
New finds of Olenekian, Early Triassic, trematosaurid amphibians and prolocophonid reptiles from Poland
TOMASZ SULEJ, MARCIN MACHALSKI, and MATEUSZ TAŁANDA
Sulej, T., Machalski, M., and Tałanda, M. 2024. New finds of Olenekian, Early Triassic, trematosaurid amphibians and prolocophonid reptiles from Poland. Acta Palaeontologica Polonica 69 (1): 49–56.
The continental Lower Triassic (Middle Buntsandstein) siliciclastic deposits exposed along the margins of the Holy Cross Mountains, Poland, yield locally abundant vertebrate footprints and bones. Although the footprints have been described in a number of studies, providing, for example, new insights into the origin of dinosaurs, there are few studies focusing on the bones. Here, we describe new amphibian and reptile material from the Buntsandstein fluvial sandstones of early Olenekian age exposed at Stryczowice on the north-eastern margin of the Holy Cross Mountains. These finds include fragmentary cranial specimens referred to as Trematosauridae gen. et sp. indet. and Procolophonidae gen. et sp. indet. Faunal differences between Stryczowice and the best-known Polish Olenekian vertebrate-bearing site of Czatkowice 1 near Kraków support heterogeneity in the Early Triassic vertebrate distribution across Pangea.
Key words: Temnospondyli, Parareptilia, tooth, parasphenoid, maxilla, Olenekian, Triassic, Poland.
Tomasz Sulej [sulej@twarda.pan.pl; ORCID: https://orcid.org/0000-0002-8839-2736 ] and Marcin Machalski [mach@twarda.pan.pl; ORCID: https://orcid.org/0000-0002-1121-4093 ], Institute of Paleobiology PAS, Twarda 51/55, 00-818 Warsaw, Poland.
Mateusz Tałanda [m.talanda@uw.edu.pl; ORCID: https://orcid.org/0000-0003-3358-9539 ], Institute of Evolutionary Biology, Faculty of Biology, Biological and Chemical Research Centre, University of Warsaw, Żwirki i Wigury 101, 02-089 Warsaw, Poland.
Received 9 October 2023, accepted 31 January 2024, published online 5 March 2024.
Copyright © 2024 T. Sulej et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Continental Lower Triassic (Middle Buntsandstein in lithostratigraphic scheme) siliciclastic deposits of Olenekian age are exposed along the margins of the Holy Cross Mountains, Poland (Senkowiczowa and Ślączka 1962; Senkowiczowa 1970; Mader and Barczuk 1985; Mader and Rdzanek 1985; Ptaszyński and Niedźwiedzki 2006; Brusatte et al. 2011). Locally, these strata yield abundant vertebrate footprints and bones.
The vertebrate footprints from the Middle Buntsandstein succession have been described in a number of studies (e.g., Fuglewicz et al. 1990; Ptaszyński 2000; Niedźwiedzki and Ptaszyński 2007; Klein and Niedźwiedzki 2012), some providing new insights into the origin of dinosaurs (Brusatte et al. 2011; Niedźwiedzki et al. 2013).
In contrast, there are few studies on vertebrate skeletal material from these deposits (Sulej and Niedźwiedzki 2009, 2013; Skrzycki et al. 2018), although the vertebrate remains, largely assigned to Temnospondyli, are common at some levels of the Buntsandstein succession (the „Labyrinthodontidae Beds” of Senkowiczowa and Ślączka 1962).
Here we describe new vertebrate material recovered from the fluvial Middle Buntsandstein sandstones (the “Labyrinthodontidae Beds”) exposed at Stryczowice, in the north-eastern margin of the Holy Cross Mts., Poland (Fig. 1). This is the same locality that yielded abundant arthropod trackways, possibly of notostracan origin (Machalski and Machalska 1994). The material studied is fragmentary and includes a labyrinthodont tooth, a skull fragment of a trematosaurid amphibian (identified as Trematosauridae gen. et sp. indet.) and a jaw fragment of a prolocophonid reptile (identified as Procolophonidae gen. et sp. indet.). These new records add to our knowledge of regional Early Triassic vertebrate faunas and communities, which are poorly understood in the Germanic Basin due to the sparse skeletal record (Sues et al. 2022).
A diverse fauna of small vertebrates has been found on the edge of this basin in a bone breccia of early late Olenekian age that infilled paleokarst structures developed in Carboniferous limestones at the locality of Czatkowice 1 near Kraków, southern Poland (Paszkowski 2009; Borsuk-Białynicka 2018; Borsuk-Białynicka et al. 2003; Cook and Trueman 2009; Evans and Borsuk-Białynicka 1998). The Czatkowice 1 bone breccia originated in an upland environment, which significantly contrasted with the lowland floodplain setting of the Stryczowice section described here. Therefore, the present study creates an opportunity to compare the distibution of amphibians and reptiles in not far distant but different Olenekian environments of the Germanic Basin.
Institutional abbreviations.—MSB (Museum Schloss Bernburg, Germany; PIN, Paleontological Institute, Russian Academy of Sciences, Moscow, Russia; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
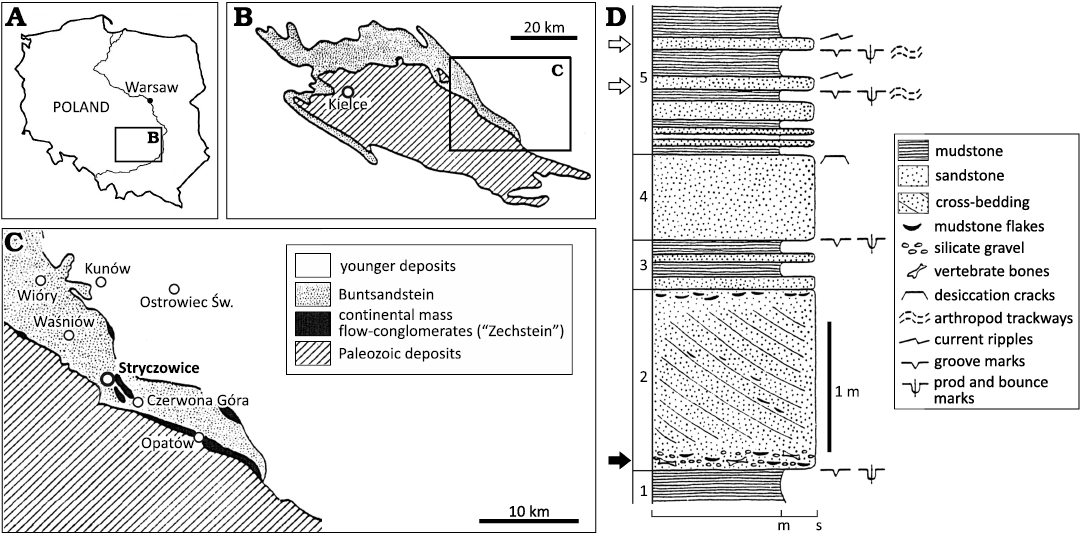
Fig. 1. The geological setting of the vertebrate-bearing locality at Stryczowice. A. Location of the Holy Cross Mountains in Poland. B. Simplified geological map of the Holy Cross Mountains with Paleozoic core and range of the Bundsandstein exposures. C. Location of the Stryczowice locality on the geological map of the north-eastern margin of the Holy Cross Mountains with Paleozoic deposits, continental mass flow-conglomerates traditionally referred to as the “Zechstein”, Buntsandstein, and younger deposits. D. Log of the Stryczowice quarry with location of the bone-bearing basal conglomerate marked by a black arrow, and location of the levels with arthropod trackways marked by white arrows; numbers 1–5 mark units discussed in text. Figure adopted from Machalski and Machalska 1994: fig. 1.
Geological setting
A small inactive quarry is located on a hill south of Stryczowice, a village south-west of the town of Ostrowiec Świętokrzyski at the north-eastern margin of the Holy Cross Mountains (Fig. 1; outcrop 32 in Mader and Barczuk 1985: fig. 5). The outcrop section, which is heavily overgrown and largely inaccessible today, is composed of intercalated sandstone and mudstone beds, with a distinct decrease in the thickness of the sandstones upward in the section (Fig. 1D).
Stratigraphically, the quarry section at Stryczowice was assigned by Machalski and Machalska (1994) to the “Labirynthodontidae Beds” of the Middle Buntsandstein. According to Brusatte et al. (2011), the conchostracans and palynomorphs indicate an early Olenekian age for the Stryczowice succession. In terms of sedimentology, the section at Stryczowice was studied in detail by Machalski and Machalska (1994), who interpreted it as a typical cyclothem of a weakly braided river system, which characterised the depositional environment of this part of the Buntsandstein succession (Mader and Rdzanek 1985; Fuglewicz et al. 1990). The massive cross-bedded sandstone bed with the basal conglomerate containing vertebrate remains, quartz gravel and mudstone intraclasts (Fig. 1D: unit 2) was interpreted by these authors as the main channel deposit. According to Machalski and Machalska (1994), the intercalating mudstones and sandstones in the upper part of the section (Fig. 1D: units 3–5) were deposited in a floodplain environment. The mudstones were deposited in stagnant water (i.e., ponds and/or lakes), whereas the intercalated sands accumulated as crevasse-splay deposits during periodic floods.
The osteological material described here was recovered by MM during the mid 1980s from the conglomerate at the bottom of the massive sandstone bed in the lower portion of the section (Fig. 1D: unit 2, the conglomerate is marked by a black arrow). In addition to the specimens described in the present study, the presence of distinctive cross sections of the “labyrinthodont” teeth was noted during exploitation of the conglomeratic level. All the fossils studied are represented by isolated fragmentary skeletal elements, pointing, along with their occurence in a basal conglomerate, to the allochthonous nature of the fossil assemblage. This admixture of fossils was probably formed as the result of lateral migration of a river channel leading to exhumation, reworking and redeposition of remains originally buried in the floodplain deposits. The allochthonous and mixed character of the Stryczowice taphocoenosis is also supported by the fact that trematosaurids were largely fully-aquatic predators (Schoch and Millner 2000), whereas procolophonids are regarded as exclusively terrestrial animals (de Braga 2003; Cisneros 2008; Botha-Brink and Smith 2012).
The bone material from the Stryczowice quarry has not been described so far. As far as trace fossils are concerned, only arthropod (presumably notostracan) walking trackways assigned to the ichnospecies “Diplichnites” triassicus have been reported to date from the Stryczowice quarry (Machalski and Machalska 1994). They were recorded on the soles of two crevasse-splay sandstone beds (Fig. 1D: white arrows). No vertebrate footprints were recorded from the Stryczowice quarry by Machalski and Machalska (1994). However, the vertebrate footprints reported from Stryczowice by Brusatte et al. (2011) probably come from a location close to the quarry succession (see Brusatte et al. 2011: electronic supplementary material).
The material described here is housed at the Institute of Paleobiology, Polish Academy of Sciences (abbreviated ZPAL V. 78).
Systematic palaeontology
Class Amphibia Gray, 1825
Order Temnospondyli Zittel, 1888
Family Trematosauridae Watson, 1919
Trematosauridae gen. et sp. indet.
Figs. 2, 3B.
Material.—ZPAL V. 78/3, parasphenoid from lower Olenekian (Lower Triassic) of Stryczowice, north-eastern margin of the Holy Cross Mountains, Poland.
Description.—An almost complete parasphenoid ZPAL V. 78/3 is preserved in a conglomerate slab in two pieces (part and counterpart) with the bone visible in cross-section (Fig. 2). The anterior part of cultriform process and the left part of the parasphenoid plate are broken (Fig. 3B1).
A large area for the exoccipital is visible on the posterolateral corner of the dorsal side. It has a similar shape, size and texture to those of Inflectosaurus amplus Shishkin, 1960 (PIN 953/100; Shishkin 1968; Novikov 2007). The posterior part is divided on three belts by distinct ridges. The opening for the intracranial branch of the internal carotid is located more anteriorly than in Trematosaurus brauni Burmeister, 1849 (Schoch 2019) and Inflectosaurus amplus (Shishkin 1968; PIN 953/100). The posterior aspect of the pterygoid area is not visible in dorsal view, unlike in Inflectosaurus amplus (Shishkin 1968; PIN 953/100). The entrance foramen for the palatine nerve, which is also the exit for the posterior branch of the palatine artery, is well visible only in Inflectosaurus amplus (PIN 953/100). This foramen is much larger in Trematosaurus galae Novikov, 2010 (Novikov 2010: fig. 1d) and has a large ornamented area, which is absent in the new Polish specimen.
The long, irregular area for suture with the pterygoid is well visible on the ventral side of the parasphenoid plate. Its ventral edge is bent inwards (Fig. 3) as in Trematosaurus brauni (Schoch 2019). The posterior edge of the parasphenoid plate is slightly bent inwards, similar to Inflectosaurus amplus (Novikov 2007). A small opening is visible in the central area of the parasphenoid plate, which was not reported in any trematosaurid so far.
The cultriform process is T-shaped in cross-section with shallow grooves on both sides. Its ventral and lateral edges are sharp. This morphology is very similar to that observed in Trematosaurus brauni (Schoch 2019) and unlike Angusaurus Getmanov, 1989, which has a very narrow cultriform process (Fernández-Coll et al. 2019). The posterior aspect of the lateral edge of the cultriform process is slightly convexly curved compared to the same feature in Trematosaurus brauni and Inflectosaurus amplus, which is strongly curved.
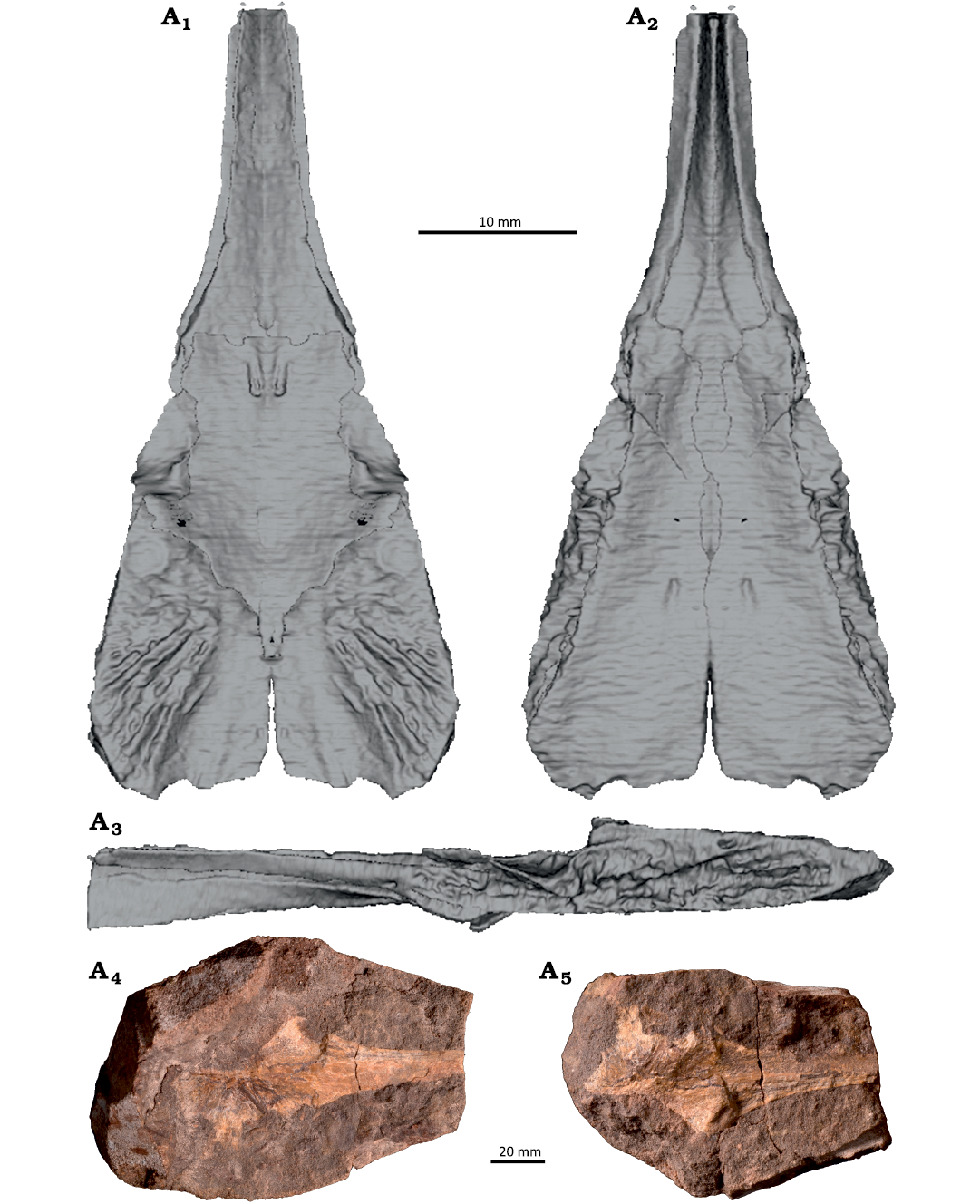
Fig. 2. Parasphenoid of Trematosauridae gen. et sp. indet. from Lower Triassic of Stryczowice, Holy Cross Mountains, Poland. ZPAL V. 78/3, reconstruction in dorsal (A1), ventral (A2), and lateral (A3) views, original specimen, part and counterpart (A4, A5). The restoration was based on the CT scan by supplementing the missing fragment with a mirror image of the preserved part (Fig. 3B).
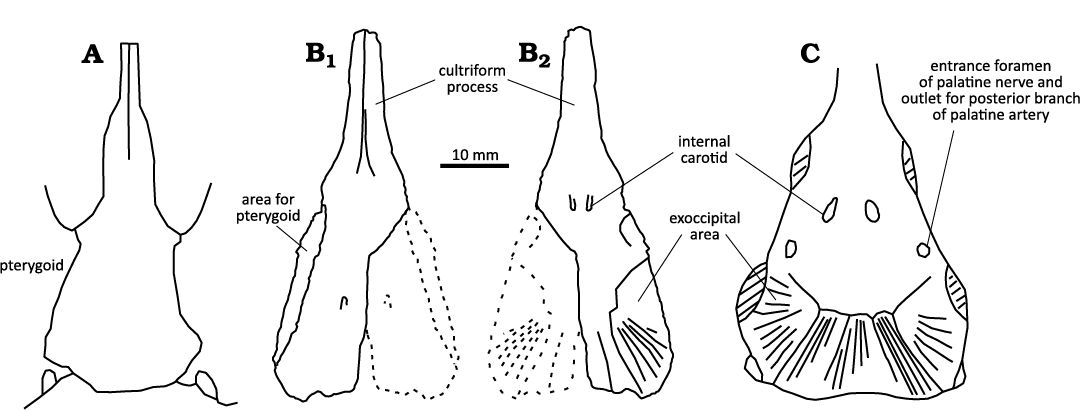
Fig. 3. Comparison of the parasphenoid of Trematosauridae gen. et sp. indet. with specimens from Germany and Russia. A. Trematosaurus brauni Burmeister, 1849, Merkel’s Quarry, Germany, ?late Olenekian, MSB G 366 in ventral view (modified from Schoch 2019: fig. 7B). B. Trematosauridae gen. et sp. indet., Stryczowice, Holy Cross Mountains, Poland, early Olenekian{?}, ZPAL V. 78/3 in ventral (B1) and dorsal (B1) views. C. Inflectosaurus amplus Shishkin, 1960, Bolshoe Bogdo locality, late Olenekian, PIN 953/100 in dorsal view (modified from Shishkin 1968: fig. 2c).
Remarks.—The Germanic Basin trematosaurids were originally described based on material from two localities: Merkel’s Quarry and Kappel in the Black Forest, Germany (Schoch and Werneburg 1998; Schoch and Milner 2000). Up until 2009, the geographically closest sites to the Holy Cross Mountains that yielded termatosaurid material were in Eastern Europe (Shishkin 1968; Novikov 2007, 2010). Sulej and Niedźwiedzki (2009) then reported the first trematosaurid from Poland from the Wióry locality, dated as Olenekian (Early Triassic). Later, Sulej and Niedźwiedzki (2013) described the capitosauroid Parotosuchus ptaszynski Sulej & Niedźwiedzki, 2013, from the same locality, based on fragments of a cranium and mandible. Globally, trematosaurids are also known from Middle and Late Triassic-aged deposits from Madagascar (Lehman 1966, 1979) and South Africa (Watson 1919; Welles 1993; Shishkin and Welman 1994).
Temnospondyli indet.
Fig. 4A.
Material.—ZPAL V. 78/2, tooth with broken tip from lower Olenekian (Lower Triassic) of Stryczowice, north-eastern margin of the Holy Cross Mountains, Poland.
Description.—Only the base of the crown of a tooth (ZPAL V. 78/2) was preserved in the conglomerate (Fig. 4A). In their comprehensive comparison of amphibian and reptile teeth from the Middle Triassic, Schoch et al. (2018) showed that temnospondyl teeth were the same for the entire Lower Triassic material. Based on the fact that deep grooves are limited to the basal aspect of the tooth crown and are aligned parallel to the apicobasal axis of the tooth crown, the features present in ZPAL V. 78/2 are most consistent with this tooth being assigned to a temnospondyl. The lack of the tip of the tooth complicates any taxonomical ascription below the subordinal level.
Class Reptilia Laurenti, 1768
Order Procolophonomorpha Romer, 1964
Family Procolophonidae Lydekker in Nicholson and Lydekker, 1889
Procolophonidae gen. et sp. indet.
Fig. 4B.
Material.—ZPAL V. 78/1, maxilla with broken posterior part from lower Olenekian (Lower Triassic) of Stryczowice, north-eastern margin of the Holy Cross Mountains, Poland.
Description.—Maxilla (ZPAL V. 78/1) is broken into two parts embedded in rock matrix, one preserving most of the bone and three teeth, whereas the other has three teeth and the posterior piece of the same bone (Fig. 4B). The lateral surface of the bone is poorly preserved and provides little anatomical information. The lateral surface is flat with no visible postnarial maxillary depression. The maxillary foramen opens anteroventrally near the ventral margin of the bone and near the facet for the premaxilla. The facet is well preserved and oriented anteromedially, although it is still partially covered by sediment. This anterior margin of the bone is blunt and dorsoventrally tall. There is no emargination for the naris. The bone is anteroposteriorly straight and labiolingually broad in ventral view. The posterior aspect of the maxilla is broken and incomplete.
The teeth are visible only in lateral view and in cross-section, which limits their description to some extent. The preserved portion of the maxilla bears ten tooth positions, but only six teeth remain in place (4th to 8th, and 10th maxillary teeth). All teeth are subtriangular in lateral view with broad bases (Fig. 4). Their tips are rather blunt, but it is not visible due to rock matrix if they have one or two cusps. The teeth gradually increase in size posteriorly up to seventh tooth, but the differences are modest. The enamel has a rather featureless surface, with no visible grooves or other structures. The first tooth was virtually circular in cross-section (orthogonal diameters of 0.88 mm vs 0.89 mm). The following teeth gradually increase in labiolingual breath, which becomes much larger than their mesiodistal length. The sixth tooth is the posteriormost one for which the measurement is possible. It is 1.63 mm wide labiolingually and 1.16 mm long mesiodistally. After the seventh tooth they seem to slightly decrease in size but the rock matrix precludes exact measurements.

Fig. 4. Temnospondyli indet. (A) and Procolophonidae gen. et sp. indet (B) from Lower Triassic of Stryczowice, Holy Cross Mountains, Poland. A. ZPAL V. 78/2, tooth. B. ZPAL V. 78/1, maxilla in lateral (B1) and ventral (B2) views; teeth in lateral view (B3, B5); cross-section of maxillary teeth in dorsal view (B4).
Remarks.—Up to four different procolophonids were described from roughly coeval (Olenekian) paleokarst bone breccia recovered from the Czatkowice 1 site (Borsuk-Białynicka and Lubka 2009). Procolophonids found there have much lower maxillary tooth counts (5–6 and 7), and the teeth are more heterodont in size compared to those from Stryczowice. Procolina teresae Borsuk-Białynicka & Lubka, 2009, has a much more pronounced anterior slope of the maxilla and its teeth are less triangular. The latter is also true for Procolophonidae gen. indet. II and III from Czatkowice 1. Procolophonidae gen. indet. I from Czatkowice 1 has subcircular teeth in cross-section throughout the entire maxilla, whereas the Stryczowice specimen has transversely expanded the posterior teeth. Therefore we can exclude assignment of the new material to any of the taxa from Czatkowice 1.
Numerous other procolophonids were described from the Buntsandstein of Germany (Huene 1912; Säilä 2008; Sues and Reisz 2008). The Stryczowice material cannot be assigned to Sclerosaurus armatus Meyer in Fischer, 1857, because the latter has a lower tooth count (7) and its teeth have long-axes oriented mesiolingually (Sues and Reisz 2008) rather than lingually. The comparison to Anomoidon liliensterni Huene, 1939, is fairly limited due to its poorly preserved maxillae (Säilä 2008), but the teeth seem to be much more heterodont in terms of size compared to the Stryczowice material. Koiloskiosaurus coburgensis Huene, 1912, has a similar tooth count (9 or 10) in the maxilla and its teeth are expanded transversely (Huene 1912) as in the Stryczowice material.
Procolophonids are also very diverse in the Lower Triassic of Russia (Ivakhnenko 1979; Ivakhnenko and Kurochkin 2008; Spencer and Benton 2000). The specimen from Stryczowice differs from specimens assigned to species of Tichvinskia, Orenburgia, and Kapes in having a higher tooth count (Ivakhnenko 1973, 1975, 1983; Spencer and Benton 2000). Phaanthosaurus (= Contritosaurus) has 10 rather uniform teeth in its maxilla but also possesses a large fossa surrounding the external naris (Ivakhnenko 1974, 1979; Spencer and Benton 2000). The latter feature seems to be absent in the Stryczowice specimen.
To sum up, the Stryczowice material can be distinguished from all species mentioned above except maybe Koiloskiosaurus coburgensis Huene, 1912, described from the Buntsandstein of Germany. However, more detailed comparisons between the two are difficult as the material of Koiloskiosaurus coburgensis requires redescription (Sues and Reisz 2008) and we did not study these specimens personally. Therefore, their possible conspecificity requires further study.
Concluding remarks
The vertebrate fauna described from Czatkowice 1 is of similar age (Olenekian, Early Triassic) to the material recovered from the Stryczowice section. The Czatkowice 1 locality is located just about 100 km southwest of Stryczowice. The absence of trematosaurids in Czatkowice 1 was already discussed by Shishkin and Sulej (2009). They stated that at Czatkowice 1 the accumulation of bones in the palaeocaves developed in the upland area and was more conducive to gathering terrestrial vertebrates than water-dependent amphibians (see Cook and Trueman 2009 for a detailed taphonomical study of the Czatkowice 1 assemblages). However, we observe that procolophonid faunas also differ between Czatkowice 1 and other Germanic Basin sites, including the Stryczowice site. These animals were terrestrial, so the taphonomic reasoning presented above cannot be applied to them. Although the new material is fragmentary, our preliminary observations suggest that the distribution of these two groups was strongly dependent on local palaeoecological, rather than only taphonomic, conditions. Indeed, the presence of a trematosaurid and a prolocophonid similar to species of Koiloskiosaurus link Stryczowice to the far more distant localities in Germany than to the Czatkowice 1 site. This is consistent with studies from better sampled areas that found that the distribution of Early Triassic vertebrates was heterogeneous across Pangea (Shishkin et al. 2006; Sidor et al. 2013).
This study adds a new record of procolophonids, a group of highly successful Permo-Triassic reptiles (Cisneros 2008). Up to four distinct taxa are known from the Czatkowice 1 locality and they differ from the three taxa from the Buntsandstein and from more eastern basins (Borsuk-Białynicka and Lubka 2009; Huene 1912; Säilä 2008; Sues and Reisz 2008; Ivakhnenko 1979; Ivakhnenko and Kurochkin 2008; Spencer and Benton 2000). It shows their relatively high diversity in that part of Pangea just a few million years after the end of the Permian.
Acknowledgements
We thank Krzysztof Karczewski and Szymon Łukasiewicz (both Wojskowa Akademia Techniczna, Warsaw, Poland) for CT scanning the specimen ZPAL V. 78/3 and Marian Dziewiński (ZPAL) for photographs of the specimens. We also thank Sergi López Torres (Institute of Evolutionary Biology, University of Warsaw, Poland) for his comments and linguistic help, and two reviewers Michael Benton (Bristol University, UK) and Hans-Dieter Sues (Smithsonian National Museum of Natural History, Washington, USA) for their constructive criticism.
References
Borsuk-Białynicka, M. 2018. Diversity of diapsid fifth metatarsals from the Lower Triassic karst deposits of Czatkowice, southern Poland—functional and phylogenetic implications. Acta Palaeontologica Polonica 63: 417–434. Crossref
Borsuk-Białynicka, M. and Lubka, M. 2009. Procolophonids from the Early Triassic of Poland. Palaeontologia Polonica 65: 107–144.
Borsuk-Białynicka, M., Maryańska, T., and Shishkin, M.A. 2003. New data on the age of the bone breccia from the locality Czatkowice 1 (Cracow Upland, Poland). Acta Palaeontologica Polonica 48: 153–155.
Botha-Brink, J. and Smith, R.M.H. 2012. Palaeobiology of Triassic procolophonids, inferred from bone microstructure. Comptes Rendus Palevol 11: 419–433. Crossref
Brusatte, S., Niedźwiedzki, G., and Butler, R. 2011. Footprints pull origin and diversification of dinosaur stem lineage deep into Early Triassic. Proceedings of the Royal Society B, Biological Sciences 278: 1107–1113. Crossref
Burmeister, H. 1849. Die Labyrinthodonten aus dem bunten Sandstein bei Bernburg. I. Abtheilung Trematosaurus. 69 pp. Reimer, Berlin.
Cisneros, J.C. 2008. Phylogenetic relationships of procolophonid parareptiles with remarks on their geological record. Journal of Systematic Palaeontology 6: 345–366. Crossref
Cook, E. and Trueman, C.N. 2009. Taphonomy and geochemistry of a vertebrate microremains assemblage from the Early Triassic fissure deposits at Czatkowice 1, southern Poland. Palaeontologia Polonica 65: 17–30.
de Braga, M. 2003. The postcranial skeleton, phylogenetic position, and probable lifestyle of the Early Triassic reptile Procolophon trigoniceps. Canadian Journal of Earth Sciences 40: 527–556. Crossref
Evans, S.E. and Borsuk-Białynicka, M. 1998. A stem-group frog from the Early Triassic of Poland. Acta Palaeontologica Polonica 43: 573–580.
Fernández-Coll, M., Arbez, T., Bernardini, F., and Fortuny, J. 2019. Cranial anatomy of the Early Triassic trematosaurine Angusaurus (Temnospondyli: Stereospondyli): 3D endocranial insights and phylogenetic implications. Journal of Iberian Geology 45: 269–286. Crossref
Fischer , L.H. 1857. Über Sclerosaurus armatus Meyer, eine neue Saurier-Gattung aus dem Bunten Sandstein bei Warmbach gegenüber Rheinfelden. Neues Jahrbuch für Mineralogie, Geologie. Geognosie und Petrefakten-Kunde 1857: 136–140.
Fuglewicz, R., Ptaszyński, T., and Rdzanek, K. 1990. Lower Triassic footprints from the Świętokrzyskie (Holy Cross) Mountains, Poland. Acta Palaeontologica Polonica 35: 109–164.
Getmanov, S.N. 1989. Triassic amphibians of the East European platform (family Benthosuchidae Efremov). Trudy Paleontologičeskogo Instituta AN SSSR 236: 1–102.
Gray, J.E. 1825. A synopsis of the genera of reptiles and Amphibia, with a description of some new species. Annals of Philosophy 10: 193–217.
Huene, F. von 1912. Die Cotylosaurier der Trias. Palaeontographica 59: 69–102.
Huene, F. von. 1939. Ein neuer Procolophonide aus dem deutschen Buntsandstein. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilage-Band, Abteilung B 81: 501–511.
Ivakhnenko, M.F. 1973. Skull structure in the Early Triassic procolophonian Tichvinskia vjatkensis [in Russian]. Paleontologičeskij žurnal 1973 (4): 74–83.
Ivakhnenko, M.F. 1974. New data on the procolophonids from the Early Triassic of USRR [in Russian]. Paleontologičeskij žurnal 1974 (3): 68–74.
Ivakhnenko, M.F. 1975. On the generic contents of the Procolophonidae from the Early Triassic of the Cis−Uralian region [in Russian]. Paleontologičeskij žurnal 1975 (1): 88–93.
Ivakhnenko, M.F. 1979. Permian and Triassic procolophonoids of the Russian platform [in Russian]. Trudy Paleontologičeskogo Instituta AN SSSR 164: 1–80.
Ivakhnenko, M.F. 1983. New procolophonids of the Eastern Europe [in Russian]. Paleontologičeskij žurnal 1983 (2): 130–133.
Ivakhnenko, M.F. and Kurochkin, Y.N. (eds.) 2008. Fossil Vertebrates of Russia and Adjacent Countries. Fossil Reptiles and Birds. Part 1. 348 pp. GEOS, Moscow.
Klein, H. and Niedźwiedzki, G. 2012 Revision of the Lower Triassic tetrapod ichnofauna from Wióry, Holy Cross Mountains, Poland. New Mexico Museum of Natural History and Science, Bulletin 56: 1–62.
Laurenti, J.N. 1768. Specimen medicum, exhibens synopsin reptilium emendatam cum experimentis circa venena et antidota reptilium austracorum, quod authoritate et consensu. 217 pp. Joan Thomae, Vienna. Crossref
Lehman, J.-P. 1966. Nouveaux stégocéphales de Madagascar. Annales de Paléontologie 52: 115–139.
Lehman, J.-P. 1979. Nouveaux Trematosaures de Madagascar, les Stegocephales Malgaches et leur Paleoecologie. Annales de Paléontologie 65: 35–53.
Machalski, M. and Machalska, K. 1994. Arthropod trackways “Diplichnites” triassicus (Linck, 1943) from the Lower Triassic (Buntsandstein) fluvial deposits of the Holy Cross Mts. Acta Geologica Polonica 44: 267–275.
Mader, D. and Barczuk, A. 1985. Gravelly to sandy braidplain deposition in the Czerwona Góra Beds and Stryczowice Beds (Middle Buntsandstein) of the northeastern Holy Cross Mountains (Poland). In: D. Mader (ed.), Aspects of Fluvial Sedimentation in the Lower Triassic Buntsandstein of Europe. Lecture Notes in Earth Sciences 4: 351–396. Crossref
Mader, D. and Rdzanek, K. 1985. Sandy braidplain deposition with minor pedogenesis in the Labyrinthodontidae Beds (Middle Buntsandstein) of the Northeastern Holy Cross Mountains (Poland). In: D. Mader (ed.), Aspects of Fluvial Sedimentation in the Lower Triassic Buntsandstein of Europe. Lecture Notes in Earth Sciences 4: 281–317. Crossref
Nicholson, H.A. and Lydekker, R. 1889. A Manual of Palaeontology for the Use of Students. Third Edition. Volume II. 735 pp. William Blackwood and Sons, Edinburgh.
Niedźwiedzki, G. and Ptaszyński, T. 2007. Large Chirotheriidae tracks in the Early Triassic of Wióry, Holy Cross Mountains, Poland. Acta Geologica Polonica 57: 325–342.
Niedźwiedzki, G., Brusatte, S.L., and Butler, R.J. 2013. Prorotodactylus and Rotodactylus tracks: an ichnological record of dinosauromorphs from the Early–Middle Triassic of Poland. Geological Society, London, Special Publications 379 (1): 319–351. Crossref
Novikov, I.V. 2007. New data on trematosauroid labyrinthodonts of eastern Europe: 1. Genus Inflectosaurus Shishkin, 1960. Paleontological Journal 41: 167–174. Crossref
Novikov, I.V. 2010. New data on trematosauroid labyrinthodonts of Eastern Europe: 2. Trematosaurus galae sp. nov.: cranial morphology. Paleontological Journal 44: 457–467. Crossref
Paszkowski, M. 2009. The Early Triassic karst of Czatkowice 1, southern Poland. Palaeontologia Polonica 65: 7–16.
Ptaszyński, T. 2000. Lower Triassic vertebrate footprints from Wióry, Holy Cross Mountains, Poland. Acta Palaeontologica Polonica 45: 151–194.
Ptaszyński, T. and Niedźwiedzki, G. 2006. Pstry piaskowiec w Górach Świętokrzyskich: chronostratygrafia i korelacja litostratygraficzna z basenem turyńskim. Przegląd Geologiczny 54: 525–533.
Romer, A.S. 1964. Diadectes an amphibian? Copeia 4: 718–719. Crossref
Säilä, L. 2008. The osteology and affinities of Anomoiodon liliensterni, a procolophonid reptile from the Lower Triassic Bundsandstein [sic] of Germany. Journal of Vertebrate Paleontology 28: 1199–1205. Crossref
Schoch, R.R. 2019. Osteology of the temnospondyl Trematosaurus brauni Burmeister, 1849 from the Middle Buntsandstein of Bernburg, Germany. Palaeodiversity 12: 41–63. Crossref
Schoch, R.R. and Milner, A.R. 2000. Stereospondyli: Stem-Stereospondyli, Rhinesuchidae, Rhytidostea, Trematosauroidea, Capitosauroidea. 164 pp. F. Pfeil, Munich.
Schoch, R.R. and Werneburg, R. 1998. The Triassic labyrinthodonts from Germany. Zentralblatt für Geologie und Paläontologie Teil I 1998: 629–650.
Schoch, R.R., Ullmann, F., Rozynek, B., Ziegler, R., Seegis, D., and Sues, H.D. 2018. Tetrapod diversity and palaeoecology in the German Middle Triassic (Lower Keuper) documented by tooth morphotypes. Palaeobiodiversity and Palaeoenvironments 98: 615–638. Crossref
Senkowiczowa, H. 1970. Mesozoic stratigraphy of the Holy Cross Mts. area. Prace Instytutu Geologicznego 56: 7–41.
Senkowiczowa, H. and Ślączka, A. 1962. The Bunter on the northern border of the Holy Cross Mts. Rocznik Polskiego Towarzystwa Geologicznego 22: 313–338.
Shishkin, M.A. 1960 New Triassic trematosaurid Inflectosaurus amplus. Paleontologičeskij žurnal 1960 (2): 130–148.
Shishkin, M.A. 1968. On the cranial arterial system of the labyrinthodonts. Acta Zoologica 49: 1–22. Crossref
Shishkin, M.A. and Sulej, T. 2009. The Early Triassic temnospondyls of the Czatkowice 1 tetrapod assemblage. Palaeontologia Polonica 65: 31–77.
Shishkin, M.A. and Welman, J. 1994. A new find of Trematosuchus (Amphibia, Temnospondyli) from the Cynognathus zone of South Africa. Palaeontologia Africana 31: 39–49.
Shishkin, M.A., Sennikov, A.G., Novikov, I.V., and Ilyina, N.V. 2006. Differentiation of tetrapod communities and some aspects of biotic events in the Early Triassic of Eastern Europe. Paleontological Journal 40: 1–10. Crossref
Sidor, C.A., Vilhena, D.A., Angielczyk, K.D., Huttenlocker, A.K., Nesbitt, S.J., Peecook, B.R., Steyer, J.S., Smith, R.M.H., and Tsuji, L.A. 2013. Provincialization of terrestrial faunas following the end-Permian mass extinction. Proceedings of the National Academy of Sciences 110: 8129–8133. Crossref
Skrzycki, P., Niedźwiedzki, G., and Tałanda, M. 2018. Dipnoan remains from the Lower–Middle Triassic of the Holy Cross Mountains and northeastern Poland, with remarks on dipnoan palaeobiogeography. Palaeogeography, Palaeoclimatology, Palaeoecology 496: 332–345. Crossref
Spencer, P.S. and Benton, M.J. 2000. Procolophonoids from the PermoTriassic of Russia. In: M.J. Benton, M.A. Shishkin, D. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 160–176. Cambridge Univerisity Press, Cambridge.
Sues, H.-D. and Reisz, R. R. 2008. Anatomy and phylogenetic relationships of Sclerosaurus armatus (Amniota: Parareptilia) from the Buntsandstein (Triassic) of Europe. Journal of Vertebrate Paleontology 28: 1031–1042. Crossref
Sues, H.-D., Ezcurra, M.D., and Schoch, R.R. 2022. Eifelosaurus triadicus Jaekel, 1904, a “forgotten” reptile from the Upper Buntsandstein (Triassic: Anisian) of the Eifel region, Germany. Paläontologische Zeitschrift 96: 275–287. Crossref
Sulej, T. and Niedźwiedzki, G. 2009. Kapitozaury i trematozaury (płazy tarczogłowe) z wczesnego triasu Wiór w Górach Świętokrzyskich. In: Konferencja Paleontologiczna: Kręgowce kopalne, morfologia, systematyka, ewolucja, 3–5 XII 2009 Wrocław, 58–59. Instytut Biologii Uniwersytetu Przyrodniczego, Wrocław.
Sulej, T. and Niedźwiedzki, G. 2013. A new large capitosaurid temnospondyl amphibian from the Early Triassic of Poland. Acta Palaeontologica Polonica 58: 65–75.
Watson, D.M.S. 1919. The structure, evolution and origin of the Amphibia. The “Orders” Rachitomi and Stereospondyli. Philosophical Transactions of the Royal Society of London B 209: 1–73. Crossref
Welles, S.P. 1993. A review of the lonchorhynchine trematosaurs (Labyrinthodontia), and a description of a new genus and species from the Lower Moenkopi Formation of Arizona. PaleoBios 14: 1–24.
Acta Palaeontol. Pol. 69 (1): 49–56, 2024
https://doi.org/10.4202/app.01109.2023