
A well-preserved vertebra provides new insights into rebbachisaurid sauropod caudal anatomical and pneumatic features
GUILLERMO J. WINDHOLZ, JUAN D. PORFIRI, DOMENICA DOS SANTOS, FLAVIO BELLARDINI, and MATHEW J. WEDEL
Windholz, G.J., Porfiri, J.D., Dos Santos, D., Bellardini, F., and Wedel, M.J. 2024. A well-preserved vertebra provides new insights into rebbachisaurid sauropod caudal anatomical and pneumatic features. Acta Palaeontologica Polonica 69 (1): 39–47.
Rebbachisauridae is a clade of sauropod dinosaurs whose maximum diversification and abundance are known from the Cretaceous of South America. We describe an anterior caudal vertebra, MDPA-Pv 007, from the Upper Cretaceous of Argentine Patagonia, whose characters allow it to be referred to this clade. Also, two phylogenetic analyses reinforce the referral of the new material more exclusively to Rebbachisaurinae. We analyze pneumatic structures using the first CT scans of a caudal element of a rebbachisaurid. The excellent preservation of MDPA-Pv 007, combined with CT images, allows us to document external fossae and foramina that connect to larger internal chambers, constituting unambiguous evidence of pneumaticity. The centrum of MDPA-Pv 007 is camerate, with large interconnected internal chambers; this is accompanied by a neural arch with wide and deep fossae. Caudal pneumaticity has a complex phylogenetic distribution among neosauropods. This feature may have evolved independently in diplodocoids and titanosauriforms, or it could be ancestral for Neosauropoda but secondarily lost in a few lineages. Future investigations, taking advantage of new technologies, will provide insights into the phylogenetic distribution and paleobiological implications of pneumaticity in sauropod dinosaurs and other fossil archosaurs.
Key words: Dinosauria, Sauropoda, Diplodocoidea, Rebbachisauridae, pneumaticity, Cretaceous, Neuquén Basin, Argentina.
Guillermo J. Windholz [gwindholz@unrn.edu.ar; ORCID: https://orcid.org/0000-0002-3240-2392 ] and Flavio Bellardini [flaviobellardini@gmail.com; ORCID: https://orcid.org/0000-0003-1616-8548 ], Universidad Nacional de Río Negro, Instituto de Investigación en Paleobiología y Geología, Río Negro, Argentina; IIPG, UNRN, Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), Av. Roca 1242, (R8332EXZ) General Roca, Río Negro, Argentina; Museo Carlos Ameghino, Belgrano 1700, Paraje Pichi Ruca (predio Marabunta), Cipolletti, Río Negro, 8300, Argentina.
Juan D. Porfiri [jporfiri@gmail.com; ORCID: https://orcid.org/0000-0003-1720-5961 ] and Domenica Dos Santos [domenicasantos@gmail.com; ORCID: https://orcid.org/0000-0002-3247-4660 ], Museo del Desierto Patagónico, Calle 6 S/N, Añelo, Neuquén, 8305, Argentina; Museo de Ciencias Naturales, Universidad Nacional del Comahue, Buenos Aires 1400, Neuquén, 8300, Argentina; Catedra de Reptiles Mesozoicos, Facultad de Ingeniería, Universidad Nacional del Comahue, Buenos Aires 1400, Neuquén, 8300, Argentina.
Mathew J. Wedel [mathew.wedel@gmail.com; ORCID: https://orcid.org/0000-000 1-6082-3103 ] College of Osteopathic Medicine of the Pacific and College of Podiatric Medicine, Western University of Health Sciences, Pomona, California, 91711, USA.
Received 31 August 2023, accepted 19 January 2024, published online 9 February 2024.
Copyright © 2024 Guillermo J. Windholz et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Rebbachisauridae is a clade of sauropod dinosaurs that probably originated in the Jurassic but which flourished during the Cretaceous (Salgado et al. 2022). Fossils of rebbachisaurids are primarily known from Africa, Europe and South America. A record of a rebbachisaurid from Asia has recently been reported (Averianov and Sues 2021), but its taxonomic assignment has been questioned (Lerzo et al. 2021), and the enigmatic North American taxon Maraapunisaurus (formerly Amphicoelias) fragillimus has been referred to Rebbachisauridae, albeit on the basis of missing material (Carpenter 2018). Despite the broad paleogeographic distribution of the group, the greatest diversity and abundance of rebbachisaurid sauropods are known from the Cretaceous of South America, especially in Neuquén Basin from Argentina (Calvo and Salgado 1995; Bonaparte 1996, 1997; Dalla Vecchia 1998; Sereno et al. 1999; Carvalho et al. 2003; Gallina and Apesteguía 2005; Salgado et al. 2006; Apesteguía 2007; Taylor and Naish 2007; Carballido et al. 2010; Torcida et al. 2011; Salgado et al. 2012; Fanti 2013, 2015; Ibiricu et al. 2013, 2015; Canudo et al. 2018; Lindoso et al. 2019; Bellardini et al. 2022).
The Candeleros Formation (Cenomanian, Upper Cretaceous) presents extensive outcrops in Argentine Patagonia (Garrido 2010). This stratigraphic unit has provided a diverse fauna of fossil reptiles and other continental vertebrates from the Neuquén Basin, among which the sauropod dinosaurs stand out. Candeleros Formation sauropods include the titanosaur Andesaurus delgadoi (Calvo and Bonaparte 1991) and the rebbachisaurids Limaysaurus tessonei (Calvo and Salgado 1995), Nopcsaspondylus alarconensis (Apesteguía 2007), and Rayososaurus agrioensis (Bonaparte 1996; Carballido et al. 2010). The anterior caudal vertebrae described herein, MDPA-Pv 007, is from the Candeleros Formation and is referred to the family Rebbachisauridae.
Birds are the only clade of extant amniotes with postcranial skeletal pneumaticity (PSP). This means that the interior of the bones is invaded by diverticulae from the lungs and air sacs of the respiratory system; these pneumatic diverticulae induce bone resorption to produce internal cavities or chambers filled with air. Among fossil vertebrates, unambiguous PSP is known only in pterosaurs and saurischian dinosaurs, including sauropods (Britt 1997; Wedel 2003, 2005; O’Connor 2006; Claessens et al. 2009). Among sauropods, rebbachisaurids have been characterized as having highly pneumatic vertebrae, based on external fossae and foramina (Fanti et al. 2015) revealed by natural fractures (Windholz et al. 2022a) as well as computed tomography (CT) scans of dorsal vertebrae of Nigersaurus taqueti (Sereno et al. 2007) and Katepensaurus goicoecheai (Ibiricu et al. 2013). Here we present the first CT scans of a caudal element of a rebbachisaurid. The vertebra described herein records exquisite laminar architecture, which alongside the CT scan images, provides new anatomical and paleobiological information on rebbachisaurid sauropods.
Institutional abbreviations.—MAU, “Museo Municipal Argentino Urquiza”, Rincón de los Sauces, Argentina; MDPA, Museo del Desierto Patagónico de Añelo, Argentina; MIWG, Museum of Isle of Wight Geology (now Dinosaur Isle Visitor Centre), Sandown, Isle of Wight, UK; NHMUK, Natural History Museum, London, UK; UNPSJB, Universidad Nacional de la Patagonia San Juan Bosco, Comodoro Rivadavia, Argentina.
Other abbreviations.—acdl, anterior centrodiapophyseal lamina; amedl, anterior medial lamina; cdf, centrodiapophyseal fossa; cpol, centropostzygapophyseal lamina; cprl, centroprezygapophyseal laminae; nc, neural canal; pcdl, posterior centrodiapophyseal lamina; pmedl, posterior medial lamina; pocdf, postzygapophyseal centrodiapophyseal fossa; pocdf-l, postzygapophyseal centrodiapophyseal fossa lamina; podl, postzygodiapophyseal lamina; posdf, postzygapophyseal spinodiapophyseal fossa; prcdf, prezygapophyseal centrodiapophyseal fossa; prcdf-l, prezygapophyseal centrodiapophyseal fossa lamina; prdl, prezygodiapophyseal lamina; prsdf, prezygapophyseal spinodiapophyseal fossa; PSP, postcranial skeletal pneumaticity; pz, postzygapophyses; spof, spinopostzygapophyseal fossa; sprl, spinoprezygapophyseal laminae.
Material and methods
The element described herein, MDPA-Pv 007, is a previously unpublished rebbachisaurid caudal vertebra from the Sierra Chata locality (Neuquén Province) (Fig. 1), Candeleros Formation, Cenomanian (Upper Cretaceous) (Garrido 2010). It is housed in the “Museo del Desierto Patagónico de Añelo”, located in Añelo city, Neuquén Province, Argentina. The serial position was inferred by comparison with the well-preserved vertebral caudal sequences of Limaysaurus (Calvo and Salgado 1995; Salgado et al. 2004). Based on the size and development of the wing-like transverse processes, we interpret this element as one of the most anterior caudal vertebrae.
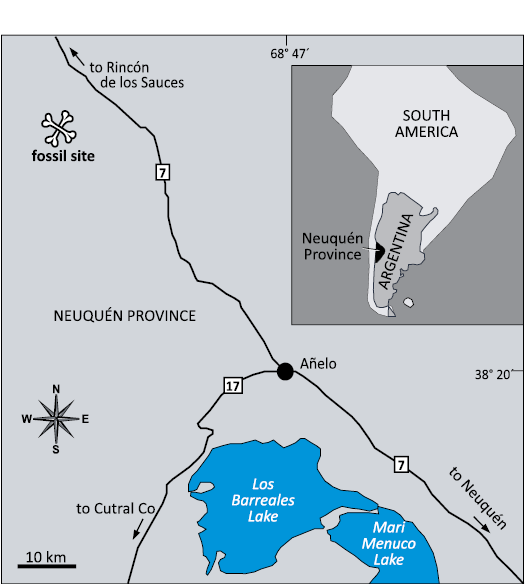
Fig. 1. Maps showing location of Sierra Chata fossil site within Neuquén Province, Argentina.
The nomenclature for sauropod vertebral laminae (Wilson 1999, 2012) and fossae (Wilson et al. 2011) are adopted in osteological description. In addition, the osteological correlates linked to pneumaticity are described, on both the external and internal surfaces of the bone. The internal anatomy has been characterized from CT images, representing the first published CT scan of a rebbachisaurid caudal vertebra. Here we follow the nomenclature for pneumatic structures proposed by Wedel et al. (2000), Wedel (2003) and O’Connor (2006). The CT scans were carried out in “Clínica de Imágenes SA” located in Neuquén city, Neuquén Province, Argentina. Scans were obtained using a SIEMENS Somatom Definition Edge CT scanner, with a slice thickness of 0.75 mm at 0.8 mm intervals.
Results
External anatomy.—MDPA-Pv 007 is well preserved, possibly the 1st or 2nd caudal vertebra. The centrum is platycoelous, slightly wider than high, with a subcircular outline in anterior and posterior views (Fig. 2A1–A6). In lateral view, the centrum is anteroposteriorly short and lacks lateral fossae or pleurocoels. However, the centrum has several foramina irregularly distributed on its lateral and lateroventral surfaces (Fig. 2A7–A9). Such foramina are present on caudal vertebrae of many other neosauropod species (e.g., belonging to Malawisaurus, Rocasaurus, Saltasaurus, and Tornieria) including rebbachisaurids belonging to Tataouinea and Comahuesaurus (Carballido et al. 2012; Cerda et al. 2012; Wedel and Taylor 2013; Fanti et al. 2015; Ibiricu et al. 2015; Zurriaguz and Cerda 2017). The ventral surface is concave, with the posteroventral margin narrower and more ventrally prominent than the anteroventral one in lateral view. The entrance to the neural canal has an elliptical outline, taller dorsoventrally than wide.
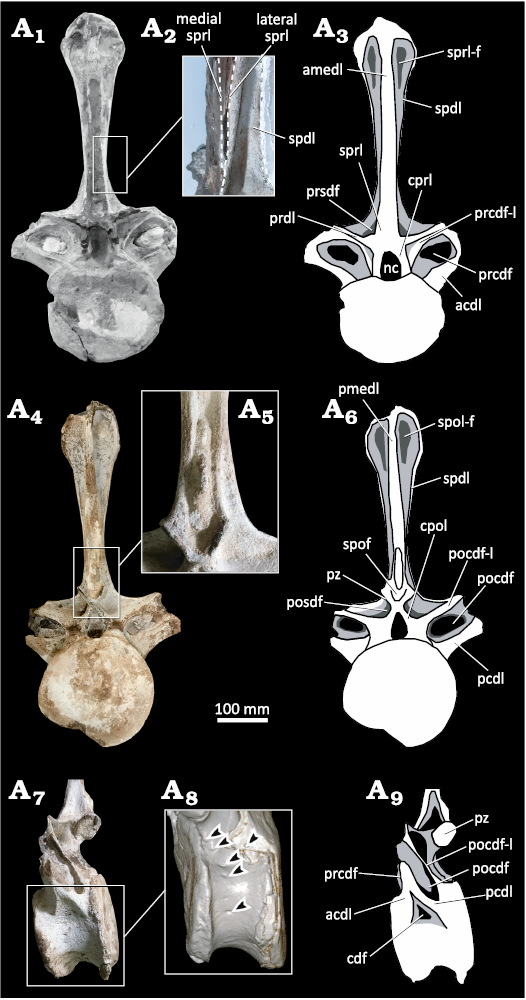
Fig. 2. Rebbachisauridae indet. (MDPA-Pv 007) from the Sierra Chata locality (Candeleros Formation) Cenomanian (Upper Cretaceous). Anterior caudal vertebra in anterior (A1, A3), posterior (A4, A6), and left lateral (A7, A9) views. Close ups showing lateral spinal laminae (A2), accessory bony lamina located inside of spof (A5), foramina in the lateral surface of the centrum, arrowheads indicate the presence of foramina (A8). Abbreviations: acdl, anterior centrodiapophyseal lamina; amedl, anterior medial lamina; cdf, centrodiapophyseal fossa; cpol, centropostzygapophyseal lamina; cprl, centroprezygapophyseal laminae; nc, neural canal; pcdl, posterior centrodiapophyseal lamina; pmedl, posterior medial lamina; pocdf, postzygapophyseal centrodiapophyseal fossa; pocdf-l, postzygapophyseal centrodiapophyseal fossa lamina; posdf, postzygapophyseal spinodiapophyseal fossa; prcdf, prezygapophyseal centrodiapophyseal fossa; prcdf-l, prezygapophyseal centrodiapophyseal fossa lamina; prdl, prezygodiapophyseal lamina; prsdf, prezygapophyseal spinodiapophyseal fossa; pz, postzygapophyses; spof, spinopostzygapophyseal fossa; spdl, spinodiapophyseal lamina; spol-f, spinopostzygapophyseal lamina fossa; sprl, spinoprezygapophyseal laminae; sprl-f, spinoprezygapophyseal lamina fossa.
The transverse processes are dorsoventrally tall, anteroposteriorly short, and inclined dorsally 25° from the horizontal axis. Transverse processes laterodorsally oriented in anterior caudal vertebrae is a condition also presents in other non-diplodocid diplodocoids as Rebbachisauridae and Dicraeosauridae (e.g., Janensch 1929; Calvo and Salgado 1995; Salgado et al. 2004; Mannion and Barrett 2013; Ibiricu et al. 2013, 2015; Windholz et al. 2022b). In MDPA-Pv-007, the transverse processes are composed of two bone bars: a ventral, robust and laterodorsally oriented bar; and a dorsal, slender and lateroventrally inclined bar (Fig. 2A1–A6). These bars converge and contact apically, enclosing a system of neural laminae, fossae and fenestrae. The presence of a composite and perforated transverse process is an atypical condition within Sauropoda, being recorded in the first caudal vertebrae of Limaysaurus tessonei, and considered as an autapomorphic feature by Calvo and Salgado (1995). Despite sharing this morphological feature, the comparisons between specimens included into Limaysaurus tessonei and MDPA-Pv-007 are limited by the poorly preserved anterior caudal vertebrae of the former, which lack the set of neural structures seen in MDPA-Pv-007.
In anterior view, the neural arch presents a wide, elliptic and laterodorsally elongated prezygapophyseal centrodiapophyseal fossa (prcdf), which is dorsally framed by a thin and prominent prezygodiapophyseal lamina (prdl) and ventrally by a robust anterior centrodiapophyseal lamina (acdl). Close to the medial margin of the prcdf, there is a wide prezygapophyseal centrodiapophyseal fossa-lamina (prcdf-l). Obliquely oriented prcdf-l are a feature also present in anterior caudal vertebrae of rebbachisaurids Itapeuasaurus cajapioensis, Katepensaurus goicoecheai, Rebbachisauridae indet. NHMUK R36636 and Rebbachisauridae indet. MIWG 5384 (Mannion et al. 2011; Mannion and Barrett 2013; Ibiricu et al. 2013, 2015; Lindoso et al. 2019). Articular facets of the prezygapophyses are not preserved. In lateral view, the base of neural arch bears a shallow triangular-shaped centrodiapophyseal fossa (cdf). In posterior view, the neural arch bears well-developed posterior centrodiapophyseal (pcdl) and postzygapophyseal centrodiapophyseal fossa (pocdf-l) laminae. These laminae frame a wide postzygapophyseal centrodiapophyseal fossa (pocdf). Additionally, postzygapophyseal centrodiapophyseal fossa (pocdf-l), centropostzygapophyseal (cpol) and postzygodiapophyseal (podl) laminae, delimit a broad, triangular postzygapophyseal spinodiapophyseal fossa (posdf). The articular facets of postzygapophyses diverge ventrolaterally at an angle of 290°. The vertebra lacks a hyposphenal ridge, as in the anterior caudal vertebrae of some titanosauriforms and most rebbachisaurids (Mannion et al. 2011).
The neural spine is simple, unlike the anterior caudal vertebrae of some diplodocids (whose anterior caudal neural spines are slightly bifurcate distally, e.g., Barosaurus and Diplodocus, Wedel and Taylor 2013); and tall, at least twice as tall as the vertebral centrum (Table 1), as occurs in most diplodocoids, including rebbachisaurid members. The neural spine is petal-shaped, with a proximal transversal constriction and presents an abrupt distal expansion (triangular lateral process), which is nearly twice as wide transversely as the neural spine base. The transversal constriction of the neural spine is proximal, close to the base of the spine, as in several rebbachisaurids (e.g., Amazonsaurus maranhensis, Itapeuasaurus cajapioensis, Limaysaurus tessonei, Tataouinea hannibalis, and Rebbachisauridae indet. MAU-PV-EO-666), and differing from Demandasaurus darwini, Rebbachisauridae indet. UNPSJB-PV 580 and Rebbachisauridae indet. MIWG 5384, where the minimum width is more distal, over the mid-height of the spine (Fanti et al. 2013, 2015; Bellardini et al. 2022). On the other hand, triangular lateral processes are also present in caudal vertebrae of Demandasaurus darwini, Katepensaurus goicoecheai, Nigersaurus taqueti, Rebbachisaurus garasbae, Zapalasaurus bonapartei, Rebbachisauridae indet. MIWG 5384, and Rebbachisauridae indet. MAU-PV-EO-666 (Mannion et al. 2011; Whitlock 2011; Ibiricu et al. 2013). In anterior view, the neural spine has a robust anterior medial lamina (amedl) and a pair of bifid spinoprezygapophyseal laminae (medial and lateral sprl) that medially frame wide prezygapophyseal spinodiapophyseal fossa (prsdf). In posterior view the posterior medial lamina (pmedl) is promiment. Unlike caudal vertebrae of other rebbachisaurid sauropods, MDPA-Pv 007 bears an accessory bony lamina located inside the spinopostzygapophyseal fossa (spof). In lateral view, the neural spine is directed backwards as occurs in most anterior caudal vertebrae of rebbachisaurids (Pereda Suberbiola et al. 2003; Gallina and Apesteguía 2005; Torcida et al. 2011; Ibiricu et al. 2013, 2015). Furthermore, the neural spine of MDPA-Pv 007 is straight rather than curved, as in anterior caudal vertebra of specimens belonging to Amazonsaurus maranhensis (Carvalho et al. 2003), Itapeuasaurus cajapioensis (Lindoso et al. 2019), Katepensaurus goicoecheai (Ibiricu et al. 2013), Limaysaurus tessonei (Calvo and Salgado 1995; Salgado et al. 2004), and Tataouinea hannibalis (Fanti et al. 2013). In anterior view, the apex of the neural spine is convex, as in most of Rebbachisauridae (e.g., Amazonsaurus maranhensis, Demandasaurus darwini, Nigersaurus taqueti, Rebbachisaurus garasbae, Rebbachisauridae indet. MIGW 5384; Sereno et al. 1999; Carvalho et al. 2003; Mannion et al. 2011; Torcida et al. 2011; Wilson and Allain 2015), differing from Zapalasaurus bonapartei and Rebbachisauridae indet. MAU-PV-EO-666 whose the dorsal surface of the spine is slightly transversely concave.
Table 1. Measurements (in mm) of new specimen MDPA-Pv 007.
|
Total height |
395 |
|
Centrum length |
95 |
|
Neural spine height |
315 |
|
Maximum neural spine width |
110 |
|
Minimum neural spine width |
35 |
|
Anterior centrum height |
150 |
|
Anterior centrum width |
165 |
|
Posterior centrum height |
145 |
|
Posterior centrum width |
155 |
|
Minimum ventral width of the centrum |
130 |
|
Postzygapophyses length |
40 |
|
Postzygapophyses width |
30 |
Internal anatomy.—The ventral region of the centrum is thick and solid (Fig. 3). However, the dorsal half of the vertebral centrum is invaded by large, interconnected, enclosed camerae (Figs. 3, 4A1–A4). The presence of interconnecting large internal chambers (camerae) and relatively thick-walled centra was termed camerate (Britt 1997; Wedel 2003). This form of vertebral pneumatization differs substantially from the typical “camellate” internal structures (Britt 1997) displayed by caudal vertebrae of some saltasaurines as Rocasaurus and Saltasaurus (Cerda et al. 2012; Zurriaguz and Cerda 2017), “procamerate” condition of diplodocid Tornieria (as seen in Wedel and Taylor 2013: fig. 1); and the apneumatic centra of the basal lithostrotian Malawisaurus (Wedel 2009: fig. 2) and derived dicraeosaurids as Pilmatueia (Windholz et al. 2019). The internal camerae in the centrum of MDPA-Pv 007 communicate with foramina on the external surface (Figs. 2A7–A9, 3A4).
The above described internal camerae of the centrum connect internally with the neural arch (Fig. 4A6); however, no pneumatic foramina connect to the neural canal, as has been documented in the brachiosaurid Giraffatitan (Schwarz and Fritsch 2006) and the saltasaurine Ibirania (Aureliano et al. 2021). The region lateral to the neural canal bears internal camerae that are connected to each other, and simultaneously, with the external surface of the bone (Fig. 4A1–A3). Neural fossae are broad and deep, reaching almost to the axial plane of the element (Fig. 4A7, A8). The neural spine presents deep fossae, resulting in a tetraradiate outline in cross section. Internally, it presents a variation in bone tissue density that corresponds to small chambers that connect with the rest of the neural arch (Fig. 4A9, A10).
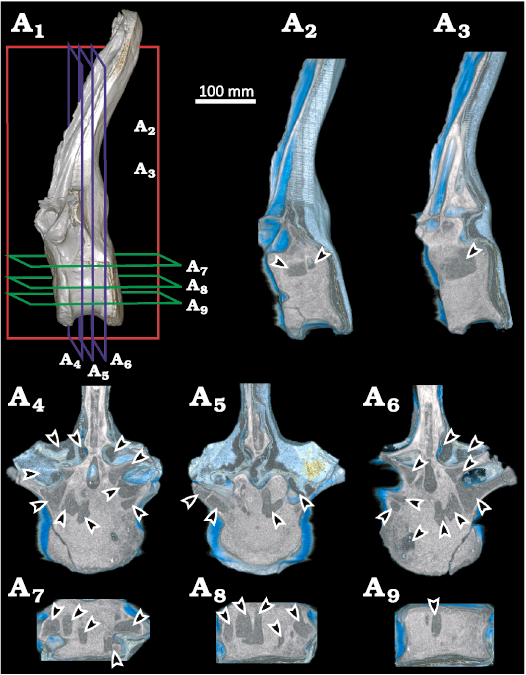
Fig. 3. Three-dimensional reconstruction of Rebbachisauridae indet. (MDPA-Pv 007) from the Sierra Chata locality (Candeleros Formation) Cenomanian (Upper Cretaceous). Vertebra in lateral view (A1), parasagittal sections (A2, A3), transverse sections (A4–A6), frontal sections (A7–A9). Arrowheads show the presence of pneumatic internal cameras.
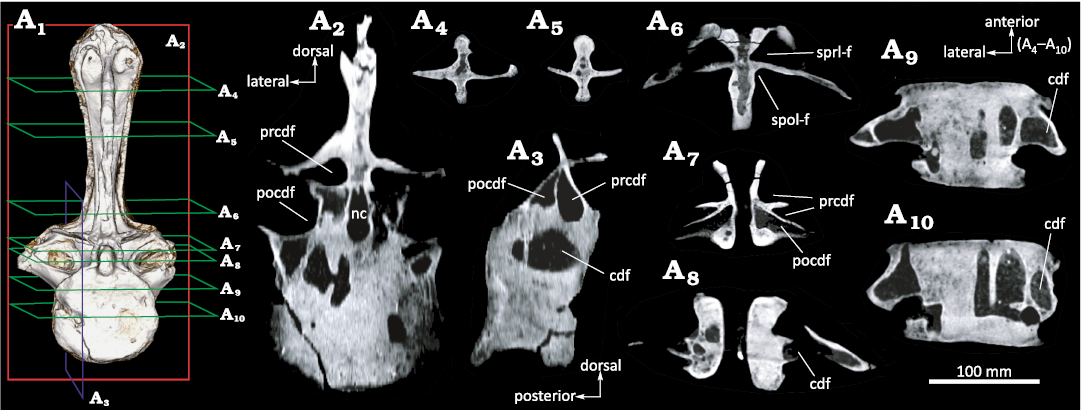
Fig. 4. Selected computed tomographic sections of Rebbachisauridae indet. (MDPA-Pv 007) from the Sierra Chata locality (Candeleros Formation) Cenomanian (Upper Cretaceous). Vertebra in anterior view (A1), transverse section taken at mid-length of the element (A1), parasagittal section (A3), frontal sections (A4–A10). Abbreviations: cdf, centrodiapophyseal fossa; nc, neural canal; pocdf, postzygapophyseal centrodiapophyseal fossa; prcdf, prezygapophyseal centrodiapophyseal fossa; spol-f, spinopostzygapophyseal lamina fossa; sprl-f, spinoprezygapophyseal lamina fossa.
Phylogenetic context in Bellardini et al. (2022; Appendix 1).—We coded 23 characters observed on MDPA-Pv 007 into the character matrix of Bellardini et al. (2022) to determine its phylogenetic affinities. The data matrix has 418 characters distributed in 95 OTUs (Operational Taxonomic Units) counting MDPA-Pv 007. A heuristic tree search was carried out using TNT v.1.6 software (Goloboff and Catalano 2016). The cladistic analysis retrieved 390 most parsimonious trees (MPTs) of 1429 steps, consistency index (CI) = 0.341; retention index (RI) = 0.692. The obtained trees underwent an extra round of branch swapping (TBR), resulting in 10000 trees of 1429 steps. MDPA-Pv 007 was recovered well nested among the rebbachisaurids (Fig. 5A). In general, Rebbachisauridae is well-resolved in this analysis. However, two small polytomies are recovered, but they do not involve the specimen under study; one occurs among the basal rebbachisaurids (Comahuesaurus, Lavocatisaurus, Zapalasaurus, and Histriasaurus) and the other one among more derived genera (Demandasaurus, Nigersaurus, Tataouinea, and Rebbachisaurus).
The inclusion of MDPA-Pv 007 among the rebbachisaurids is based on the absence of the hyposphenal ridge (character 241, shared state with some macronarians; among rebbachisaurids: apomorphic state in Demandasaurus and Tataouinea; unknown in Histriasaurus, Rayososaurus, and Rebbachisaurus); and the presence of the spinodiapophyseal lamina (character 244, shared with Lognkosauria; unknown in rebbachisaurids Comahuesaurus, Lavocatisaurus, Zapalasaurus, Histriasaurus, Rayososaurus, and Limaysaurus) in anterior caudal vertebrae. At the same time, the analysis shows MDPA-Pv 007 as the most basal member of the subfamily Rebbachisaurinae. Thus, MDPA-Pv 007 is recovered in sister taxon to a clade containing Katepensaurus and all more derived rebbachisaurids. This is based on the presence of triangular lateral processes in the neural spines of anterior and middle caudal vertebrae (character 235, also present in Losillasaurus and unknown in the Rebbachisaurus).
Phylogenetic context in Windholz et al. (2022b; Appendix 2).—We coded 34 characters observed on MDPA-Pv 007, resulting in a matrix of 494 characters distributed in 40 OTUs. The cladistic analysis was carried out with the same software and parameters as the previous analysis, retrieved 4 MPTs of 1595 steps (CI = 0.345 and RI = 0.547). Rebbachisauridae is well-resolved in this analysis, with MDPA-Pv 007 well nested among them as a Rebbachisaurinae member (Fig. 5B).
The inclusion of MDPA-Pv 007 among rebbachisaurids is based on the presence of “petal” shaped caudal neural spines (character 304); anterior caudal neural spine, not including arch, more than 1.5 times centrum height (character 312); and transverse processes of anterior caudal vertebrae directed laterodorsally (character 322). Furthermore, MDPA-Pv 007 shares with other Rebbachisaurinae members the presence of triangular lateral processes in caudal neural spines (character 303).
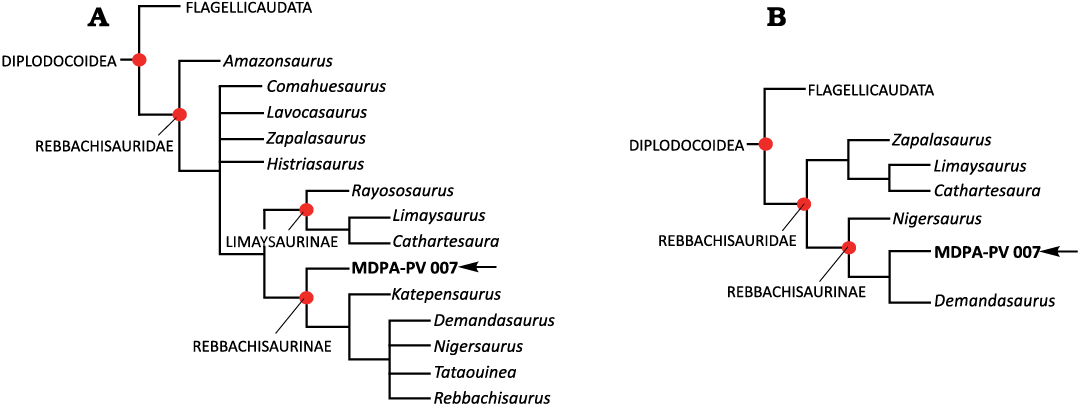
Fig. 5. Simplified strict consensus showing the position of new specimen MDPA-Pv 007 among rebbachisaurids coded in: Bellardini et al. 2022 (A) and Windholz et al. 2022b (B).
Discussion
MDPA-Pv 007 presents characters that allow it to be referred to the diplodocoid sauropods, including anterior caudal vertebrae with broad, “wing-like” transverse processes, and the presence of a tall neural spine, at least twice as tall as the respective vertebral centrum. Furthermore, it shows affinities with the family Rebbachisauridae, such as laterodorsally oriented transverse processes, and lobed-shaped neural spines with a tetraradiate outline in cross section (Whitlock 2011; Bellardini et al. 2022). The phylogenetic analyses reinforces the referral of the new material to Rebbachisauridae, and more exclusively to Rebbachisaurinae. MDPA-Pv 007 shares synapomorphies with other rebbachisaurids, but differs from all other known rebbachisaurids in having an accessory bony lamina located inside the spinopostzygapophyseal fossa (spof). However, given that MDPA-Pv 007 is a single isolated element, it seems prudent to treat it as Rebbachisaurinae indet., as opposed to erecting a new taxon. Additional specimens may further elucidate the specimen’s phylogenetic and taxonomic status. Nevertheless, MDPA-Pv 007 suggests that the diversity of rebbachisaurids in the Candeleros Formation is greater than currently recognized.
MDPA-Pv 007 adds to the known fossil record of rebbachisaurid genera from the Candeleros Formation, so far represented by Limaysaurus (Calvo and Salgado 1995), Nopcsaspondylus (Apesteguía 2007), Rayososaurus (Bonaparte 1996; Carballido et al. 2010) and some remains of Rebbachisauridae indet. (Paulina Carabajal et al. 2016). These records, plus those from the Alcântara Formation (Lindoso et al. 2019), Bajo Barreal Formation (Ibiricu et al. 2012, 2015), and Huincul Formation (Calvo and Salgado 1995; Gallina and Apesteguía 2005; Apesteguía et al. 2010; Haluza et al. 2012; Bellardini et al. 2022) show that representatives of this family constituted one of the most diverse sauropod clades during the Cenomanian of South America. This caudal vertebra is the first record of a rebbachisaurid sauropod in the Sierra Chata locality. We noted that, despite the extensive record of rebbachisaurids, MDPA-Pv 007 is possibly the best preserved anterior caudal vertebra of the family. The excellent preservation of MDPA-Pv 007, combined with CT images, allows us to document external fossae and foramina that connect to larger internal chambers. Together, these features constitute unambiguous evidence of pneumaticity (O’Connor 2006), and the best documentation to date of caudal pneumaticity in rebbachisaurids (see Wedel and Taylor 2013 and references therein).
The centrum of MDPA-Pv 007 is camerate, with large interconnected internal chambers and extensive fossae on the neural arch. The dorsal half of the centrum and the lower portion of the neural arch are extensively pneumatized, but internal pneumatization of both the centrum and the neural arch and spine is less extensive than in the caudal vertebrae of saltasaurine sauropods such as Rocasaurus and Saltasaurus (Cerda et al. 2012; Zurriaguz and Cerda 2017). Both the external foramina and the internal pneumatic chambers in MDPA-Pv 007 are bilaterally asymmetric, as often occurs in other sauropods with caudal pneumaticity (Wedel and Taylor 2013). This asymmetry is consistent with the hypothesis that the pneumatization process was essentially opportunistic (Witmer 1997), possibly as a result of developing pneumatic diverticulae following inherently variable blood vessels (Taylor and Wedel 2021). Although most rebbachisaurids are small- to medium-sized sauropods, they were still large terrestrial animals; e.g., estimates for Rebbachisaurus garasbae suggest a body mass of 7915–12 015 kg (Wilson and Allain 2015), while the estimated length for Tataouinea hannibalis was 12 meters (Fanti et al. 2015); Maraapunisaurus fragillimus would have been one of the largest known sauropods if it was as large as claimed (Carpenter 2018). Extensive skeletal pneumaticity, as shown by the new specimen and by other rebbachisaurids, is consistent with the hypothesis that by reducing bone mass, the muscular energy necessary to propel the body would consequently be reduced (O’Connor 2009; Ibiricu et al. 2015).
Caudal pneumaticity has a complex phylogenetic distribution among neosauropods (Wedel and Taylor 2013). Pneumatic tail vertebrae have been documented in numerous brachiosaurids (Giraffatitan, Cedarosaurus, Venenosaurus) and titanosaurians, especially saltasaurids (Neuquensaurus, Rocasaurus, and Saltasaurus), and in virtually all diplodocoids, including Haplocanthosaurus, diplodocids, and rebbachisaurids, but excluding dicraeosaurids (Cerda et al. 2012; Fanti et al. 2013; Wedel and Taylor 2013). Caudal pneumaticity may have evolved independently in diplodocoids and titanosauriforms, or it could be ancestral for Neosauropoda but secondarily lost in a few lineages (e.g., Dicraeosauridae, Camarasaurus). We note that even in clades in which caudal pneumaticity is the norm, certain taxa lack it, presumably through phylogenetic reversals; examples include Nigersaurus among Rebbachisauridae and Opisthocoelicaudia among derived titanosaurs. It should be noted, however, that there are few broad studies of caudal pneumaticity in sauropods. Future investigations, taking advantage of new technologies, will hopefully allow us to further unravel the phylogenetic distribution and paleobiological implications of pneumaticity in sauropod dinosaurs and other fossil archosaurs.
Conclusions
We present a new caudal vertebra MDPA-Pv 007 from Candeleros Formation (Cenomanian, Upper Cretaceous) Patagonia, Argentina. External anatomy indicates unequivocal affinities to Rebbachisauridae, and two phylogenetic analyses recover the specimen as a member of the more exclusive clade Rebbachisaurinae. The new specimen differs from all other known rebbachisaurids in having an accessory bony lamina located inside the spinopostzygapophyseal fossa (spof). However, given that MDPA-Pv 007 is a single isolated element, we prefer to treat it as Rebbachisaurinae indet. Also, paleobiological aspects of caudal pneumaticity of a new rebbachisaurid specimen were studied. This specimen preserves unambiguous evidence of pneumaticity and is the best documentation to date of pneumaticity in rebbachisaurid caudal vertebrae. This research complements the few studies of caudal pneumaticity in neosauropods, and helps to elucidate its complex phylogenetic distribution. Finally, in light of new findings and taking advantage of new technologies, we encourage continuing investigating the phylogenetic distribution and paleobiological implications of pneumaticity in sauropod dinosaurs and other fossil archosaurs.
Acknowledgements
We thank to Dario López and Federico Poblete (both Universidad Nacional del Comahue, Neuquén, Argentina) for their assistance in the field works. We are also grateful to team of “Clínica de Imágenes S.A.” especially to D. Leguiza, G. Diez, M. Gac, M. Maisonave, and M. Miñana who made the CT scans possible. We also express gratitude to the Patrimonial Direction of Neuquén Province for granting permission to move the fossil. We are grateful for the constructive reviews and comments of John Whitlock (Mount Aloysius College, Cresson, USA) and Brooks Britt (Brigham Young University, Provo, USA), and the Acta Palaeontologica Polonica editorial board.
References
Apesteguía, S. 2007. The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina). Gondwana Research 12: 533–546. Crossref
Apesteguía, S., Gallina, P.A., and Haluza, A. 2010. Not just a pretty face: anatomical peculiarities in the postcranium of rebbachisaurids (Sauropoda: Diplodocoidea). Historical Biology 22: 165–174. Crossref
Averianov, A. and Sues, H.D. 2021. First rebbachisaurid sauropod dinosaur from Asia. PLOS ONE 16: e0246620. Crossref
Aureliano, T., Aline, M., Ghilardi, B.A., Navarro, M.A., Fernandes, F., Ricardi, B., and Wedel, M.J. 2021. Exquisite air sac histological traces in a hyperpneumatized nanoid sauropod dinosaur from South America. Scientific Reports 11: 24207. Crossref
Bellardini, F., Carballido, J.L., Filippi, L.S., Baiano, M.A., and Garrido, A.C. 2022. New rebbachisaurid remains from the Huincul Formation (Middle Cenomanian–Early Turonian) of the Central Neuquén Basin, Patagonia, Argentina. Publicación Electrónica de la Asociación Paleontológica Argentina 21: 1–24. Crossref
Bonaparte, J.F. 1996. Cretaceous tetrapods of Argentina. Münchner Geowissenschaftliche Abhandlungen 30: 73–130.
Bonaparte, J.F. 1997. Rayososaurus agrioensis Bonaparte, 1995. Ameghiniana 34: 1–116.
Britt, B.B. 1997. Postcranial pneumaticity. In: P.J. Currie and K. Padian (eds.), The Encyclopedia of Dinosaurs, 590–593. Academic Press, San Diego.
Calvo, J.O. and Bonaparte, J.F. 1991. Andesaurus delgadoi gen. et. sp. nov. (Saurischia-Sauropoda), dinosaurio Titanosauridae de la Formación Rio Limay (Albiano–Cenomaniano), Neuquén, Argentina. Ameghiniana 28: 303–310.
Calvo, J.O. and Salgado, L. 1995. Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian–Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia 11: 13–33.
Canudo, J.I., Carballido, J.L., Garrido, A., and Salgado, L. 2018. A new rebbachisaurid sauropod from the Aptian–Albian, Lower Cretaceous Rayoso Formation, Neuquén, Argentina. Acta Palaeontologica Polonica 63: 1–13. Crossref
Carballido, J.L., Garrido, A.C., Canudo, J.I., and Salgado, L. 2010. Redescription of Rayososaurus agrioensis Bonaparte (Sauropoda, Diplodocoidea), a rebbachisaurid from the early Late Cretaceous of Neuquén. Geobios 43: 493–502. Crossref
Carballido, J.L., Salgado, L., Pol, D., Canudo, J.I., and Garrido, A.C. 2012. A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Basin; evolution and biogeography of the group. Historical Biology 24: 631–654. Crossref
Carpenter, K. 2018. Maraapunisaurus fragillimus, n.g. (formerly Amphicoelias fragillimus), a basal rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado. Geology of the Intermountain West 5: 227–244. Crossref
Carvalho, I.S., dos Santos Avilla, L., and Salgado, L. 2003. Amazonsaurus maranhensis gen. and sp. nov. (Sauropoda, Diplodocoidea) from the Lower Cretaceous (Aptian–Albian) of Brazil. Cretaceous Research 24: 697–713. Crossref
Cerda, I.A., Salgado, L., and Powell, J.E. 2012. Extreme postcranial pneumaticity in sauropod dinosaurs from South America. Paläontologische Zeitschrift 86: 441–449. Crossref
Claessens, L.P., O’Connor, P.M., and Unwin, D.M. 2009. Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism. PLoS ONE 4: e4497. Crossref
Dalla Vecchia, F.M. 1998. Remains of Sauropoda (Reptilia, Saurischia) in the Lower Cretaceous (Upper Hauterivian/Lower Barremian) limestones of SW Istria (Croatia). Geologia Croatia 51: 105–134.
Fanti, F., Cau, A., Cantelli, L., Hassine, M., and Auditore, M. 2015. New information on Tataouinea hannibalis from the Early Cretaceous of Tunisia and implications for the tempo and mode of rebbachisaurid sauropod evolution. PLOS ONE 10: e0123475. Crossref
Fanti, F., Cau, A., Hassine, M., and Contessi, M. 2013. A new sauropod dinosaur from the Early Cretaceous of Tunisia with extreme avian-like pneumatization. Nature Communications 4: 1–7. Crossref
Gallina, P.A. and Apesteguía, S. 2005. Cathartesaura anaerobica gen. and sp. nov., a new rebbachisaurid (Dinosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Río Negro, Argentina. Revista del Museo Argentino de Ciencias Naturales 7: 153–166. Crossref
Garrido, A.C. 2010. Estratigrafía del Grupo Neuquén, Cretácico Superior de la Cuenca Neuquina (Argentina): nueva propuesta de ordenamiento litoestratigráfico. Revista del Museo Argentino de Ciencias Naturales Nueva Serie r 12: 121–177. Crossref
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Haluza, A., Canale, J.I., Otero, A., Pérez, L.M., and Scanferla, C.A. 2012. Changes in vertebral laminae across the cervicodorsal transition of a well-preserved rebbachisaurid (Dinosauria, Sauropoda) from the Cenomanian of Patagonia, Argentina. Journal of Vertebrate Paleontology 32: 219–224. Crossref
Ibiricu, L.M., Casal, G.A., Lamanna, M.C., Martínez, R.D., Harris, J.D., and Lacovara, K.J. 2012. The southernmost records of Rebbachisauridae (Sauropoda: Diplodocoidea), from early Late Cretaceous deposits in central Patagonia. Cretaceous Research 34: 220–232. Crossref
Ibiricu, L.M., Casal, G.A., Martínez, R.D., Lamanna, M.C., Luna, M., and Salgado, L. 2013. Katepensaurus goicoecheai, gen. et sp. nov., a Late Cretaceous rebbachisaurid (Sauropoda, Diplodocoidea) from central Patagonia, Argentina. Journal of Vertebrate Paleontology 33: 1351–1366. Crossref
Ibiricu, L.M., Casal, G.A., Martínez, R.D., Lamanna, M.C., Luna, M., and Salgado, L. 2015. New material of Katepensaurus goicoecheai (Sauropoda: Diplodocoidea) and its significance for the morphology and evolution of Rebbachisauridae. Ameghiniana 52: 430–446. Crossref
Janensch, W. 1929. Die Wirbelsaule der Gattung Dicraeosaurus. Palaeontographica Supplement 7: 39–133.
Lerzo, L.N., Carballido, J.L., and Gallina, P.A. 2021. Rebbachisaurid sauropods in Asia? A re-evaluation of the phylogenetic position of Dzharatitanis kingi from the late Cretaceous of Uzbekistan. Publicación Electrónica de la Asociación Paleontológica Argentina 21: 18–27.
Lindoso, R.M., Medeiros, M.A.A., Carvalho de Souza, I., Pereira, A.A., Mendes, I.D., Iori, F.V., and Silva, T.C.M. 2019. A new rebbachisaurid (Sauropoda: Diplodocoidea) from the middle Cretaceous of northern Brazil. Cretaceous Research 104: 104191. Crossref
Mannion, P.D. and Barrett, P.M. 2013. Additions to the sauropod dinosaur fauna of the Cenomanian (early Late Cretaceous) Kem Kem beds of Morocco: palaeobiogeographical implications of the mid-Cretaceous African sauropod fossil record. Cretaceous Research 45: 49–59. Crossref
Mannion, P.D., Upchurch, P., and Hutt, S. 2011. New rebbachisaurid (Dinosauria: Sauropoda) material from the Wessex Formation (Barremian, Early Cretaceous), Isle of Wight, United Kingdom. Cretaceous Research 32: 774–780. Crossref
O’Connor, P.M. 2006. Postcranial pneumaticity: an evaluation of soft-tissue influences on the postcranial skeleton and the reconstruction of pulmonary anatomy in archosaurs. Journal of Morphology 267: 1199–1226. Crossref
O’Connor, P.M. 2009. Evolution of archosaurian body plans: skeletal adaptations of an air-sac-based breathing apparatus in birds and other archosaurs. Journal of Experimental Zoology. Part A, Ecological Genetics and Physiology 311: 629–646. Crossref
Paulina Carabajal, A., Canale, J.I., and Haluza, A. 2016. New rebbachisaurid cranial remains (Sauropoda, Diplodocoidea) from the Cretaceous of Patagonia, Argentina, and the first endocranial description for a South American representative of the clade. Journal of Vertebrate Paleontology 36: e1167067. Crossref
Pereda Suberbiola, J., Torcida, F., Izquierdo, L.A., Huerta, P., Montero, D., and Pérez, G. 2003. First rebbachisaurid dinosaur (Sauropoda, Diplodocoidea) from the early Cretaceous of Spain: palaeobiogeographical implications. Bulletin de la Société géologique de France 174: 471–479. Crossref
Salgado, L., Canudo, J.I., Garrido, A.C., and Carballido, J.L. 2012. Evidence of gregariousness in rebbachisaurids (Dinosauria, Sauropoda, Diplodocoidea) from the Early Cretaceous of Neuquén (Rayoso Formation), Patagonia, Argentina. Journal of Vertebrate Paleontology 32: 603–613. Crossref
Salgado, L., Carvalho de Souza, I., and Garrido, A.C. 2006. Zapalasaurus bonapartei, un nuevo dinosaurio saurópodo de la Formación La Amarga (Cretácico Inferior), noroeste de Patagonia, Provincia de Neuquén, Argentina. Geobios 39: 695–707. Crossref
Salgado, L., Gallina, P.A., Lerzo, L.N., and Canudo, J.I. 2022. Highly specialized diplodocoids: the Rebbachisauridae. In: A. Otero, J.L. Carballido, and D. Pol (eds.), South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution, 165–208. Springer International Publishing, Cham. Crossref
Salgado, L., Garrido, A., Cocca, S.E., and Cocca, J.R. 2004. Lower Cretaceous rebbachisaurid sauropods from Cerro Aguada del León (Lohan Cura Formation), Neuquén Province, northwestern Patagonia, Argentina. Journal of Vertebrate Paleontology 24: 903–912. Crossref
Schwarz, D. and Fritsch, G. 2006. Pneumatic structures in the cervical vertebrae of the Late Jurassic Tendaguru sauropods Brachiosaurus brancai and Dicraeosaurus. Eclogae Geologicae Helvetiae 99: 65–78. Crossref
Sereno, P.C., Beck, A.L., Dutheil, D.B., Larsson, H.C.E., Lyon, G.H., Moussa, B., Sadleir, R.W., Sidor, C.A., Varricchio, D.J., Wilson, G.P., and Wilson, J.A. 1999. Cretaceous Sauropods from the Sahara and the Uneven Rate of Skeletal Evolution among Dinosaurs. Science 286: 1342–1347. Crossref
Sereno, P.C., Wilson, J.A., Witmer, L.M., Whitlock, J.A., Maga, A., Ide, O., and Rowe, T.A. 2007. Structural extremes in a Cretaceous dinosaur. PLoS ONE 2: e1230. Crossref
Taylor, M.P. and Naish, D. 2007. An unusual new neosauropod dinosaur from the Lower Cretaceous Hastings Beds Group of East Sussex, England. Palaeontology 50: 1547–1564. Crossref
Taylor, M.P. and Wedel, M.J. 2021. Why is vertebral pneumaticity in sauropod dinosaurs so variable? Qeios 1G6J3Q.5. Crossref
Torcida, F.B., Canudo, J.I., Huerta, P., Montero, D., Suberbiola, X.P., and Salgado, L. 2011. Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica 56: 535–552. Crossref
Wedel, M.J. 2003. The evolution of vertebral pneumaticity in sauropod dinosaurs. Journal of Vertebrate Paleontology 23: 344–357. Crossref
Wedel, M.J. 2005. Postcranial skeletal pneumaticity in sauropods and its implications for mass estimates. In: J.A. Wilson and K. Curry-Rogers (eds.), The Sauropods: Evolution and Paleobiology, 201–228. University of California Press, Berkeley. Crossref
Wedel, M.J. 2009. Evidence for bird-like air sacs in saurischian dinosaurs. Journal of Experimental Zoology 311:611–628. Crossref
Wedel, M.J. and Taylor, M.P. 2013. Caudal pneumaticity and pneumatic hiatuses in the sauropod dinosaurs Giraffatitan and Apatosaurus. PLOS ONE 8: e78213. Crossref
Wedel, M.J., Cifelli, R.L., and Sanders, R.K. 2000. Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma. Journal of Vertebrate Paleontology 20: 109–114. Crossref
Whitlock, J.A. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society 161: 872–915. Crossref
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639–653. Crossref
Wilson, J.A. 2012. New vertebral laminae and patterns of serial variation in vertebral laminae of sauropod dinosaurs. Contributions from the Museum of Paleontology, University of Michigan 32: 91–110.
Wilson, J.A. and Allain, R. 2015. Osteology of Rebbachisaurus garasbae Lavocat, 1954, a diplodocoid (Dinosauria, Sauropoda) from the early Late Cretaceous-aged Kem Kem beds of southeastern Morocco. Journal of Vertebrate Paleontology 35: e1000701. Crossref
Wilson, J.A., D’Emic, M.D., Ikejiri, T., Moacdieh, E.M., and Whitlock, J.A. 2011. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. PLoS ONE 6: e17114. Crossref
Windholz, G.J., Coria, R.A., and Zurriaguz, V.L. 2019. Vertebral pneumatic structures in the Early Cretaceous sauropod dinosaur Pilmatueia faundezi from northwestern Patagonia, Argentina. Lethaia 53: 369–381. Crossref
Windholz, G.J., Carballido, J.L., Coria, R.A., Zurriaguz, V.L., and Rauhut, O.W. 2022a. How pneumatic were the presacral vertebrae of dicraeosaurid (Sauropoda: Diplodocoidea) dinosaurs? Biological Journal of the Linnean Society 138: 103–120. Crossref
Windholz, G.J., Coria, R.A., Bellardini, F., Baiano, M.A., Pino, D., Ortega, F., and Currie P.J. 2022b. On a dicraeosaurid specimen from the Mulichinco Formation (Valanginian, Neuquén Basin) of Argentina and phylogenetic relationships of the South American dicraeosaurids (Sauropoda, Diplodocoidea). Comptes Rendus Palevol 21: 991–1019. Crossref
Witmer, L.M. 1997. The evolution of the antorbital cavity of archosaurs: a study in soft-tissue reconstruction in the fossil record with an analysis of the function of pneumaticity. Journal of Vertebrate Paleontology 17: 1–76. Crossref
Zurriaguz, V.L. and Cerda, I.A. 2017. Caudal pneumaticity in derived titanosaurs (Dinosauria: Sauropoda). Cretaceous Research 73: 14–24. Crossref
Character scores for new specimen MDPA-Pv 007 added to the matrix proposed by Bellardini et al. (2022).
??????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????000140001131000?1101000????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????1
Character scores for new specimen MDPA-Pv 007 added to the matrix proposed by Windholz et al. (2022b).
?????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????111?0001?0100000??001100?11010010001100??????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????????
Acta Palaeontol. Pol. 69 (1): 39–47, 2024
https://doi.org/10.4202/app.01104.202