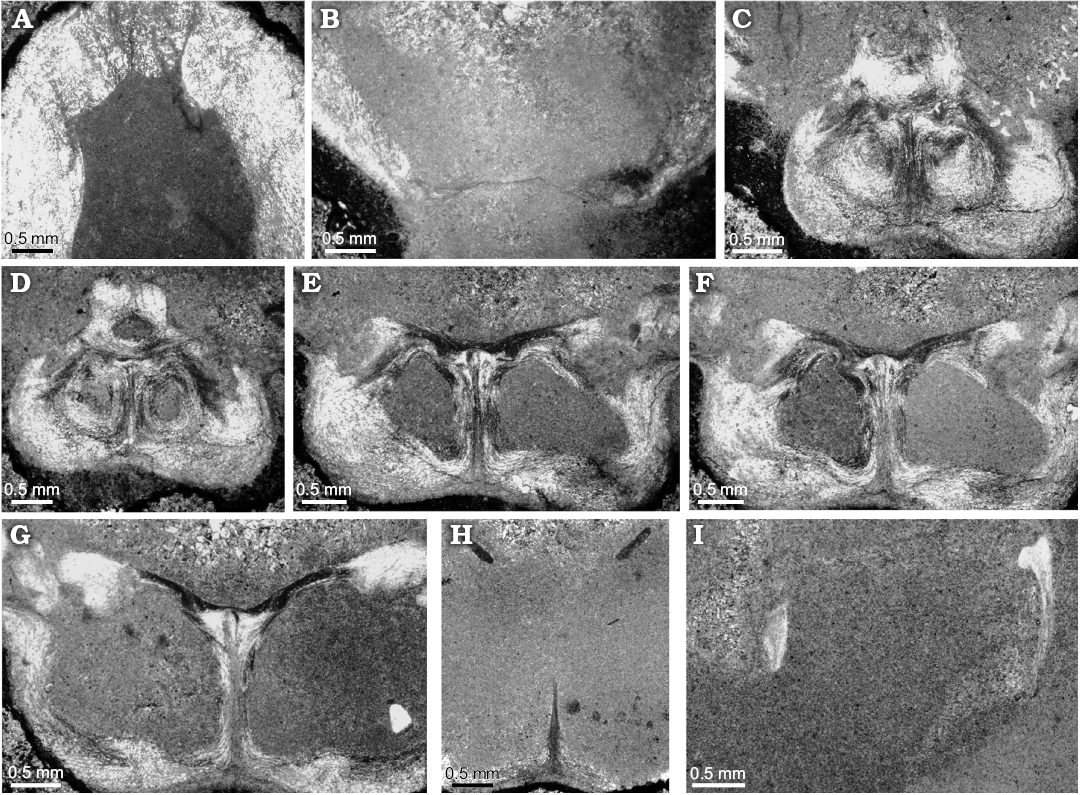
Fig. 5. Microphotographs of acetate peels from zeillerid brachiopod Misunithyris goyi gen. et sp. nov. (BQ-AH2.2) from the lower Ladinian of the South-Iberian Triassic platform, the Arroyo Hurtado section. A. Section at 1.00 mm from the apex showing dental lamellae enveloped in a thick-shelled wall. B. Section at 2.60 mm from the apex showing the disposition of deltidial plates. C, D. Sections at 3.30 and 3.50 mm from the apex, respectively, showing the first stages of hinge plates and earlier cardinalia and the evolution of the cardinal process, clearly striated and raised by a high cardinal platform. E, F. Sections at 4.10 and 4.30 mm from the apex, respectively, showing the cardinal area with the position of crural bases and the articulation system. G–I. Partial sections at 4.90, 5.70, and 7.10 mm from the apex, respectively, showing the evolution of the crural architecture and dorsal median septum development.
Remarks.—In addition to the main biometric ratios (Table 1), intraspecific variability mainly lies in the number of ribs present in each valve, depending on the bifurcation or intercalation. The smooth posterior stage is also variable between ½ and ¼ of the shell. A single specimen shows an exceptionally wide sulcus giving to the anterior commissure an almost rectimarginate aspect, but analyzing in detail this individual the sinus can be noticed from the early stages in the posterior areas of the shell. Thus, even width of sulcus is rather stable, it ranges from mid-width in the BQ-TA1.2 specimen up to wider developments (BQ-AH2.1 and BQ-CL1.1) occupying nearly the entire anterior margin. On the other hand, maximum width can be shifted toward the anterior third, resulting in a more trigonal dorsal outline.
Table 1. Main biometric measurements (in mm) and ratios of the studied specimens of Misunithyris goyi gen. et sp. nov. L, total length; W, total width; T, total thickness; R, ribs on each valve; Rp, ribbing pattern (p, primary rib; s, secondary rib, referred to bifurcate or intercalated ribs).
|
Specimen |
L |
W |
T |
W/L |
T/L |
T/W |
R |
Rp |
|
BQ-CL1.1 |
17.56 |
12.78 |
10.83 |
0.73 |
0.62 |
0.85 |
8 |
7p 2s |
|
BQ-CL1.2 |
13.00 |
12.44 |
8.20 |
0.96 |
0.63 |
0.66 |
9 |
6p 2s |
|
BQ-TA1.1 |
17.41 |
16.60 |
~8.44 |
0.95 |
0.48 |
0.51 |
9 |
9p |
|
BQ-TA1.2 |
28.34 |
24.43 |
17.01 |
0.86 |
0.60 |
0.70 |
13 |
9p 2s |
|
BQ-AH2.1 |
21.74 |
15.77 |
15.51 |
0.72 |
0.71 |
0.98 |
11 |
9p 4s |
|
BQ-AH2.2 |
16.64 |
13.54 |
~10.90 |
0.81 |
0.65 |
0.80 |
11 |
9p 2s |
As it is discussed above in the supraspecific assignment, this species shows high singularity in the external and internal traits, probably because it is an endemic taxon. The closer affinity is shown with the Lower Jurassic (mostly anteriorly) multicostulate zeilleriids Tauromenia polymorpha, T. brevicostata, and Fimbriothyris spp. (Seguenza 1885; Dubar 1942; Elmi et al. 2003; Alméras et al. 2007; Baeza-Carratalá and García Joral 2012), mainly recorded in the southern part of Western Tethys and even with the Middle Jurassic Eudesiidae representatives recorded in both Tethyan margins and the Mid-East platforms (e.g., Alméras 1987; Cooper 1989; Mukherjee et al. 2000; Alméras et al. 2010b), having in common with these last taxa several internal features (cardinal process present in several taxa, long median septum) but not sharing the brachidium architecture and the ribs covering the whole surface of the shell in the Middle Jurassic taxa.
Among the Triassic representatives, the closer affinities are found in Menathyris wilsoni Feldman, 2013 from Israel, virtually contemporary with the Betic material, sharing the sulcate shape of the shell, the marginal ribbing pattern and comparable internal structure, except for the cardinal process, absent in M. wilsoni; however, sulcus is shallower and narrower, anterior commissure is generally rectimarginate and ribs are simple without bifurcation or intercalation in such species. Curious similarity concerning the internal structure is observed in one specimen of Coenothyris with a central umbonal chamber in the cardinal process, similarly to the Betic material, and several Aulacothyroides species developing wide cardinal process, all of them depicted by Senkowiczowa and Popiel-Barczyk (1996) in the Anisian–Ladinian from Poland.
Stratigraphic and geographic range.—In the Betic Range, this new species has been recorded together with biostratigraphical marker of the Gevanites epigonus Biochronozone (Pérez-Valera F. 2005; Pérez-Valera et al. 2011; Pérez-Valera 2015) of the Fassanian (early Ladinian).
Discussion
Paleobiogeographical implications.—During the Triassic Period, brachiopod bioprovinces are mainly defined for the Late Triassic, where the taxonomic diversity was higher, being therefore the biochoremas more definite, with up to five high-rank biochoremas subdivided in several bioprovinces (Dagys 1993; Manceñido 2002). In the Ladinian, brachiopods were less diverse and their record is very rare in several regions and therefore their biogeographic differentiation pattern is unclear (Dagys 1993). In fact, in several Tethyan basins such as the Caucasus, brachiopods are totally absent during the Ladinian (Ruban 2006a, b) leading to postulate a severe crisis on brachiopod fauna likely as a results of an unsteady recovery after the end-Permian event (Ruban 2017).
The low-latitude Tethyan brachiopod faunas or Southern Tethyan Sub-Realm (cf. Dagys 1993; Manceñido 2002) comprise several minor-rank biochoremas usually referred not only for establishing the brachiopod fauna affinities but also for cephalopods, conodonts, bivalves and other groups (Hirsch 1987; Parnes et al. 1985; Pérez-López et al. 2003; Pérez-Valera 2015). Thus, the affinity of the faunal components from these Southern Tethyan basins can be discriminated, at least, into the Germanic, Tethyan, and Sephardic bioprovinces (Fig. 6).
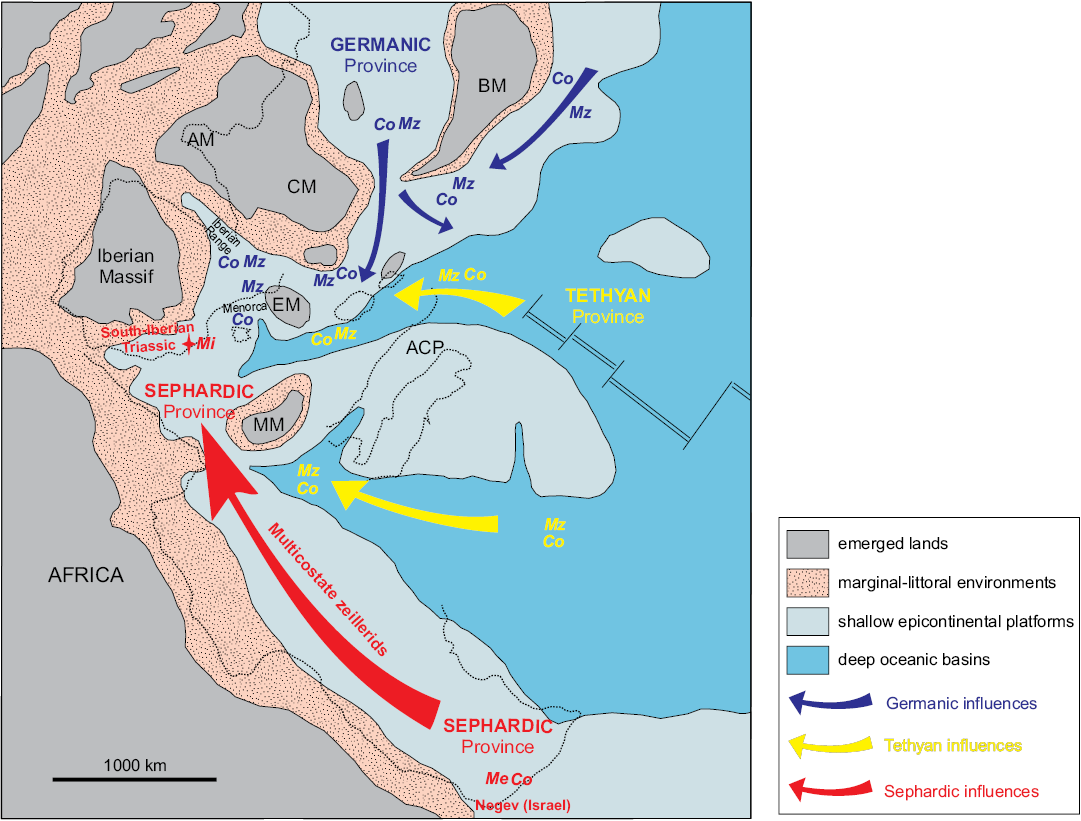
Fig. 6. Paleogeographical map of the Ladinian times (modified after Pérez-López and Pérez-Valera 2007), showing the distribution of taxa highly related to, or common with the brachiopod fauna inhabiting the Middle Triassic peri-Iberian platforms system. Color of arrows and taxa symbolizes the inferred paleobiogeographical affinities. ACP, Apennine Carbonate Platform; AM, Armorican Massif; BM: Bohemian Massif; CM, Central Massif; EM, Ebro Massif; MM, Meso-Mediterranean Massif; Mi, Misunithyris; Me, Menathyris; Mz, Mentzelia; Co, Coenothyris. Occurrences mainly based on Schmidt (1935), Siblík (1972, 1988, 2001), Popiel-Barczyk and Senkowiczowa (1989), Dagys (1993), Török (1993), Kaim (1997), Torti and Angiolini (1997), Pálfy (2003), Feldman (2005, 2013), Ruban (2010), Escudero-Mozo et al. (2015), among others (see text for details). Dotted line shows palaeogeographic position of the currently emerged land.
The attribution of the studied material to a new genus and species seems to point out to the presence of endemic brachiopod fauna in the Betic Range. On the other hand, independently from the argued supra-generic assignment of Misunithyris, this genus undoubtedly corresponds to an unusual and scarce morphotype within the multicostate zeillerids. The unique morphology of this group makes it a suitable tool for paleobiogeographic correlations and biostratigraphical calibrations (e.g., Ager and Walley 1977; Alméras 1987; Baeza-Carratalá and García Joral 2012). In this sense, Misunithyris finds its closer counterpart within the multicostate zeilleriids in the coeval Menathyris Feldman, 2013, if we compare the internal and external diagnostic features of both genera. Menathyris was recorded in the early Ladinian from Negev (southern Israel), integrated into the Sephardic bioprovince (Feldman 2013).
Thus, a Sephardic affinity can be suggested for the new brachiopod fauna recorded in the Betic Range, since in addition to the possible link to the Israeli zeilleriids (Fig. 6), it has to be taken into account that, in the Ladinian, this bioprovince is typified by the low faunal diversity and high degree of endemicity developed in epicontinental seas (Page 1996), just as in the Betic case. This endemicity is not only applicable for the brachiopods but also for the nekto-planktonic biota affinities (e.g., Tozer 1981; Pérez-Valera 2015). The geographical/reproductive isolation assumed for the nekto-planktonic fauna would become even more evident in groups with short larval stages such as brachiopods.
Conversely to the Sephardic province, the Germanic and Tethyan bioprovinces are more prolific in brachiopod diversity in the entire Triassic (Dagys 1993), recording a variety of rhynchonellid and spiriferinid taxa and, in the Ladinian, even emerging new taxonomic groups (Dagys 1993) such as Pennospiriferinidae or Angustothyrididae. Significant blooms of Koninckinacea, Dielasmatoidea, Neoretziinae, Aulacothyropsidae, and Vectellinae also occurred in this timespan in both bioprovinces (e.g., Bittner 1890, 1892; Siblík 1972, 1998; Dagys 1974; Popiel-Barczyk and Senkowiczowa 1989; Vörös and Pálfy 1989; Pálfy 1991, 2003; Pálfy and Török 1992; Senkowiczowa and Popiel-Barczyk 1993; Kaim 1997; Torti and Angiolini 1997; Hagdorn and Sandy 1998; Benatov 2001; Klug et al. 2005; Ruban 2010).
As with ammonites, the biogeographic boundaries of Sephardic bioprovince in the Ladinian are unclear toward the North of the Betic Range (Perez-Valera 2016). In the closest peri-Iberian epicontinental platforms, Middle Triassic brachiopods are only represented in the Levantine Sector of the Iberian Range through the monospecific pervasive Lingularia bursts (Márquez-Aliaga et al. 1999, 2007; Escudero-Mozo et al. 2015; Giannetti et al. 2017), and very few specimens attributed to Coenothyris (Schmidt 1935; Márquez-Aliaga et al. 1994; Escudero-Mozo et al. 2015) all of them mainly Ladinian in age, plus the Anisian Mentzelia from Menorca and the Catalonian Coastal Range (Schmidt 1935; Escudero-Mozo et al. 2015).
Consequently, it can be inferred that influences from the epicontinental seas of the Germanic or Tethyan provinces could occasionally have taken place in the Northern peri-Iberian platforms by means the immigrants Coenothyris and Mentzelia (Fig. 6). Nevertheless, both Coenothyris and Mentzelia can be regarded as ubiquitous taxa since, even they are more common in the Alpine Muschelkalk facies, they are wide-world distributed in basins such as Northern Caucasus (Ruban 2010), Hungary (Török 1993; Pálfy 2003); Southern Alps of NW-Italy (Torti and Angiolini 1997); Northern Alps and Slovakia (Siblík 1972, 1988, 2001), Poland (Popiel-Barczyk and Senkowiczowa 1989; Senkowiczowa and Popiel-Barczyk 1993; Kaim 1997; Szulc 2000), and even in the Sephardic bioprovince from Israel (Feldman 2005). In this sense, it has to be taken into account that Feldman (2005) separated the endemic Coenothyris oweni from those of the Germanic bioprovince. For this reason, the occurrence of both genera (Coenothyris and Mentzelia) in the peri-Iberian platform system is not fully indicative to establish the boundary of the Sephardic vs. Tethyan/Germanic influences (Fig. 6).
Finally, it is substantiated that the multicostate zeilleriids counterparts of Misunithyris exceeded the end-Triassic extinction event. In the Early Jurassic the genera Tauromenia and Calpella continued inhabiting the wide and shallow platforms developed in low-latitude seas of the Western Tethys as deduced from their record in the High/Middle Atlas (Dubar 1942), Western Algeria (Elmi et al. 2003; Alméras et al. 2007), Eastern Sicily and Calabria (Di Stefano 1887; Taddei-Ruggiero and Vörös 1987), Apennines (Pozza 1989), Gibraltar (Owen and Rose 1997), Cephalonia (Manceñido 1993a) and Eastern Subbetic (Baeza-Carratalá and García Joral 2012). These occurrences are mostly uppermost Sinemurian–lowermost Pliensbachian in age. The exceptional record of Tauromenia from the South Armorican Massif in the Uptonia jamesoni Zone is slightly younger (Alméras et al. 2010a). On the other hand, the genus Fimbriothyris, besides its Mediterranean distribution, shows a more profuse presence in the NW-European platforms from the late Pliensbachian onwards (Deslongchamps 1856; Alméras 1987).
In fact, this restricted geographical distribution makes this group a useful paleobiogeographical tool. In the Early Jurassic, multicostate zeilleriids distribution reveals an E-W dispersion pattern from the intra-Tethyan basins to the Atlantic areas (Ager and Walley 1977; Baeza-Carratalá and García Joral 2012). Thus, it can be emphasized that multicostate zeilleriids were not recorded in the NW-European basins during Triassic and earliest Jurassic times (except for the youngest South Armorican Massif record, Alméras et al. 2010a, which fit well in the E-W route). They disappeared from the Mediterranean Betic Ranges when the epicontinental/epioceanic duality became more evident from the early Pliensbachian onwards. The ulterior Middle Jurassic diversification of these forms and their migration to the NW-European platforms typified by the fully costate Eudesia or Flabellothyris, could have taken place concurrently with the well-documented general spreading of Mediterranean brachiopod fauna to the NW-European epicontinental seas. This migration was early detected by several Middle Jurassic incursions and chiefly persisted in the Late Jurassic (cf. Vörös 1988, 2005; Manceñido 1993b).
Suggestions on the origin of multicostate zeilleriids.—The uncommon occurrence of ribbed forms amidst zeilleriids resulted in a controversy as for the origin of multicostate zeillerids. Alméras (1987) put forward a comprehensive synthesis on this subject, postulating feasible relationships between the fully costate genera Eudesia King, 1850 and Flabellothyris, Deslongchamps, 1884, widespread in the Western Tethys during the Bajocian–Callovian interval, considering the Lower Jurassic Fimbriothyris Deslongchamps, 1884 (including some species currently assigned to Tauromenia Seguenza, 1885) as the possible ancestor of this group.
In fact, a polyphyletic origin for the Middle Jurassic ribbed forms was hypothesized (Alméras 1987; Alméras and Fauré 2008). These authors explained the diversification and evolutionary trends of this small clade in different biochoremas. The diversification took place on the one hand in the NW-European bioprovince, and on the other in the Indo-Arabian and North-African basins, all of them regarding as star-point the genus Fimbriothyris. Subsequent studies (Mukherjee et al. 2000) invoked a rapid migration to explain the distribution of the Middle Jurassic forms, better than a polyphyletic origin of Eudesia from Flabellothyris.
Regardless their mono-versus polyphyletic origin, most of the authors (except for Delance 1974) seem to agree in Fimbriothyris as possible ancestor or, at least, as the possible Lower Jurassic counterpart of this clade (Ager and Walley 1977; Alméras 1987; Mukherjee et al. 2000; Alméras and Fauré 2008). An entirely ribbed and bifurcate extreme form was depicted by Deslongchamps (1863) within the intraspecific variability of the genus Fimbriothyris, which was postulated as possible predecessor of this group (Alméras 1987; Alméras and Fauré 2008).
The new genus herein erected shares significant internal and external features such as the ribbing pattern, cardinal process, or dental plates structure with this possible evolutionary lineage (Table 2). Taking into account the above paleobiogeographical analysis (Fig. 6), from an anagenetic perspective it can be suggested that the Triassic stock made up by Misunithyris gen. nov. (probably together with Menathyris Feldman, 2013) might represent the precursor linkage taxon of this lineage which had its initial homeland in the low-latitude Tethyan basins during Triassic, such as the Sephardic bioprovince.
Table 2. Comparison of main attributes on Misunithyris gen. nov. with those of the Early–Middle Jurassic multicostate zeilleriid representatives.
|
Taxon |
Diagnostic features |
Stratigraphic distribution |
||||
|
Ribbing pattern |
Dental plates |
Cardinal process |
Dorsal median septum |
Brachidium |
||
|
Misunithyris |
anterior; radiate disposition; bifurcate; (often) intercalate |
thick-shelled, short, enveloped, subparallel |
present |
well-developed and long |
long-looped |
Ladinian |
|
Tauromenia |
anterior; radiate disposition; simple; (rare) bifurcate |
slender, short; ventrally divergent |
absent |
long, with large septalium |
long-looped |
(Rhaetian?) Sinemurian–Lower Pliensbachian |
|
Calpella |
anterior; radiate disposition; simple |
present, unknown orientation |
unknown |
prominent and long |
unknown |
Sinemurian |
|
Fimbriothyris |
anterior; rare on lateral slopes; simple |
slender, long; subparallel |
absent |
relatively short, blade-like |
poorly known |
upper Sinemurian–upper Pliensbachian |
|
Eudesia |
fully costae shell; radiate disposition; (often) bifurcate |
enveloped in callus, large, arcuate |
present |
moderately long and high |
long-looped |
upper Bajocian– |
|
Flabellothyris |
fully costae shell; radiate disposition; (not rare) bifurcate and intercalate |
slender, short; ventrally divergent |
absent |
short |
long-looped |
upper Bajocian–Callovian |
Conclusions
We report first record of brachiopod fauna in the Triassic from the Betic Range, as well as the oldest occurrence amidst Mesozoic brachiopod assemblages in this domain. The new finding is assigned to a new genus and species (Misunithyris goyi), which reveals a very characteristic external and internal structure, emphasizing the distinctive dental lamellas, a prominent cardinal process and its ribbing pattern. These features together with the septal development and the brachidium architecture make feasible its arrangement within the superfamily Zeillerioidea, evidencing high similarity with the multicostate zeilleriids stock. The most plausible supra-generic position is found within family Zeilleriidae, although several features are also shared with several zeilleriid families, not discarding the erection of a new subfamily in a future including other taxa with similar structure.
The occurrence of Misunithyris goyi in the Betic Range has been accurately established in the Gevanites epigonus Biochronozone of the Fassanian (lower Ladinian) and it can be used therefore as a valuable correlation and calibration tool.
A Sephardic paleobiogeographic affinity is suggested, since this fauna is so far endemic to the Betic Range and finds its possible counterparts within the multicostate zeilleriids integrated into the epicontinental platforms of the Sephardic bioprovince.
From an anagenetic perspective, it is suggested that this new genus might represent the precursor stock of the Early–Middle Jurassic multicostate zeillerids, probably originated in the low-latitude Tethyan basins (such as the Sephardic bioprovince) during the Triassic.
Acknowledgements
We appreciate the insightful comments and suggestions of the reviewers Maurizio Gaetani (State University of Milan, Italy) and Howard R. Feldman (American Museum of Natural History, New York, USA). We gratefully thank Milos Siblík (Czech Academy of Sciences, Prague, Czech Republic), Attila Vörös (Hungarian Natural History Museum, Budapest, Hungary) and Fernando García Joral (Complutense University of Madrid, Spain) for their valuable advices and enthusiastic help with literature. This research was supported by projects CGL2015-66604-R (MINECO, Government of Spain), the Research Groups VIGROB-167 (University of Alicante), and RNM 325 (Junta de Andalucía).
References
Ager, D.V. 1965. The adaptation of Mesozoic brachiopods to different environments: Palaeogeography, Palaeoclimatology, Palaeoecology 1: 143–172. Crossref
Ager, D.V. and Walley, C.D. 1977. Mesozoic brachiopod migrations and the opening of the North Atlantic. Palaeogeography, Palaeoclimatology, Palaeoecology 21: 85–99. Crossref
Alméras, Y. 1987. Origin and evolution of the Jurassic Multicostate Zeileriid Brachiopods Eudesia King and Flabellothyris Deslongchamps. Palaeogeography, Paleoclimatology, Palaeoecology 59: 261–268. Crossref
Alméras, Y. and Fauré, P. 2008. Le genre Flabellothyris Deslongchamps, 1884 Ses différentes espèces et leur variabilité morphologique [Brachiopoda, Zeilleriidae]. Bulletin de la Société d’histoire naturelle de Toulouse 144: 51–71.
Alméras, Y., Becaud, M., and Cougnon, M. 2010a. Brachiopodes liasiques de la bordure sud du Massif Armoricain. Bulletin de la Société des Sciences Naturelles de l’Ouest de la France 1 (Supplément hors-série): 1–131.
Alméras Y., Cougnon, R., Énay, R., and Mangold, C. 2010b. Brachiopodes du Jurassique Inférieur et Moyen d’Arabie Saoudite Centrale. Paléontologie, Biostratigraphie et Paléoenvironnements, échelles chronostratigraphiques. Documents des Laboratoires de Géologie Lyon 168: 1–247.
Alméras, Y., Elmi, S., and Fauré, P. 2007. Les Brachiopodes Liasiques d’Algérie Occidentale. Documents des Laboratoires de Géologie Lyon 163: 3–241.
Baeza-Carratalá, J.F. 2011. New Early Jurassic brachiopods from the Western Tethys (Eastern Subbetic, Spain) and their systematic and paleobiogeographic affinities. Geobios 44: 345–360. Crossref
Baeza-Carratalá, J.F. 2013. Diversity patterns of Early Jurassic brachiopod assemblages from the westernmost Tethys (Eastern Subbetic): Palaeogeography, Palaeoclimatology, Palaeoecology 381–382: 76–91. Crossref
Baeza-Carratalá, J.F. and García Joral, F. 2012. Multicostate zeillerids (Brachiopoda, Terebratulida) from the Lower Jurassic of the Eastern Subbetic (SE Spain) and their use in correlation and paleobiogeography. Geologica Acta 10: 1–12.
Baeza-Carratalá, J.F. and García Joral, F. 2014. Crural bases position as a structural criterion for supraspecific diagnosis of Early Jurassic zeilleriid brachiopods. Acta Palaeontologica Polonica 59: 651–661.
Baker, P.G. 2006. Zeillerioidea. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology. Part H. Brachiopoda. Revised, 5: Rhynchonelliformea (part), 2163–2188. Geological Society of America, Boulder and University of Kansas, Lawrence.
Balanyá, J.C., Crespo-Blanc, A., Díaz-Aspiroz, M., Expósito, I., and Luján, M. 2007. Structural trend line pattern and strain partitioning around the Gibraltar Arc accretionary wedge: Insights as to the mode of orogenic arc building. Tectonics 26: 1–19. Crossref
Balini, M., Lucas, S.G., Jenks, J.F., and Spielmann, J.A. 2010. Triassic ammonoid biostratigraphy: An overview. Geological Society, London, Special Publications 334: 221–262. Crossref
Benatov, S. 2001. Brachiopod biostratigraphy of the Middle Triassic in western Bulgaria and comparison with elsewhere in Europe. Systematics Association, Special Volume Series 63: 384–393.
Bittner, A. 1890. Brachiopoden der alpinen Trias. Abhandlungen der kaiserlich-königlichen geologischen Reichsanstalt 14: 1–325.
Bittner, A. 1892. Brachiopoden der alpinen Trias. Nachtrag 1. Abhandlungen der kaiserlich-königlichen geologischen Reichsanstalt 17: 1–40.
Chen, J., Tong, J., Song, H., Luo, M., Huang, Y., and Xiang, H. 2015. Recovery pattern after the Permian/Triassic crisis in South China. Palaeogeography, Palaeoclimatology, Palaeoecology 433: 91–105. Crossref
Chen, Z.Q., Kaiho, K., and George, A.D. 2005. Early Triassic Recovery of the Brachiopod Faunas from the End-Permian Mass Extinction: A Global Review. Palaeogeography, Palaeoclimatology, Palaeoecology 224: 232–269. Crossref
Cooper, G.A. 1983. The Terebratulacea (Brachiopoda), Triassic to Recent: A study of the Brachidia (Loops). Smithsonian Contributions to Paleobiology 50: 1–445. Crossref
Cooper, G.A. 1989. Jurassic brachiopods of Saudi Arabia. Smithsonian Contributions to Paleobiology 65: 1–213. Crossref
Dagys, A.S. 1974. Triasovye brahiopodi (morfologiâ, sistema, filogeniâ, stratigrafičeskoe značenie i biogeografiâ). 387 pp. Izdatel’stvo Nauka (Sibirskoe Otdelenie), Novosibirsk.
Dagys, A.S. 1993. Geographic differentiation of Triassic brachiopods. Palaeogeography, Palaeoclimatology, Palaeoecology 100: 79–87. Crossref
Delance, J.H. 1974. Zeilleridés du Lias d’Europe Occidentale (Brachiopodes). Université de Dijon, Mémoires de Géologie 2: 1–406.
Deslongchamps, E.E. 1863. Paléontologie Française ou description des animaux invertébrés Fossiles de la France. Terrain Jurassique, 1. Brachiopodes. 448 pp. Masson and Fils, Paris.
Deslongchamps, E.E. 1856. Notes sur deux nouvelles Térébratules du Lias moyen de Précigné (Sarthe). Mémoires Société Linnéenne de Normandie 10: 302–305.
Di Stefano, G. 1887. Sul Lias inferiore di Taormina e de’ suoi dintorni. Giornale della Societa di Scienze Naturale ed Economiche di Palermo 18: 46–184.
Dubar, G. 1942. Études paléontologiques sur le Lias du Maroc. Brachiopodes. Térébratules et Zeilléries multiplisées. Notes et Mémoires du Service Géologique du Maroc 57: 25–53.
Dulai, A. 2001. Diversification of Mediterranean Early Jurassic brachiopods after the end-Triassic mass extinction—new results from Hungary. In: C.H.C. Brunton, L.R.M Cocks, and S.L. Long (eds.), Brachiopods: Past and Present. The Systematic Association, Special Volume Series 63: 411–423. Crossref
Elmi, S., Alméras, Y., Benhamou, M., Mekahli, L., and Marok, A. 2003. Biostratigraphie des brachiopodes et âge Carixien (Pliensbachien inférieur) des calcaries à grands bivalves en Algérie occidentale. Geóbios 36: 695–706. Crossref
Escudero-Mozo, M.J., Márquez-Aliaga, A., Goy, A., Martín-Chivelet, J., López-Gómez, J., Márquez, L., Arche, A., Plasencia, P., Pla, C., Marzo, M., and Sánchez-Fernández, D. 2015. Middle Triassic carbonate platforms in the eastern Iberia: Evolution of their fauna and palaeogeographic significance in the western Tethys. Palaeogeography, Palaeoclimatology, Palaeoecology 417: 236–260. Crossref
Feldman, H.R. 2005. Paleoecology, taphonomy, and biogeography of a Coenothyris community (Brachiopoda, Terebratulida) from the Triassic (Upper Anisian–Lower Ladinian) of Israel. American Museum Novitates 3479: 1–19. Crossref
Feldman, H.R. 2013. Menathyris wilsoni (Brachiopoda), new genus and species from the Middle Triassic (Ladinian) of Southern Israel. Annales Societatis Geologorum Poloniae 83: 81–85.
Gaetani, M. 2016. Brachiopods from the type-section of the Bithynian substage (Anisian, Middle Triassic, Northwestern Turkey). Rivista Italiana Paleontologia Stratigrafia 122: 61–76.
García-Dueñas, V., Balanyá, J.C., and Martínez-Martínez, J.M. 1992. Miocene extensional detachments in the outcropping basement of the northern Alboran Basin (Betics) and their tectonic implications. Geo-Marine Letters 12: 88–95. Crossref
García-Hernández, M., López-Garrido A.C., Rivas, P., Sanz de Galdeano, C., and Vera, J.A. 1980. Mesozoic palaeogeographic evolution of the External Zones of the Betic Cordillera. Geologie in Mijnbouw 59: 155–168.
Giannetti, A., Tent-Manclús, J.E., and Baeza-Carratalá, J.F. 2017. New evidence of nearshore Mid-Triassic Zoophycos: morphological and palaeoenvironmental characterization. Facies 63: 1–12. Crossref
Gould, S.J. and Calloway, C.B. 1980. Clams and brachiopods-ships that pass in the night. Paleobiology 6: 383–396. Crossref
Hagdorn, H. and Sandy, M.R. 1998. Color banding in the Triassic terebratulid brachiopod Coenothyris from the Muschelkalk of Central Europe. Journal of Paleontology 72: 11–28. Crossref
Hallam, A. and Wignall, P.B. 1997. Mass Extinction and their Aftermath. 302 pp. Oxford University Press, Oxford.
Hirsch, F. 1987. Bio-stratigraphy and correlation of the marine Triassic of the Sephardic Province. Cuadernos de Geología Ibérica 11: 815–826.
Jiménez de Cisneros, D. 1923. La Fauna de los estratos de “Pygope Aspasia” Menegh. del Liásico Medio del Rincón de Egea en el W de la provincia de Murcia: Junta para Ampliación de Estudios e Investigaciones Científicas. Trabajos del Museo Nacional de Ciencias Naturales. Serie Geológica 30: 1–55.
Kaesler, R.L. and Selden, P.A. (eds.) 1997–2007. Treatise on Invertebrate Paleontology. Part H, Brachiopoda, Revised, vols. 1–6. Geological Society of America, Boulder and University of Kansas, Lawrence.
Kaim, A. 1997. Brachiopod-bivalve assemblages of the Middle Triassic Terebratula Beds, Upper Silesia, Poland. Acta Palaeontologica Polonica 42: 333–359.
Ke, Y., Shen, S.-Z., Shi, G.R., Fan, J.-X., Zhang, H., Qiao, L., and Zeng, Y. 2016. Global brachiopod paleobiogeographical evolution from Changshingian (Late Permian) to Rhaetian (Late Triassic). Palaeogeography, Palaeoclimatology, Palaeoecology 448: 4–25. Crossref
Klug, C., Schatz, W., Korn, D., and Reisdorf, A.G. 2005. Morphological fluctuations of ammonoid assemblages from the Muschelkalk (Middle Triassic) of the Germanic Basin. Indicators of their ecology, extinctions, and immigrations. Palaeogeography, Palaeoclimatology, Palaeoecology 221: 7–34. Crossref
Laborda-López, C., Aguirre, J., Donovan, S.K., Navas-Parejo, P., and Rodríguez, S. 2015. Fossil assemblages and biostratigraphy of metamorphic rocks of the Nevado-Filábride Complex from the Águilas tectonic arc (SE Spain). Spanish Journal of Palaeontology 30: 275–292.
Lonergan, L. and White, N. 1997. Origin of the Betic-Rif mountain belt. Tectonics 16: 504–522. Crossref
Manceñido, M.O. 1993a. Early Jurassic Brachiopods from Greece: a review. In: J. Pálfy and A. Vörös (eds.), Mesozoic Brachiopods of Alpine Europe, 79–100. Hungarian Geological Society, Budapest.
Manceñido, M.O. 1993b: First record of Jurassic nucleatid brachiopods from the southwest Pacific with comments on the global distribution of the group. Palaeogeography, Palaeoclimatology, Palaeoecology 100: 189–207. Crossref
Manceñido, M.O. 2002. Paleobiogeography of Mesozoic brachiopod faunas from Andean–Patagonian areas in a global context. Geobios 35: 176–192. Crossref
Martín-Algarra, A. and Vera, J.A. 2004. La Cordillera Bética y las Baleares en el contexto del Mediterráneo Occidental. In: J.A. Vera (ed.), Geología de España, 352–354. SGE, IGME, Madrid.
Márquez-Aliaga, A., Arche, A., Brito, J.M., Calvet, F., Goy, A., López-Gómez, J., and Márquez, L. 1994. El Muschelkalk en el Anticlinal de Calanda (Teruel, España): Estratigrafía y contenido paleontológico. Boletín Real Sociedad Española de Historia Natural (Sección Geológica) 89: 145–166.
Márquez-Aliaga, A., Emig, C.C., and Brito, J.M. 1999. Triassic Lingulide brachiopods from the Iberian Range (Spain). Geobios 32 (6): 815–821. Crossref
Márquez-Aliaga, A., Emig, C.C., and López-Gómez, J. 2007. Triassic Lingularia (Brachiopoda) from Moya (SE Iberian Ranges, Spain). Resúmenes 23 Jornadas de Paleontología, Caravaca de la Cruz: 121–122.
Márquez-Aliaga, A., Goy, A., and Gómez, J.J. 2001. Bivalvos del Ladiniense en la región de Riópar (Albacete, España). Publicaciones del seminario de Paleontología de Zaragoza 5: 144–152.
Mukherjee, D., Bardhan, S., and Ghosh, D.N. 2000. Evolution and migration of zeillerid brachiopods: A new record of Eudesia King from the Middle Jurassic of Kutch, India. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 215: 347–364. Crossref
Owen, E.F. and Rose, E.P.F. 1997. Early Jurassic Brachiopods from Gibraltar, and their Tethyan affinities. Palaeontology 40: 497–513.
Page, K.N. 1996. Mesozoic ammonoids in space and time. Triassic Provincialism. In: N. Landman, K. Tanabe, and R.A. Davis (eds.), Ammonoid Paleobiology. Topics in Geobiology, 770–775. Springer, New York. Crossref
Pálfy, J. 1991. Paleoecological significance of Anisian (Middle Triassic) brachiopod assemblages from the Balaton Highland (Hungary). In: D.I Mackinnon, D.E. Lee, and J. Campbell (eds.), Brachiopods Through Time, 241–246. Balkeema, Rotterdam.
Pálfy, J. 2003. The Pelsonian brachiopod fauna of the Balaton Highland. In: A. Vörös (ed.), The Pelsonian Substage on the Balaton Highland (Middle Triassic, Hungary). Geologica Hungarica Serie Palaeontologica 55: 139–158.
Pálfy, J. and Török, A. 1992. Comparison of Alpine and Germanotype Middle Triassic brachiopod faunas from Hungary, with remarks on Coenothyris vulgaris (Schlotheim, 1820). Annales Universitas Scientarium Budapestinensis Rolando Eötvös Nominatae, Sectio Geologica 29: 303–323.
Parnes, A., Benjamini, C., and Hirsch, F. 1985. Some aspects of Triassic ammonite biostratigraphy, paleoenvironments and paleobiogeography in southern Israel. Journal of Paleontology 59: 656–666.
Payne, J.L., Lehrmann, D.J., Wei, J., Orchard, M.J., Schrag, D.P., and Knoll, A.H. 2004. Large perturbations of the carbon cycle during recovery from the end-Permian extinction. Science 305: 506–509. Crossref
Pérez-López, A. 1998. Epicontinental Triassic of the Southern Iberian Continental Margin (Betic Cordillera, Spain). In: G.H. Bachmann and I. Lerche (eds.), Epicontinental Triassic. Zentralblatt für Geologie und Paläontologie 1: 1009-1031.
Pérez-López, A. and Pérez-Valera, F. 2007. Palaeogeography, facies and nomenclature of the Triassic units in the different domains of the Betic Cordillera (S Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 254: 606–626. Crossref
Pérez-López, A., López-Garrido, A.C., Márquez-Aliaga, A., De Galdeano, C.S., and García-Tortosa, F.J. 2003. Ladinian carbonates of the Cabo Cope Unit (Betic Cordillera, SE Spain): a tethys-maláguide palaeogeographic gateway. Facies 48: 1–7. Crossref
Pérez-López, A., Márquez, L., and Pérez-Valera, F. 2005. A foraminiferal assemblage as a biovevent marker of the main Ladinian transgressive stage in the Betic Cordillera, Southern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 224: 217–231. Crossref
Pérez-Valera, F. 2005. Estratigrafía y tectónica del Triásico Sudibérico en el sector oriental de la Cordillera Bética. 301 pp. Unpublished Ph.D. Thesis, Universidad de Granada, Granada.
Pérez-Valera, F. and Pérez-López, A. 2007. Stratigraphy and sedimentology of Muschelkalk carbonates of the Southern Iberian Continental Palaeomargin (Siles and Cehegín Formations, Southern Spain). Facies 54: 61–87. Crossref
Pérez-Valera, F. and Pérez-López, A. 2008. Stratigraphy and sedimentology of Muschelkalk carbonates of the Southern Iberian Continental Palaeomargin (Siles and Cehegín Formations, Southern Spain). Facies 54: 61–87. Crossref
Pérez-Valera, J.A. 2005. Ammonoideos y bioestratigrafía del Triásico Medio (Anisiense superior-Ladiniense) en la sección de Calasparra (sector oriental de la Cordillera Bética, Murcia, España). Coloquios de Paleontología 55: 125–161.
Pérez-Valera, J.A. 2015. Ammonoideos y bioestratigrafía del Triásico medio (Ladiniense) del sector oriental de la Cordillera Bética. 489 pp. Unpublished Ph.D. Thesis, Universidad Complutense de Madrid, Madrid.
Pérez-Valera, J.A., Barroso Barcenilla, F., Goy, A., and Pérez-Valera, F. 2017. Nautiloids from the Muschekalk facies of the Southiberian Triassic (southern Spain). Journal of Systematic Palaeontology 15: 171–191. Crossref
Pérez-Valera, J.A., Goy, A., Pérez-Valera, F., and Pérez-López, A. 2011. Ammonoid biostratigraphy of Muschelkalk carbonates (Ladinian) ofthe South-Iberian Triassic (Betic External Zones, S. Spain). In: A. Maate (ed.), Livret des Résumés du Sixième Réunion du Groupe Marocain du Permien et du Trias, Tétouan, 20–21 May, 2011, 23–24. University of Tétouan, Tétouan.
Plasencia, P., Márquez-Aliaga, A., Pérez-Valera, F., Pérez-Valera, J.A., Goy, A., López-Gómez, J., Márquez, L., and Pérez-López, A. 2007. Registro paleontológico del Triásico Medio de Calasparra (Murcia). In: J.C. Braga, A. Checa, and M. Company (eds.), Libro de Resúmenes de las XXIII Jornadas Sociedad Española de Paleontología, 187–188. Copicentro Granada S.L., Granada.
Platt, J., Allerton, S., Kirker, A., and Platzman, E. 1995. Origin of the western Subbetic arc (South Spain): palaeomagnetic and structural evidence. Journal of Structural Geology 17: 765–775. Crossref
Popiel-Barczyk, E. and Senkowiczowa, H. 1989. Representatives of the genus Coenothyris Douville, 1879 from the Terebratula Bed (Upper Muschelkalk) of the Holy Cross Mts, Central Poland. Acta Geologica Polonica 39: 93–111.
Pozza, G.C. 1989. Spiriferina del Lias inferiore di Cingoli (Marche, Italia). Bolletino della Società Paleontologica Italiana 28: 49–62.
Raup, D.M. and Sepkoski, J.J. 1982. Mass extinctions in the marine fossil record. Science 215: 1501–1503. Crossref
Ruban, D.A. 2006a. Diversity changes of the Brachiopods in the Northern Caucasus: a brief overview. Acta Geologica Hungarica 49: 57–71. Crossref
Ruban, D.A. 2006b. Diversity dynamics of the Triassic marine biota in the Western Caucasus (Russia): a quantitative estimation and a comparison with the global patterns. Revue de Paleobiologie 25: 699–708.
Ruban, D.A. 2010. The Permian/Triassic mass extinction among brachiopods in the Northern Caucasus (northern Palaeo-Tethys): A tentative assessment. Geobios 43: 355–363. Crossref
Ruban, D.A. 2017. Examining the Ladinian crisis in light of the current knowledge of the Triassic biodiversity changes. Gondwana Research 48: 285–291. Crossref
Sandy, M. 1995. Early Mesozoic (Late Triassic–Early Jurassic) Tethyan brachiopod biofacies: Possible evolutionary intra-phylum niche replacement within the Brachiopoda. Paleobiology 21: 479–495. Crossref
Schmidt, M. 1935. Fossilien der Spanischen trias. Abhängungen Heidelberg Akademie Wissenschaften Mathematische Naturkunde 22: 1–140.
Seguenza, G. 1885. Monografia delle Spiriferina dei varii piani del Lias Messinese. Bollettino della Società Geologica Italiana 4: 377–497.
Senkowiczowa, H. and Popiel-Barczyk, E. 1993. Brachiopods from Terebratula Bed from the Upper Muschelkalk in the Grotow P-ll borehole (SW Poland). Geological Quarterly 37: 579–592.
Senkowiczowa, H. and Popiel-Barczyk, E. 1996. Some Terebratulida (Brachiopoda) from the Muschelkalk sediments in the Holy Cross Mts. Geological Quarterly 40: 443–466.
Sepkoski, J.J. Jr. 1996. Competition in macroevolution. In: D. Jablonski, J. Erwin, and J.H. Lipps (eds.), Evolutionary Paleobiology, 211–255. University of Chicago Press, Chicago.
Shen, S.-Z., Zhang, H., Li, W.-Z., Mu. L., and Xie, J.-F. 2006. Brachiopod diversity patterns from Carboniferous to Triassic in South China. Geological Journal 41: 345–361. Crossref
Siblík, M. 1972. Anisian Spiriferida and Terebratulida from the Slovak Karst Region. Geologické Práce Spravy 59: 179–202.
Siblík, M. 1988. Brachiopoda mesozoica. A. Brachiopoda triadica. Catalogus Fossilium Austriae. 145 pp. Verlag der Österreichischen Akademie der Wissenschaften, Wien.
Siblík, M. 1998. A contribution to the brachiopod fauna of the “Oberrhätkalk” (northern Calcareous Alps, Tyrol-Salzburg). Jahrbuch der geologischen Bundesanstalt 141: 73–95.
Siblík, M. 2001. Brachiopoda mesozoica. A. Brachiopoda triadica. Supplementum. Catalogus Fossilium Austriae. 45 pp. Verlag der Österreichischen Akademie der Wissenschaften, Wien.
Smirnova, T.N. 2008. Ontophylogenetic studies of the brachiopods of the Order Terebratulida. Paleontological Journal 42: 805–824. Crossref
Szulc, J. 2000. Middle Triassic evolution of the northern Peri-Tethys area as influenced by early opening of the Tethys Ocean. Annales Societatis Geologorum Poloniae 70: 1–48.
Taddei-Ruggiero, E. and Vörös, A. 1987. Paleobiogeographical evaluation of Calabrian Liassic brachiopods. Rendiconti della Società Geologica Italiana 9: 235–242.
Tomasových, A. 2006. A new Early Jurassic rhynchonellid brachiopod from the western Tethys and implications for systematics of rhynchonellids from the Triassic–Jurassic Boundary. Journal of Paleontology 80: 212–228. Crossref
Török, Á. 1993. Brachiopod beds as indicators of storm events: an example from the Muschelkalk of southern Hungary. In: J. Pálfy and A. Vörös (eds.), Mesozoic Brachiopods of Alpine Europe, 161–172. Hungarian Geological Society, Budapest.
Torti, V. and Angiolini, L. 1997. Middle Triassic brachiopods from Val Parina, Bergamasc Alps, Italy. Rivista Italiana di Paleontologia e Stratigrafia 103: 149–172.
Tozer, E.T. 1981. Triassic Ammonoidea: Geographic and Stratigraphic distribution. In: M.R House and J.R. Senior (eds.), The Ammonoidea. The Systematics Association, Special Volume 18: 397–431.
Vörös, A. 1988. Conclusions on Brachiopoda. In: M. Rakus, J. Dercourt, and A.E.M. Nairn (eds.), Evolution of the Northern Margin of the Tethys. Mémoires de la Société Géologique de France (N.S.) 154: 79–83.
Vörös, A. 2005. The smooth brachiopods of the Mediterranean Jurassic: refugees or invaders? Palaeogeography, Palaeoclimatology, Palaeoecology 223: 222–242. Crossref
Vörös, A. and Pálfy, J. 1989. The Anisian/Ladinian boundary in the Vászoly section (Balaton Highland, Hungary). Fragmenta Mineralogica et Palaeontologica 14: 17–27.
Zakharov, Y.D. and Popov, A.M. 2014. Recovery of brachiopod and ammonoid faunas following the end-Permian crisis: additional evidence from the Early Triassic of Russian Far East and Khazakhstan. Journal of Earth Sciences 25: 1–40. Crossref
Acta Palaeontol. Pol. 63 (1): 71–85, 2018
https://doi.org/10.4202/app.00415.2017