

Taxonomic and stratigraphic update of the material historically attributed to Megalosaurus from Portugal
ELISABETE MALAFAIA, PEDRO MOCHO, FERNANDO ESCASO, IVAN NARVAÉZ, and FRANCISCO ORTEGA
Malafaia, E., Mocho, P., Escaso, F., Narvaéz, I., and Ortega, F. 2024. Taxonomic and stratigraphic update of the material historically attributed to Megalosaurus from Portugal. Acta Palaeontologica Polonica 69 (2): 127–171.
The first paleontological works on Mesozoic vertebrates from Portugal, carried out from the end of the 19th and the first half of the 20th century, provided the discovery of significant collections of vertebrate fossils. These collections are particularly relevant because they include several specimens collected from different regions of the Lusitanian Basin (some of the sites are currently inaccessible), whose fossil record is poorly known. Theropod remains are relatively scarce and generally consist of fragmentary material, mostly assigned to the megalosaurid Megalosaurus from the Middle Jurassic of England, the first dinosaur to be named and a “wastebasket” taxon used by many scientists to identify theropod material. The studied fossils mostly consist of isolated teeth and vertebrae collected from Upper Jurassic levels of the coastal region, with also some material from Lower and Upper Cretaceous strata from the central and northern sectors of the Lusitanian Basin. Here specimens attributed to Megalosaurus from different Portuguese institutions are reviewed and their taxonomic affinity and stratigraphic context are updated. Most specimens actually belong to different theropod groups, including several isolated teeth from different Upper Jurassic localities here assigned to Ceratosaurus, Torvosaurus, and Allosaurus, as well as an isolated tooth from the Lower Cretaceous that is attributed to an indeterminate allosauroid. Other theropod remains consist mostly of vertebral fragments of indeterminate avetheropods and allosauroids. Elements of other dinosaur groups are also represented, including a few vertebrae here referred to stegosaurians and iguanodontians, as well as a vertebra and some appendicular remains attributed to sauropods. Two vertebrae assigned to thalattosuchians were also identified. The study of this collection allows to better characterize the diversity of Late Jurassic dinosaur faunas from different areas of the Lusitanian Basin and provides some data on the poorly known Cretaceous fossil record of theropods from Portugal.
Key words: Crocodylomorpha, Dinosauria, Thalattosuchia, Stegosauria, Iguanodontia, Sauropoda, Eusauropoda, Titanosauriformes,Theropoda, Ceratosauria, Megalosauridae, Avetheropoda, Allosauroidea, Lusitanian Basin, Upper Jurassic, Cretaceous.
Elisabete Malafaia [ efmalafaia@ciencias.ulisboa.pt; ORCID: https://orcid.org/0000-0003-4894-4257 ], Instituto Dom Luiz, Faculdade de Ciências, Universidade de Lisboa, Campo Grande, 1749-016 Lisboa, Portugal; Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, Avda. Esparta s/n., E-28232 Las Rozas de Madrid, Spain.
Pedro Mocho [pdmocho@ciencias.ulisboa.pt; ORCID: https://orcid.org/0000-0002-3348-5572 ], Instituto Dom Luiz, Faculdade de Ciências, Universidade de Lisboa, Campo Grande, 1749-016 Lisboa, Portugal; Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, Avda. Esparta s/n., E-28232 Las Rozas de Madrid, Spain; Dinosaur Institute, Natural History Museum of Los Angeles County, 900 Exposition Blvd, 90007 CA, Los Angeles, USA.
Fernando Escaso [fescaso@ccia.uned.es; ORCID: https://orcid.org/0000-0001-7642-1555 ], Ivan Narvaéz [inarvaez @ccia.uned.es; ORCID: https://orcid.org/0000-0003-0114-7058 ], and Francisco Ortega [fortega@ccia.uned.es; ORCID: https://orcid.org/0000-0002-7431-354X ], Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, Avda. Esparta s/n., E-28232 Las Rozas de Madrid, Spain.
Received 23 October 2023, accepted 4 March 2024, published online 11 April 2024.
Copyright © 2024 E. Malafaia et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The first paleontological works on Mesozoic vertebrates from Portugal were associated with the activities of the Geological Commission, whose main objective was the geological mapping of the country. The first record of osteological dinosaurian remains from the Upper Jurassic of Portugal is attributed to the geologist Carlos Ribeiro and consists of some isolated theropod teeth found in 1863 during prospecting works in the Upper Jurassic strata on the Lourinhã Coast (Lapparent and Zbyszewski 1957; Ortega et al. 2006). One of these teeth is currently deposited in the paleontological collections of the Museu Nacional de História Natural e da Ciência of the University of Lisbon and is labeled as coming from “Coupe du Vale do Portinheiro à Carrasqueira”, which is an area near the locality of Porto Dinheiro, in the municipality of Lourinhã. This tooth was described by Lapparent and Zbyszewski (1957) as part of the material used for the description of a putative new species, Megalosaurus “pombali” and later assigned to Theropoda indet. (Carrano et al. 2012). The first study about the fossil record of vertebrates from the Mesozoic of Portugal was published by Henri-Émile Sauvage (1896, 1897–1898). In these publications, several osteological remains were identified as belonging to dinosaurs (including theropods, sauropods, and iguanodontids) and other vertebrates collected in different Upper Jurassic and Cretaceous localities of the Lusitanian Basin. Most of these specimens are currently housed in the collections of the Museu Nacional de História Natural e da Ciência and the Museu Geológico, although a few have been lost. From 1942, Georges Zbyszewski conducted a series of systematic paleontological prospecting in different Upper Jurassic sequences of the Lusitanian Basin, mainly in the coastal region, between Foz do Arelho (Caldas da Rainha) and Santa Cruz (Torres Vedras). Following these works, a partial skeleton of a dinosaur was collected in the locality of Pedras Muitas (Baleal, Peniche), which was identified as belonging to the stegosaur Omosaurus (Zbyszewski 1946), currently synonym of Dacentrurus. Since the early 1950s, Zbyszewski and the French paleontologist Albert Felix de Lapparent carried out a series of studies on dinosaurs from the Lusitanian Basin, which were summarized in the first monograph on dinosaurs from Portugal (Lapparent and Zbyszewski 1957). Several specimens of different dinosaurian groups were described, including theropods, sauropods, ornithopods, and thyreophorans. All theropod material was assigned to the megalosaurid Megalosaurus from the Middle Jurassic of England (e.g., Buckland 1824; Day and Barrett 2004; Benson et al. 2008; Benson 2009, 2010a), including a putative new species M. “pombali” based on some isolated teeth and few vertebrae from different localities of the region of Lourinhã, Batalha, and Pombal (Lapparent and Zbyszewski 1957). Some of these vertebrae have been reinterpreted as belonging to sauropods (Mocho et al. 2016).
Megalosaurus is the first dinosaur to be named in the scientific literature and was described based on material collected from Middle Jurassic (Bathonian) levels at Stonesfield, England (Buckland 1824). It occupied a central position in early studies of theropod dinosaurs and over time several specimens from different ages worldwide were attributed to this taxon (Benson 2008, 2010a; Carrano et al. 2012; Rauhut et al. 2016). Most of this material has never been described comprehensively or studied in a modern context and Megalosaurus became a “wastebasket” and poorly understood taxon that included material of different theropod clades, as well as other dinosaurian groups, ranging from the Upper Triassic to the latest Cretaceous (e.g., Benson et al. 2008; Benson 2010a; Carrano et al. 2012; Rauhut et al. 2016). Although the systematics of Megalosaurus is still debated (Allain and Chure 2002; Day and Barrett 2004; Carrano et al. 2012), the thorough description of the type series allowed the development of a phylogenetic framework for the recognition of the clade Megalosauroidea (Benson 2008; 2010a; Benson et al. 2008; Carrano et al. 2012). This clade includes early branching tetanuran theropods from Middle Jurassic to Upper Cretaceous strata worldwide (e.g., Rauhut 2005; Li et al. 2009; Rauhut and Lopez-Arbarello 2009; Carrano et al. 2012; Serrano-Martínez et al. 2015; Rauhut et al. 2016). Megalosauroids are particularly abundant and diverse in the Middle Jurassic of Europe and most European theropods from that time belong to this clade (e.g., Taquet and Welles 1977; Allain 2001, 2002, 2005; Allain and Chure 2002; Benson 2008, 2010a, b; Benson et al. 2008; Sadleir et al. 2008; Carrano et al. 2012; Rauhut et al. 2016). The Upper Jurassic record of megalosauroids is less diverse and so far restricted to North America (e.g., Madsen 1976b; Britt 1991; Bakker et al. 1992; Hanson and Makovicky 2014) and Europe (e.g., Mateus and Antunes 2000b; Gascó et al. 2012; Rauhut et al. 2012, 2016, 2018, 2020; Cobos et al. 2014; Hendrickx and Mateus 2014; Vullo et al. 2014; Malafaia et al. 2017c), beside some putative dental material of spinosaurids from Africa (Buffetaut 2011, but see Rauhut 2011 and Hendrickx et al. 2019 for the taxonomic affinity of this isolated tooth). Spinosauridae, traditionally considered as one of the main megalosauroid clades (but see Rauhut and Pol 2019 for a different interpretation), is well represented in Cretaceous levels of Gondwanan landmasses (e.g., Sereno et al. 1998; Sues et al. 2002; Bittencourt and Kellner 2004; Dal Sasso et al. 2005; Smith et al. 2006; Barrett et al. 2011; Kellner et al. 2011; Ibrahim et al. 2014; Evers et al. 2015; Candeiro et al. 2018; Schade et al. 2023), as well as in several localities from the northern hemisphere, being particularly well represented in different European areas (e.g., Viera and Torres 1995; Charig and Milner 1997; Buffetaut 2007; Canudo et al. 2008; Mateus et al. 2011; Alonso and Canudo 2016; Malafaia et al. 2020a, b; Barker et al. 2021; Mateus and Estravís-López 2022; Isasmendi et al. 2023) and Asia (e.g., Buffetaut and Ingavat 1986; Allain et al. 2012; Samathi et al. 2021).
Here, we describe a set of osteological material attributed to Megalosaurus collected in different localities from the Upper Jurassic and Cretaceous of the Lusitanian Basin. This material is part of the historical collections currently deposited in the Museu Municipal Leonel Trindade (Leonel Trindade Municipal Museum) of Torres Vedras as well as in the Museu Geológico (Geological Museum) and the Museu Nacional de História Natural e da Ciência of Lisbon. Beside the historical importance of this material, there is also a scientific interest, as some of the localities that yielded the material are no longer accessible. The revision of this historical material provides additional information on the fossil record of the Lusitanian Basin, which is particularly interesting for our knowledge on the Upper Cretaceous, as well as the Upper Jurassic of the Bombarral and Mamede sub-basins. We discuss the taxonomic identification of this material and update their stratigraphical context. This study allows the reinterpretation of this material as belonging to different dinosaurian groups, including theropods, sauropods, thyreophorans, and ornithopods, as well as crocodylomorphs.
Institutional abbreviations.—MG, Museu Geológico, Lisbon, Portugal; MMLT, Museu Municipal Leonel Trindade, Torres Vedras, Portugal; MNHN/UL, Museu Nacional de História Natural, Lisbon, Portugal (currently part of the MUHNAC, Museu Nacional de História Natural e da Ciência, University of Lisbon).
Other abbreviations.—AL, apical length; CA, crown angle; CBL, crown base length; CBR, crown base ratio; CBW, crown base width; CH, crown height; CHR, crown height ratio; DA, distoapical denticle density; DAVG, average distal denticle density; DB, distobasal denticle density; DC, distocentral denticle density; DSDI, denticle size density index; MA, mesioapical denticle density; MAVG, average mesial denticle density; MB, mesiobasal denticle density; MC, mesiocentral denticle density; MCL, mid-crown length; MCR, mid-crown ratio; MCW, mid-crown width; MDE, mesiobasal denticles extent; MSL, mesial serrated carina length.
Geological setting
The Lusitanian Basin is a marginal basin of west-central Portugal, whose origin is associated with the opening of the North Atlantic Ocean (e.g., Wilson 1975; Leinfelder 1987; Azerêdo et al. 1998; Rasmussen et al. 1998). Most of the basin fill is Jurassic in age, but Middle–Upper Triassic (?Ladinian–Carnian) to Upper Cretaceous strata also occur with a Cenozoic covering (e.g., Rocha et al. 1996; Azerêdo et al. 1998; Rey 1999; Kullberg et al. 2013). Betic compressional movements in the Cenozoic (probably during the Miocene) allowed the exposition of a great part of the Mesozoic sequences (Azerêdo et al. 1998). It is one of the basins on the North Atlantic margins with a greatest surficial exposition of the Mesozoic sequences. Thus, it has attracted researchers of different areas, with particular relevance for studies on the paleogeographic evolution of the peri-North Atlantic and Tethys realms (Kullberg et al. 2006). The fossil record of vertebrates from the Lusitanian Basin is well represented in the Upper Jurassic units (e.g., Dantas 1990; Antunes and Mateus 2003; Escaso et al. 2014; Mateus et al. 2009, 2014; Malafaia et al. 2015, 2020b; Mocho et al. 2017b, 2019), with abundant occurrences in the coastal region of the central sector, which is defined by the area between the Nazaré and Tejo faults (Fig. 1). These units represent a third rifting episode marked by intense subsidence, associated to terrigenous and prograding sedimentation that progressively filled the basin (e.g., Wilson 1988; Hill 1988; Rey 1992; Kullberg et al. 2006).
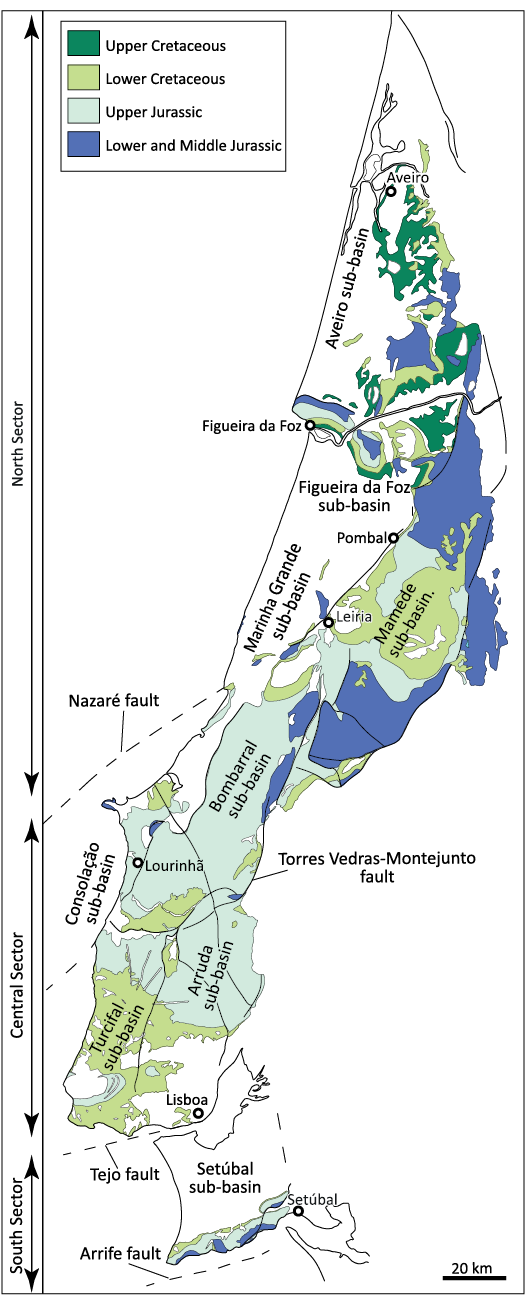
Fig. 1. Geological map (adapted from Oliveira et al. 1992) showing the Mesozoic levels and the main structures that delimit the different sectors (sensu Ribeiro et al. 1996 and Kullberg et al. 2006) and sub-basins (sensu Fürsich et al. 2021) of the Lusitanian Basin, Portugal.
The sedimentary sequences from the Upper Jurassic to the Lower Cretaceous (middle Oxfordian to Berriasian) have been interpreted as representing three main depositional stages: (i) the first, from the middle to late Oxfordian, dominated by marine and lacustrine carbonates, representing a widespread rise of relative sea-level; (ii) the second, from the latest Oxfordian to the late Kimmeridgian, marked by a maximum siliciclastic influx that corresponds to the progradation of terrestrial systems into the basin due to the increase of accommodation space created by intense tectonic subsidence; and (iii) the third, from the late Kimmeridgian to the early Berriasian, representing a passive phase during which the deposition of fluvial–deltaic sequences occurred after a short term sea-level rise (at the late Kimmeridgian), with widespread deposition of carbonate shelf sediments. These sequences are mainly represented by the upper levels of the Alcobaça and Lourinhã formations in the central sector of the basin (Pena dos Reis et al. 1996; Rasmussen et al. 1998; Alves et al. 2002; Kullberg et al. 2013). The sedimentation of the end of the Jurassic reflects the progradation of siliciclastic deposits originating from the northeastern, north, and northwestern margins of the basin to the central axis as the result of the decrease in the tectonic subsidence (Pena dos Reis et al. 2011). After the late Berriasian, the sedimentation is mostly represented by marine deposits, associated with different transgressive episodes, following the progressive oceanization of the basin (Rasmussen et al. 1998; Kullberg et al. 2013). Some carbonate-rich sequences with upwards intercalations of siliciclastic beds identified offshore indicate that these Cretaceous deposits would be well represented in the basin (Alves et al. 2003). However, Miocene inversion would contribute to the uplift and erosion of most of the post-Jurassic units (Rasmussen et al. 1998). From the late Aptian onwards, a passive margin was established in northwestern Iberia, and deposition ceased in most parts of the central and southern sectors, but not in the northern sector that records important subsidence (Rasmussen et al. 1998; Alves et al. 2002).
During the Upper Jurassic rifting phase, the Lusitanian Basin was differentiated in three main sectors (Fig. 1) with different subsidence rates and the development of fault-bounded and diapir-bounded sub-basins (e.g., Rocha and Soares 1984; Rasmussen et al. 1998; Kullberg et al. 2006). The central sector has traditionally been divided into three sub-basins (Arruda, Bombarral-Alcobaça, and Turcifal) based on isopachs and facies distribution (e.g., Leinfelder 1987; Montenat et al. 1988; Pena dos Reis et al. 1996, 2000; Alves et al. 2002). More recently, Taylor et al. (2014) defined a new sub-basin (Consolação) in the western part of the central sector of the Lusitanian Basin and Fürsich et al. (2021) subdivided the Bombarral-Alcobaça sub-basin (sensu Montenat et al. 1988) in a southern Bombarral and a northern Mamede sub-basins, which are delimited by the Porto de Mós and Leiria diapirs (Fig. 1). The central sector of the Lusitanian Basin, delimited to the north by the Nazaré fault and extending to the region of Lisbon, had maximum subsidence rates during the Upper Jurassic. The sedimentary units mainly correspond to distal fluvio-deltaic and coastal environments (Figs. 2–5) and have the richest fossil record of terrestrial vertebrates in the basin (e.g., Lapparent and Zbyszewski 1957; Dantas 1990; Martin and Krebs 2000; Antunes and Mateus 2003; Ortega et al. 2009). In the southern sector, between Lisbon and Arrábida, any Upper Jurassic osteological remains of terrestrial vertebrates are known, but some Lower Cretaceous localities (Fig. 6) have provided scarce material of dinosaurs and other vertebrates (e.g., Sauvage 1897–1898; Lapparent and Zbyszewski 1957; Jonet 1981; Galton 1994, 1996; Buffetaut 2007; Mateus et al. 2011; Figueiredo et al. 2015; Malafaia et al. 2020a; Mateus and Estravíz-López 2022). During the last decades of the 19th and the 20th centuries, Upper Cretaceous strata in the northern sector (Fig. 7) provided some fragmentary osteological material of different terrestrial vertebrates, mostly represented by isolated elements attributed to turtles, crocodylomorphs, and dinosaurs (e.g., Carrington da Costa 1940; Antunes and Pais 1978; Jonet 1981; Antunes and Sigogneau-Russell 1991; Galton 1994, 1996).
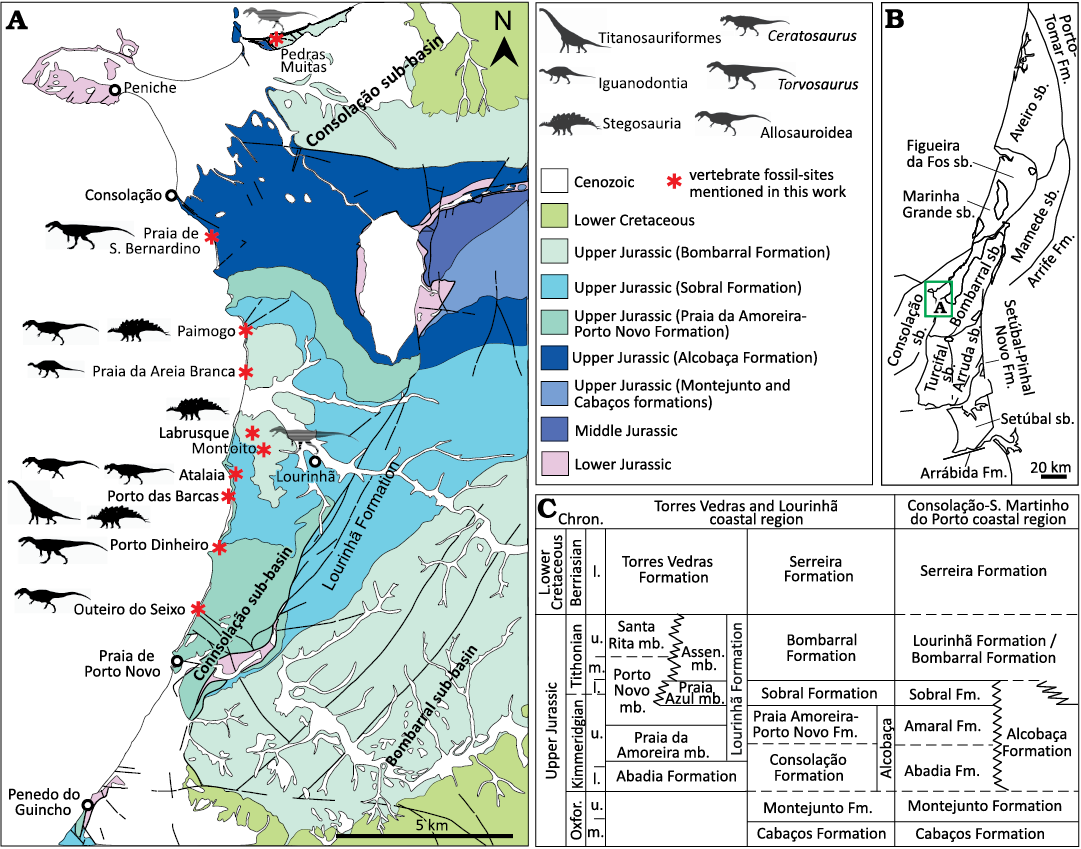
Fig. 2. Geological and stratigraphical context of fossil sites in the Consolação and Bombarral sub-basins, Portugal. A. Geological map of coastal region of Lourinhã and Peniche with localities yielding fossil remains studied in this work (modified from Camarate França et al. 1960; Manuppella et al. 1999a). B. Map of the Lusitanian Basin with the main structural elements and the different sub-basins (sensu Fürsich et al. 2021). C. Simplified stratigraphy of the Upper Jurassic and Lower Cretaceous sequences in the Torres Vedras and Lourinhã costal region and Consolação-S.Martinho do Porto coastal region based on Hill (1988), Manuppella et al. (1999a) and Azerêdo et al. (2010). Abbreviations: Assen., Assenta; Chron., Chronostratigraphy; Fm., Formation; mb., member; l., lower; m., middle; Oxfor., Oxfordian; sb., sub-basin; u., upper.
Material and methods
Material.—We have studied a set of dental and postcranial remains collected between the end of the 19th century and the first half of the 20th century (between about 1863 and the 1950s) from different Upper Jurassic, Lower, and Upper Cretaceous localities of the Lusitanian Basin, which were first attributed to Megalosaurus. This material mostly consists of isolated elements, particularly tooth fragments and vertebrae (see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app69-Malafaia_etal_SOM.pdf), which are currently in the paleontological collections of the MG and MUHNAC of Lisbon, and the MMLT of Torres Vedras.
Comparative methodology and terminology.—All the specimens are described and discussed based on first-hand analysis by all authors. We use Romerian directional descriptors (e.g., anterior, posterior) rather than the veterinarian terminology (e.g., cranial, caudal). The anatomical, positional, directional, and morphometric nomenclatures for the tooth crowns followed those proposed by Smith et al. (2005) and Hendrickx et al. (2015a).
Discriminant Function Analysis.—The morphometric analyses were performed to explore the taxonomic distribution of the isolated theropod teeth using a modified dataset provided by Hendrickx et al. (2020b). Eight measurements were taken using a digital caliper (CBL, CBW, CH, AL, MCL, MCW, MDE, and MSL), following the methodology proposed by Hendrickx et al. (2015a). Only Upper Jurassic and Cretaceous taxa with teeth of apical length greater than 12 mm were included in the analyses to reduce potential noise into the statistical analysis and because all the studied specimens have larger tooth crowns. After a first analysis with all taxa, some groups (Masiakasaurus, Baryonyx, Suchomimus, Megaraptor, Australovenator, Fukuiraptor, Velociraptor, Zanabazar, and Zapsalis) that were well separated from all the other morphospaces in the Discriminant Analysis and that were clearly not related with the studied specimens were eliminated to allow a better visualization of the results. Some taxa with fewer than five specimens (e.g., Arcovenator, Aucasaurus, Skorpiovenator, Carnotaurus, and Neovenator) were also excluded from the database as the size of the sample may influence the results of the statistical analyses (Huberty and Olejnik 2006). Denticle density data (DA, DB, DC, MA, MC, and MB) were collected using a microscope Leica Z6 with incorporated camera Leica DMC4500 (see SOM 2). All data were log-transformed prior the analyses to better reflect a normally distributed multivariate dataset (Smith et al. 2005). Statistical discriminant analyses were conducted using the software Past v4 (Hammer et al. 2001). Thirteen variables were used for these analyses (CBL, CBW, CH, AL, MCL, MCW, MDE, MA, MC, MB, DA, DB, DC). A log(x+1) correction was applied to MDE to account for the specimens in which the mesial carina extends to the cervix and an arbitrary value of 100 denticles per 5 mm was used for unserrated carinae (see Young et al. 2019 and Hendrickx et al. 2020b).
Cladistic analysis.—To support the identification of the isolated teeth we additionally performed a series of cladistic analyses based on the data matrices published by Hendrickx et al. (2020b), in which we codified the studied tooth morphotypes from the Lusitanian Basin (see SOM 3 and 4 for the TNT file and the extended strict consensus tree of 30 most parsimonious trees recovered in the cladistic analysis of the tooth-crown-based data matrix enforcing constrains). The tooth crown MNHN/UL.8628 (belonging to the Morphotype 2) and the specimens grouped in the Morphotype 3 (MG 8781, MG 8775, MG 4817, MNHN/UL.EPt.019, and MNHN/UL.EPt.004) were codified as mesial teeth. All the other specimens were considered as lateral teeth. A first cladistic analysis was conducted using the dentition-based matrix (146 morphological dental characters) with the positive constraints defined by Hendrickx et al. (2020b) and considering the studied tooth morphotypes as floating terminals. A second analysis was performed using the same dataset without constraints and a third analysis based on the tooth-crown-only data matrix consisting of tooth-based characters (characters 38 to 121 and 141 to 146, see Hendrickx et al. 2020b). However, because the strict consensus trees obtained on the second and third analyses show poor resolution, with all taxa placed in a large polytomies, only the results obtained in the first analysis are here discussed. The different morphotypes were codified using Mesquite 3.7 (Maddison and Maddison 2011) and then imported to TNT 1.6 (Goloboff and Morales 2023). The search strategy of each analysis followed the protocol used by Hendrickx et al. (2020b) and started with a combination of the tree-search algorithms: Wagner trees, TBR branch swapping, sectorial searches, Ratchet (the perturbation phase was stopped after 20 substitutions) and Tree Fusing (5 rounds), until 100 hits of the same minimum tree length were achieved. The best trees obtained were subjected to a final round of TBR branch swapping through the TNT command “bb”. Consistency (CI) and retention (RI) indices were obtained using the STATS.RUN command. A first analysis including all the studied morphotypes (except the tooth crown fragment MG 73, which is very fragmentary and distorted) was carried, but because some of them may be instable and therefore reduce the resolution of the results, we performed different analyses for each morphotype.
Systematic palaeontology
Archosauria Cope, 1869
Pseudosuchia Zittel, 1887–1890
Crocodylomorpha (Hay, 1930) sensu Walker 1970
Thalattosuchia (Fraas, 1901) sensu Young and Andrade 2009
Thalattosuchia indet.
Fig. 8.
Previous identifications: These vertebrae were described by Lapparent and Zbyszewski (1957) and assigned to Megalosaurus insignis (Lapparent and Zbyszewski 1957: 23; pl. 13: 27 and pl. 30: 121).
Material.—Two cervical vertebrae (MG 4829; Fig. 8) from Fervença, about 2 km to the north of Alcobaça, Portugal. In this region sediments mainly from the Bombarral Formation crop out, but there is a narrow area to the Oeste of Fervença where levels of the Alcobaça Formation also emerge (Fig. 3). It is, however, likely that MG 4829 comes from the Alcobaça Formation based on the description of the location provided by Lapparent and Zbyszewski (1957) and the carbonate material associated to the surface of the specimens. The age range of this formation has been considered as extending from the earliest to the latest Kimmeridgian (Azerêdo et al. 2010; Kullberg et al. 2013; Fürsich et al. 2021).

Fig. 8. Cervical vertebrae attributed to a thalattosuchian crocodyliforms from the Kimmeridgian Alcobaça Formation, Fervença (Alcobaça region, Portugal). A. MG 4829a. B. MG 4829b. Right lateral (A1, B1), left lateral (A2, B2), anterior (A3, B3), dorsal (anterior to the right, A4, B4), ventral (anterior to the left, A5, B5), and posterior (A6, B6) views.
Description.—The material consists of two small vertebral centra (see SOM 2) lacking the neural arches and transverse processes. The centra are elongate, with thick anterior and posterior margins, giving them an hourglass shape outline (Fig. 8). Their length is approximately 1.5 times their height (centrum length between 49 and 53 mm; centrum height between 33 and 40 mm). In lateral view, the ventral margin is slightly concave, forming a weakly marked sagittal ventral keel. Although severely damaged, the base of a robust process is visible in the lateral surfaces, projecting anteroposteriorly and delimiting a deep fossa ventrally approximately at the mid-height of the centrum. This process is interpreted as the base of the ramus of the parapophysis. Both vertebrae have circular to slightly dorsoventrally elongated and flat articular surfaces. The neural canal is broad. The bases of the pedicles of the neural arch are preserved and occupy almost the entire anteroposterior length of the dorsal margin of the centrum. Fragments of the diapophyses projecting from the mid-length of the dorsal margin of the centrum are preserved in both specimens.
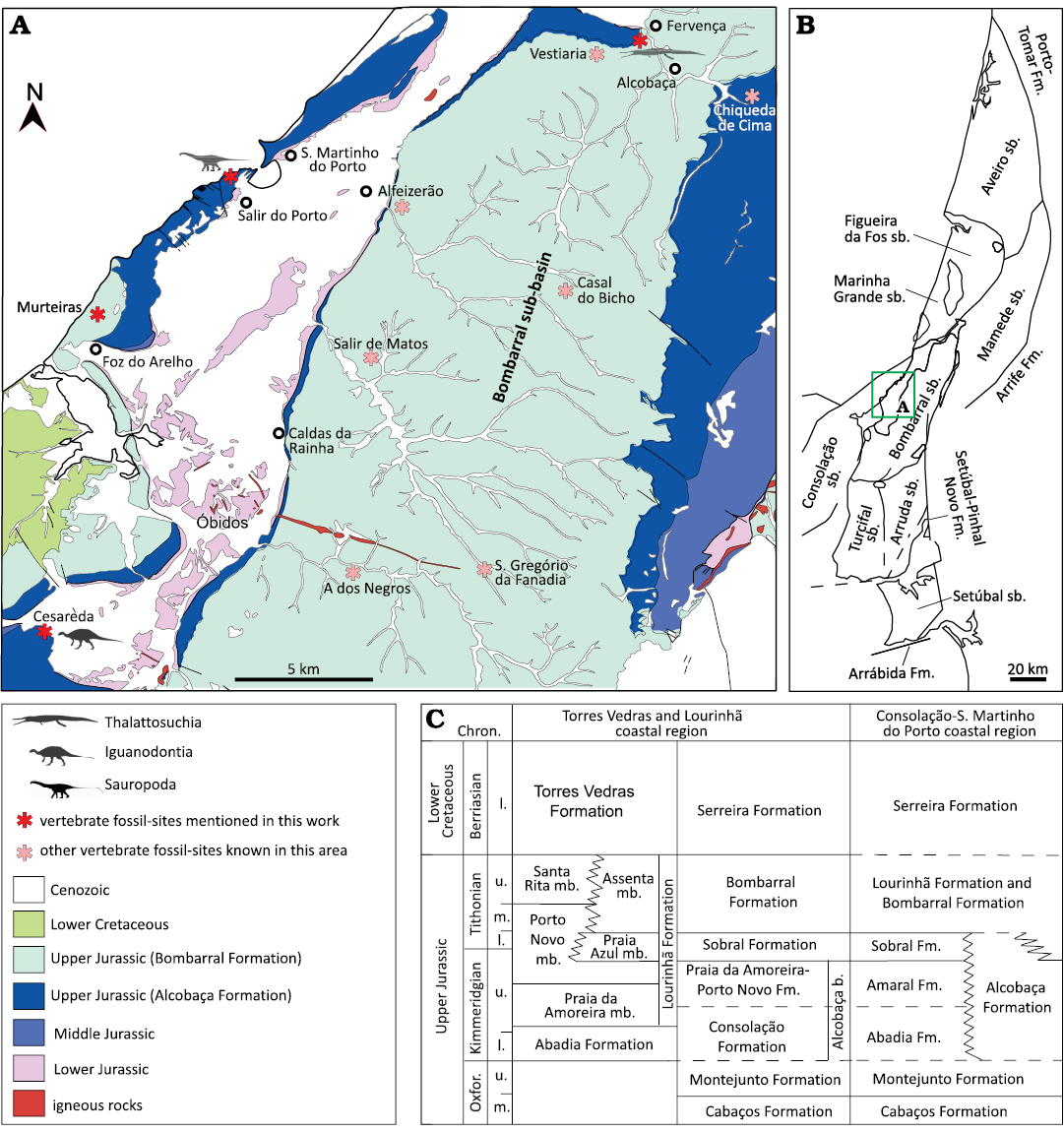
Fig. 3. Geological and stratigraphical context of fossil sites in the northern part of the Consolação sub-basin and in the Bombarral sub-basin. A. Geological map of the region between Caldas da Rainha and Alcobaça with localities yielding fossil remains studied in this work (modified from Zbyszewski and Moitinho de Almeida 1960; Camarate França and Zbyszewski 1963). B. Map of the Lusitanian Basin with the main structural elements and the different sub-basins (sensu Fürsich et al. 2021). C. Simplified stratigraphy of the Upper Jurassic and Lower Cretaceous sequences in the Consolação-S. Martinho, Torres Vedras and Lourinhã coastal regions. The stratigraphy for Torres Vedras and Lourinhã based on Hill (1988) and Manuppella et al. (1999a) and for the littoral of Consolação-S. Martinho do Porto based on Manuppella et al. (1999a) and Azerêdo et al. (2010). Abbreviations: Chron., Chronostratigraphy; Fm., Formation; mb., member; l., lower; m., middle; Oxfor., Oxfordian; sb., sub-basin; u., upper.
Remarks.—Given the preservation of the centra, it is difficult to provide a detailed comparison and to be sure where in the vertebral series they originated from. Nevertheless, the broad articular surfaces, the position of the parapophyses and the lack of supporting facets for the haemal arches allow to discard them as caudal vertebrae and suggest that they come from the cervical column. The hourglass shape outline and circular, rather than hexagonal morphology of the articular surfaces, indicate that these vertebrae can be attributed to Thalattosuchia. This group is particularly abundant in the upper Kimmeridgian of Portugal (Young et al. 2014; Johnson 2020, and references therein), and both teleosaurid and metriorhynchid thalattosuchians have been previously reported in the area of Fervença (Bardet et al. 2008).
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Thyreophora Nopcsa, 1915
Stegosauria Marsh, 1877a
Stegosauria indet.
Figs. 9–11.
Previous identifications: The label associated with MG 8798 says Megalosaurus pombali, but the specimen was not mentioned in the description of the species by Lapparent and Zbyszewski (1957). The dorsal vertebra MG 4823 is labeled as Megalosaurus sp. but it was neither figured by Sauvage (1897–1898) nor by Lapparent and Zbyszewski (1957). MG 4827 was described and figured by Lapparent and Zbyszewski (1957: 25; pl. 13: 29 and pl. 18: 50) and interpreted as belonging to an anterior dorsal vertebra of M. pombali. The anterior caudal vertebra (MNHN/UL.EPt.020) was described and figured by Lapparent and Zbyszewski (1957: 25–26, fig. 6b) as also part of the material assigned to M. pombali. Another partial vertebra from the collections of the Faculdade de Ciências was also figured by Lapparent and Zbyszewski (1957: 25–26, fig. 6a) but we could not find it in the MUHNAC collections. MNHN/UL.EPt.027 is mentioned by Lapparent and Zbyszewski (1957: 22) as part of the material assigned to Megalosaurus insignis.
Material.—A cervical and a dorsal vertebrae (MG 8798; Fig. 9), two dorsal vertebrae (MG 4823; Fig. 10A and MG 4827; Fig. 10B), a partial anterior caudal vertebra (MNHN/UL.EPt.020; Fig. 11A), and a centrum of a posterior caudal vertebra (MNHN/UL.EPt.027; Fig. 11B). MG 8798 and MNHN/UL.EPt.020 came from Porto das Barcas (Lourinhã, Portugal) and based on the description of the locality provided by Lapparent and Zbyszewski (1957) were probably collected in sediments of the Sobral Formation, corresponding to the Kimmeridgian–Tithonian interval (Manuppella et al. 1999a; Mateus et al. 2017). MG 4823 came from Paimogo (Lourinhã, Portugal) and likely came from Kimmeridgian–Tithonian Sobral Formation as well. MG 4827 is labelled Porto das Barcas (Lourinhã, Portugal), but Lapparent and Zbyszewski (1957) instead detailed that it came from Torrinhas, in the municipality of Batalha, in their description and figuration of the specimen. Outcrops of sediments from the Kimmeridgian Alcobaça Formation are present in this area (Azerêdo et al. 2010; Kullberg et al. 2013; Fürsich et al. 2021). MNHN/UL.EPt.027 was collected at Casal de Labrusque (Lourinhã, Portugal) from sediments of the Bombarral Formation, which is Tithonian in age (Manuppella et al. 1999a). See Figs. 2 and 4 for location of the fossil sites where these vertebrae were collected.
Description.—MG 8798 includes two poorly preserved vertebral centra (Fig. 9A, B). The most complete centrum (Fig. 9A), which corresponds to a cervical vertebra is elongated and slightly longer than high (see SOM 2). In left lateral view, the centrum has a shallow concavity and there is a fragment of a robust process anteriorly in the dorsal part (Fig. 9A2). The articular facets of the centrum are slightly expanded relative to the width of the centrum and have circular contours. The anterior facet (Fig. 9A3) seems slightly wider mediolaterally than the posterior facet, but this may be related with taphonomic distortion. Both facets are concave and a well-marked concavity is present in the middle of both anterior and posterior articular facets but being more pronounced in the posterior one (Fig. 9A3, A6). The ventral surface has a stout longitudinal ridge extending along the entire length of the centrum, which is more developed in the posterior part (Fig. 9A5). The centrum has a V-shape section due to the presence of this robust ventral ridge. Dorsally, the specimen preserves fragments of the base of the pedicles of the neural arch, which extend along the entire length of the dorsal margin of the centrum. The neural canal is broad, deep, and expands toward both the anterior and posterior ends (Fig. 9A4). The other vertebra (Fig. 9B) is poorly preserved, with highly distorted and incomplete centrum. The latter is short and almost as high as long. In lateral view, the centrum is slightly concave with the articular facets weakly expanded. Only one articular facet (probably the anterior one) is relatively well-preserved. This facet is concave and circular in outline. Dorsally, the specimen preserves fragments of the pedicles of the neural arch, which extend along the entire length of the centrum. The neural canal is broad mediolaterally, being about half of the mediolateral width of the anterior articular facet of the centrum. This specimen is interpreted as an anterior dorsal vertebra based on the absence of parapophyses on the centrum or the preserved fragment of the neural arch.

Fig. 9. Cervical (MG 8798a) and dorsal (MG 8798b) vertebrae attributed to indeterminate stegosaurian dinosaurs from the Kimmeridgian–Tothonian Sobral Formation, Porto das Barcas (Lourinhã region, Portugal). A. MG 8798a. B. MG 8798b. Right lateral (A1, B1), left lateral (A2, B2), anterior (A3, B3), dorsal (anterior to the left, A4, B4), ventral (anterior to the right, A5, B5), and posterior (A6, B6) views.

Fig. 10. Dorsal vertebrae attributed to indeterminate stegosaurian dinosaurs from the Kimmeridgian–Tithonian Sobral Formation, Paimogo (Lourinhã region, Portugal) and from the Kimmeridgian Alcobaça Formation, Torrinhas (Batalha region, Portugal). A. MG 4823. B. MG 4827. Left lateral (A1, B1), right lateral (A2, B2), anterior (A3, B3), ventral (anterior to the right, A4, B4), dorsal (anterior to the left, A5, B5), and posterior (A6, B6) views.

Fig. 11. Caudal vertebrae attributed to indeterminate stegosaurian dinosaurs from the Kimmeridgian–Tithonian Sobral Formation and Tithonian Bombarral Formation, Porto das Barcas and Casal de Labrusque (Lourinhã region, Portugal). A. Anterior caudal vertebra, MNHN/UL.EPt.020. B. Centrum of a posterior caudal vertebra, MNHN/UL.EPt.027. Left lateral (A1, B1), right lateral (A2, B2), dorsal (anterior to the right, A3), ventral (anterior to the left, A4, B5), anterior (A5, B3), and posterior (A6, B4) views.
MG 4823 is a dorsal vertebra with a well-preserved centrum and missing the neural arch (Fig. 10A). In lateral view the centrum is rectangular and slightly higher than long (see SOM 2). The lateral surface is slightly concave, with weakly expanded articular facets that are mostly flat, with subcircular contour (slightly higher than wide). The ventral surface is somewhat concave in lateral view and is mostly rounded in ventral view, with no visible groove or crest (Fig. 10A4). Dorsally, the specimen preserves the pedicles of the neural arch, which occupy the entire length of the centrum. The rough surface of the pedicles seems to indicate that the neural suture was open. Another dorsal vertebra (MG 4827) is represented by the relatively well-preserved centrum (Fig. 10B). It is rectangular in lateral view and higher than long. The lateral surface of the centrum has a shallow depression that occupies almost the entire dorsal part and is delimited dorsally by a well-marked longitudinal ridge (Fig. 10B1, B2). The articular facets are poorly preserved, but the most complete and presumably the posterior one has a subcircular outline, is slightly higher dorsoventrally than the mediolaterally wide, and is slightly concave (Fig. 10B6). In ventral view, the centrum is mostly rounded, approximately as wide as long, and with a well-marked transverse concavity (Fig. 10B4). The neural canal is broad mediolaterally with mostly parallel lateral margins.
The anterior caudal vertebra (MNHN/UL.EPt.020) is represented by the well-preserved centrum, including a fragment of the left transverse process and the pedicles of the neural arch (Fig. 11A). The centrum is short, with the height almost twice its length. In lateral and ventral views, the centrum is slightly concave due to the weakly expanded articular facets. A fragment of the transverse process projecting laterally is present in the dorsal end of the left lateral surface of the centrum. This process is situated in the posterior part of the centrum and projects slightly to the rear (Fig. 11A3). The anterior articular facet is mostly flat and has a subcircular outline (Fig. 11A5). The posterior facet also has a subcircular shape but is slightly concave and has a well-marked peripheral groove bounding the contour of the facet (Fig. 11A6). The ventral margin has a rough surface adjacent to the anterior articular facet, which is interpreted as the surface for the haemal arch (Fig. 11A4, A5). Dorsally, the neural arch is broken at the level of the pedicles exposing the neural canal, which is relatively narrow and circular (Fig. 11A3). The centrum, interpreted as a posterior caudal vertebra (MNHN/UL.EPt.027), is poorly preserved with an iron cover that obscures most of the morphology (Fig. 11B). The centrum is short and has a triangular shape section, strongly mediolaterally compressed ventrally. The articular facets are slightly concave and have a triangular outline, also being somewhat constricted ventrally (Fig. 11B3, B4). The ventral margin adjacent to the posterior articular facet has a well-developed facet, which is interpreted as the surface receiving the haemal arch (Fig. 11B5).
Remarks.—One of the elements included in MG 8798 corresponds to a centrum of a cervical vertebra. A centrum that is longer than wide and bears gently concave articular facets characterize stegosaurian anterior (4 to 7 according to Galton 1991) or anterior and mid-cervical centra in the case of Miragaia (Mateus et al. 2009). This condition is present in other stegosaurs such as Stegosaurus (Gilmore 1914; Maidment et al. 2015), Kentrosaurus (Hennig 1915), Dacentrurus (Cobos and Gascó 2013; Escaso 2014; Allain et al. 2022), and Miragaia (Mateus et al. 2009; Allain et al. 2022). In addition, the presence of a similar concavity in both articular facets could relate this vertebra with the neck cervical vertebra 7 (Galton 1991). A ventral keel is shared with other stegosaurs such as Stegosaurus (Gilmore 1914; Maidment et al. 2015) and Dacentrurus (Galton 1991; Escaso 2014). The other element included in MG 8798 is a dorsal vertebral centrum characterized by having similar length and width, a condition typical of the middle and posterior dorsal vertebrae of Dacentrurus (Cobos and Gascó 2013). The dorsal vertebrae MG 4823 and MG 4827 are characterized by having centra wider than long, a condition shared with stegosaurs described in the Portuguese Jurassic record such as Dacentrurus (Galton 1991; Allain et al. 2022) and Miragaia (Costa and Mateus 2019). This condition is exclusive of these two stegosaur genera (see Allain et al. 2022) and the North African taxon Adratiklit (Maidment et al. 2020). MNHN/UL.EPt.020 is an anterior caudal vertebra based on its short centrum length, its circular contour, and the presence of prominent transverse processes (Galton 1991; Galton and Upchurch 2004). The transverse process bears a dorsal process, which is a condition commonly seen in stegosaurs such as Dacentrurus (Galton 1991; Cobos et al. 2010), Stegosaurus (Gilmore 1914; Maidment et al. 2015), Hesperosaurus (Carpenter et al. 2001) and Kentrosaurus (Henning 1915). However, Dacentrurus, Miragaia and Kentrosaurus show a moderately developed dorsal process (Henning 1915; Galton 1991; Cobos et al. 2010; Costa and Mateus 2019). As for MNHN/UL.EPt.027, the absence of transverse processes in the centrum is a condition of the posterior region of the caudal series. Based on the incompleteness of this set of vertebrae and their combination of features we prefer to refer all of them to Stegosauria indet.
Ornithopoda Marsh, 1881
Iguanodontia Dollo, 1888
Iguanodontia indet.
Figs. 12, 13.
Previous identifications: The label associated with the sacral vertebra (MG 8786) identifies it as belonging to Megalosaurus pombali, but this specimen was not mentioned by Lapparent and Zbyszewski (1957) in the description of the species. The caudal vertebra (MG 4825) was described by Lapparent and Zbyszewski (1957: pl. 25: 87) and assigned to Megalosaurus insignis by these authors. The two articulated caudal vertebrae (MG 4820) were described and figured by Lapparent and Zbyszewski (1957: 23; pl. 14: 34 and pl. 15: 40) and assigned to M. insignis. This specimen was associated with a sequence of five articulated caudal vertebrae collected in the same area (Lapparent and Zbyszewski 1957: fig. 4) and currently housed in the collections of the MUHNAC. Based on a preliminary redescription of this last specimen, Escaso et al. (2017) suggested that it belonged to an ornithopod dinosaur closely related to Ankylopollexia. The posterior caudal centrum (MNHN/UL.EPt.024) was mentioned by Lapparent and Zbyszewski (1957: 23) as part of the same set as the two articulated vertebrae (MG 4820) and the sequence of five caudal vertebrae (MNHN/UL.EPt.028).
Material.—A sacral vertebra (MG 8786; Fig. 12A), an isolated dorsal vertebra (MG 4825; Fig. 12B), two articulated anterior caudal vertebrae (MG 4820; Fig. 12C), and a posterior caudal centrum (MNHN/UL.EPt.024; Fig. 13). MG 8786 comes from the Kimmeridgian Alcobaça Formation (Azerêdo et al. 2010; Kullberg et al. 2013; Fürsich et al. 2021) of the locality of Torrinhas, in the municipality of Batalha (Portugal) (Fig. 4). MG 4825 comes from Cesareda, near the locality of Olho Marinho of the municipality of Óbidos (Portugal) (Fig. 3), where outcrops from the Kimmeridgian Alcobaça Formation are exposed (Azerêdo et al. 2010; Fürsich et al. 2021). The label associated with MG 4820 indicates the locality of Casais da Pedreira (Lourinhã, Portugal), but Lapparent and Zbyszewski (1957) argued that the specimen comes from Praia da Areia Branca, in the coastal region of Lourinhã (Fig. 2). The specimen was probably collected in sediments of the Tithonian Bombarral Formation (Manuppella et al. 1999a). MNHN/UL.EPt.024 was also collected in Praia da Areia Branca.
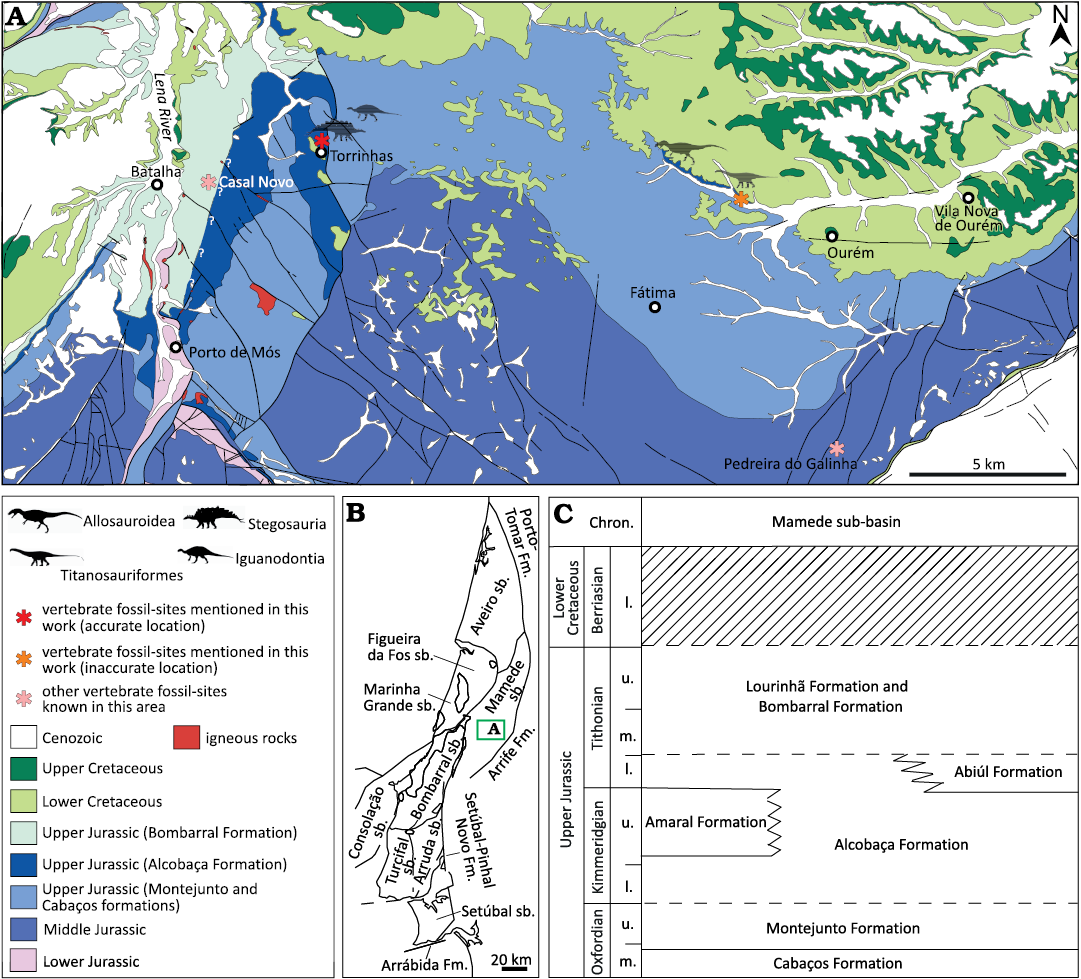
Fig. 4. Geological and stratigraphical context of fossil sites in the Mamede sub-basin. A. Geological map of the region of Batalha and Ourém with localities yielding fossil remains studied in this work (modified from Teixeira et al. 1968; Manuppella et al. 2000). B. Map of the Lusitanian Basin with the main structural elements and the different sub-basins (sensu Fürsich et al. 2021). C. Simplified stratigraphy of the Upper Jurassic sequences in the Mamede sub-basin based on Kullberg et al. (2013) and Fürsich et al. (2021). Abbreviations: Chron., Chronostratigraphy; Fm., Formation; mb., member; l., lower; m., middle; sb., sub-basin; u., upper.
Description.—The sacral vertebra (MG 8786) lacks the neural arch, but the centrum is complete and well-preserved (Fig. 12A). In lateral view, the centrum is slightly elongated, being somewhat longer than high (see SOM 2). The lateral surface is shallowly concave and the articular facets are somewhat expanded. The articular facets have circular contours and are almost flat and highly rugose, with a series of grooves and ridges that extend divergently from the central part (Fig. 12A3, A4). A transverse groove is present in the dorsal part of both anterior and posterior facets. In the dorsal margin of the centrum there is a pair of rough surfaces adjacent to the lateral margin of the anterior and posterior facets, which are interpreted as the sutures for the sacral ribs. The posterior suture is a small concave surface with an oval shape and the anterior one is much broader extending to almost the mid-length of the centrum (Fig. 12A1). The neural canal is mediolaterally broad and bears a pair of small foramina in the posterior part (Fig. 12A6). In ventral view, the centrum is rectangular, broad mediolaterally, and has a shallow longitudinal concavity that extends for most of the length (Fig. 12A5). The ventral surface is concave in lateral view. The dorsal vertebra (MG 4825) preserves only the centrum, which is subquadrangular in lateral view, almost as high as it is long (Fig. 12B). The lateral surface is slightly concave, and the ventral margin is almost straight. The articular facets are subcircular (possibly due to distortion as the left margin is broken) and almost flat. The dorsal end preserves fragments of the pedicles of the neural arch, which extend along the entire length of the centrum. The neural canal is broad mediolaterally and rectangular, slightly expanded adjacent to the articular facets (Fig. 12B6). In ventral view, the centrum is mostly circular with a low longitudinal ridge.
MG 4820 is represented by two articulated caudal vertebrae that preserve the centra and parts of the neural arch (Fig. 12C). The centra are subquadrangular in lateral view, slightly higher than long, with shallow transverse concavities. The left lateral surface has a well-marked foramen in the ventral part of the centrum of both vertebrae. The anterior articular facet has a circular contour, but the posterior facet is oval, much higher than wide, and both are slightly concave. The ventral surface has well-developed facets for the haemal arches adjacent to the ventral margin of both anterior and posterior articular facets. The ventral surface has a shallow longitudinal groove, which is delimited by low crests extending between the facets for the haemal arches (Fig. 12C4). The neural arches preserve fragments of the transverse processes, the base of the neural spines, and partial pre- and post-zygapophyses. The transverse processes are incomplete, but the preserved fragments suggest that they were robust and projecting laterally from almost the entire length of the dorsal margin of the centrum (Fig. 12C2). The ventral surface of the transverse processes is slightly concave longitudinally. Only the base of the neural spines is preserved, so their morphology is mostly unknown. From the preserved fragment can be inferred that these processes project from the middle of the neural arch. In the most anterior of the two vertebrae, where the spine is more complete, it seems to project somewhat posteriorly. In dorsal view, there is a well-marked concavity between the transverse process and the neural spine. The prezygapophyses are short and project anterodorsally with the articular facets facing medially, whereas the postzygapophyses face laterally. In dorsal view, the prezygapophyses are mostly parallel with a mediolateral width approximately equal to the width of the centrum.

Fig. 12. Vertebrae attributed to indeterminate iguanodontian dinosaurs from the Kimmeridgian Alcobaça Formation and the Tithonian Bombarral Formation, Portugal. A. Sacral vertebra, MG 8786, Torrinhas (Batalha region). B. Dorsal vertebra, MG 4825, Cesareda (Óbidos region). C. Two articulated anterior caudal vertebrae, MG 4820, Casais da Pedreira (Lourinhã region). Left lateral (A1–C1), right lateral (A2–C2), anterior (A3–C3), posterior (A4, B4, C6), ventral (A5, B5, C4), and dorsal (A6, B6, C5) views.
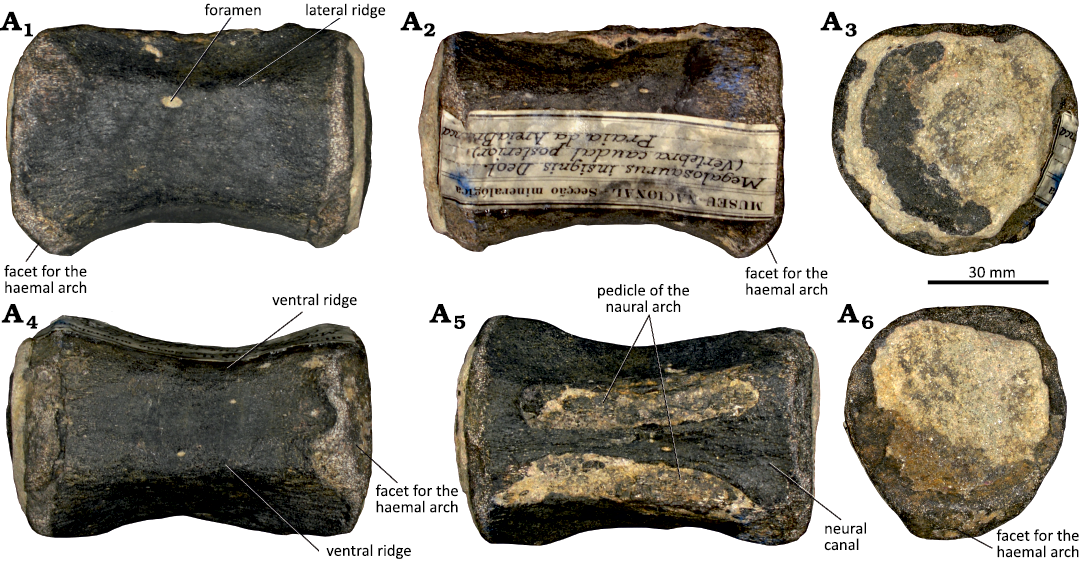
Fig. 13. Posterior caudal centrum (MNHN/UL.EPt.024) attributed to an iguanodontian dinosaur from the Tithonian Bombarral Formation, Praia de Areia Branca (Lourinhã region, Portugal) in right lateral (A1), left lateral (A2), anterior (A3), ventral (anterior to the left, A4), dorsal (anterior to the left, A5), and posterior (A6) views.
The posterior caudal centrum (MNHN/UL.EPt.024) is elongated, rectangular in lateral view and with a slightly concave ventral margin (Fig. 13). The lateral surface has a longitudinal ridge extending almost the entire length of the centrum in the dorsal part, just ventral to the pedicles of the neural arch (Fig. 13A1). A relatively large foramen is placed adjacent to this ridge near the mid-length of the centrum. The articular facets are slightly expanded. The anterior facet is subcircular, whereas the posterior one is oval, slightly dorsoventrally higher than its mediolateral width. The anterior articular facet of the centrum is slightly concave as is also probably the posterior one, although the presence of a layer of sediment does not allow this to be confirmed. The ventral surface has a shallow and mediolaterally broad longitudinal concavity that is delimited laterally by low longitudinal crests (Fig. 13A4). A well-developed surface for the haemal arch is present in the ventral margin of the posterior articular facet of the centrum (any surface for the haemal arch is visible near the anterior articular facet). The centrum has a somewhat quadrangular section due to the presence of the ventral and the previously described lateral ridges. The neural arch is absent and broken at the level of the base of the pedicles, exposing the neural canal, which is narrow but strongly expands to the anterior and posterior ends (much more markedly to the posterior part).
Remarks.—MG 8786 corresponds to a sacral vertebra showing a similar morphology of the mid-sacrals of other Late Jurassic iguanodontians as Cumnoria or Uteodon, with centrum articular facets wider than tall (Carpenter and Wilson 2008; Maidment et al. 2022). Like the latter, the ventral surface of the centrum is slightly grooved. MG 4825 is a centrum of a dorsal vertebra similar to those of other Late Jurassic iguanodontian such as Camptosaurus, Cumnoria, and Uteodon (Gilmore 1909; Carpenter and Wilson 2008; Maidment et al. 2022). As in the latter, the ventral side of this centrum retains a smooth ventral keel (Carpenter and Wilson 2008). The morphology of MG 4820 is similar to those of anterior caudal vertebra of Late Jurassic iguanodontian ornithopods as Camptosaurus, Cumnoria, Draconyx, or Dryosaurus (Gilmore 1909; Galton 1981; Maidment et al. 2022; Rotatori et al. 2022). As is the case of the anterior caudal vertebrae of Camptosaurus, Cumnoria, Uteodon, Eousdryosaurus, or Dryosaurus, the transverse processes are in the center of the arch surface (Gilmore 1909; Galton 1981; Carpenter and Wilson 2008; Escaso et al. 2014; Maidment et al. 2022). As in these taxa, except Eousdryosaurus, the centra of MG 4820 are equally long as high. Despite its incompleteness the orientation of the neural spine is similar to those of Camptosaurus, Uteodon, Eousdryosaurus, or Dryosaurus in being inclined posteriorly (Gilmore 1909; Galton 1981; Carpenter and Wilson 2008; Escaso et al. 2014). MNHN/UL.EPt.024 is similar to the posterior caudal vertebrae of ornithopod dinosaurs such as Cumnoria or Dryosaurus (Galton 1981; Maidment et al. 2022). Based on this combination of features, the vertebrae discussed here are assigned to Iguanodontia indet.
Sauropoda Marsh, 1878
Sauropoda indet.
Previous identifications: The partial metacarpal (MNHN/UL.EPt.026) was interpreted by Lapparent and Zbyszewski (1957: 23 and 26, fig. 5) as a fragment of a femur assigned to Megalosaurus insignis. The fragments of the caudal vertebrae (MG 8782) were mentioned by Lapparent and Zbyszewski (1957: 23) as part of the material attributed to M. insignis.
Material.—A fragment of the distal part of a metacarpal (MNHN/UL.EPt.026; SOM 5: fig. S1A1–A5) and three fragments of middle or posterior caudal centra (MG 8782; SOM 5: fig. S1B1–B3, C1–C2, and D1–D2). MNHN/UL.EPt.026 came from Porto das Barcas (Lourinhã, Portugal) and was probably collected from levels of the Kimmeridgian–Tithonian Sobral Formation (Fig. 2) (Manuppella et al. 1999a; Mateus et al. 2017). The vertebrae MG 8782 are labeled as coming from Salir do Porto, in the municipality of Caldas da Rainha (Portugal) (Fig. 3). They were probably collected in the coastal region of Salir do Porto where an extensive sedimentary sequence interpreted as belonging to the Alcobaça Formation is exposed (Azerêdo et al. 2010; Kullberg et al. 2013; Fürsich et al. 2021).
Description.—MG 8812 is reinterpreted here as a fragment of the distal end of a metacarpal, which preserves the articular surface and a small part of the diaphysis (SOM 5: fig. S1A1–A5). The articular surface is 125 mm long mediolaterally and 93 mm wide anteroposteriorly. In anterior view, the diaphysis is broad mediolaterally and the element expands distally. The articular surface extends onto the anterior surface of the metacarpal, especially medially. The lateral surface is concave distally and has a well-developed pit for the lateral ligaments. The medial surface on the contrary does not have a ligament pit but only a shallow concavity. In posterior view, the diaphysis is narrower than the anterior surface and has a low longitudinal crest near the mid-width of the diaphysis. Distally there is a shallow and narrow groove between the articular condyles. In distal view, the articular surface is rounded and rectangular, with the anterior margin slightly longer than the posterior one. The distal surface is perpendicular relatively to the axis of the shaft. MG 8782 include three fragments of mid-posterior caudal centra (SOM 5: fig. S1B1–B3, C1, C2, D1, D2). These fragments are too incomplete to allow a detailed description. The internal bone tissue is solid and there are no vascular foramina. The interpreted anterior and posterior articular surfaces are both concave. The ventral surface of the centrum is transversely flat.
Remarks.—The incompleteness of MG 8812 makes it difficult to assign the element to a more accurate position in the manus. The relative symmetrical shape of the distal end and the absence of any deflection of the distal surface suggest that this fragment may belong to a metacarpal II, III or IV. However, the presence of a developed ligament pit only in the lateral side of the distal end, might indicate that this element more likely corresponds to a metacarpal II or IV than to a metacarpal III. The extension of the articular surface to the anterior margin of the shaft is common in non-titanosauriform sauropods (some titanosauriforms might present this feature for metacarpal IV; D’Emic 2012; Mannion et al. 2013). In the absence of more relevant anatomical data, this element is referred to Sauropoda indet. The mid-posterior caudal vertebrae of MG 8782 are too fragmentary to allow a detailed comparison and more precise systematic approach and are also referred to Sauropoda indet.
Eusauropoda Upchurch, 1995
Eusauropoda indet.
Fig. 14.
Previous identifications: The fragment of the humerus (MG 8812) was described and figured by Lapparent and Zbyszewski (1957: 23, pl. 14: 37) and interpreted as a fragment of an ulna of Megalosaurus insignis.
Material.—A portion of the proximal end of a right humerus (MG 8812; Fig. 14), labeled as coming from Vila Nova de Ourém (Portugal). The sedimentary levels around the locality of Vila Nova de Ourém consist mostly of calcareous deposits of Lower Cretaceous age and Oxfordian sedimentary levels of the Cabaços and Montejunto formations to the south and northeast (Fig. 4). However, there is a narrow area to the northeast of Ourém where levels interpreted as belonging to the Alcobaça Formation crops out (Manuppellla et al. 2000; Mocho et al. 2017b). The dinosaur fossil remains labeled as coming from Ourém or Vola Nova de Ourém were probably collected from these levels, which are Kimmeridgian in age (Fürsich et al. 2021).
Description.—MG 8812 is a poorly preserved proximal end of a right humerus, lacking the most proximal part, but preserving a taphonomically fractured and medially displaced deltopectoral crest (Fig. 14). The anterior surface is widely concave and laterally bordered by the deltopectoral crest. This crest is constricted dorsoventrally, and its posterior face preserves a strong posterolateral bulge, which possibly corresponds to the site for M. scapulohumeralis anterior (Upchurch et al. 2015). The posterior face is transversely convex with no visible ridge. The shaft is elliptical, anteroposteriorly compressed, and the eccentricity (mediolateral to anteroposterior width ratio at midshaft) of the preserved portion of the shaft is 2.78.

Fig. 14. Portion of the proximal end of a right humerus (MG 8812) attributed to an indeterminate eusauropod dinosaur from the Kimmeridgian Alcobaça Formation, Ourém region (Portugal) in posterior (A1) and anterior (A2) views.
Remarks.—MG 8812 is characterized by the presence of a posterolateral bulge posterior to the deltopectoral crest, which is present in many titanosauriforms and turiasaurs (e.g., Mateus et al. 2014; Upchurch et al. 2015; Mannion et al. 2019). The dorsoventrally constricted deltopectoral crest seems to be exclusive of turiasaurs and brachiosaurids (e.g., Royo-Torres et al. 2006; Mateus et al. 2014; Mocho et al. 2017a). This combination of features allows us to attribute this incomplete humerus to an indeterminate eusauropod.
Neosauropoda Bonaparte, 1986
Macronaria Wilson and Sereno, 1998
Titanosauriformes Salgado et al., 1997
Titanosauriformes indet.
Fig. 15.
Previous identifications: The anterior caudal vertebra (MMLT.002528) was described by Lapparent and Zbyszewski (1957: pl. 13: 31–33) as a posterior dorsal vertebra of Megalosaurus pombali. Later, Mocho et al. (2017b: 23) reinterpreted this vertebra as an anterior caudal vertebra of an indeterminate sauropod.
Material.—A partial anterior caudal vertebra (MMLT.002528; Fig. 15) from Porto das Barcas (Lourinhã, Portugal) and was probably collected from levels of the Sobral Formation (Fig. 2), which is Kimmeridgian–Tithonian in age (Manuppella et al. 1999a; Mateus et al. 2017).
Description.—The anterior caudal vertebra (MMLT.002528) is represented by a relatively well-preserved centrum as well as parts of the neural arch, including the pedicles, and the transverse processes (Fig. 15). The centrum is robust, short, and high, with slightly expanded articular surfaces. The anterior articular surface is oval in outline and strongly concave. The posterior one is incomplete but seems to be more triangular, mediolaterally larger dorsally than ventrally, flat to slightly convex near the edges. The lateral surface of the centrum is concave anteroposteriorly and slightly concave below the transverse processes, which are placed in the dorsal part of the lateral surface. Small and shallow vascular foramina pierce the lateral and ventral surfaces. The transverse processes extend to the lateral surface of the neural arch. They are incomplete but the preserved fragments suggest that they would be robust as the base occupies almost the entire length of the centrum. In dorsal view, the transverse processes are directed posteriorly. The ventral surface is transversely flat and slightly concave near the facets for the haemal arch (the anterior one is not preserved) differing from the well-developed ventral concavity of many diplodocoids and titanosaurs (e.g., Wilson 2002; Upchurch et al. 2004; Tschopp et al. 2015). The centrum has a V-shape cross-section that is narrower ventrally than dorsally. The ventral margin of the posterior articular surface has a well-developed and subtriangular facet for the haemal arch. Dorsally, only the base of the pedicles of the neural arch is preserved and they are placed in the anterior part of the dorsal margin of the centrum. The neural canal is broad and circular.
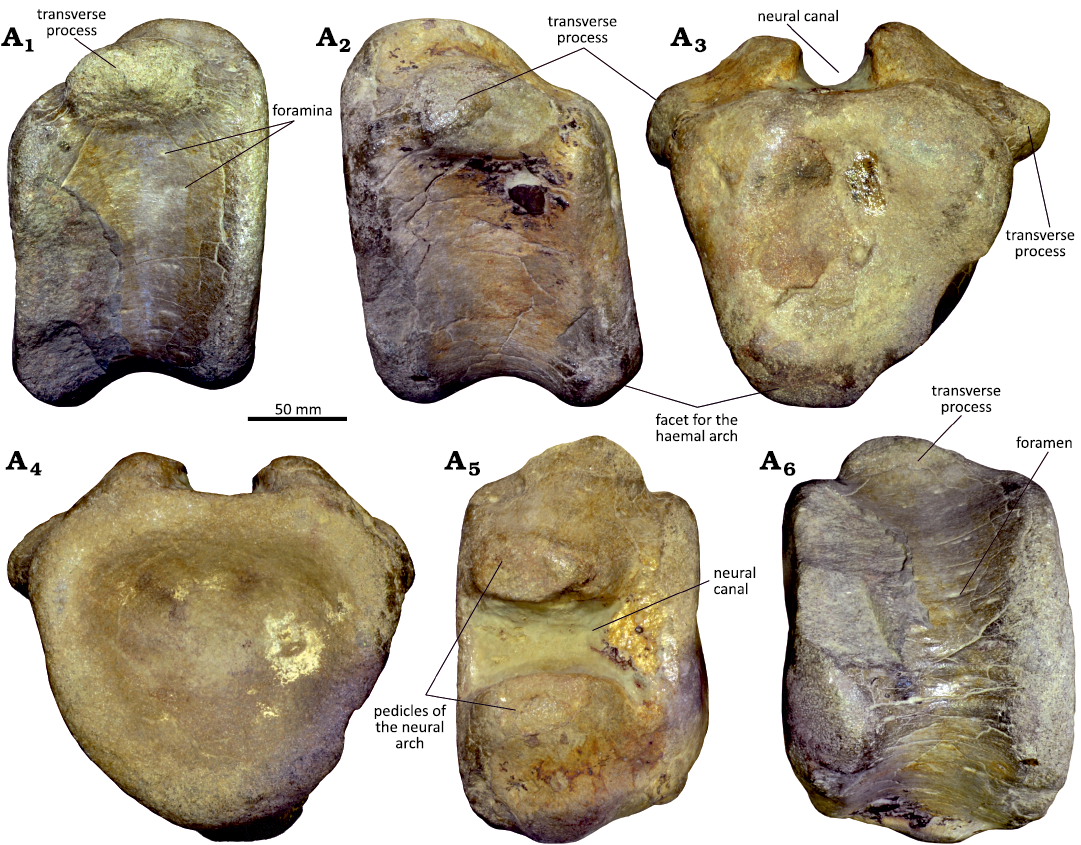
Fig. 15. Partial anterior caudal vertebra (MMLT.002528) attributed to an indeterminate Titanosauriformes sauropod from the Kimmeridgian–Tithonian Sobral Formation, Porto das Barcas (Lourinhã region, Portugal) right lateral (A1), left lateral (A2), posterior (A3), anterior (A4), dorsal (anterior to the left, A5), and ventral (anterior to the right, A6) views.
Remarks.—The anterior caudal vertebra (MMLT.002528) is characterized by a flat posterior articular surface as in many non-titanosaurian macronarians such as Camarasaurus supremus (Osborn and Mook 1921), Lourinhasaurus alenquerensis (Mocho et al. 2014), Brachiosaurus altithorax (D’Emic 2012), Lusotitan atalaiensis (Mocho et al. 2017a) and Aragosaurus ischiaticus (Royo-Torres et al. 2014). The presence of small vascular foramina in the lateral surface of the centrum is shared by many Titanosauriformes and they are present in Lusotitan (Mannion et al. 2013; Mocho et al. 2017b). However, some non-titanosauriform taxa, including some diplodocoids and non-neosauropods, also have small vascular foramina in their anterior-middle centra (Mannion et al. 2013, 2019). Posteriorly oriented transverse processes are also common in titanosauriformes and some non-neosauropod eusauropods (e.g., Mannion and Calvo 2011; Mannion et al. 2013, 2019). This combination of features seems to be exclusive of titanosauriforms, supporting the attribution of this anterior caudal vertebra to Titanosauriformes indet.
Theropoda Marsh, 1881
Theropoda indet.
SOM 5: fig. S2.
Previous identifications: Most of the tooth crown fragments are labeled as Megalosaurus insignis and some of them were mentioned by Sauvage (1897–1898), Lapparent and Zbyszewski (1957), and Mateus (2005), although none of these authors figured them. The only exceptions are: (i) a specimen collected in the region of Pombal (MG 8778; SOM 5: fig. S2A1–A4), which is part of the material used for the description of Megalosaurus pombali (Lapparent and Zbyszewski 1957: 25; pl. 28: 105) and (ii) the tooth fragment from Viso (MG 73; SOM 5: fig. S2E1–E5) that is part of a set of three tooth fragments described by Sauvage (1897–1898: 40, pl. 8: 16) as belonging to Megalosaurus sp. Later, Lapparent and Zbyszewski (1957: 27, pl. 12: 4, 5, 20) attributed these three specimens (including MG 73) as belonging to Megalosaurus cf. pannoniensis. The ungual phalanges (MNHN/UL.EPt.025a and b) were figured by Lapparent and Zbyszewski (1957: 23, pl. 12: 18, 22) and attributed to Megalosaurus sp.
Material.—Several fragments of tooth crowns (MG 14, 4812, 4815, 4816, 8776, 8778; SOM 5: fig. S2A1–A4; MG 25199, MNHN/UL.EPt.022, SOM 5: fig. S2B1–B6; MG 73, SOM 5: fig. S2E1–E5) and two small ungual phalanges (MNHN/UL.EPt.025, SOM 5: fig. S2C1–C4, D1–D4). The label associated to MG 4812 indicates that this specimen comes from the “Lusitanian” of Ourém (Portugal) (Fig. 4) and was probably collected from Kimmeridgian levels of the Alcobaça Formation (Fürsich et al. 2021). The tooth fragments (MG 4815) were collected at Murteiras, in the municipality of Foz do Arelho (Portugal), from sedimentary rocks of the Tithonian Bombarral Formation (Fig. 3) (Manuppella et al. 1999a). The tooth fragments (MG 14) are labeled as coming from a locality southwest of Vermoil, in the municipality of Pombal (Portugal). The sedimentary levels around this locality correspond mainly to Lower Cretaceous deposits (Fig. 5), but a large section of the “Complexo de vale de Lagares” crops out southeast to Vermoil (Teixeira et al. 1968). These levels have been recently included in the Alcobaça Formation (Fürsich et al. 2021). It is probable that the tooth fragments were collected from these Kimmeridgian levels of the Alcobaça Formation. Other tooth fragments (MG 4816, 8778, 25199) are labeled as coming from Pombal (Portugal), but more precise geographic information is not available for these specimens. MG 8776 was collected at Atalaia (Lourinhã, Portugal), from Kimmeridgian–Tithonian deposits of the Sobral Formation (Fig. 2). A tooth crown (MNHN/UL.EPt.022) and the ungual phalanges (MNHN/UL.EPt.025) came from Porto Dinheiro (Lourinhã, Portugal) and were probably collected in upper Kimmeridgian–lowermost Tithonian levels of the Praia da Amoreira-Porto Novo Formation (Fig. 2) (Manuppella et al. 1999a). Finally, MG 73 was collected at the locality of Viso, in the region of Montemor-o-Velho (Coimbra, Portugal). This specimen came from a fossil site found at km 20 of the railway (Sauvage 1897–1998), where a predominantly sandstone sequence of the “Sandstones and Mudstones of Viso” (Barbosa et al. 2008) crops out (Fig. 7). The study of the palynological record in some areas within this unit (here referred to as “Viso Formation”) indicates an age ranging from the Santonian to the Campanian with possible extension into the early Maastrichtian (Barbosa et al. 2008).
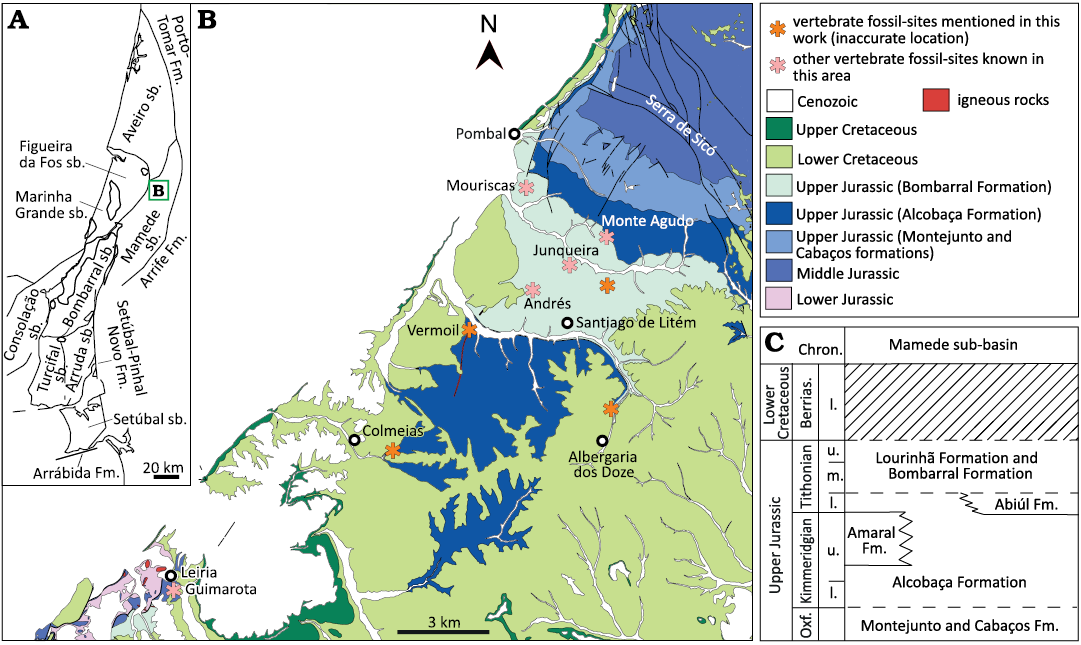
Fig. 5. Geological and stratigraphical context of fossil sites in the Mamede sub-basin. A. Map of the Lusitanian Basin with the main structural elements and the different sub-basins (sensu Fürsich et al. 2021). B. Geological map map of the region of Pombal and Leiria with localities yielding fossil remains studied in this work (modified from Teixeira et al. 1968; Manuppella et al. 1978). C. Simplified stratigraphy of the Upper Jurassic sequences in the Mamede sub-basin based on Kullberg et al. (2013) and Fürsich et al. (2021). Abbreviations: Berrias., Berriasian; Chron., Chronostratigraphy; Fm., Formation; mb., member; l., lower; m., middle; Oxf., Oxfordian; sb., sub-basin; u., upper.
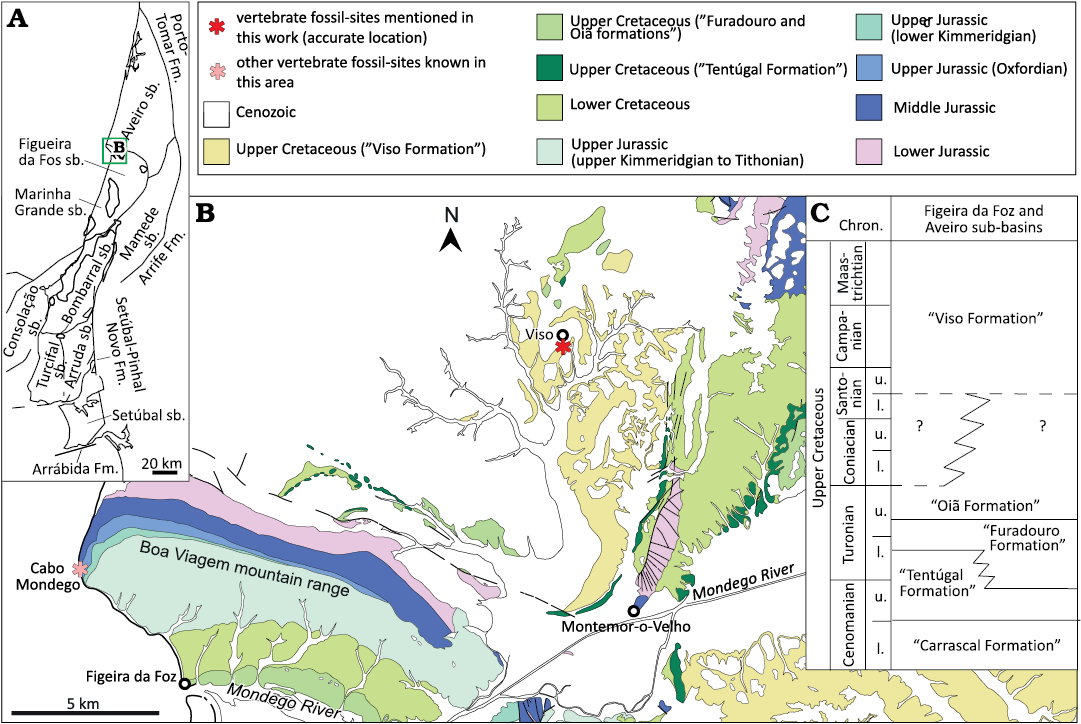
Fig. 7. Geological and stratigraphical context of the Figeira de Foz and Aveiro region. A. Map of the Lusitanian Basin with the main structural elements and the different sub-basins (sensu Fürsich et al. 2021). B. Geological map of Figueira da Foz and Aveiro region (Aveiro sub-basin) with localities yielding fossil remains studied in this work (modified from Rocha et al. 1981 and Barbosa et al. 2008). C. Simplified stratigraphy of the Upper Cretaceous sequences in the Figueira da Foz and Aveiro sub-basins based on Barbosa et al. (2008). Abbreviations: Chron., Chronostratigraphy; Fm., Formation; mb., member; l., lower; m., middle; sb., sub-basin; u., upper.
Description.—With the exception of one specimen (MNHN/UL.EPt.22), which is an almost complete tooth crown (SOM 5: fig. S2B1–B6), all the other specimens are particularly incomplete and distorted. MNHN/UL.EPt.22 is covered by a layer of sediment obscuring most of the denticles and the enamel ornamentation, preventing them to be described. This specimen corresponds to a relatively large (AL = 22.55), elongated (CHR = 2.32) and moderately compressed (CBR = 0.64) crown. It is triangular in lateral view and slightly recurved distally with the mesial margin strongly convex, but the distal margin is mostly straight. The distal carina is visible in a small part of the central sector of the distal margin and bears a large number of small denticles (21 denticles per 5 mm). The latter are rectangular (slightly longer mesiodistally than wide apicobasally), extend perpendicular to the carina, and with a symmetrically convex external margin (SOM 5: fig. S2B5). The tooth fragment from Viso (MG 73) is represented by a relatively well-preserved tooth crown without the apex (SOM 5: fig. S2E1–E5). Despite the absence of the latter, the preserved fragment suggests that it would be relatively elongated relative to the basal mesiodistal length. The crown is labiolingually compressed (CBR = 0.5), with a lanceolate basal section. The distal carina has poorly preserved denticles extending to the cervix and the mesial carina, which seems to have also denticles, but it is incomplete, and their extension cannot be determined. The distal carina is centrally placed on the distal surface. Interdenticular sulci, marginal and transverse undulations are not visible in the preserved fragment of the crown. Both labial and lingual surfaces are slightly convex. The enamel is mostly smooth or has a very subtle irregular ornamentation.
The ungual phalanges (MNHN/UL.EPt.025a and b) are slightly recurved ventrally and have well-developed lateral grooves, which are mostly symmetrical and extend along the entire length (SOM 5: fig. S2C1–C4, D1–D4). The proximal end of both phalanges is missing, which makes it difficult to determine their anatomical position and to discuss their taxonomy in more detail. MNHN/UL.EPt.025a (SOM 5: fig. S2C1–C4) has a triangular section, with a rounded dorsal surface and a mostly flat ventral margin whereas the other ungual, MNHN/UL.EPt.025b (SOM 5: fig. S2D1–D4) is much narrower mediolaterally with a blade-like section.
Remarks.—Most of the tooth crown fragments are incomplete, but several of them show evidence of denticles and can therefore be attributed to indeterminate theropods. MNHN/UL.EPt.22 is a relatively large denticulated ziphodont teeth with mesial carina that appears to terminate above the cervix. These features together with the currently known record of Late Jurassic theropods from the Lusitanian Basin indicate that this specimen likely belong to an indeterminate averostran theropod. However, we have not been able to find any other feature that would allow a more precise identification. The reclassification rate of the performed discriminant analysis is 70.91% and the average of cases correctly classified is 60% for non-abelisauroid Ceratosauria (Genyodectes, 78%; Ceratosaurus, 41%), 62% for Abelisauroidea (Abelisaurus, 60%; Indosuchus, 50%; Majungasaurus, 75%), 92% for Megalosauroidea (Marshosaurus, 100%; Torvosaurus, 84%), 53% for Allosauroidea (Sinraptor, 73%; Allosaurus, 55%; Acrocanthosaurus, 69%; Carcharodontosaurus, 47%; Giganotosaurus, 71%; Mapusaurus, 43%), 71% for Tyrannosauroidea (Guanlong, 83%; Raptorex, 71%; Alioramus, 100%; Gorgosaurus, 42%; Daspletosaurus, 58%; Albertosaurus, 54%; Tyrannosaurus, 91%), and 65% for Dromaeosauridae (Deinonychus, 75%; Dromaeosaurus, 55%). Result of this analysis classifies the tooth crown fragment MG 73 as belonging to Gorgosaurus (see Table 1) and in the plot with the distribution of the different theropods it falls in the intersection of the morphospace of several large sized taxa, including Acrocanthosaurus, Torvosaurus, and Indosuchus (Fig. 16). This specimen is a medium size (estimated CH around 25 mm), ziphodont tooth, with an oval basal section and poorly preserved mesial and distal denticles in both mesial and distal surfaces.
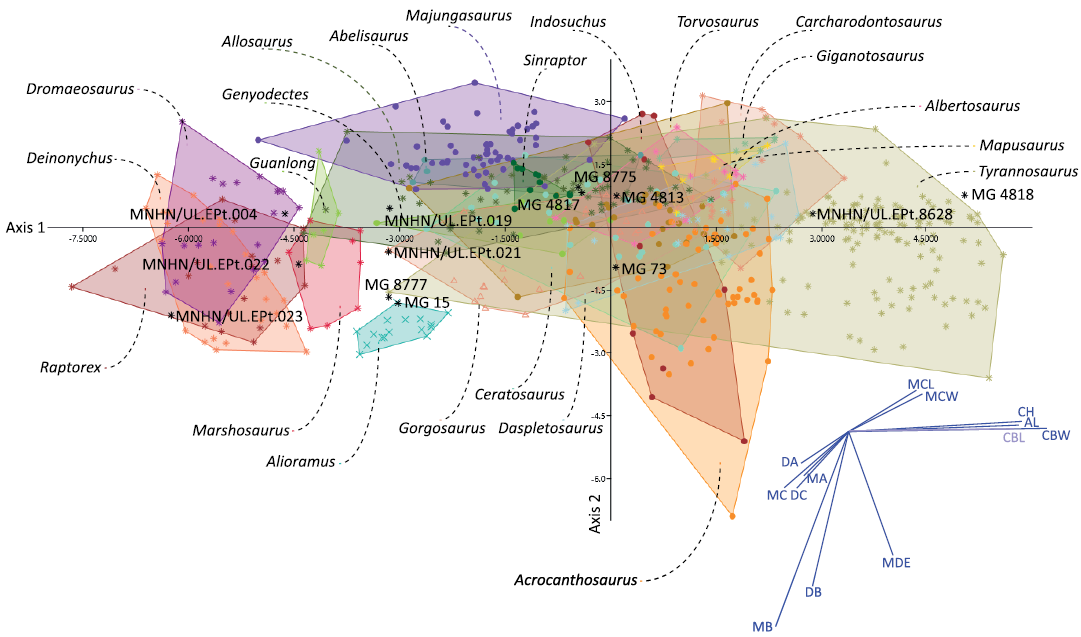
Fig. 16. Graphical results of the Linear Discriminant Analysis (LDA) of 581 tooth crowns belonging to twenty-two theropod taxa and the isolated specimens from the Lusitanian Basin studied in this work. Eigenvalue of axis 1 = 8.2989 (which accounts for 67.28% of the total variation) and eigenvalue of axis 2 = 1.129 (which accounts for 9.152% of the total variation). AL, apical length; CBW, crown base width; CH, crown height; DA, distoapical denticle density; DB, distobasal denticle density; DC, distocentral denticle density; MA, mesioapical denticle density; MB, mesiobasal denticle density; MC, mesiocentral denticle density; MCL, mid-crown length; MCW, mid-crown width; MDE, mesiobasal denticles extent.
The fossil record of continental vertebrates known from the Upper Cretaceous of the Lusitanian Basin is scarce and currently restricted to three areas in the northern sector of the basin: the region of Aveiro and Viso, in the Aveiro sub-basin, and Taveiro, in the Figueira da Foz sub-basin. Theropod dinosaurs are represented, beside the specimens first described by Sauvage (1897–1898) and Lapparent and Zbyszewski (1957), by a collection of mostly small-sized isolated teeth, some caudal vertebrae (first interpreted as belonging to pterosaurs) and few ungual phalanges interpreted as belonging to different coelurosaurian groups (Antunes and Sigogneau-Russell 1991, 1992; Galton 1994, 1996; Antunes and Mateus 2003). However, there are also some larger tooth crown fragments that have been attributed to “Megalosauridae” (e.g., Antunes and Sigogneau-Russell 1991; Galton 1996). This material is part of a collection provisionally deposited in the Centro de Estudos Geológicos of the Universidade Nova de Lisboa (Antunes and Mateus 2003), but unfortunately, we were unable to have yet access to it. Due to the poor preservation of MG 73, a more precise taxonomic identification is not possible, and it is therefore attributed here to a medium or large-sized indeterminate theropod taxon.
The ungual phalanges belong to very small individuals, but their taxonomic discussion is difficult. They have a relatively strong ventral curvature and a triangular cross-section, which together with the presence of well-developed and mostly symmetrically placed lateral grooves suggest that these ungual phalanges probably belong to a pedal digit of an indeterminate theropod. However, their attribution to a more inclusive clade is not possible mostly because the proximal end is missing in both phalanges.
Ceratosauria Marsh, 1884
Genus Ceratosaurus Marsh, 1884
Type species: Ceratosaurus nasicornis Marsh, 1884, from the late Kimmeridgian–early Tithonian Brushy Basin member of the Morrison Formation in Garden Park, Colorado, USA.
cf. Ceratosaurus sp. Marsh, 1884
Fig. 17.
Previous identifications: MG 8777 is associated with a label that says Megalosaurus pombali, Ribamar, but was described by Lapparent and Zbyszewski (1957) as coming from Atalaia and it was assigned to Megalosaurus insignis (Lapparent and Zbyszewski 1957: 22, pl. 12: 9). The tooth crown fragment MNHN/UL.Ept.021 is associated with a label with the identification M. insignis and possibly corresponds to the specimen from Porto das Barcas mentioned by Lapparent and Zbyszewski (1957: 22) as belonging to this species.
Material.—Two tooth crowns (MG 8777, Fig. 17A; MNHN/UL.EPt.021, Fig. 17B). MG 8777 was found in Atalaia and MNHN/UL.Ept.021 came from Porto das Barcas. Both localities belonging to the municipality of Lourinhã (Portugal) and the two specimens were probably collected from upper Kimmeridgian–lower Tithonian sediments of the Sobral Formation (Fig. 2) (Manuppella et al. 1999a).
Description.—MG 8777 is represented by a relatively well-preserved tooth crown and preserves a small fragment of the root (Fig. 17A). The crown is quite elongated (CHR = 2.8) and strongly labiolingually compressed (CBR = 0.5), with a lanceolate basal section. The crown is slightly recurved distally in lateral view. The mesial margin is strongly convex whereas the distal one is slightly concave, and the apex is positioned distal to the distal end of the mesial surface. The labial surface is slightly convex, whereas the lingual surface is mostly flat. Subtle transverse undulations are visible on the lingual surface. The enamel has a faint ornamentation consisting of a series of poorly marked ridges and grooves mostly apicobasally oriented (braided texture sensu Hendrickx et al. 2015a). There is a shallow longitudinal concave surface adjacent to the distal carina on the lingual surface. A short irregular surface of the enamel (spalled surface sensu Hendrickx et al. 2015a) is present in the apical end of the crown and extends slightly into the labial and lingual surfaces. The presence of these enamel flaking in theropod teeth have been interpreted as resulting from forces produced during contact between the crown and food (Schubert and Ungar 2005; Hendrickx et al. 2015a). Both the mesial and distal carinae are denticulated. They are mostly straight and centrally positioned in the mesial and distal surfaces, respectively. In the distal carina the denticles extend to the cervix, but in the mesial carina they end at about two thirds of the height of the crown from the tip. There are 19 and 18 denticles per 5 mm in the central and basal sectors of the mesial carina, respectively. In the distal carina there are 15 denticles per 5 mm in the apical and central sectors and 19 denticles per 5 mm in the denticulated basal sector (see SOM 2). The mesial denticles are subquadrangular, slightly apically inclined, and with convex distal margins. The mesial carina is poorly preserved apically so the number and morphology of the denticles in this part cannot be known but the central denticles are relatively well-preserved. The denticles become much smaller mesiodistally to the basal end of the denticulated carina. The distal denticles are rectangular, longer mesiodistally than apicobasally, with convex distal margins, and project perpendicularly to the carina. They become much narrower apicobasally to the crown base and mesiodistally shorter apically, ending slightly below the apex. Between the distal denticles, there are well-defined interdenticular sulci projecting towards the base of the crown. The interdenticular sulci are visible on both lingual and labial surfaces, being more evident in the central and basal sectors of the carina.

Fig. 17. Tooth crowns attributed to cf. Ceratosaurus sp. from the Kimmeridgian–Tithonian Sobral Formation, Atalaia and Porto das Barcas (Lourinhã region, Portugal). A. MG 8777. B. MNHN/UL.EPt.021. Labial (A1, B1), distal (A2, B2), mesial (A3, B3), and lingual (A4, B4) views; basal cross-section (A5, A6, B5); detail of the distal denticles (B6).
MNHN/UL.Ept.021 is represented by a small fragment of the basal part of a tooth crown (Fig. 17B). Although being very incomplete, the preserved fragment (preserved CH = 19.56 mm; estimated CH = 35.66 mm) indicates that this specimen belongs to a relatively large sized taxon (see SOM 2). The crown is highly compressed labiolingually (CBR = 0.53) and has a lanceolate basal cross section with a strongly convex labial surface and a flat lingual surface. The lingual surface is slightly concave longitudinally adjacent to the distal carina. The distal carina has denticles extending to the cervix, but denticles are absent in the basal sector of the mesial one carina (a small section of denticles is visible about 13 mm above the cervix). There are 12 denticles per 5 millimeters in the basal part of the distal carina. The distal denticles are rectangular, much higher mesiodistally than apicobasally and have strongly convex external margins (Fig. 17B6). They are separated by relatively large interdenticular spaces, which project into the lateral surface of the crown and delimit well-developed and obliquely oriented caudae projecting from the base of the denticles. A few short marginal undulations are visible on the lingual surface adjacent to the distal carina. As in the previously described specimen, the enamel has a subtle braided texture, which is more visible in the lingual surface adjacent to the distal carina.
Table 1. Classification of the isolated teeth based on the performed Discriminant Analysis (first column) and proposed taxonomic identification.
|
Specimen |
Classification LDA |
Taxonomic attribution |
|
MG 8777 |
Marshosaurus |
cf. Ceratosaurus sp. |
|
MNHN/UL.EPt.021 |
Genyodectes |
cf. Ceratosaurus sp. |
|
MNHN/UL.EPt.023 |
Dromaeosaurus |
Megalosauridae indet. |
|
MG 4818 |
Mapusaurus |
cf. Torvosaurus gurneyi |
|
MG 4813 |
Ceratosaurus |
cf. Torvosaurus gurneyi |
|
MNHN/UL.EPt.8628 |
Albertosaurus |
cf. Torvosaurus gurneyi |
|
MNHN/UL.EPt.004 |
Dromaeosaurus |
Allosaurus sp. |
|
MNHN/UL.EPt.019 |
Genyodectes |
Allosaurus sp. |
|
MG 8775 |
Allosaurus |
Allosaurus sp. |
|
MG 4817 |
Allosaurus |
Allosaurus sp. |
|
MG 15 |
Marshosaurus |
Allosauroidea indet. |
|
MG 73 |
Gorgosaurus |
Theropoda indet. |
Remarks.—The discriminant analysis assigned MG 8777 as belonging to Marshosaurus and MNHN/UL.Ept.021 to Genyodectes (see Table 1). In the plot, there is a large overlap of the morphospaces occupied by several taxa of medium and large-sized theropods with ziphodont dentition. Both specimens fall outside the morphospace of all defined groups but closer to those occupied by tyrannosaurids, but MG 8777 is also close to the mosphospaces of Allosaurus and Genyodectes (Fig. 16). The cladistic analysis of the dentition-based data matrix with constraints and including all studied morphotypes found 30 Most Parsimonious Trees (MPTs) (CI = 0.201, RI = 0.456, L = 1332). The strict consensus tree from this MPTs (CI = 0.194, RI = 0.432, L = 1379; Fig. 18) is relatively well-resolved, except for one branch comprising megalosaurid and spinosaurid taxa that are mostly placed in a large polytomy. In this analysis the specimens MG 8777 and MNHN/UL.Ept.021 (Morphotype 1) are placed in this polytomy as the sister taxon of Piatnitzkysaurus (Fig. 18A). The results of the analysis pruning a priory all the morphotypes from the Lusitanian Basin but Morphotype 1 found 1 MPT (CI = 0.203, RI = 0.462, L = 1320). The Megalosauroidea clade is well resolved, but the placement of Morphotype 1 is similar to that obtained in the analysis with all morphotypes, being placed within the megalosauroid clade as the sister taxon of Piatnitzkysaurus (Fig. 18B). The crowns of the two specimens (MG 8777 and MNHN/UL.Ept.021) are strongly labiolingually compressed (CBR = 0.53), a common dental feature of the lateral teeth in several theropods, including Ceratosaurus, Abelisaurus, Majungasaurus, Marshosaurus, and Torvosaurus (Hendrickx et al. 2020b). Some non-abelisauroid ceratosaurians, such as Ceratosaurus and Genyodectes, usually have blade-shaped lateral crowns with CBR lower than 0.5, but some teeth may have higher ratios (Rauhut 2004; Hendrickx et al. 2015b, 2020b). The crown of MG 8777 is strongly elongated (CHR = 2.35), which is higher than the mean ratio for most theropod lateral tooth crowns, except those of Genyodectes, Torvosaurus, Acrocanthosaurus, and Giganotosaurus (Hendrickx et al. 2015b, 2020b). Both mesial and distal carinae have a high number of small denticles with a DSDI ratio near 1.2 (in MG 8777), meaning that the number of denticles in the central sector of the mesial carina is slightly higher than the number of denticles in the central sector of the distal carina. This ratio is usually less than or equal to 1 in the lateral teeth of most theropod taxa, except for Ceratosaurus, in which in some lateral teeth this ratio is higher than 1.2 (Hendrickx et al. 2020b). As in some lateral teeth of Ceratosaurus, both MG 8777 and MNHN/UL.ePt.021 have a flattened lingual margin and a longitudinal concave surface adjacent to the distal carina in the lingual surface. The presence of a concave area adjacent to the mesial and distal carinae has been proposed as representing a neoceratosaurian synapomorphy as this feature is shared by Ceratosaurus, Genyodectes, and at least some abelisaurids (Rauhut 2004). Morphotype 1 shows some differences relative to the dentition of Ceratosaurus, including the extension of the mesial carina, which usually reaches the cervix or very close to it (Hendrickx et al. 2015b; Malafaia et al. 2017a). However, in some ceratosaurian taxa, such as Genyodectes, similarly to the condition in both MG 8777 and MNHN/UL.EPt.021, the mesial carina of at least some lateral tooth crowns is restricted to the apical half (Rauhut 2004; Christophe Hendrickx, personal communication 2023). Despite these differences and the results of the cladistic analyses, the specimens grouped in Morphotype 1 are here tentatively assigned to Ceratosaurus based on the presence of a concave surface adjacent to the distal carina (a possible synapomorphy for Ceratosauria), the symmetrical shape of the crow (with both mesial and distal carinae positioned centrally), the strongly labiolingual compression of the crown, and the relatively high number of denticles.

Fig. 18. Strict consensus trees resulting from a cladistic analysis performed on a dentition-based data matrix and forcing the constrains defined by Hendrickx et al. (2020a) with the five tooth morphotypes from the Lusitanian Basin (A), simplified strict consensus tree resulting from the cladistic analysis pruning a priori all morphotypes from the Lusitanian Basin but Morphotype 1 (B), and simplified strict consensus tree resulting from the analysis pruning a priori all morphotypes from the Lusitanian Basin but MNHN/UL.EPt.023 (C).
Tetanurae Gauthier, 1986
Megalosauroidea (Fitzinger, 1843) sensu Benson 2010a
Megalosauridae (Fitzinger, 1843) sensu Benson 2010a
Megalosauridae indet.
Fig. 19.
Previous identifications: The tooth crown (MNHN/UL.EPt.023) was attributed to Megalosaurus insignis by Lapparent and Zbyszewski (1957: 22).
Material.—One small tooth crown (MNHN/UL.EPt.023; Fig. 19) from Porto Dinheiro (Lourinhã, Portugal) and was possibly collected from upper Kimmeridgian–lowermost Tithonian levels of the Praia da Amoreira-Porto Novo Formation (Fig. 2) (Manuppella et al. 1999a).
Description.—MNHN/UL.EPt.023is represented by a relatively well-preserved small crown (CH = 10.24 mm) of a lateral tooth, which preserves a fragment of the root. The crown is relatively low (CHR = 1.85) and highly compressed labiolingually (CBR = 0.54). It is triangular in lateral view and slightly recurved distally, with a convex mesial margin, a slightly concave distal margin, and an apex positioned beyond the level of the distal margin. The labial surface is convex, the lingual surface is flat, and the basal section is lanceolate shaped. Both mesial and distal margins are denticulated, straight and placed in the distal and mesial margins, respectively. The carinae have several small denticles that extend to the cervix in the distal carina, but they end at about the mid-height of the crown in the mesial carina. There are 22, 23, and 25 denticles per 5 mm in the apical, central, and basal sectors of the distal carina, respectively. The mesial carina is broken in the central and basal portions so only the apical denticles can be described. There are 28 denticles per 5 mm in the apical part of the mesial carina (see SOM 2). The distal denticles are subquadrangular in the apical part of the distal carina (Fig. 19A1) but become rectangular (slightly longer mesiodistally than the apicobasal wide) to the crown base (Fig. 19A7). They have convex external margins and are placed perpendicular to the carina. The mesial denticles are quadrangular and perpendicularly to the carina. Both mesial and distal denticles extend to the apex, forming a continuous serrated margin. The distal denticles become smaller to the base and the apex of the crown. A small flat surface adjacent to the distal carina is visible in the lingual surface. The enamel has an ornamentation formed by subtle ridges and crests oriented vertically (braided texture sensu Hendrickx et al. 2015a).
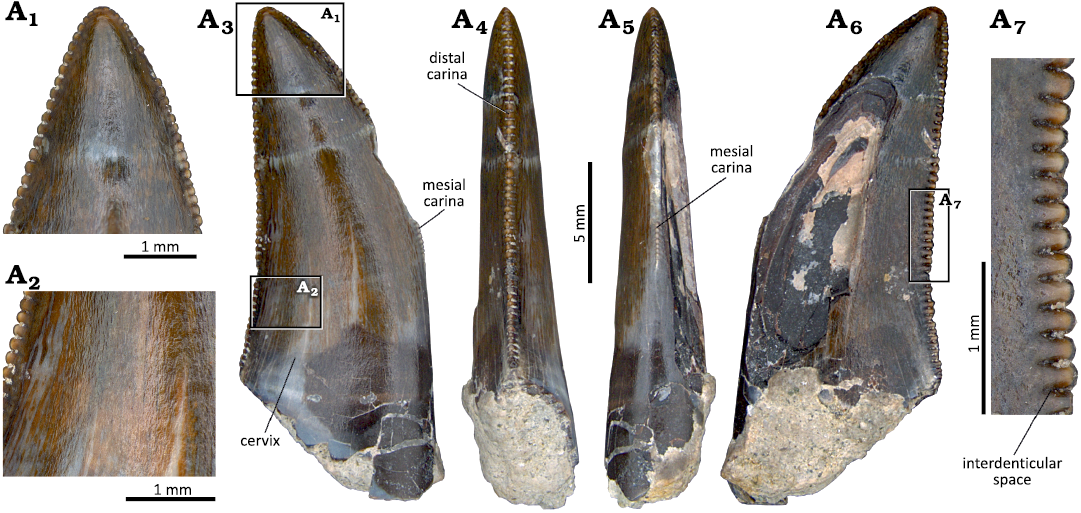
Fig. 19. Tooth crown (MNHN/UL.EPt.023) attributed to an indeterminate megalosauroid theropod from the upper Kimmeridgian–lowermost Tithonian Praia da Amoreira-Porto Novo Formation, Porto Dinheiro (Lourinhã region, Portugal) in labial (A3), distal (A4), mesial (A5), and lingual (A6) views; detail of the mesial and distal denticles in the apical part of the crown (A1), detail of the enamel ornamentation (A2), detail of the distal denticles in the central sector of the carina (A7).
Remarks.—The results of the discriminant analysis classify MNHN/UL.EPt.023 as belonging to Dromaeosaurus (see Table 1). In the plot obtained by the LDA analysis it falls at the limit of the morphospace occupied by Raptorex, which has a great overlap with those of Dromaeosaurus and Deinonychus (Fig. 16). This result may be related to the small size of the specimen and the high density of denticles in both carinae. However, in several dromaeosaurid taxa, the lateral teeth lack serrated carinae or, if present, the mesial carina has a characteristic twist, as occur in Dromaeosaurus (Currie et al. 1990; Currie 1995; Hendrickx et al. 2015b). In addition, the lateral teeth of most dromaeosaurids have a wide apicobasally elongated concavity near the base of the labial surface (Hendrickx et al. 2015b and references herein). The cladistic analysis of the dentition-based data matrix with constraints and including all studied morphotypes recovered the tooth crown (MNHN/UL.EPt.023) in a polytomy within a poorly-resolved Megalosauroidea clade (Fig. 18A). The results of the analysis pruning a priori all the morphotypes from the Lusitanian Basin but MNHN/UL.EPt.023 found 7 MPTs (CI = 0.204, RI = 0.465, L = 1315). The consensus tree from these MPTs (CI = 0.203, RI = 0.462, L = 1320) recovered two clades within Megalosauroidea, a well-resolved Spinosauridae and a second group with megalosauridae taxa mostly placed in polytomy, also including MNHN/UL.EPt.023 (Fig. 18C). This specimen has a combination of features shared with lateral teeth of megalosaurids, including: (i) the braided and oriented texture of the enamel; (ii) the morphology of the distal denticles, which are subquadrangular to subrectangular, with short to well-developed interdenticular sulci and symmetrically convex external margins; (iii) the centrally positioned mesial and distal carinae on their mesial and distal surfaces, respectively; and (iv) a mesial carina not reaching the cervix. However, this specimen has a much higher denticles density in both mesial and distal carinae than is typical of megalosaurids. A higher number of denticles in the lateral teeth of juvenile individuals relative to that present in adult forms has been identified in some deeply nested theropod taxa (e.g., Mapusaurus, Canale et al. 2014), but the opposite pattern has been described for Coelophysis and Tyrannosaurus (Long and McNamara 1997; Buckley and Currie 2014). Based on this combination of features and the results of the cladistic analysis, MNHN/UL.EPt.023 is interpreted here as a lateral tooth of a juvenile megalosaurid theropod and the relatively high density of denticles is proposed as been related with ontogeny. Moreover, based on its paleogeographic and stratigraphic distribution it may be related to Torvosaurus gurneyi.
Genus Torvosaurus Galton & Jensen, 1979
Type species: Torvosaurus tanneri Galton & Jensen, 1979, from the late Kimmeridgian–early Tithonian Brushy Basin member of the Morrison Formation at Colorado, USA.
cf. Torvosaurus gurneyi Hendrickx & Mateus, 2014
Fig. 20.
Previous identifications: MNHN/UL.EPt.8628 was described and figured by Lapparent and Zbyszewski (1957: 25, pl. 12: 17) and possibly corresponds to one of the first dinosaur remains identified in Portugal, which was mentioned by Carlos Ribeiro in 1863 (Mateus 2005). This specimen is part of the material used to describe the putative new species Megalosaurus pombali. The fragment of tooth crown (MG 4813) was mentioned by Lapparent and Zbyszewski (1957) and assigned to Megalosaurus insignis (Lapparent and Zbyszewski 1957: 22). MG 4818 is labeled as Megalosaurus pombali, but we have not found any reference to this tooth fragment in previous publications.
Material.—Two tooth crown fragments (MG 4813, Fig. 20B; MG 4818, Fig. 20C) and a large mesial tooth (MNHN/UL.EPt.8628, Fig. 20A). MG 4813 was found in Montoito (Lourinhã, Portugal), in Tithonian deposits of the Bombarral Formation (Manuppella et al. 1999a). MG 4818 comes from Praia de S. Bernardino (Peniche, Portugal) from Kimmeridgian levels of the Alcobaça Formation (Fürsich et al. 2021). MNHN/UL.EPt.8628 came from Porto Dinheiro (Lourinhã, Portugal) and was collected in upper Kimmeridgian–lowermost Tithonian levels of the Praia da Amoreira-Porto Novo Formation (Manuppella et al. 1999a). See Fig. 2 for the location of these specimens.
Description.—MNHN/UL.8628 corresponds to an almost complete tooth crown, without the apex, but with a fragment of the root preserved (Fig. 20A). It corresponds to a large lateral tooth (the preserved fragment is 75 mm in high). The crown is slightly compressed labiolingually (CBR = 0.59), with a lanceolate basal section. Both lingual and labial surfaces are convex. In lateral view, the mesial surface is also convex, but the distal margin is mostly straight to slightly concave. The mesial and distal carinae are denticulated, but the mesial one is mostly worn, so is not possible to describe the morphology of the denticles. However, it is possible to verify that the mesial carina ends well above the base of the crown while the distal carina extends to the cervix. The distal carina is placed in the distal surface and is straight along the entire preserved length. There are 9 and 14 denticles per 5 mm in the central and basal sectors of the distal carina respectively (see SOM 2). The denticles are rectangular (larger mesiodistally than apicobasally) and project perpendicularly to the carina. They have slightly convex external margins and become smaller to the base of the crown. The interdenticular space is relatively wide, but there are not interdenticular sulci between the denticles at least in the preserved part of the crown. Subtle transverse and marginal undulations are present in both labial and lingual surfaces. The enamel has a very weak irregular ornamentation (but it is possible that the surface is somewhat worn out).
MG 4813 and 4818 correspond to fragments of the basal part of two relatively large tooth crowns, with estimated CH around 80 mm (Fig. 20B, C). Despite fragmentary, these specimens show morphology similar to the previously described tooth, including the shape of the basal cross-section, the size of the preserved fragment of the crown base (see SOM 2), the morphology and number of denticles, as well as the shape and relative extension of the mesial and distal carinae. In MG 4818 the distal carina is straight and placed in the distal surface, but the mesial one seems to project slightly into the lingual surface to the base of the crown. The mesial denticles are quadrangular, with slightly convex distal margins. The denticles are separated by narrow interdenticular spaces and become smaller to the base of the crown. The distal denticles are subquadrangular with slightly convex external margins and are separated by broad interdenticular spaces in MG 4813. On the other hand, the distal denticles of MG 4818 are rectangular and much longer mesiodistally than apicobasally. A clear demarcation between the operculum and the radix is present in all preserved denticles of both mesial and distal carinae. Transverse and marginal undulations are absent in the preserved fragment of the crown in both specimens, but MG 4818 has subtle oblique interdenticular sulci adjacent to some distal denticles on the lingual surface. The enamel has irregular, very subtle texture on both lingual and labial surfaces.

Fig. 20. Tooth crowns attributed to the megalosauroid theropod cf. Torvosaurus gurneyi Hendrickx & Mateus, 2014 from different Kimmeridgian to Tithonian localities of Portugal. A. MNHN/UL.EPt.8628, Porto Dinheiro (Lourinhã region). B. MG 4813, Montoito (Lourinhã region). C. MG 4818, Praia de S. Bernardino (Peniche region). Labial (A1, B1, C1), lingual (A2, B2, C2), distal (A5, B4, C4), and mesial views (A6, B3, C3); detail of the distal denticles in the central sector of the carina (A3); basal cross-section (A4, B5, C6, C7); cross-section at mid-height of the crown (C5).
Remarks.—MG 4818 and 4813 are incomplete and distorted, but the size of the preserved fragment suggests that these specimens likely belong to a very large theropod taxon. The size of these specimens (with CH > 75 mm) is only comparable to the crowns of Torvosaurus among the Late Jurassic theropod taxa currently known in the Portuguese fossil record. The number and morphology of the denticles as well as the extension of the carinae are also similar to those described for the lateral and mesial teeth of Torvosaurus (Hendrickx et al. 2015b, 2020b; Malafaia et al. 2017a). The results of the discriminant analysis classify MG 4818 as belonging to Mapusaurus, MG 4813 to Ceratosaurus, and MNHN/UL.EPt.8628 to Albertosaurus (see Table 1). The plot obtained in the LDA shows MG 4818 and MNHN/UL.Ept.8628 within or close to the morphospace of Tyrannosaurus, while MG 4813 is in the intersection of the morphospace of different large theropods, including abelisaurids, Torvosaurus, carcharodontosaurids, and Tyrannosaurus (Fig. 16). These results can be explained by the large size of the specimens and the high percentage of missing variables, particularly those related to denticles density. The cladistic analysis of the dentition-based data matrix with constraints and including all studied morphotypes recovered the specimens here grouped in Morphotype 2 in a polytomy within a poorly-resolved Megalosauroidea clade (Fig. 18A). The results of the analysis pruning a priori all the morphotypes from the Lusitanian Basin but Morphotype 3 found 3 MPTs (CI = 0.203, RI = 0.463, L = 1318). The consensus tree from these MPTs (CI = 0.198, RI = 0.444, L = 1355) is similar to that obtained in the previous analysis also recovering Morphotype 2 within a large polytomy with several megalosauridae taxa (Fig. 21A). MNHN/UL.Ept.8628 possibly corresponds to a mesial tooth crown based on its elongation, relatively low labiolingual compression (CBR = 0.6) and the presence of mesial carina restricted to the apical part of the crown. Only the number of denticles on the central and basal sectors of the distal carina of MNHN/UL.Ept.8628 can be determined, and it is similar to the average number of distal denticles on Torvosaurus, also sharing with this taxon similar denticle shape and the centrally positioned carinae (see Hendrickx et al. 2020b). Furthermore, despite the absence of the crown apex, the preserved part indicates that the specimen would belong to a particularly large theropod taxon (CH > 75 mm). Except for Torvosaurus, no other taxon currently known in the Upper Jurassic fossil record of the Lusitanian Basin has tooth crowns of this height. Based on this combination of features, Morphotype 2 is here referred to Torvosaurus, which is also consistent with the results of the cladistic analyses. Moreover, based on the paleogeographic and stratigraphic distribution, these specimens most likely belong to the Portuguese species Torvosaurus gurneyi Hendrickx & Mateus, 2014.
Avetheropoda Paul, 1988
Avetheropoda indet.
Fig. 22.
Previous identifications: MG 4822 is labeled as belonging to Megalosaurus, but we have not found reference to this specimen in previous publications.
Material.—A small dorsal vertebra (MG 4822; Fig. 22) labeled as coming from Paimogo, in the municipality of Lourinhã (Portugal) and probably comes from the Sobral Formation (Fig. 2), which is Kimmeridgian–Tithonian in age (Manuppella et al. 1999a).
Description.—The specimen is represented by the well-preserved centrum, but the neural arch is missing (Fig. 22). It corresponds to a very small specimen, with 31 mm in length (see SOM 2). The centrum is spool-shaped in lateral view with strongly projected articular facets and is slightly elongated, being a little longer than high. There is a shallow depression in the lateral surface near the dorsal margin of the centrum. The neurocentral suture is open and extends along all the dorsal length of the centrum, slightly projecting into the lateral margin near the articular facets. It is strongly rough with a series of transverse grooves and ridges. The articular facets of the centrum are subcircular and slightly concave. The ventral surface is rounded, without any groove or ridge and strongly concave in lateral view. The neural canal is narrow, but slightly expanded near the articular facets.
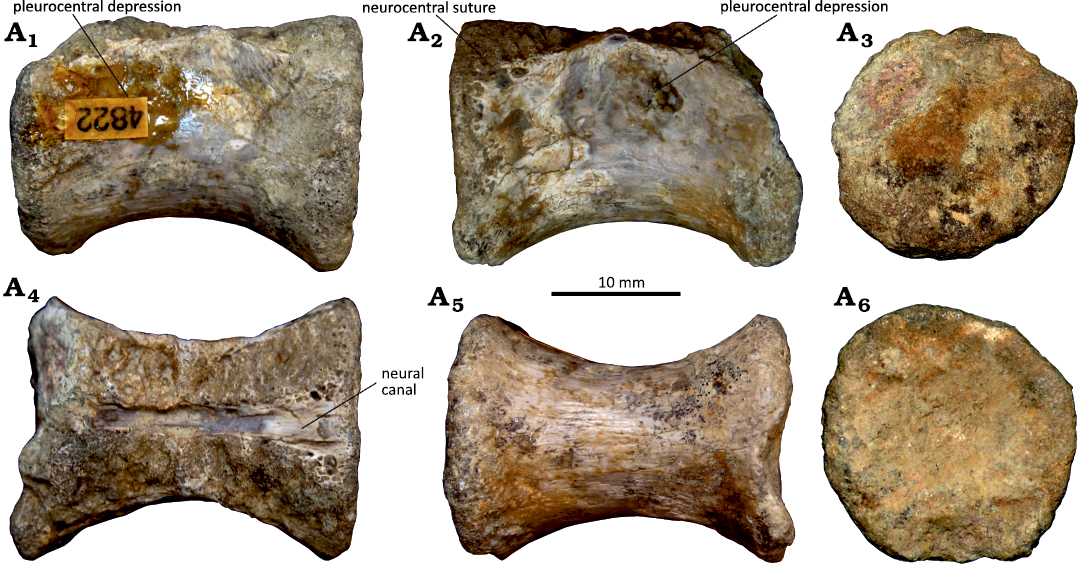
Fig. 22. Small dorsal vertebra of a juvenile indeterminate avetheropod (MG 4822) from the Kimmeridgian–Tithonian Sobral Formation, Paimogo (Lourinhã region, Portugal) in ?right lateral (A1), ?left lateral (A2), ?posterior (A3), dorsal (?anterior to the right, A4), ventral (?anterior to the left, A5), and ?anterior (A6) views.
Remarks.—The small vertebral centrum (MG 4822) is relatively elongated (length to height ratio is approximately 1.6) and is separated from the neural arch by an open neurocentral suture, suggesting that it belongs to a juvenile individual (Irmis 2007; Verrière et al. 2022). No evidence of parapophyses or ventral keel is visible in the centrum, indicating that this specimen likely corresponds to a vertebra from the middle or posterior part of the dorsal series. The centrum is spool-shaped, being strongly constricted dorsally by two well-developed pleurocentral depressions lacking pleurocoel. A strong constriction of the centrum is a feature first used to describe Allosaurus fragilis (Marsh 1877b). However, similar well-developed pleurocentral depressions are also present in the dorsal vertebrae of several other allosauroids, including Asfaltovenator, Metriacanthosaurus, Sinraptor, Concavenator, and Neovenator (Currie and Zhao 1993; Brusatte et al. 2008; Carrano et al. 2012; Cuesta et al. 2019; Rauhut and Pol 2019). Moreover, well-developed depressions are also present in some coelurosaurian theropods (Carpenter et al. 2005). Based on this combination of features and the fragmentary nature of the specimen, we assign this dorsal vertebra to an indeterminate avetheropod.
Allosauroidea (Marsh, 1878) sensu Sereno 1998
Allosauroidea indet.
Figs. 23, 24, SOM 5: fig. S3.
Previous identifications: MG 15 and another tooth crown fragment (currently lost) from Cabo Espichel were described by Sauvage (1897–1898: pl. 5: 4, 5) and assigned to Megalosaurus aff. superbus. Later, Lapparent and Zbyszewski (1957: pl. 12: 7) described the lost specimen and identified it as belonging to M. superbus. The sacral and anterior caudal vertebrae (MG 4824) were described by Lapparent and Zbyszewski (1957) and were assigned to Megalosaurus insignis (Lapparent and Zbyszewski 1957: 22–23, pl. 15: 42, pl. 30: 123). The almost complete posterior caudal vertebrae (MG 4831a) are part of the material described by Sauvage (1897–1898) as belonging to the putative new sauropod species Morosaurus marchei (Sauvage 1897–1898: 29, pl. 4: 6–8). Later, Lapparent and Zbyszewski (1957) reinterpreted this vertebra as belonging to M. insignis (Lapparent and Zbyszewski 1957: 23, pl. 13: 28). The other fragments of caudal centra (MG 4813b and MG 4828) were also attributed to M. insignis by Lapparent and Zbyszewski (1957: 23).
Material.—An incomplete tooth crown (MG 15; Fig. 23), a sacral and an anterior caudal vertebrae (MG 4824a, b; Fig. 24A, SOM 5: fig. S3A), a fragment of a posterior caudal centrum (MG 4828; SOM 5: fig. S3B), and two posterior caudal vertebrae (MG 4831a, b; Fig. 24B). MG 15 came from the locality of Boca do Chapim (Cabo Espichel, Sesimbra, Portugal), where a sedimentary sequence that corresponds to the middle Hauterivian to the middle Barremian interval is exposed (Fig. 6). These levels correspond mainly to marly sandstone deposits first interpreted by Sauvage (1897–1898) and Choffat (1904) as the Almargem Beds, considered to be Aptian to early Albian in age. Later, Rey (1972) proposed the name Papo Seco Formation for this sequence and showed that they instead correspond to lower Barremian deposits (Rey et al. 2003; Buffetaut 2007; Dinis et al. 2008). MG 4824, 4828, 4831 came from near the village of Ourém (Portugal) and were probably collected in Kimmeridgian sediments of the Alcobaça Formation (Fig. 4) (Fürsich et al. 2021).
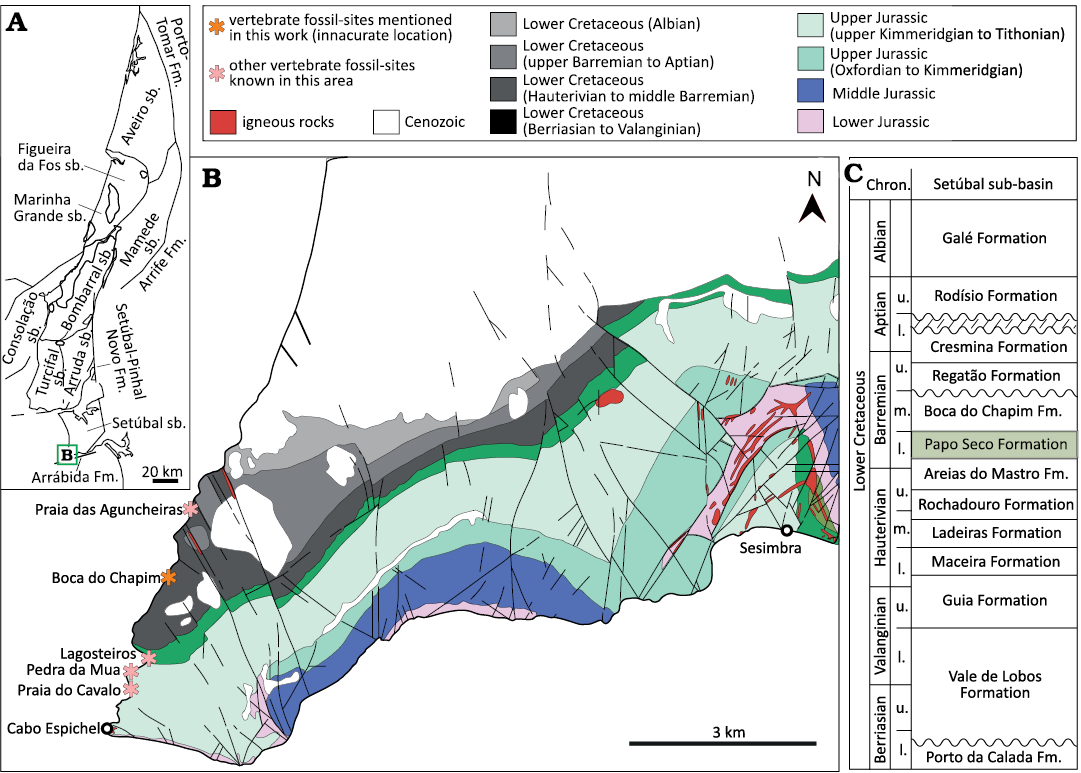
Fig. 6. Geological and stratigraphical context of fossil sites in the Setúbal sub-basin. A. Map of the Lusitanian Basin with the main structural elements and the different sub-basins (sensu Fürsich et al. 2021). B. Geological map of the Sesimbra to Cabo Espichel region with localities yielding fossil remains studied in this work (modified from Manuppella et al. 1999b). C. Simplified stratigraphy of the Lower Cretaceous sequences in the Setúbal sub-basin based on Manuppella et al. (1999b). The stratigraphic position of the Papo Seco Formation from which the studied fossils were collected is marked in green. Abbreviations: Chron., Chronostratigraphy; Fm., Formation; mb., member; l., lower; m., middle; sb., sub-basin; u., upper.
Description.—MG 15 is represented by a small relatively well-preserved tooth crown (Fig. 23). It is apicobasally short (CHR = 1.55) and strongly labiolingually compressed (CBR = 0.5), with a lenticular basal section. The crown is slightly recurved distally in lateral view, with the mesial margin strongly convex and the distal margin weakly concave. Both labial and lingual surfaces are slightly convex. Both mesial and distal carinae are denticulated, straight, and centrally positioned, being placed on the mesial and distal surfaces, respectively. The denticles extend to the cervix or close to it in both mesial and distal carinae. There are 15 denticles per 5 mm in the central portion of the distal carina (see SOM 2). The mesial carina is poorly preserved so the number and morphology of the denticles cannot be described. The distal denticles are quadrangular at mid-crown in the central sector of the carina and become rectangular (slightly longer mesiodistally than apicobasally) towards the base of the crown. They have slightly convex external margins and are separated by narrow interdenticular space. The mesial carina is incomplete, with only some denticles of the apical end of the crown being relatively well preserved. These denticles are short, subquadrangular in outline, with slightly convex external margins and extending to the apex. Interdenticular sulci are absent as well as marginal or transverse undulations. A few mesiodistally oriented transverse bands of different colors are present across the lingual or labial surfaces. The enamel has a well-marked ornamentation, consisting of a series of ridges and grooves that are mostly apicobasally oriented (braided texture sensu Hendrickx et al. 2015a).
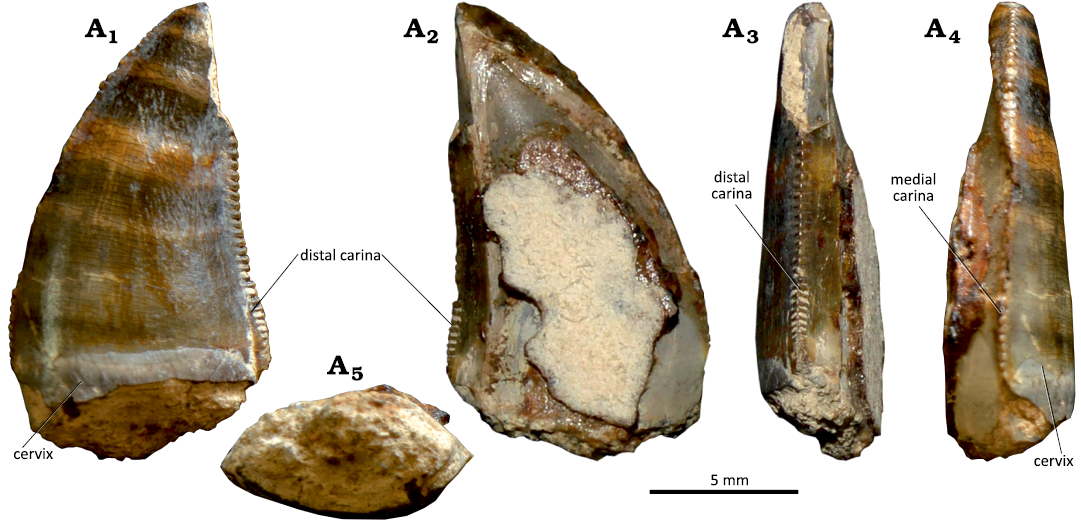
Fig. 23. Tooth crown fragment (MG 15) attributed to an indeterminate allosauroid theropod from the Lower Cretaceous of Cabo Espichel (Setúbal region, Portugal), in labial or lingual (A1, A2), distal (A3), and mesial (A4) views, and basal cross-section (A5).
The sacral vertebra (MG 4824a) is represented by a relatively well-preserved centrum, but the neural arch is completely lost (Fig. 24A). The centrum is subquadrangular in lateral view, being almost has high as long. It is spool-shaped with strongly expanded articular facets and a well-marked pleurocentral depression on the dorsolateral surface. Only the posterior articular facet of the centrum is relatively well-preserved. This facet is flat, with a subcircular outline, slightly higher dorsoventrally than the mediolateral width (see SOM 2). The ventral surface is rounded without any visible groove or ridge. The neural arch is broken at the level of the pedicles, exposing the neural canal, which is relatively wide. In the posterior end of the dorsal surface of the centrum there is a large intervertebral foramen opening laterally from the neural canal. The anterior caudal vertebra (MG 4824b) is represented by a somewhat distorted and fractured centrum, which is slightly longer than it is high (SOM 5: fig. S2A1–A5). In lateral view, the centrum is spool-shaped, with strongly expanded articular facets, and strongly concave. The articular facets are poorly preserved, so their morphology is mostly unknown. The most complete facet is slightly concave and seems subcircular in outline. The ventral surface is concave in lateral view and has a well-marked longitudinal ridge, which probably extends along the entire length of the centrum, as it is visible in all the preserved part almost reaching the articular facets (SOM 5: fig. S2A3).
The posterior caudal vertebra (MG 4831a) preserves the centrum and part of the neural arch (Fig. 24B). The centrum is strongly elongated, being more than twice as long as its dorsoventral height. In lateral view, it is somewhat concave ventrally and the articular facets are slightly expanded. This vertebra is from the posterior part of the tail based on the absence of transverse processes, although there is a reminiscent small longitudinal crest that is delimited ventrally by a well-marked groove. The articular facets of the centrum are strongly concave and circular in outline. The neural arch is incomplete and only preserves a fragment of the base of the prezygapophyses that project anteriorly almost parallel to the dorsal margin of the centrum. The neural arch is slightly shorter anteroposteriorly than the centrum. A small fragment of the neural spine is preserved between the prezygapophyses. The lateral surface of the neural arch is slightly concave and there is a pair of longitudinal crests extending along the dorsolateral margin, connecting the base of the pre- and postzygapophyses. In dorsal view the neural arch is narrower than the centrum but expands slightly to the anterior end. The ventral surface of the centrum is mostly rounded and has a short but relatively well-marked longitudinal groove near the mid-length. Facets for the haemal arches are present adjacent to the ventral margin of the articular facets of the centrum, with the anterior facet being much more marked than the posterior one. The anterior facet for the haemal arch is mediolaterally elongated and has a shallowly concave anterior surface that is crossed by a low transverse crest near the mid-length. The other fragments of posterior caudal vertebrae (MG 4831 and MG 4828) are represented by the anterior and posterior parts of the centra (SOM 5: fig. S2B1–B4). The morphology of the preserved fragments is similar to that of the previously described vertebra, except for the facet for the haemal arches, which is slightly less developed. The centra have V-shaped cross-section and are strongly constricted ventrally. In the preserved fragment of MG 4828 the ventral margin seems to have a low crest adjacent to the articular facet. The articular facets are concave and oval (slightly higher dorsoventrally than the mediolateral width).
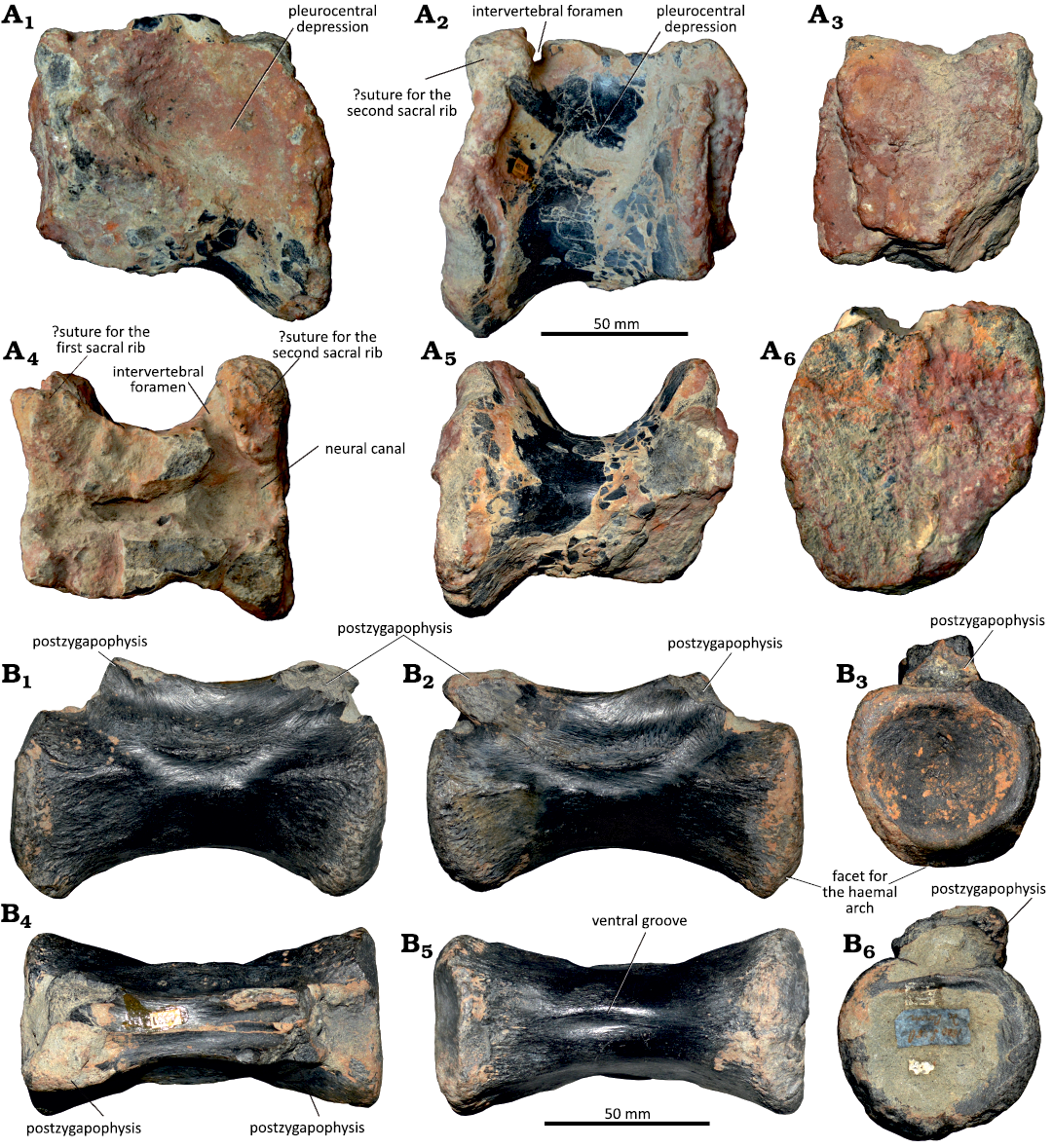
Fig. 24. Sacral and caudal vertebrae attributed to indeterminate allosauroid theropods from Kimmeridgian Alcobaça Formation, Ourém region (Portugal). A. Sacral vertebra, MG 4824a. B. Posterior caudal vertebra, MG 4831a. Left lateral (A1, B2), right lateral (A2, B1), anterior (A3, B6), dorsal (anterior to the left, A4, B4), ventral (anterior to the right, A5, B5), and posterior (A6, B3) views.
Remarks.—Result of the discriminant analysis classifies MG 15 as belonging to Marshosaurus (see Table 1). The plot with the distribution of the different theropod taxa shows that MG 15 falls close to the polygon occupied by Alioramus (Fig. 16). The cladistic analysis of the dentition-based data matrix with constraints and including all studied morphotypes recovered the tooth crown (MG 15) in polytomy with Allosaurus within a relatively well-resolved Allosauroidea clade (Fig. 18A). The results of the analysis pruning a priory all the morphotypes from the Lusitanian Basin but MG 15 found 3 MPTs (CI = 0.204, RI = 0.464, L = 1316). The position of MG 15 in the consensus tree from these MPTs (CI = 0.203, RI = 0.463, L = 1319) is similar to that obtained in the previous analysis (Fig. 21B). The tooth crown is strongly compressed labiolingually, with a triangular shape in lateral view and is slightly recurved distally. These features together with the absence of flutes in the labial and lingual surfaces allow to distinguish MG 15 from the teeth of spinosaurids, which are currently the only theropod known in the Lower Cretaceous fossil record of Cabo Espichel based on dental and non-dental material (Buffetaut 2007; Mateus et al. 2011; Figueiredo et al. 2015; Malafaia et al. 2020a; Mateus and Estraviz-López 2022). The specimen can also be distinguished from dromaeosaurids and other coelurosaurian theropods by the non-constricted crown with straight denticulated mesial and distal carinae that extend to the cervix and are centrally positioned on the mesial and distal margins, respectively (e.g., Zinke 1998; Larson 2008; Hendrickx et al. 2015b). MG 15 has a lanceolate basal cross-section, whereas the lateral teeth of most dromaeosaurids and tyrannosaurids tend to have wide apicobasally elongated concavities on the base of the labial surface (or in both labial and lingual surfaces), so that basal cross section of the crowns has a characteristic reniform or figure-of-eight outline (Brusatte et al. 2009; Gianechini et al. 2011; Hendrickx et al. 2015b). The possibility that this tooth belongs to an abelisauroid taxon can also be discarded based on the strongly concave distal surface, the symmetrically convex external margin of the denticles (instead the hooked or asymmetrically convex denticles present in some abelisaurid taxa), and the braided texture of the enamel (e.g., Hendrickx et al. 2015b, 2020b; Canale et al. 2016). The specimen from Cabo Espichel has a combination of dental features compatible with the morphology of the lateral dentition of allosauroids, sharing the morphology and number of denticles, the slightly recurved profile and the strong labiolingual compression of the crown (e.g., Hendrickx et al. 2015b and references herein). It resembles the maxillary teeth of the lectotype of Erectopus superbus from the Albian of eastern France based on the symmetrically oval shape of the basal cross-section and the extension of the mesial carina along the entire height of the crown (Allain 2005). In Neovenator salerii from the Barremian of southern England, the mesial carina of the lateral teeth occupies only about one third of the crown height (as seems to be also the case in Concavenator corcovatus from the Barremian of Cuenca, in Spain) and the basal cross-section is asymmetrical (Brusatte et al. 2008; Cuesta et al. 2018). However, the specimen from the Lusitanian Basin differs from the maxillary teeth of E. superbus by the weak elongation of the crown.
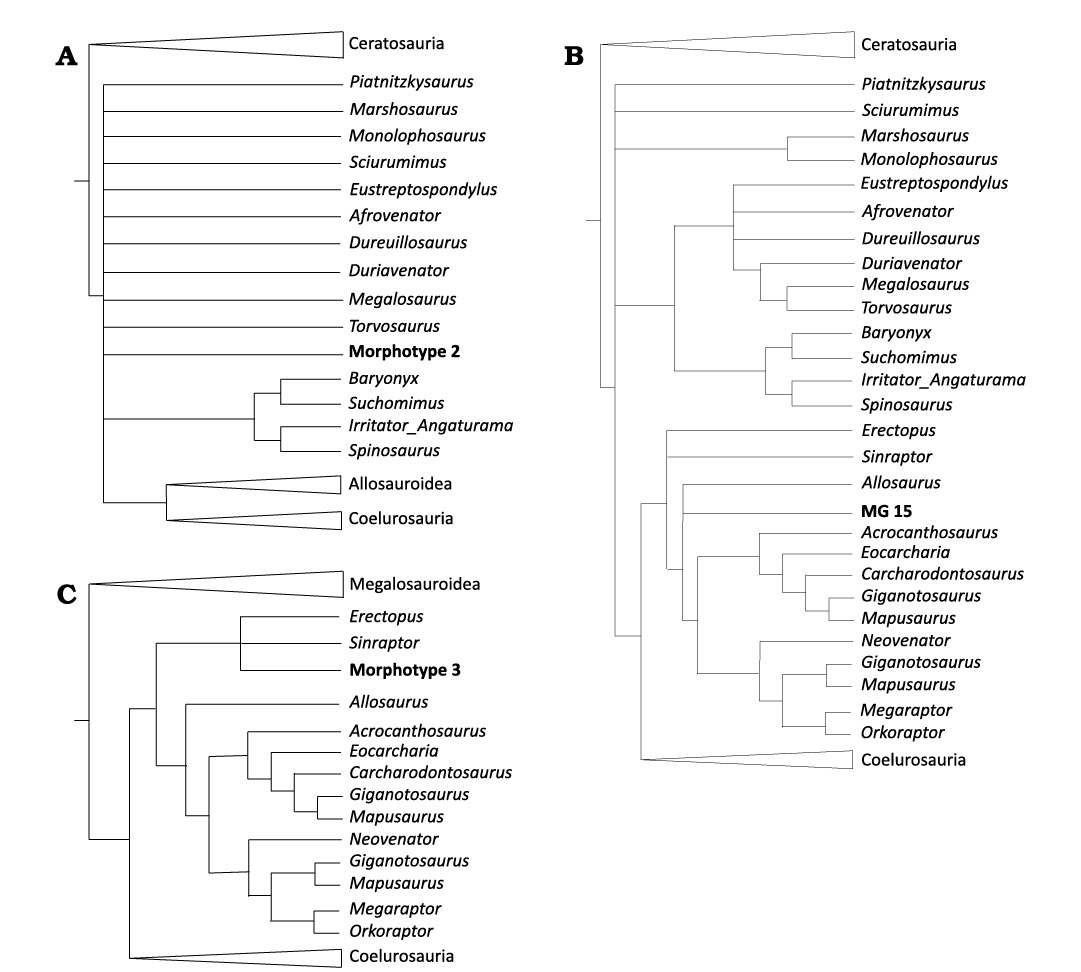
Fig. 21. Simplified strict consensus trees resulting from a cladistic analysis performed on a dentition-based data matrix and forcing the constrains defined by Hendrickx et al. (2020a) pruning a priori all morphotypes from the Lusitanian Basin but Morphotype 2 (A), simplified strict consensus tree resulting from the analysis pruning a priori all morphotypes from the Lusitanian Basin but MG 15 (B), and simplified strict consensus tree resulting from the analysis pruning a priori all morphotypes from the Lusitanian Basin but Morphotype 3 (C).
Based on this combination of features and the results of the cladistic analysis, MG 15 is here interpreted as belonging to an allosauroid theropod. The Lower Cretaceous of Cabo Espichel is well known for the important ichnological record of dinosaurs and other vertebrates found in different Upper Jurassic and Lower Cretaceous localities (e.g., Antunes 1976; Carvalho and Santos 1993; Dantas et al. 1994; Lockley et al. 1994; Santos 2003, 2008; Figueiredo et al. 2021). Osteological remains are scarcer, but some significant material attributed to different dinosaurian groups have been found (e.g., Sauvage 1897–1898; Lapparent and Zbyszewski 1957; Buffetaut 2007; Mateus et al. 2011; Figueiredo et al. 2015, 2017; Malafaia et al. 2020a; Mateus and Estraviz-López 2022). Apart from this specimen and another (currently lost) ziphodont tooth described by Sauvage (1897–1898) and Lapparent and Zbyszewski (1957), the osteological fossil record of theropods from this region consists mostly of some cranial fragments, several isolated teeth, and postcranial material of spinosaurids (Buffetaut 2007; Mateus et al. 2011; Figueiredo et al. 2015; Malafaia et al. 2020a; Mateus and Estraviz-López 2022). Likewise, some isolated and incomplete osteological fragments attributed to indeterminate theropods, dromaeosaurids, and avian theropods have been described from these levels (Figueiredo et al. 2017, 2018). If the allosauroid affinity of the tooth fragment (MG 15) is correct, it would represent the first record for the presence of this clade in the Lower Cretaceous of the Lusitanian Basin.
MG 4824a is interpreted as possibly representing the first sacral vertebra, as the centrum has a small surface for articulation with the sacral ribs dorsally at the level of the neural arch. In more posterior sacral vertebrae, these articulations extend ventrally into the lateral surface of the centra (e.g., Madsen 1976a; Benson 2010a). The centrum seems to be unfused to other sacral vertebrae, which may also support the identification as a dorsosacral element or may indicate that it corresponds to an immature individual (Rauhut 2005; Brusatte et al. 2008). The sacral vertebrae are fused in Neovenator and Concavenator, but are unfused in Acrocanthosaurus and Sinraptor, while fusion is variable in Allosaurus (Madsen 1976a; Currie and Zhao 1993; Harris 1998; Brusatte et al. 2008; Cuesta et al. 2019). The centrum has a well-developed pleurocentral depression with no pleurocoels, as in the sacral vertebrae of several allosauroid theropods, such as Allosaurus, Neovenator, Sinraptor, and Tyrannotitan (Gilmore 1920; Madsen 1976a; Currie and Zhao 1993; Novas et al. 2005; Brusatte et al. 2008).
MG 4824b is interpreted as an anterior caudal vertebra due to the relatively elongated centrum, which is slightly compressed laterally (V-shaped in cross-section) and has moderately concave lateral surfaces. The presence of a longitudinal crest in the ventral surface of anterior caudal vertebrae is a feature unknown in most early-branching tetanurans and neoceratosaurs, which instead usually have distinct longitudinal grooves (Britt 1991; Madsen and Welles 2000; Brusatte et al. 2008; Cuesta et al. 2019). The presence of a ventral ridge in some anterior caudal vertebrae has been described in some allosauroids, including Neovenator and Lusovenator (Brusatte et al. 2008; Malafaia et al. 2020b). Even though the specimen is incomplete and distorted, it shows a combination of features compatible with allosauroid theropods. The three posterior caudal vertebrae (MG 4831a, b and MG 4828) show similar features and are interpreted here as possibly belonging to the same taxon (or even to the same individual). The centrum of MG 4831a is elongated and belongs to a vertebra of the posterior sector of the tail based on the absence of transverse processes, and a particularly reduced neural spine. The elongation of the centrum, with a height to length ratio near 0.3 is similar to that of the mid-posterior caudal vertebrae of several allosauroids (Gilmore 1920; Madsen 1976a; Mateus 1998; Brusatte et al. 2008; Malafaia et al. 2017b; Cuesta et al. 2019). The mid-posterior caudal vertebrae of most other early branching theropods have a more robust shape, with less elongated centra with height to length ratios > 0.5 (Gilmore 1920; Britt 1991; Sadleir et al. 2008). One exception is Elaphrosaurus from the Upper Jurassic of the Tendaguru Formation (Tanzania), which has also strongly elongated mid-posterior caudal vertebrae, but in this taxon the centra are wider than high (Rauhut and Carrano 2016), whereas in allosauroids they are approximately as high as wide.
Allosauridae Marsh, 1878 sensu Sereno 1998
Genus Allosaurus Marsh, 1877b
Type species: Allosaurus fragilis Marsh, 1877, from the Kimmeridgian-Tithonian Brushy Basin member of the Morrison Formation at Garden Park, Colorado, USA.
Allosaurus sp.
Fig. 25, SOM 5: fig. S4.
Previous identifications: MG 8775 is associated with a label with the attribution to Megalosaurus pombali probably corresponding to the specimen from Ribamar mentioned (but not figured) by Lapparent and Zbyszewski (1957) in the description of the species (Lapparent and Zbyszewski 1957: 25). MG 4817 is associated with a label with the identification Megalosaurus insignis from Paimogo, but it is also part of a set with another tooth fragment collected in Atalaia. None of the teeth attributed to this taxon by Lapparent and Zbyszewski (1957) were referred as having been collected at Paimogo so it is possible that this specimen came from Atalaia. MNHN/UL.EPt.019 was described and figured by Sauvage (1897–1898: pl. 5: 3), who referred it to M. insignis. MG 8781 is labeled as pertaining to Megalosaurus, but we have not found any reference to this specimen in previous publications. Finally, MNHN/UL.EPt.004 does not have any information about the identification and we could not find any reference to it on previous publications. Any information about the specimen could also be found in the database of the museum, probably because it is part of the collections that were partially destroyed in the fire of 1978.
Material.—Five tooth crowns (MG 8781, SOM 5: fig. S4B; MG 8775, Fig. 25B; MG 4817, Fig. 25A; MNHN/UL.EPt.019, Fig. 25C; MNHN/UL.EPt.004, SOM 5: fig. S4A). MG 8775 is from Outeiro do Seixo, near Ribamar, in the municipality of Lourinhã (Portugal), probably coming from upper Kimmeridgian–lowermost Tithonian levels of the Praia da Amoreira-Porto Novo Formation (Fig. 2) (Manuppella et al. 1999a). MG 4817 is associated with a label saying that it came from Paimogo, but it is also part of a set with another tooth fragment with the location Atalaia, both from the municipality of Lourinhã (Portugal). In both cases, the specimen was probably collected in sedimentary deposits of the Sobral Formation (Fig. 2) which is Kimmeridgian–Tithonian in age (Manuppella et al. 1999a). MG 8781 is labeled as coming from Pedras Muitas, near Baleal, in the municipality of Peniche (Portugal) (Fig. 2). This area is near the contact between the Alcobaça and Bombarral formations. However, the vertebrate fossil site of Pedras Muitas is marked in the Bombarral Formation on the geological map (Camarate França et al. 1960). If MG 8781 came from the same locality, it would have been collected from sedimentary rocks of the Bombarral Formation, which is Tithonian in age (Manuppella et al. 1999a). MNHN/UL.EPt.019 came from Pombal (Portugal), but there is no more specific information about where it was found. Finally, MNHN/UL.EPt.004 is part of the collections of MUHNAC and is associated with a label saying that it was probably collected from Lower Cretaceous levels at the locality of Cabo Espichel (Sesimbra, Portugal). However, its preservation is very different from other material from these levels, and it is a dental morphotype that has not yet been identified in the Lower Cretaceous of the Lusitanian Basin. It is more likely that this specimen came from an unknown Late Jurassic locality.
Description.—MG 4817 (Fig. 25A) and MG 8775 (Fig. 25B) are relatively well-preserved tooth crowns, which are triangular in lateral view and quite elongated (CHR = 2.25 for MG 4817 and 2.13 for MG 8775). The basal section is subcircular, with a CBR close to one in MG 8775. The crown is slightly recurved, with the mesial surface somewhat convex, the distal surface is straight, and the apex is placed at the level of the mesial carina. In MG 8775 the lingual surface is weakly concave in mesial view, whereas it is apically convex in MG 4817 (Fig. 25A4, B2). The labial surface is strongly convex, whereas the lingual surface has a convex centrally placed surface, but it is mostly flat adjacent to the distal margin and has a well-marked longitudinal concavity adjacent to the mesial carina. This longitudinal concavity is bounded mesially by the carina that projects labially (particularly in MG 4817). The enamel has a relatively well-marked irregular ornamentation and subtle transverse undulations are visible on both lingual and labial surfaces. In MG 8775, a well-developed spalled surface is present on the apex and extends on both lingual and mesial surfaces. The mesial and distal carinae are serrated and the denticles extend to the cervix in the distal carina and at least to the basal part of the mesial carina (the mesial surface is incomplete to the base). The distal carina is straight and placed on the distal surface, whereas the mesial carina is strongly twisted, being placed on the labial surface near the apex, but strongly twists to the base, being placed almost entirely in the lingual surface in the basal half of the crown. In MG 4817 there are 11 denticles per 5 mm in the apical and central sectors of the mesial carina and approximately 10 denticles per 5 mm in the basal sector of the mesial and distal carinae. In MG 8775 there are 10.5 and 12 denticles per 5 mm in the central and basal sectors of the mesial carina and 10, 10.5, and 11 denticles per 5 mm in the apical, central, and basal sectors of the distal carina, respectively (see SOM 2). The denticles of both mesial and distal carinae of MG 4817 are subquadrangular, with mostly straight to slightly convex external margins, whereas in MG 8775 they are rectangular, slightly longer mesiodistally than the apicobasal width. In MG 4817 there are neither interdenticular sulci between the denticles nor marginal and transverse undulations visible on the lingual and labial surfaces of the crown. However, in MG 8775 well-marked interdenticular sulci curving towards the base of the crown are present adjacent to the distal denticles in the lingual surface, but not between the mesial denticles.

Fig. 25. Mesial tooth crowns attributed to allosauroid theropod Allosaurus sp. from different Upper Jurassic localities of Portugal. A. MG 4817, ?Atalaia (Lourinhã region). B. MG 8775, Ribamar (Lourinhã region). C. MNHN/UL.EPt.019, unknown locality (Pombal region). Lingual (A1, B1, C1), labial (A2, B4, C5), distal (A3, B3, C2), and mesial (A4, B2, C4), views; basal cross-section (B5, C3); detail of the distal denticles from the central and basal sector of the crown respectively (C6, C7); detail of the mesial denticles from the central and basal sector of the crown respectively (C8, C9).
MNHN/UL.EPt.04 is an almost complete crown, missing the apex and a fragment of the lingual surface (SOM 5: fig. S4A1–A8). It has a similar morphology to the previously described specimens but is slightly shorter apicobasally (CHR = 1.58). The labial surface is strongly convex (at least the preserved part, all the surface adjacent to the distal margin is missing) and the lingual surface has a longitudinal convexity that is centrally positioned and delimited distally by a well-marked concave area adjacent to the distal carina. Due to this morphology of the lingual surface the base of the crown most likely had a J-shaped cross-section. The distal carina is centrally positioned on the distal surface of the crown in its apical portion, but strongly twists into the lingual surface to the base so it mostly faces lingually. The distal denticles are poorly preserved and only the morphology of those from the basal part of the crown can be described. They are rectangular with asymmetrically convex external margins (slightly pointing apically). The mesial denticles are subquadrangular, becoming more rectangular (larger mesiodistally than apicobasally) to the base of the crown. They have mostly symmetrically convex external margins, except those from the basal sector, which have somewhat apically pointed external convexities. No marginal or transverse undulations are visible, but the enamel has a well-marked ornamentation consisting of an irregular (non-oriented) texture that is more visible in the lingual surface adjacent to the distal carina. Finally, MG 8781 corresponds to a fragment of the apical part of a tooth crown (SOM 5: fig. S4B1–B5). It is poorly preserved but has a similar morphology to the previously described specimens, including the presence of a convex longitudinal surface in the central part of the lingual surface. The specimens MG 4817, 8775, 8781 and MNHN/UL.EPt.04 are interpreted as mesial tooth crowns based on the poor labiolingual compression, the strongly twisted mesial carina with a well-marked longitudinal groove, and the presence of a convex longitudinal surface in the central part of the lingual surface (see Hendrickx et al. 2020a).
MNHN/UL.EPt.019 is an almost complete and well-preserved crown (only the apical end is missing), interpreted here as a transitional tooth (Fig. 25C). The crown is relatively low (CHR = 2.14) and slightly compressed labiolingually (CBR = 0.61). In lateral view, it is triangular and slightly recurved distally, but the distal margin is mostly straight, and the mesial margin is slightly convex. The labial surface is flat for most of its width and convex adjacent to the mesial margin. The lingual surface is slightly convex but is weakly concave transversely because it projects somewhat lingually. The crown base section is oval to subcircular. The mesial and distal carinae are denticulated and both extend to the base of the crown (the mesial carina is somewhat incomplete, and the exact extension cannot be determined). The distal carina is centrally positioned in the distal margin and is mostly straight in distal view, but projects slightly lingually to the base of the crown. The mesial carina is centrally positioned on the mesial margin in the apical end of the crown but progressively twists to the lingual surface to the base. There is a narrow longitudinal sulcus on the lingual surface adjacent to the mesial carina, which projects for most of the height of the crown but becomes shallower apically. Transverse undulations are well marked in both lingual and labial surfaces, being slightly more evident lingually in the central part of the crown. A shallow longitudinal concavity is visible in the lingual surface adjacent to the distal carina, but it is restricted to the basal end of the crown (Fig. 25C1). The enamel has a subtle and irregular (non-oriented) ornamentation. There are 17.5 and 12 denticles per 5 mm in the central section of the mesial and distal carinae, respectively (see SOM 2). The distal denticles are mostly quadrangular (mesiodistally as long as the apicobasal width), with symmetrically convex external margins (Fig. 25C6, C7). They are separated by narrow interdenticular sulci that project only slightly into the surface of the crown. The denticles decrease in size to the base of the crown and the external margin becomes asymmetrically convex (pointing apically). The mesial denticles are rectangular (slightly larger apicobasally than mesiodistally). The external margin is also symmetrically convex, and the denticles become smaller towards the base of the crown (Fig. 25C8, C9). The mesial denticles are separated by narrow interdenticular sulci, which do not extend on the surface of the crown.
Remarks.—The discriminant analysis classifies two specimens (MG 8775, 4817) as belonging to Allosaurus, MNHN/UL.EPt.019 as Genyodectes and MNHN/UL.EPt.004 as Dromaeosaurus (see Table 1). The plot shows similar results for the distribution of these specimens, but three of the specimens fall within the morphospace of Allosaurus and only MNHN/UL.EPt.004 is placed with Dromaeosaurus (Fig. 16). The cladistic analysis of the dentition-based data matrix with constraints and including all studied morphotypes recovered the specimens here grouped in Morphotype 3 at the base of the Allosauroidea clade in polytomy with Sinraptor and Erectopus (Fig. 18A). The results of the analysis pruning a priory all the morphotypes from the Lusitanian Basin but Morphotype 3 found 2 MPTs (CI = 0.204, RI = 0.464, L = 1316). The consensus tree from these MPTs (CI = 0.203, RI = 0.463, L = 1318) shows a better resolved Allosauroidea clade and similarly to the previous analysis, Morphotype 3 is allied to Sinraptor and Erectopus (Fig. 21C). However, this result should be interpreted with caution due to the similarity on both discrete and quantitative features of the dentition of Sinraptor and Allosaurus that show almost indistinguishable mesial and lateral tooth morphology (Hendrickx et al. 2020a). The synapomorphies found in the analysis supporting the position of Morphotype 3 within a group with Sinraptor and Erectopus include the straight distal margin of the crown in lateral view and the presence of short interdenticular sulci, which are features also present in Allosaurus (see Hendrickx et al. 2020a, b).
There is no evidence of flutes in the lingual surface, which is a feature present in the mesial teeth of some ceratosaurians, such as Masiakasaurus and Ceratosaurus (Madsen and Welles 2000; Carrano et al. 2012; Soto and Perea 2008; Rauhut 2011). The crowns are poorly compressed labiolingually (CBR close to 1, in MG 8775, or higher than 0.6), with both mesial and distal carinae extending to the cervix. The mesial carina strongly twists lingually to the base of the crown and the lingual surface has a longitudinal centrally positioned convexity, resulting in a characteristic J-shaped cross-sectional outline. This combination of features is exclusive to the teeth from the mesial dentition of Allosaurus and Sinraptor (Hendrickx et al. 2015b, 2020a). Based on this combination of features, together with their paleogeographic and stratigraphic distribution, the specimens of Morphotype 3 are here interpreted as a mesial teeth of Allosaurus. In three of the described specimens (MG 8775, 4817 and MNHN/UL.EPt.019), the mesial carina is strongly twisted lingually to the base of the crown (particularly marked in MG 4817), defining a longitudinal groove in the lingual surface adjacent to the carina that extends for most of the height of the crown. On the other hand, the mesial carina of MNHN/UL.EPt.019 is much less twisted lingually and the crown is more compressed labiolingually, with an oval cross-section of the base. These features suggest that this specimen is a transitional tooth (from a position between the most mesial and lateral teeth, possibly a first or second maxillary tooth).
Discussion
The osteological remains studied here are mostly fragmentary and isolated but represent a relatively diverse fauna of Late Jurassic dinosaurs, which is particularly well-documented for the coastal region of the Lusitanian Basin. Also, several specimens come from the region of Pombal, Leiria, Batalha, and Ourém (belonging to the Mamede sub-basin). Most of this material was collected from Tithonian levels of the Bombarral Formation (25 specimens) and from upper Kimmeridgian strata of the Praia da Amoreira-Porto Novo Formation (18 specimens). Of the material first assigned to Megalosaurus only 50% actually belongs to theropods, whereas 12% corresponds to sauropods, 9% to stegosaurs, and 8% to ornithopods, beside two small cervical vertebrae of a thalattosuchian crocodylomorph (see SOM 1). Sauropods are represented by the proximal end of a right humerus of an indeterminate eusauropod, three fragments of caudal vertebrae of diplodocines (Mocho et al. 2016), and a partial anterior caudal vertebra of an indeterminate titanosauriform. Also, a distal end of a metacarpal and several vertebral fragments of indeterminate sauropods were described. Material assigned to indeterminate stegosaurs includes several fragments of dorsal and caudal vertebrae. Ornithopods are represented by some isolated cervical and caudal vertebrae assigned to indeterminate iguanodontians and a sequence of caudal vertebrae referred to Ankylopollexia sp. (Escaso et al. 2017). Finally, theropods are mainly represented by isolated teeth referred to as cf. Ceratosaurus sp., Torvosaurus gurneyi, and Allosaurus sp., beside some dorsal, sacral, and caudal vertebrae assigned to indeterminate avetheropods and allosauroids.
To support the taxonomic identification of the isolated theropod teeth, we conducted a series of analyses based on both morphometric and cladistic methods. The results of the morphometric analyses showed to be inconsistent for most of the studied specimens, except for some specific morphotypes (e.g., mesial tooth crowns of Allosaurus). Limitations of the morphometric analysis (DFA) on the identification of isolated theropod teeth have been identified in the literature (e.g., Gerke and Wings 2016; Hendrickx et al. 2020b). The use of size-dependent variables, together with possible differences in the methodology applied to measure the tooth crowns and the fragmentary state of the specimens may strongly influence the results of the DFA (Hendrickx et al. 2020b). The cladistic analysis shows somewhat more consistent results. However, well-resolved topologies were only obtained enforcing constrains and even in these analyses some branches still poorly defined, possibly reflecting high degree of homoplasy in the dentition morphology of those taxa and/or missing data (Hendrickx et al. 2020a, b).
The assemblage of Late Jurassic theropods here identified is compatible with the diversity currently described in the Lusitanian Basin. The record of allosauroid theropods is the most abundant and diverse in this assemblage, being represented in different areas, including the Consolação and Mamede sub-basins. These theropods seem to predominate in the Mamede sub-basin, where currently no evidence of Torvosaurus has yet been identified. Also, a small tooth crown from the area of Cabo Espichel, which is here assigned to an indeterminate allosauroid, may represent the first record of this clade in the Lower Cretaceous of Portugal. The fossil record of Late Cretaceous theropods currently known from the Lusitanian Basin is scarce, but among the material first referred to Megalosaurus are some tooth crown fragments belonging to an indeterminate medium or large sized form and three ungual phalanges assigned to coelurosaurs (Galton 1994).
The Lusitanian Basin has yielded one of the most extensive fossil collections of Late Jurassic dinosaurs and other vertebrates from Europe. The similarity with coeval faunas of the North American Morrison Formation has been well-documented, with several shared genera (e.g., Stegosaurus, Escaso et al. 2007; Ceratosaurus, Mateus and Antunes 2000a; Malafaia et al. 2015; Torvosaurus, Mateus and Antunes 2000b; Hendrickx and Mateus 2014; Malafaia et al. 2017c; Allosaurus, Pérez-Moreno et al. 1999; Mateus et al. 2006). Other European Upper Jurassic fossil records are scarce (particularly for theropod dinosaurs) and mostly consist of fragmentary material with uncertain phylogenetic interpretation (e.g., Allain and Pereda Suberbiola 2003; Canudo and Ruiz-Omeñaca 2003; Ruiz-Omeñaca et al. 2008; Lubbe et al. 2009; Cobos et al. 2014; Evers and Wings 2020). Isolated theropod teeth are possibly the most abundant dinosaurian remains in the Upper Jurassic of Europe. Despite most of these records need to be revised based on more recent methodologies, some similarities with the assemblage of theropod tooth morphotypes from the Upper Jurassic of the Lusitanian Basin have been identified, particularly from the upper Kimmeridgian–Berriasian of the Villar del Arzobispo Formation, in Teruel (Spain) and different localities of northern Germany (e.g., Gascó et al. 2012; Cobos et al. 2014; Gerke and Wings 2016).
Conclusions
We have studied a collection of osteological remains from the Upper Jurassic and Cretaceous of the Lusitanian Basin, initially attributed to Megalosaurus, reinterpreting their taxonomic identification, and updating their stratigraphic context. This study allows reinterpretation of this material as belonging to different vertebrates, including thalattosuchian crocodyliforms, and several dinosaur groups represented by stegosaurian and iguanodontian ornithischians, sauropods (including indeterminate eusauropod and titanosauriforms), and various theropod taxa. Remains of theropod dinosaurs are the most abundant in the analyzed sample, represented mainly by isolated teeth and vertebrae here attributed to cf. Ceratosaurus, Torvosaurus, and Allosaurus. Besides, a small dorsal vertebra is interpreted as belonging to a juvenile indeterminate avetheropod and some sacral and caudal vertebrae are assigned to indeterminate allosauroids. Several isolated teeth referred to Allosaurus from different parts of the tooth row, including the most mesial and the transition sectors, witness of changes in the dentition morphology for this taxon.
Most of the specimens come from Upper Jurassic (Kimmeridgian to upper Tithonian) levels cropping out in the coastal sector of the Lusitanian Basin, generally from localities in the area of Lourinhã, Peniche, and Caldas da Rainha (Consolação and Bombarral sub-basins) or from the region of Pombal, Leiria, Batalha, and Ourém (Mamede sub-basin). As already established from other records, the Upper Jurassic sedimentary levels of the Consolação sub-basin and the coastal region of the Bombarral sub-basins have provided the highest diversity of dinosaurs. This study suggests that allosauroid theropods predominate in the Mamede sub-basin, where currently no records of other taxa that are well represented in the Consolação and Bombarral sub-basins, such as Torvosaurus (and possibly Ceratosaurus), have been identified. This distribution may indicate some paleoenvironmental biases in different sectors of the Lusitanian Basin. There are also few isolated teeth of theropod dinosaurs collected in Lower Cretaceous (lower Barremian) localities in the region of Cabo Espichel (Setúbal sub-basin) and in the Upper Cretaceous (Campanian–Maastrichtian) of the Aveiro sub-basin. The material of the latter is particularly interesting because most of the localities where it was found are no longer available and the fossil record in these levels is poorly known. One of the teeth collected in the locality of Boca do Chapim is interpreted here as possibly belonging to an allosauroid theropod, which would be the first record of this clade in the Lower Cretaceous of the Lusitanian Basin.
Acknowledgements
We thank Jorge Sequeira (MG), Liliana Póvoas and Judite Alves (both MUHAC), and Carlos Anunciação (MMLT) for access to the specimens. We also thank to Roberto Keller (MUHNAC) for access the microscope to study and photograph the specimens and to Bruno Maggia (Faculdade de Ciências, Universidade de Lisboa, Portugal) for his help on the treatment of the images, as well as reviewers Ronan Allain (Muséum National d’Histoire Naturelle, Paris, France) and Cristophe Hendrickx (CONICET-Fundación Miguel Lillo, Tucumán, Argentina) for their comments and suggestions, which significantly improved the quality of this work. We acknowledge the Willi Hennig Society and Øyvind Hammer (Natural History Museum, University of Oslo, Norway) for the use of TNT and Past3, respectively. This work was supported by the Fundação para a Ciência e a Tecnologia (FCT) under Project CEECIND/01770/2018 (https://doi.org/10.54499/CEECIND/01770/2018/CP1534/CT0004); and I.P./MCTES through national funds (PIDDAC)-UIDB/50019/2020 (https://doi.org/10.54499/UIDB/50019/2020), and LA/P/0068/2020 (https://doi.org/10.54499/LA/P/0068/2020).
References
Allain, R. 2001. Redescription de Streptospondylus altdorfensis, le dinosaure théropode de Cuvier, du Jurassique de Normandie. Geodiversitas 23: 349–367.
Allain, R. 2002. Discovery of megalosaur (Dinosauria, Theropoda) in the middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae. Journal of Vertebrate Paleontology 22: 548–563. Crossref
Allain, R. 2005. The postcranial anatomy of the megalosaur Dubreuillosaurus valesdunensis (Dinosauria Theropoda) from the middle Jurassic of Normandy, France. Journal of Vertebrate Paleontology 25: 850–858. Crossref
Allain, R. and Chure, D.J. 2002. Poekilopleuron bucklandii, the theropod dinosaur from the Middle Jurassic (Bathonian) of Normandy. Palaeontology 45: 1107–1121. Crossref
Allain, R. and Pereda Suberbiola, X. 2003. Dinosaurs of France. Comptes Rendus Palevol 2: 27–44. Crossref
Allain, R., Vullo, R., Rozada, L., Anquetin, J., Bourgeais, R., Goedert, J., Lasseron, M., Martin, J.E., Pérez-García, A., Peyre De Fabrègues, C., Royo-Torres, R., Augier, D., Bailly, G., Cazes, L., Despres, Y., Gailliègue, A., Gomez, B., Goussard, F., Lenglet, T., Vacant, R., Mazan, and Tournepiche, J.-F. 2022. Vertebrate paleobiodiversity of the Early Cretaceous (Berriasian) Angeac-Charente Lagerstätte (southwestern France): implications for continental faunal turnover at the J/K boundary. Geodiversitas 44: 683–752. Crossref
Allain, R., Xaisanavong, T., Richir, P., and Khentavong, B. 2012. The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early Cretaceous of Laos. Naturwissenschaften 99: 369–377. Crossref
Alonso, A. and Canudo, J.I. 2016. On the spinosaurid theropod teeth from the early Barremian (Early Cretaceous) Blesa Formation (Spain). Historical Biology 28: 823–834. Crossref
Alves, T.M., Gawthorpe, R.L., Hunt, D.W., and Monteiro, J.H. 2002. Jurassic tectono-sedimentary evolution of the Northern Lusitanian Basin (offshore Portugal). Marine and Petroleum Geology 19: 727–754. Crossref
Alves, T.M, Manuppella, G., Gawthorpe, R.L., Hunt, D.W., and Monteiro, J.H. 2003. The depositional evolution of diapir- and fault-bounded rift basins: examples from the Lusitanian Basin of West Iberia. Sedimentary Geology 162: 273–303. Crossref
Antunes, M.T. 1976. Dinossáurios eocretácicos de Lagosteiros, Ciências da Terra UNL 1: 1–35.
Antunes, M.T. and Mateus, O. 2003. Dinosaurs of Portugal. Palevol 2: 77–95. Crossref
Antunes, M.T. and Pais, J. 1978. Notes on deposits from Taveiro stratigraphy, paleontology, age, paleoecology. Ciências da Terra UNL 4: 109–128.
Antunes, M.T. and Sigogneau-Russell, D. 1991. New data on dinosaurs from the Upper Cretaceous of Portugal. Comptes Rendus de l’Académie des Scieces Paris 313 (Serie 11): 113–119.
Antunes, M.T. and Sigogneau-Russell, D. 1992. Fauna of small dinosaurs from the Portuguese late Cretaceous. Comunicações dos Serviços Geológicos de Portugal 78 (1): 49–62.
Azerêdo, A.C., Cabral, M.C., Martins, M.J., Loureiro, I.M., and Inês, N. 2010. Stratigraphic study of a new outcrop of the Cabaços Formation (Oxfordian) in the Serra do Bouro region (Caldas da Rainha). Comunicações Geológicas 97: 5–22.
Azerêdo, A.C., Ramalho, M.M., and Wright, P.V. 1998. The Middle–Upper Jurassic disconformity in the Lusitanian Basin, Portugal: preliminary facies analysis and evidence for palaeoclimatic fluctuation. Cuadernos de Geología Ibérica 24: 99–119.
Bakker, R.T., Siegwarth, J.D., Kralis, D., and Filla, J. 1992. Edmarka rex: A new, gigantic theropod dinosaur from the middle Morrison Formation, Late Jurassic of the Como Bluff outcrop region. Hunteria 2: 1–24.
Barbosa, B.P., Soares, A.F., Rocha, R.B., Manuppella, G., and Henriques, M.H. 2008. Geological map of Portugal at a scale of 1:50.000: explanatory news of the sheet 19-A (Cantanhede). Departamento de Geologia, Instituto Nacional de Engenharia, Tecnologia e Inovação, Lisboa.
Bardet, N., Pereda Suberbiola, X., and Ruiz-Omeñaca, J.I. 2008. Mesozoic marine reptiles from the Iberian Peninsula. Geo-Temas 10: 1245–1248.
Barker, C.T., Hone, D.W.E., Naish, D., Cau, A., Lockwood, J.A.F., Foster, B., Clarkin, C.E., Schneider, P., and Gostling, N.J. 2021. New spinosaurids from the Wessex Formation (Early Cretaceous, UK) and the European origins of Spinosauridae. Scientific Reports 11: 1–15. Crossref
Barrett, P.M., Benson, R.B.J., Rich, T.H. and Vickers-Rich, P. 2011. First spinosaurid dinosaur from Australia and the cosmopolitanism of Cretaceous dinosaur faunas. Biology Letters 7: 933–936. Crossref
Benson, R.B.J. 2008. A redescription of “Megalosaurus” hesperis (Dinosauria, Theropoda) from the Inferior Oolite (Bajocian, Middle Jurassic) of Dorset, United Kingdom. Zootaxa 1931: 57–67. Crossref
Benson, R.B.J. 2009. An assessment of variability in theropod dinosaur remains from the Bathonian (Middle Jurassic) of Stonesfield and New Park quarry, UK and taxonomic implications for Megalosaurus bucklandii and Iliosuchus incognitus. Palaeontology 52: 857–877. Crossref
Benson, R.B.J. 2010a. A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods. Zoological Journal of the Linnean Society 158: 882–935. Crossref
Benson, R.B.J. 2010b. The osteology of Magnosaurus nethercombensis (Dinosauria, Theropoda) from the Bajocian (Middle Jurassic) of the United Kingdom and a re-examination of the oldest records of tetanurans. Journal of Systematic Palaeontology 8: 131–146. Crossref
Benson, R.B.J., Barrett, P.M., Powell, H.P., and Norman, D.B. 2008. The taxonomic status of Megalosaurus bucklandii (Dinosauria, Theropoda) from the Middle Jurassic of Oxfordshire, UK. Palaeontology 51: 419–424. Crossref
Bittencourt, J.S. and Kellner, A.W.A. 2004. On a sequence of sacrocaudal theropod dinosaur vertebrae from the Lower Cretaceous Santana Formation, northeastern Brazil. Arquivos do Museu Nacional, Rio de Janeiro 62: 309–320.
Bonaparte, J.F. 1986. The dinosaurs (carnosaurs, allosaurids, sauropods, cetiosaurids) from the Middle Jurassic of Cerro Cóndor (Chubut, Argentina). Annales de Paléontologie 72: 247–386.
Britt, B. 1991. Theropods of Dry Mesa Quarry (Morrison Formation, Late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri. In: B.J. Kowallis and K. Seeley (eds.), Young University Geology Studies 37, 1–72. Ocean Press, Provo.
Brusatte, S.L., Benson, R.B.J., and Hutt, S. 2008. The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden Group (Barremian) of the Isle of Wight. Monograph of the Palaeontographical Society 162: 1–75.
Brusatte, S., Carr, T.D., Erickson, G.M., Bever, G.S., and Norell, M.A. 2009. A long-snouted, multihorned tyrannosaurid from the Late Cretaceous of Mongolia. PNAS 106: 17261–17266. Crossref
Buckland, W. 1824. Notice on the Megalosaurus or great fossil lizard of Stonesjield. Transactions of the Geological Society of London 2-1: 390–396. Crossref
Buckley, L.G. and Currie, P.J. 2014. Analysis of intraspecific and ontogenetic variation in the dentition of Coelophysis bauri (Late Triassic), and implications for the systematics of isolated theropod teeth. In: S.G. Lucas, R. Sullivan, and L.H. Tanner (eds.), New Mexico Museum of Natural History & Science, Bulletin 63: 1–54.
Buffetaut, E. 2007. The spinosaurid dinosaur Baryonyx (Saurischia, Theropoda) in the Early Cretaceous of Portugal. Geology Magazine 144: 1021–1025. Crossref
Buffetaut, E. 2011. An early spinosaurid dinosaur from the Late Jurassic of Tendaguru (Tanzania) and the evolution of the spinosaurid dentition. Oryctos 10: 1–8.
Buffetaut, E. and Ingavat, R. 1986. Unusual theropod dinosaur teeth from the Upper Jurassic of Phu Wiang, northeastern Thailand. Revue de Paleobiology 5: 217–220.
Camarate França, J. and Zbyszewski, G. 1963. Geological map of Portugal at a scale of 1:50000: explanatory news of the sheet 26-B (Alcobaça). Serviços Geológicos de Portugal, Lisboa.
Camarate França, J., Zbyszewski, G., and Almeida, F.M. 1960. Geeologic map of Portugal at a scale of 1:50 000: explanatory news of the sheet 26-C (Peniche). Serviços Geológicos de Portugal, Lisboa.
Canale, J.I., Apesteguía, S., Gallina, P.A., Gianechini, F.A., and Haluza, A. 2016. The oldest theropods from the Neuquén Basin: Predatory dinosaur diversity from the Bajada Colorada Formation (Lower Cretaceous: Berriasian–Valanginian), Neuquén, Argentina. Cretaceous Research 71: 63–78. Crossref
Canale, J.I., Novas, F.E., Salgado, L., and Coria, R.A. 2014. Cranial ontogenetic variation in Mapusaurus roseae (Dinosauria: Theropoda) and the probable role of heterochrony in carcharodontosaurid evolution. Paläontologische Zeitschrift 89: 983–993. Crossref
Candeiro, C.R., Gil, L.M., and de Castro, P.E.P. 2018. Large-sized theropod Spinosaurus: an important component of the carnivorous dinosaur fauna in southern continents during the Cretaceous. Bulletin de la Société Géologique de France, Earth Sciences Bulletin 189: 1–9. Crossref
Canudo, J.I. and Ruiz-Omeñaca, J.I. 2003. Direct remains of theropod dinosaurs (excluding birds) in Spain. In: F.P. Lorent, M.M.R. Molina, P.R. Carrera (eds.), Dinosaurios y otros reptiles mesozoicos en España, Ciencias de la Tierra 26, 347–373. Universidad de La Rioja, Instituto de Estudios Riojanos, La Rioja.
Canudo, J.I., Gasulla, J.M., Gómez-Fernández, D., Ortega, F., Sanz, J.L., and Yagüe, P. 2008. Primera evidencia de dientes aislados atribuidos a Spinosauridae (Theropoda) en el Aptiano inferior (Cretácico Inferior) de Europa: Formación Arcillas de Morella (España). Ameghiniana 45: 649–662.
Carpenter, K. and Wilson, Y. 2008. A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb. Annals of Carnegie Museum 76: 227–264. Crossref
Carpenter, K., Miles, C.A., and Cloward, K. 2001. New primitive stegosaur from the Morrison Formation, Wyoming. In: K. Carpenter (ed.), The Armored Dinosaurs, 55–75. Indiana University Press, Bloomington.
Carpenter, K., Miles, C., Ostrom, J.H., and Cloward, K. 2005. Redescription of the small maniraptoran theropods Ornitholestes and Coelurus from the Upper Jurassic Morrison Formation of Wyoming. In: K. Carpenter (ed.). The Carnivorous Dinosaurs, 49–71. Indiana University Press, Bloomington.
Carrano, M.T., Benson, R.B.J., and Sampson, S.D. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology 10: 211–300. Crossref
Carrington da Costa, J. 1940. A new chelonian fossil. Comunicações dos Serviços Geológicos de Portugal 21: 107–125.
Carvalho, A.M.G. and Santos, V.F. 1993. Dinosaur footprints from Sesimbra. Sesimbra Cultural 3: 10–14.
Charig, A.J. and Milner, A.C. 1997. Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey. Bulletin of the Natural History Museum. Geology Series 53: 11–70.
Choffat, P. 1904. The Cretaceous of the Arrabida and Ericeira regions. Comunicações de Comissão dos Serviços Geológicos de Portugal 6: 1–44.
Cobos, A. and Gascó, F. 2013. New vertebral remains of the stegosaurian dinosaur Dacentrurus from Riodeva (Teruel, Spain). Geogaceta 53: 17–20.
Cobos, A., Lockley, M.G., Gascó, F., Royo-Torres, R., and Alcalá, L. 2014. Megatheropods as apex predators in the typically Jurassic ecosystems of the Villar del Arzobispo Formation (Iberian Range, Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 399: 31–41. Crossref
Cobos, A., Royo-Torres, R., Luque, L., Alcalá, L., and Mampel, L. 2010. An Iberian stegosaurs’ paradise: the Villar del Arzobispo Formation (Tithonian–Berriasian) in Teruel (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 293: 223–236. Crossref
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the American Philosophical Society 14 (1): 1–252. Crossref
Costa, F. and Mateus, O. 2019. Dacentrurine stegosaurs (Dinosauria): A new specimen of Miragaia longicollum from the Late Jurassic of Portugal resolves taxonomical validity and shows the occurrence of the clade in North America. PLoS ONE 14: e0224263. Crossref
Cuesta, E., Ortega, F., and Sanz, J.L. 2019. Axial osteology of Concavenator corcovatus (Theropoda; Carcharodontosauria) from the Lower Cretaceous of Spain. Cretaceous Research 95: 106–120. Crossref
Cuesta, E., Vidal, D., Ortega, F., and Sanz, J.L. 2018. The cranial osteology of Concavenator corcovatus (Theropoda; Carcharodontosauria) from the Lower Cretaceous of Spain. Cretaceous Research 91: 176–194. Crossref
Currie, P.J. 1995. New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). Journal of Vertebrate Paleontology 15: 576–591. Crossref
Currie, P.J. and Zhao, X.-J. 1993. A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Canadian Journal of Earth Sciences 30: 2037–2081. Crossref
Currie, P.J., Rigby, J.K.J., and Sloan, R.E. 1990. Theropod teeth from the Judith River Formation of southern Alberta, Canada. In: K. Carpenter and P.J. Currie (eds.), Dinosaur Systematics: Approaches and Perspectives, 107–125. Cambridge University Press, Cambridge. Crossref
Dantas, P. 1990. Dinosaurs from Portugal. Gaia 2: 17–26.
Dantas, P., dos Santos, V.F., Lockley, M.G., and Meyer, C.A. 1994. Footprint evidence for limping dinosaurs from the Upper Jurassic of Portugal. Gaia 10: 43–48.
Dal Sasso, C., Maganuco, S., Buffetaut, E., and Mendez, M.A. 2005. New information on the skull of the enigmatic theropod Spinosaurus, with remarks on the size and affinities. Journal of Vertebrate Paleontology 25: 888–896. Crossref
Day, J.J. and Barrett, P.M. 2004. Material referred to Megalosaurus (Dinosauria: Theropoda) from the Middle Jurassic of Stonesfield, Oxfordshire, England: one taxon or two? Proceedings of the Geologists’ Association 115: 359–366. Crossref
D’Emic, M.D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society 166: 624–671. Crossref
Dinis, J.L., Rey, J., Cunha, P.P., Callapez, P., and Pena dos Reis, R. 2008. Stratigraphy and allogenic controls of the western Portugal Cretaceous: an updated synthesis. Cretaceous Research 29: 772–780. Crossref
Dollo, L. 1888. Iguanodontidae et Camptonotidae. Comptes Rendus de l’Académie des Sciences 106: 775–777.
Escaso, F. 2014. Evolutionary History of the Ornithischia (Dinosauria) From the Upper Jurassic of Portugal. 289 pp. Unpublished Ph.D. Thesis, Universidad Autónoma de Madrid, Madrid.
Escaso, F., Malafaia, E., Mocho, P., Narváez, I., and Ortega, F. 2017. “Megalosaurus insignis” from Praia de Areia Branca (Lourinhã, Portugal): is it Theropoda or Ornithopoda? In: A.B. Díaz, F. Barroso-Barcenilla, and P.C. Tonicher (eds.), Livro de Resumos da XXII Bienal da Real Sociedad Española de Historia Natural, 227. Real Sociedad Española de Historia Natural, Madrid.
Escaso, F., Ortega, F., Dantas, P., Malafaia, E., Silva, B., Gasulla, J.M., Mocho, P., Narváez, I., and Sanz, J.L. 2014. A new dryosaurid ornithopod (Dinosauria, Ornithischia) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology 34: 1102–1112. Crossref
Escaso, F., Ortega, F., Dantas, P., Malafaia, E., Pimentel, N.L., Pereda-Suberbiola, X., Sanz, J.L., Kullberg, J.C., Kullberg, M.C., and Barriga, F. 2007. New evidence of shared dinosaur across Upper Jurassic Proto-North Atlantic: Stegosaurus from Portugal. Naturwissenschaften 94: 367–374. Crossref
Evers, S.W. and Wings, O. 2020. Late Jurassic theropod dinosaur bones from the Langenberg Quarry (Lower Saxony, Germany) provide evidence for several theropod lineages in the central European archipelago. PeerJ 8: e8437. Crossref
Evers, S.W., Rauhut, O.W.M., Milner, A.C., McFeeters, B., and Allain, R. 2015. A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco. PeerJ 3: e1323. Crossref
Figueiredo, S.D., de Carvalho, C.N., Cunha, P.P., and Sousa Carvalho, I.S. 2021. New dinosaur tracks from the Lower Barremian of Portugal (Areia do Mastro Formation, Cape Espichel). Journal of Geoscience and Environment Protection 9: 84–96. Crossref
Figueiredo, S., Rosina, P., and Figuti, L. 2015. Dinosaurs and other vertebrates from the Papo-Seco Formation (Lower Cretaceous) of southern Portugal. Journal of Iberian Geology 41: 301–314. Crossref
Figueiredo, S., Rosina, P., and Gomes, M. 2018. New remains of a avian dinosaur from the Lower Cretaceous of Portugal (Areias do Mastro-Sesimbra). Revista de Geo-História e Pré-História 2: 66–69.
Figueiredo, S., Strantzali, I.B., Gomes, M., Pimenta, R., and Santos, M. 2017. Preliminary data of new dinosaurs and turtles remains from the basal deposits of the Papo-Seco Formation in Areias do Mastro´s (Cabo Espichel, Portugal). Arnava 6 (1): 117–130.
Fitzinger, L. 1843. Systema Reptilium. Fasciculus primus Amblyglossae. Vindobonae. 106 + vi pp. Braumüller und Seidel, Viena. Crossref
Fraas, E. 1901. The sea crocodiles (Thalattosuchia n. g.) are a new saurian group from the Jurassic formation. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 57: 409–418.
Fürsich, F.T., Schneider, S., Werner, W., Lopez-Mir, B., and Pierce, C.S. 2021. Life at the continental-marine interface: palaeoenvironments and biota of the Alcobaça Formation (Late Jurassic, Central Portugal), with a formal definition of the unit appended. Palaeobiodiversity and Palaeoenvironments 102: 265–329. Crossref
Galton, P.M. 1981. Dryosaurus, a hypsilophodontid dinosaur from the Upper Jurassic of North America and Africa. Postcranial skeleton. Paläontologische Zeitschrift 55: 271–312. Crossref
Galton P.M. 1991. Postcranial remains of the stegosaurian dinosaur Dacentrurus from the Upper Jurassic of France and Portugal. Geologica et Paleontologica 25: 299–327.
Galton, P.M. 1994. Notes on Dinosauria and Pterodactylia from the Cretaceous of Portugal. Neues Jahrbuch für Geologie and Paläontologie Abhandlungen 194: 253–267. Crossref
Galton, P.M. 1996. Notes on Dinosauria from the Upper Cretaceous of Portugal. Neues Jahrbuch für Geologie and Paläontologie Monatshefte 1996 (2): 83–90. Crossref
Galton, P.M. and Jensen, J.A. 1979. A new large theropod dinosaur from the Upper Jurassic of Colorado. Brigham Young University Geology Studies 26: 1–12.
Galton, P.M. and Upchurch, P. 2004. Stegosauria. In: D.B. Weishampel, P. Dodson, and H. Osmólska (ed.), The Dinosauria, 2nd Edition, 343–362. University of California Press, Berkeley. Crossref
Gascó, F., Cobos, A., Royo-Torres, R., Alcalá, L. and Mampel, L. 2012. Theropod teeth diversity from Villar del Arzobispo Formation (Tithonian–Berriasian) at Riodeva (Teruel, Spain). Palaeobiodiversity and Palaeoenvironments 92: 273–286. Crossref
Gauthier, J. 1986. Saurischian monophyly and the origin of birds. In: K. Padian (ed.), The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8: 1–55.
Gerke, O. and Wings, O. 2016. Multivariate and cladistic analyses of isolated teeth reveal sympatry of theropod dinosaurs in the Late Jurassic of Northern Germany. PLoS ONE 11: e0158334. Crossref
Gianechini, F.A., Makovicky, P.J., and Apesteguía, S. 2011. The teeth of the unenlagiine theropod Buitreraptor from the Cretaceous of Patagonia, Argentina, and the unusual dentition of the Gondwanan dromaeosaurids. Acta Palaeontologica Polonica 56: 279–290. Crossref
Gilmore, C.W. 1909. Osteology of the Jurassic reptile Camptosaurus, with a revision of the species of the genus, and a description of two new species. Proceedings of the United States National Museum 36: 197–332. Crossref
Gilmore, C.W. 1914. Osteology of the armoured Dinosauria in the United States National Museum, with special reference to the genus Stegosaurus. Bulletin of the United States National Museum 89: 1–143. Crossref
Gilmore, C.W. 1920. Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus. Bulletin of the United States National Museum 110: 1–154. Crossref
Goloboff, P.A. and Morales, M.E. 2023. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39: 144–153. Crossref
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. Past: Paleontological Statistics Software Package for education and data analysis. Palaeontologia Electronica 4 (1): 1–9.
Hanson, M. and Makovicky, P.J. 2014. A new specimen of Torvosaurus tanneri originally collected by Elmer Riggs. Historical Biology 26: 775–784. Crossref
Harris, J.D. 1998. A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiogeographic implications, based on a new specimen from Texas. Bulletin of the New Mexico Museum of Natural History & Science 13: 1–75.
Hay, O.P. 1930. Second Bibliography and Catalogue of the Fossil Vertebrata of North America. Volume 2. 990 pp. Carnegie Institute, Washington.
Hendrickx, C. and Mateus, O. 2014. Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian theropods. PLoS ONE 9: e88905. Crossref
Hendrickx, C., Mateus, O., and Araújo, R. 2015a. A proposed terminology of theropod teeth (Dinosauria, Saurischia). Journal of Vertebrate Paleontology 35: e982797. Crossref
Hendrickx, C., Mateus, O., and Araújo, R. 2015b. The dentition of megalosaurid theropods. Acta Palaeontologica Polonica 60: 627–642. Crossref
Hendrickx, C., Mateus, O., Araújo, R., and Choiniere, J. 2019. The distribution of dental features in non-avian theropod dinosaurs: Taxonomic potential, degree of homoplasy, and major evolutionary trends. Palaeontologia Electronica 22.3.74: 1–110. Crossref
Hendrickx, C., Stiegler, J., Currie, P.J., Han, F., Xu, X., Choiniere, J.N., and Wu, X.-C. 2020a. Dental anatomy of the apex predator Sinraptor dongi (Theropoda: Allosauroidea) from the Late Jurassic of China. Canadian Journal of Earth Sciences 57: 1127–1147. Crossref
Hendrickx, C., Tschopp, E. and Ezcurra, M.D. 2020b. Taxonomic identification of isolated theropod teeth: the case of the shed tooth crown associated with Aerosteon (Theropoda: Megaraptora) and the dentition of Abelisauridae. Cretaceous Research 108: 104312. Crossref
Hennig, E. 1915. Kentrosaurus aethiopicus, the stegosauridae of Tendaguru. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin 1915: 219–247.
Hill, G. 1988. The Sedimentology and Lithostratigraphy of the Upper Jurassic Lourinhã Formation, Lusitanian Basin, Portugal. 292 pp. Unpublished Ph.D. Thesis, Open University, Milton Keynes.
Huberty, C.J. and Olejnik, S. 2006. Applied MANOVA and Discriminant Analysis. 2nd Edition. John Wiley & Sons, Hoboken. Crossref
Ibrahim, N., Sereno, P.C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D.M., Zouhri, S., Myhrvold, N., and Iurino, D.A. 2014. Semiaquatic adaptations in a giant predatory dinosaur. Science 345: 1613–1616. Crossref
Irmis, R.B. 2007. Axial skeleton ontogeny in the Parasuchia (Archosauria: Pseudosuchia) and its implications for ontogenetic determination in archosaurs. Journal of Vertebrate Paleontology 27: 350–361. Crossref
Isasmendi, E., Navarro-Lorbés, P., Sáez-Benito, P., Viera, L.I., Torices, A., and Pereda-Suberbiola, X. 2023. New contributions to the skull anatomy of spinosaurid theropods: Baryonychinae maxilla from the Early Cretaceous of Igea (La Rioja, Spain). Historical Biology 35: 909–923. Crossref
Johnson, M.M., Young, M.T., and Brusatte, S.L. 2020. The phylogenetics of Teleosauroidea (Crocodylomorpha, Thalattosuchia) and implications for their ecology and evolution. PeerJ 8: e9808. Crossref
Jonet, S. 1981. Contribution to the study of vertebrates from the Portuguese Cretaceous and especially from the Cenomanian of Extremadura. Comunicações dos Serviços Geológicos de Portugal 67: 10l–306.
Kellner, A.W.A., Azevedo, S.A.K., Machado, E.B., de Carvalho, L.B., and Henriques, D.D.R. 2011. A new dinosaur (Theropoda, Spinosauridae) from the Cretaceous (Cenomanian) Alcântara Formation, Cajual Island, Brazil. Anais da Academia Brasileira de Ciências 83 (1): 99–108. Crossref
Kullberg, J.C., Rocha, R.B., Soares, A.F., Rey, J., Terrinha, P., Azerêdo, A.C., Callapez, P., Duarte, L.V., Kullberg, M.C., Martins, L., Miranda, J.R., Alves, C., Mata, J., Madeira, J., Mateus, O., Moreira, M., and Nogueira, C.R. 2013. The Lusitanian Basin: stratigraphy, paleogeography and tectonics. In: R. Dias, A. Araújo, P. Terrinha, and J.C. Kullberg (eds.), Geologia de Portugal no contexto da Ibéria, 989–1141. Escolar Editora, Lisboa.
Kullberg, J.C., Rocha, R.B., Soares, A.F., Rey, J., Terrinha, P., Callapez, P., and Martins, L. 2006. The Lusitanian Basin: stratigraphy, paleogeography and tectonics. In: R. Dias, A. Araújo, P. Terrinha, and J.C. Kullberg (eds.), Geologia de Portugal no contexto da Ibérica, 317–368. Universidade de Évora, Évora.
Lapparent, A.F. and Zbyszewski, G. 1957. The dinosaurs from Portugal. Memoire Services Géologiques du Portugal 2 (nouvelle série): 1–131.
Larson, D.W. 2008. Diversity and variation of theropod dinosaur teeth from the uppermost Santonian Milk River Formation (Upper Cretaceous), Alberta: a quantitative method supporting identification of the oldest dinosaur tooth assemblage in Canada. Canadian Journal of Earth Sciences 45: 1455–1468. Crossref
Leinfelder, R.R. 1987. Multifactorial control of sedimentation patterns in an ocean marginal basin: the Lusitanian Basin (Portugal) during the Kimmeridgian and Tithonian. Geologische Rundschau 76: 599–631. Crossref
Li, F., Peng, G., Yong, Y., Jiang, S., and Huang, D. 2009. A new carnosaur from the Late Jurassic of Qianwei, Sichuan, China. Acta Geologica Sinica 83: 1203–1214.
Lockley, M.G., Meyer, C.A., and dos Santos, V.F. 1994. Trackway evidence for a herd of juvenile sauropods from the Late Jurassic of Portugal. Gaia 10: 27–35.
Long, J.A. and McNamara, K.J. 1997. Heterochrony. In: P.J. Currie and K. Padian (eds.), The Encyclopedia of Dinosaurs, 311–317. Academic Press, San Diego.
Lubbe, T., Richter, U., and Knötschke, N. 2009. Velociraptorine dromaeosaurid teeth from the Kimmeridgian (Late Jurassic) of Germany. Acta Palaeontologica Polonica 54: 401–408. Crossref
Maddison, W.P. and Maddison, D.R. 2011. Mesquite: A Modular System for Evolutionary Analysis. Version 2.75 [available online, http://mesquiteproject.org].
Madsen, J.H., Jr. 1976a. Allosaurus fragilis: a revised osteology. Utah Geological Survey Bulletin 109: 1–163.
Madsen, J.H., Jr. 1976b. A second new theropod dinosaur from the Late Jurassic of east central Utah. Utah Geology 3 (1): 51–60. Crossref
Madsen, J.H., Jr. and Welles, S.P. 2000. Ceratosaurus (Dinosauria, Theropoda). A revised osteology. Miscellaneous Publication, Utah Geological Survey 00-2: 1–80.
Maidment, S.C.R., Brassey, C., and Barrett, P.M. 2015. The postcranial skeleton of an exceptionally complete individual of the plated dinosaur Stegosaurus stenops (Dinosauria: Thyreophora) from the Upper Jurassic Morrison Formation of Wyoming, U.S.A. PLoS ONE 10: e0138352. Crossref
Maidment, S.C.R., Chapelle, K.E.J., Bonsor, J.A., Button, D., and Barrett, P. 2022. Osteology and relationship of Cumnoria prestwichii (Ornithischia: Ornithopoda) from the Late Jurassic of Oxfordshire, UK. Monograph of the Palaeontographical Society 176: 1–55. Crossref
Maidment, S.C.R., Raven, T.J., Ouarhache, D., and Barrett, P.M. 2020. North Africa’s first stegosaur: Implications for Gondwanan thyreophoran dinosaur diversity. Gondwana Research 77: 82–97. Crossref
Malafaia, E., Escaso, F., Mocho, P., Serrano-Martínez, A., Torices, A., Cachão, M., and Ortega, F. 2017a. Analysis of diversity, stratigraphic and geographical distribution of isolated theropod teeth from the Upper Jurassic of the Lusitanian Basin, Portugal. Journal of Iberian Geology 43: 257–291. Crossref
Malafaia, E., Gasulla, J.M., Escaso, F., Narvaéz, I., and Ortega, F. 2020a. An update of the spinosaurid (Dinosauria: Theropoda) fossil record from the Lower Cretaceous of the Iberian Peninsula: distribution, diversity, and evolutionary history. Journal of Iberian Geology 46: 431–444. Crossref
Malafaia, E., Mocho, P., Escaso, F., and Ortega, F. 2020b. A new carcharodontosaurian theropod from the Lusitanian Basin: evidence of allosauroid sympatry in the European Late Jurassic. Journal of Vertebrate Paleontology 40 (1): e1768106. Crossref
Malafaia, E., Mocho, P., Escaso, F., and Ortega, F. 2017b. A juvenile allosauroid theropod (Dinosauria, Saurischia) from the Upper Jurassic of Portugal. Historical Biology 29: 654–676. Crossref
Malafaia, E., Mocho, P., Escaso, F., and Ortega, F. 2017c. New data on the anatomy of Torvosaurus and other remains of megalosauroid (Dinosauria, Theropoda) from the Upper Jurassic of Portugal. Journal of Iberian Geology 43: 33–59. Crossref
Malafaia, E., Ortega, F., Escaso, F. and Silva, B. 2015. New evidence of Ceratosaurus (Dinosauria: Theropoda) from the Late Jurassic of the Lusitanian Basin, Portugal. Historical Biology 27: 938–946. Crossref
Mannion, P.D. and Calvo, J.O. 2011. Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian–early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: implications for titanosaur systematics. Zoological Journal of the Linnean Society 163: 155–181. Crossref
Mannion, P.D., Upchurch, P., Barnes, R.N., and Mateus, O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriformes. Zoological Journal of the Linnean Society 168: 98–206. Crossref
Mannion, P.D., Upchurch, P., Schwarz, D., and Wings, O. 2019. Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: phylogenetic and biogeographic implications for eusauropod dinosaur evolution. Zoological Journal of the Linnean Society 185: 784–909. Crossref
Manuppella, G., Antunes, M.T., Costa Almeida, C.A., Azerêdo, A.C., Barbosa, B., Cardoso, J.L., Crispim, J.A., Duarte, L.V., Henriques, M.H., Martins, L.T., Ramalho, M.M., Santos, V.F., and Terrinha, P. 2000. Geological Map of Portugal at Scale of 1:50 000: Explanatory Notice of the Sheet 27-A (Vila Nova de Ourém). Departamento de Geologia, Instituto Geológico e Mineiro, Lisboa.
Manuppella, G., Antunes, M.T., Pais, J., Ramalho, M.M., and Rey, J. 1999a. Geological Map of Portugal at Scale of 1:50 000: Explanatory Notice of the Sheet 30-A (Lourinhã). Departamento de Geologia, Instituto Geológico e Mineiro, Lisboa.
Manuppella, G., Antunes, M.T., Pais, J., Ramalho, M.M., and Rey, J. 1999b. Geological Map of Portugal at Scale of 1:50 000: Explanatory Notice of the Sheet 38-B (Setúbal). Departamento de Geologia, Instituto Geológico e Mineiro, Lisboa.
Manuppella, G., Zbyszewski, G., and Veiga Ferreira, O. 1978. Geological Map of Portugal at Scale of 1:50 000: Explanatory Notice of the Sheet 23-A (Pombal). Serviços Geológicos de Portugal, Lisboa.
Marsh, O.C. 1877a. New order of extinct Reptilia (Stegosauria) from the Jurassic of the Rocky Mountains. American Journal of Science 14 (Series 3): 513–514. Crossref
Marsh, O.C. 1877b. Notice of new dinosaurian reptiles from the Jurassic Formation. American Journal of Science 14 (Series 3): 514–516. Crossref
Marsh, O.C. 1878. Principal characters of American Jurassic dinosaurs. Part I. American Journal of Science 16 (Series 3): 411–416. Crossref
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Part V. American Journal of Science 21 (Series 3): 417–423. Crossref
Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs. Part VIII. The order Theropoda. American Journal of Science 27 (Series 3): 329–340. Crossref
Martin, T. and Krebs, B. 2000. Guimarota. A Jurassic Ecosystem. 155 pp.Verlag Dr. Friedrich Pfeil, München.
Mateus, O. 1998. Lourinhanosaurus antunesi, a new Upper Jurassic allosauroid (Dinosauria: Theropoda) from Lourinhã, Portugal. Memórias da Academia das Ciências de Lisboa 37: 111–124.
Mateus, O. 2005. Dinosaurs of the Upper Jurassic of Portugal, with Emphasis on Saurischians. 377 pp. Unpublished Ph.D. Thesis, Universidade Nova de Lisboa, Lisboa.
Mateus, O. and Antunes, M.T. 2000a. Ceratosaurus sp. (Dinosauria: Theropoda) in the Late Jurassic of Portugal. In: Abstracts, 31st International Geological Congress. Rio de Janeiro.
Mateus, O. and Antunes, M.T. 2000b. Torvosaurus sp. (Dinosauria: Theropoda) in the Late Jurassic of Portugal. In: Abstracts Book I Congresso Ibérico de Paleontología, XVI Jornadas de la Sociedad Española de Paleontología, 115–117. Universidade de Évora, Évora.
Mateus, O. and Estravíz-López, D. 2022. A new theropod dinosaur from the early Cretaceous (Barremian) of Cabo Espichel, Portugal: Implications for spinosaurid evolution. PLoS ONE 17: e0262614. Crossref
Mateus, O., Araújo, R., Natário, C., and Castanhinha, R. 2011. A new specimen of the theropod dinosaur Baryonyx from the early Cretaceous of Portugal and taxonomic validity of Suchosaurus. Zootaxa 2827: 54–68. Crossref
Mateus, O., Dinis, J., and Cunha, P. P. 2017. Upper Jurassic to Lowermost Cretaceous of the Lusitanian Basin, Portugal—landscapes where dinosaurs walked. Ciências da Terra 34: 618–634. Crossref
Mateus, O., Maidment, S.C.R., and Christiansen, N.A. 2009. A new long-necked “sauropod-mimic” stegosaur and the evolution of the plated dinosaurs. Proceedings of the Royal Society of London Series B 276: 1815–1821. Crossref
Mateus, O., Mannion, P.D., and Upchurch, P. 2014. Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology 34: 618–634. Crossref
Mateus, O., Walen, A., and Antunes, M.T. 2006. The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. New Mexico Museum of Natural History and Science Bulletin 36: 123–129.
Mocho, P., Royo-Torres, R., and Ortega, F. 2014. Phylogenetic reassessment of Lourinhasaurus alenquerensis, a basal Macronaria (Sauropoda) from the Upper Jurassic of Portugal. Zoological Journal of the Linnean Society 170: 875–916. Crossref
Mocho, P., Royo-Torres, R., and Ortega, F. 2017a. New data of the Portuguese brachiosaurid Lusotitan atalaiensis (Sobral Formation, Upper Jurassic). Historical Biology 29: 789–817. Crossref
Mocho, P., Royo-Torres, R., and Ortega, F. 2019. A new macronarian sauropod from the Upper Jurassic of Portugal. Journal of Vertebrate Paleontology 39 (1): e1578782. Crossref
Mocho, P., Royo-Torres, R., Escaso, F., Malafaia, E., de Miguel Chaves, C., Narváez, I., Pérez-García, A., Pimentel, N., Silva, B.C., and Ortega, F. 2017b. Upper Jurassic sauropod record in the Lusitanian Basin (Portugal): Geographical and lithostratigraphical distribution. Palaeontologia Electronica 20.2.27A: 1–50. Crossref
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., and Ortega, F. 2016. Systematic review of Late Jurassic sauropods from the Museu Geológico collections (Lisboa, Portugal). Journal of Iberian Geology 42: 227–250.
Montenat, C., Guery, F., and Jamet, M. 1988. Mesozoic evolution of the Lusitanian Basin: comparison with the adjacent margin. In: G. Boillot, E.L. Winterer et al. (ed.), Proceedings of the Ocean Drilling Program, Scientific Results 103: 757–775. Crossref
Nopcsa, F. 1915. The dinosaurs of the Transylvanian parts of Hungary. Mitteilungen aus dem Jahrbuche der Kgl. Ungarischen Geologischen Reichanstalt 23: 1–24.
Novas, F.E., de Valais, S., Vickers-Rich, P., and Rich, T. 2005. A large Cretaceous theropod from Patagonia, Argentina, and the evolution of carcharodontosaurids. Naturwissenschaften 92: 226–230. Crossref
Oliveira, J.T., Pereira, H., Ramalho, M., and Antunes, M.T. 1992. Geological Map of Portugal, at the Scale of 1:500 000. Serviços Geológicos de Portugal, Lisboa.
Ortega, F., Escaso, F., Gasulla, J.M., Dantas, P., and Sanz, J.L. 2006. Dinosaurs of the Iberian Peninsula. Estudios Geológicos 62 (1): 219–240. Crossref
Ortega, F., Malafaia, E., Escaso, F., Pérez-García, A., and Dantas, P. 2009. Faunas of reptiles from the Upper Jurassic of Portugal. Paleolusitana 1: 43–56.
Osborn, H.F. and Mook, C.C. 1921. Camarasaurus, Amphicoelias, and other sauropods of Cope. Memoirs of the American Museum of Natural History New Series 3: 249–387.
Owen, R. 1842. Report on British fossil reptiles. Part II. Reports of the British Association for the Advancement of Sciences 11: 60–204.
Paul, G.S. 1988. Predatory Dinosaurs of the World: A Complete Illustrated Guide. 464 pp. Simon & Schuster, New York.
Pena dos Reis, R.P.B., Dinis, J.L., Cunha, P.M.R., and Trincão, P. 1996. Upper Jurassic sedimentary infill and tectonics of the Lusitanian Basin (western Portugal). In: A.C. Riccardi (ed.), Advances in Jurassic Research, GeoResearch Forum 1–2, 377–386. Transtec Publications, Zurich.
Pena dos Reis, R.P.B., Pimentel, N.L., and Garcia, A.J.V. 2011. The Lusitanian Basin, stratigraphic analysis and geodynamic evolution. Boletim de Geociencias Petrobras 19: 23–52.
Pena dos Reis, R.P.B., Proença Cunha, C.P., Dinis, J.L., and Trincão, P.R. 2000. Geologic evolution of the Lusitanian Basin (Portugal) during the late Jurassic. GeoResearch Forum 6: 345–356.
Pérez-Moreno, B.P., Chure, D.J., Pires, C., Silva, C.M.D., Santos, V.D., Dantas, P., Póvoas, L., Cachão, M., Sanz, J.L., and Galopim de Carvalho, A.M. 1999. On the presence of Allosaurus fragilis (Theropoda: Carnosauria) in the Upper Jurassic of Portugal: first evidence of an intercontinental dinosaur species. Journal of the Geological Society 156: 449–452. Crossref
Rasmussen, E.S., Lomholt, S., Andersen, C., and Vejbaek, O.V. 1998. Aspects of the structural evolution of the Lusitanian Basin in Portugal and the shelf and slope area offshore Portugal. Tectonophysics 300: 199–225. Crossref
Rauhut, O.W.M. 2004. Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia. Journal of Vertebrate Paleontology 24: 894–902. Crossref
Rauhut, O.W.M. 2005. Osteology and relationships of a new theropod dinosaur from the Middle Jurassic of Patagonia. Palaeontology 48: 87–110. Crossref
Rauhut, O.W.M. 2011. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Special Papers in Palaeontology 86: 195–239.
Rauhut, O.W.M. and Carrano, M.T. 2016. The theropod dinosaur Elaphrosaurus bambergi Janensch, 1920, from the Late Jurassic of Tendaguru, Tanzania. Zoological Journal of the Linnean Society 178: 546–610. Crossref
Rauhut, O.W.M. and López-Arbarello, A. 2009. Considerations on the age of the Tiouaren Formation (Iullemmeden Basin, Niger, Africa): Implications for Gondwanan Mesozoic terrestrial vertebrate faunas. Palaeogeography, Palaeoclimatology, Palaeoecology 271: 259–267. Crossref
Rauhut, O.W.M. and Pol, D. 2019. A probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs. Scientific Reports 9: 18826. Crossref
Rauhut, O.W.M., Foth, C., Tischlingerd, H., and Norell, M.A. 2012. Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany. PNAS 109: 11746–11751. Crossref
Rauhut, O.W.M., Hübner, T.R., and Lanser, K.-P. 2016. A new megalosaurid theropod dinosaur from the late Middle Jurassic (Callovian) of north-western Germany: Implications for theropod evolution and faunal turnover in the Jurassic. Palaeontologia Electronica 19.2.26A: 1–65. Crossref
Rauhut, O.W.M., Piñuela, L., Castanera, D., García-Ramos, J.C., and Cela, I.S. 2018. The largest European theropod dinosaurs: remains of a gigantic megalosaurid and giant theropod tracks from the Kimmeridgian of Asturias, Spain. PeerJ 6: e4963. Crossref
Rauhut, O.W.M., Schwermann, A.H., Hübner, T.R., and Lanser, K.-P. 2020. The oldest record of the genus Torvosaurus (Theropoda: Megalosauridae) from the Callovian Ornatenton Formation of north-western Germany. Geologie und Paläontologie in Westfalen 93: 1–13.
Rey, J. 1972. Geological research on the Lower Cretaceous of Extremadura (Portugal). Mémoires du Service géologique du Portugal 21: 1–477.
Rey, J. 1992. The Lower Cretaceous lithostratigraphic units of the Lisbon region. Comunicações dos Serviços Geológicos de Portugal 78: 103–24.
Rey, J. 1999. Lower Cretaceous Depositional Sequences in the Cascais Area. In: European Palaeontological Association Workshop, Field Trip A, 1–57. Lisboa.
Rey, J., Graciansky, P.C. de, and Jacquin, T. 2003. Depositional sequences in the Lower Cretaceous of the Lusitano Basin. Comunicações del Instituto Geologico e Minero 90: 15–42.
Ribeiro, A., Silva, J.B., Cabral, J., Dias, R., Fonseca, P., Kullberg, M.C., Terrinha, P., and Kullberg, J.C. 1996. Tectonics of the Lusitanian Basin. Final Report, Proj. MILUPOBAS, ICTE/GG/GeoFCUL. Lisboa.
Rocha, R.B. and Soares, A.F. 1984. Some thoughts on Jurassic sedimentation in the western Mesocenozoic rim of Portugal. Memórias e Notícias 97: 133–142.
Rocha, R.B., Manuppella, G., Mouterde, R., Ruget, C., and Zbyszewski, G. 1981. Geological Map of Portugal at the Scale of 1:50 000: Explanatory News of the Sheet 19-C (Figueira da Foz). Serviços Geológicos de Portugal, Lisboa.
Rocha, R.B., Marques, B.L., Kullberg, J.C., Caetano, P.C., Lopes, C., Soares, A.F., Duarte, L.V., Marques, J.F. and Gomes, C.R. 1996. The 1st and 2nd rifting phases of the Lusitanian Basin: stratigraphy, sequence analysis and sedimentary evolution. Final Report C.E.C. Proj. MILUPOBAS, Lisboa.
Rotatori, F.M., Moreno-Azanza, M., and Mateus, O. 2022. Reappraisal and new material of the holotype of Draconyx loureiroi (Ornithischia: Iguanodontia) provide insights on the tempo and modo of evolution of thumb-spiked dinosaurs. Zoological Journal of the Linnean Society 195: 125–156. Crossref
Royo-Torres, R., Cobos, A., and Alcalá, L. 2006. A giant European dinosaur and a new sauropod clade. Science 314: 1925–1927. Crossref
Royo-Torres, R., Upchurch, P., Mannion, P.D., Mas, R., Cobos, A., Gascó, F., Alcalá, L., and Sanz, J.L. 2014. The anatomy, phylogenetic relationships and stratigraphic position of the Tithonian–Berriasian Spanish sauropod dinosaur Aragosaurus ischialus. Zoological Journal of the Linnean Society 71: 623–655. Crossref
Ruiz-Omeñaca, J.I., Piñuela, P., García-Ramos, J.C., and Canudo, J.I. 2008. Carnivorous dinosaur teeth (Saurischia; Theropoda) from the Late Jurassic of Asturias, north Spain. In: Huerta Hurtado, P. and Fernández-Baldor, F.T. (eds,), Actas de las IV Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno Salas de los Infantes, Burgos, 273–291. Colectivo Arqueológico y Paleontológico de Salas, Salas de los Infantes.
Sadleir, R., Barrett, P.M., and Powell, H.P. 2008. The anatomy and systematics of Eustreptospondylus oxoniensis, a theropod dinosaur from the middle Jurassic of Oxfordshire, England. Monograph of the Palaeontographical Society 160: 1–82. Crossref
Salgado, L., Coria, R.A., and Calvo, J.O. 1997. Evolution of titanosaurid sauropods. I: phylogenetic analysis based on the postcranial evidence. Ameghiniana 34: 3–32.
Samathi, A., Sander, P.M., and Chanthasit, P. 2021. A spinosaurid from Thailand (Sao Khua Formation, Early Cretaceous) and a reassessment of Camarillasaurus cirugedae from the Early Cretaceous of Spain. Historical Biology 33: 3480–3494. Crossref
Santos, V.F. 2003. Dinosaur Tracks in the Jurassic and Cretaceous of Portugal. Paleobiological and Paleoecological Considerations. 365 pp. Unpublished Ph.D. Thesis, Facultad de Ciencias, Universidad Autónoma de Madrid, Madrid.
Santos, V.F. 2008. Dinosaur Footprints from Portugal. 124 pp. Museu Nacional de História Natural, Lisboa.
Seeley, H.G. 1887. On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London 43: 165–171. Crossref
Sauvage, H.E. 1896. Crocodilians and dinosaurians from the Mesozoic of Portugal. Bulletin de la Société géologique de France 24: 46–49.
Sauvage, H.E. 1897–1898. Fossil vertebrates from Portugal. Contributions to the study of fish and reptiles from the Jurassic and Cretacic. Direction des Travaux Géologiques du Portugal 29: 1–46.
Sereno, P.C. 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 210: 41–83. Crossref
Sereno, P.C., Beck, A.L., Dutheil, D.B., Gado, B., Larsson, H.C.E., Lyon, G.H., Marcot, J.D., Rauhut, O.W.M., Sadleir, R.W., Sidor, C.A., Varricchio, D.D., Wilson, G.P., and Wilson, J.A. 1998. A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids. Science 282: 1298–1302. Crossref
Serrano-Martínez, A., Vidal, D., Sciscio, L., Ortega, F., and Knoll, F. 2015. Isolated theropod teeth from the Middle Jurassic of Niger and the early dental evolution of Spinosauridae. Acta Palaeontologica Polonica 61: 403–415. Crossref
Schade, M., Rauhut, O.W.M., Foth, C., Moleman, O., and Evers, S.W. 2023. A reappraisal of the cranial and mandibular osteology of the spinosaurid Irritator challengeri (Dinosauria: Theropoda). Palaeontologia Electronica 26: a17. Crossref
Schubert, B.W. and Ungar, P.S. 2005. Wear facets and enamel spalling in tyrannosaurid dinosaurs. Acta Palaeontologica Polonica 50: 93–99.
Smith, J.B., Lamanna, M.C., Mayr, H., and Lacovara, K.J. 2006. New information regarding the holotype of Spinosaurus aegyptiacus Stromer, 1915. Journal of Paleontology 80: 400–406. Crossref
Smith, J.B., Vann, D.R., and Dodson, P. 2005. Dental morphology and variation in theropod dinosaurs: implications for the taxonomic identification of isolated teeth. The Anatomical Record 285A: 699–736. Crossref
Soto, M. and Perea, D. 2008. A ceratosaurid (Dinosauria, Theropoda) from the Late Jurassic–Early Cretaceous of Urugay. Journal of Vertebrate Paleontology 28: 439–444. Crossref
Sues, H.-D., Frey, E., Martill, D.M., and Scott, D.M. 2002. Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil. Journal of Vertebrate Paleontology 22: 535–547. Crossref
Taquet, P. and Welles, S.M. 1977. Redescription du crane de dinosaure théropode de Dives (Normandie). Annales de Paléontologie 63: 191–206.
Taylor, A.M., Gowland, S., Leary, S., Keogh, K.J., and Martinius, A.W. 2014. Stratigraphical correlation of the Lourinhã Formation in the Consolação Sub-basin (Lusitanian Basin), Portugal. Geological Journal 49: 143–162. Crossref
Teixeira, C., Zbyszewski, G., Torres de Assunção, C., and Manuppell, G. 1968. Geological Map of Portugal at the Scale of 1:50 000: Explanatory News of the Sheet 23-C (Leiria). Serviços Geológicos de Portugal, Lisboa.
Tschopp, E., Mateus, O., and Benson, R. 2015. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ 3: e857. Crossref
Upchurch, P. 1995. Evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London B 349: 365–390. Crossref
Upchurch, P., Barrett, P.M., and Dodson, P. 2004. Sauropoda. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, Second Editon, 259–322. University of California Press, Oakland. Crossref
Upchurch, P., Mannion, P.D., and Taylor, M.P. 2015. The anatomy and phylogenetic relationships of “Pelorosaurus” becklesii (Neosauropoda, Macronaria) from the Early Cretaceous of England. PLoS ONE 10: e0125819. Crossref
Verrière, A., Fröbisch, N.B., and Fröbisch, J. 2022. Regionalization, constraints, and the ancestral ossification patterns in the vertebral column of amniotes. Scientific Reports 12: 1–12. Crossref
Viera, L.I. and Torres, J.A. 1995. The presence of Baryonyx walkeri (Saurischia, Theropoda) in the Weald of La Rioja (Spain). Nota previa. MUNIBE 47: 57–61.
Vullo, R., Abit, D., Ballévre, M., Billon-Bruyat, J.-P., Bourgeais, R., Buffetaut, É., Daviero-Gomez, V., Garcia, G., Gomez, B., Mazin, J.-M., Morel, S., Néraudeau, D., Pouech, J., Rage, J.-C., Schnyder, J., and Tong, H. 2014. Palaeontology of the Purbeck-type (Tithonian, Late Jurassic) bonebeds of Chassiron (Oléron Island, western France). Comptes Rendus Palevol 13: 421–441. Crossref
Walker, A.D. 1970. A revision of the Jurassic reptile Hallopus victor (Marsh), with remarks on the classification of crocodiles. Philosophical Transactions of the Royal Society of London 257 (Series B): 323–372. Crossref
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 217–276. Crossref
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology 18 (Supplement 2): 1–68. Crossref
Wilson, R.C.L. 1988. Mesozoic development of the Lusitanian Basin, Portugal. Revista de la Sociedad Geológica Española 1: 393–407.
Wilson, R.C.L. 1975. Atlantic opening and Mesozoic continental margin basins of Iberia. Earth and Planetary Science Letters 25: 33–43. Crossref
Young, C.M.E., Hendrickx, C., Challands, T.J., Foffa, D., Ross, D.A., Butler, I.B., and Brusatte, S.L. 2019. New theropod dinosaur teeth from the Middle Jurassic of the Isle of Skye, Scotland. Scottish Journal of Geology 55: 7–19. Crossref
Young, M.T. and Andrade, M.B. 2009. What is Geosaurus? Redescription of Geosaurus giganteus (Thalattosuchia, Metriorhynchidae) from the Upper Jurassic of Bayern, Germany. Zoological Journal of the Linnean Society 157: 551–585. Crossref
Young, M.T., Andrade, M.B., Cornée, J.-J., Steel, L., and Foffa, D. 2014. Re-description of a putative Early Cretaceous “teleosaurid” from France, with implications for the survival of metriorhynchids and teleosaurids across the Jurassic–Cretaceous Boundary. Annales de Paléontologie 100: 165–174. Crossref
Zbyszewski, G. 1946. Bones of Omosaurus discovered near Baleal (Peniche). Comunicações dos Serviços Geológicos de Portugal 28: 135–144.
Zbyszewski, G. and Moitinho de Almeida, F. 1960. Geological Map of Portugal at the Scale of 1:50 000: Explanatory News of the Sheet 26-D (Caldas da Rainha). Serviços Geológicos de Portugal, Lisboa.
Zinke, J. 1998. Small theropod teeth from the Upper Jurassic coal mine of Guimarota (Portugal). Paläontologische Zeitschrift 72: 179–189. Crossref
Zittel, K.A. 1887–1890. Handbuch der Paläontologie 1. Abteilung: Paläozoologie 3. 900 pp. Druck und Verlag von R. Oldenbourg, München.
Acta Palaeontol. Pol. 69 (2): 127–171, 2024
https://doi.org/10.4202/app.01113.2023