
Youngest record of the extinct walrus Ontocetus emmonsi from the Early Pleistocene of South Carolina and a review of North Atlantic walrus biochronology
SARAH J. BOESSENECKER, ROBERT W. BOESSENECKER, and JONATHAN H. GEISLER
Boessenecker, S.J., Boessenecker, R.W., and Geisler, J.H. 2018. Youngest record of the extinct walrus Ontocetus emmonsi from the Early Pleistocene of South Carolina and a review of North Atlantic walrus biochronology. Acta Palaeontologica Polonica 63 (2): 279–286.
The extinct North Atlantic walrus Ontocetus emmonsi is widely reported from Pliocene marine deposits in the eastern USA (New Jersey, Florida), Belgium, Netherlands, Great Britain, and Morocco. Ontocetus was slightly larger than the modern walrus Odobenus rosmarus, may have had wider climatic tolerances (subtropical), and likely originated in the western North Pacific before dispersing through the Arctic. Owing to geochronologic uncertainties in the North Atlantic Plio-Pleistocene walrus record, it is unclear whether Ontocetus and Odobenus overlapped in time and thus may have competed, or whether the two were temporally separate invasions of the North Atlantic. A new specimen of Ontocetus emmonsi (CCNHM-1144) from the Austin Sand Pit (Ridgeville, South Carolina, USA) is a complete, well-preserved left tusk that is proximally inflated and oval in cross-section, relatively short (maximum length: 369 mm) and markedly curved (radius of arc of curvature 197 mm). Globular dentine is present, confirming assignment to Odobenini; proportions and curvature identify the specimen as Ontocetus emmonsi rather than Odobenus. Hitherto unstudied deposits in the Austin Sand Pit lack calcareous macro and microinvertebrates, but vertebrate biochronology provides some temporal resolution. The co-occurrence of a giant beaver (Castoroides sp.) and a snaggletooth shark (Hemipristis serra) indicate an age of 1.1–1.8 Ma (Early Pleistocene) and correlation with the Lower Pleistocene Waccamaw Formation. The vertebrate assemblage is named the Ridgeville Local Fauna. The composition of the marine mammal assemblage from the Austin Sand Pit is intermediate between that of the lower Pliocene Yorktown Formation (North Carolina, USA) and the modern North Atlantic fauna. This record reported here is the youngest of Ontocetus emmonsi from the Atlantic Coastal Plain. A review of North Atlantic Plio-Pleistocene walrus records reveals no overlap between extinct Ontocetus and extant Odobenus—suggesting independent dispersal to the North Atlantic and a lack of competition.
Key words: Mammalia, Carnivora, Odobenidae, Ontocetus, Odobenus, Pliocene, Pleistocene, North Atlantic.
Sarah J. Boessenecker [michaliess@cofc.edu], Mace Brown Museum of Natural History, College of Charleston, Charleston, South Carolina, 29424 USA.; School of Museum Studies, University of Leicester, Leicester LE1 7RF, UK.
Robert W. Boessenecker [boesseneckerrw@cofc.edu] (corresponding author), Department of Geology and Environmental Geosciences, College of Charleston, Charleston, South Carolina, 29424 USA; University of California Museum of Paleontology, Berkeley, California, 94720 USA.
Jonathan H. Geisler [jgeisler@nyit.edu], Department of Anatomy, New York Institute of Technology, Old Westbury, New York 11568-8000 USA.
Received 3 January 2018, accepted 26 March 2018, available online 8 June 2018.
Copyright © 2018 S.J. Boessenecker et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The extinct Pliocene walrus Ontocetus emmonsi is a tusked walrus (family Odobenidae, tribe Odobenini) closely related to the extant walrus Odobenus rosmarus and the extinct eastern North Pacific toothless walrus Valenictus (Deméré 1994a; Kohno and Ray 2008). Unlike extant Odobenus rosmarus, fossils of Ontocetus are known from temperate and subtropical latitudes (Kohno and Ray 2008), similar to its close relative Valenictus (Deméré 1994b; Boessenecker 2017). Ontocetus was roughly 15% larger than modern walrus (Odobenus rosmarus, Deméré 1994a). Relative to Odobenus, the tusks of Ontocetus are generally more curved, proportionally shorter and proximally inflated with deeper lingual furrows. Ontocetus emmonsi is reported from the Eastern North Atlantic Coastal Plain (New Jersey through Florida), the North Sea (Belgium, Netherlands, Great Britain) and Morocco (Kohno and Ray 2008), indicating a circum North Atlantic distribution. Early studies assumed some form of ancestor-descendant relationship between Ontocetus and Odobenus, and an Atlantic origin for modern walruses, followed by a Pleistocene reinvasion of the north Pacific by the Pacific walrus subspecies Odobenus rosmarus divergens (Repenning and Tedford 1977; Repenning et al. 1979; Tomida 1989; Miyazaki et al. 1995). However, a new wealth of walrus fossils indicate a North Pacific center of origin for the Odobenidae (Deméré 1994a; Kohno et al. 1995a, b; Miyazaki et al. 1995), and a central American Seaway dispersal of walruses was revised by Kohno et al. (1995b) and Kohno and Ray (2008). Early Pliocene fossils from of Japan suggest a western North Pacific origin for Ontocetus, followed by a dispersal through the Arctic (Kohno et al. 1995b: 125, 1998). Similarly, fossils of Odobenus in Japan appear to predate those from the North Atlantic, suggesting a North Pacific center of origin for Odobenus as well (Kohno et al. 1995b). Several questions regarding Recent walrus evolution remain, including (i) when exactly did Ontocetus become extinct?; (ii) when did Ontocetus and Odobenus first arrive in the Atlantic?; and (iii) did Ontocetus and Odobenus overlap and compete with one another? We report a new specimen from the Austin Sand Pit in Ridgeville, South Carolina, USA and review the biochronology of the North Atlantic walrus fossil record as it pertains to these questions. Updated geochronology of walrus-bearing deposits on the Atlantic Coastal Plain invites a new hypothesis for walrus biogeography in the North Atlantic.
Institutional abbreviations.—CCNHM, Mace Brown Museum of Natural History, College of Charleston, USA; ChM PV, Charleston Museum (vertebrate paleontology collection), USA; SC, South Carolina State Museum, Columbia, USA.
Other abbreviations.—FAD, first appearance datum; LAD, last appearance datum; NALMA, North American Land Mammal Age.
Geological setting
The Austin Sand Pit is a now-inundated sandpit in Ridgeville, Dorchester County, South Carolina, USA prospected by amateur fossil collectors in the 1990s but never seriously studied by stratigraphers or paleontologists. The Austin deposit consists of a basal bluish-gray sandy, unconsolidated silt (~60 cm thick) with a basal bone and phosphate nodule lag overlying the lower unconformable contact with the Ashley Formation (early Oligocene; Weems et al. 2014); the silty unit is overlain by an unconsolidated tan-gray medium-coarse quartzose sand (Fig. 1). A stratigraphic section was never formally recorded for the original pit while open in the 1990s. A new sand pit located 100 m to the southeast, freshly dug in 2017 has a similar stratigraphic succession for the Austin deposit, permitting re-analysis of the unit. In the new pit, it consists of a basal bluish-white clay (50-cm thick; base not exposed) overlain by a richly fossiliferous granule-pebble conglomeratic sand (20–50 cm thick) with abundant phosphate nodules and with a sharp, irregular lower contact, overlain in turn by a fine-grained white sand (~1 m thick). The Austin deposit is overlain by 2–3 m of brown-purple clay and sandy interbeds of the Penholoway Formation (Middle Pleistocene; Weems et al. 2014).
The identity of this unit is unknown, though it overlies the Givhan’s Ferry Member of the Ashley Formation (fossiliferous fine-grained calcarenite) and is overlain by the Middle Pleistocene Penholoway Formation (back-barrier estuarine clay with basal quartzose sand and pebble lag; Fig. 1). The Ashley Formation is late early Oligocene in age and dated to 26.57–29.0 Ma based on Sr isotope ratios (Weems et al. 2016); locally the Penholoway Formation is approximately 700–900 Ka in age based on calcareous nannofossils and U-Th ratios (Middle Pleistocene; Weems et al. 1994, 2014). The Austin deposit lacks calcareous micro- and macrofossils, precluding precise dating. However, some biochronological control is afforded by vertebrate fossils. A diverse assemblage of 52 taxa is recorded (Table 1) including sharks, rays, bony fish, alligators, turtles, whales, birds, dolphins, land mammals, and the walrus Ontocetus; we propose the name Ridgeville Local Fauna for the assemblage. A maximum age is provided by two partial incisors of giant beavers (CCNHM 1924, CCNHM 1925) identifiable as Castoroides sp. owing to well-developed longitudinal grooves and ridges in the incisor enamel. Castoroides has a first appearance datum (FAD) of 1.8 Ma in the Bermont Formation at Leisey Shell Pits 1 and 3 in Florida (Morgan and Hulbert 1995; Bell et al. 2004; Morgan 2005), indicating correlation with the early Irvingtonian North American Land Mammal Age (NALMA). A minimum age is provided by teeth of the snaggletooth shark Hemipristis serra (CCNHM 1926, ChM PV 5132, ChM PV 5135, ChM PV 5136, ChM PV 5178), which is known from upper Oligocene through Pleistocene deposits of the Atlantic Coastal Plain (Cicimurri and Knight 2009; Scudder et al. 1995). The last appearance datum (LAD) of Hemipristis serra is 1.1 Ma, based on the minimum age of strata within the Leisey Shell Pit (pits 1A and 3B) of Florida, the youngest known occurrence for this species (Morgan and Hulbert 1995: 30; Scudder et al. 1995: 266–267). The FAD of Castoroides in concert with the LAD of Hemipristis serra constrains the age of the fossiliferous deposit in the Austin Sand Pit to 1.1–1.8 Ma (Early Pleistocene, Irvingtonian NALMA correlative). The deposit is too young to be mapped as either the Pliocene Raysor Formation (3.1–3.6 Ma; Tseng and Geisler 2016, and references therein) or the Goose Creek Limestone (3.1–3.6 Ma; Tseng and Geisler 2016, and references therein). Further, both the Raysor Formation and Goose Creek Limestone are highly calcareous and fossiliferous; this deposit entirely lacks calcareous microfossils and macroinvertebrates. The Austin deposit is best tentatively identified as the Plio-Pleistocene Waccamaw Formation, which is mapped nearby (Weems et al. 2014) and reported from auger holes 4.5 km to the northeast (= auger hole RI-6 of Weems et al. 1987, 1994). Further northeast, the Waccamaw Formation is preliminarily dated to earliest Pleistocene (1.55–2.8 Ma) based on 86Sr/87Sr ratios from exposures in southeastern North Carolina (MacGregor et al. 2011).
Another locality that may be related to the Austin Sand Pit is the Walrus Ditch Local Fauna (Fields et al. 2012), a locality 20 km southwest in Summerville, South Carolina, USA which has yielded a large vertebrate assemblage including Ontocetus emmonsi (Kohno and Ray 2008) and terrestrial mammals (Fields et al. 2012). Biochronologically significant land mammals include the horse Nannipus and the porcupine Erethizon, constraining the age to 2.6–2.2 Ma (Fields et al. 2012: 8); this led Fields et al. (2012) to identify the Walrus Ditch Local Fauna as occurring within the Waccamaw Formation. The deposit in the Austin Sand Pit is entirely younger than the Walrus Ditch deposit, and it is possible that the Walrus Ditch deposit may represent the lower Waccamaw and the Austin deposit may represent the upper Waccamaw. Further geologic and paleontologic studies of the newly excavated pit will permit further observations on the stratigraphy, taphonomy, and paleontology of the ?Waccamaw Formation.
Chronostratigraphic framework, subdivision of the Pleistocene, and the definition of the Pliocene–Pleistocene boundary follows Gradstein et al. (2012).
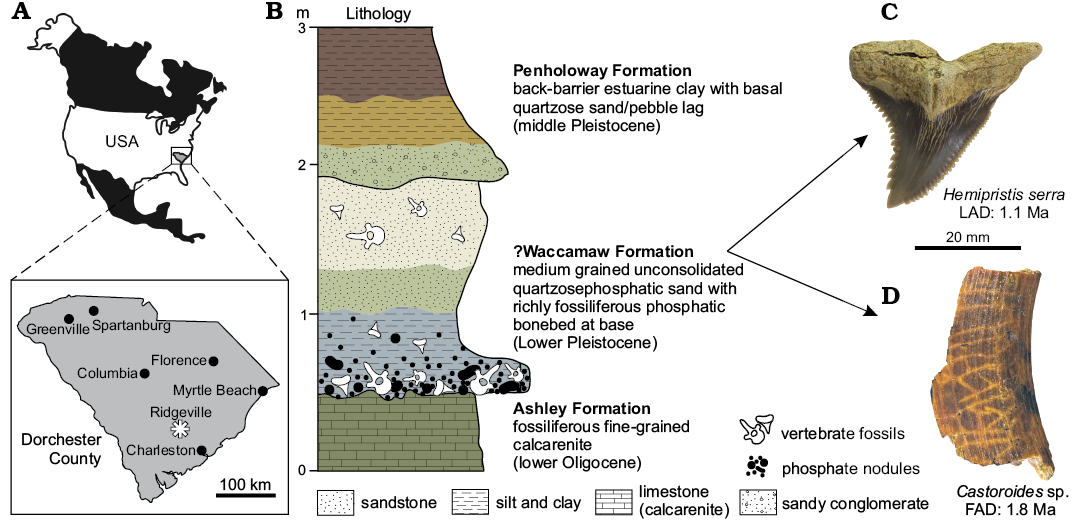
Fig. 1. A. Map showing the location of South Carolina and Austin Sand Pit near Ridgeville in Dorchester County (asterisked). B. Generalized stratigraphic column of the Austin Sand Pit with biostratigraphically significant vertebrate fossils from the ?Waccamaw Formation (C, D). Colors approximate sediments exposed in the Austin Sand Pit.
Systematic palaeontology
Mammalia Linnaeus, 1758
Carnivora Bowditch, 1821
Pinnipedia Illiger, 1811
Odobenidae Allen, 1880
Odobeninae Mitchell, 1968
Odobenini Deméré, 1994
Genus Ontocetus Leidy, 1858
Type species: Ontocetus emmonsi Leidy, 1859; Yorktown Formation, North Carolina, Pliocene.
Ontocetus emmonsi Leidy, 1858
Fig. 2.
Material.—CCNHM 1144, a complete left canine, collected by Matthew Swilp (Shamokin, Pennsylvania, USA) in the 1990s from the ?Waccamaw Formation exposed in the now-inundated Austin sand pit, Ridgeville, South Carolina, USA. A second tusk of Ontocetus emmonsi (SC 98.51.1) was referred by Kohno and Ray (2008) but not figured or described and is not considered further in this study.
Description.—CCNHM-1144 is a complete, well-preserved left canine (linear length 369 mm; anterior curvilinear length 345 mm; posterior curvilinear length 477 mm), which is markedly curved posteriorly (radius of arc of curvature 197 mm; Fig. 3). The tusk evenly tapers distally and bears a blunt tip with a posterolingual oval-shaped wear facet (35 mm long and 19 mm wide). Though expected, there is no obvious dentine exposed in the wear facet. The tusk is oval in cross-section and compressed transversely (transverse basal diameter 67 mm; anteroposterior basal diameter 90 mm). Two deep longitudinal furrows are present labially, the posterior furrow being slightly deeper and extending further distally; these furrows become diffuse distally, and terminate 54 mm from the apex. Though mostly curved in the parasagittal plane, the tusk is slightly twisted about its longitudinal axis from proximal to distal so that the tip is rotated and slightly reverted lingually. Longitudinal furrows are also present lingually, though they are diffuse and less deeply incised, terminating only 239 mm from the apex. The position of the gum line is unclear. However, subparallel anteroposterior scratches are present lingually but are absent from the proximal 150 mm of the tusk. If these were produced in vivo (e.g., sediment abrasion during benthic feeding) it would suggest the gum line was positioned at about 150 mm from the proximal end of the tusk. External annular growth rings are not visible under low angle light.
The pulp cavity is open and forms the widest part of the tusk, indicating continuous growth. The shape of the pulp cavity is oval, and the depth is 82 mm; the globular orthodentine core is developed within as a proximally tapering cone. Individual bead-like dentine globules are visible proximally within the pulp cavity, confirming assignment of CCNHM 1144 to Odobenini (Deméré 1994a).
The tusk lacks evidence of post-mortem abrasion; irregular, circular-oval crater-like pits (2–16 mm diameter, 0.7–1 mm deep) are evenly distributed proximo-distally along the tusk and several occur inside the pulp cavity. Pitting is more common on the labial side and often coalesced so that large areas are missing cementum. The margins of the pits are smooth and lack evidence of fracturing, resembling enamel cupping reported in dolphin teeth by Loch et al. (2013).
Remarks.—CCNHM 1144 is identifiable as Ontocetus emmonsi owing to its strong curvature, marked distal tapering, transverse compression, and deeply incised longitudinal furrows (Kohno and Ray 2008: 45). This specimen has a transverse width:anteroposterior diameter ratio of 0.74, which is relatively wide for Ontocetus emmonsi (Fig. 3). However, the deeply incised labial grooves and small radius of the arc of curvature (Fig. 3) clearly differentiate this specimen from Valenictus (Deméré 1994b) and Odobenus (Kohno and Ray 2008), and are characteristic features identifying this specimen as Ontocetus emmonsi.
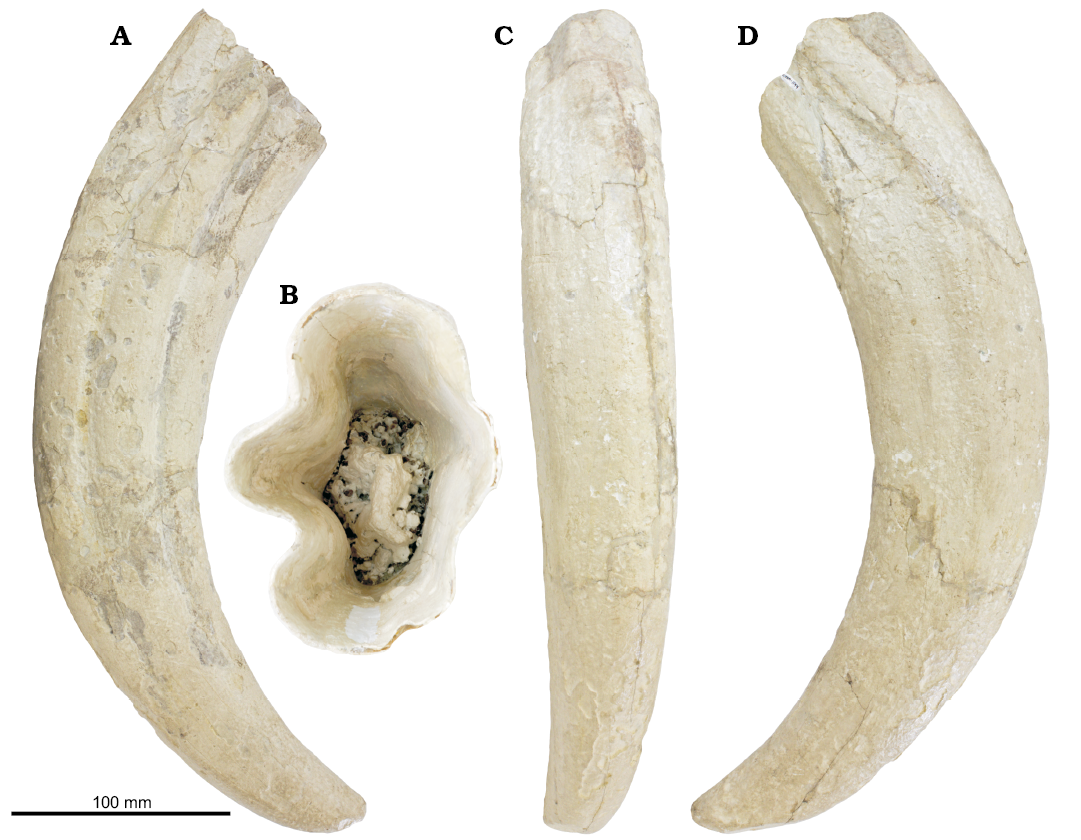
Fig. 2. Right canine tusk of odobenid walrus Ontocetus emmonsi (CCNHM 1144) from the Lower Pleistocene ?Waccamaw Formation, Austin Sand Pit, South Carolina, in lingual (A), proximal (B), anterior (C), and labial (D) views.

Fig. 3. Scatterplot of ratio of proximal transverse width/anteroposterior length versus radius of the arc of curvature for tusks of Odobenus and Ontocetus, including CCNHM 1144. Original plot modified from Kohno and Ray (2008: fig. 27).
North Atlantic walrus biochronology
A review of fossil occurrences and available stratigraphic literature was implemented to evaluate the exact chronology of walruses in the North Atlantic. This review emphasized specimens identifiable to Ontocetus or Odobenus and ignored specimens identifiable only to the subfamily Odobeninae or tribe Odobenini. In many cases, fossil occurrences were reported long before formal stratigraphic study commenced, were reported without provenance, or were specimens discovered ex situ and thus isolated from geological context (see discussions in Ray 1960, 1975; Kohno and Ray 2008). Rather than rely on outdated age data published in the original articles reporting walrus specimens, in all cases where specimens were reported with stratigraphic provenance, the most current literature on the stratigraphy and geochronology of these strata was used to compile geochronologic ranges for each occurrence (e.g., Parham et al. 2012: 350–352).
The majority of Ontocetus specimens from the Atlantic Coastal Plain were collected from the Yorktown Formation (lower Pliocene) of North Carolina and Virginia (Berry and Gregory 1906; Kohno and Ray 2008). The geochronologically earliest Ontocetus are reported from the Bone Valley Formation (Palmetto Fauna, Mio-Pliocene) in Florida Morgan (1994). The geochronologically latest appearance of Ontocetus is reported from the ?Waccamaw Formation at the Austin Sand Pit (Ridgeville Local Fauna), dating to 1.8–1.1 Ma (this study).
This new specimen and the biochronologic dating of the ?Waccamaw Formation, in concert with updated dates for the Palmetto Fauna, provides an updated maximum geochronologic range for Ontocetus emmonsi of 5.8–1.1 Ma. Our survey of published record of Odobenus failed to identify any reliable fossil records of the extant genus before the late Middle Pleistocene in the North Atlantic. An Odobenus-like mandible from the “Scaldisian sands” (= Lillo Formation) was referred to Odobenus koninckii by Van Beneden (1877), possibly representing a Pliocene occurrence of Odobenus in the Atlantic (Deméré, 1994a). However, recognition of a singular Pliocene Atlantic occurrence awaits redescription and stratigraphic reassessment of this specimen. All specimens with reliable stratigraphic provenance are a maximum of 0.24 Ma (Marine isotope stage 7), supporting the conclusion of Kohno and Ray (2008: 63) that no walruses are known from the Middle Pleistocene of the Atlantic basin. Despite the range extension for Ontocetus, there is a nearly 1 Ma gap between the Ontocetus LAD (1.1 Ma) and the Odobenus FAD (0.24 Ma) in the North Atlantic (Fig. 4).

Fig. 4. Biochronology of the walruses Ontocetus and Odobenus in the North Atlantic basin. Yellow vertical bar denotes temporal gap between the last occurrence of Ontocetus and the first occurrence of Odobenus in the North Atlantic. See SOM 1, 2 (Supplementary Online Material available at http://app.pan.pl/SOM/app63-Boessenecker_etal_SOM.pdf) for discussion of the geochronologic age of each occurrence. Abbreviations: E, early; L, late; M, middle.
Discussion
The marine mammal assemblage of the ?Waccamaw Formation at the Austin Sand Pit (Table 1) is notable in that the taxonomic composition is intermediate between the lower Pliocene Yorktown Formation (e.g., Ontocetus emmonsi, Kogiidae gen. nov., cf. Kogiopsis, Balaenula sp., Balaenoptera sp., “Balaenoptera” borealina, Herpetocetus sp.; Kohno and Ray 2008; Whitmore and Barnes 2008; Whitmore and Kaltenbach 2008) and the modern fauna (Eschrichtius sp., Megaptera sp.). Cf. Astadelphis sp. is preliminarily recorded from the Plio-Pleistocene ?Waccamaw Formation, a first for North America; Astadelphis has been previously reported only from the late Pliocene (Piacenzian) of Italy (Bianucci 1996). Additional petrosals of cf. Astadelphis (CCNHM 1908) have also been collected ex situ from Folly Beach, South Carolina, USA likely originating from the Pliocene Goose Creek Limestone. Further study of this chronologically unique marine mammal assemblage may shed light on the emergence of the modern marine mammal fauna (e.g., Whitmore 1994; Boessenecker 2013).

CCNHM 1144 is now the youngest record of Ontocetus worldwide. Despite the range extension of Ontocetus to the Early Pleistocene, the available fossil record indicates Ontocetus and Odobenus are separated by 0.9 Ma (Fig. 4). Similarly, in the western North Pacific Ontocetus and Odobenus are not contemporaneous; Ontocetus has a LAD of 3.6 Ma (Kohno et al. 1998; Kimura et al. 2007) and Odobenus has a FAD of 2.7 Ma (Kohno et al. 1995b) and are separated by approximately 1 my (Kohno et al. 1995b, 1998). We hypothesize that Odobenus and Ontocetus represent independent (early Pliocene, Late Pleistocene, respectively) dispersals of tusked walruses (Odobenini) into the North Atlantic from the North Pacific, and did not directly compete in either ocean basin according to present biochronologic evidence.
The FAD of Ontocetus in the Atlantic basin is from the Palmetto Fauna of the Bone Valley Formation (Morgan 1994; Kohno and Ray 2008). Land mammals from the Palmetto Fauna correspond with the latest Hemphillian (Hh4) NALMA (Wallace and Hulbert 2013), indicating an age of 5.8–4.6 Ma (Tedford et al. 2004). Because this does not significantly predate the opening of the Bering Strait (5.4–5.5 Ma; Gladenkov et al. 2002), Ontocetus most likely dispersed through the Arctic (Kohno et al. 1995b; Kohno and Ray 2008).
We hypothesize that Ontocetus and Odobenus were completely temporally disjunct within the Atlantic basin, but note that the Middle Pleistocene record is likely incomplete, and similarly note an equivalent 1 my gap between the two lineages in the western North Pacific record (e.g., Kohno et al. 1995b, 1998; Miyazaki et al. 1995). Few fossils of Odobenus exist from high latitudes, suggesting either collection bias or perhaps suggesting that Odobenus was not cold-adapted until geologically recent times. The earliest high-latitude fossil occurrences of Odobenus date to the latest Middle Pleistocene (~127 Ka) of Amchitka Island, Alaska, USA associated with a cold temperate foraminiferal assemblage (Whitmore and Gard 1977). This occurrence notably post-dates specimens from Virginia, indicating an earlier trans-Arctic dispersal (Fig. 4). Furthermore, dispersal may have taken place entirely during interglacial periods. We note that at present, the majority of the fossil record of Odobenus is associated with ice-free regions. This fact, in concert with evidence of temperate latitude haulout and breeding sites as recent as the 18th century (McLeod et al. 2014) may have implications for of modern walrus conservation as several populations are being affected by rapid loss of sea ice driven by warming climate (Laidre et al. 2008). More field exploration of Pliocene–Middle Pleistocene fossil localities within the Arctic Circle (e.g., Gubik Formation, Alaska) with special attention towards walrus fossils and associated paleoclimatic indicators is necessary to further evaluate the timing of dispersal and timing of cold climate adaptation within the Odobenus lineage.
Conclusions
We report a new specimen of Ontocetus emmonsi from a previously unstudied vertebrate fossil assemblage from the Austin Sand Pit in Ridgeville, South Carolina, consisting of a complete, well-preserved tusk. The large size, curvature, labial grooves, and narrow cross-section indicate it is the extinct walrus Ontocetus rather than extant Odobenus. A brief stratigraphic description of the unstudied deposit is provided, and the rich vertebrate assemblage (52 taxa) is herein named the Ridgeville Local Fauna. Biochronologically significant vertebrate fossils from the locality, namely the giant beaver Castoroides sp. and the snaggletooth shark Hemipristis serra, constrain the age of the deposit to 1.8–1.1 Ma (latest Pliocene to Early Pleistocene), permitting correlation of the deposit at the Austin Sand Pit with the Waccamaw Formation. This is the youngest record of Ontocetus emmonsi. A comprehensive survey of up to date stratigraphic literature for walrus-bearing deposits in the North Atlantic found a range of 5.8–1.1 Ma for Ontocetus emmonsi and also indicates that no Atlantic fossil occurrences of Odobenus are older than 244 Ka. Given available evidence, Odobenus and Ontocetus were temporally separated in the North Atlantic for approximately 1 my and thus did not compete for resources and reflect separate early Pliocene (Ontocetus) and Middle–Late Pleistocene (Odobenus) arctic dispersals into the North Atlantic from the North Pacific rather than in-situ evolution. Further study of Plio-Pleistocene walrus biogeography may shed light on contemporary environmental pressures faced by walruses owing to the rapidly warming Arctic climate.
Acknowledgements
We would like to thank Matthew Swilp (Shamokin, Pennsylvania, USA) for collecting CCNHM-1144 and Mace Brown (Mount Pleasant, South Carolina, USA) for donating the specimen to CCNHM, Matthew Gibson and Jessica Peragine (ChM) for assistance with ChM collections, and Thomas Deméré (San Diego Natural History Museum, USA), Naoki Kohno (National Museum of Nature and Science, Tsukuba, Japan), Morgan Churchill (University of Wisconsin Osh Kosh, Wisconsin, USA), Robert Weems (Gainesville, Virginia, USA), and Scott Harris (College of Charleston, South Carolina, USA) for discussions on fossil walruses and South Carolina stratigraphy. We would also like to thank the late Kurt Austin, James Austin (Ridgeville, South Carolina, USA), Mark Bunce (Ladson, South Carolina, USA), Ashby Gale (James Island, South Carolina, USA), and Jordan Taylor (Summerville, South Carolina, USA) for access to the site for further collection and study as well as field assistance. Thanks to Leonard Dewaele (University of Ghent, Belgium) and N. Kohno and the editor Olivier Lambert (Institut royal des Sciences naturelles de Belgique, Brussels, Belgium), for constructive reviews.
References
Bell, C.J., Lundelius, E.L., Barnosky, A.D., Graham, R.W., Lindsay, E.H., Ruez, D.R., Semken, H.A., Webb, S.D., and Zakrzewski, R.J. 2004. The Blancan, Irvingtonian, and Rancholabrean Mammal Ages. In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology, 232–314. Columbia University Press, New York.
Berry, E.W. and Gregory, W.K. 1906. Prorosmarus alleni, a new genus and species of walrus from the upper Miocene of Yorktown, Virginia.American Journal of Science 21: 444–450. Crossref
Bianucci, G. 1996. The Odontoceti (Mammalia, Cetacea) from Italian Pliocene systematics and phylogenesis of Delphinidae. Palaeontolographia Italica 83: 73–167.
Boessenecker, R.W. 2013. A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, Part II: Pinnipeds and cetaceans. Geodiversitas 35: 815–940. Crossref
Boessenecker, R.W. 2017. A new early Pliocene record of the toothless walrus Valenictus (Carnivora, Odobenidae) from the Purisima Formation of Northern California. PaleoBios 34: 1–6.
Cicimurri, D.J. and Knight, J.L. 2009. Late Oligocene sharks and rays from the Chandler Bridge Formation, Dorchester County, South Carolina, USA. Acta Palaeontologica Polonica 54: 627–647. Crossref
Deméré, T.A. 1994a. The family Odobenidae: a phylogenetic analysis of fossil and living taxa. Proceedings of the San Diego Society of Natural History 29: 99–123.
Deméré, T.A. 1994b. Two new species of fossil walruses (Pinnipedia: Odobenidae) from the Upper Pliocene San Diego Formation, California. Proceedings of the San Diego Society of Natural History 29: 77–98.
Fields, S.E., McDonald, H.G., Knight, J.L., and Sanders, A.E. 2012. The ground sloths (Pilosa) of South Carolina. PalArch’s Journal of Vertebrate Paleontology 9 (3): 1–18.
Gladenkov, A.Y., Oleinik, A.E., Marincovich, L., and Barinov, B.K. 2002. A refined age for the earliest opening of Bering Strait. Palaeogeography, Palaeoclimatology, Palaeoecology 183: 321–328. Crossref
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M. 2012. The Geologic Time Scale.1176 pp. Elsevier, Boston.
Kimura, T., Narita, K., Fujita, T., and Hasegawa, Y. 2007. A new species of Eubalaena (Cetacea: Mysticeti: Balaenidae) from the Gonda Formation (latest Miocene–early Pliocene) of Japan. Bulletin of the Gunma Museum of Natural History 11: 15–27.
Kohno, N. and Ray, C.E. 2008. Pliocene walruses from the Yorktown Formation of Virginia and North Carolina, and a systematic revision of the North Atlantic Pliocene walruses. Virginia Museum of Natural History Special Publication 14: 39–80.
Kohno, N., Barnes, L.G., and Hirota, K. 1995a. Miocene fossil pinnipeds of the genera Prototaria and Neotherium (Carnivora; Otariidae; Imagotariinae) in the North Pacific Ocean: Evolution, relationships and distribution. The Island Arc 3: 285–308. Crossref
Kohno, N., Narita, K., and Hakuichi, K. 1998. An early Pliocene odobenid (Mammalia: Carnivora) from the Joshita Formation, Nagano Prefecture, central Japan. Research Reports of the Shinshushinmachi Fossil Museum 1: 1–7.
Kohno, N., Tomida, Y., Hasegawa, Y., and Furusawa, H. 1995b. Pliocene tusked odobenids (Mammalia: Carnivora) in the Western North Pacific, and their paleobiogeography. Bulletin of the National Science Museum, Tokyo Series C (Geology & Paleontology) 21: 111–131.
Laidre, K.L., Stirling, I., Lowry, L.F., Wiig, Ø., Heide-Jørgensen, M.P., and Ferguson, S.H. 2008. Quantifying the sensitivity of arctic marine mammals to climate-induced habitat change. Ecological Applications 18: S97–S125. Crossref
Loch, C., Grando, L.J., Schwass, D.R., Kieser, J.A., Fordyce, R.E., and Simões-Lopes, P.C. 2013. Dental erosion in South Atlantic dolphins (Cetacea: Delphinidae): a macro and microscopic approach. Marine Mammal Science 29: 338–347. Crossref
MacGregor, D.A., Harris, W.B., Dietl, G.P., and Kelley, P.H. 2011. Strontium isotopic dating of the Waccamaw Formation at Acme, NC, and the Duplin Formation at Tar Heel, NC: A Plio-Pleistocene research progress report. Southeastern Section Geological Society of America Abstracts with Programs 43 (2): 4.
McLeod, B.A., Frasier, T.R., and Lucas, Z. 2014. Assessment of the extirpated maritimes walrus using morphological and ancient DNA analysis. PLoS ONE 9 (6): 399–569. Crossref
Miyazaki, S., Horikawa, H., Kohno, N., Hirota, K., Kimura, M., Hasegawa, Y., Tomida, Y., Barnes, L.G., and Ray, C.E. 1995. Summary of the fossil record of pinnipeds of Japan, and comparisons with that from the eastern North Pacific. The Island Arc 3: 361–372. Crossref
Morgan, G.S. 1994. Miocene and Pliocene marine mammal faunas from the Bone Valley Formation of central Florida. Proceedings of the San Diego Society of Natural History 29: 239–268.
Morgan, G.S. 2005. The great American biotic interchange in Florida. Bulletin of the Florida Museum of Natural History 45: 271–311.
Morgan, G.S. and Hulbert, R.C. 1995. Overview of the geology and vertebrate biochronology of the Leisey Shell Pit local fauna, Hillsborough County, Florida. Bulletin of the Florida Museum of Natural History 37: 1–92.
Parham, J.F., Donoghue, P.C.J., Bell, C.J., Calway, T.D., Head, J.J., Holroyd, P.A., Inoue, J.G., Irmis, R.B., Joyce, W.G., Ksepka, D.T., Patane, J.S.L., Smith, N.D., Tarver, J.E., Tuinen, M.v., Yang, Z., Angielczyk, K.D., Greenwood, J.M., Hipsley, C.A., Jacobs, L., Makovicky, P.J., Muller, J., Smith, K.T., Theodor, J.M., Warnock, R.C.M., and Benton, M.J. 2012. Best practices for justifying fossil calibrations. Systematic Biology 61: 346–359. Crossref
Ray, C.E. 1960. Trichecodon huxleyi (Mammalia: Odobenidae) in the Pleistocene of Southeastern United States. Bulletin of the Museum of Comparative Zoology 122: 129–142.
Ray, C.E. 1975. The relationships of Hemicaulodon effodiens Cope 1869 (Mammalia: Odobenidae). Proceedings of the Biological Society of Washington 26: 281–304.
Repenning, C.A. and Tedford, R.H. 1977. Otarioid seals of the Neogene. U.S. Geological Survey Professional Paper 992: 1–93.
Repenning, C.A., Ray, C.E., and Grigorescu, D. 1979. Pinniped biogeography. In: J. Gray and A.J. Boucot (eds.), Historical Biogeography, Plate Tectonics, and the Changing Environment, 357–369. Stanford University Press, California.
Scudder, S., Simons, E., and Morgan, G.S. 1995. Chondrichthyes and Osteichthyes from the early Pleistocene Leisey Shell Pit local fauna, Hillsborough County, Florida. Bulletin of the Florida Museum of Natural History 37: 251–272.
Tedford, R.H., Albright, L.B., III,, Barnosky, A.D., Ferrusquia-Villafranca, H., R.M. Jr., Swisher, C.C., III,, Voorhies, M.R., Webb, S.D., and Whistler, D.P. 2004. Mammalian biochronology of the Arikareean through Hemphillian interval (Late Oligocene through Early Pliocene epochs). In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology, 169–231. Columbia University Press, New York.
Tomida, Y. 1989. A new walrus from the middle Pleistocene of the Boso Peninsula, Japan, and its implication on odobenid paleobiogeography. Bulletin of the National Science Museum, Tokyo, Series C 15: 109–119.
Tseng, Z.J. and Geisler, J.H. 2016. The first fossil record of borophagine dogs (Mammalia, Carnivora) from South Carolina, U.S.A. Journal of Vertebrate Paleontology 36: e1062022. Crossref
Van Beneden, P.J. 1877. Description des ossements fossiles des environs d’Anvers. Annales du Musée royal d’histoire naturelle de Belgique 1: 1–88.
Wallace, S.C. and Hulbert, R.C. 2013. A new machairodont from the Palmetto Fauna (Early Pliocene) of Florida, with comments on the origin of the Smilodontini (Mammalia, Carnivora, Felidae). PLoS ONE 8 (3): e56173. Crossref
Weems, R.E., Lemon, E.M. Jr., and Nelson, M.S. 1994. Geology of the Pringletown, Ridgeville, Summerville, and Summerville Northwest 7.5 minute quadrangles, Berkeley, Charleston, and Dorchester Counties, South Carolina. US Geological Survey Miscellanous Investigations Map I-2502: 1–9.
Weems, R.E., Lemon, E.M. Jr., Nelson, M.S., Gohn, G.S., and Houser, B.B. 1987. Detailed sections from auger holes and outcrops in the Pringletown, Ridgeville, Summerville Northwest, and Summerville quadrangles, South Carolina. US Geological Survey Open File Report 87-524: 1–89.
Weems, R.E., Lewis, W.C., and Lemon, E.M. Jr. 2014. Surficial geologic map of the Charleston region, Berkeley, Charleston, Colleton, Dorchester, and Georgetown Counties, South Carolina. US Geological Survey Open-File Report 2013-1030. U.S. Geological Survey, Reston. Crossref
Weems, R.E., Bybell, L.M., Edwards, L.E., Lewis, W.C., Self-Trail, Albright, L.B., Cicimurri, D.J., Harris, W.B., Osborne, J.E., and Sanders, A.E. 2016. Stratigraphic revision of the Cooper Group and the Chandler Bridge and Edisto Formations in the coastal plain of South Carolina. South Carolina Geology 49: 1–24.
Whitmore, F.C. 1994. Neogene climatic change and the emergence of the modern whale fauna of the North Atlantic Ocean. Proceedings of the San Diego Society of Natural History 29: 223–227.
Whitmore, F.C. and Barnes, L.G. 2008. The Herpetocetinae, a new subfamily of extinct baleen whales (Mammalia, Cetacea, Cetotheriidae). Virginia Museum of Natural History Special Publication 14: 141–180.
Whitmore, F.C. and Gard, L.M.J. 1977. Steller’s sea cow (Hydrodamalis gigas) of late Pleistocene age from Amchitka, Aleutian Islands, Alaska. U.S. Geological Survey Professional Paper 1036: 1–19.
Whitmore, F.C. and Kaltenbach, J.A. 2008. Neogene Cetacea of the Lee Creek Phosphate Mine, North Carolina. Virginia Museum of Natural History Special Publication 14: 181–269.
Acta Palaeontol. Pol. 63 (2): 279–286, 2018
https://doi.org/10.4202/app.00454.2018