
New taxa, records, and data for vesicomyid bivalves from Cenozoic strata of the North Pacific region
Steffen Kiel, Kazutaka Amano, and James L. Goedert
Kiel, S., Amano, K., and Goedert, J.L. 2023. New taxa, records, and data for vesicomyid bivalves from Cenozoic strata of the North Pacific region. Acta Palaeontologica Polonica 68 (2): 297–320.
New discoveries of Cenozoic deep-water hydrocarbon seep deposits and continued collecting at previously documented sites in the North Pacific region have resulted in additional fossils of vesicomyid bivalves and necessitate a systematic review. We report five new vesicomyid species, including four species from western Washington State, USA: Isorropodon humptulipsense sp. nov. from middle to upper Eocene strata of the Humptulips Formation and the “Siltstone of unit B”, the oldest record for Isorropodon, Pleurophopsis thieli sp. nov. from upper Eocene to lowermost Oligocene strata of the Lincoln Creek, Makah, and Pysht formations, and Pliocardia? guthrieorum sp. nov. and Squiresica plana sp. nov. from Oligocene strata of the Lincoln Creek and Pysht formations. The new species Squiresica yooni sp. nov. is from the Middle Miocene Duho Formation in South Korea. We report possibly the as-yet oldest Vesicomya from a lower Oligocene seep deposit in the Lincoln Creek Formation in western Washington. Pliocardia kawadai was previously only known from Lower to Middle Miocene strata in Japan; with our new record from the Lower to Middle Miocene Astoria Formation in western Washington, this species represents the first fossil vesicomyid species with a trans-Pacific distribution. The large and elongated Pleurophopsis chinookensis is restricted to upper Eocene strata; previous Oligocene records are shown to belong to other species.
Key words: Bivalvia, chemosymbiosis, deep-sea, Japan, South Korea, USA, Washington.
Steffen Kiel [steffen.kiel@nrm.se; ORCID: https://orcid.org/0000-0001-6281-100X ], Swedish Museum of Natural History, Department of Palaeobiology, Box 50007, 10405 Stockholm, Sweden.
Kazutaka Amano [amano@juen.ac.jp; ORCID: https://orcid.org/0000-0002-8320-8661 ], Joetsu University of Education, Joetsu 943-8512, Japan.
James L. Goedert [jamesgoedert@outlook.com; ORCID: https://orcid.org/0000-0002-0257-7255 ], Burke Museum of Natural History and Culture, University of Washington, Seattle, Washington 98195, USA.
Received 17 February 2023, accepted 26 May 2023, available online 21 June 2023.
Copyright © 2023 S. Kiel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Vesicomyid bivalves are common in deep-water habitats around the world, especially around hydrothermal vents, hydrocarbon seeps, and sunken whale carcasses (Krylova and Sahling 2010). Most of them live in symbiosis with sulfide–oxidizing bacteria, hosted in their gills, from which they derive most of their nutrition (Fisher 1990). More than 100 extant species have been described, and more than 40 fossil species, ranging back in time to the middle Eocene (Amano and Kiel 2007; Krylova and Sahling 2010; Amano et al. 2022). However, there are still gaps in our understanding of taxonomic and phylogenetic relationships, especially among the geochronologically early members of the family.
Cenozoic deep-water strata of the North Pacific region proved to be particularly rich in vesicomyid fossils, including the oldest record of the family (Amano and Kiel 2007; Hryniewicz 2022). On the east coast of the North Pacific Ocean, most vesicomyid fossils are from western Washington State, USA (e.g., Goedert and Squires 1990, 1993; Squires and Goedert 1991; Goedert and Campbell 1995), with a few additional records from California (Crickmay 1929; Woodring 1938; Squires 1991; Squires and Gring 1996). Farther north, vesicomyid fossils are known from Alaska (Kanno 1971), including the oldest record of the genus Calyptogena (Amano and Kiel 2007; Kiel and Amano 2010). Along the western coast of the North Pacific, Japan has a rich and well-investigated record of fossil vesicomyids with 19 described species (Amano et al. 2013, 2019a). One species has been reported as Calyptogena cf. elongata from South Korea (Yoon 1976a) and there are some potential records described as species of Macrocallista Meek, 1876, from Miocene rocks of eastern Kamchatka (Krishtofovich 1969).
In the present contribution, we use extensive collections made in western Washington over the last few decades to describe new vesicomyid species and to clarify the identity of those previously described. Furthermore, the identity of the South Korean “Calyptogena cf. elongata” is clarified, the largest specimens of the Japanese species Hubertschenckia ezoensis are illustrated, and the biogeographic history and relationships of these species are discussed.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:251DFB5C-149C-4629-B960-7128B1B34AE8
Institutional abbreviations.—CSUN, California State University at Northridge, California, USA; LACM and LACMIP, Natural History Museum of Los Angeles County, Invertebrate Paleontology, Los Angeles, California, USA; NRM, Swedish Museum of Natural History, Stockholm, Sweden; UWBM, University of Washington, Burke Museum of Natural History and Culture, Seattle, Washington, USA.
Other abbreviations.—aams, anterior adductor muscle scar; H, height; L, length; LV, left valve; pams, posterior adductor muscle scar; RV, right valve; W, width.
Material and methods
Specimens were mechanically extracted from the rock matrix using hammers and chisels, and pneumatic fossil preparation tools. Casts were made using colored silicone rubber. Some fossil specimens were used to produce polished surfaces. Specimens were coated with ammonium chloride prior to photography, unless noted otherwise. One articulated specimen was scanned with an Xradia Versa 520 x-ray microscope at the Stockholm University Brain Imaging Center (SUBIC). Most specimens are deposited at the Swedish Museum of Natural History (NRM), some are stored in the Natural History Museum of Los Angeles County (LACMIP) and the Burke Museum, University of Washington (UWBM).
Localities
Washington State, USA.—The material from Washington State, USA (Fig. 1), was collected by JLG over the last decades; some of it jointly with SK. GPS coordinates are given using the WGS84 datum (World Geodetic System). Standard locality descriptions, based upon US Geological Survey topographic maps, refer to section, township, and range according to the USA Public Land Survey System.
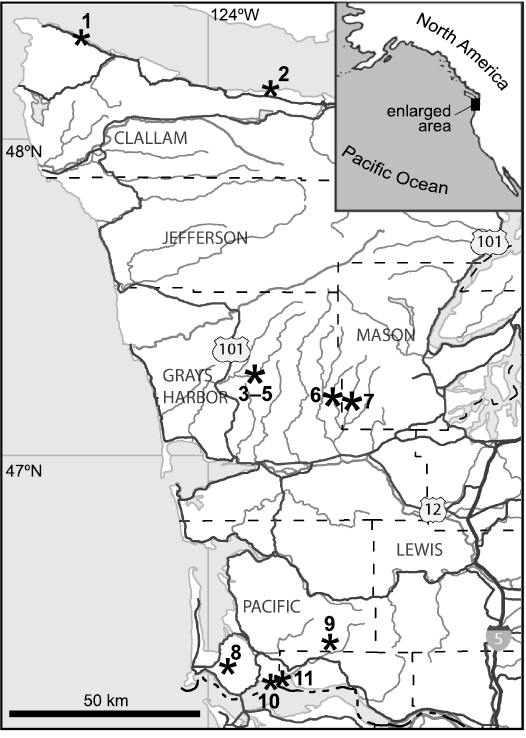
Fig. 1. Map showing localities in western Washington State, USA. 1, coastal sites between Rasmussen Creek and Jansen Creek (Makah Formation); 2, Whiskey Creek site (Pysht Formation); 3–5, sites LACMIP loc. 12385, CSUN loc. 1583, “East Fork Bridge site” (Humptulips Formation); 6, UWBM loc. B7452 in the Canyon River (Lincoln Creek Formation); 7, SR2 and SR4, Satsop River (Lincoln Creek Formation); 8, Bear River seep deposit; 9, West Fork of Grays River (Siltstone of Unit B); 10, Frankfort (Astoria Formation); 11, sites around Rocky Point (Astoria Formation).
Astoria Formation: The material reported here was collected from the Lower to Middle Miocene, thin-bedded mudstone unit of the Astoria Formation (“Tasu” of Wolfe and McKee 1968; Unit III of Wolfe and McKee 1972; Bald Ridge unit of Wells 1989) cropping out along the north shore of the Columbia River in Pacific County. Fossils were found in float seep limestone and concretions (Amano and Kiel 2007; Kuechler et al. 2012) found on intertidal platforms at three sites between the ghost town of Frankfort in the west and the mouth of Deep River in the east. These include the previously published site LACMIP 6132 just to the east of Frankfort (Kiel 2010b) and two new sites just east and west of Rocky Point. The concretions contain mostly the vesicomyid Pliocardia kawadai and are accompanied by small-sized specimens (up to 40 mm in length) of the thyasirid Conchocele spp. and a few indeterminable gastropods. Paleodepths for these strata are poorly constrained, ranging 16–650 m or possibly deeper based on foraminiferans (e.g., Wolfe and McKee 1968, 1972), and bottom water temperatures may have been near zero (Qu et al. 2017). Deposition was in part by turbidity flows (Wolfe and McKee 1972). Other fossils are generally rare, but include a few crustaceans and a variety of terrestrial plants (Berglund and Goedert 1992), propeamussiid bivalves coined “mud pectens” (Wolfe and McKee 1972), pteropod gastropods (Squires 1989), and a marine mammal (Peredo et al. 2018). The seep deposits are not common but are conspicuous because they contain abundant, densely packed, large and mostly articulated bivalves.
Bear River seep deposit: We document further specimens of Pleurophopsis chinookensis from its type locality, a well-described, upper Eocene seep deposit at Bear River (Goedert and Squires 1990; Squires and Goedert 1991; Goedert and Benham 2003; Kiel 2006; Kiel and Amano 2013). There is much more research to be done on this site but currently, Weyerhaeuser Company owning the land does not allow access.
Humptulips Formation: This formation consists mainly of upper middle Eocene deep-water mudstone (Rau 1984, 1986; Prothero et al. 2001; Goedert et al. 2013). The only molluscan fossils not found in seep deposits were rare wood-fall taxa (Kiel 2008b). The material reported here comes from four different blocks of seep carbonate found along, or in the vicinity of, the Humptulips River: (i) LACMIP locality 12385, a large seep deposit with a diverse fauna (Goedert and Squires 1990; Squires and Goedert 1991; Kiel 2006; Kiel and Amano 2013; Hryniewicz et al. 2017); (ii) CSUN loc. 1583, a seep deposit also known as the “Raui site” for its most conspicuous gastropod, Humptulipsia raui (Goedert and Kaler, 1996) (Saul et al. 1996; Goedert and Kaler 1996; Squires and Goedert 1996; Kiel 2008a; Jakubowicz et al. 2020); (iii) a seep deposit named “East Fork Bridge site” in a recent isotopic study (Jakubowicz et al. 2020), with a fauna consisting mainly of an as-yet unidentified lucinid bivalve and the vesicomyid reported herein; (iv) an as-yet undescribed block found as float on the same gravel bar (at 47.2469°N, 123.8409°W) as the previous deposit, but with a nodular, micritic lithology, with narrow vugs filled by calcite spar, and a diverse fauna including worm tubes, thyasirid bivalves, and the herein reported vesicomyid. Much more work remains to be done at these sites, but currently Rayonier Corporation owning the land no longer allows access.
Lincoln Creek Formation: This formation is composed of the upper Eocene to lowest Miocene strata, mainly deep-water deposits, and crops out between the southern foothills of the Olympic Mountains and the Columbia River. The material reported here is from four seep deposits, from the Oligocene part of the formation, exposed along the Canyon and Satsop rivers as mapped by Rau (1966). The in situ deposits from the Satsop River were previously described as SR2 (= UWBM loc. B6702; earliest Oligocene age) and SR4 (= LACMIP loc. 17426; late Oligocene age) (Peckmann et al. 2002). Those from the Canyon River were blocks found as float on gravel bars along the river, among them several from UWBM loc. B7452 that include specimens of Bathymodiolus inouei Amano and Jenkins, 2011b, and Lucinoma sp. (Kiel and Amano 2013) in addition to the vesicomyid reported herein; the other was found at about 47.267522° N, 123.526238° W and is probably of late Oligocene age. Some of the Satsop and Canyon river in situ seep deposits are very small in size (e.g., Peckmann et al. 2002; Kiel 2006; Yamaguchi et al. 2016) and because the rivers generally flow perpendicular to strike and dip with the oldest strata exposed upstream, the ages given for deposits found as float should be regarded as minimums because there is the possibility they are older.
Makah Formation: The Makah Formation crops out along the shore of the Strait of Juan de Fuca in northwestern Washington (Snavely et al. 1980). Between Whiskey Creek and the Lyre River are extensive outcrops of deep-water siltstone with abundant concretions, originally mapped (Brown and Gower 1958; Tabor and Cady 1978) as the middle member of the Twin River Formation. The middle member of the Twin River Formation was later named the Makah Formation by Snavely et al. (1980), although they do not specifically mention the strata exposed between Whiskey Creek and the Lyre River. These rocks yield a crab, Macroacaena alseana (Rathbun, 1932), only found in upper Eocene rocks in Oregon and Washington (Feldman 1989; Tucker 1998), but except for foraminifers (Rau 1964), a marine bird (e.g., Goedert and Cornish 2002), and a terrestrial plant (Crabtree and Miller 1989), other fossils have not yet been described. Rocks between the Lyre River and Whiskey Creek were later mapped in error as the Pysht Formation by Nesbitt et al. (2010) (Elizabeth A. Nesbitt, personal communication to JLG, 2011). Seep carbonates are present as float along these coastal outcrops and include thyasirids and possibly a vesicomyid (discussed herein), but these carbonates have not yet been fully investigated.
Farther to the west, seep carbonate from the Shipwreck Point area west of the Sekiu River contains vesicomyids (Goedert and Campbell 1995) and these are further discussed herein. Two float seep carbonate blocks containing vesicomyids were sampled from coastal outcrops of the uppermost Eocene or lowermost Oligocene Jansen Creek Member between the mouth of Rasmussen Creek eastward to a point about 500 m southeast of Jansen Creek. The Jansen Creek Member is an olistostrome (Snavely et al. 1980) and contains both deep and shallow water marine deposits. This olistostrome contains a complex of seep carbonates that are either in situ within large (up to 100 m long; Snavely et al. 1980) blocks of marine sediment, allochthonous (Goedert and Campbell 1995), or perhaps even in part precipitated within and on top of the rubble pile after submarine slumping, but this awaits further analyses and detailed mapping. Goedert and Squires (1993) reported vesicomyids from a turbidite deposit belonging to the Jansen Creek Member. The Jansen Creek Member seep carbonates yield a distinct species of Bathymodiolus that differs from other known fossil bathymodiolins in western Washington (Kiel and Amano 2013). Thyasirids were reported from Jansen Creek Member seep deposits (Hryniewicz et al. 2017) as were provannid gastropods (Squires 1995). A recently described seep site has abundant echinoids (Müller et al. 2023).
Pysht Formation: Sampling sites for this study include the highly fossiliferous, upper Eocene Whiskey Creek seep deposit (Goedert et al. 2003; Peckmann et al. 2003; Hryniewicz et al. 2017) probably from the basal part of the formation. Additional specimens of a previously published vesicomyid from an lower upper Oligocene seep deposit at LACMIP loc. 15621, in the lower part of the Pysht Formation west of the mouth of Murdock Creek, Clallam County (Goedert and Squires 1993) are re-evaluated.
Siltstone of unit B: Several float carbonate concretions and an in situ site from this upper Eocene rock unit (Wolfe and McKee 1968) were collected along West Fork of Grays River, upstream from the mouth of Beaver Creek in Pacific County. Although this siltstone is locally highly fossiliferous, the fossils have not yet been described in detail (i.e., Wolfe and McKee 1968, 1972; Kiel 2010a). Limacinid pteropods were documented by Garvie et al. (2020). A deep water crab, Macroacaena alseana from this rock unit was discussed by Feldman (1989). This crab fossil is also found in the upper Eocene Hoko River and Makah formations (between Whiskey Creek and the Lyre River, see discussion above) in northwestern Washington and the upper Eocene Nestucca Formation in Oregon (Feldman 1989; Tucker 1998). Based on benthic foraminifers, the deposition of this siltstone unit was estimated to have occurred at depths of ca. 150–750 m (Wolfe and McKee 1968, 1972). In addition to the vesicomyids reported herein, an unidentified thyasirid was reported from these seep deposits (Hryniewicz et al. 2017).
South Korea.—Duho Formation: The Middle Miocene Duho Formation of the Euichang Group in the Pohang Basin, is exposed in and around Pohang City, near the southeast coast of Korea (Yun 1986). It is composed of deep-water mudstone and has produced a diverse fauna of deep-water taxa, including various mollusks (Yoon 1976a, b), brittlestars (Seong et al. 2009; Ishida et al. 2022), sharks (Yun 2020, 2021; Malyshkina et al. 2022), and a pleuronectiform flatfish (Ko 2016). The specimens documented here were collected by the late Professor Sun Yoon at a hill-side cut for the foundation of a house, about 400 m southeast of Goedong-dong, Daesong-myeon, Yeongil-gun, South Korea. This is loc. 18 of Yoon (1976b).
Japan.—Poronai Formation: Five specimens of Hubertschenckia ezoensis were collected by the late Saburo Kanno, who was an Emeritus Professor at the University of Tsukuba and Joetsu University of Education, from the upper Eocene Poronai Formation. The exact locality is unknown but was most likely either in Yubari City or Mikasa City, in central Hokkaido.
Systematic palaeontology
Class Bivalvia Linnaeus, 1758
Subclass Heteroconchia Gray, 1854
Order Venerida Gray, 1854
Family Vesicomyidae Dall and Simpson, 1901
Subfamily Vesicomyinae Dall and Simpson, 1901
Genus Vesicomya Dall, 1886
Type species: Callocardia atlantica Smith, 1885, by original designation; Recent, North Atlantic Ocean.
Vesicomya? sp. 1
Fig. 2A–G.
Material.—One articulated specimen (NRM Mo 204811) from the SR2 seep deposit of Peckmann et al. (2002) [= UWBM loc. B6702], in the lower Oligocene part of the Lincoln Creek Formation, western Washington State, USA.
Dimensions (in mm).—NRM Mo 204811: L = 17.0, H = 14.5, W = 12.8.
Description.—Large for genus, well inflated; umbones large, blunt, strongly raised, displaced anteriorly to approximately 30% of total shell length; anterior margin truncate, acutely rounded; ventral margin very slightly convex; posterodorsal margin gently sloping, accompanied by submarginal ridge and weak sulcus to its anterior; lunular incision short, heart-shaped; escutcheon short, lanceolate; surface with irregular growth lines. Hinge plate narrow, with teeth oriented subparallel to dorsal margin, as is typical for Vesicomya.
Vesicomya? sp. 2
Fig. 2H–K.
Material.—One articulated specimen and two isolated RV (NRM Mo 204701–03) from the SR4 seep deposit of Peckmann et al. (2002) [= UWBM loc. 17426], in the upper Oligocene part of the Lincoln Creek Formation, western Washington State, USA.
Dimensions (in mm).—NRM Mo 204701: L = 6.5, H = 5.0; NRM Mo 204702: L = 5.6; NRM Mo 204703: L = 4.3.
Description.—Average size for genus, well inflated; umbones subcentral, raised, prosogyrate; posterior margin angular due to weak ridge from umbones to posteroventral corner; surface with irregular growth lines; lunular incision deeply incised, broad, short.

Fig. 2. The small vesicomyid bivalves Vesicomya? sp. 1 and sp. 2, from Oligocene seep deposits in the Lincoln Creek Formation, western Washington State, USA. A. Vesicomya? sp. 1 from the early Oligocene SR2 site, Satsop River. NRM Mo 204811, left side view (A1) and dorsal view on LV and dorsal side of actual specimen (A2); A3, A4, X-ray section through the hinge area, with our interpretations; A5–A7, 3D renderings of the specimen. B, C. Vesicomya? sp. 2 from the late Oligocene SR4 site, Satsop River. B. NRM Mo 204701, disarticulated specimen, view on RV with naticid drill hole on the umbo (B1) and on the LV showing the truncate posterior margin (B2). C. NRM Mo 204702, RV in anterior view, showing inflation and lunular incision (C1) and dorsal view, showing lunular incision (C2).
Remarks.—Vesicomya? sp. 1 and sp. 2 reported here are potentially the oldest fossil records of the Vesicomya (sensu stricto). Unfortunately, in neither of the two species the pallial line or muscle scars are visible, thus some doubt about their assignments to Vesicomya remain (cf., Cosel and Salas 2001; Krylova et al. 2018). The only other fossil species that can confidently be assigned to Vesicomya sensu stricto is Vesicomya margotae Beets, 1953, from the Upper Miocene of Indonesia (Buton Island, Beets 1953) and the Pliocene of the Philippines (Leyte Island, Kiel et al. 2020a).
A lower Oligocene record of a putative Vesicomya sp. was published by Goedert and Campbell (1995: 26, fig. 3), a single valve approximately 12 mm long with a convexity of 4.8 mm, from the Makah Formation. However, that specimen is more elongate than a typical Vesicomya and the hinge is not observable in the only known specimen. If the Makah Formation fossil is indeed a vesicomyid, it is more likely a juvenile Squiresica or Isorropodon.
Subfamily Pliocardiinae Woodring, 1925
Genus Isorropodon Sturany, 1896
Type species: Isorropodon perplexum Sturany, 1896, by monotypy; Recent, eastern Mediterranean Sea.
Isorropodon humptulipsense sp. nov.
Fig. 3.
1996 Vesicomya sp.; Squires and Goedert 1996: 270.
1996 Vesicomya sp.; Goedert and Kaler 1996: 67.
2007 Archivesica cf. tschudi (Olsson, 1931); Amano and Kiel 2007: 282, figs. 24–29.
ZooBank LCID: urn:lsid:zoobank.org:act:A03AC3DA-5702-4AA3-8099-D69460782E1F.
Etymology: After the Humptulips Formation, the type horizon of the species.
Type material: Holotype: NRM Mo 204723 from CSUN loc. 1583, an internal mold showing muscle scars and pallial line. Paratypes: NRM Mo 204722 from CSUN loc. 1583; NRM Mo 204724, 25 from LACMIP loc. 12385; NRM Mo 204733, from as-yet undescribed boulders found near the East Fork Bridge site (reported here as site (iv) for the Humptulips Formation). Eocene, Washington State, USA.
Type locality: The seep deposit at CSUN loc. 1583, Washington State, USA.
Type horizon: Humptulips Formation, Eocene.
Material.—39 specimens; 29 specimens from Humptulips Formation: two specimens from CSUN loc. 1583 (NRM Mo 204722, 23), 9 specimens from LACMIP loc. 12385 (NRM Mo 204724–32), 10 specimens from a block found near the East Fork Bridge site, (NRM Mo 204733–42), 8 specimens from the East Fork Bridge site (NRM Mo 204753–60), and 10 specimens from Siltstone of unit B, Grays River mudstone (NRM Mo 204743–52). Eocene, Washington State, USA.
Dimensions.—See Table 1.
Table 1. Measurements (in mm) of Isorropodon humptulipsense sp. nov. ** holotype; * paratype; na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
Locality |
|
NRM Mo 204722* |
20.6 |
15.3 |
11.8 |
CSUN loc. 1583 |
|
NRM Mo 204723** |
31.0 |
23.8 |
15.8 |
CSUN loc. 1583 |
|
NRM Mo 204724* |
11.0 |
9.6 |
7.0 |
LACMIP loc. 12385 |
|
NRM Mo 204725* |
13.8 |
11.4 |
11.7 |
LACMIP loc. 12385 |
|
NRM Mo 204726 |
18.3 |
11.9 |
na |
LACMIP loc. 12385 |
|
NRM Mo 204727 |
15.2 |
12.0 |
7.1 |
LACMIP loc. 12385 |
|
NRM Mo 204728 |
15.3 |
11.7 |
9.0 |
LACMIP loc. 12385 |
|
NRM Mo 204729 |
12.8 |
10.8 |
7.8 |
LACMIP loc. 12385 |
|
NRM Mo 204730 |
12.1 |
8.5 |
5.7 |
LACMIP loc. 12385 |
|
NRM Mo 204731 |
11.1 |
8.8 |
6.0 |
LACMIP loc. 12385 |
|
NRM Mo 204732 |
8.4 |
6.8 |
4.5 |
LACMIP loc. 12385 |
|
NRM Mo 204733* |
22.8 |
18 |
13.7 |
near East Fork Bridge |
|
NRM Mo 204734 |
34.6 |
24.7 |
na |
near East Fork Bridge |
|
NRM Mo 204735 |
17.0 |
13.5 |
11.0 |
near East Fork Bridge |
|
NRM Mo 204737 |
16.2 |
12.0 |
na |
near East Fork Bridge |
|
NRM Mo 204739 |
12.6 |
12.3 |
na |
near East Fork Bridge |
|
NRM Mo 204740 |
11.2 |
11.7 |
na |
near East Fork Bridge |
|
NRM Mo 204741 |
10.6 |
7.6 |
na |
near East Fork Bridge |
|
NRM Mo 204743 |
30.3 |
20.0 |
na |
Grays River |
|
NRM Mo 204744 |
23.5 |
18.8 |
na |
Grays River |
|
NRM Mo 204745 |
32.0 |
25.0 |
19.6 |
Grays River |
|
NRM Mo 204746 |
30.8 |
22.8 |
19.0 |
Grays River |
|
NRM Mo 204747 |
25.0 |
20.8 |
17.8 |
Grays River |
|
NRM Mo 204748 |
25.0 |
19.0 |
15.2 |
Grays River |
|
NRM Mo 204749 |
24.5 |
19.5 |
na |
Grays River |
|
NRM Mo 204750 |
25.5 |
22.0 |
13.3 |
Grays River |
|
NRM Mo 204751 |
na |
21.5 |
16.6 |
Grays River |
|
NRM Mo 204753 |
na |
21.4 |
15.0 |
East Fork Bridge site |
|
NRM Mo 204755 |
19.5 |
14.6 |
na |
East Fork Bridge site |
|
NRM Mo 204756 |
19.7 |
16.0 |
12.0 |
East Fork Bridge site |
Diagnosis.—Well-inflated shells reaching nearly 35 mm in length, umbones elevated, prosogyrate; lunule deeply incised; broad escutcheon bordered by blunt ridges; pallial line distinct, anteriorly narrow, posteriorly broadening, curving upward, either evenly or with slight indentation, no pallial sinus; RV hinge with elongate cardinal teeth 1 and 3a running parallel to shell margin, 3b pointing posteroventrally; LV hinge with cardinal teeth 2a and 2b elongate, parallel to shell margin, 2b with small posterior peg.
Description.—Shells reaching nearly 35 mm in length and 25 mm in height, inflated, umbones elevated, strongly prosogyrate, at anterior 16–18% of shell length; lunule deeply incised, elongate-heart shaped, escutcheon broad, demarcated by blunt ridges; anterior adductor muscle scar bean-shaped, fused with pedal retractor scar; pallial line entire, distinct, anteriorly narrow, broadening posteriorly, starting at posterodorsal corner of anterior adductor muscle scar, running close and parallel to ventral shell margin, posteriorly curving upward, either evenly or with slight indentation, to join posterior adductor muscle scar at its base; no pallial sinus; distinct interior ridge running from umbones to ventral side of posterior adductor muscle scar; hinge with three cardinals in each valve; RV with elongate cardinal teeth 1 and 3a running parallel to shell margin, 3b bifid with raised edges, pointing posteroventrally; in LV cardinal teeth 2a and 2b elongate, parallel to shell margin, 2b with small posterior peg, cardinal tooth 4b thin, short.
Remarks.—For a more detailed description see Amano and Kiel (2007). The small, circular and deep subumbonal pit figured earlier (Amano and Kiel 2007: figs. 24, 25) was likely an artifact because we have not seen it in additional specimens. Small-sized specimens reaching 2.5 mm in length were reported as “unidentified vesicomyid bivalve” from the middle Eocene Weatherwax seep deposit of the Humptulips Formation (Hybertsen and Kiel 2018); these oval specimens have an almost Vesicomya-like hinge dentition and might be juveniles of Isorropodon humptulipsense sp. nov.
The pallial line of Isorropodon humptulipsense shows some variation in thickness and in its posterior-most part before reaching the posterior adductor muscle scar. In some specimens the pallial line curves evenly upward into the posterior adductor muscle scar (Fig. 3B, G), in others, it shows a slight indentation (but not a sinus) on its way upward (Fig. 3F). Other species of Isorropodon also show some variation of this feature. In I. megadesmum Oliver, Rodrigues, and Cunha, 2011, the pallial line joins the posterior adductor muscle scar after a sinuous swing (Oliver et al. 2011: fig. 9C); in I. bigoti Cosel and Salas, 2001 (their fig. 55) and I. curtum Cosel and Salas, 2001 (their fig. 59) both join after a straight section followed by an angulation of the pallial line, and this angulation can point either posteriorly (I. bigoti) or anteriorly (I. curtum). In I. nyeggaense Krylova, Gebruk, Portnova, Todt, and Haflidason, 2011 (Krylova et al. 2011: figs. 6D, 7D), the pallial line just has a straight section before reaching the posterior adductor muscle scar.
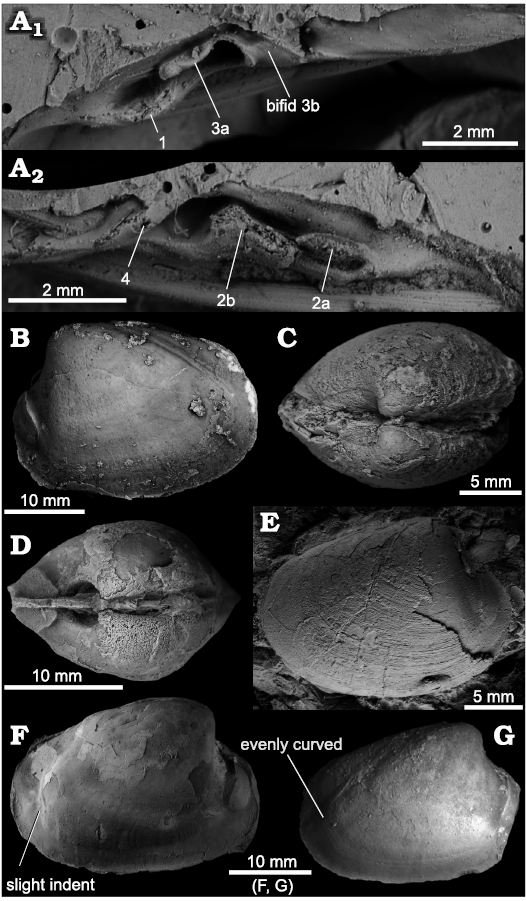
Fig. 3. The vesicomyid bivalve Isorropodon humptulipsense sp. nov., from middle to upper Eocene strata in western Washington State, USA. A. Rubber cast of the paratype, NRM Mo 204722, Humptulips Formation, CSUN loc. 1583, hinge area, RV (A1), LV (A2). B. Holotype, NRM Mo 204723, Humptulips Formation, CSUN loc. 1583, internal mold, view on LV. C. NRM Mo 204724, Humptulips Formation, LACMIP loc. 12385, articulated specimen, view on anterodorsal margin showing lunular incision. D. Paratype, NRM Mo 204733, Humptulips Formation, near East Fork Bridge site, view on anterodorsal margin and anterior adductor muscle scars. E. Paratype, NRM Mo 204744, Siltstone Unit B along Grays River, specimen with partially preserved shell. F. Paratype, NRM Mo 204743, Siltstone Unit B along Grays River, internal mold showing pallial line with slight indentation on RV. G. Internal mold, USNM 534952, Humptulips Formation, CSUN loc. 1583, view on RV with pallial line curving evenly into posterior adductor muscle scar (re-illustrated from Amano and Kiel 2007).
Compared to other species of Isorropodon, I. humptulipsense is relatively large and has a distinct lunular incision. Isorropodon bigoti reaches only 21 mm in length, has a straighter ventral margin, and an indistinct lunular incision. The type species I. perplexum reaches a maximum of 15 mm in length, its hinge teeth are arranged mostly parallel to each other in contrast to the radiating teeth of I. humptulipsense, and it lacks a lunular incision. Isorropodon curtum is not only much smaller, it also has a shorter shell. Of similar size as I. humptulipsense is I. striatum, but its hinge teeth are arranged in a parallel fashion and its lunular incision is indistinct. Isorropodon megadesmum reaches only 12 mm in length and has an indistinct lunular incision, and I. nyeggaense: is up to 21 mm long, is less inflated than I. humptulipsense, and has an indistinct lunular incision. Isorropodon arguinense Hoffman, Cosel, and Freiwald, 2019, is smaller and much more elongate than I. humptulipsense (i.e., Hoffman et al. 2019). The Miocene I. frankfortense Kiel and Amano, 2007 (originally introduced as I. frankfortensis) from the Astoria Formation in southwestern Washington State is much smaller and has a more convex ventral margin than I. humptulipsense, and has the hinge teeth arranged in a parallel fashion (Amano and Kiel 2007).
Somewhat similar is Cytherocardia cytheroides (Mayer, 1868), type of the little-known genus Cytherocardia Sacco, 1900, from Miocene of Italy (Mayer 1868; Sacco 1900), which has been suggested to be a member of Vesicomyidae (Janssen and Krylova 2012). The shells are oval with strong, elevated, prosogyrate umbones, thus somewhat have a similar shape to Isorropodon humptulipsense (i.e., Janssen and Krylova 2012: pl. 5: 19). However, the taxonomically important dentition of the right valve of C. cytheroides remains unknown, as are internal features such as pallial line and muscle scars. We find the allocation of Cytherocardia to the Vesicomyidae doubtful, and the lack of a lunular incision in C. cytheroides clearly distinguishes it from Isorropodon humptulipsense.
Stratigraphic and geographic range.—Seep carbonate deposits within the middle Eocene Humptulips Formation and upper Eocene Siltstone of unit B, Washington State, USA.
Genus Pliocardia Woodring, 1925
Type species: Anomalocardia bowdeniana Dall, 1903, by original designation; Pliocene, Bowden Formation, Jamaica (Dall 1903).
Pliocardia kawadai (Aoki, 1954)
Figs. 4, 5.
1954 Lamelliconcha kawadai sp. nov.; Aoki 1954: 36–37, pl. 2: 1–13, 22.
1962 Vesicomya kawadai (Aoki); Kamada 1962: 88–89, pl. 8: 2a–b.
2001 Vesicomya kawadai (Aoki); Amano et al. 2001: 192, figs. 3–5, 8–11.
2007 Vesicomya kawadai (Aoki); Amano et al. 2007: figs. 3D, E, G, J.
2012 Pliocardia kawadai (Aoki); Amano and Kiel 2012: 80–83, figs. 2–4, 6, 7, 9–12.
2019 Pliocardia kawadai (Aoki); Amano et al. 2019b: fig. 4 C, E, J–O.
2022 Pliocardia kawadai (Aoki); Amano et al. 2022: fig. 10.9 f, g.
Material.—Eight specimens from LACM 6132 (NRM Mo 204715–18, 21, LACMIP 6132.2–6132.4); seven specimens from northeast of Rocky Point (NRM Mo 204709–13, 19, 20); 27 mostly fragmentary specimens from west of Rocky Point (NRM Mo 204714 [1 specimen], 204812 [26 specimens]). Miocene, Astoria Formation, Bald Ridge unit, north shore of the Columbia River, Pacific County, Washington State, USA.
Dimensions.—See Table 2.
Table 2. Measurements (in mm) of Pliocardia kawadai (Aoki, 1954). na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
Locality |
|
NRM Mo 204709 |
39.7 |
25.0 |
na |
Frankfort |
|
NRM Mo 204710 |
27.4 |
19.8 |
na |
Frankfort |
|
NRM Mo 204711 |
49.8 |
39.6 |
na |
Frankfort |
|
NRM Mo 204712 |
36.0 |
30.4 |
na |
Frankfort |
|
NRM Mo 204714 |
40.0 |
24.5 |
18.4 |
Rocky Point |
|
NRM Mo 204719 |
34.0 |
20.5 |
18.4 |
Frankfort |
|
NRM Mo 204720 |
31.7 |
22.4 |
17.4 |
Frankfort |
|
NRM Mo 204715 |
37.0 |
22.7 |
18.8 |
LACM 6132 |
|
NRM Mo 204716 |
34.6 |
25.2 |
na |
LACM 6132 |
|
NRM Mo 204717 |
36.0 |
22.3 |
19.0 |
LACM 6132 |
|
NRM Mo 204718 |
34.0 |
32.5 |
27.3 |
LACM 6132 |
|
NRM Mo 204721 |
31.6 |
22.8 |
20.0 |
LACM 6132 |
|
LACMIP 6132.2 |
39.5 |
24.7 |
23.3 |
LACM 6132 |
|
LACMIP 6132.3 |
45.0 |
33.8 |
25.2 |
LACM 6132 |
|
LACMIP 6132.4 |
37.0 |
23.2 |
na |
LACM 6132 |
Remarks.—The specimens of Pliocardia kawadai from the Lower to Middle Miocene Columbia River seep deposits resemble those from the Lower Miocene Honya Formation of Joban coalfield in Japan in every aspect (Aoki 1954; Kamada 1962). The topotype specimens subsequently reported (Amano and Kiel 2012) show no discernable difference in hinge structure compared with the specimens from Washington, though they appear slightly less inflated and have sharper (more angular) posterodorsal ridges. The Japanese specimens of Pliocardia kawadai reach 43.1 mm in length (i.e., Amano et al. 2019b), which is similar to the average size of the specimens from Washington, with the exception of one specimen that is nearly 50 mm long.
As pointed out previously (Amano and Kiel 2012), the most similar extant species is the West Pacific P. crenulomarginata (Okutani, Kojima, and Iwasaki, 2002), which differs from P. kawadai mainly by having less prosogyrate umbones and fine crenulations on the posterior margin.
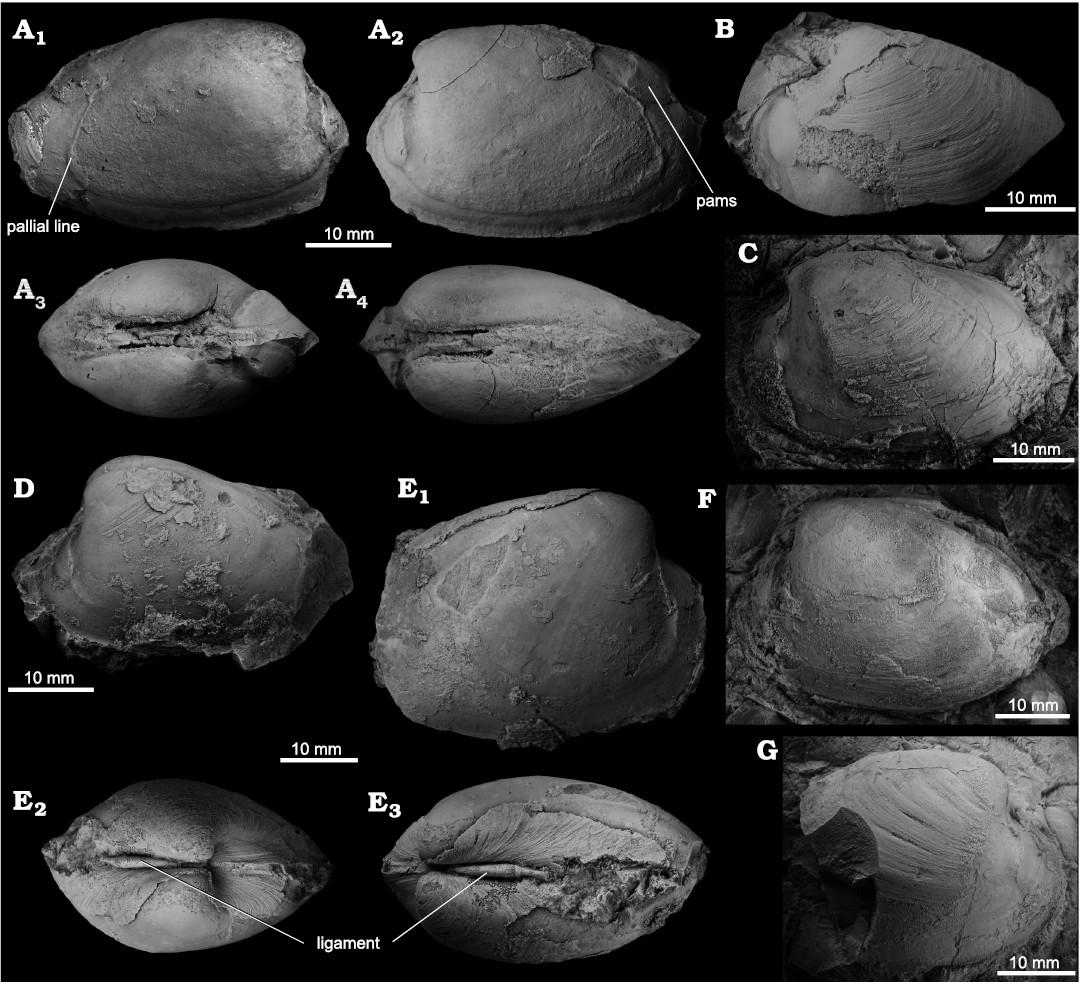
Fig. 4. The vesicomyid bivalve Pliocardia kawadai (Aoki, 1954), from Middle Miocene seep deposits of the Astoria Formation in western Washington State, USA. A. NRM Mo 204714, Rocky Point, internal mold showing muscle scars and pallial line. B. NRM Mo 204719, Frankfort, specimen with partially preserved shell (LV) showing the fine, irregular growth lines and the anterior adductor muscle scar. C. NRM Mo 204709, Frankfort, mostly an internal mold, showing the anterior adductor muscle scar and the posterior ridge. D. NRM Mo 204720, Frankfort, mostly an internal mold, note naticid drill hole on posterodorsal margin. E. LACMIP 6132.2, LACMIP loc. 6132, specimen with partially preserved shell; view of RV showing muscle scars (E1); anterodorsal view showing elongate lunular incision (E2); posterodorsal view showing ligament (E3). F. NRM Mo 204711, Frankfort, internal mold of left valve, showing pallial line and posterior adductor muscle scar. G. NRM Mo 204712, Frankfort, specimen with partially preserved shell (RV) showing the fine, irregular growth lines and the anterior adductor muscle scar.
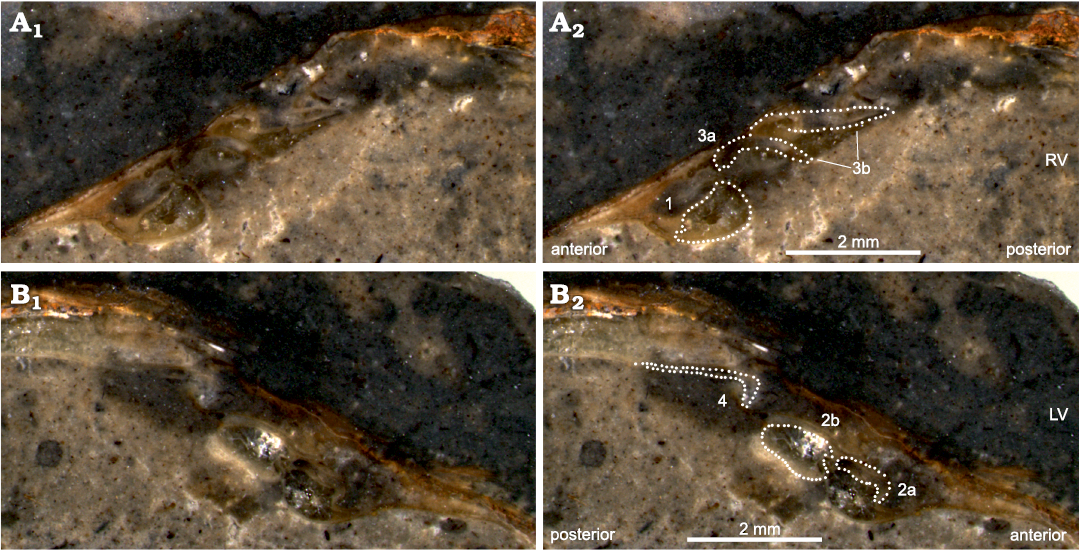
Fig. 5. The vesicomyid bivalve Pliocardia kawadai (Aoki, 1954), from Middle Miocene seep deposit at Frankfort of the Astoria Formation in western Washington State, USA. NRM Mo 204713, polished sections of the hinge area in the RV (A) and LV (B). Photographs (A1, B1) and their interpretation (A2, B2).
Stratigraphic and geographic range.—Lower–Middle Miocene of Hokkaido and Honshu, Japan, and seep deposit in southwestern Washington State, USA.
Pliocardia? guthrieorum sp. nov.
Fig. 6.
ZooBank LCID: urn:lsid:zoobank.org:act:EEDC380D-91CD-44E9-A722-6096971A8D9C.
Etymology: In honor of Lloyd S. and Kay Guthrie (Olympia, USA), for their help over several fieldtrips that produced some of these specimens.
Type material: Holotype: NRM Mo 204785 (Fig. 6G); paratypes: NRM Mo 204779–82, 204784, 204813, 14, UWBM IP 117930, 31. All specimens from the type locality and horizon.
Type locality: UWBM loc. B7452 in the Canyon River in western Washington State, USA.
Type horizon: Lincoln Creek Formation, Oligocene.
Material.—73 specimens from UWBM loc. B7452 (NRM Mo 204779–89, 92, 93, 204813, 14, UWBM IP 117930–31), and four from UWBM loc. B6702 (NRM Mo 204790), all Lincoln Creek Formation. One specimen from the Makah Formation, west of Whiskey Creek (NRM Mo 204791). Oligocene, Washington State, USA.
Dimensions.—See Table 3.
Table 3. Measurements (in mm) of Pliocardia? guthrieorum sp. nov. ** holotype; * paratype; na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
Locality |
|
NRM Mo 204779* |
48.6 |
37.5 |
22.7 |
UWBM loc. B7452 |
|
NRM Mo 204780* |
37.0 |
24.0 |
14.8 |
UWBM loc. B7452 |
|
NRM Mo 204781* |
47.2 |
31.5 |
na |
UWBM loc. B7452 |
|
NRM Mo 204784* |
46.0 |
30.4 |
19.4 |
UWBM loc. B7452 |
|
NRM Mo 204785** |
46.5 |
29.8 |
19.0 |
UWBM loc. B7452 |
|
NRM Mo 204788 |
44.9 |
39.4 |
16.8 |
UWBM loc. B7452 |
|
NRM Mo 204789 |
16.8 |
12.4 |
7.2 |
UWBM loc. B7452 |
|
NRM Mo 204791 |
24.0 |
16.2 |
na |
W of Whiskey Creek |
|
NRM Mo 204813* |
30- |
25+ |
15.8 |
UWBM loc. B7452 |
|
NRM Mo 204814* |
32+ |
17.5+ |
13.5 |
UWBM loc. B7452 |
|
UWBM 117930 |
36.1 |
21.8 |
13.4 |
UWBM loc. B7452 |
|
UWBM 117931 |
47.0 |
33.0 |
18+ |
UWBM loc. B7452 |
Diagnosis.—Shell oval to elongate-oval, moderately inflated, reaching nearly 50 mm in length; blunt, submarginal posterior ridge; umbones prominent, elevated, prosogyrate; lunular incision very elongate, often indistinct; ligament short.
Description.—Shell medium-sized, broadly oval-elongate, moderately inflated (max. L = 48.6 mm, H = 37.5 mm, W = 22.7 mm); umbones elevated, prosogyrate, positioned anterior at about 21% of total shell length; blunt ridge close to posterodorsal margin; elongate lunular incision present but not very deep; short and narrow ligament, no escutcheon. Anterior adductor muscle scar D-shaped, posterior margin shallow; pallial line starting at posteroventral corner of anterior adductor scar, extending mostly parallel to ventral shell margin, near posterior end of shell bending upward to continue nearly straight into the base of the posterior adductor muscle scar; posterior adductor muscle scar round, with weak but distinct ridge running from umbo to ventral margin where it meets the pallial line. Hinge plate narrow, with three teeth in each valve; teeth in RV subparallel to dorsal shell margin, 1 thin, elongate, just anterior of umbo, 3a and 3b both short, thin, originating underneath umbo; LV with thin 2a, subparallel to dorsal shell margin, 2b nearly perpendicular to 2a, thick and short.
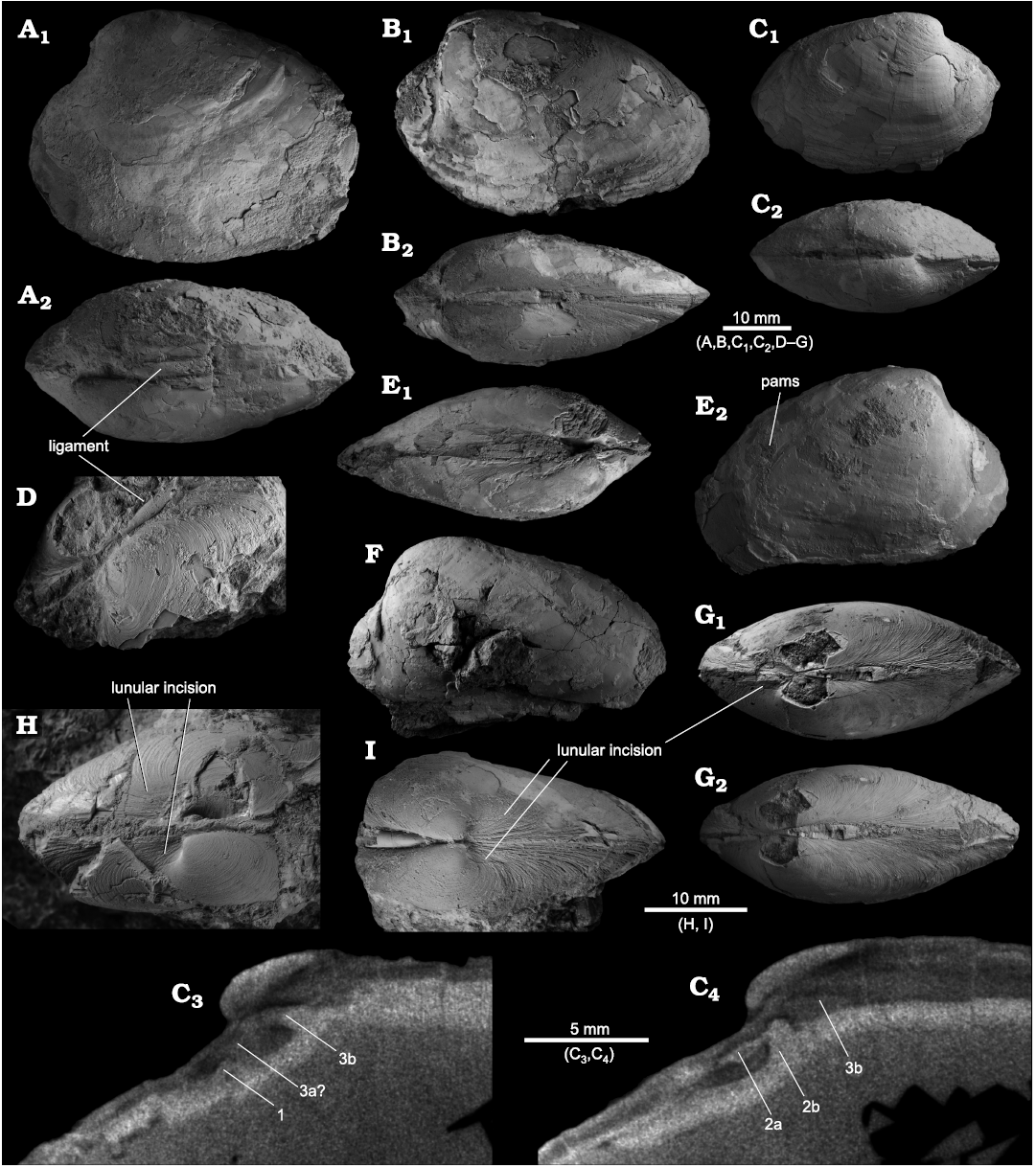
Fig. 6. The vesicomyid bivalve Pliocardia? guthrieorum sp. nov., from the Oligocene seep deposits at UWMB loc. B7452, Lincoln Creek Formation, western Washington State, USA. A. Paratype, NRM Mo 204779, large specimen showing outline, inflation, and ligament; left valve (A1) and dorsal (A2) views. B. Paratype, NRM Mo 204784, showing outline, inflation, and anterior adductor muscle scar; left valve (B1) and dorsal (B2) views. C. Paratype, NRM Mo 204780, small, elongate specimen; right valve (C1) and dorsal (C2) views; digital sections through the hinge area, with our interpretation of the hinge teeth indicated (C3, C4). D. Paratype, NRM Mo 204782, showing ligament. E. Holotype, NRM Mo 204785, showing inflation (E1) and both adductor muscle scars and the pallial line in the RV (E2). F. Paratype, NRM Mo 204781, large specimen with particularly large and blunt umbo. G. Paratype, NRM Mo 204814, small specimen in dorsal view, showing inflation and lunular incision (G1) and the lack of an escutcheon (G2). H. Paratype, NRM Mo 204786, showing lunular incision. I. Paratype, NRM Mo 204813, showing lunular incision. Abbreviation: pams, posterior adductor muscle scar.
Remarks.—All specimens are from the Oligocene part of the Lincoln Creek Formation in the Canyon and Satsop rivers area. One specimen from limestone deposits found in upper Eocene rocks mapped as Makah Formation, just west of Whiskey Creek, may also belong here. It is a small (c. 25 mm long) right valve that externally resembles Pliocardia? guthrieorum, but it is embedded in rock matrix so neither hinge nor pallial line or muscle scars are visible, hampering a reliable identification.
We hesitantly assign this species to Pliocardia because it resembles some of the larger-sized extant species assigned to this genus, as well as for example P. kawadai reported above. However, virtually all species of Pliocardia are more strongly inflated than Pliocardia? guthrieorum. Furthermore, our interpretation of the hinge dentition is based on x-ray scanning of a specimen with an only moderately preserved hinge, and some uncertainty remains, in particular about cardinal 3a. Extant species of the genus Wareniconcha Cosel and Olu, 2009, have shells with similar minimal inflation, but they have more convex ventral margins and less elevated umbones, and they have an escutcheon (Smith 1906; Prashad 1932; Cosel and Olu 2009; Krylova and Sahling 2010), which Pliocardia? guthrieorum lacks.
Stratigraphic and geographic range.—Upper Eocene? to Oligocene seep deposits; Olympic Peninsula, western Washington State, USA.
Genus Squiresica Hybertsen, Goedert, and Kiel, 2022
Type species: Archivesica knapptonensis Amano and Kiel, 2007, by original designation; Oligocene, western Washington State, USA (Hybertsen et al. 2022).
Remarks.—Some species of Squiresica show a dimorphism in shell shape. The posterior part of the shell either has an evenly rounded posterior margin (i.e., S. yooni sp. nov., Fig. 9C2) or a sloping posterodorsal margin (i.e., S. yooni sp. nov., Fig. 9A3, B2). This dimorphism has been reported from extant vesicomyid taxa such as Calyptogena (Krylova and Sahling 2006). Although it occurs in fossil vesicomyids too (SK personal observation), it is rarely explicitly described.
Squiresica cf. knapptonensis (Amano and Kiel, 2007)
Fig. 7.
Material.—Eleven numbered specimens (NRM Mo 204773– 78; 204805–09) and numerous additional fragments (NRM Mo 204810) from the Eocene Siltstone of Unit B in the Grays River area, western Washington State, USA.
Dimensions.—See Table 4.
Table 4. Measurements (in mm) of Squiresica cf. knapptonensis (Amano and Kiel, 2007). na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
Locality |
|
NRM Mo 204773 |
23.6 |
12.0 |
na |
Grays River |
|
NRM Mo 204774 |
21.7 |
10.0 |
na |
Grays River |
|
NRM Mo 204775 |
23.5 |
12.3 |
8.0 |
Grays River |
|
NRM Mo 204776 |
13.5+ |
10.7 |
na |
Grays River |
|
NRM Mo 204777 |
21.7 |
12.2 |
na |
Grays River |
|
NRM Mo 204778 |
23.7 |
11.5 |
na |
Grays River |
|
NRM Mo 204807 |
16.3 |
10.0 |
na |
Grays River |
|
NRM Mo 204808 |
18.0 |
12.0 |
na |
Grays River |
Remarks.—The visible features of the late Eocene age specimens from the Siltstone of unit B fall within the variation seen in the early to late Oligocene age specimens of S. knapptonensis from various deposits of the Lincoln Creek Formation (Amano and Kiel 2007; Hybertsen et al. 2022). However, their hinge dentition remains unknown and therefore we assign these specimens only preliminarily to S. knapptonensis.
Stratigraphic and geographic range.—Upper Eocene, Siltstone of unit B, southwestern Washington State, USA.
Squiresica plana sp. nov.
Fig. 8.
1993 Calyptogena chinookensis Squires and Goedert, 1991; Goedert and Squires 1993: fig. 5.
1995 Calyptogena (Calyptogena) chinookensis Squires and Goedert, 1991; Goedert and Campbell 1995: 24, table 1.
Zoobank LCID: urn:lsid:zoobank.org:act:82DEC36A-45E3-438D-9A F0-99BBD8F1A5D4.
Etymology: From Latin plana, flat; in reference to the minimal inflation of the species.
Type material: Holotype: articulated specimen with shell material (NRM Mo 204704). Paratypes: single valves or semi-articulated specimens embedded in rock matrix (NRM Mo 204705, 06, 204770–72) from the type locality and horizon.
Type locality: UWBM loc. B7452, Canyon River, Washington State, USA.
Type horizon: The Lincoln Creek Formation, Oligocene.
Material.—Four specimens (NRM Mo 204769–72) and numerous fragments from Canyon River (UWBM loc. B7452), five specimens from LACMIP loc. 17426 (NRM Mo 204704–08); all Lincoln Creek Formation. About ten fragmentary specimens showing external features only (NRM Mo 204834) from LACMIP loc. 15621, Pysht Formation. Oligocene, Washington State, USA.
Diagnosis.—Small for genus, minimally inflated, lunular incision very indistinct, ligament short, anterior adductor muscle scar circular.
Description.—Small, elongate, little inflated shells reaching 21 mm in length; umbones only slightly elevated, slightly prosogyrate, pointed, situated anteriorly at about 20% total shell length; surface covered by fine, dense growth increments; escutcheon and ligament short, narrow, no lunular incision. Anterior adductor muscle scar roundish with acute dorsal side; pallial line starting just anterior of posteroventral corner of anterior adductor muscle scar, running parallel to ventral margin, at posterior end curving upward into circular posterior adductor muscle scar. Hinge plate moderately wide, LV with three radiating teeth, cardinal tooth 2a thin, pointing anteriorly, 2b thick, short, pointing posteroventrally, cardinal tooth 4b short, thin, subparallel to 2b; subumbonal pit small, triangular.

Fig. 7. The vesicomyid bivalve Squiresica cf. knapptonensis (Amano and Kiel, 2007) from upper Eocene seep deposits of the Siltstone Unit B along the West Fork of Grays River in western Washington State, USA. A. NRM Mo 204773, internal mold of RV showing adductor muscle scars and pallial line. B. NRM Mo 204808, internal mold of semi-articulated specimen; view on RV (B1); close-up on lunular incision (B2). C. NRM Mo 204776, internal mold of LV showing anterior adductor muscle scar, onset of pallial line, and the short ligament. D. NRM Mo 204775, internal mold of RV showing adductor muscle scars and pallial line. E. NRM Mo 204809, anterior half of RV showing external shell sculpture. F. NRM Mo 204805, view on internal mold of posterior adductor muscle scar. G. NRM Mo 204807I, internal mold of semi-articulated specimen, view on LV. H. NRM Mo 204806, close-up on lunular incision of LV.

Fig. 8. The vesicomyid bivalve Squiresica plana sp. nov., from Oligocene seep deposits in the Lincoln Creek Formation in western Washington State, USA. Specimens from UWBM loc. B7452 (A–C) and LACMIP loc. 17426 (= SR4) (D–F). A. Paratype, NRM Mo 204772, exterior of LV showing fine growth increments. B. Paratype, NRM Mo 204771, exterior of LV showing shell outline and fine growth increments. C. Paratype, NRM Mo 204770, close-up on anterior part of RV showing near absence of a lunular incision. D. Holotype, NRM Mo 204704, articulated specimen with preserved shell; lateral view on RV (D1) and dorsal view, showing inflation and ligament (D2). E. Paratype, NRM Mo 204705, specimen with partially preserved shell, showing anterior adductor muscle scar and onset of pallial line. F. Paratype, NRM Mo 204706, semi-articulated specimen; F1, close-up on LV hinge area; F2, lateral view on LV.
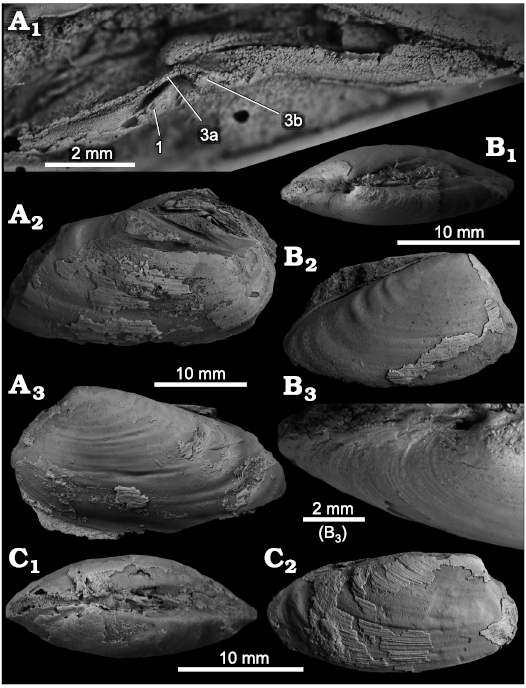
Fig. 9. The vesicomyid bivalve Squiresica yooni sp. nov., from the Middle Miocene Duho Formation, loc. 18 of Yoon (1976b), South Korea. A. Holotype, NRM Mo 204829, an articulated specimen with preserved cast of RV hinge area; close-up on hinge area (A1), view on right valve and exposed hinge of left valve (A2), and view on left valve (A3). B. Paratype, NRM Mo 204832, articulated specimen, mostly an internal mold; dorsal view showing inflation (B1); view on exterior of RV (B2); close-up on anterior part of LV showing the lack of a lunular incision (B3). C. Paratype, NRM Mo 204831, articulated specimen with some shell preserved; dorsal view showing inflation (C1); lateral view on RV showing fine growth increments (C2).
Dimensions.—See Table 5.
Table 5. Measurements (in mm) of Squiresica plana sp. nov. ** holotype; * paratype; na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
Locality |
|
NRM Mo 204704** |
14.2 |
7.0 |
4.4 |
LACMIP loc. 17426 |
|
NRM Mo 204705* |
15.7 |
10.0 |
5.4 |
LACMIP loc. 17426 |
|
NRM Mo 204706* |
12.0 |
7.8 |
4.0 |
LACMIP loc. 17426 |
|
NRM Mo 204707 |
14.8 |
9.0 |
5.0 |
LACMIP loc. 17426 |
|
NRM Mo 204708 |
12.8 |
7.6 |
4.0 |
LACMIP loc. 17426 |
|
NRM Mo 204769 |
20.8 |
8.7 |
na |
UWBM loc. B7452 |
|
NRM Mo 204770* |
12.4+ |
7.8 |
na |
UWBM loc. B7452 |
|
NRM Mo 204771* |
14.5 |
6.3 |
na |
UWBM loc. B7452 |
|
NRM Mo 204772* |
7.5 |
3.3 |
na |
UWBM loc. B7452 |
Remarks.—Squiresica plana sp. nov. differs from Squiresica knapptonensis mainly by having a smaller, less inflated shell with posteriorly directed 2b tooth and very indistinct, elongate and narrow lunular incision. The Miocene Squiresica yooni sp. nov. from Korea described below is more inflated and has a wider escutcheon. Oligocene specimens previously reported as Calyptogena chinookensis Squires and Goedert, 1991, from the Pysht and Makah formations in northwestern Washington State (Goedert and Squires 1993; Goedert and Campbell 1995) have the general shape and size of S. plana and are here considered as belonging to this species. They are quite different from the large type specimens of Pleurophopsis chinookensis (discussed below) that reach 80 mm in length.
Stratigraphic and geographic range.—Seep deposits in the Oligocene parts of the Pysht, and Lincoln Creek formations, western Washington State, USA.
Squiresica yooni sp. nov.
Fig. 9.
1976 Calyptogena cf. elongata Dall; Yoon 1976a: 13, pl. 3: 3, 7, 9.
Zoobank LCID: urn:lsid:zoobank.org:act:74536D5A-3B34-4763-B4E0 -89FC7DD33241.
Etymology: In honor of Sun Yoon, who donated the illustrated specimens to the late Professor Kanno, who in turn passed them on to KA.
Type material: Holotype: partially damaged internal mold, showing hinge characters (NRM Mo 204829). Paratypes: three articulated specimens, mostly internal molds (NRM Mo 204830–32) from the type locality and horizon.
Type locality: Loc. 18 of Yoon (1976b), hillside cut for the foundation of house building, about 400 m southeast of Goedong-dong, Daesong-myeon, Yeongil-gun, South Korea.
Type horizon: Duho Formation, uppermost part of Yeonil Group, Middle Miocene.
Material.—The type material only.
Dimensions.—See Table 6.
Table 6. Measurements (in mm) of Squiresica yooni sp. nov. from loc. 18 of Yoon (1976b), South Korea; ** holotype; * paratype.
|
Specimen no. |
Length |
Height |
Width |
|
NRM Mo 204829** |
30.0 |
17.0 |
8.2 |
|
NRM Mo 204830* |
24.4 |
13.0 |
8.0 |
|
NRM Mo 204831* |
19.8 |
9.6 |
6.0 |
|
NRM Mo 204832* |
19.4 |
11.2 |
6.0 |
Diagnosis.—Small Squiresica having some rough commarginal ribs in posterior surface, umbo strongly displaced anteriorly, convex ventral margin, very indistinct lunular incision, and rather strong cardinal tooth 1 of RV.
Original description.—“Shell small to medium in size, ovate, transversely elongated, thin. Beaks low, small, situated at about one-fourth of shell length anteriorly. No lunule, but escutcheon long, excavated; ligament external. Antero-dorsal margin short; anterior end rather narrowly rounded; posterodorsal margin long, almost straightly sloping down and passing into posterior end which is more narrowly rounded than the anterior one; ventral margin almost straight except for the both ends. Surface sculptured-with fine, somewhat rough, concentric growth lines.” (Yoon 1976a).
Supplemental description.—Hinge plate narrow, RV hinge with strong, elongate, forward-directed cardinal tooth 1, cardinal tooth 3a very thin, long, parallel to dorsal margin, cardinal tooth 3b short, bifid with raised edges, radiating posteroventrally, subumbonal pit elongate-triangular, very distinct. Slight dimorphism in shell outline: posterior margin either evenly rounded or with sloping posterodorsal margin.
Remarks.—According to Yoon (1976b), specimens of Squiresica yooni sp. nov. occur in dense aggregations of articulated valves in the siltstone of the type locality, associated with specimens of a thyasirid identified as Conchocele bisecta. Yoon’s (1976a) largest specimen was 34.5 mm long.
The main differences between Squiresica yooni and S. knapptonensis (Amano and Kiel, 2007) are the shorter anterior portion of the shell, the stronger cardinal tooth 1, and the virtual lack of a lunular incision in S. yooni. The Eocene–Oligocene Squiresica plana sp. nov. described above differs from S. yooni by being smaller (length up to 20.8 mm in S. plana vs. up to 30 mm in P. yooni), possessing a more distinct lunular incision and having more parallel dorsal and ventral shell margins.
Notably similar to Squiresica yooni regarding size, elongation, and inflation is the early Oligocene age Pleurophopsis lithophagoides Olsson, 1931, from Peru, which is also similar to the Oligocene Squiresica knapptonensis. With the pallial sinus and lunule of P. lithophagoides being unknown, its relation to Squiresica remains uncertain (i.e., Kiel et al. 2020b; Hybertsen et al. 2022).
Stratigraphic and geographic range.—Middle Miocene Duho Formation, South Korea.
Genus Pleurophopsis Van Winkle, 1919
Type species: Pleurophopsis unioides Van Winkle, 1919, by monotypy; Middle Miocene, Trinidad.
Pleurophopsis chinookensis (Squires and Goedert, 1991)
Fig. 10.
1990 Calyptogena n. sp.; Goedert and Squires 1990: fig. 20p, o.
1991 Calyptogena chinookensis Squires and Goedert, 1991: 414, figs. 2.7–2.11.
non 1993 Calyptogena chinookensis Squires and Goedert; Goedert and Squires 1993: 53, figs. 3–5.
non 1995 Calyptogena (Calyptogena) chinookensis Squires and Goedert; Goedert and Campbell 1995: 24, table 1.
2007 Adulomya chinookensis (Squires and Goedert, 1991); Amano and Kiel, 2007: 279, figs. 19–23.
Material.—15 specimens (NRM Mo 204815–20) from the upper Eocene Bear River seep deposit, Washington State, USA.
Dimensions.—See Table 7.
Table 7. Measurements (in mm) of Pleurophopsis chinookensis (Squires and Goedert, 1991) from the Bear River seep deposit. na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
|
NRM Mo 204815 |
65.8 |
25.5 |
na |
|
NRM Mo 204816 |
71.3 |
25.9 |
na |
|
NRM Mo 204817 |
70.4 |
33.6 |
23.4 |
|
NRM Mo 204818 |
78.4 |
28.2 |
na |
Remarks.—To supplement previous documentations and descriptions of this species (Squires and Goedert 1991; Amano and Kiel 2007), here we illustrate (i) features of the shell surface (Fig. 10A) including the absence of a lunular incision (Fig. 10E1, E2); (ii) a fully articulated specimen in dorsal view to show its true inflation (Fig. 10C); (iii) the short ligament (Fig. 10A2); (iv) the small, elongate anterior pedal retractor scar that is distinct from the adductor muscle scar (Fig. 10C); and (v) the hinge of a right valve showing two strong, radiating cardinal teeth, namely cardinals 1 and 3b, and a large and deep subumbonal pit (Fig. 10E3). This species was originally reported from middle to upper Eocene strata. Oligocene specimens previously assigned to P. chinookensis differ from those from the upper Eocene type locality by being notably smaller, less inflated, having a different arrangement of the hinge teeth, and possessing a lunular incision. These specimens belong to either Squiresica knapptonensis (Amano and Kiel, 2007) or S. plana sp. nov., as discussed above.

Fig. 10. The vesicomyid bivalve Pleurophopsis chinookensis (Squires and Goedert, 1991), from the upper Eocene Bear River seep deposit in western Washington State, USA. A. NRM Mo 204815, semi-articulated specimen with preserved shell material; view on LV (A1); dorsal view showing short ligament (A2). B. NRM Mo 204816, largely an internal mold, view on LV showing anterior adductor muscle scar. C. NRM Mo 204817, closely articulated specimen in dorsal view, showing true width. D. NRM Mo 204818, view on RV of internal mold showing internal posterior ridge. E. NRM Mo 204819, anterior part of RV with preserved shell, showing lack of lunular incision (E1, E2) and hinge dentition (E3). Abbreviations: aprs, anterior pedal retractor scar; sub. pit., subumbonal pit.
Stratigraphic and geographic range.—Humptulips Formation and Bear River seep deposit, Middle to upper Eocene, southwestern Washington State, USA.
Pleurophopsis thieli sp. nov.
Figs. 11, 12.
2002 Vesicomya (Calyptogena) sp.; Peckmann et al. 2002: 861, fig. 3C.
2003 Vesicomya (Calyptogena) sp.; Goedert et al. 2003: pl. 42: 6, 7.
Zoobank LCID: urn:lsid:zoobank.org:act:C3DAEBBE-2844-4BDB-BEA4-3C1C01A76A69.
Etymology: In honor of Bruce Thiel (Portland, Oregon), in recognition of his help with fieldwork related to this project.
Type material: Holotype: articulated internal mold showing pallial line and muscle scars (NRM Mo 204761). Paratypes: mostly internal molds (NRM Mo 204762–65, 204826) from Jansen Creek Member; articulated or semi-articulated specimens with shell material (Mo 204794–797) and internal molds (Mo 204798–802 from Whiskey Creek seep deposit. Upper Eocene to lower Oligocene, Washington State, USA.
Type locality: Shoreline exposures approximately 800 m southeast of the mouth of Rasmussen Creek, Clallam County, Washington State, USA.
Type horizon: Jansen Creek Member of the Makah Formation, lower Oligocene.
Material.—About 25 specimens and fragments from the Whiskey Creek site, Pysht Formation (NRM Mo 204794–204802, UWBM 97305, 06). About 34 specimens from the Jansen Creek Member of the Makah Formation, including four from near Rasmussen Creek (NRM Mo 204761–64) and about 30 from near the mouth of Jansen Creek, many in butterfly preservation (NRM Mo 204765–68, 204828). Nine specimens from UWBM loc. B6702 (= SR2 in Peckmann et al. 2002), Lincoln Creek Formation (NRM Mo 204826, 27). Upper Eocene to lower Oligocene, Washington State, USA. UWBM 97038 was previously reported as Vesicomya (Calyptogena) sp. by Peckmann et al. (2002).
Diagnosis.—Shell average size for genus, inflation slight to moderate, posterior side tapering, often bent downward due to concave ventral margin; anterior adductor muscle scar bean or D-shaped; posterior adductor muscle scar rather small, displaced anteriorly, near posterodorsal margin; hinge with two strong, radiating teeth in right valve and three in left valve; subumbonal pit large and deep.
Description.—Shell elongate, up to 57.6 mm long; umbones anterior, at about 20% of total shell length; blunt, low ridge running from umbones to posterior margin; no lunular incision or escutcheon; anterior margin evenly to acutely rounded; anterior half of ventral margin straight, posterior half slightly concave, giving shell curved appearance; posterodorsal margin either straight and slightly tapering (Fig. 11D), or anterior half straight and parallel to ventral margin, and posterior half more strongly inclined toward the ventral (Fig. 12E); posterior margin evenly rounded. Anterior adductor muscle scar bean to D-shaped with posterior edge, pallial line starting at its posteroventral corner, extending subparallel to ventral shell margin, and bending upward and anteriorly near the posterior shell margin, to merge into ventral margin of posterior adductor muscle scar; posterior adductor muscle scar positioned at about 30% of total shell length, relatively small, with broad trapezoid area including the pedal retractor scar to its anterior; hinge plate short and moderately broad, with two strong, radiating cardinal teeth in each valve; RV with short cardinal tooth 1 originating under umbo, running oblique to anterodorsal shell margin, cardinal tooth 3a reduced, 3b originating under umbo, short, running oblique to posterodorsal shell margin; LV with short, thin cardinal tooth 2a running subparallel to anterodorsal shell margin, 2b thick, triangular or bifid, directly under umbo, 4b very short and thin, or reduced; oval subumbonal pit present in each valve.
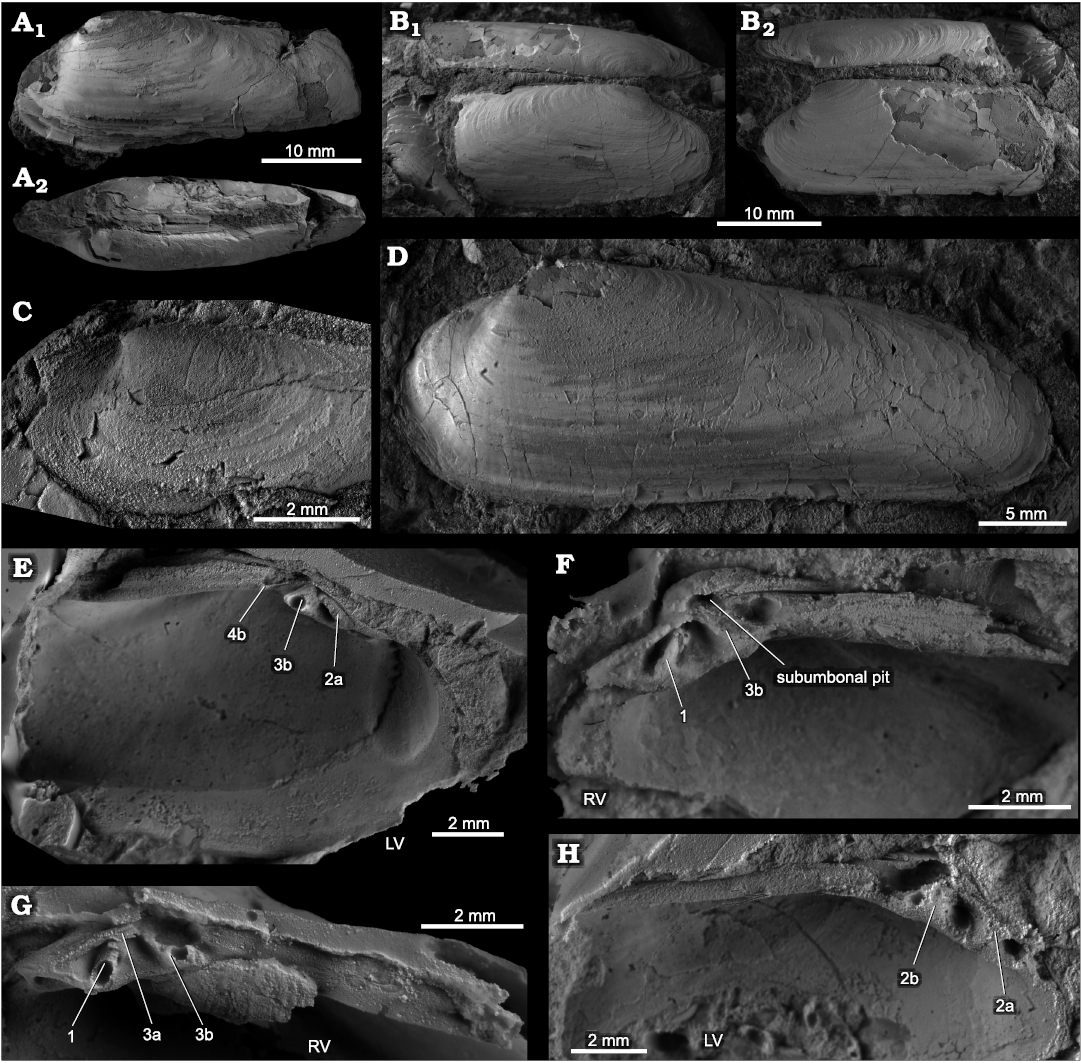
Fig. 11. The vesicomyid bivalve Pleurophopsis thieli sp. nov., from the upper Eocene Whiskey Creek seep deposit in the Pysht Formation in western Washington State, USA. A. NRM Mo 204797, small articulated specimen, showing inflation; left valve (A1) and dorsal (A2) views. B. NRM Mo 204795, semi-articulated specimen; left valve (B1) and right valve (B2) views. C. NRM Mo 204796, small left valve, showing lack of lunular incision. D. NRM Mo 204794, LV showing shell outline. E. NRM Mo 204799, hinge area and anterior adductor muscle scar of LV. F. NRM Mo 204800, hinge plate of right valve. G. NRM Mo 204801, hinge plate and ligament nymph of RV. H. NRM Mo 204802, hinge plate and ligament nymph of LV. All specimens are paratypes.
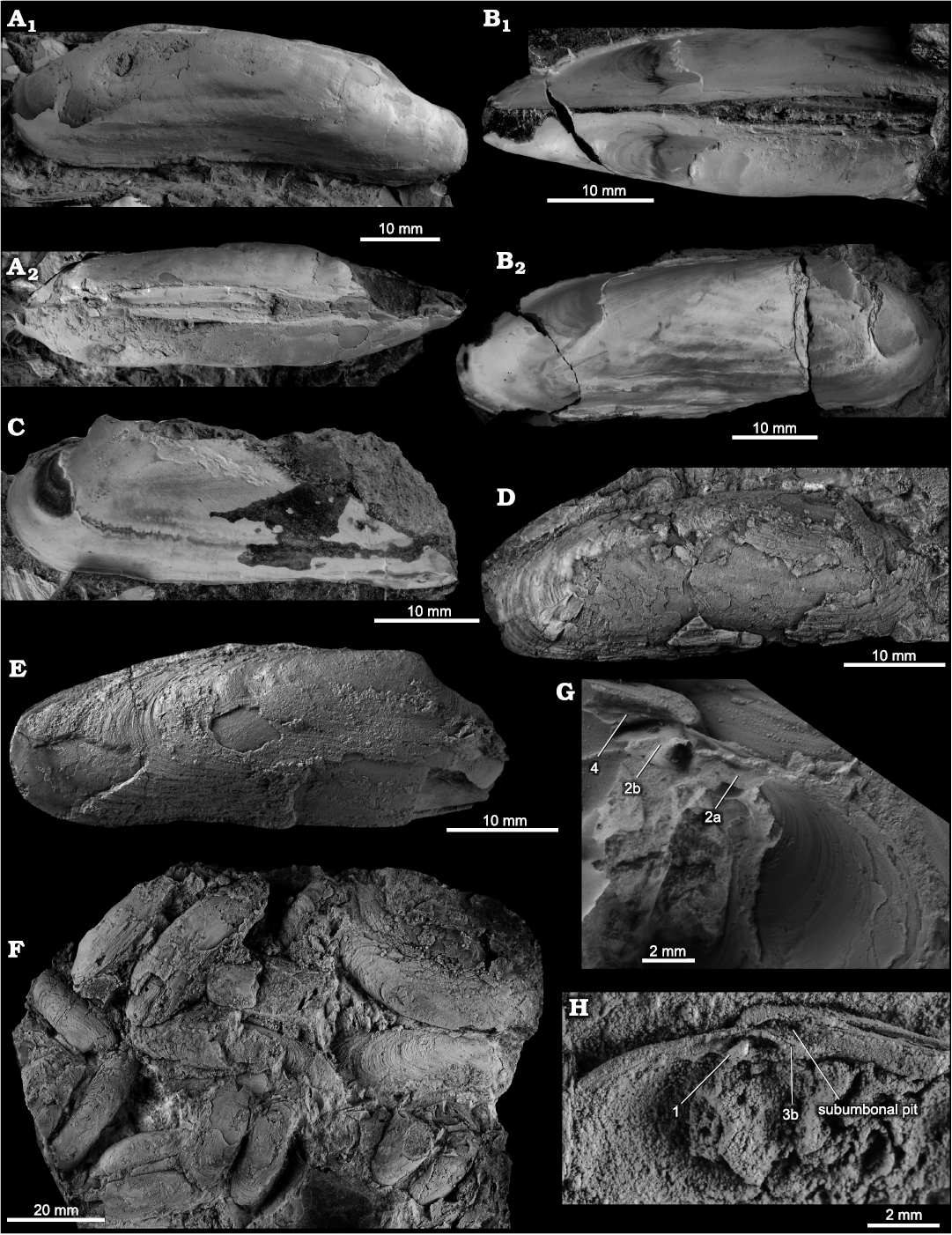
Fig. 12. The vesicomyid bivalve Pleurophopsis thieli sp. nov., from lower Oligocene, western Washington State, USA. Jansen Creek Member, Makah Formation (A–C, G); “Big slide”, Jansen Creek Member, Makah Formation (D–F); UWBM loc. B6702, Lincoln Creek Formation (H). A. Paratype, NRM Mo 204762, internal mold of articulated specimen, showing anterior adductor muscle scar (aams), inflation, and long ligament; left valve (A1) and dorsal (A2) views. B. Holotype, NRM Mo 204761, internal mold of articulated specimen, showing muscle scars and pallial lin; dorsal view of posterior part (B1) and right valve (B2). C. Paratype, NRM Mo 204764, internal mold of LV, showing anterior adductor muscle scar and course of anterior part of pallial line. D. Paratype, NRM Mo 204768, RV with preserved shell material, showing external sculpture. E. Paratype, NRM Mo 204765, RV with preserved shell material, showing external sculpture. F. Paratype, NRM Mo 204828, block with numerous specimens in butterfly preservation. G. Paratype, NRM Mo 204763, rubber cast of LV hinge area. H. Paratype, NRM Mo 204826, anterior part of RV showing hinge dentition. B and C not coated with ammonium chloride.
Dimensions.—See Table 8.
Table 8. Measurements (in mm) of Pleurophopsis thieli sp. nov. ** holotype; * paratype; na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
Locality |
|
NRM Mo 204794* |
35.7 |
13.5 |
na |
Whiskey Creek |
|
NRM Mo 204795* |
31.3 |
11.4 |
na |
Whiskey Creek |
|
NRM Mo 204796* |
9.2 |
4.5 |
na |
Whiskey Creek |
|
NRM Mo 204797* |
26.0 |
10.4 |
na |
Whiskey Creek |
|
NRM Mo 204798* |
38.8 |
16.0 |
na |
Whiskey Creek |
|
NRM Mo 204761** |
57.0 |
20.0 |
19.0 |
Jansen Creek area |
|
NRM Mo 204762* |
57.6 |
16.4 |
15.0 |
Jansen Creek area |
|
NRM Mo 204764* |
45.6 |
na |
na |
Jansen Creek area |
|
NRM Mo 204765* |
44.4 |
16.8 |
na |
Jansen Creek area |
|
NRM Mo 204766 |
37.8 |
13.3 |
na |
Jansen Creek area |
|
NRM Mo 204767 |
36.6 |
14.0 |
na |
Jansen Creek area |
|
NRM Mo 204768 |
43.8 |
15.0 |
na |
Jansen Creek area |
Remarks.—Most similar to Pleurophopsis thieli sp. nov. is Pleurophopsis chitanii (Kanehara, 1937) from Lower to Middle Miocene strata in Japan (Kanehara 1937; Hirayama 1973; Amano and Jenkins 2007; Amano and Kiel 2011). The main difference is in the posterior adductor muscle scar, which is smaller and more anteriorly positioned in P. thieli, whereas P. chitanii has a very large posterior adductor muscle scar in an almost terminal posterior position.
Specimens resembling P. thieli have been reported as P. chitanii from presumably Lower to Middle Miocene strata on Kayak Island, Alaska (Kanno 1971). In our view, the identity and relation of the Kayak Island specimens to either P. chitanii or P. thieli are somewhat uncertain. The size and morphological range of the specimens sketched and illustrated by Kanno (1971) suggests that they perhaps belong to more than one species. One illustrated specimen (Kanno 1971: pl. 7: 6) shows a pams that appears to be in a somewhat intermediate position between those seen in P. chitanii and P. thieli. Confusingly, although Kanno (1971) lists P. chitanii only from USGS loc. M1769 in the upper part of the Yakataga Formation on Kayak Island, Plafker (1974) lists only one mollusk from that locality, a sareptid bivalve. Plafker (1974) does, however, list P. chitanii from the upper and lower Yakataga Formation at four other localities on Kayak Island. Plafker (1974) lists Calyptogena n. sp.? from five additional localities, one of which (USGS loc. 15861), approximately 1.8 km east of Cape St. Peter is apparently the same, or very near loc. M1769 according to the map. Additional collecting effort is needed to clarify how many species of Pleurophopsis (or possibly other vesicomyid genera) are present in the strata on Kayak Island, including the upper and lower Katalla Formation because Plafker (1974) also recorded ?Hubertschenckia and a “vesicomyacid” from those strata. We also do not at present know anything about the local detailed geologic context for each of the sites listed and mapped by Plafker (1974); much more could be learned if these are discreet ancient seep sites. According to Addicott (1976: 123), the age of the Yakataga Formation on Kayak Island is Early Miocene, whereas the exposures on the mainland are Middle Miocene.
Pleurophopsis thieli differs from the late Eocene P. chinookensis by having a lower shell, more curved outline, resulting from its often slightly concave ventral margin and its sloping posterodorsal margin, whereas P. chinookensis has nearly straight and subparallel posterodorsal and posteroventral margins. The Middle Miocene type species P. unioides also has straighter and subparallel posterodorsal and posteroventral margins compared to P. thieli, and furthermore has larger and more round anterior and posterior adductor muscle scars (Van Winkle 1919; Kiel 2007; Krylova et al. 2010).
Stratigraphic and geographic range.—Seep deposits within the Pysht and Makah formations, upper Eocene to lowermost Oligocene, western Washington State, USA.
Genus Hubertschenckia Takeda, 1953
Type species: Tapes ezoensis Yokoyama, 1890, by monotypy; Poronai Formation, upper Eocene, Hokkaido, Japan.
Remarks.—A species named Hubertschenckia quisquiliae Hu, 1995, was reported from Eocene–Oligocene strata in south-central Taiwan, near Yakou village along Nankeng road (Hu 1995: 2020–2021, pl. 369: 1, 5; pl. 370: 9, 16; pl. 371: 1, 6, 8, 9). The shell was described as follows: “The shell is unequal triangular in shape, moderately swollen, with a short and concave front, a long and slight arc in the back, and a slightly boat-bottom-shaped ventral side. The umbo is located in front of the vertical midline, protruding and slightly inclined to the front. The front end of the shell is short and small, and the rear end is long and round. The surface is sculptured by evenly spaced commarginal ribs.” (Hu 1995, translated from Chinese by Liping Liu).
The illustrated specimens, some of which were examined by SK in the National Museum of Natural Sciences in Taichung (NMNS 007556), bear some resemblance to Hubertschenckia ezoensis in outline, showing a large, round anterior adductor muscle scar and reaching 70 mm in length. Some have distinctive, equally spaced commarginal ribs with smooth, rounded surfaces (similar to those of an Astarte). Although these ribs match those on the sketch reconstruction by Hu (1995: 2021, text-fig. 7), several illustrated specimens are far more elongated (like H. ezoensis) than on Hu’s (1995) sketch reconstruction. Whether all of those specimens belong to the same species, and whether any of the specimens belong to Hubertschenckia, remains questionable.
Hubertschenckia snatolensis Sinelnikova in Sinelnikova et al., 1991, was described from the middle Eocene Snatol Formation in western Kamchatka (Sinelnikova et al. 1991: 157, pl. 39: 14, pl. 40: 20). The species was described as “Shell elongate, flat, rather small, with almost parallel dorsal and ventral margin, and much anteriorly located umbo. Umbo very wide and weakly bending. Sculpture only consisting of growth lines. Hinge ill-preserved, but two cardinal teeth well visible on each valve. Pallial sinus wide, very short and high, with round top, located near muscle scar.Measurements (in mm); length = 35, height = 19, thickness = 8. Comparison; This species is distinguished from Hubertschenckia ezoensis (Yok.) by having much smaller size, longer and more flat shell.” (Sinelnikova et al. 1991, translated from Russian by KA).
The description and illustrations of this species are insufficient to accept or reject affinities to Vesicomyidae. Considering its size and outline, the specimens resemble Squiresica rather than Hubertschenckia; however, the reported two cardinal teeth in each valve would preclude a placement in either of these genera.
As noted in the discussion of Pleurophopsis thieli, an unidentified species questionably assigned to Hubertschenckia was listed from upper Oligocene to Lower Miocene localities in the Katalla Formation on Kayak Island, Alaska (Plafker 1974).
Hubertschenckia ezoensis (Yokoyama, 1890)
Fig. 13.
1890 Tapes ezoënsis sp. nov.; Yokoyama 1890: 197, pl. 25: 6–8.
non 1928 Meretrix (Macrocallista) ezoensis (Yokoyama). Yokoyama 1928: 77, pl. 8: 1.
1951 Tapes (new gen.?) ezoensis Yokoyama; Minato and Uozumi 1951: 150, pl. 13: 108a–c.
1953 Hubertschenckia ezoensis (Yokoyama); Takeda 1953: 85, pl.13: 5.
1960 Hubertschenckia ezoensis (Yokoyama); Oyama et al. 1960: 188–189, pl. 58: 2a–d.
non 1989 Hubertschenckia ezoensis (Yokoyama); Honda 1989: 79: 13, 14.
2007 Hubertschenckia ezoensis (Yokoyama); Amano and Kiel 2007: 275–277, figs. 6–12.
2007 Hubertschenckia ezoensis (Yokoyama); Amano and Jenkins 2007: fig. 2B.
2013 Hubertschenckia ezoensis (Yokoyama); Amano et al. 2013: figs. 6K–M.
2016 Hubertschenckia ezoensis (Yokoyama); Nobuhara et al. 2016: figs. 2–16.
2016 Hubertschenckia ezoensis (Yokoyama); Kiel et al. 2016: fig. 1G.
2022 Hubertschenckia ezoensis (Yokoyama); Amano et al. 2022: figs. 10.9a, k.
Material.—Five large specimens (NRM Mo 204821–25) from the upper Eocene Poronai Formation in Hokkaido, Japan.
Dimensions.—See Table 9.
Table 9. Measurements (in mm) of Hubertschenckia ezoensis (Yokoyama, 1890) from the Poronai Formation; na, measurement not possible.
|
Specimen no. |
Length |
Height |
Width |
|
NRM Mo 204821 |
55.8+ |
38.8 |
25.5 |
|
NRM Mo 204822 |
85.0 |
44.0 |
na |
|
NRM Mo 204823 |
na |
65.0 |
na |
|
NRM Mo 204824 |
90.4 |
52.6 |
na |
|
NRM Mo 204825 |
92.4 |
50.5 |
na |
Remarks.—We illustrate several very large specimens of Hubertschenckia ezoensis to further document the characteristics of this species and their variability. The largest previously published specimen measured 74.6 mm in length (Amano and Kiel 2007); the largest specimen reported here exceeds 90 mm in length. One of our specimens has a height of 65 mm. Considering the L/H ratio of ca. 1.8 known from other specimens of Hubertschenckia ezoensis, this specimen could have been almost 120 mm in length, making it the largest Eocene vesicomyid known to date.
Stratigraphic and geographic range.—Upper Eocene to lower Oligocene, Hokkaido, Japan.

Fig. 13. The vesicomyid bivalve Hubertschenckia ezoensis (Yokoyama, 1890), from the upper Eocene Poronai Formation in Hokkaido, Japan. A. NRM Mo 204823, very large RV; external sculpture (A1) and hinge (A2) views. B. NRM Mo 204821, small articulated specimen; showing ligament (B1) and external ornament and posterior ridge (B2). C. NRM Mo 204824, internal mold of large RV, showing anterior adductor muscle scar; view on right valve (C1) and close-up on hinge area (C2). D. NRM Mo 204825, large articulated specimen, mostly an internal mold; view on right valve (D1) and left valve (D2). E. NRM Mo 204822, hinge of LV.
Discussion
The two Vesicomya? spp. reported here from the Oligocene are potentially the geologically oldest members of this genus. They would significantly extend the fossil record of Vesicomya compared to the next oldest records from Upper Miocene rocks of Buton Island, Indonesia (Beets 1953). Vesicomya is considered the sister taxon to all other vesicomyids (Johnson et al. 2017). Small, thin-shelled deep-water bivalves such as Vesicomya are particularly prone to have an incomplete fossil record (Valentine et al. 2006), and hence an even longer fossil record could be expected.
The new records and species of Squiresica, S. plana and S. yooni, expand the stratigraphic range of this genus, from the upper Eocene to the mid-Miocene. They also expand the geographic range of the genus, which now includes both sides of the North Pacific Ocean. The genus is not known from seep deposits of similar age in the Caribbean region, despite a relatively good fossil record (Gill et al. 2005; Kiel and Hansen 2015).
The decrease in vesicomyid species diversity in western Washington State after the Oligocene (Fig. 14) is likely a facies bias because most Neogene sediments in this region were deposited in increasingly shallower water (Kiel 2010c), where vesicomyids are uncommon. The Lower Miocene deep-water strata on Kayak Island, Alaska (Plafker 1974), may therefore be uniquely informative in this respect.
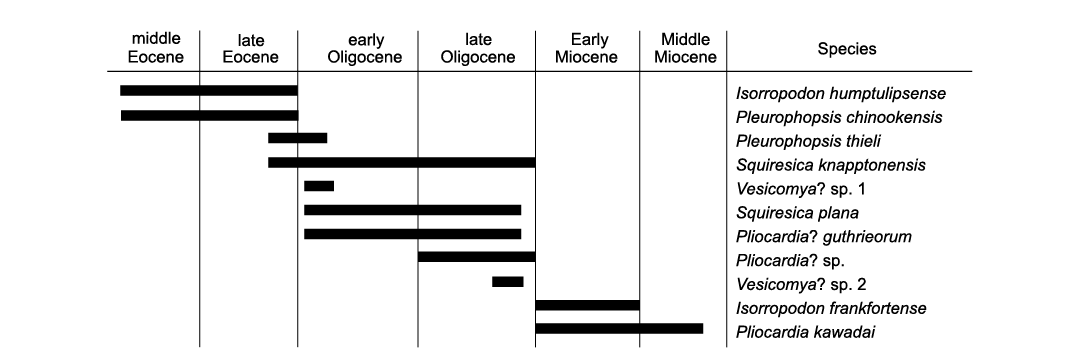
Fig. 14. Stratigraphic ranges of vesicomyid bivalve species in western Washington State, USA (reported herein and by Amano and Kiel 2007).
Several vesicomyid species today have broad geographic ranges including trans-Pacific occurrences (Kojima et al. 2004; Audzijonyte et al. 2012). There is no evidence for shared species or even genera between the eastern and western North Pacific during the Eocene and most of the Oligocene. The late Oligocene–Early Miocene Pleurophopsis matsumotoi Amano, Miyajima, Jenkins, and Kiel, 2019, from Japan is the first western Pacific occurrence of a vesicomyid genus that occurred earlier only in the eastern Pacific Ocean (Amano et al. 2019a). The first vesicomyid species with a trans-Pacific distribution is the Early to Middle Miocene Pliocardia kawadai, initially described from Japan and here reported from western Washington State, USA. However, the Oligocene record of seep deposits and vesicomyids of Japan is rather poor, with only three reported occurrences: Hubertschenckia ezoensis from upper Eocene to possibly lower Oligocene seep carbonates in southern Honshu (Amano et al. 2013) and from lower Oligocene seep deposit in eastern Hokkaido (Amano and Jenkins 2013; Kiel et al. 2016), and an upper Oligocene to Lower Miocene carbonate nodule with Pleurophopsis matsumotoi (Amano et al. 2019a). Thus, increased sampling in the Paleogene of Japan could potentially change this picture of shared, trans-Pacific vesicomyid species.
The fossil record of Late Miocene and younger deep-water faunas in western North America becomes rather poor, and a thorough comparison with the much better record of Japan is not feasible. Calyptogena first appears in the Oligocene of Alaska and is known from Japan from the Late Miocene and younger rocks (Kiel and Amano 2010; Amano et al. 2019a), suggesting a Northeastern Pacific origin of this genus. Furthermore, the Plio-Pleistocene Archivesica gibbera (Crickmay, 1929) and A. lasia (Woodring, 1938) from California (e.g., Squires 1991) seem to have quite close relatives in coeval strata in Japan, such as the Pliocene A. kannoi Amano and Kiel, 2010. Archivesica is regarded as having a warm-water origin (Amano and Jenkins 2011a; Amano et al. 2019a), and with Archivesica being unknown from Oligocene–Miocene deposits in the Caribbean region (Gill et al. 2005; Kiel and Hansen 2015) but present in Miocene to Pleistocene deposits of the Philippines (Kiel et al. 2020a, 2022), a western Pacific source area for the East Pacific, present-day Archivesica seems plausible.
Among other families of chemosymbiotic bivalves, the eastern and western North Pacific regions shared the bathymodiolin mussel Bathymodiolus inouei Amano and Jenkins, 2011, the lucinid Lucinoma acutilineatum (Conrad, 1849), and the thyasirid Conchocele taylori Hickman, 2015, during the late Eocene to Oligocene (Majima et al. 2005; Kiel 2010c; Amano and Jenkins 2011b; Kiel and Amano 2013; Hickman 2015; Hryniewicz et al. 2017). Whether the Neogene Conchocele bisecta (Conrad 1849) indeed occurs on both sides of the North Pacific (Weaver 1942; Majima et al. 2005; Kiel 2010a), or represents an as-yet undifferentiated garbage-bin taxon, remains to be tested.
Conclusions
Five new species and one new genus of Vesicomyidae are described from Cenozoic deep-water deposits in the North Pacific region. These include the oldest known vesicomyid, the middle to late Eocene Isorropodon humptulipsense, and the oldest records of the genera Squiresica (late Eocene), and possibly Vesicomya (early Oligocene). Our new findings document a rapid early morphological diversification among vesicomyids, starting with the short, strongly inflated Isorropodon humptulipsense and the elongate, inflated Pleurophopsis chinookensis in the middle Eocene. By the late Eocene, two further morphologies had appeared: the small, little inflated, elongated Squiresica, and the large, oval-elongate, moderately inflated Hubertschenckia. During the Eocene and most of the Oligocene there was apparently little faunal exchange among vesicomyids between the eastern and western North Pacific. Colonization from east to west appears to have commenced in the late Oligocene with the genus Pleurophopsis, and the Early to Middle Miocene occurrence of Pliocardia kawadai in both Japan and western Washington State is currently the oldest record of a vesicomyid species with a trans-Pacific distribution.
Acknowledgements
We are extremely grateful for the continuing cooperation of Green Diamond Resource Company allowing access to localities on their land in western Washington, and we thank Mark Golliet (Shelton, Washington, USA) for facilitating this access. Bruce Thiel (Portland, Oregon, USA) is thanked for assisting with fieldwork in the Grays River area in Washington, Ron Thielen (Naselle, Washington, USA) for allowing SK and JLG to use his kayaks to access sites along the shore of Columbia River, Ron Eng (UWBM) provided specimen and locality numbers, Engla Nordin (Stockholm and Gothenburg, Sweden) for her skillful preparation of rubber casts, Liping Liu (NRM) for translating the Chinese description of Hubertschenckia quisquiliae, Tunhe Zhou (SUBIC, Stockholm, Sweden) for operating the x-ray microscope, Therese Flink (Stockholm, Sweden) for help with the Dragonfly software, and Shih-Wei Wang (National Museum of Natural Science, Taichung, Taiwan) for providing access to specimens in his care. We also thank the reviewers Elena Krylova (Institute of Oceanology RAS) and Tom DeVries (UWBM) for the insightful comments that helped to improve the quality of our study. Field work by SK was financially supported by the Swedish Research Council (Vetenskapsrådet) through grant 2016-03920.
References
Addicott, W.O. 1976. On the significance of the bivalve Acila gettysburgensis (Reagan) in middle Tertiary chronostratigraphy of the Pacific Coast. The Veliger 19: 121–124.
Amano, K. and Jenkins, R.G. 2007. Eocene drill holes in cold-seep bivalves of Hokkaido, northern Japan. Marine Ecology 28: 108–114. Crossref
Amano, K. and Jenkins, R.G. 2011a. Fossil record of extant vesicomyid species from Japan. Venus 69: 163–176.
Amano, K. and Jenkins, R.G. 2011b. New fossil Bathymodiolus (s. l.) (Mytilidae, Bivalvia) from Oligocene seep-carbonates in eastern Hokkaido, Japan, with remarks on the evolution of Bathymodiolus (s. l.). The Nautilus 125: 29–35.
Amano, K. and Jenkins, R.G. 2013. A new species of Provanna (Gastropoda: Provannidae) from an Oligocene seep deposit in eastern Hokkaido, Japan. Paleontological Research 17: 325–329. Crossref
Amano, K. and Kiel, S. 2007. Fossil vesicomyid bivalves from the North Pacific region. The Veliger 49: 270–293.
Amano, K. and Kiel, S. 2010. Taxonomy and distribution of fossil Archivesica (Vesicomyidae, Bivalvia) in Japan. The Nautilus 124: 155–165.
Amano, K. and Kiel, S. 2011. Fossil Adulomya (Vesicomyidae, Bivalvia) from Japan. The Veliger 51: 76–90.
Amano, K. and Kiel, S. 2012. Two Neogene vesicomyid species (Bivalvia) from Japan and their biogeographic implications. The Nautilus 126: 79–85.
Amano, K., Hamuro, T., Hamuro, M., and Fujii, S. 2001. The oldest vesicomyid bivalves from the Japan Sea borderland. Venus 60: 189–198.
Amano, K., Jenkins, R.G., Sako, Y., Ohara, M., and Kiel, S. 2013. A Paleogene deep-sea methane-seep community from Honshu, Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 387: 126–133. Crossref
Amano, K., Kiel, S., Hryniewicz, K., and Jenkins, R.G. 2022. Bivalvia in ancient hydrocarbon seeps. In: A. Kaim, N.H. Landman, and J.K. Cochran (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 267–321. Crossref
Amano, K., Little, C.T.S., and Inoue, K. 2007. A new Miocene whale-fall community from Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 247: 236–242. Crossref
Amano, K., Miyajima, Y., Jenkins, R.G., and Kiel, S. 2019a. The Miocene to Recent biogeographic history of vesicomyid bivalves in Japan, with two new records of the family. The Nautilus 133: 48–56.
Amano, K., Miyajima, Y., Nakagawa, K., Hamuro, M., and Hamuro, T. 2019b. Chemosymbiotic bivalves from the lower Miocene Kurosedani Formation in Toyama Prefecture, central Honshu, Japan. Paleontological Research 23: 208–219. Crossref
Aoki, S. 1954. Mollusca from the Miocene Kabeya Formation, Joban coal-field, Fukushima Prefecture, Japan. Science Reports of the Tokyo Kyoiku Daigaku, Section C 17: 23–40.
Audzijonyte, A., Krylova, E.M., Sahling, H., and Vrijenhoek, R.C. 2012. Molecular taxonomy reveals broad trans-oceanic distributions and high species diversity of deep-sea clams (Bivalvia: Vesicomyidae: Pliocardiinae) in chemosynthetic environments. Systematics and Biodiversity 10: 403–415. Crossref
Beets, C. 1953. Reconsideration of the so-called Oligocene fauna in the asphaltic deposits of Buton (Malay Archipelago). Leidse Geologische Mededelingen 17: 237–258.
Berglund, R.E. and Goedert, J.L. 1992. A new species of Cancer (Decapoda: Brachyura) from the Miocene Astoria Formation in Washington. Burke Museum Contributions in Anthropology and Natural History 9: 1–11.
Brown, R.D.J. and Gower, H.D. 1958. Twin River Formation (redefinition), northern Olympic Peninsula, Washington. American Association of Petroleum Geologists Bulletin 42: 2492–2512. Crossref
Conrad, R.A. 1849. Fossils from northwestern America. In: J.D. Dana (ed.), U.S. Exploration Expedition, 1838–1842, under Charles Wilkes. Geology, Volume 10, 723–728 (appendix). C. Sherman, Philadelphia.
Cosel, R. von and Olu, K. 2009. Large Vesicomyidae (Mollusca: Bivalvia) from cold seeps in the Gulf of Guinea off the coasts of Gabon, Congo and northern Angola. Deep-Sea Research II 56: 2350–2379. Crossref
Cosel, R. von and Salas, C. 2001. Vesicomyidae (Mollusca: Bivalvia) of the genera Vesicomya, Waisiuconcha, Isorropodon and Callogonia in the eastern Atlantic and the Mediterranean. Sarsia 86: 333–366. Crossref
Crabtree, D.R. and Miller, C.N.J. 1989. Pityostrobus makahensis, a new species of silicified pinaceous seed cone from the middle Tertiary of Washington. American Journal of Botany 76: 176–184. Crossref
Crickmay, C.H. 1929. On a new pelecypod Calyptogena gibbera. The Canadian Field-Naturalist 43: 93.
Dall, W.H. 1886. Diagnoses of new species of mollusks from the west coast of America. Proceedings of the U.S. National Museum of Natural History 18: 7–20.
Dall, W.H. 1903. Contributions to the Tertiary fauna of Florida, with especial reference to the silex beds of Tampa and the Pliocene beds of the Caloosahatchie River, including in many cases a complete revision of the generic groups treated of and their American species. Part VI. Concluding the work. Wagner Free Institute of Science of Philadelphia, Transactions 3: xiv + 1219–1654. Crossref
Dall, W.H. and Simpson, C.T. 1901. The Mollusca of Porto Rico. United States Fishery Commission, Bulletin 20: 351–524.
Feldman, R.A. 1989. Lyreidus alseanus Rathbun from the Paleogene of Washington and Oregon, U.S.A. Annals of Carnegie Museum 58: 61–70. Crossref
Fisher, C.R. 1990. Chemoautotrophic and methanotrophic symbioses in marine invertebrates. Reviews in Aquatic Sciences 2: 399–436.
Garvie, C.L., Goedert, J.L., and Janssen, A.W. 2020. Paleogene and late Cretaceous Pteropoda (Mollusca, Gastropoda, Heterobranchia) from North America. Zootaxa 4782: 1–115. Crossref
Gill, F.L., Harding, I.C., Little, C.T.S., and Todd, J.A. 2005. Palaeogene and Neogene cold seep communities in Barbados, Trinidad and Venezuela: An overview. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 191–209. Crossref
Goedert, J.L. and Benham, S.R. 2003. Biogeochemical processes at ancient methane seeps: The Bear River site in southwestern Washington. Geological Society of America Field Guide 4: 201–208. Crossref
Goedert, J.L. and Campbell, K.A. 1995. An Early Oligocene chemosynthetic community from the Makah Formation, northwestern Olympic Peninsula, Washington. The Veliger 38: 22–29.
Goedert, J.L. and Cornish, J. 2002. A preliminary report on the diversity and stratigraphic distribution of the Plotopteridae (Pelecaniformes) in Paleogene rocks of Washington State, USA. In: Z. Zhou and F. Zhang (eds.), 5th Symposium of the Society of Avian Paleontology and Evolution, 63–76. Beijing Science Press, Beijing.
Goedert, J.L. and Kaler, K.L. 1996. A new species of Abyssochrysos (Gastropoda: Loxonematoidea) from a Middle Eocene cold-seep carbonate in the Humptulips Formation, western Washington. The Veliger 39: 65–70.
Goedert, J.L. and Squires, R.L. 1990. Eocene deep-sea communities in localized limestones formed by subduction-related methane seeps, southwestern Washington. Geology 18: 1182–1185. Crossref
Goedert, J.L. and Squires, R.L. 1993. First Oligocene record of Calyptogena (Bivalvia: Vesicomyidae). The Veliger 36: 72–77.
Goedert, J.L., Peckmann, J., Benham, S.R., and Janssen, A.W. 2013. First record of the Eocene pteropod Heliconoides nitens (Gastropoda: Thecosomata: Limacinidae) from the Pacific Basin. Proceedings of the Biological Society of Washington 126: 72–82. Crossref
Goedert, J.L., Thiel, V., Schmale, O., Rau, W.W., Michaelis, W., and Peckmann, J. 2003. The late Eocene “Whiskey Creek” methane-seep deposit (western Washington State) Part I: Geology, palaeontology, and molecular geobiology. Facies 48: 223–240. Crossref
Gray, J.E. 1854. A revision of the arrangement of the families of bivalve shells (Conchifera). Annals and Magazine of Natural History, Series 2 13: 408–418. Crossref
Hickman, C.S. 2015. Paleogene marine bivalves of the deep-water Keasey Formation in Oregon, part III: The heteroconchs. PaleoBios 32: 1–44. Crossref
Hirayama, K. 1973. Molluscan Fauna from the Miocene Hiranita Formation, Chichubu Basin, Saitama Prefecture, Japan. Tohoku University, Science Reports, 2nd series (Geology) Special Volume 6 (Hatai Memorial Volume): 163–177.
Hoffman, L., Cosel, R. von, and Freiwald, A. 2019. New and little-known Pliocardiinae (Bivalvia, Vesicomyidae) from the continental slope off Mauritania. Basteria 83: 151–157.
Honda, Y. 1989. Paleogene molluscan faunas from the Kushiro coal field, eastern Hokkaido. Science Reports of the Tohoku University, Sendai, Japan, 2nd Series (Geology) 60: 1–137.
Hryniewicz, K. 2022. Ancient hydrocarbon seeps of the world. In: A. Kaim, N.H. Landman, and J.K. Cochran (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 571–647. Crossref
Hryniewicz, K., Amano, K., Jenkins, R.G., and Kiel, S. 2017. Thyasirid bivalves from Cretaceous and Paleogene cold seeps. Acta Palaeontologica Polonica 62: 705–728. Crossref
Hu, C.-H. 1995. Molluscan fossils from various localities in Taiwan Island (continued) [in Chinese]. Taiwan Molluscan Fossils 4: 1927–2166.
Hybertsen, F. and Kiel, S. 2018. A middle Eocene seep deposit with silicified fauna from the Humptulips Formation in western Washington State, USA. Acta Palaeontologica Polonica 63: 751–768. Crossref
Hybertsen, F., Goedert, J.L., and Kiel, S. 2022. Squiresica, a new fossil genus of the chemosymbiotic bivalve family Vesicomyidae from western North America. Acta Palaeontologica Polonica 67: 703–709. Crossref
Ishida, Y., Thuy, B., Nam, G.-S., Martynov, A., Fujita, T., and Kim, J.-H. 2022. A new species of Ophiura (Echinodermata, Ophiuroidea) from Miocene deep-sea deposits in the Pohang Basin, Korea. Paleontological Research 26: 18–30. Crossref
Jakubowicz, M., Kiel, S., Goedert, J.L., Dopieralska, J., and Belka, Z. 2020. Fluid expulsion system and tectonic architecture of the incipient Cascadia convergent margin as revealed by Nd, Sr and stable isotope composition of mid-Eocene methane seep carbonates. Chemical Geology 558: 119872. Crossref
Janssen, R. and Krylova, E.M. 2012. Bivalves of the family Vesicomyidae from the Neogene Mediterranean basin (Bivalvia: Vesicomyidae). Archiv für Molluskenkunde 141: 87–113. Crossref
Johnson, S.B., Krylova, E.M., Audzijonyte, A., Sahling, H., and Vrijenhoek, R.C. 2017. Phylogeny and origins of chemosynthetic vesicomyid clams. Systematics and Biodiversity 15: 346–360. Crossref
Kamada, Y. 1962. Tertiary marine Mollusca from the Joban coal-field, Japan. Special Papers, Palaeontological Society of Japan 8: 1–187.
Kanehara, K. 1937. Miocene shells from the Joban coal field. Bulletin of the Imperial Geological Survey of Japan 27: 1–21.
Kanno, S. 1971. Tertiary molluscan fauna from the Yakataga District and adjacent areas of southern Alaska. Palaeontological Society of Japan, Special Papers 16: 1–154.
Kiel, S. 2006. New records and species of mollusks from Tertiary cold-seep carbonates in Washington State, USA. Journal of Paleontology 80: 121–137. Crossref
Kiel, S. 2007. Status of the enigmatic fossil vesicomyid bivalve Pleurophopsis. Acta Palaeontologica Polonica 52: 639–642.
Kiel, S. 2008a. An unusual new gastropod genus from an Eocene hydrocarbon seep in Washington State, USA. Journal of Paleontology 82: 188–191. Crossref
Kiel, S. 2008b. Fossil evidence for micro- and macrofaunal utilization of large nekton-falls: examples from early Cenozoic deep-water sediments in Washington State, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 267: 161–174. Crossref
Kiel, S. 2010a. An Eldorado for paleontologists: the Cenozoic seep deposits of western Washington State, USA. In: S. Kiel (ed.), The Vent and Seep Biota. Topics in Geobiology 33: 433–448. Crossref
Kiel, S. 2010b. On the potential generality of depth-related ecologic structure in cold-seep communities: Cenozoic and Mesozoic examples. Palaeogeography, Palaeoclimatology, Palaeoecology 295: 245–257. Crossref
Kiel, S. 2010c. The fossil record of vent and seep mollusks. In: S. Kiel (ed.), The Vent and Seep Biota. Topics in Geobiology 33: 255–278. Crossref
Kiel, S., Aguilar, Y.M., and Kase, T. 2020a. Mollusks from Pliocene and Pleistocene seep deposits in Leyte, Philippines. Acta Palaeontologica Polonica 65: 589–627. Crossref
Kiel, S., Amano, K., and Jenkins, R.G. 2016. Predation scar frequencies in chemosymbiotic bivalves at an Oligocene seep deposit and their potential relation to inferred sulfide tolerances. Palaeogeography, Palaeoclimatology, Palaeoecology 453: 139–145. Crossref
Kiel, S. and Amano, K. 2010. Oligocene and Miocene vesicomyid bivalves from the Katalla district in southern Alaska, USA. The Veliger 51: 76–84.
Kiel, S. and Amano, K. 2013. The earliest bathymodiolin mussels: Evaluation of Eocene and Oligocene taxa from deep-sea methane seep deposits in western Washington State, USA. Journal of Paleontology 87: 589–602. Crossref
Kiel, S. and Hansen, B.T. 2015. Cenozoic methane-seep faunas of the Caribbean region. PLoS ONE 10: e0140788. Crossref
Kiel, S., Fernando, A.G., Magtoto, C.Y., and Kase, T. 2022. Mollusks from Miocene hydrocarbon-seep deposits in the Ilocos-Central Luzon Basin, Luzon Island, Philippines. Acta Palaeontologica Polonica 67: 917–947. Crossref
Kiel, S., Hybertsen, F., Hyžný, M., and Klompmaker, A.A. 2020b. Mollusks and a crustacean from early Oligocene methane-seep deposits in the Talara Basin, northern Peru. Acta Palaeontologica Polonica 65: 109–138. Crossref
Ko, J.Y. 2016. The description of the flat fish (Pleuronectiformes) fossils from the Miocene Duho Formation, Pohang Yeonam-dong in Korea and its implication. Journal of the Korean Earth Science Society 37: 1–10. Crossref
Kojima, S., Fujikura, K., and Okutani, T. 2004. Multiple trans-Pacific migrations of deep-sea vent/seep-endemic bivalves in the family Vesicomyidae. Molecular Phylogenetics and Evolution 32: 396–406. Crossref
Krishtofovich, L.V. 1969. Miocene Mollusca from eastern Kamchatka. Trudy VNIGRI, Paleontlogical Series 4: 172–211.
Krylova, E.M. and Sahling, H. 2006. Recent bivalve molluscs of the genus Calyptogena (Vesicomyidae). Journal of Molluscan Studies 72: 359–395. Crossref
Krylova, E.M. and Sahling, H. 2010. Vesicomyidae (Bivalvia): current taxonomy and distribution. PLoS ONE 5: e9957. Crossref
Krylova, E.M., Gebruk, A.V., Portnova, D.A., Todt, C., and Haflidason, H. 2011. New species of the genus Isorropodon (Bivalvia: Vesicomyidae: Pliocardiinae) from cold methane seeps at Nyegga (Norwegian Sea, Vøring Plateau, Storrega Slide). Journal of the Marine Biological Association of the UK 91: 1135–1144. Crossref
Krylova, E.M., Sahling, H., and Borowski, C. 2018. Resolving the status of the families Vesicomyidae and Kelliellidae (Bivalvia: Venerida), with notes on their ecology. Journal of Molluscan Studies 84: 69–91. Crossref
Krylova, E.M., Sahling, H., and Janssen, R. 2010. Abyssogena: a new genus of the family Vesicomyidae (Bivalvia) from deep water vents and seeps. Journal of Molluscan Studies 76: 107–132. Crossref
Kuechler, R.R., Birgel, D., Kiel, S., Freiwald, A., Goedert, J.L., Thiel, V., and Peckmann, J. 2012. Miocene methane-derived carbonates from southwestern Washington, USA, and a model for silicification at seeps. Lethaia 45: 259–273. Crossref
Linnaeus, C. von 1758. Systema Naturae (tenth edition). 824 pp. Laurrentii Salvii, Holmiae.
Majima, R., Nobuhara, T., and Kitazaki, T. 2005. Review of fossil chemosynthetic assemblages in Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 86–123. Crossref
Malyshkina, T.P., Ward, D.J., Nazarkin, M.V., Nam, G.-S., Kwon, S.-H., Lee, J.-H., Kim, T.-W., Kim, D.-K., and Baek, D.-S. 2022. Miocene Elasmobranchii from the Duho Formation, South Korea. Historical Biology [published online https://doi.org/10.1080/08912963.2022.2110870]. Crossref
Mayer, M.C. 1868. Description de coquilles fossiles des terrains tertiaires supérieurs (suite). Journal de Conchyliologie 16: 102–112.
Meek, F.B. 1876. A report on the invertebrate Cretaceous and Tertiary fossils of the Upper Missouri Country. US Geological Survey of the Territories, Annual Report 9: 1–629.
Minato, M. and Uozumi, S. 1951. Illustrated Cenozoic fossils in North Japan. Tapes from the Poronai Formation and Calyptogena from Morai Formation [in Japanese]. Cenozoic Research 9: 150–151.
Müller, S.P., Kroh, A., Birgel, D., Goedert, J.L., Kiel, S., and Peckmann, J. 2023. Mass occurrence of echinoids in an Oligocene hydrocarbon-seep limestone from the Olympic Peninsula, Washington State, USA. Geological Magazine 160: 941–954. Crossref
Nesbitt, E.A., Martin, R.A., Carroll, N.P., and Grieff, J. 2010. Reassessment of the Zemorrian foraminiferal stage and Juanian molluscan stage north of the Olympic Mountains, Washington State and Vancouver Island. Newsletters on Stratigraphy 43: 275–291. Crossref
Nobuhara, T., Hitomi, S., and Shirotori, Y. 2016. Predatory drill intensity on deep-sea bivalves from the Eocene Poronai Formation, Hokkaido, Japan―Comparison between the cold-seep site and its surrounding normal mud [in Japanese]. Geoscience Reports of Shizuoka University 43: 13–20.
Okutani, T., Kojima, S., and Iwasaki, N. 2002. New and known vesicomyid bivalves recently collected from the western and central Nankai Trough off Shikoku and Honshu, by Deep Sea Research Systems of Japan Marine Science and Technology Center. Venus 61: 129–140.
Oliver, P.G., Rodrigues, C.F., and Cunha, M.R. 2011. Chemosymbiotic bivalves from the mud volcanoes of the Gulf of Cadiz, NE Atlantic, with descriptions of new species of Solemyidae, Lucinidae and Vesicomyidae. Zookeys 113: 1–38. Crossref
Olsson, A.A. 1931. Contributions to the Tertiary paleontology of northern Peru: Part 4, The Peruvian Oligocene. Bulletins of American Paleontology 17: 97–264.
Oyama, K., Mizuno, A., and Sakamoto, T. 1960. Illustrated Handbook of Japanese Paleogene Molluscs. 244 pp. Geological Survey of Japan, Tokyo.
Peckmann, J., Goedert, J.L., Heinrichs, T., Hoefs, J., and Reitner, J. 2003. The late Eocene “Whiskey Creek” methane-seep deposit (western Washington State) Part II: petrology, stable isotopes, and biogeochemistry. Facies 48: 241–254. Crossref
Peckmann, J., Goedert, J.L., Thiel, V., Michaelis, W., and Reitner, J. 2002. A comprehensive approach to the study of methane-seep deposits from the Lincoln Creek Formation, western Washington State, USA. Sedimentology 49: 855–873. Crossref
Peredo, C.M., Uhen, M.D., and Nelson, M.D. 2018. A new kentriodontid (Cetacea: Odontoceti) from the early Miocene Astoria Formation and a revision of the stem delphinidan family Kentriodontidae. Journal of Vertebrate Paleontology 38: e1411357. Crossref
Plafker, G. 1974. Preliminary geologic map of Kayak and Wingham islands, Alaska. USGS Open-file Map 74–82. Crossref
Prashad, B. 1932. The Lamellibranchia of the Siboga expedition. Systematic part II (exclusive of the Pectinidae). Siboga Expeditie 53C: 1–351.
Prothero, D.R., Armentrout, J.M., and Pearson, P. 2001. Magnetic stratigraphy of the upper Middle Eocene (Ulatisian-Narizian) Humptulips Formation, Grays Harbor County, Washington. In: D.R. Prothero (ed.), Magnetic Stratigraphy of the Pacific Coast Cenozoic, 96–104. The Pacific Section SEPM, Santa Fe Springs.
Qu, Y., Teichert, B., Birgel, D., Goedert, J.L., and Peckmann, J. 2017. The prominent role of bacterial sulfate reduction in the formation of glendonite: a case study from Paleogene marine strata of western Washington State. Facies 63: 10. Crossref
Rathbun, M.J. 1932. New species of fossil Raninidae from Oregon. Journal of the Washington Academy of Sciences 22: 239–242.
Rau, W.W. 1964. Foraminifera from the Northern Olympic Peninsula, Washington. US Geological Survey Professional Paper 374G: G1–G33. Crossref
Rau, W.W. 1966. Stratigraphy and Foraminifera of the Satsop River area, southern Olympic Peninsula, Washington. State of Washington Division of Mines and Geology Bulletin 53: 1–66.
Rau, W.W. 1984. The Humptulips Formation—a new Eocene formation of southwest Washington. Washington Geologic Newsletter 12: 1–5.
Rau, W.W. 1986. Geologic Map of the Humptulips Quadrangle and Adjacent Areas, Grays Harbor County, Washington, Map GM–33. Washington State Department of Natural Resources, Olympia.
Sacco, F. 1900. I molluschi dei terreni terziarii del Piemonte e della Liguria. Parte XXVIII (Isocardiidae, Cyprinidae, Veneridae, Petricolidae, Cyrenidae e Sphaeriidae). 98 pp. Carlo Clausen, Torino.
Saul, L.R., Squires, R.L., and Goedert, J.L. 1996. A new genus of cryptic lucinid? bivalve from Eocene cold seeps and turbidite-influenced mudstone, western Washington. Journal of Paleontology 70: 788–794. Crossref
Seong, M.N., Kong, D.-Y., Lee, B.J., and Lee, S.-J. 2009. Cenozoic brittle stars (Ophiuroidea) from the Hagjeon Formation and the Duho Formation, Pohang Basin, Korea. Economic and Environmental Geology 42: 367–376.
Sinelnikova, V.N., Oleinik, A.E., Gladenkov, Y.B., and Titova, L.V.E. 1991. Systematic descriptions. Molluscs [in Russian]. Doklady Akademii Nauk SSSR 467: 119–161, 172–182.
Smith, E.A. 1885. Report on the Lamellibranchiata collected by H.M.S. Challenger during the years 1873–1876. Reports of the Scientific Results of the Challenger Expedition, Zoology 13: 1–341.
Smith, E.A. 1906. Natural History Notes from R.I.M.S. “Investigator” Series III. No.10. On Mollusca from the Bay of Bengal and the Arabian Sea. Annals and Magazine of Natural History 18: 157–175 and 245–264. Crossref
Snavely, P.D.J., Niem, A.R., MacLeod, N.S., Pearl, J.E., and Rau, W.W. 1980. Makah Formation—a deep-marginal-basin sequence of late Eocene and Oligocene age in the northwestern Olympic Peninsula, Washington. US Geological Survey Professional Paper 1162: 1–28. Crossref
Squires, R.L. 1989. Pteropods (Mollusca: Gastropoda) from Tertiary formations of Washington and Oregon. Journal of Paleontology 63: 443–448. Crossref
Squires, R.L. 1991. New morphologic and stratigraphic information on Calyptogena (Calyptogena) gibbera Crickmay, 1929, (Bivalvia: Vesicomyidae), from the Pliocene and Pleistocene of southern California. The Veliger 34: 73–77.
Squires, R.L. 1995. First fossil species of the chemosynthetic-community gastropod Provanna: Localized cold-seep limestones in Upper Eocene and Oligocene rocks, Washington. The Veliger 38: 30–36.
Squires, R.L. and Goedert, J.L. 1991. New Late Eocene mollusks from localized limestone deposits formed by subduction-related methane seeps, southwestern Washington. Journal of Paleontology 65: 412–416. Crossref
Squires, R.L. and Goedert, J.L. 1996. A new species of Thalassonerita? (Gastropoda: Neritidae?) from a Middle Eocene cold-seep carbonate in the Humptulips Formation, Western Washington. The Veliger 39: 270–272.
Squires, R.L. and Gring, M.P. 1996. Late Eocene chemosynthetic? bivalves from suspect cold seeps, Wagonwheel Mountain, central California. Journal of Paleontology 70: 63–73. Crossref
Sturany, R. 1896. Mollusken I (Prosobranchier und Opisthobranchier; Scaphopoden; Lamellibranchier) gesammelt von S.M. Schiff “Pola” 1890–94. Denkschriften der mathematisch-naturwissenschaftlichen Klasse der Kaiserlichen Akademie der Wissenschaften 63: 1–36.
Tabor, R.W. and Cady, W.M. 1978. Geologic map of the Olympic Peninsula. US Geological Survey Miscellaneous Investigations Series Map I-994: scale 1:125 000. U.S. Geological Survey, Washington, DC
Takeda, H. 1953. The Poronai Formation (Oligocene Tertiary) of Hokkaido and South Sakhalin and its fossil fauna. Studies on Coal Geology 3: 1–103.
Tucker, A.B. 1998. Systematics of the Raninidae (Crustacea: Decapoda: Brachyura), with accounts of three new genera and two new species. Proceedings of the Biological Society of Washington 111: 320–371.
Valentine, J.W., Jablonski, D., Kidwell, S.M., and Roy, K. 2006. Assessing the fidelity of the fossil record by using marine bivalves. Proceedings of the National Academy of Sciences of the USA 103: 6599–6604. Crossref
Van Winkle, K. 1919. Remarks on some new species from Trinidad. Bulletins of American Paleontology 8: 19–27.
Weaver, C.E. 1942. Paleontology of the marine Tertiary formations of Oregon and Washington. University of Washington Publications in Geology 5: 1–789.
Wells, R.E. 1989. Geologic map of the Cape Disappointment—Naselle River area, Pacific and Wahkiakum counties, Washington. US Geological Survey Miscellaneous Investigations Map I–1832. U.S. Geological Survey, Washington, DC.
Wolfe, E.W. and McKee, E.H. 1968. Geology of the Grays River Quadrangle, Wahkiakum and Pacific counties, Washington. Geologic Map GM-4. Washington State Department of Natural Resources, Olympia.
Wolfe, E.W. and McKee, E.H. 1972. Sedimentary and igneous rocks of the Grays River quadrangle, Washington. United States Geological Survey Bulletin 1335: 1–70.
Woodring, W.P. 1925. Miocene Mollusca from Bowden Jamaica, pelecypods and scaphopods. Carnegie Institution of Washington Publications 336: 1–564. Crossref
Woodring, W.P. 1938. Lower Pliocene mollusks and echinoids from the Los Angeles basin, California, and their inferred environment. US Geological Survey Professional Paper 190: 1–67. Crossref
Yamaguchi, T., Goedert, J.L., and Kiel, S. 2016. Marine ostracodes from Paleogene hydrocarbon seep deposits in Washington State, USA and their ecological structure. Geobios 49: 407–422. Crossref
Yokoyama, M. 1890. Versteinerungen aus der japanischen Kreide. Palaeontographica 36: 159–202.
Yokoyama, M. 1928. Mollusca from the oil-field of the island of Taiwan. Report of the Imperial Geological Survey of Japan 101: 1–112.
Yoon, S. 1976a. Geology and paleontology of the Tertiary Pohang basin, Pohang district, Korea. Part 2, Paleontology (Mollusca) No. 1, systematic description of Bivalvia. The Journal of the Geological Society of Korea 12: 1–22.
Yoon, S. 1976b. Geology and paleontology of the Tertiary Pohang basin, Pohang district, Korea. Part 2, Paleontology (Mollusca) No. 2, systematic description of Scaphopoda and Gastropoda, with descriptions of fossil localities. The Journal of the Geological Society of Korea 12: 63–78.
Yun, C.-G. 2020. New example of Cosmopolitodus hastalis (Lamniformes, Lamnidae) from the Miocene of South Korea Zoodiversity 54: 433–438. Crossref
Yun, C.-G. 2021. First deep-sea shark fossil teeth from the Miocene of South Korea. Zoodiversity 55: 225–232. Crossref
Yun, H. 1986. Emended stratigraphy of the Miocene formations in the Pohang Basin, Part 1. Journal of the Paleontological Society of Korea 2: 54–69.
Acta Palaeontol. Pol. 68 (2): 297–320, 2023
https://doi.org/10.4202/app.01061.2023