
Oligocene plant assemblage from Rębiszów, Lower Silesia: First “volcanic flora” from Poland
RAFAŁ KOWALSKI, GRZEGORZ WOROBIEC, ELŻBIETA WOROBIEC, and KATARZYNA KRAJEWSKA
Kowalski, R., Worobiec, G., Worobiec, E., and Krajewska, K. 2020. Oligocene plant assemblage from Rębiszów, Lower Silesia: First “volcanic flora” from Poland. Acta Palaeontologica Polonica 65 (2): 273–290.
Fossil plant macroremains preserved in laminated diatomites from Łysa Góra near Rębiszów, Lower Silesia, have been documented for the first time. The fossil assemblage consists mostly of leaves, but fruits, seeds and sporadic flowers also occur. Forty-three identified taxa represent nineteen plant families: Aceraceae, Berberidaceae, Betulaceae, Cornaceae, Cupressaceae, Elaeocarpaceae, Ericaceae, Fagaceae, Lauraceae, ?Leguminosae, ?Meliaceae, Magnoliaceae, Pinaceae, Rhamnaceae, Rosaceae, Salicaceae, Tilioideae, Ulmaceae, Vitaceae, and one incertae sedis. The prevalence of entire margined leaves, mostly represented by Majanthemophyllum basinerve and Daphnogene cinnamomifolia, and a significant presence of Calocedrus suleticensis and Liriodendron haueri, are characteristic features of the Rębiszów flora. The composition of the plant assemblage points to a mesophytic forest with some riparian elements and suggests favourable, warm climatic conditions. Radiometrically dated basalts overlying fossiliferous layers and composition of palynoflora suggest at least Chattian (late Oligocene) age. The lithology and floristic composition link the Rębiszów flora with the so-called volcanic floras of the Nerchau-Flörsheim or Kleinsaubernitz floristic complex (Oligocene) from Germany and Czech Republic with Suletice-Berand flora being especially close.
Key words: Acrogymnospermae, Angiospermae, sporomorphs, volcanic floras, diatomites, Paleogene, Poland.
Rafał Kowalski [rafal.kowalski@mz.pan.pl] and Katarzyna Krajewska (passed away 2011), Museum of the Earth in Warsaw, Polish Academy of Sciences, Al. Na Skarpie 27, 00-488 Warsaw, Poland.
Grzegorz Worobiec [g.worobiec@botany.pl] and Elżbieta Worobiec [e.worobiec@botany.pl], W. Szafer Institute of Botany, Polish Academy of Sciences, ul. Lubicz 46, 31-512 Kraków, Poland.
Received 8 October 2019, accepted 9 January 2020, available online 16 April 2020.
Copyright © 2020 R. Kowalski et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Localities where the late Paleogene plant remains occur are quite common in Central Europe, but fossil plant assemblages which were formed in connection with volcanic activity, so-called volcanic floras (Kvaček and Walther 1998), are rare. These floras, however, have particular relevance for our knowledge of Central European Paleogene vegetation. The importance of these type of floras is due to the fact that they represent zonal, upland vegetation and, often, they can be radiometrically dated. They also frequently provide exceptionally well preserved macro- and microfossils, both of plants and animals. The Oligocene volcanic floras are connected with distribution of the Central European Tertiary intraplate volcanic province (Birkenmajer et al. 2011) and have been documented so far from Germany and Czech Republic. Lower Silesia (SW Poland) belongs to the north-eastern part of this volcanic province as a part of the Bohemo-Silesian volcanic belt though Paleogene fossil macrofloras have not yet been reported from this area.
The fossil plant remains documented here were discovered at Łysa Góra (Urwista), located in the western part of Lower Silesia, ca. 2 km south of Rębiszów village (Fig. 1) by Stanisław Dyjor in 1979. The first attempts to investigate this fossil flora were undertaken much later, in the early 2000s by Katarzyna Krajewska. Krajewska and Dyjor planned a comprehensive collaborative paper about fossil flora from Rębiszów. Unfortunately, their untimely death thwarted this plan.
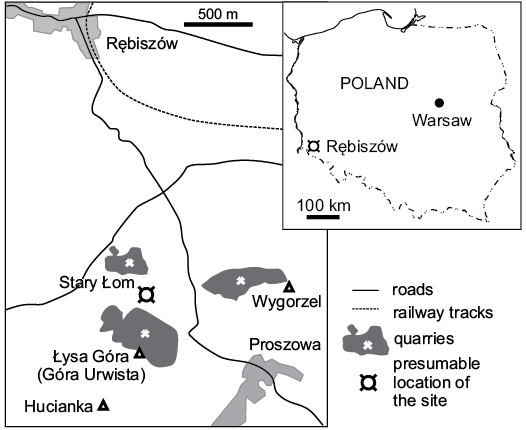
Fig. 1. Location of the Łysa Góra near Rębiszów, Lower Silesia, Poland.
Considering the uniqueness of the Rębiszów flora among other Cenozoic floras from Poland, and its significance to knowledge of the Oligocene volcanic floras of Central Europe, we decided to continue the investigations of Krajewska and Dyjor. This paper also commemorates Katarzyna Krajewska as a paleobotanist as well as the efforts she made in studying this flora.
Some data on the site location and local geology presented in the Geological settings originate from a draft by Stanisław Dyjor given in a letter to Katarzyna Krajewska dated March 14, 2006. However, we also present an alternative view on the general geology of this locality based on more recent studies on the geology of the Rębiszów locality. The taxonomic composition within the assemblage was preliminarily recognised by Krajewska and prepared in the form of a list of indicated specimens. She investigated some of the remains in consultation with Zlatko Kvaček in 2004.
In preparation of the present treatment, we carefully reexamined (RK and GW) the whole collection to evaluate the taxonomic determinations made by Krajewska. Most of the carpological remains in the collection that remained unrecognised or unnoticed by Krajewska were studied during the preparation of this paper. Preliminary palynological analysis of the deposits containing the macroremains was conducted by Aleksandra Kohlman-Adamska and Maria Ziembińska-Tworzydło (unpublished material), but while preparing this report new and detailed palynological investigations were made (EW) to supplement the preliminary results.
This paper briefly documents current knowledge on the Łysa Góra fossil assemblage, including palynological analysis, interpretation of the vegetation and paleoclimate, and comparison with other European assemblages of the same age and age determination. In recognition of the work already done on this collection we decided to include Katarzyna Krajewska posthumously as a co-author of this paper.
Institutional abbreviation.—KRAM-P: Palaeobotanical collections of W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, Poland.
Geological settings
Volcanic vents in the Rębiszów area are connected with the Rębiszów fault zone, where a tectonic contact with the gneisses and granitic gneisses of the Izera complex and the Neoproterozoic Stara Kamienica mica schist can be observed (Birkenmajer et al. 2011).
According to Birkenmajer et al. (2011), volcanic activity in Lower Silesia began in the Rupelian (early Oligocene) and continued with variable intensity to Zanclean (early Pliocene). One of the latest radiometric datings suggests that volcanic activity in the Rębiszów area took place in the late Oligocene. Dates obtained from ankaratrites exposed in the Łysa Góra quarry are of ca. 25.3±1.0 Ma, and those exposed in Grudza quarry, ca. 4 km East of the Rębiszów village, are of ca. 26.0±1.0 Ma (Birkenmajer et al. 2011).
According to Stanisław Dyjor (unpublished material, a letter to Katarzyna Krajewska, 14.03.2006), the layers with plant remains which he found were exposed somewhere in or near the “lower quarry”. Despite the imprecise localisation, we are almost certain that the “lower quarry” could equate to an abandoned quarry that is located within a hill called “Łysa Góra” (Fig. 1). Unfortunately, despite two excursions to search this area, we were unable to find the exact position of the site where the fossils were collected. Below, we summarise the interpretation of the local geological settings left by Stanisław Dyjor. According to him, some of the basaltic vents or lava flows in the Rębiszów area are covered with small patches of various Cenozoic sedimentary rocks, which were deposited in a small lake. The lake probably formed after the stream was restricted by lava flow. Layers with plant remains are a part of a sedimentary series that originally filled the paleovalley, although much of their northern part was eroded in the Pleistocene. Sedimentary series were deposited between the lava flow on the southern margin and gneisses on the northern erosional margin of the valley, and consisted mostly of loamy sand and gravel with tuff and tuffite intercalations in the lower part, and diatomite with fossil flora in the upper part of the profile. Mass development of diatoms was probably induced by silica-rich water. The high-silica content probably resulted from percolation of waters through silica-rich volcanic rocks.
We have, however, considerable doubts about the above presented interpretation. The chronological order proposed by Stanisław Dyjor (unpublished material) is in contradiction with the technical report on the geological setting of the basalt resources in Rębiszów, which we had the opportunity to see during our visit to the main office of Pri Bazalt S.A. mining company and information provided by Adamczyk (2008). Both of the mentioned sources clearly state that sedimentary rocks with fossiliferous layers found in the study area are overlain by basalt. According to Adamczyk (2008), Cenozoic sedimentary series in the Rębiszów area were preserved only due to the protective basalt layer. This alternative interpretation suggests that sedimentary rocks are older than the basalts, in this case providing the upper age limit of the Rębiszów fossil flora.
During an excursion (31.07.2019) to an abandoned quarry located on the Urwista and Odarte Skały hills, RK and Olaf Tietz and Joerg Büchner (both Senckenberg Museum of Natural History, Görlitz, Germany) discovered three new sites where sedimentary sequences are exposed. According to Olaf Tietz and Joerg Büchner (personal communication 2019), the lithological characteristics and position of the sedimentary sequences in relation to the overlying basalts suggest that these deposits may represent maar lake infill rather than the infill of a lake formed by the damming of a river by a lava flow.
Thus, the fossil plant assemblage from Rębiszow represents a so-called volcanic flora sensu Walther (2005), which is similar to the Oligocene floras from Northern Bohemia, Czechia and Upper Lusatia, Germany (Akhmetiev et al. 2009). These assemblages were usually preserved in diatomaceous deposits of maar crater lakes, such as late Oligocene fossil flora from Kleinsaubernitz (Walther 1999, 2005).
The genesis and lithology of the deposits with a plant assemblage from Rębiszów (diatomaceous sediments in a volcanogenic lake) are also similar to the late Eocene Florissant fossil beds from Colorado, USA, yielding the renowned plant and animal fossil Lagerstätte (Meyer et al. 2004).
Material and methods
Fossil leaves from Rębiszów are mostly impressions, although sometimes traces of coalified tissues are preserved, especially in the case of fruits or cones. Considering the state of preservation, leaf remains usually lacking cuticles were identified based only on their macromorphology. The studied fossil material (almost 600 rock samples) are housed in the W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków (KRAM-P 128/1–595). The studied fossil material from Rębiszów is probably the only such material obtained from the Łysa Góra (Urwista) locality. To our knowledge, no more material can be collected from the locality discovered by Stanisław Dyjor because the fossiliferous layer is no longer accessible.
Two samples from the sediments containing plant macroremains were used for the palynological study. They were prepared in the laboratory of the W. Szafer Institute of Botany PAS, using HCl, KOH, and HF (Moore et al. 1991). Ten slides were studied and in each sample more than 1000 sporomorphs (pollen grains and spores), as well as all co-occurring non-pollen palynomorphs, were counted.
Results
Plant macroremains assemblage.—We recognised 43 taxa among the plant macroremains from Rębiszów, but 36 are illustrated in this paper (Figs. 2–8, Table 1). Most numerous are angiosperm remains, but gymnosperms also represent a significant group. In respect of the systematic diversity the flowering plants also prevail and are represented by 19 families, including Aceraceae (Figs. 3D, 4F), Berberidaceae (Fig. 4G, K), Betulaceae (Figs. 3A, F, 4I, 5A, 7B), Cornaceae, Elaeocarpaceae (Fig. 6E), Ericaceae, Fagaceae (Fig. 6A–C, F–H), Lauraceae (Fig. 4A–E), ?Leguminosae (Fig. 4J), ?Meliaceae (Fig. 7A, E), Magnoliaceae (Fig. 3C), Rhamnaceae (Fig. 5B), Rosaceae (Figs. 7D, 8B), Salicaceae (Fig. 6D), Tilioideae (Fig. 3E), Ulmaceae (Fig. 3B), and Vitaceae (Fig. 5E, G). Conifers are represented by Cupressaceae: Calocedrus suleticensis (Brabenec, 1909) Kvaček, 1999 (Fig. 2A), Cunninghamia cf. miocenica Ettingshausen, 1872 (Fig. 2C), Cryptomeria cf. rhenana Kilpper, 1968 (Fig. 2D1, D2) and Pinaceae: Tsuga cf. moenana Kirchheimer, 1935 (Fig. 4H) and Tsuga sp. (Fig. 2B).

Fig. 2. Conifers impressions from Łysa Góra near Rębiszów, Poland, late Oligocene. A. Calocedrus suleticensis (Brabenec, 1909) Kvaček, 1999, KRAM-P 128/38, twig. B. cf. Tsuga sp., KRAM-P 128/50, twigs. C. Cunninghamia cf. miocenica Ettingshausen, 1872, KRAM-P 128/251, cone scale. D. Cryptomeria cf. rhenana Kilpper, 1968, KRAM-P 128/1; twigs (D1), cone (D2). Scale bars: A, B, D1, 10 mm; C, D2, 5 mm.

Fig. 3. Angiosperms impressions from Łysa Góra near Rębiszów, Poland, late Oligocene. A. Alnus julianiformis (Sternberg, 1823) Kvaček and Holy, 1974, KRAM-P 128/347, leaf. B. Ulmus fischeri Heer, 1856, KRAM-P 128/149, leaf. C. Liriodendron haueri Ettingshausen, 1869, KRAM-P 128/318, fruit. D. Acer cf. hercynicum Mai, 1978, KRAM-P 128/118, fruit. E. Craigia bronnii (Unger, 1845) Kvaček, Bůžek, and Manchester, 1991, KRAM-P 128/203, fruit. F. Alnus kefersteinii (Goeppert, 1838) Unger, 1845, KRAM-P 128/44, infructescence. Scale bars: A, B, F, 10 mm; C, D, 5 mm; E, 2 mm.

Fig. 4. Angiosperms impressions from Łysa Góra near Rębiszów, Poland, late Oligocene. A. cf. Laurophyllum acutimontanum Mai, 1963, leaf, KRAM-P 128/436. B–D. Daphnogene cinnamomifolia (Brongniart, 1822) Unger, 1850 forma lanceolata sensu Kvaček and Walther (1995), leaves. B. KRAM-P 128/131. C. KRAM-P 128/397. D. KRAM-P 128/298. E. Daphnogene cinnamomifolia (Brongniart, 1822) Unger, 1850 forma cinnamomifolia sensu Kvaček and Walther (1995), KRAM-P 128/256, leaf. F. Acer cf. polymorphoides Mai, 1987, KRAM-P 128/415, fruit. G. Mahonia sp., KRAM-P 128/35, leaf. H. Tsuga cf. moenana Kirchheimer, 1935, KRAM-P 128/75, cone. I. Ostrya atlantidis Unger, 1850, KRAM-P 128/318, fruit. J. cf. Leguminosites sp. or cf. Rhodomyrtophyllum sp., KRAM-P 128/88, leaf. K. Mahonia sp., KRAM-P 128/291, leaf. Scale bars: A–E, 10 mm; H, G, 5 mm; K, 3 mm; F, H, J, 2.5 mm; I, 1 mm.

Fig. 5. Angiosperms impressions from Łysa Góra near Rębiszów, Poland, late Oligocene. A. Carpinus sp. vel Ostrya sp., KRAM-P 128/546, leaf. B. Paliurus favonii Unger, 1847, KRAM-P 128/165, fruit. C, D, F. Majanthemophyllum basinerve (Rossmässler, 1840) Knobloch and Kvaček, 1996, leaves. C. KRAM-P 128/571. D. KRAM-P 128/289. F. KRAM-P 128/212. E. Vitis sp. or Ampelopsis sp., KRAM-P 128/205, leaf. G. Ampelopsis cf. rotundata Chandler, 1925, KRAM-P 128/109, seed. Scale bars: A, C–F, 10 mm; B, G, 1 mm.

Fig. 6. Angiosperms impressions from Łysa Góra near Rębiszów, Poland, late Oligocene. A. cf. Trigonobalanopsis rhamnoides (Rossmässler, 1849) Walther and Kvaček, 1988, KRAM-P 128/305, leaf. B. Trigonobalanopsis exacantha (Mai, 1970) Kvaček and Walther, 1988, KRAM-P 128/185, cupules. C. Quercus sp. or Castanopsis sp., KRAM-P 128/313, fruit. D. Populus cf. zaddachii Heer, 1959, KRAM-P 128/511, leaf. E. Sloanea artocarpites (Ettingshausen, 1869) Kvaček and Hably, 2001, KRAM-P 128/433, leaf. F, G. cf. “Quercus” bavarica Knobloch and Kvaček, 2004, leaves. F. KRAM-P 128/79. G. KRAM-P 128/212. H. Eotrigonobalanus furcinervis (Rossmässler, 1840) Walther and Kvaček, 1989, KRAM-P 128/234, leaf. Scale bars: A–C, 5 mm; D–H, 10 mm.

Fig. 7. Angiosperms impressions from Łysa Góra near Rębiszów, Poland, late Oligocene. A, E. cf. Cedrela macrophylla Andreánszky, 1955, leaves. A. KRAM-P 128/570. E. KRAM-P 128/516. B. Alnus cf. gaudinii (Heer, 1856) Knobloch and Kvaček, 1976, KRAM-P 128/390, leaf. C. cf. Acer tricuspidatum Bronn, 1838, KRAM-P 128/388, leaf. D. Prunus cf. scharfii Gregor, 1978, KRAM-P 128/352, endocarp. Scale bars: A–C, E, 10 mm; D, 1 mm.

Fig. 8. Angiosperms impressions from Łysa Góra near Rębiszów, Poland, late Oligocene. A. Dombeyopsis lobata Unger, 1850, KRAM-P 128/462, leaf. B. cf. Rosa lignitum Heer, 1869, KRAM-P 128/401, leaf. Scale bars 5 mm.
The plant assemblage is dominated by entire-margined leaves, which significantly overweigh toothed leaves, representing ca. 80% of the whole leaf assemblage. Among those, the most common species is Majanthemophyllum basinerve (Rossmässler, 1840) Knobloch and Kvaček, 1996 (ca. 46% of all leaves), which is a taxon of uncertain affinity. Representatives of Lauraceae (ca. 22% of all leaves) and Fagaceae (ca. 11% of all leaves) are less abundant in the Rębiszów flora. Among Lauraceae, the most common is Daphnogene cinnamomifolia (Brongniart, 1822) Unger, 1851 (ca. 96%), but the form D. lanceolata sensu Kvaček and Walther (1995) prevails over the form D. cinnamomifolia. Fagaceae are represented by Eotrigonobalanus furcinervis (Rossmässler, 1840) Walther and Kvaček, 1989, Trigonobalanopsis exacantha (Mai, 1970) Kvaček and Walther, 1988, cf. Trigonobalanopsis rhamnoides (Rossmaessler, 1840) Walther and Kvaček, 1988, cf. “Quercus” bavarica Knobloch and Kvaček, 2004, and Quercus sp. or Castanopsis sp.
The most common among toothed leaves are Alnus cf. gaudinii (Heer, 1856) Knobloch and Kvaček, 1976 and Alnus sp. (ca. 65%). Aside from leaf remains, Alnus Miller, 1754 is also represented by infructescence of A. kefersteinii (Goeppert, 1838) Unger, 1845. A very common element of the Rębiszów assemblage are fruits of Liriodendron haueri Ettingshausen, 1869 (more than 53 specimens). Characteristic but less common is Craigia bronnii (Unger, 1845) Kvaček, Bůžek, and Manchester, 1991 (ca. 11 specimens). Elements such as Ulmus sp., Ostrya atlantidis Unger, 1850, Mahonia sp., Dombeyopsis lobata Unger, 1850, Acer sp., Carpinus sp. vel Ostrya sp., Sloanea artocarpites (Ettingshausen, 1869) Kvaček and Hably, 2001, Populus cf. zaddachii Heer, 1859, cf. Rosa lignitum Heer, 1869, cf. Leguminosites sp. (or cf. Rhodomyrtophyllum sp.), and Vitis sp. or Ampelopsis sp. (arranged according to the number of specimens, see also Table 1) are sparse.
Table 1. Taxa recorded among leaf and fruit/seeds remains from Rębiszów. N, number of specimens.
| |
Type of remain |
N |
Specimens |
|
Cupressaceae |
|||
|
Calocedrus suleticensis (Brabenec, 1909) Kvaček, 1999 |
leaf |
71 |
1, 3, 11, 14, 32, 38, 43–45, 48, 52, 78, 79, 99, 101, 109, 119, 121, 122, 124, 139, 141, 143, 161, 209, 213, 214, 237, 238, 240, 258, 261, 263, 277, 291, 293, 298, 305, 307, 308, 318, 329, 335, 337, 343, 350, 361, 363, 366, 378, 397, 411, 417, 426, 437, 470, 472, 508, 519, 526–529, 531, 558, 562, 564, 567, 570, 571, 578 |
|
Cryptomeria cf. rhenana Kilpper, 1968 |
leaf, cone |
16 |
1, 40, 95, 142, 159, 200, 202, 239, 311, 321, 331, 354, 377, 386, 391, 442 |
|
Cunninghamia cf. miocenica Ettingshausen, 1872 |
scale |
1 |
251 |
|
Pinaceae |
|||
|
Tsuga cf. moenana Kirchheimer, 1935 |
cone |
5 |
33, 75, 245, 311, 351 |
|
cf. Tsuga sp. |
leaf |
1 |
50 |
|
Aceraceae |
|||
|
cf. Acer tricuspidatum Bronn, 1838 |
leaf |
1 |
388 |
|
Acer cf. hercynicum Mai, 1978 |
fruit |
1 |
118 |
|
Acer cf. polymorphoides Mai, 1987 |
fruit |
1 |
415 |
|
Acer sp. |
leaf |
1 |
501, |
|
fruit |
2 |
27, 558 |
|
|
Betulaceae |
|||
|
Alnus cf. gaudinii (Heer, 1856) Knobloch and Kvaček, 1976 |
leaf |
1 |
390 |
|
Alnus julianiformis (Sternberg, 1823) Kvaček and Holy, 1974 |
leaf |
1 |
347 |
|
?Alus rostaniana Saporta emend. Mai and Walther |
leaf |
1 |
290 |
|
Alnus kefersteinii (Goeppert, 1838) Unger, 1845 |
infructe-scence |
7 |
42, 44, 275–277, 298, 570 |
|
Aluns sp. |
leaf |
37 |
45, 49, 83, 112, 187, 196, 197, 198, 233, 262, 264, 290, 293, 297, 317, 338, 341, 343, 347, 355, 375, 386, 390, 391, 395, 429, 440, 444, 457, 461, 467, 507, 508, 509, 512, 527, 585 |
|
Ostrya atlantidis Unger, 1850 |
fruit |
3 |
120, 318, 526 |
|
Carpinus sp. or Ostrya sp. |
leaf |
1 |
546 |
|
Berberidaceae |
|||
|
Mahonia sp. |
leaf |
3 |
34, 35, 291 |
|
Cornaceae |
|||
|
Cornus cf. gorbunovii (Dorofeev, 1963) Negru, 1972 |
fruit |
1 |
51 |
|
Elaeocarpaceae |
|||
|
Sloanea artocarpites (Ettingshausen, 1869) Kvaček and Hably, 2001 |
leaf |
1 |
433 |
|
Ericaceae |
|||
|
Ericaceae gen. et sp. indet. |
fruit |
2 |
408, 417 |
|
Fagaceae |
|||
|
Eotrigonobalanus furcinervis (Rossmässler, 1840) Walther and Kvaček, 1989 |
leaf |
2 |
234, 241 |
|
Trigonobalanopsis exacantha (Mai, 1970) Kvaček and Walther, 1988 |
fruit |
5 |
49, 52, 185, 332, 346 |
|
cf. Trigonobalanopsis rhamnoides (Rossmässler, 1840) Walther and Kvaček, 1988 |
leaf |
16 |
12, 78, 80, 92, 105, 118, 134, 138, 150, 151, 180, 305, 307, 397, 547, 570 |
|
cf. “Quercus” bavarica Knobloch and Kvaček, 2004 |
leaf |
20 |
64, 79, 89, 93, 99, 195, 202, 216, 235, 259, 303, 349, 379, 437, 440, 519, 528, 542, 547, 563 |
|
Quercus sp. or Castanopsis sp. |
fruit |
1 |
313 (part and counterpart) |
|
Lauraceae |
|||
|
Daphnogene cinnamomifolia (Brongniart, 1822) Unger, 1850 forma cinnamomifolia sensu Kvaček and Walther (1995) |
leaf |
3 |
256, 280, 285 |
| |
Type of remain |
N |
Specimens |
|
Daphnogene cinnamomifolia (Brongniart, 1822) Unger, 1850 forma lanceolata sensu Kvaček and Walther (1995) |
leaf |
72 |
1, 4, 10, 11, 15, 28, 32, 50, 52, 66, 78, 79, 80, 82, 94, 101, 108, 113, 127, 131, 151, 160, 178, 181–183, 201, 202, 215, 237, 238, 257, 263–265, 268, 273, 280, 285, 293, 296, 298, 305, 316, 322, 325, 327, 338, 349, 356, 359, 361, 365, 376, 391, 397, 418, 433, 436, 440, 456, 459, 470, 471–475, 496, 502, 524, 525, 537, 547, 565, 576, 588, 592 |
|
cf. Laurophyllum acutimontanum Mai, 1963 |
leaf |
3 |
129, 190, 436 |
|
?Leguminosae |
|||
|
cf. Leguminosites sp or cf. Rhodomyrtophyllum sp. |
leaf/ |
1 |
88 |
|
Magnoliaceae |
|||
|
Liriodendron haueri Ettingshausen, 1869 |
fruit or seed |
53 |
10, 13, 14, 22, 26, 78, 98, 99, 101, 156, 162, 169, 188, 207, 209, 211, 212, 218, 265, 293, 318, 321, 360, 376, 378, 387, 400, 402, 403, 407, 413, 414, 423, 428, 439, 444, 462, 465, 470, 487, 500, 519, 522, 525, 527, 528, 529, 542, 543, 563, 568, 571, 576 |
|
?Meliaceae |
|||
|
cf. Cedrela macrophylla Andreánszky, 1955 |
leaf |
3 |
516, 569, 570 |
|
Rhamnaceae |
|||
|
Paliurus favonii Unger, 1847 |
fruit |
1 |
165, 170 (part and counterpart) |
|
Rosaceae |
|||
|
Prunus cf. scharfii Gregor, 1978 |
fruit |
2 |
39, 352 |
|
cf. Rosa lignitum Heer, 1869 |
leaf/ leaflet |
1 |
401 |
|
Salicaceae |
|||
|
Populus cf. zaddachii Heer, 1859 |
leaf |
1 |
511, 513 (part and counterpart) |
|
Tilioideae |
|||
|
Craigia bronnii (Unger, 1845) Kvaček, Bůžek and Manchester, 1991 |
fruit |
11 |
83, 158, 203, 264, 319, 378, 413, 440, 543, 574, 582 |
|
Dombeyopsis lobata Unger, 1850 |
leaf |
3 |
490, 584, 585 |
|
Ulmaceae |
|||
|
Ulmus fischeri Heer, 1856 |
leaf |
1 |
149 |
|
Ulmus sp. |
leaf |
5 |
94, 118, 264, 443, 529 |
|
Vitaceae |
|||
|
Ampelopsis cf. rotundata Chandler, 1925 |
seed |
1 |
109 |
|
Vitis sp. or Ampelopsis sp. |
leaf |
1 |
205, 207 (part and counterpart) |
|
Incertae sedis |
|||
|
Majanthemophyllum basinerve (Rossmässler, 1840) Knobloch and Kvaček, 1996 |
leaf/ leaflet |
162 |
1, 6, 10–12, 18, 23, 25, 29–32, 43, 58, 63, 65, 74, 76, 77, 79, 80, 84–87, 95, 98, 100, 102, 109, 114, 115, 132, 133, 135, 136, 139, 141, 144, 150, 158, 165, 171–173, 181, 188, 197, 202, 203, 209–212, 215, 220, 221, 223, 224, 232, 235, 240, 244, 247, 248, 251, 255, 260, 265, 270, 285, 288, 289, 292, 293, 295, 298, 300, 301, 302, 304, 307, 313, 315, 316, 318–322, 324, 333, 337, 340, 342, 345, 348–350, 357, 359, 367, 372, 375, 376, 377, 379, 384, 391, 392, 397, 413, 421, 429, 433, 436, 442, 444, 450, 452, 453, 456, 464, 468, 476, 480, 483, 485, 490, 498, 502, 503, 510, 513, 515, 518, 522, 526, 528, 531, 533–535, 541–543, 547, 549, 551, 553–555, 562, 563, 569–571, 576, 582, 589, 591, 595 |
Palynoflora.—Palynological analysis revealed rich and well-preserved palynoflora. A total of 88 fossil-species were identified, including 19 species of spores, 15 species of pollen grains of gymnosperms, and 54 species of angiosperms. Palaeotropical and palaeotropical/warm-temperate elements dominate the spore-pollen spectra (Table 2).
Table 2. Spores, pollen grains, non-pollen palynomorphs, and palynoclasts recorded in palynological samples from the Rębiszów. Taxonomy and botanical affinity according to Stuchlik et al. (2001, 2002, 2009, 2014). The following paleofloristical elements have been distinguished: palaeotropical (P), including: tropical (P1) and subtropical (P2), “arctotertiary” (A), including: warm-temperate (A1) and temperate (A2), and cosmopolitan (P/A). ×, unassigned.
|
Fossil taxa |
Botanical affinity |
Element |
Sample 1 |
Sample 2 |
|
Spores |
||||
|
Baculatisporites sp. |
Osmundaceae: Osmunda Linnaeus, 1753 |
P/A |
2 |
1 |
|
Cicatricosisporites chattensis Krutzsch, 1961 |
Schizaeaceae: Anemia Swartz, 1806 |
P1 |
1 |
|
|
Concavisporites pseudopartitus Krutzsch, 1959 |
Gleicheniaceae? |
P |
2 |
|
|
Cryptogrammasporis sp. |
Pteridaceae: Cryptogramma Brown in Franklin, 1823 |
A1 |
1 |
|
|
Distancoraesporis sp. |
Sphagnaceae: Sphagnum Linnaeus, 1753 |
P/A |
1 |
|
|
Echinatisporis sp. |
Selaginellaceae: Selaginella P. de Beauvois, 1804 |
P/A |
1 |
1 |
|
Foveotriletes semifovearis Krutzsch, 1967 |
unknown |
unknown |
1 |
|
|
Laevigatosporites haardtii (Potonié and Venitz, 1934) Thomson and Pflug, 1953 + Laevigatosporites sp. |
Polypodiaceae, Davalliaceae, and other ferns |
P/A |
9 |
1 |
|
Leiotriletes maxoides Krutzsch, 1962 + L. wolffii Krutzsch, 1962 + Leiotriletes sp. |
Lygodiaceae and other ferns |
P |
12 |
1 |
|
Polypodiaceoisporites sp. |
Pteridaceae: Pteris Linnaeus, 1753 |
P/A1 |
1 |
|
|
Retitriletes sp. |
Lycopodiaceae: Lycopodium Linnaeus, 1753 |
P/A |
2 |
2 |
|
Selagosporis sp. |
Lycopodiaceae: Huperzia Bernhardi, 1801 |
P/A |
2 |
|
|
Triplanosporites sp. |
unknown |
P |
3 |
1 |
|
Verrucatosporites alienus (Potonié, 1931) Thomson and Pflug, 1953 + V. favus (Potonié, 1931) Thomson and Pflug, 1953 + Verrucatosporites sp. |
Davalliaceae, Dennstaedtiaceae, and other ferns |
P/A |
3 |
3 |
|
Pollen grains of gymnosperms |
||||
|
Cathayapollis wilsonii (Sivak, 1976) Ziembińska-Tworzydło in Stuchlik et al., 2002 + Cathayapollis sp. |
Pinaceae: Cathaya Chun and Kuang, 1962 |
A1 |
1 |
6 |
|
Cunninghamiaepollenites sp. |
Cupressaceae: Cunninghamia Brown in Richard, 1826 |
A1 |
4 |
|
|
Cupressacites sp. |
Cupressaceae |
A1 |
7 |
5 |
|
Inaperturopollenites concedipites (Wodehouse, 1933) Krutzsch, 1971 + I. dubius (Potonié and Venitz, 1934) Thomson and Pflug, 1953 |
Cupressaceae: Taxodium Richard, 1810, Glyptostrobus Endlicher, 1847 |
P2/A1 |
590 |
317 |
|
Piceapollis sp. |
Pinaceae: Picea Dietrich, 1824 |
A |
2 |
11 |
|
Pinuspollenites labdacus (Potonié, 1931) Raatz, 1937 ex Potonié, 1958 + Pinuspollenites sp. |
Pinaceae: Pinus Linnaeus, 1753 |
A |
7 |
51 |
|
Sciadopityspollenites crassus Krutzsch, 1971 ex Kohlman-Adamska, 1993 + Sciadopityspollenites sp. |
Sciadopityaceae: Sciadopitys Siebold and Zuccarini, 1842 |
A1 |
3 |
1 |
|
Sequoiapollenites gracilis Krutzsch, 1971 + Sequoiapollenites sp. |
Cupressaceae: Sequoia Endlicher, 1847, Sequoiadendron Buchholz, 1939, Metasequoia Hu and Cheng, 1948, Cryptomeria Don, 1838 |
A1 |
59 |
32 |
|
Zonalapollenites verrucatus Krutzsch, 1971 ex Ziembińska-Tworzydło, 1974 + Zonalapollenites sp. |
Pinaceae: Tsuga (Endlicher, 1847) Carrière, 1855 |
A1 |
29 |
76 |
|
Pollen grains of angiosperms |
||||
|
Aceripollenites sp. |
Sapindaceae: Acer Linnaeus, 1753 |
A1 |
4 |
2 |
|
Alnipollenites verus (Potonié, 1931) Potonié, 1931 |
Betulaceae: Alnus Miller, 1754 |
P2/A |
6 |
20 |
|
Araliaceoipollenites sp. |
Araliaceae |
P/A1 |
1 |
|
|
Arecipites sp. |
Arecaceae, Butomaceae, Ammarylidaceae, Araceae |
P/A |
1 |
|
|
Boehlensipollis hohli Krutzsch, 1962 |
Elaeagnacea, Lythraceae |
P2/A1 |
3 |
|
|
Carpinipites carpinoides (Pflug in Thomson and Pflug, 1953) Nagy, 1985 |
Betulaceae: Carpinus Linnaeus, 1753 |
P2/A1 |
10 |
25 |
|
Caryapollenites simplex (Potonié, 1931) Raatz, 1937 ex Potonié, 1960 |
Juglandaceae: Carya Nuttall, 1818 |
A1 |
4 |
3 |
|
Cornaceaepollis satzveyensis (Pflug in Thomson and Pflug, 1953) Ziembińska-Tworzydło in Ziembińska-Tworzydło et al., 1994 ex Jansonius, Hills, and Hartkopf-Fröder, 1998 |
Mastixiaceae |
P1 |
14 |
4 |
|
Fossil taxa |
Botanical affinity |
Element |
Sample 1 |
Sample 2 |
|
Cupuliferoipollenites oviformis (Potonié, 1934) Potonié, 1951 ex Potonié, 1960 + C. pusillus (Potonié, 1934) Potonié, 1951 ex Potonié, 1960 |
Fagaceae: Castanea
Miller, 1754, Castanopsis |
P2/A1 |
45 |
134 |
|
Cyrillaceaepollenites exactus (Potonié, 1931) Potonié, 1960 + C. brühlensis (Thomson in Potonié et al., 1950) Durska in Stuchlik et al., 2014 |
Cyrillaceae, Clethraceae |
P |
5 |
16 |
|
Dicolpopollis middendorfii (Potonié, 1931) Krutzsch, 1961 |
Arecaceae |
P |
|
1 |
|
Edmundipollis edmundii (Potonié, 1931) Konzalová, Słodkowska and Ziembińska-Tworzydło in Stuchlik et al., 2014 |
Mastixiaceae |
P1 |
2 |
1 |
|
Ericipites roboreus (Potonié, 1931) Krutzsch, 1970 + Ericipites sp. |
Ericaceae |
P/A |
3 |
3 |
|
Faguspollenites sp. |
Fagaceae: Fagus Linnaeus, 1753 |
A |
2 |
3 |
|
Fraxinipollis sp. |
Oleaceae: Fraxinus Linnaeus, 1753 |
P/A |
5 |
5 |
|
Fususpollenites fusus (Potonié, 1931) Kedves, 1978 |
Fagaceae: Trigonobalanus Forman, 1962 s.l. (incl. Colombobalanus Nixon and Crepet, 1989) |
P1 |
36 |
119 |
|
Ilexpollenites margaritatus (Potonié, 1931) Thiergart, 1938 |
Aquifoliaceae: Ilex Linnaeus, 1753 |
P2 |
4 |
|
|
Intratriporopollenites insculptus Mai, 1961 + Intratriporopollenites sp. |
Malvaceae: Tilioideae (incl. Craigia Smith and Evans, 1921) |
P/A1 |
5 |
1 |
|
Juglanspollenites verus Raatz, 1937 |
Juglandaceae: Juglans Linnaeus, 1753 |
A1 |
5 |
8 |
|
Magnoliaepollenites neogenicus (Krutzsch, 1970) Mohr, 1984 |
Magnoliaceae: Magnolia Linnaeus, 1753 |
P/A1 |
|
1 |
|
Meliaceoidites sp. |
Meliaceae |
P |
2 |
|
|
Momipites quietus (Potonié, 1931) Nichols, 1973 |
Juglandaceae: Engelhardia Leschenault and Blume, 1826, Alfaroa Standley, 1927, Oreomunnea Oersted, 1856 |
P |
2 |
2 |
|
Myricipites sp. |
Myricaceae |
P2/A |
1 |
4 |
|
Nyssapollenites contortus (Pflug and Thomson, 1953) Nagy, 1985 |
Nyssaceae: Nyssa Linnaeus, 1753 |
P2/A1 |
3 |
|
|
Oligopollis bilateralis Krutzsch, 1959 |
unknown |
unknown |
1 |
|
|
Quercoidites
microhenricii (Potonié, 1931) Potonié, Thomson and
Thiergart, 1950 ex Potonié, 1960 |
Fagaceae: Quercoideae |
P2/A1 |
11 |
12 |
|
Quercopollenites sp. |
Fagaceae: Quercus Linnaeus, 1753 |
P2/A1 |
1 |
|
|
Parthenopollenites formosus (Mamczar, 1960) Worobiec in Stuchlik et al., 2014 |
Vitaceae |
P/A1 |
4 |
4 |
|
Platanipollis ipelensis (Pacltová, 1966) Grabowska in Ziembińska-Tworzydło et al., 1994 |
Platanaceae: Platanus Linnaeus, 1753 |
P/A1 |
1 |
|
|
Platycaryapollenites sp. |
Juglandaceae: Platycarya Siebold and Zuccarini, 1843 |
P2/A1 |
|
1 |
|
Polyatriopollenites sp. |
Juglandaceae: Pterocarya Kunth, 1824 |
A1 |
7 |
7 |
|
Reevesiapollis triangulus (Mamczar, 1960) Krutzsch, 1970 |
Malvaceae: Reevesia Lindley, 1827 |
P |
1 |
|
|
Salixipollenites sp. |
Salicaceae: Salix Linnaeus, 1753 |
A |
2 |
3 |
|
Sapotaceoidaepollenites sp. |
Sapotaceae |
P |
4 |
4 |
|
Styraxipollis sp. |
Styracaceae: Styrax Linnaeus, 1753 |
P/A1 |
4 |
|
|
Symplocoipollenites sp. |
Symplocaceae: Symplocos Jacquin, 1760 |
P |
1 |
|
|
Tricolporopollenites fallax (Potonié, 1934) Krutzsch in Krutzsch et al., 1960 + T. liblarensis (Thomson in Potonié et al., 1950) Hochuli, 1978 + T. quisqualis (Potonié, 1934) Krutzsch, 1954 |
Fabaceae |
P/A |
13 |
85 |
|
Tricolporopollenites pseudocingulum (Potonié, 1931) Thomson and Pflug, 1953 |
?Fagaceae, ?Styracaceae |
P/A1 |
3 |
1 |
|
Tricolporopollenites spinus Krutzsch, 1962 |
unknown |
P |
2 |
|
|
Tricolporopollenites staresedloensis Krutzsch and Pacltová in Krutzsch, 1969 |
Hamamelidaceae: Parrotia Meyer, 1831, Distylium Siebold and Zuccarini, 1841 |
P2 |
11 |
7 |
|
Tricolporopollenites theacoides (Roche and Schuler, 1976) Kohlman-Adamska and Ziembińska-Tworzydło, 2000 |
Theaceae |
P |
1 |
|
|
Fossil taxa |
Botanical affinity |
Element |
Sample 1 |
Sample 2 |
|
Tricolporopollenites villensis (Thomson in Potonié et al., 1950) Thomson and Pflug, 1953 + T. dolium (Potonié, 1931) Thomson and Pflug, 1953 |
?Fagaceae |
unknown |
34 |
13 |
|
Tricolporopollenites sp. |
?Fagaceae |
P/A |
3 |
1 |
|
Trivestibulopollenites betuloides Pflug in Thomson and Pflug, 1953 |
Betulaceae: Betula Linnaeus, 1753 |
A |
1 |
7 |
|
Ulmipollenites undulosus Wolff, 1934 |
Ulmaceae: Ulmus Linnaeus, 1753 |
A |
2 |
1 |
|
Vitispollenites tener Thiele-Pfeiffer, 1980 |
Vitaceae: Vitis Linnaeus, 1753 |
P2/A1 |
5 |
4 |
|
Other pollen grains |
unknown |
unknown |
9 |
8 |
|
Sum of spores and pollen grains |
1027 |
1019 |
||
|
Selected non-pollen palynomorphs and palynoclasts |
||||
|
Botryococcus braunii Kützing, 1849 |
Dictyosphaeriaceae: Botryococcus braunii Kützing, 1849 |
× |
5 |
3 |
|
Diagonalites diagonalis Krutzsch and Pacltová, 1990 |
Zygnemataceae: Mougeotia laetevirens (Braun 1855) Wittrock, 1877 type |
× |
|
1 |
|
Ovoidites
elongatus (Hunger, 1952) Krutzsch, 1959 |
Zygnemataceae: Spirogyra Link in Nees, 1820 |
× |
1 |
1 |
|
?fragment of microsporangium |
?Salviniaceae: Salvinia Séguier, 1754 |
× |
|
1 |
|
Leaf-spines |
Ceratophyllaceae: Ceratophyllum Linnaeus, 1754 |
× |
|
3 |
|
Spores of fungi |
fungi |
× |
2 |
1 |
|
Total |
1035 |
1029 |
||
Among gymnosperms, the most common are pollen grains of Cupressaceae (fossil-genera Inaperturopollenites Pflug and Thomson, 1953 and Sequoiapollenites Thiergart, 1958), Tsuga (Endlicher) Carrière, 1855 (mainly fossil-species Zonalapollenites verrucatus Krutzsch, 1971) and bisaccate Pinaceae. Angiosperms are dominated by pollen grains of Fagaceae, including small tricolporate grains: Cupuliferoipollenites oviformis (Potonié, 1934) Potonié, 1951, C. pusillus (Potonié, 1934) Potonié, 1951, Fususpollenites fusus (Potonié, 1931) Kedves, 1978), Tricolporopollenites villensis (Thomson, 1950) Thomson and Pflug, 1953, T. dolium (Potonié, 1931) Thomson and Pflug, 1953, Quercoidites microhenricii (Potonié, 1931) Potonié, Thomson, and Thiergart, 1950, Q. henricii (Potonié, 1931) Potonié, Thomson, and Thiergart, 1950, and a few specimens of Faguspollenites Raatz, 1937. The pollen of Betulaceae: mainly Alnipollenites verus (Potonié, 1931) Potonié, 1931 and Carpinipites carpinoides (Pflug, 1953) Nagy, 1985); Cyrillaceae/Clethraceae: mainly Cyrillaceaepollenites exactus (Potonié, 1931) Potonié, 1960); Fabaceae: Tricolporopollenites fallax (Potonié, 1934) Krutzsch, 1960, T. liblarensis (Thomson, 1950) Hochuli, 1978, and T. quisqualis (Potonié, 1934) Krutzsch, 1954); Hamamelidaceae: Tricolporopollenites staresedloensis Krutzsch and Pacltová, 1969); Juglandaceae; Mastixiaceae: Cornaceaepollis satzveyensis (Pflug, 1953) Ziembińska-Tworzydło, 1994; Sapotaceae and Vitaceae are also common. In addition, pollen grains of the families Aquifoliaceae: Ilexpollenites margaritatus (Potonié, 1931) Thiergart, 1938; Ericaceae; Meliaceae; Nyssaceae: Nyssapollenites contortus (Pflug and Thomson, 1953) Nagy, 1985; Oleaceae; Salicaceae; Tilioideae: mainly Intratriporopollenites insculptus Mai, 1961; Sapindaceae: Aceripollenites Nagy, 1969; Ulmaceae, and others are present.
Cryptogams are represented mainly by ferns from the families Lygodiaceae, Osmundaceae, Pteridaceae, Schizaeaceae, and others, plus a few spores of Lycopodiaceae and Selaginellaceae. Among non-pollen palynomorphs, freshwater algae (Botryococcus Kützing, 1849 and zygospores of Zygnemataceae) are present. Spores of fungi are almost absent. In addition, some leaf-spines of plants with submerged leaves (Alismataceae, Hydrocharitaceae vel Ceratophyllum Linnaeus, 1754) and probably a fragment of a microsporangium of Salvinia Séguier, 1754, were encountered.
Discussion
Taphonomy and vegetation reconstruction.—In view of the lithology of the fossiliferous layer and preservation of the plant remains, as well as composition of the plant assemblage, we can interprete the sedimentary environment as calm, and most probably related to a small water body (maar lake?). This is confirmed by the presence of Botryococcus algae and zygospores of Zygnemataceae (Spirogyra Link, 1820, and Mougeotia Agardh, 1824). Extant representatives of Zygnemataceae occur in shallow, stagnant waters, which warm up easily, and these algae produce resting cells (e.g., zygospores) that enable them to survive through unfavourable growth conditions, e.g., desiccation (Worobiec 2014 and literature cited therein). The presence of leaf-spines of plants with submerged leaves also points to the occurrence of submerged (entirely under water) aquatic plants, such as Hydrocharitaceae or Ceratophyllum commonly found in ponds, marshes, and quiet streams. Remains of the water fern Salvinia are another example of floating aquatics growing in standing waters.
Well-preserved leaves and sparse flowers suggest that the plant remains were carried by wind and water only over a short distance and originated from vegetation surrounding the basin (see e.g., Ferguson 1985).
Based on the floristic composition of the Rębiszów fossil assemblage we can distinguish three types of vegetation. The vast majority of the fossil remains represents plants of the mesophytic forest around the sedimentary basin. The angiosperm elements of this vegetation are as follows (arranged according to frequency): Majanthemophyllum basinerve (Rossmässler, 1840) Knobloch and Kvaček, 1996, Daphnogene cinnamomifolia (Brongniart, 1822) Unger, 1851, Liriodendron haueri Ettingshausen, 1869; Fagaceae (Trigonobalanopsis cf. “Quercus” bavarica), Craigia bronnii (Unger, 1845) Kvaček, Bůžek, and Manchester, 1991, Sloanea artocarpites (Ettingshausen, 1869) Kvaček and Hably, 2001, Ostrya sp., Acer sp. div, Mahonia sp., cf. Laurophyllum acutimontanum Mai, 1963, cf. Leguminosites sp. or cf. Rhodomyrtophyllum sp., and cf. Rosa lignitum Heer, 1869. Conifers, especially Calocedrus Kurz, 1873, but also Cunninghamia Brown, 1826, Cryptomeria Don, 1838, and Tsuga (Endlicher, 1847) Carrière, 1855, were also important components of the Oligocene vegetation of Rębiszów. The tree layer of that mesophytic forest can probably be subdivided into the upper (canopy) and lower tree layer (understory). Based on their modern habits, we can infer that the upper canopy was composed of Cunninghamia, Cryptomeria, Liriodendron Linnaeus, 1753, Fagaceae, and Sloanea Linnaeus, 1753, whereas in the understory, trees and shrubs such as Calocedrus, Daphnogene Unger, 1850, Craigia Smith and Evans, 1921, Ostrya Scopoli, 1760, Acer Linnaeus, 1753, Laurophyllum Goeppert, 1853, Mahonia Nuttall, 1818, and probably Rosa Linnaeus, 1753 may have occurred. Lianas are modestly represented by a single leaf of Vitis Linnaeus, 1753 or Ampelopsis Michaux, 1803 and one seed impression of Ampelopsis cf. rotundata Chandler, 1925. Some authors suggest that Majanthemophyllum Weber, 1852, which is a dominant element in the Rębiszów fossil assemblage, could be a climber (Walther 1999), but this assumption lacks conclusive evidence. Palynological analysis supports the results obtained from the investigations of plant macroremains. Many fossil taxa from the spore-pollen assemblage have their counterparts in macroremains. In addition, based on the palynoflora, several elements could be added or some details could be provided. For example, we could add to the list of taxa: Fagus Linnaeus, 1753, Mastixiaceae, and Sapotaceae. Fossil macroremains of the herbaceous layer have not been found. Similarly, palynological analysis shows almost exclusively pollen of trees and shrubs. On the other hand, various spores of cryptogams, including ferns and lycopods, have been recorded. These cryptogams could be elements of the forests’ herbaceous layer, or it is probable that they grew on the lake margin.
Riparian taxa were probably confined on the shore of the water body and were not so abundant. The main component among the riparian taxa was probably Alnus Miller, 1754, presumably accompanied by Craigia, Ulmus Linnaeus, 1753, Acer, Populus Linnaeus, 1753, and Vitis vel Ampelopsis. Palynological analysis also revealed the presence of Carya Nuttall, 1818, Fraxinus Linnaeus, 1753, Pterocarya Kunth, 1824, and Salix Linnaeus, 1753. Presumably, also Daphnogene cinnamomifolia may have played a part, as it was suggested that it might tolerate riparian conditions (Kvaček and Walther 1998). On the other hand, according to Mai and Walther (1991) Populus zaddachii Heer, 1859 might be a component of a mesophytic forest as well. Species typical for swamp communities were not found among the macroremains from Rębiszów. However, as in both palynological samples taken from Rębiszów, pollen grains of Taxodium/Glyptostrobus type prevail, and we could not exclude the presence of swamp communities in the vicinity. The absence of swamp plant macroremains in this fossil assemblage reflect the absence or scarcity of temporarily inundated areas around the basin. Besides, this could also be related with the fact that volcanic floras typically represent the mesophytic climax vegetation (Walther 2005; Akhmetiev et al. 2009). Thus, the results of palynological analysis increased our knowledge on the diversity of plant communities occurring in the Rębiszów area during the Oligocene.
Moreover, the experience derived from the autecology of modern relatives of fossil plants shows that many extant species thrive in a relatively wide range of habitats (Reed 1988). Therefore, we believe that the majority of those species found in Rębiszów were not confined to one particular vegetation type (compare Worobiec and Szynkiewicz 2016), but we could contend that, generally, it was of mixed mesophytic forest type.
Palaeoclimate.—The composition of the assemblage of plant macroremains from Rębiszów, especially the presence of representatives of genera Calocedrus Kurz, 1873, Craigia Smith and Evans, 1921, Cunninghamia R. Brown, 1826, Daphnogene Unger, 1850, Laurophyllum Goeppert, 1853, Sloanea Linnaeus, 1753, and Trigonobalanopsis Kvaček and Walther, 1988 points to subtropical (sensu Belda et al. 2014) climatic conditions in this part of the Oligocene in Lower Silesia. Recent counterparts of Rębiszów vegetation could be found in the Mixed Mesophytic Forests of China (Wang 1961).
Composition of spore-pollen spectra (domination of palaeotropical and palaeotropical/warm-temperate elements) also points to warm, subtropical climatic conditions.
Comparison between the Rębiszów flora and Central European Oligocene floras and age evaluation.—Taxonomic composition clearly links the Rębiszów flora with the late Paleogene, so-called volcanic floras from the neighboring regions of the Czech Republic and SE Germany (Kvaček and Walther 2001, 2004; Walther 2005; Akhmetiev et al. 2009). With only 43 taxa, it is relatively poorly diversified in relation to other coeval fossil floras. Comparing the composition of the Rębiszów flora and some Oligocene floras from Kundratice, Seifhennersdorf, Markvartice, Suletice-Berand, and Kleinsaubernitz, several common elements can be indicated: Daphnogene cinnamomifolia forma lanceolata Kvaček and Walther, 1995, D. cinnamomifolia forma cinnamomifolia Kvaček and Walther, 1995, Laurophyllum acutimontanum Mai, 1963, Craigia bronnii (Unger, 1845) Kvaček, Bůžek, and Manchester, 1991, Sloanea artocarpites (Ettingshausen, 1869) Kvaček and Hably, 2001 (Kvaček and Walther 2001). Also, Rosa lignitum Heer, 1869, Populus zaddachii Heer, 1859, and Liriodendron haueri Ettingshausen, 1869 quite often occur in those floras. However, Mahonia Nuttall, 1818 is relatively sparse and has been documented only in some sites, including early Oligocene of Bechlejovice (Kvaček and Walther 2004) and late Oligocene of Suletice-Berand (Kvaček and Walther 1995). One of the characteristic features of the Rębiszów flora is an abundance of Calocedrus suleticensis (Brabenec, 1909) Kvaček, 1999, which has been reported so far only from the late Oligocene of Matrý, near Sebuzín (Kvaček et al. 2018) and the early Oligocene of Holý Kluk (Radoń et al. 2006) both Czechia, the early Miocene of Kymi, Greece (Kvaček 1999), the early Oligocene of Eger-Kiseged, Hungary (Kvaček and Hably 1998), and the late Oligocene of Suletice-Berand (Kvaček and Walther 1995). The occurrence of C. suleticensis is rather important. According to Kvaček and Walther (1998, 2001), it starts to appear in early/late Oligocene floras of the Nerchau-Flörsheim floristic complex. The domination of Majanthemophyllum basinerve (Rossmässler, 1840) Knobloch and Kvaček, 1996 in Rębiszów is unique in comparison to other Oligocene volcanic floras. Majanthemophyllum basinerve is only sparsely reported from Europe, it is known from the late Eocene (Staré Sedlo; Knobloch et al. 1996), and can provide only a little help in determining the age of the Rębiszów flora.
Considering the comparison mentioned above, the Rębiszów flora shows the closest relations only with some Oligocene floras. The high proportion of the laurophyllous element, and especially the presence of Calocedrus suleticensis, links the Rębiszów flora with early/late Oligocene floras of the Nerchau-Flörsheim floristic complex (especially Suletice) (Bůžek et al. 1976; Kvaček and Walther 1995, 1998, 2001). On the other hand, Majanthemophyllum basinerve has not been reported in floras of the Nerchau-Flörsheim floristic complex.
The composition of the Rębiszów palynoflora also clearly points to its Oligocene age. Crucial for the stratigraphic position of the studied flora is the presence of indicative taxa, such as Boehlensipollis hohli Krutzsch, 1962 (recorded in our samples and in the previous unpublished results by Kohlman-Adamska and Ziembińska-Tworzydło) and Cupanieidites eucalyptoides Krutzsch, 1962 (unpublished results reported by Kohlman-Adamska and Ziembińska-Tworzydło, announced here with permission of the authors). The ranges of these taxa are confined to the Rupelian–Eochattian (Krutzsch et al. 1992) or the Rupelian–early Chattian (Grabowska and Słodkowska 2003). The Rupelian has relatively rich palynological documentation due to the presence of lignites of the 5th Czempiń group of seams in the Polish Lowlands (Słodkowska 2004; Kasiński and Słodkowska 2016). In contrast, the Chattian palynofloras are very rare (Grabowska and Piwocki 1975). The Rębiszów spore-pollen assemblage is generally similar to the palynofloras from the 5th Czempiń seam (Kasiński and Słodkowska 2016). Nevertheless, the lack of some “older” taxa (for example Aglaoreidia cyclops Erdtman, 1960, and Cicatricosisporites dorogensis Potonié and Gelletich, 1933), and the presence of some “new” elements (for example Faguspollenites Raatz, 1937) rather point to the early Chattian age of the Rębiszów assemblage.
The Rębiszów flora is the first volcanic flora found in Poland and second Oligocene macroflora known from Poland. The Oligocene macroremain assemblage found in the marine Jasło limestone at Sobniów, southern Poland, consists of only 36 specimens of plant macroremains, including a few leaves (or leaflets) of dicotyledons (?Palaeocarya sp., Laurophyllum princeps (Heer, 1856) Krausel and Weyland, 1951, Theaceae, and Leguminosae (“Leguminosentypus”) type sensu Berger (1955), monocotyledons, and predominat marine algae remains (Zastawniak and Worobiec 1997). The assemblage from Sobniów (marine deep water limestone deposits), however, represents a sedimentary environment completely different to the Rębiszów flora (shallow, freshwater lake). The Rębiszów flora is definitely richer, better preserved and accompanied by rich palynoflora. The fact that the fossil assemblage from Rębiszów represents the easternmost locality among the known Oligocene volcanic floras of Central Europe makes this site unique and of great value, and complements our knowledge of the Oligocene floras of this part of Europe.
Conclusions
The fossil plant assemblage form Rębiszów is the second Oligocene macroflora known from Poland. The Oligocene age of the deposits from this site is confirmed by the radiometrically dated basalts covering fossiliferous layers (ca. 25 Ma) and the presence of pollen Boehlensipollis hohli that constrains the possible age to the Rupelian–Chattian. The lithology, preservation of the plant remains, and the composition of the plant assemblage suggest that the sedimentary environment was calm, and most probably took place in a small and shallow, arguably maar lake. Based on the floristic composition three types of vegetation could be deduced from the Rębiszów flora. Most of the fossil remains represent plants of the mesophytic forest surrounding the sedimentary basin. Riparian vegetation was probably confined to the lake shore. Species typical for swamp communities were not found among macroremains. However, in palynological samples pollen grains of Taxodium/Glyptostrobus were found, suggesting the presence of swamp communities probably in a distant neighborhood. The composition of the assemblage of plant macroremains and spore-pollen spectra point to subtropical climatic conditions. Recent counterparts of Rębiszów vegetation could be found in Mixed Mesophytic Forests of China.
In light of the sedimentary environment and floristic composition, it is clear that the Rębiszów flora corresponds to so-called volcanic floras of the early/late Oligocene. Among them, it is closest to floras of the Nerchau-Flörsheim or Kleinsaubernitz floristic complex. When considering the lithology, preservation and floristic composition, Rębiszów shows a close similarity to the early/late Oligocene fossil plant locality, Suletice-Berand (Kvaček and Walther 1995). The Rębiszów flora is the first volcanic flora found in Poland. A conclusive explanation of the position of the basalts in relation to the fossiliferous layers, sedimentary environment of deposits and detailed analysis of the palaeoclimate requires further studies of the Rębiszów floras. Considering the state of preservation of the plant remains and sedimentary environment of the newly found sites described and noted here, they may be productive for additional discoveries of well-preserved plant and animal fossils.
Acknowledgements
We thank Paweł Raczyński (University of Wrocław, Poland) who helped to arrange and participated in a first excursion to “Stary Łom” quarry in May, 2019. We also thank Izabela Sawicka (Pri Bazalt S.A. quarry manager, Rębiszów, Poland) for granting admission and friendly reception during our fieldwork. Special thanks are due to Steven Manchester (Florida Museum of Natural History, Gainesville, USA) and Torsten Utescher (University of Bonn, Bonn, Germany) for their helpful comments to the manuscript. This work was supported by the W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, Poland through the statutory funds (GW, EW).
References
Adamczyk, Z. 2008. Ewolucja wulkanizmu kenozoicznego Przedgórza Rębiszowskiego (Dolny Śląsk) oraz jej wpływ na charakter petrograficzny i jakość surowca bazaltowego. Zeszyty Naukowe Politechniki Śląskiej 1775: 1–245.
Akhmetiev, M., Walther, H., and Kvaček, Z. 2009. Mid-latitude Palaeogene floras of Eurasia bound to volcanic settings and palaeoclimatic events—experience obtained from the Far East of Russia (Sikhote-Alin’) and Central Europe (Bohemian Massif). Acta Musei Nationalis Pragae, B, Historia Naturalis 65: 61–129.
Belda, M., Holtanová, E., Halenka, T., and Kalvová, J. 2014. Climate classification revisited: from Köppen to Trewartha. Climate Research 59: 1–13. Crossref
Berger, W. 1955. Neue Ergebnisse zur Klima und Vegetationsgeschichte des Europäischen Jungtertiärs. Bericht über das Geobotanische Forschungsinstitut Rübel in Zürich (for 1954): 12–29.
Birkenmajer, K., Pécskay, Z., Grabowski, J., Lorenc, M.W., and Zagożdżon, P.P. 2011. Radiometric dating of the Tertiary volcanics in Lower Silesia, Poland. VI. K-Ar palaeomagnetic data from basaltic rocks of the West Sudety Mountains and their northern foreland. Annales Societatis Geologorum Poloniae 81: 115–131.
Bůžek, C., Holý, F., and Kvaček, Z. 1976. Tertiary flora from the volcanogenic series at Markvartice and Veselícko near Česká Kamenice (České středohorí Mts.). Sborník geologických věd. Paleontologie 18: 69–132.
Ferguson, D. 1985. The origin of leaf-assemblages—new light on an old problem. Review of Palaeobotany and Palynology 46: 117–188. Crossref
Franklin, J. 1823. Narrative of a Journey to the Shores of the Polar Sea in the Years 1819, 20, 21 and 22, with an Appendix on Various Subjects Relating to Science and Natural History Illustrated by Numerous Plates and Maps. 768 pp. John Murray, London. Crossref
Grabowska, I. and Piwocki, M. 1975. The age and origin of the Toruń clays from the vicinity of Toruń defined on the basis of palynological and lithological observations. Biuletyn Instytutu Geologicznego 284: 41–70.
Grabowska, I. and Słodkowska, B. 2003. Paleogen. In: S. Dybova-Jachowicz and A. Sadowska (eds.), Palinologia, 177–182. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków.
Jansonius, J., Hills, L.V., and Hartkopf-Fröder, C. 1998. Genera File of Fossil Spores. Supplement 12. File 5057. T2N 1N4. Special Publication, Department of Geology, University of Calgary, Calgary.
Kasiński, J.R. and Słodkowska, B. 2016. Factors controlling Cenozoic anthracogenesis in the Polish Lowlands. Geological Quarterly 60: 959–974.
Knobloch, E., Konzalova, M., and Kvaček, Z. 1996. Die obereozäne Flora der Staré Sedlo-Schichtenfolge in Böhmen (Mitteleuropa). Rozpravy Českého geologického ústavu 49: 1–260.
Krutzsch, W. 1969. Über einige stratigraphisch wichtige Longaxoner-Pollen aus dem mitteleuropäischen Alttertiär. Geologie 18: 472–487.
Krutzsch, W., Pchalek, J., and Spiegler, D. 1960. Tieferes Paläozän (?Montien) in Westbrandenburg. Proceedings of the 21st International Geological Congress, Part 6, 135–143. Kopenhagen.
Krutzsch, W., Blumenstengel, H., Kiesel, Y., and Rüffle, L. 1992. Paläobotanische Klimagliederung des Alttertiärs (Mitteleozän bis Oberoligozän) in Mitteldeutschland und das Problem der Verknüpfung mariner und kontinentaler Gliederungen (klassischer Biostratigraphien-paläobotanisch-ökologische Klimastratigraphie-Evolut. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 186: 137–253.
Kvaček, Z. 1999. An ancient Calocedrus (Cupressaceae) from the European Tertiary. Flora 194: 237–248.
Kvaček, Z. and Hably, L. 1998. New plant elements in the Tard Clay Formation from Eger-Kiseged. Acta Palaeobotanica 38: 5–23. Crossref
Kvaček, Z. and Walther, H. 1995. The Oligocene volcanic flora of Suletice-Berand near Ústí nad Labem, North Bohemia—a review. Acta Musei Nationalis Pragae, B, Historia Naturalis 50: 25–54.
Kvaček, Z. and Walther, H. 1998. The Oligocene volcanic flora of Kundratice near Litoměřice, České Středohoří volcanic complex (Czech Republic)—a review. Acta Musei Nationalis Pragae, B, Historia Naturalis 54: 1–42.
Kvaček, Z. and Walther, H. 2001. The Oligocene of Central Europa and the developement of forest vegetation in space and time based on megafossils. Palaeontographica Abteilung B 259: 125–148.
Kvaček, Z. and Walther, H. 2004. Oligocene flora of Bechlejovice at Děčín from the neovolcanic area of the České Středohoří Mountains, Czech Republic. Acta Musei Nationalis Pragae, B, Historia Naturalis 60: 9–60.
Kvaček, Z., Teodoridis, V., and Radoń, M. 2018. Review of the late Oligocene flora of Matrý near Sebuzín (České Středohoří Mts., the Czech Republic). Fossil Imprint 74: 292–316. Crossref
Mai, D.H. and Walther, H. 1991. Die oligozänen und untermiozänen Floren NW Sachsens und des Bitterfelder Raumes. Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden 38: 1–230.
Meyer, H.W., Veatch, S.W., Cook, A., Nelson, E.P., and Erslev, E.A. 2004. Field guide to the paleontology and volcanic setting of the Florissant fossil beds, Colorado. In: E.P. Nelson and E.A. Erslev (eds.), Field Trips in the Southern Rocky Mountains, USA: Geological Society of America Field Guide 5, 151–166. Geological Society of America. Crossref
Moore, P.D., Webb, J.A., and Collinson, M.E. 1991. Pollen Analysis. 216 pp. Blackwell Publishing Company, Oxford.
Nees, C.G. 1820. Horae physicae Berolinenses collectae ex symbolis virorum doctorum H. Linkii...; edicuravit Christianus Godof. Nees ab Esenbeck. xii + 123 pp. Sumtibus Adolphi Marcus, Bonnae.
Potonié, R., Thomson, P.W., and Thiergart, F. 1950. Zur Nomenklatur und Klassifikation der neogenen Sporomorphae (Pollen und Sporen). Geologishes Jahrbuch 65: 35–70.
Radoń, M., Kvaček, Z., and Walther, H. 2006. Oligocene plant megafossils and environment from the newly recovered locality of the Holý Kluk Hill near Proboštov (České Středohoří Moutains, Czech Republic). Acta Universitatis Carolinae, Geologica 47: 95–124.
Reed, P.B.J. 1988. National list of plant species that occur in wetlands: national summary. U.S. Fish and Wildlife Service, Biological Reports. 88: 1–112.
Richard, L.C-M. 1826. Commentatio botanica de Conifereis et Cycadeis, characteres genericos singulorum utriusque familiae, et figuris analyticis eximiè ab auctore ipso ad naturam delineatis ornatos complectens. Opus posthumum ab Achille Richard, filio, Med. doctore, botanices in Academiae Parisiensi professore, perfertum et in lucem editum. xv + 212 pp. Sumptibus J.G. Cottae. Ex typographia Pauli Renouard Parisini, Stutgardiae.
Słodkowska, B. 2004. Palynological studies of the Paleogene and Neogene deposits from the Pomeranian Lakeland area (NW Poland). Polish Geological Institute Special Papers 14: 1–116.
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowska, I., Słodkowska, B., Ważyńska, H., and Sadowska, A. 2009. Atlas of Pollen and Spores of the Polish Neogene. Volume 3—Angiosperms (1). 233 pp. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków.
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowska, I., Słodkowska, B., Worobiec, E., and Durska, E. 2014. Atlas of Pollen and Spores of the Polish Neogene. Volume 4—Angiosperms (2). 466 pp. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków.
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowska, I., Ważyńska, H., and Sadowska, A. 2002. Atlas of Pollen and Spores of the Polish Neogene. Volume 2. Gymnosperms. 238 pp. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków.
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowska, I., Ważyńska, H., Słodkowska, B., and Sadowska, A. 2001. Atlas of Pollen and Spores of the Polish Neogene. Volume 1. Spores. 158 pp. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków.
Thomson, P.W. and Pflug, H. 1953. Pollen und Sporen des mitteleuropäischen Tertiärs. Palaeontographica Abteilung B 94: 1–138.
Walther, H. 1999. Die Tertiärflora von Kleinsaubernitz bei Bautzen. Palaeontographica Abteilung B 249: 63–174.
Walther, H. 2005. Vulkanische Floren der südöstlichen Oberlausitz und des böhmischen Mittelgebirges – Zeugen der tertiären Waldentwicklung in Mitteleuropa. Berichte der Naturforschenden Gesellschaft Der Oberlausitz 13: 21–34.
Wang, C.-W. 1961. The Forests of China, with a Survey of Grassland and Desert Vegetation. 313 pp. Maria Moors Cabot Foundation, Cambridge.
Worobiec, E. 2014. Fossil zygospores of Zygnemataceae and other microremains of freshwater algae from two Miocene palaeosinkholes in the Opole region, SW Poland. Acta Palaeobotanica 54: 113–157. Crossref
Worobiec, G. and Szynkiewicz, A. 2016. Neogene wetland vegetation based on a leaf assemblage from the Bełchatów Lignite Mine (Central Poland). Acta Palaeobotanica 56: 441–497. Crossref
Zastawniak, E. and Worobiec, G. 1997. Szczątki roślin towarzyszące ichtiofaunie w oligoceńskich wapieniach jasielskich w Sobniowie koło Jasła (summary: Plant remains accompanying ichtyofauna in the Oligocene Jasło limestones at Sobniów near Jasło (southern Poland). Przegląd Geologiczny 45: 875–879.
Ziembińska-Tworzydło, M., Grabowska, I., Kohlman-Adamska, A., Skawińska, K., Słodkowska, B., Stuchlik, L., Sadowska, A., and Ważyńska, H. 1994. Taxonomical revision of selected pollen and spores taxa from Neogene deposits. In: L. Stuchlik (ed.), Neogene Pollen Flora of Central Europe 1. Acta Palaeobotanica (Supplementum 1): 5–30.
Acta Palaeontol. Pol. 65 (2): 273–290, 2020
https://doi.org/10.4202/app.00686.2019