
A new Late Ordovician bubble-headed trilobite species from South West Wales and its implications for biostratigraphy
MANUS G. LEIDI, LUCY M.E. MCCOBB, PATRICK D. MCDERMOTT, and ALAN W. OWEN
Leidi, M.G., McCobb, L.M.E., McDermott, P.D., and Owen, A.W. 2024. A new Late Ordovician bubble-headed trilobite species from South West Wales and its implications for biostratigraphy. Acta Palaeontologica Polonica 69 (1): 57–72.
A new species of bubble-headed trilobite, Staurocephalus oliveae sp. nov., is described from 250 specimens collected from a recently discovered locality in the Sholeshook Limestone Formation (upper Katian, Upper Ordovician; Ashgill, Cautleyan Zone 1 equivalent) in the village of Llanddowror, Carmarthenshire. Although no fully articulated specimens were found, the material is the most abundant and well-preserved of any Ordovician Staurocephalus species known from Britain and Ireland, and enables the complicated 3D structure of its cephalon to be reconstructed. Staurocephalus oliveae sp. nov. is the earliest representative of the genus recorded in Avalonia, following its presumed migration into the area from Laurentia or Baltica. Recognition of this Cautleyan species resurrects the potential usefulness of the long established, widely distributed, Staurocephalus clavifrons as a latest Katian (= Rawtheyan) indicator.
Key words: Trilobita, Phacopida, Encrinuridae, Staurocephalus, Late Ordovician, biostratigraphy, Wales, Great Britain.
Manus G. Leidi [LeidiM@cardiff.ac.uk; ORCID: https://orcid.org/0009-0009-8726-5784], School of Biosciences, Cardiff University, Cardiff CF10 3AX, Wales, UK.
Lucy M.E. McCobb [lucy.mccobb@museumwales.ac.uk; ORCID: https://orcid.org/0000-0002-8035-3337 ] (corresponding author), Department of Natural Sciences, Amgueddfa Cymru-National Museum Wales, Cardiff CF10 3NP, Wales, UK.
Patrick D. McDermott [ rock_nibbler@outlook.com], Flat 4, 12 Quay St., Carmarthen, SA31 3JT, Wales, UK.
Alan W. Owen [alan.owen@glasgow.ac.uk; ORCID: https://orcid.org/0000-0002-6558-5498 ], School of Geographical and Earth Sciences, University of Glasgow, Glasgow G12 8QQ, Scotland, UK.
Received 9 November 2023, accepted 5 February 2024, published online 6 March 2024.
Copyright © 2024 M.G. Leidi et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Staurocephalus was a long-ranging genus of “bubble-headed” trilobite (Phacopida, Encrinuridae), with the two oldest known species recorded from the lower Katian (uppermost Caradoc; Upper Ordovician) of Maine, USA (Owen and Bruton 2012) and the Oslo Region of Norway (Owen and Bruton 1980). It was an Ashgill (late Katian) immigrant from Laurentia or Baltica into the Anglo-Welsh area and Ireland and was the only member of the Staurocephalinae to survive into the Silurian (Owen 2007; Adrain 2013: 308). One species, S. clavifrons Angelin, 1854, has long been used as a marker for the uppermost Katian and possibly Hirnantian (both Upper Ordovician) in Sweden (Jaanusson 1963, Nielsen et al. 2023), Denmark and Poland (Kielan 1960) but its application has been questioned on the basis of occurrences of Staurocephalus in Britain (Price 1973b). The material described herein provides some resolution to this matter.
A locality discovered by one of us (PDMcD) around 20 years ago at Talfan Farm near the village of Llanddowror, Carmarthenshire, South West Wales (Fig. 1; loc. 6 of Cocks 2014) has yielded a large number of specimens of a new species of Staurocephalus from the lower part of the Sholeshook Limestone Formation. The formation is largely correlated with the Cautleyan to lowest Rawtheyan stages of the Anglo-Welsh regional Ashgill Series (Fig. 2). The base of the Ashgill lies somewhere in the middle of the international Katian Stage (see Bergström et al. 2009) and thus the Sholeshook Limestone is upper Katian in global terms. The lowest part of the formation at Pengawse Hill near Whitland (see Fig. 1C) contains conodonts of the Amorphognathus ordovicicus Biozone (Ferretti et. al. 2014) and chitinozoans from the upper part of the Fungochitina spinifera Biozone (Vandenbroucke et al. 2008). As noted by Vandenbroucke et al. (2008: 209), the latter supports the correlation close to the Pusgillian/Cautleyan boundary (upper Katian) indicated by the trilobites (Zalasiewicz et al. 1995). The formation at Pengawse Hill overlies shales of the Mydrim Shales Formation with graptolites of the “Normalograptus proliferation interval”, the upper part of which might extend into the Pleurograptus linearis Biozone (Vandenbroucke et al. 2008: 209). As noted below, the trinucleid trilobite Tretaspis moeldenensis moeldenensis Cave, 1960, is present in the section at Talfan Farm indicating an earliest Cautleyan age for the occurrence of Staurocephalus at Talfan Farm, possibly slightly younger than the base of the formation at Pengawse Hill. It is the oldest recorded occurrence of the genus from Avalonia.
Although there are no complete exoskeletons and the very convex cephalon is subject to varying degrees of deformation, the Sholeshook Limestone specimens of Staurocephalus oliveae sp. nov. provide the most complete information on the morphology of any of the Ordovician occurrences of the genus in Britain and Ireland and, with the possible exception of S. clavifrons, the most complete of any Ordovician species of Staurocephalus so far described.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:BAF4499F-2ED3-4DB8-AE0C-3A3F58A5A280
Institutional abbreviations.—BIRUG, Lapworth Museum of Geology, University of Birmingham, UK; GLAHM, Hunterian Museum, University of Glasgow, UK; NMW, National Museum Wales, Cardiff, UK; SM, Sedgwick Museum of Earth Sciences, University of Cambridge, UK.
Other abbreviations.—exsag., exsagittal; L, lateral glabellar lobe; max., maximum; S, lateral glabellar furrow; sag., sagittal; tr., transverse.
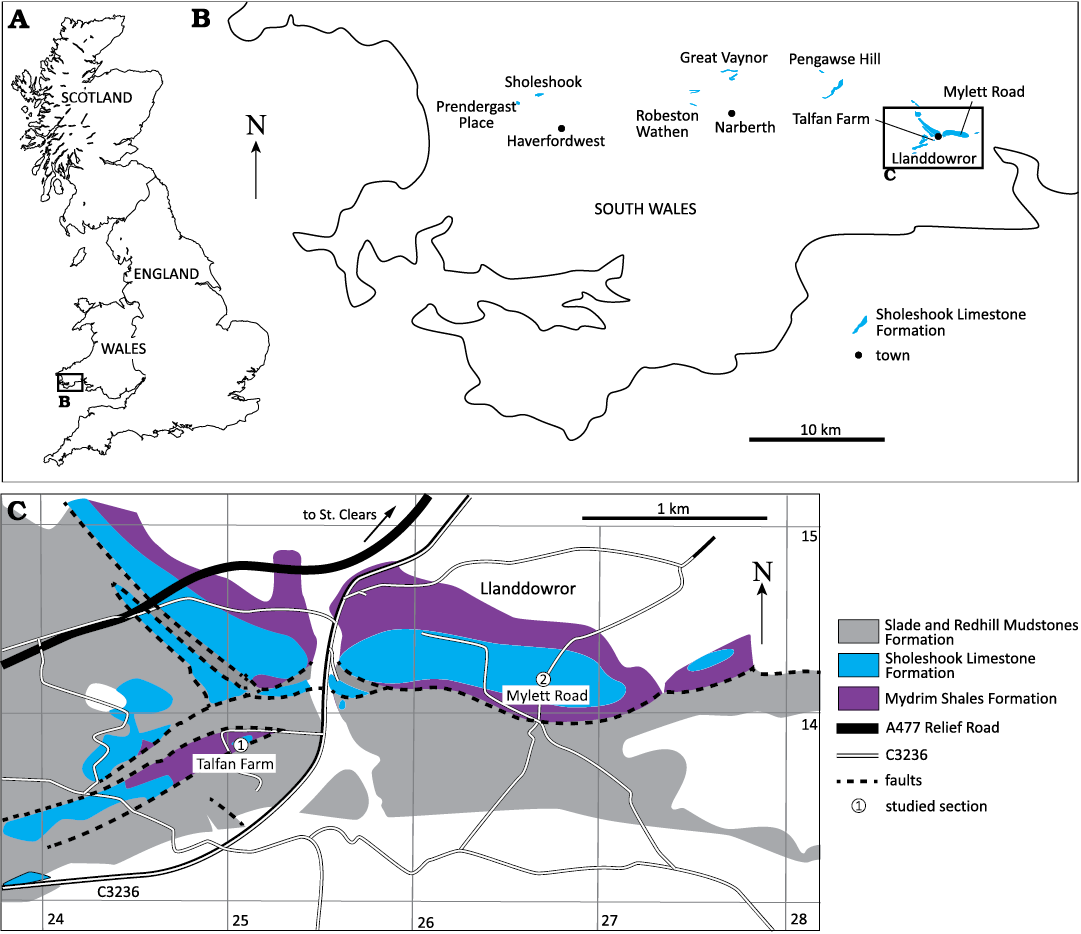
Fig. 1. Maps showing the location of the Talfan Farm fossil locality in Great Britain (A) and South West Wales (B). C. Geological map of the area around Llanddowror (greatly revised from BGS 1:50 000 Sheet 229 (Carmarthen), 1975 and Price (1973a: fig. 4) on the basis of recent field observations by PDMcD. Numbers on the x and y axis are co-ordinates (eastings and northings) on the UK National Grid. Talfan Farm is located at latitude 51°47’41’’N and longitude 4°32’27’’W.
Geological setting
The Sholeshook Limestone Formation is an upper Katian (Upper Ordovician) rock unit a little over 60 m thick in its most complete exposure near Sholeshook, comprising calcareous mudstones, siltstones and sandstones intercalated with some purer limestone horizons (Price 1973a; Owens 2000). It crops out in three main areas in South West Wales within a 30 km, roughly west-east trending band: at Sholeshook and Prendergast in the Haverfordwest area, at Robeston Wathen, and around the village of Llanddowror near St. Clears, south-west of Carmarthen (Fig. 1A–C). This study is based largely on fossils collected from the lower part of the formation at Talfan Farm near Llanddowror (Fig. 1D). In this area, the Sholeshook Limestone is represented by basal calcareous mudstones, which gradually coarsen upwards into coarse calcareous sandstone that makes up the bulk of the formation (Price 1973a). The majority of specimens at Talfan Farm were collected from a vertical section from three horizons in the basal Sholeshook Limestone at 1, 2, and 3 m above the boundary with the underlying Mydrim Shales Formation. A small number of additional specimens were from two spot samples, namely a deep 1 m diameter circular pit and a 1×2 m trench, both of which exposed the top of the Mydrim Shales Formation and the basal 1–3 m of the Sholeshook Limestone Formation. All five sampled horizons yielded very similar fossil faunas.
Marr and Roberts (1885) introduced the term “Sholeshook Limestone”, and Marr (1907) placed the formation at the base of his newly proposed Ashgill Series, though a subsequent revision (Ingham and Wright 1970) redefined the base of the series at a lower horizon. The Sholeshook Limestone has long been recognized for its diverse trilobite fauna, with Price (1974, 1977, 1980b, 1981) documenting in detail over fifty species. Price (1973a, 1980a) established the age of the Sholeshook Limestone Formation based on correlation of the trilobite species present with those in the shelly biozones in the type area for the Ashgill Series, the Cautley and Dent areas of northern England (Ingham 1966; Ingham and Wright 1970, 1972), with particular emphasis on the sequence of species of the trinucleid trilobite Tretaspis. The Upper Ordovician chitinozoan biozonal scheme established by Vandenbroucke (2008) for the largely siliciclastic successions of England and Wales incorporates the Ashgill zones documented in northern England by Vandenbroucke et al. (2005), which in turn, enhance the potential for international correlation of many of the shelly zones (Fig. 2). Apart from the basal few metres at Pengawse Hill (Vandenbroucke et al. 2008), chitinozoans have not been sampled from the Sholeshook Limestone.
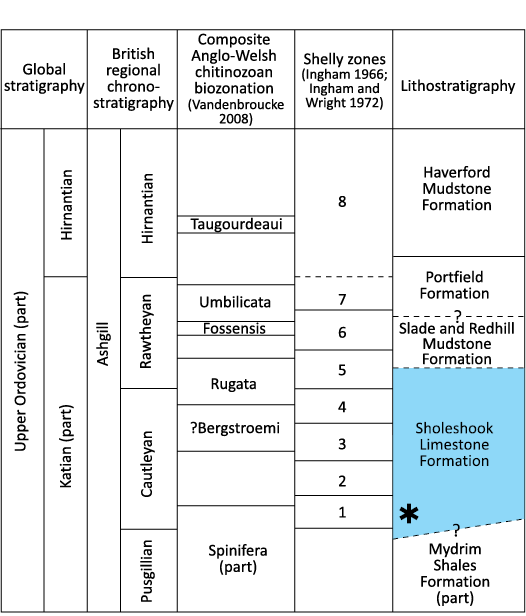
Fig. 2. Stratigraphical column showing the age of the Sholeshook Limestone Formation (in blue) and the underlying and overlying formations together with the shelly and chitinozoan biozonations of the Anglo-Welsh succession. Asterisk shows approximate age of the Talfan Farm locality, in the lower part of the formation (equivalent to Cautleyan Zone 1).
The basal Sholeshook Limestone was correlated with the upper part of Cautleyan Zone 1 of Ingham (1966) by Price (1973a) with the top of the formation having a low Rawtheyan (Zone 5, upper Katian) age (Price 1980a; Fig. 2).
Price (1973a: 243) recognized a stratigraphical gap between the Cautleyan (upper Zone 1) base of the Sholeshook Limestone and the underlying Mydrim Shales Formation at Mylet Road, Llanddowror (Fig. 1C), with the latter placed in the Caradoc Onnian Stage (now substage) (lower Katian) by Cave (1960) based on the graptolite fauna found in its highest beds. Hence, there appears to be a hiatus in the Mylet Road succession encompassing the Ashgill Pusgillian Stage and likely also parts of the Onnian and Cautleyan Zone 1 (Owens 2000: 159–161). However, further west at Pengawse Hill, Whitland (Fig. 1C), Zalasiewicz et al. (1995) used graptolite evidence to show a continuous succession through the top of the Mydrim Shales Formation and basal Sholeshook Limestone, with no apparent gap, and they suggested that the base of the Sholeshook Limestone may range down into the Pusgillian, a correlation subsequently supported by Vandenbroucke et al. (2008: 209). Hence, it appears that the base of the Sholeshook Limestone Formation is diachronous. The contact with the overlying Slade and Redhill Mudstone Formation (Fig. 2) is conformable, with some trilobite species ranging through the boundary. Price (1973a) argued that the base of the Slade and Redhill Mudstone Formation also appears to be diachronous, and part of the formation may be contemporaneous with the Sholeshook Limestone in its western outcrops although we consider that this may not prove to be the case.
Based on sedimentological evidence and the trilobite genera present, Price (1980b) interpreted the depositional setting of the Sholeshook Limestone as a relatively high energy environment, somewhere on the middle to upper part of the slope between the platform edge and basin. A diverse non-trilobite fauna has also been reported from the formation (e.g., Paul 1973, 1982, 1997; Donovan and Paul 1985; Barrs 2013; Ferretti et al. 2014; Cocks 2014) and the Talfan Farm locality has yielded abundant echinoderms (crinoids, cystoids, and coronates; Lanc et al. 2015; McDermott and Paul 2015) and brachiopods (Cocks 2014), along with rarer corals (Peel and McDermott 2016), bryozoans, gastropods, cephalopods, and ostracods. The presence of the trinucleid trilobite Tretaspis moeldenensis moeldenensis indicates an early Cautleyan (Zone 1) age.
Price (1973a, 1980b) recorded rare occurrences of Staurocephalus clavifrons (later S. cf. clavifrons) from various levels throughout the Sholeshook Limestone Formation, and from the basal Slade and Redhill Mudstone Formation (see below).
Material and methods
Staurocephalus oliveae sp. nov. is described based on 250 specimens recently collected, with permission, from Talfan Farm and now housed in the Palaeontology collections of the National Museum of Wales, Cardiff. The collection encompasses all sclerites apart from the hypostome. No complete articulated exoskeletons were recovered and most of the highly convex cranidial specimens have undergone some degree of deformation or crushing, but the large number of specimens available collectively provides a comprehensive view of the original morphology of the species. The majority of specimens are preserved as internal moulds, with small numbers of external moulds and internal/external mould part/counterpart pairs.
Specimens from the National Museum of Wales collections were whitened with ammonium chloride before photographing using a Canon 80D camera attached to a Leica Z6 microscope; those from other museum collections were photographed unwhitened in accordance with the loan conditions of those institutions. Multiple images were taken of each specimen at different depths of field, which were then stacked to produce composite images using Helicon Focus software. Photographs were edited and photographic figures compiled using Adobe Photoshop Elements 2022. Some external moulds were cast in latex to reveal details of the external sculpture and were consolidated prior to casting using a 5% w/w solution of Paraloid B-72 adhesive in Acetone.
Systematic palaeontology
Class Trilobita Walch, 1771
Order Phacopida Salter, 1864
Suborder Cheirurina Harrington & Leanza, 1957
Family Encrinuridae Angelin, 1854
Subfamily Staurocephalinae Prantl & Přibyl, 1947
Remarks.—Although given full family rank by many earlier workers (e.g., Kielan 1957; Evitt and Tripp 1977; Holloway 1980; Owen and Bruton 1980; Tomczykowa 1987), the concept of Staurocephalidae was largely based on the distinctive “bubble headed” morphology of derived members of the group. We concur with Adrain (2013: 307–308) in regarding Staurocephalinae as a subfamily of Encrinuridae, on the basis of shared morphological traits of protaspid and early meraspid stages of less derived staurocephalines such as Libertella, encrinurines, and “advanced” cybelines (see Edgecombe et al. 1988).
Genus Staurocephalus Barrande, 1846
Type species: Staurocephalus murchisoni Barrande, 1846, from the Liteň Formation (Wenlock) of Bohemia, Czech Republic.
Emended diagnosis.—Glabella with subparallel-sided posterior portion bearing short S1 and S2; S3 extends across glabella; frontal lobe greatly inflated and overhanging anterior margin of cephalon. Anterior part of rostral plate expanded and continuous with frontal glabellar lobe; short fixigenal spine present in most species. Thorax of 10 segments. Pygidium with 3 pairs of spines. Emended from Curtis and Lane (1997: 50)
Staurocephalus oliveae sp. nov.
Figs. 3, 4, 5, ?6A.
non1885 Staurocephalus globiceps Portlock; Marr and Roberts 1885: 481 [= Staurocephalus cf. clavifrons].
?1938 Staurocephalus cf. murchisoni Barrande; Whittington 1938: 451.
?1973 Staurocephalus clavifrons Angelin; Price 1973a: tables 1–3, non 7 [= Staurocephalus cf. clavifrons].
?1980 Staurocephalus cf. clavifrons Angelin; Price 1980b: 866 (Sholeshook Limestone specimens only), pl. 110: 18.
ZooBank LSID: urn:lsid:zoobank.org:act:99371F20-B988-498A-81E0- 55729B248609
Etymology: After Olive Potts (St. Clears, Carmarthenshire, UK), who discovered the first specimen of this species.
Type material: Holotype: almost complete cephalon, NMW 2013.11G.1 (Fig. 3A). Paratypes: almost complete cephalon with eyes preserved, NMW 2013.11G.2 (Fig. 3B); one further cephalon, NMW 2013.11G.8 (Fig. 3E); three cranidia, NMW 2013.11G.6, 7, 9 (Fig. 3C, D, F); three librigenae, NMW 2013.11G.3, 10, 12 (Fig. 4B–D); three pygidia, NMW 2013.11G.4, 11, 15 (Fig. 4F, G, H); one rostral plate, NMW 2013.11G.5 (Fig. 4A); one thoracic segment, NMW 2013.11G.13 (Fig. 4E). All from the type locality and horizon.
Type locality: Talfan farm, Llanddowror, Carmarthenshire, Wales, UK.
Type horizon: Sholeshook Limestone Formation (Cautleyan, upper Katian), Upper Ordovician.
Material.—Type material and 236 specimens (88 cephala/cranidia, 33 pygidia, 80 librigenae, 33 rostral plates, one thoracic segment; NMW 2013.11G.13, 14, 16–249) from the type locality. One cranidium, (NMW 2013.11G.250) and one pygidium (NMW 2013.11G.251) from the Sholeshook Limestone Formation at Mylet Road near Llanddowror, Carmarthenshire. Price (1980b) reported 15 cranidia of S. cf. clavifrons, ranging throughout the Sholeshook Limestone Formation into the basal Slade and Redhill Mudstone Formation, from the Sholeshook and Prendergast areas near Haverfordwest and from the Llanddowror area but not all of his specimens could be located in museum collections. The single cranidium (GLAHM: A 9751) figured by Price (1980b: pl. 110: 18), from low in the Sholeshook Limestone in Craig-y-deilo Quarry, Llanddowror, was collected by J. Keith Ingham (Glasgow, UK) and is very likely conspecific with our material (see Remarks, below). The specimen register at the Hunterian Museum, Glasgow, UK, notes those collected by J.K. Ingham in the 1960s from the Sholeshook Limestone that were studied by David Price as part of his Ph.D. research (1971), and these include the figured cranidium and six further Staurocephalus specimens (what became referred by Price 1973a: “one from Loc. 18c, four from Loc. 18d, and two from Loc. 25”). Unfortunately, these specimens cannot currently be located, but are presumably among the 15 Staurocephalus cf. clavifrons cranidia listed by Price (1980b). Two cranidia from the historic collections of the Sedgwick Museum, Cambridge, UK, collected from the Sholeshook Limestone Formation in the Sholeshook railway cutting, Haverfordwest area, were labelled as Staurocephalus globiceps (SM A31591) and Staurocephalus cf. murchisoni (SM A31592; Fig. 6A) but likely also belong in our new species. Three further fragmentary cranidia labelled “Staurocephalus globiceps” in the Sedgwick Museum collections (SM A31587–9) were collected by Marr and Roberts (1885) from a locality near Pelcomb Cross, in strata now assigned to the basal Slade and Redhill Mudstone Formation (see Price 1973a). These specimens are generally too poorly preserved for specific identification, but one possesses a prominent genal spine (Fig. 6E) and is excluded from S. oliveae sp. nov. (see below).
Diagnosis.—Differs from other congeners by possessing the following combination of characters: Staurocephalus species with right-angled genal angle, lacking a genal spine or protrusion, with four lateral denticles anterior to the genal angle on the fixigena, decreasing in size posteriorly. Librigenae with 13–17 marginal denticles; mode and mean = 15. Inflated anterior glabellar lobe making up around two-thirds of preglabellar glabellar length (sag.). Pygidium around one fifth wider (tr.) than long. Inner of three pairs of spines making up just under two-fifths of total pygidial length. Cephalon (including rostral plate and librigenae) with dense covering of fine to medium granules, median occipital tubercle centrally located or on the posterior half of the occipital ring; pygidium with dense fine granules on spines with larger granules on axial rings and pleural ribs.
Description.—No complete articulated exoskeletons known. Cephalon approximately semicircular in outline; cranidium subtriangular (Figs. 3, 5A). Cephalon max. length (sag.) mean = 6.86 mm, range = 2.39–10.08 mm (n = 62). Cephalon max. width (tr.) mean = 10.15 mm, range = 3.37–14.01 mm (n = 42). On those specimens (n = 42) that are complete enough to measure both dimensions, maximum width (tr.) (mean = 10.02 mm, range = 3.37–14.01 mm) of cephalon is about one and a half times (mean = 1.50, range = 0.75–2.18) the maximum length (sag.) (mean = 6.71 mm, range = 2.39–8.45 mm). Glabella strongly convex (tr.), consisting of a greatly inflated, subcircular anterior glabellar lobe in front of a rectangular glabellar stem. In some specimens, glabellar stem tapers gently forwards, however, in less crushed specimens (e.g., NMW 2013.11G.6; Fig. 3C), margins of stem are sub-parallel. Occipital ring accounts for about one fifth of transverse posterior width of cephalon and about a tenth of the total cephalic length (sag.) medially, tapering abaxially by about a third; separated from the posterior border by a very shallow furrow. Median tubercle present, centrally located or on the posterior half of the occipital ring (Fig. 3A1, B1, C). Occipital furrow shallow and follows the transverse convexity of the glabellar stem and occipital ring, ending in deep pits where it meets the axial furrows. Anterior to the occipital furrow is the narrow (tr.), rectangular glabellar stem, indented by the lateral glabellar furrows S1 and S2; uniformly deep, semi-circular pits. S1 located approx. one fifth of length (sag.) of the glabellar stem in front of occipital furrow; L1 small, an elongate suboval abaxially, narrowing and subparallel-sided adaxially, weakly inflated, angled forwards at around 40° to the axis. S2 located approx. 70% along the length of the glabellar stem; L2 subrectangular, shorter, less inflated and less strongly angled forward than L1; L3 a very weakly inflated, transverse area behind S3. Transglabellar (S3) furrow marks the anterior end of the glabellar stem behind the inflated anterior lobe. S3 is consistently shallow, following the transverse convexity of the glabellar stem, and ends at the deep axial furrow where there is a suggestion of a pit in some specimens although this feature is not clearly preserved in any. Inflated anterior glabellar lobe (mean sag. length: 3.75 mm, n = 20) occupies about two thirds (mean = 0.59, range = 0.50–0.77) of total sagittal glabellar length (mean: 6.31 mm, n = 20), excluding occipital ring, while being approximately three times as wide (tr.) as the glabellar stem (excluding occipital ring), with about half of its length (sag.) protruding over the anterior cephalic margin. The axial furrows are consistently deep alongside the glabellar stem, then anterior of the transglabellar furrow they follow the curvature of the inflated anterior glabellar lobe, between the anterior glabellar lobe and inflated anterior fixigena, remaining consistently deep.
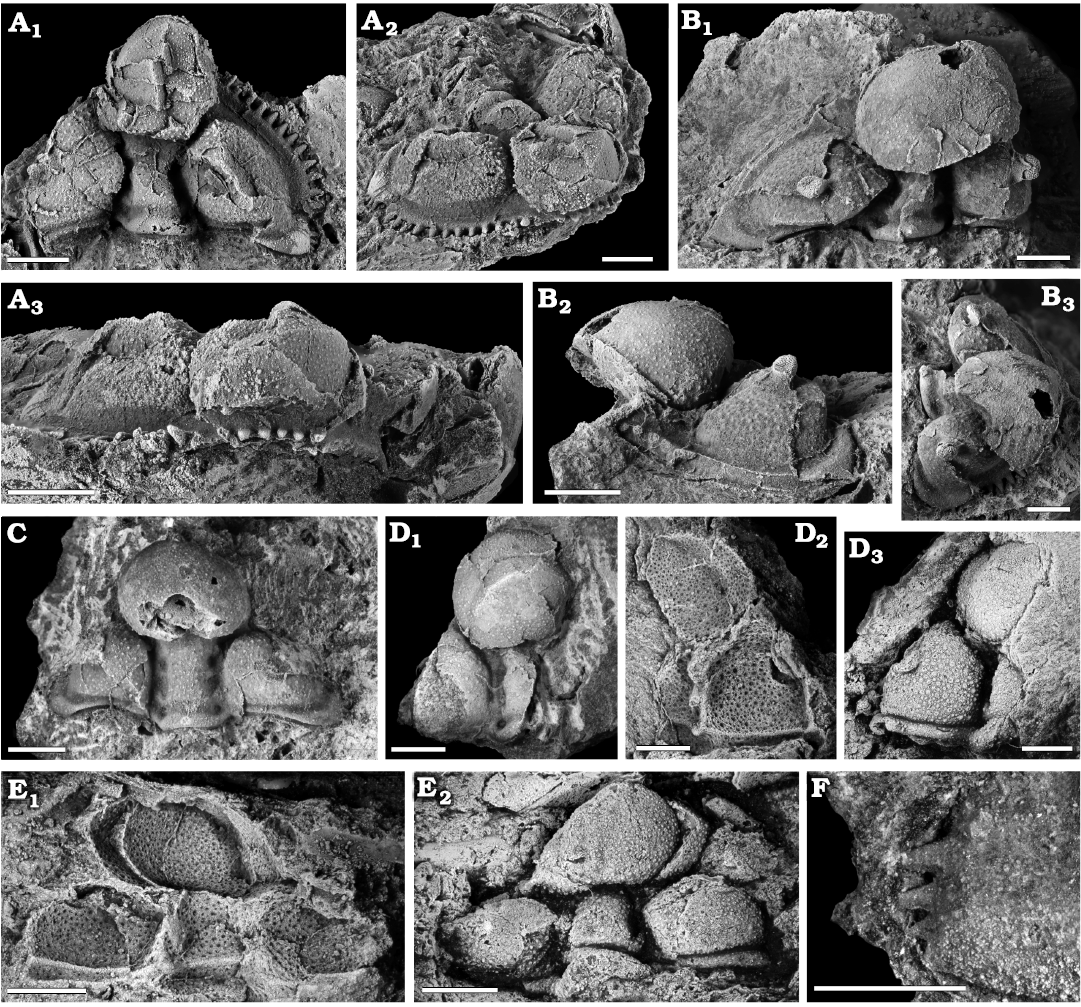
Fig. 3. Encrinurid trilobite Staurocephalus oliveae sp. nov. from the upper Katian (Upper Ordovician) Sholeshook Limestone Formation at Talfan Farm, Llanddowror, Carmarthenshire, Wales, UK. A. Holotype, NMW 2013.11G.1, cephalon in dorsal (A1), oblique anterolateral (A2), and frontal (A3) views. B. Paratype, NMW 2013.11G.2, cephalon in dorsal (B1), oblique anterolateral (B2), and dorsolateral (B3) views. C. Paratype, NMW 2013.11G.6, cranidium in dorsal view. D. Paratype, NMW 2013.11G.7, partial cranidium, dorsal view of internal mould (D1), ventral view of external mould (D2), and latex cast of external mould (D3). E. Paratype, NMW 2013.11G.8, cephalon, external mould in ventral view (E1) and latex cast of external mould (E2). F. Paratype, NMW 2013.11G.9, genal angle of cranidium, showing marginal denticles decreasing in size posteriorly. Scale bars 2 mm, except F 1 mm.
Fixigenae occupy about four fifths of the posterior (tr.) cephalic width. Palpebral lobe semicylindrical, bounded by a shallow furrow; located at the approximate mid width (tr.) of the fixed cheek, slightly posterior to level of S2 furrow. Fixigenal field inflated, sloping down steeply anteriorly, highest parts positioned above the level of the glabellar stem and commonly reaching half the preserved height of the anterior glabellar lobe in posterior view, although in uncrushed specimens the anterior glabellar lobe was probably somewhat higher. The posterior border is weakly convex (exsag.), extending transversely (sag.) from the occipital ring to behind the palpebral lobe beyond which it widens (exsag.) markedly towards the genal angle where it joins the posterior end of the lateral cephalic border at a right angle. No genal spine present, however four small denticles present on lateral border, increasing in size anteriorly and continuous with the larger denticles of the librigena. Posterior border furrow shallow, broadening markedly abaxially alongside the broadening of the posterior border.
The posterior branch of the facial suture is directed transversely from the palpebral lobe curving very gently forwards, until it reaches the lateral border where it curves slightly posteriorly. The anterior branch of the facial suture extends parallel to the sagittal line, from the palpebral lobe to opposite the middle (sag.) of the anterior glabellar lobe. Here the suture curves adaxially and is obscured by the anterior glabellar lobe in dorsal view, meeting the rostral and connective sutures on the flank of the anterior cephalic “bubble”.
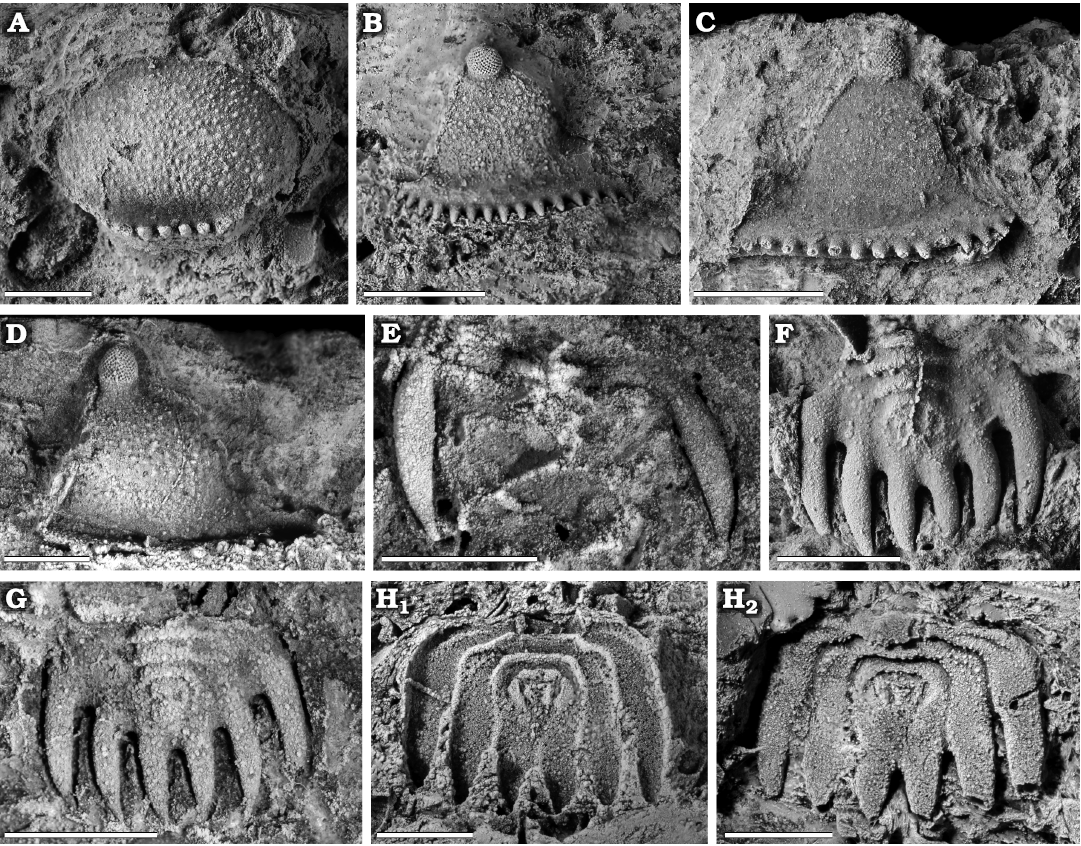
Fig. 4. Encrinurid trilobite Staurocephalus oliveae sp. nov. from the upper Katian (Upper Ordovician) Sholeshook Limestone Formation at Talfan Farm, Llanddowror, Carmarthenshire, Wales, UK. A. Paratype, NMW 2013.11G.5, rostral plate in ventral view. B. Paratype, NMW 2013.11G.3, right librigena in lateral view. C. Paratype, NMW 2013.11G.10, left librigena in lateral view. D. Paratype, NMW 2013.11G.12, right librigena in lateral view. E. Paratype, NMW 2013.11G.13, thoracic segment in dorsal view. F. Paratype, NMW 2013.11G.4, pygidium in dorsal view. G. Paratype, NMW 2013.11G.11, pygidium in dorsal view. H. Paratype, NMW 2013.11G.15, pygidium in ventral view of external mould (H1) and latex cast of external mould (H2). Scale bars 2 mm.
Librigena triangular, maximum length subparallel to the lateral border approximately two fifths longer than width normal to this; librigenal field also has a maximum length subparallel to lateral border two fifths longer than width normal to the field (Fig. 4B–D). Librigenae are a little less convex in comparison to the fixigenae. Short (exsag.) eye lies at summit of inflated field, which descends relatively steeply abaxially, separated from it by a shallowing of the slope. Visual surface of eye sub-oval, convex, taller than wide, set on top of a socle of around half its height. Visual surface containing over 120 small hexagonal lenses, with over 16 around the base, (Fig. 4D; NMW 2013.11G.12, max. length 5.94 mm, max. width 4.31 mm), arranged in hexagonal packing. Librigenal border furrow uniformly broad and shallow. Librigenal border comprises a very narrow, flat-lying, very weakly swollen ridge, barely distinguished from the border furrow, and a row of denticles, each longer than the width of the ridge. On complete specimens (n = 36), 13–17 denticles are present with a mean of 15 (rounded from 14.6) and mode of 15 (Fig. 7). The number of denticles is independent of the size of the librigena. Portion of librigena anterior of inflated field comprises a low-lying subtrapezoidal area, the anteriormost part of which is upturned to form a subtriangular flap, the librigenal precranidial lobe (Figs. 4B, C, 5A2). Anterior edge of precranidial lobe has a very gentle forwards curvature that matches the curve of the anterolateral margin of the rostral plate, the two librigenal precranidial lobes fitting with the rostral plate along the connective sutures to form the underside of the inflated cephalic lobe (“bubble”).
Rostral plate sub-circular, split into two distinct parts (Fig. 4A). Anterior part of plate lies beneath the anterior glabellar lobe along the rostral suture and is inflated, matching the convexity of anterior glabellar lobe and forming the anteroventrally directed part of the anterior cephalic “bubble” (see Fig. 5). Separated by a transverse furrow, the posterior section consists of a set of six denticles. Behind the denticles there is short, flat ventral section, of similar length (sag.) to the denticles and continuing in the same plane as them.
Hypostome not known.
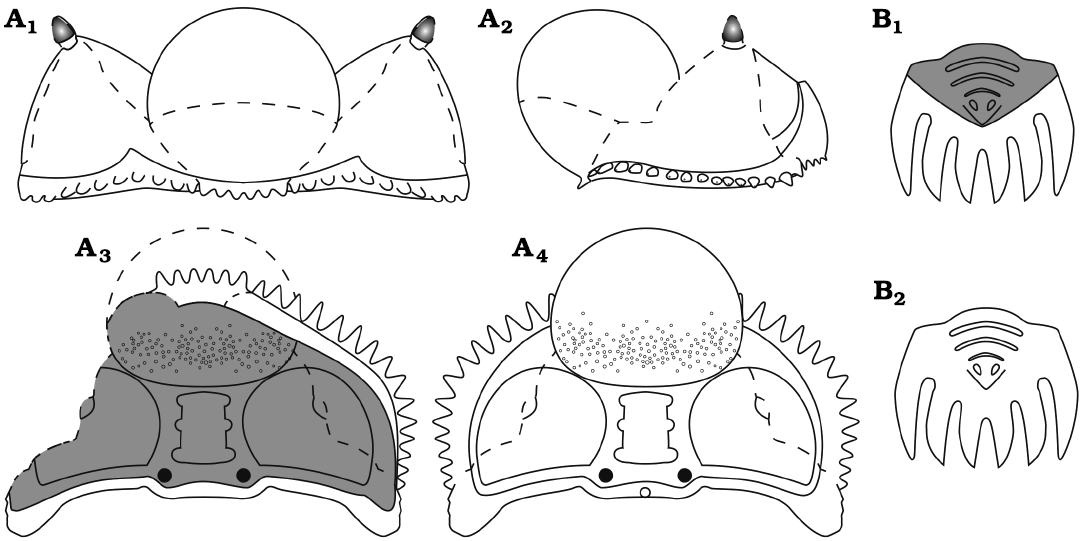
Fig. 5. Reconstruction drawings of the encrinurid trilobite Staurocephalus oliveae sp. nov. A. Cephalon, in anterior (A1), left lateral (A2), ventral (A3), and dorsal (A4) views. B. Pygidium, in ventral (B1) and dorsal (B2) views. Cephalon has a mean maximum width (tr.) of 10.15 mm; pygidium has a mean maximum width (tr.) of 3.96 mm.
Internal mould of cranidium (including rostral plate) apart from the furrows and borders densely and evenly covered in relatively small granules. Latex casts of external moulds reveal the granules to be larger and more densely packed on external exoskeletal surfaces, although the number of granules remains consistent (Fig. 3D3, E2). Additionally, the larger granules shown on latex casts vary in size. Internal moulds and latex casts of external moulds of the librigenal field have similar granulation to those of the cranidium, with occasional larger granules situated on the marginal border, in line with the denticles.
Thorax known only from internal moulds of two isolated thoracic segments from the type locality that probably belong to this species (Fig. 4E) and are consistent with segments from the posterior part of the thorax (e.g., see specimen of S. clavifrons illustrated by Kielan 1957: pl. 3: 2). Axial ring occupies about a quarter of segment width (tr.). Pleura parallel-sided over most of its length (tr.) turning posteriorly through almost a right angle at the fulcrum and extended as a posteriorly directed pleural spine, almost parallel to the sagittal line, that tapers to a sharp point over the distal 30% of its length. The granulation of these spines is denser and finer than that of the cranidium. Prominent large granules present at the fulcrum. A small number of medium granules developed into short spines visible on a small part of the axial ring of one of the specimens, but preservation too poor to determine overall sculpture.
Pygidium subquadrate in outline, being about one fifth wider (tr.) than long, including spines (Figs. 4F–H). Axis occupies approximately 50% of pygidial length (sag., including spines), and approximately 30% of the anterior pygidial width. Axis convex (tr.) anteriorly, becoming progressively less so and tapering (tr.) posteriorly. Axis has three distinct axial rings, the first two being transverse with the third arching gently forward, separated by axial ring furrows, which become relatively shallower posteriorly following the decreasing convexity of the axis. Each furrow is shallow and narrow medially, widening and becoming deeper laterally. Axis ends with a subtriangular, flat terminal piece. Axial furrows very shallow, little more than a break in slope between the convex axis and flat-lying pleural areas. Axial rings confluent with pleural ribs, the first pair of which are directed transversely for a short distance before curving posteriorly through about 90°, the second and third pairs are essentially parallel to the sagittal line. On external surface (Fig. 4H2), the transverse, inner parts of the anterior two pleural ribs bear shallow furrows directed very slightly obliquely to the anterior edge of the ribs at about one third of the distance from that edge to the posterior edge. The pleural ribs each broaden and become robust, posteriorly directed, spines; the outermost being the longest. The inner edges of the outer and middle pair of spines remains parallel to the sagittal line, with their outer edges curving gently inward, tapering to form a point. On some specimens (e.g., Fig. 4F), both inner and outer edges of the inner pair of spines are gently curved to form a more symmetrical spine, although in others the inner edge is straighter (e.g., Fig. 4G). The spines are directed parallel to the sagittal line and terminate along an essentially transverse line. The innermost pair of spines (mean = 1.41 mm, n = 4) account for around two fifths (mean = 0.42, range = 0.36–0.46, n = 4) of total length (mean = 3.37 mm, n = 4) of the pygidium.
The pygidium has a varied surface sculpture on the internal mould (Fig. 4F, G) and latex casts of external moulds (Fig. 4H2), with the spines having fine and dense granulation, relatively smaller and denser than on the cranidium. A large prominent tubercle is visible in the centre of each axial ring, similar to the occipital ring, with a row of coarse granules either side (e.g., NMW 2013.11G.11, Fig. 4G). Larger granules are also present on the pleural field, of some specimens (e.g., NMW 2013.11G.4, Fig. 4F).
Remarks.—Staurocephalus oliveae sp. nov. is most similar to Staurocephalus clavifrons Angelin, 1854, which was first described from the Staurocephalus Beds (now Ulunda Mudstone, Rawtheyan, uppermost Katian) of Ålleberg, Västergötland, Sweden (cranidium figured by Angelin 1854: pl. 34: 8; refigured by Kielan 1957: pl. 4: 1; see also Kielan-Jaworowska et al. 1991: 240). It was subsequently described in detail from an equivalent horizon in the Holy Cross Mountains, Poland, by Kielan (1957) and from the overlying Mucronaspis mucronata Zone (“Dalmanitina mucronata zone” then) there (probably Hirnantian, Upper Ordovician; but see Discussion section below) (Kielan 1960). Staurocephalus oliveae sp. nov. differs from S. clavifrons in lacking genal spines, in possessing four rather than three denticles on the lateral border of the fixigena, with these denticles increasing in size anteriorly whereas they are the same size in S. clavifrons (see e.g., Kielan 1957: fig. 2) and, in most specimens, a greater number of denticles (13–17, mean 15, cf. 13–14) on the lateral margin of the free cheeks. Some specimens also differ in having the medial occipital tubercle posteriorly (cf. centrally) placed. The cephalic and pygidial granulation of S. oliveae sp. nov. is denser and finer than that of S. clavifrons both on internal moulds and on the external surface. Staurocephalus oliveae sp. nov. may differ from S. clavifrons in the number of lenses in the eye (120 cf. 60 recorded by Kielan 1957: 165) although the size of the librigena on which this was counted by Kielan is not known.
Staurocephalus clavifrons was reported or described from Ashgill (Cautleyan and Rawtheyan, upper Katian) horizons in Wales (Whittington 1965; Price 1973a), northern England (Ingham 1974, McNamara 1979) and the Republic of Ireland (Dean 1971). However, Ingham (1977) later described some better-preserved specimens from the Rawtheyan of northern England, which showed that although they were morphologically very similar, there were differences in sculpture from the type and Polish specimens. He concluded that none of the occurrences from Britain and Ireland could confidently be placed in S. clavifrons and should instead be assigned to S. cf. clavifrons or S. aff. clavifrons. As a result, Price (1980b: 866, pl. 110: 18) described as S. cf. clavifrons the material that he earlier (1973a: 245) recorded as S. clavifrons from throughout the Sholeshook Limestone Formation and the base of the overlying Slade and Redhill Mudstone Formation. As is noted above, with the exception of the Slade and Redhill Mudstone Formation specimens, this material is here included in S. oliveae sp. nov., although questioningly so given the poor preservation of the available specimens. The posterolateral part of the left side of the cranidium from the Sholeshook Limestone at Craig-y-deilo quarry figured by Price (1980b: pl. 110: 18) appears to sweep rearwards slightly as if representing the innermost proximal part of a genal spine. However, this area is only slightly beyond the level of the palpebral lobe and is where the width (exsag.) of the border begins to expand. Examination of the specimen in the Hunterian Museum showed that the outermost part of the fixigena curves steeply downwards into the matrix and the genal area is not exposed. Price did not mention a genal spine in his description and the cranidium is similarly proportioned to ours and has a similar sculpture and is assumed to be conspecific. An historic cranidium in the Sedgwick Museum labelled Staurocephalus cf. murchisoni from the Sholeshook Limestone in the Sholeshook railway cutting near Haverfordwest, is within the size range of S. oliveae sp. nov. has similar cranidial and glabellar proportions and has similar sculpture (Fig. 6A herein). Although the genal corner is not preserved, the posterior border width (exsag.) and posterior edge of the posterior border (which does not turn sharply rearward to indicate a spine) are consistent with S. oliveae sp. nov. suggesting that it may represent that species.

Fig. 6. Historic specimens of the encrinurid trilobite Staurocephalus from museum collections. A. SM A31592, cranidium in dorsal view labelled “Staurocephalus cf. murchisoni Barrande” from the Sholeshook Limestone Formation (upper Katian, Upper Ordovician) in the Sholeshook railway cutting, Haverfordwest area, herein assigned to S. oliveae sp. nov.? B. BIRUG 48862, cranidium in dorsal view labelled “Staurocephalus cf. murchisoni” from the lower Tre-wylan Beds (Cautleyan) (upper Katian, Upper Ordovician) in the Llansantffraid-ym-Mechain district of the south-eastern Berwyn Hills, Mid Wales (Whittington 1938: 451, locality 42), herein assigned to S. oliveae sp. nov.? C, D. Specimens from the King collection, Sedgwick Museum, from the (locally) basal Ashgill mudstones (lower Rawtheyan, Ashgill Zone 5) (upper Katian, Upper Ordovician) of Aber Marchnant, Berwyn Hills, Mid Wales, originally identified as Staurocephalus cf. murchisoni by King (1923) and assigned to S. clavifrons by Whittington (1968), herein assigned to S. aff. oliveae. C. SM A39813, incomplete cranidium in dorsal view. D. SM A39814.A, incomplete cranidium in dorsal view (D1) and close up of left genal area (D2) to show large lateral denticles (arrowed). E. SM A31589, dorsal view of incomplete cranidium labelled “Staurocephalus globiceps” from the Marr collection, Sedgwick Museum, from the Slade and Redhill Mudstone Formation at Pelcomb Cross, Haverfordwest, herein assigned to S. cf. clavifrons. Note the prominent genal spine on the right side. Scale bars 1 mm, except D2 0.5 mm.
Hammann and Leone (2007: 76) discussed Ingham’s wider concerns (1977: 89) arising from the sculpture of the material of Staurocephalus from the highest Rawtheyan of the Cautley area in northern England in the light of their own specimens from the Ashgill of Sardinia and concluded that the differences that Ingham highlighted were insufficient to provide any separation from S. clavifrons. As a result, they included the material from North Wales described by Whittington (1965) and Ireland by Dean (1971) unequivocally in the synonymy of S. clavifrons and that described by Ingham (1974, 1977) from the uppermost Rawtheyan of the Cautley area of northern England questioningly so. They did, however, consider that Ingham’s (1977) S. aff. clavifrons from the lower Rawtheyan Swindale Limestone of the nearby Cross Fell Inlier differs from Angelin’s (1854) species in its dense, large, granulated cranidial tubercles of various sizes.
Whittington (1938: 451) reported Staurocephalus cf. murchisoni and Staurocephalus sp. from the Cautleyan lower Tre-wylan Beds (“Phillipsinella parabola zone”) at two localities in the Llansantffraid-ym-Mechain district of the south-eastern Berwyn Hills, Mid Wales. Two of his cranidia of Staurocephalus cf. murchisoni (Whittington locality 42; both BIRUG 48862, on same slab) were located in the Lapworth Museum, University of Birmingham. Although both specimens lack their genal areas, precluding definitive identification, their glabellar proportions and granulation are consistent with S. oliveae sp. nov. and they are tentatively assigned to the new species (Fig. 6B). Also from the Berwyn Hills, the historic King collection at the Sedgwick Museum includes three incomplete cranidia (SM A39813, 39814.a and 39814.b) and one librigena (SM A39812) from the (locally) basal Ashgill mudstones (lower Rawtheyan, Ashgill Zone 5, see Price 1980a, 1981, 1982: 61) at Aber Marchnant, originally assigned to Staurocephalus cf. murchisoni by King (1923: 497) and, presumably, are the material listed as S. clavifrons from there by Whittington (1968: table 4). The cranidia are smaller than those described here and their granulation appears coarser and less dense, but they have similar glabellar proportions and posterior margin and fixigenal corner outline (Fig. 6C, D1), although the fixigenal denticles increase in size posteriorly rather than decrease as in S. oliveae sp. nov. (Fig. 6D2). These cranidia are much closer to S. oliveae sp. nov. than to S. clavifrons and are herein reassigned to S. aff. oliveae. The librigena from the same collection possesses 12–13 marginal denticles, which would place it at the lower end of the range for S. oliveae sp. nov.
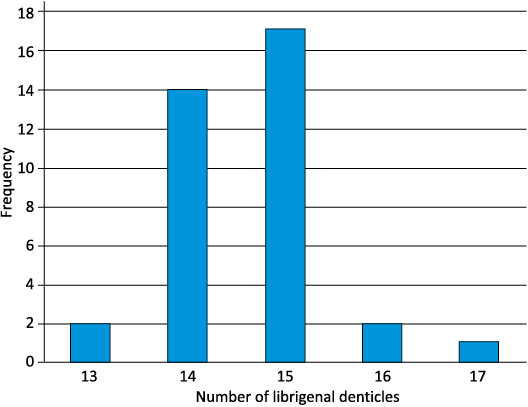
Fig. 7. Chart showing the frequency distribution of numbers of denticles on lateral margins of complete librigenae of Staurocephalus oliveae sp. nov. from the upper Katian (Upper Ordovician) Sholeshook Limestone Formation at Talfan Farm, Llanddowror, Carmarthenshire, Wales, UK. n = 36.
The absence of genal spines distinguishes S. oliveae sp. nov. from the other material previously described as, compared with or considered to have affinity to S. clavifrons from North Wales (Whittington 1965) and northern England (Ingham 1974, 1977). Dean (1971) only found one cranidium of Staurocephalus from the Ashgill Kildare Limestone in eastern Ireland, with no associated librigenae. Staurocephalus oliveae sp. nov. has a similar internal mould sculpture to Dean’s (1971: pl. 19: 1–4) cranidium and is similar in outline, glabellar proportions and overall size, although the Irish specimen is marginally narrower (tr.). However, the absence of a genal spine distinguishes S. oliveae sp. nov. from the Irish cranidium.
While details of the lateral cephalic margin are not known sufficiently in S. cf. clavifrons and S. aff. clavifrons of Ingham (1977) from the uppermost Rawtheyan and the lower Rawtheyan (Ashgill Zone 5) respectively of northern England, the librigena of material described by Whittington (1965: 53–54, pl. 15: 4–19) from the Rawtheyan Rhiwlas Limestone in North Wales as S. clavifrons has 14 denticles (cf. 13–17, mean and mode = 15, in S. oliveae sp. nov.). In addition to lacking a genal spine, S. oliveae sp. nov. also differs from the Rhiwlas Limestones specimens in having smaller, more closely spaced, granules on both the internal mould and the external surface of the cephalon. Staurocephalus cf. clavifrons of Owen and Romano (2011) from probable Cautleyan strata in eastern Ireland is too incomplete to determine whether there is a genal spine or for the numbers of denticles on the fixed or free cheeks to be determined. The specimens have the sparser but coarser external surface granulation of typical S. clavifrons but the librigena (Owen and Romano 2011: fig. 2m) also shows extremely fine pits between the granules (as in S. cf. clavifrons of Ingham 1977); the eye contains about 70 lenses. Specimens described as S. aff. murchisoni Barrande by Weir (1959) from the lower Ashgill of Slieve Bernagh in western Ireland are too poorly preserved for useful comparison with any named species.
The absence of genal spines and the surface sculpture also differentiate S. oliveae sp. nov. from material assigned to S. clavifrons from Sardinia from the upper part of the Rawtheyan Punta S’Agricola Member of the Domusnovas Formation underlying the Hirnatian Rio San Marco Formation (Loi et al. 2023) described by Hammann and Leone (2007). These differences also distinguish S. oliveae sp. nov. from material ascribed to S. clavifrons by Zhou and Zhou (1982) from Inner Mongolia from a formation correlated by them with the Rawtheyan on the basis of the trilobite fauna. There is what appears to be a tubercle at mid-length (sag.) on the occipital ring of one of Zhou and Zhou’s (1982: pl. 1: 6) figured cranidia but it is difficult to discern clearly and genal denticles are not visible on either cranidium. Details of the median occipital tubercle and genal denticles are not known from the Sardinian occurrence. Shaw (2000: 385) considered that material from the Králův Dvůr Formation (upper Rawtheyan) of Bohemia previously assigned to S. clavifrons is too poorly preserved for detailed species comparison. Although the genal area on the single incomplete cranidium known to Marek (1964: pl. 1: 3) from here is not fully exposed, it is possible that it does not possess a spine; S. oliveae sp. nov. differs from it in having markedly smaller and less dense granulation. The incomplete S. clavifrons cranidium featured by Štorch and Mergl (1989: pl. 5: 11) from the Králův Dvůr Formation has a surface sculpture like that illustrated by Marek (1964) and the details of the outermost parts of the fixigena are not preserved.
The cranidium described by Owen and Bruton (2012) from the uppermost Caradoc of Maine, USA, as S. cf. clavifrons is too incomplete to determine whether it is closest to S. clavifrons or S. oliveae sp. nov., though the granulation of the external surface is much closer to that of Angelin’s (1854) species than the fine, dense coverage of S. oliveae sp. nov.
The other named Ordovician Staurocephalus species comprise: Staurocephalus pilafrons Owen and Bruton, 1980 (uppermost Caradoc, lower Katian of the Oslo Region, Norway), Staurocephalus djarikensis Apollonov, 1974 (Ashgill, Katian of Kazakhstan), and Staurocephalus elongatus von Freyberg, 1923 (redescribed by Knüpfer 1967 from the lower Ashgill, Katian of Thuringia, Germany). Staurocephalus oliveae sp. nov. differs from S. pilafrons in having a relatively broader and shorter anterior glabellar lobe in relation to the glabellar stem, smaller granules and in lacking a large median occipital spine (although it has a prominent tubercle). Staurocephalus pilafrons also has a distinct eye ridge; the details of its genal angle or numbers of genal denticles are not known. The cranidial border (and therefore the genal angle) is not preserved in the material of S. djarikensis described by Apollonov (1974). Staurocephalus oliveae sp. nov. differs from S. djarikensis in its glabellar proportions, having a relatively wider and longer (sag.) anterior glabellar lobe taking up two thirds of the glabellar length versus around half in S. djarikensis, and a narrower glabellar stem than the Kazakh species. Staurocephalus oliveae sp. nov. differs from S. elongatus by having much finer granules on the anterior glabellar lobe and by having a differently proportioned anterior glabellar lobe, which is about one fifth wider (tr.) than long (sag.), compared to that of S. elongatus, which is about one fifth longer (sag.) than wide (tr.). Knüpfer (1967: 15–16, pl. 1: 13, 14, pl. 17: 7, 8) stated that the posterior section of the genal angle is “directed backwards”, and although he considered that it cannot be determined if there is a genal spine present similar to S. clavifrons, the figured holotype (Knüpfer 1967: pl. 17: 8) shows the left posterior margin extended strongly rearwards to form a narrow point. Staurocephalus oliveae sp. nov. clearly differs from this species in having a posterior margin that lacks any posteriorly directed portion like that seen on Knüpfer’s cranidium. Ferretti and Barnes (1997) described the conodonts from the Kalkbank Limestone from which S. elongatus was recovered, placing the formation at a level somewhere near the base of the A. ordovicicus Zone of the Katian stratigraphy that was reiterated by Kraft et al. (2023) in their recent review of the Ordovician of the Bohemian Massif. In their paper on Katian conodonts from Wales, Ferretti et al. (2014: 812) noted that: “…the Kalkbank fauna is clearly not older than the A. ordovicicus Zone and it seems likely that it is at least broadly coeval with the Sholeshook Limestone”, making it approximately contemporaneous with S. oliveae sp. nov.
In addition to revising S. clavifrons, Kielan (1957) described specimens that she termed “Staurocephalus sp. b” and “Staurocephalus sp. c” from the Ashgill of Sweden (“Staurocephalus sp. a” was assigned to S. pilafrons by Owen and Bruton 1980: 30). Staurocephalus oliveae sp. nov. differs from “Staurocephalus sp. b” described by Kielan (1957: 169, pl. 3: 1a–c) from the Upper Jonstorp Formation (Rawtheyan) in the Siljan region of Sweden, in lacking a genal spine and in commonly having more (13–17 cf. 14) marginal spines on the librigena and four rather than three on the fixigena. The single cranidium of “Staurocephalus sp. c” described by Kielan (1957: 170, pl. 4: 4) from the Jonstorp Formation in Västergötland, Sweden, is too incomplete for detailed comparison but S. oliveae sp. nov. appears to have a relatively narrower glabellar stem. Kielan (1957: pl. 4: 5) also assigned a single pygidium to the same species; S. oliveae sp. nov. has noticeably shorter spines, with its innermost pair of spines accounting for just under two fifths of maximum pygidial length (sag.), while those of “Staurocephalus. sp. c” take up just under half of maximum pygidial length (around 10% longer). Staurocephalus sp. of Weber (1948), from the Ashgill of Mamontovo, Kazakhstan (see Kielan 1957: 160) is only known from a single, poorly preserved, cranidium. Staurocephalus oliveae sp. nov. has a distinctly larger anterior glabellar lobe, which occupies two thirds of maximum glabellar length (sag.) (excluding occipital ring), while that of Staurocephalus sp. occupies just over a half.
The absence of a genal spine distinguishes S. oliveae sp. nov. from most of the Silurian species of Staurocephalus, such as the type species S. murchisoni Barrande, 1846 (Beroun, Czech Republic, Wenlock), S. susanae Thomas, 1981 (Dudley and Malvern, UK, Wenlock), S. clowni Tomczykowa, 1987 (Poland, Sheinwoodian, Wenlock), S. oarion Holloway, 1980 (Arkansas, USA, Sheinwoodian, Wenlock), S. aff. oarion Holloway, 1980 (Arkansas, USA, Wenlock), S. lagena Holloway, 1980 (Arkansas, USA, Whitwellian, Wenlock), S. aff. lagena Holloway, 1980 (USA, same locality as S. lagena), S. obsoletus (Weller, 1907) (Illinois, USA, Ludlow), S. struszi Chatterton and Campbell, 1980 (Canberra, Australia, Whitwellian, Wenlock) and S. mitchelli Chatterton and Campbell, 1980 (New South Wales, Australia, Ludlow). Both S. struszi and S. mitchelli have pygidia with proportionally smaller and flatter spines than those of S. oliveae sp. nov., which also has more lateral denticles on its free cheeks, 13–17, compared to “11 or 12” for S. struszi, and 10 for S. mitchelli. Staurocephalus mitchelli is one of the youngest species of Staurocephalus known, occurring in the Yarwood Siltstone Member of the Black Bog Shale, which Bicknell and Smith (2021) placed within the Ludfordian. Staurocephalus obsoletus is potentially of similar age, but the stratigraphical horizon (Niagaran, from Chicago Drainage Canal, near Lemont, Illinois) provided by Weller (1907) for his single specimen is too imprecise to be certain.
Another Ludfordian species, S. azuella Šnajdr, 1980, was described from the Cromus beaumonti horizon in the Kopanina Formation (see Vaněk and Valíček 2002: 52; dated as Ludlow, Ludfordian, see Lehnert et al. 2007) in the Czech Republic. Although Šnajdr (1980) listed his single specimen as an “incomplete pygidium with corroded exoskeleton”, his description and illustration focus only on the pygidium, which lacks the tips of the spines. Excluding the spines, S. oliveae sp. nov. differs in having a relatively longer pygidium (just over four fifths as wide [tr.] as long [sag.]) than that of S. azuella (twice as wide [tr.] as long [sag.]) The shape and proportions of the pygidial spines are also different, those of S. oliveae sp. nov. being slightly wider in the middle than the base and rapidly tapering to a point, with the abaxial edge of the first two pairs of spines curving gently abaxially, while the adaxial side of the spines remains straight giving these spines a conical, slightly outwardly inflated appearance. The preserved portions of the spines of S. azuella are distinctly narrower (tr.) and of more uniform width, a gently tapering rectangle in outline and appear to be directed straight rearwards.
Staurocephalus sp. of Whiteley et al. (2002) from Rochester Shale, New York, USA (Aeronian, Llandovery) is known only from a single librigena from which that of S. oliveae sp. nov. can be differentiated by having more lateral denticles, there being 12 in the American species. Like S. oliveae sp. nov., S. trichochin Kobayashi and Hamada, 1985 (Japan, Whitwellian, Wenlock) lacks genal spines but S. oliveae sp. nov. has a relatively smaller and less inflated anterior glabellar lobe as well as relatively larger fixigenae. Staurocephalus sp. of Curtis and Lane (1997, 1998) from the Aeronian, Llandovery, of northern England is too incompletely known for extensive comparisons to be made, although S. oliveae sp. nov. can be distinguished from it by having fewer denticles on the rostral plate.
Stratigraphic and geographic range.—Sholeshook Limestone Formation, upper Katian, Upper Ordovician. Llanddowror, Carmarthenshire and Haverfordwest area, Pembrokeshire, Wales, UK.
Discussion
The “bubble-headed” morphology of Staurocephalus.—With its inflated cephalic lobe, Staurocephalus is an example of what Fortey and Owens (1997) termed a “bubble-headed trilobite”. Fortey and Owens (1997) reviewed this morphology and noted that it was developed separately and in different ways in several trilobite families from the Cambrian to the Silurian. In most instances, they considered that the “bubble” was a development of all, or just the frontal part, of the glabella. These included what they termed the “Staurocephalus type” in which the anterior lobe of the glabella is inflated. This widely accepted interpretation of the condition in Staurocephalus was challenged by Tomczykowa (1987) who argued that the bulbous feature was a development of the anterior cranidial border rather than of the glabella, effectively the bubble-head condition subsequently termed the “Nepea type” by Fortey and Owens (1997). However, Edgecombe et al. (1988: 792) provided a detailed and convincing critique of Tomczykowa’s (1987) interpretation and supported the long-held interpretation of the bulbous structure comprising the frontal lobe of the glabella together with the cranidial anterior border, the precranidial lobe of each librigena and the rostral plate. This interpretation was endorsed by Kielan-Jaworowska et al. (1991: 240).
Librigenal denticle numbers through time in Staurocephalus.—The large sample size of Staurocephalus oliveae sp. nov. has shown that the number of librigenal denticles is highly variable; something that has not previously been reported in the genus. Taking those values that have been given for other species in the literature, there is some evidence for a consistent decrease in the number of denticles through time, ranging from the mean value of 15 (range 13–17) in S. oliveae sp. nov. to 10 in the late Silurian species S. susanae and S. mitchelli (Fig. 8). The number of species in which the number of denticles is known is limited but the decrease seems to be unidirectional and gradual and thus there are no markedly discrete groups of stratigraphically constrained species. At face value at least, there was a period of stasis in the number of denticles from the middle Llandovery to the early Wenlock but this may be an artefact of the lack of data from the relatively long Telychian (late Llandovery).
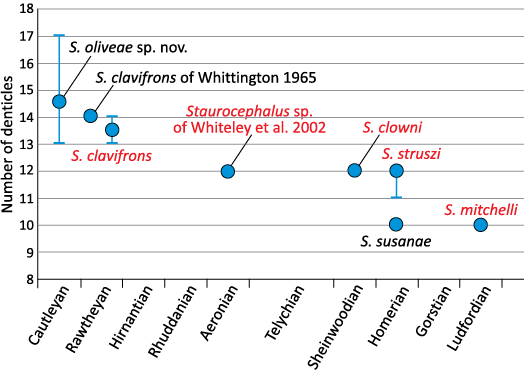
Fig. 8. Graph showing the number of lateral denticles on librigenae of various Staurocephalus species through time. Circle represents the mean, vertical lines the range where one was observed in that species. Chatterton and Campbell (1980) described S. struszi as having “twelve (possibly eleven on some specimens)” denticles. Species names in black font represent occurrences in England and Wales, those in red font occur outside Avalonia. Note that Cautleyan and Rawtheyan are regional stages from the Anglo-Welsh chronostratigraphy and belong in the upper part of the international Katian Stage. The remaining stages are international. The Telychian is approximately twice the length of the other stages (e.g., Ogg et al. 2008: fig. 1.1), hence the wider spacing allocated to it.
Implications of the new species of Staurocephalus for the upper Katian Staurocephalus clavifrons Zone.—The Staurocephalus clavifrons Zone has long been recognised in Sweden (Linnarsson 1869; Tullberg 1882; Jaanusson 1963: 112–114; Nielsen et al. 2023: 294), the Danish Island of Bornholm and Poland (Kielan 1960: 7–8, 13–16). As noted by Price (1973b: 538–539), the lower parts of the zone share many species with the underlying Eodindymene pulchra Zone, with some possibly extending lower in the succession (see also Jaanusson 1963: 133). In Sweden (the type region) and on Bornholm, the S. clavifrons Zone is succeeded by rocks of the Regional Tommarpian (= Hirnantian) Stage and is equated with the uppermost Katian (Nielsen et al. 2023: fig. 3). It is thus essentially Rawtheyan in terms of the Anglo-Welsh regional chronostratigraphy. The same effectively applies in Poland although the zonal term is no longer applied, but what is now termed the Wólka Formation that contains the S. clavifrons and E. pulchra zone faunas of Kielan (1960) and is overlain by the Hirnantian Zalesie Formation (Trela 2021, 2023). Staurocephalus clavifrons in Scandinavia and Poland has not been recorded beyond its occurrence in rocks historically ascribed to the S. clavifrons Zone and, in Poland, the overlying Hirnantian beds containing “Dalmanitina” [now Mucronaspis] mucronata (see Kielan 1960: 32, 156–157; but see also Nilsson 1979 and Nielsen et al. 2023: 296, concerning the occurrence of M. mucronata in the S. clavifrons Zone in Skåne, SW Sweden).
The occurrences of Staurocephalus had an important role in the development of Upper Ordovician stratigraphy in Wales and northern England, including the initial formulation of the Ashgill Series by Marr (1907). This culminated in the proposal by Whittington (1968: 116–117) that the Rhiwlas Limestone of North Wales should be assigned to the S. clavifrons Zone. Whittington’s definition of the zone was based on an assemblage of ten trilobite species, including S. clavifrons, and he equated it with the shelly zones 4–7 identified by Ingham (1966) in northern England. These comprised the main part of what became the Rawtheyan Stage in the revision of the Ashgill proposed by Ingham and Wright (1970).
Price (1973b) provided a critique of the S. clavifrons Zone, especially its use in Wales and England together with that of the so-called Phillipsinella parabola fauna or zone (see Ingham 1966: fig. 2; Whittington 1968: 117). He noted that the first attempted correlation of units containing Staurocephalus in Britain was by Marr and Roberts (1885: 487–488) who linked its presence in the Sholeshook Limestone with the Rhiwlas Limestone and the Staurocephalus Zone in Sweden. Price (1973a: table 2) recorded S. clavifrons, from levels throughout most of the Sholeshook Limestone; a formation that he then correlated with the lower and middle parts of the Cautleyan Stage that precedes the Rawtheyan (he subsequently identified the very uppermost part as being lowest Rawtheyan [Price 1980a]). Price (1973b: 538) noted that in addition to S. clavifrons, the Sholeshook Limestone had yielded another four of the ten species previously listed by Whittington (1968) as comprising the assemblage characterizing the S. clavifrons Zone together with material probably belonging to three other species on the list and a different species from that assigned to another of the genera by Whittington (1968). Most significantly, Price (1973a: 538) stressed that all nine species in common with, or very closely allied to, Whittington’s (1968) assemblage occur low in the Sholeshook Limestone in strata assigned to the lower Cautleyan (zones 1 & 2 of Ingham 1966). Price (1973b: 538) therefore considered that this discrepancy in age called into question the stratigraphical value of the S. clavifrons Zone.
Specimens in historical collections from all but the uppermost part of the Sholeshook Limestone are cautiously ascribed to S. oliveae sp. nov., thus extending its range through the Cautleyan. As noted in the Material and methods section (above) an incomplete cephalon (SM A31589) in the Marr collection from the overlying Slade and Redhill Mudstone Formation (probably lower Rawtheyan) at Pelcomb Cross, Haverfordwest, has a prominent genal spine (Fig. 6E) and therefore likely belongs in S. clavifrons or a closely similar species rather than S. oliveae sp. nov. The recognition herein that the species of Staurocephalus from at least the lower part of the Sholeshook Limestone (Ashgill Zone 1) and probably the entire Cautleyan, is a new species holds the prospect that S. clavifrons has more stratigraphical value than indicated by Price (1973a). As noted in the discussion of the new species above, the assignment of material to S. clavifrons from the Rhiwlas Limestone (lower Rawtheyan) by Whittington (1965) seems well founded. Specimens from northern England from the lower and uppermost Rawtheyan of the Cross Fell Inlier and Cautley areas, respectively, are closest to S. clavifrons although differing in their surface sculptures (see Ingham 1977; Hammann and Leone 2007 and above). Occurrences of the species in Sardinia (Hammann and Leone 2007), Inner Mongolia (Zhou and Zhou 1982) and possibly Ireland (Dean 1971) are in faunas of Rawtheyan (uppermost Katian) age as are those in Scandinavia and Poland noted above, with the possible extension of the species into the Hirnantian in Poland. Thus, whilst many of the taxa that occur in association with S. clavifrons are known from lower horizons in the Ashgill, the species itself may well still have great value internationally as a Rawtheyan (i.e., uppermost Katian) and possibly lower Hirnantian indicator. If this is the case, the S. clavifrons Zone in Scandinavia could be defined on the basis of the first occurrence of S. clavifrons there and, at least, most of its range. Whether such a zone would have international application would require the precise levels of the occurrences of S. clavifrons elsewhere to be more tightly constrained.
As noted above, historic specimens from the lower Tre-wylan Beds collected by Whittington (1938) in the south-eastern Berwyn Hills are tentatively assigned to S. oliveae sp. nov. Price (1977: 764–766, pl. 98: 6–9) assigned trinucleid specimens from Whittington’s (1938) locality to Tretaspis moeldenensis Cave, 1960 (sensu lato) Population B, (= T. cf. kiaeri Størmer, 1930 of Whittington 1938: 445, pl. 38: 1–3). This is consistent with the identification of T. moeldenensis moeldenensis from the Talfan Farm locality, confirming that the lower Tre-wylan Beds are of similar age (early Cautleyan, Zone 1, upper Katian). Thus S. oliveae sp. nov. may be a useful Cautleyan indicator in Wales and S. clavifrons may have very much wider stratigraphical value as a Rawtheyan indicator.
Conclusions
The discovery of a new species of Staurocephalus, S. oliveae sp. nov., in the Sholeshook Limestone (equivalent to Ashgill early Cautleyan Zone 1, upper Katian, Upper Ordovician) of South West Wales represents the oldest known occurrence of the genus in Avalonia. Recognition of this Cautleyan species resurrects the potential usefulness of Staurocephalus clavifrons as an uppermost Katian (= Rawtheyan) and possibly Hirnantian indicator.
Acknowledgements
PMcD thanks Mark Tucker and Jean Cory (both Llanddowror, Carmarthenshire, UK) for allowing access to Talfan Farm and excavating an exploratory trench, without which collection of these and many other fossils from the Sholeshook Limestone would not have been possible. Olive Potts (St. Clears, Carmarthenshire, UK) is thanked for finding the first specimen of S. oliveae sp. nov. and for helping with the illustrations. We thank Matt Riley (SE), Neil Clark (GLAHM), and Josh Dixon (BIRUG) for facilitating access to specimens in their care through collections visits, arranging loans and photography. We are grateful to Leonid Popov (Amgueddfa Cymru-Museum Wales, Cardiff, UK) for providing a translation of Apollonov’s (1974) Russian description of S. djarikensis and to Caroline Buttler (Amgueddfa Cymru- Museum Wales, Cardiff, UK) for assistance with consolidation of specimens for latex casting. We thank Wei Xin (Nanjing Institute of Geology and Palaeontology, Chinese Academy of Science, China) and an anonymous reviewer for their very helpful comments. This paper is a contribution to IGCP Project 735 Rocks and the Rise of Ordovician Life.
References
Adrain, J.M. 2013. A synopsis of Ordovician trilobite distribution and diversity. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society of London, Memoir 38: 297–336. Crossref
Angelin, N.P. 1854. Palaeontologica Scandinavica I; Crustacea Formationis Transitionis, Fasc. 2 [in Russian], 21–92. Weigel, Lund.
Apollonov, M.K. 1974. Ashgillskie trilobity Kazakhstana. 136 pp. Akademiya Nauk Kazakh SSR, Alma Ata.
Barrande, J. 1846. Notice préliminaire sur le systême Silurien et les Trilobites de Bohême. 97 pp. Hirschfeld, Leipzig. Crossref
Barrs, C. 2013. Rugose corals from the Upper Ordovician Sholeshook Limestone of southwest Wales with an assessment of the coral affinities and biofacies. Geological Journal 48: 603–619. Crossref
Bergström, S.M., Xu, C., Gutiérrez-Marco, J.C., and Dronov, A. 2009. The new chronostratigraphic classification of the Ordovician System and its relations to major regional series and stages and to the δ13C chemostratigraphy. Lethaia 42: 97–107. Crossref
Bicknell, R.D.C. and Smith, P.M. 2021. Teratological trilobites from the Silurian (Wenlock and Ludlow) of Australia. The Science of Nature 108 (58): 1–11. Crossref
Cave, R. 1960. A new species of Tretaspis from south Wales. Geological Magazine 97: 334–337. Crossref
Chatterton, B.D.E. and Campbell, K.S.W. 1980. Silurian trilobites from near Canberra and some related forms from the Yass Basin. Palaeontographica A 167: 77–119.
Cocks, L.R.M. 2014. The Late Ordovician brachiopods of southern Pembrokeshire and adjacent south-western Wales. Special Papers in Palaeontology 91: 5–89.
Curtis, N.J. and Lane, P.D. 1997. The Llandovery trilobites of England and Wales. Part 1. Monographs of the Palaeontographical Society 151 (605): 1–50. Crossref
Curtis, N.J. and Lane, P.D. 1998. The Llandovery trilobites of England and Wales. Part 2. Monograph of the Palaeontographical Society 152 (608): 51–101. Crossref
Dean, W.T. 1971. The trilobites of the Chair of Kildare Limestone (Upper Ordovician) of eastern Ireland. Part 1. Monograph of the Palaeontographical Society 125 (531): 1–60. Crossref
Donovan, S.K. and Paul, C.R.C. 1985. Coronate echinoderms from the Lower Palaeozoic of Britain. Palaeontology 28: 527–543.
Edgecombe, G.D., Speyer, S.E., and Chatterton, B.D.E. 1988. Protaspid larvae and phylogenetics of encrinurid trilobites. Journal of Paleontology 62: 779–799.
Evitt, W.R. and Tripp, R.P. 1977. Silicified Middle Ordovician trilobites from the families Encrinuridae and Staurocephalidae. Palaeontographica A 157: 109–174.
Ferretti, A. and Barnes, C.R. 1997. Upper Ordovician conodonts from the Kalkbank limestone of Thuringia, Germany. Paleontologia 40: 15–42.
Ferretti, A., Bergström, S.M., and Barnes, C.R. 2014. Katian (Upper Ordovician) conodonts from South Wales. Palaeontology 57: 801–831. Crossref
Fortey, R.A. and Owens, R.M. 1997. Bubble-headed trilobites and a new olenid example. Palaeontology 40: 451–459.
von Freyberg, B. 1923. Die fauna und gliederung der Thüringer Untersilurs. Zeitschrift der Deutschen Geologischen Gesellschaft 74 (for 1922): 237–276.
Hammann, W. and Leone, F. 2007. Trilobites from the post-Sardic (Upper Ordovician) sequence of southern Sardinia. Part 2. Beringeria 38: 3–138.
Harrington, H.J. and Leanza, A.F. 1957. Ordovician Trilobites of Argentina. Special Publications, Department of Geology, University of Kansas 1: 1–276.
Holloway, D.J. 1980. Middle Silurian trilobites from Arkansas and Oklahoma, U.S.A. Part 1. Palaeontographica A 170: 1–85. Crossref
Ingham, J.K. 1966. The Ordovician rocks in the Cautley and Dent districts of Westmorland and Yorkshire. Proceedings of the Yorkshire Geological Society 35: 455–505. Crossref
Ingham, J.K. 1974. The Upper Ordovician trilobites from the Cautley and Dent districts of Westmorland and Yorkshire. Part 2. Monograph of the Palaeontographical Society 128 (538): 59–87. Crossref
Ingham, J.K. 1977. The Upper Ordovician trilobites from the Cautley and Dent districts of Westmorland and Yorkshire. Part 3. Monograph of the Palaeontographical Society 130 (546): 89–121. Crossref
Ingham, J.K. and Wright, A.D. 1970. A revised classification of the Ashgill Series. Lethaia 3: 233–242. Crossref
Ingham, J.K. and Wright, A.D. 1972. The North of England. In: A. Williams, I. Strachan, D.A. Bassett, W.T. Dean, J.K. Ingham, A.D. Wright, and H.B. Whittington (eds.), A Correlation of the Ordovician Rocks in the British Isles. Geological Society Special Report 3: 43–49.
Jaanuson, V. 1963. Classification of the Harjuan (Upper Ordovician) rocks of the mainland of Sweden. Geologiska Föreningens i Stockholm Förhandlingar 85: 110–144. Crossref
Kielan, Z. 1957. On the trilobite family Staurocephalidae. Acta Palaeontologica Polonica 2: 155–180.
Kielan, Z. 1960. Upper Ordovician trilobites from Poland and some related forms from Bohemia and Scandinavia. Palaeontologica Polonica 11: 1–198.
Kielan-Jaworowska, Z., Bergström, J., and Ahlberg, P. 1991. Cheirurina (Trilobita) from the Upper Ordovician of Västergötland and other regions of Sweden. Geologiska Föreningens I Stockholm Förhandlingar 113: 219–244. Crossref
King, W.B.R. 1923. The Upper Ordovician rocks of the south-western Berwyn Hills. Quarterly Journal of the Geological Society 79: 671–702. Crossref
Knüpfer, J. 1967. Zur Fauna und Biostratigraphie der Ordoviciums (Gräfenthaler Schichten) in Thüringen. Freiberger Forschungshefte C220: 1–119.
Kobayashi, T. and Hamada, T. 1985. Additional Silurian trilobites to the Yokokura-Yama fauna from Shikoku, Japan. Transactions and Proceedings of the Paleontological Society of Japan 139: 206–217.
Kraft, P., Linnemann, U., Mergl, M., Bruthansová, J., Laibl, L., and Geyer, G. 2023. Ordovician of the Bohemian Massif. Geological Society, London, Special Publications 532: 433–464. Crossref
Lanc, F.A., McDermott, P.D., and Paul, C.R.C. 2015. The identity of the British Ordovician cystoid “Hemicosmites rugatus Forbes”. Geological Journal 50: 1–16. Crossref
Lehnert, O., Frýda, J., Buggisch, W., Munnecki, A., Nützel, A., Křiž, J., and Manda, S. 2007. δ13C records across the Late Silurian Lau event: New data from middle palaeo-latitudes of northern peri-Gondwana (Prague Basin, Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology 245: 227–244. Crossref
Linnarsson, J.G.O. 1869. Om Vestergötlands cambriska och siluriska aflagringar. Kongliga Vetenskaps-Akademiens Handlingar 8: 1–89.
Loi, A., Cocco, F., Oggiano, G., Funedda A., Vidal, M., Ferretti, A., Leone, F., Barca, S., Naitza, S., Ghienne, J.-F., and Pillola, G.L. 2023. The Ordovician of Sardinia (Italy): from the “Sardic Phase” to the end-Ordovician glaciation, palaeogeography and geodynamic context. Geological Society, London, Special Publications 532: 409–431. Crossref
Marek, L. 1964. Shumardia Billings, 1862 a Staurocephalus Barrande, 1846 (Trilobita) v ceském ordoviku (Shumardia Billings, 1862 and Staurocephalus Barrande, 1846 (Trilobita) from the Czech Ordovician). Časopis Národního Muzea 133: 153–154.
Marr, J.E. 1907. On the Ashgillian Series. Geological Magazine 5 (4): 59–69. Crossref
Marr, J.E. and Roberts, T. 1885. The Lower Palaezoic rocks of the neighbourhood of Haverfordwest. Quarterly Journal of the Geological Society 41: 476–490. Crossref
McDermott, P.D. and Paul, C.R.C. 2015. Coronate echinoderms from the Ordovician of the Llanddowror area, South Wales. Geological Journal 50: 173–188. Crossref
McNamara, K.J. 1979. Trilobites from the Coniston Limestone Group (Ashgill Series) of the Lake District, England. Palaeontology 22: 53–92.
Nielsen, A.T., Ahlberg, P., Ebbestad, J.O., Hammer, Ø., Harper. D.A.T., Lindskog, A., Rasmussen, C.M. and Stouge, S. 2023. The Ordovician of Scandinavia: a revised regional stage classification. Geological Society, London, Special Publications 532: 267–315. Crossref
Nilsson, R. 1979. A boring through the Ordovician–Silurian boundary in western Scania, South Sweden. Sveriges Geologiska Undersökning C 766: 1–18.
Ogg, J.G., Ogg, G.M., and Gradstein, F.M. 2008. A Concise Geologic Timescale. vi+177 pp. Cambridge University Press, Cambridge.
Owen, A.W. 2007. Trilobite diversity in Avalonia prior to the end Ordovician extinction—the peak before the trough. Palaeogeography, Palaeoclimatology, Palaeoecology 245: 264–271.
Owen, A.W. and Bruton, D.L. 1980. Late Caradoc–early Ashgill trilobites of the central Oslo Region, Norway. Palaeontological Contributions from the University of Oslo 245: 1–63.
Owen, A.W. and Bruton, D.L. 2012. The only known cyclopygid-”atheloptic” trilobite fauna from North America: the upper Ordovician fauna of the Pyle Mountain Argillite and its palaeoenvironmental significance. Geological Magazine 149: 964–988. Crossref
Owen, A.W. and Romano, M. 2011. Deep shelf trilobite biofacies from the upper Katian (Upper Ordovician) of the Grangegeeth Terrane, eastern Ireland. Geological Journal 46: 416–426. Crossref
Owens, R.M. 2000. Arenig to Ashgill in South Wales. Geological Conservation Review Series 18: 125–180.
Paul, C.R.C. 1973. British Ordovician Cystoids. Part 1. Monograph of the Palaeontographical Society 127 (536): 1–64. Crossref
Paul, C.R.C. 1982. British Ordovician Cystoids. Part 2. Monograph of the Palaeontographical Society 136 (563): 65–152. Crossref
Paul, C.R.C. 1997. British Ordovician Cystoids. Part 3. Monograph of the Palaeontographical Society 151 (604): 153–213. Crossref
Peel, J.S. and McDermott, P.D. 2016. An association of problematic corals, crinoids and parasites from the Sholeshook Limestone Formation (Ordovician) of Wales. Geological Journal 51: 212–222. Crossref
Prantl, F. and Přibyl, A. 1947. Roztrídení nekterych ceskych cheiruridu. Sborník Národního Muzea v Praze. Rada B. Přírodní Vědy 3 (1): 1–44.
Price, D. 1971. The Stratigraphy and Trilobite Fauna of the Sholeshook Limestone of South Wales. 256 pp. Unpublished Ph.D. Thesis, University of London, London.
Price, D. 1973a. The age and stratigraphy of the Sholeshook Limestone of South-West Wales. Geological Journal 8: 225–246. Crossref
Price, D. 1973b. The Phillipsinella parabola–Staurocephalus clavifrons fauna and Upper Ordovician correlation. Geological Magazine 110: 535–541. Crossref
Price, D. 1974. Trilobites from the Sholeshook Limestone (Ashgill) of South Wales. Palaeontology 17: 112–116.
Price, D. 1977. Species of Tretaspis (Trilobita) from the Ashgill Series in Wales. Palaeontology 20: 763–792.
Price, D. 1980a. A revised age and correlation for the topmost Sholeshook Limestone Formation (Ashgill) of South Wales. Geological Magazine 117: 485–489. Crossref
Price, D. 1980b. The Ordovician trilobite fauna of the Sholeshook Limestone Formation of South Wales. Palaeontology 23: 839–887.
Price, D. 1981. Tretaspis radialis Lamont and allied species. Geological Magazine 118: 289–295. Crossref
Price, D. 1982. Calymene quadrata King, 1923 and allied species of trilobites from the Ashgill Series of North Wales. Geological Magazine 119: 57–66. Crossref
Salter, J.W. 1864. A monograph of the British trilobites from Cambrian, Silurian and Devonian formations. Part 1. Monograph of the Palaeontographical Society 16 (67): 1–80. Crossref
Shaw, F.C. 2000. Trilobites of the Králův Dvůr Formation (Ordovician) of the Prague Basin, Czech Republic: Vestník Ceského geologického ústavu 75: 371–404.
Šnajdr, M. 1980. New Silurian trilobites from the Barrandian area Czechoslovakia. Věstník Ústředního ústavu geologického 55: 105–110.
Štorch, P. and Mergl, M. 1989. Králodvor/Kosov boundary and the late Ordovician environmental changes in the Prague Basin (Barrandian area, Bohemia). Sbornik Geologických Věd 44: 117–153.
Størmer, L. 1930. Scandinavian Trinucleidae with special reference to Norwegian species and varieties. Skrifter utgitt av Det Norske Videnscaps-Akademi i Oslo I. Matematisk-Naturvidenskapelige Klasse 1930 4: 1–111.
Thomas, A.T. 1981. British Wenlock trilobites. Part 2. Monograph of the Palaeontographical Society 134 (559): 57–99. Crossref
Tomczykowa, E. 1987. Taksonomia Staurocephalidae Prantl et Přibyl, 1947 w związku z morfologią wyrostka przedniego. Biuletyn Instytutu Geologicznego 354: 183–196.
Trela, W. 2021. Eustatic and local tectonic impact on the Late Ordovician–early Silurian facies evolution at SW margin of peri-Baltica (the southern Holy Cross Mountains, Poland). Geological Magazine 158: 1472–1486. Crossref
Trela, W. 2023. Stratigraphy and sedimentary record of the Ordovician system in Poland: a review. Geological Society, London, Special Publications 532: 345–373. Crossref
Tullberg, S.A. 1882. Skånes graptoliter. I: Allman ofversigtofver de siluriska bildningarna i Skåne och jemforelse med ofriga kanda samtidiga aflagringar. Sveriges Geologiska Undersökning C 50: 1–44.
Vandenbroucke, T.R.A. 2008. An Upper Ordovician chitinozoan biozonation in British Avalonia (England and Wales). Lethaia 41: 275–294. Crossref
Vandenbroucke, T.R.A., Rickards, B., and Verniers, J. 2005. Upper Ordovician chitinozoan biostratigraphy from the type Ashgill area (Cautley district) and the Pus Gill section (Dufton district, Cross Fell Inlier), Cumbria, northern England. Geological Magazine 142: 783–807. Crossref
Vandenbroucke, T.R.A., Williams, M., Zalasiewicz, J.A., Davies, J.A., and Waters, R.A. 2008. Integrated Upper Ordovician graptolite-chitinozoan biostratigraphy of the Cardigan and Whitland areas, southwest Wales. Geological Magazine 145: 199–214. Crossref
Vaněk, J. and Valíček, J. 2002. New index of the genera, subgenera, and species of Barrandian trilobites Part C–D (Silurian and Devonian). Palaeontologia Bohemiae 8: 1–74.
Walch, J.E.I. 1771. Die Naturgeschichte der versteinerungen zur erläuterung der Knorrischen Sammlung von Merkwurdigkeiten der Natur. 235 pp. Felßecker, Nürnberg.
Weber, V.N. 1948. Trilobites of the Silurian deposits of the USSR. Lower Silurian trilobites [in Russian]. Monografii po Paleontologii SSSR 69: 1–111.
Weir, J.A. 1959. Ashgillian trilobites from Co. Clare, Ireland. Palaeontology 1: 369–383.
Weller, S. 1907. The palaeontology of the Niagran Limestone in the Chicago area. Bulletin of the Natural History Survey of the Academy of Science 4 (2): 163–281.
Whiteley, T.E., Kloc, G.J., and Brett, C.E. 2002. Trilobites of New York. An Illustrated Guide. xix + 380 pp. Cornell University Press, Ithaca.
Whittington, H.B. 1938. The geology of the district around Llansantffraid ym Mechain, Montgomeryshire. Quarterly Journal of the Geological Society 94: 423–457. Crossref
Whittington, H.B. 1965. A monograph of the Ordovician trilobites of the Bala area, Merioneth. Part 2. Monograph of the Palaeontographical Society 118 (504): 33–62. Crossref
Whittington, H.B. 1968. A monograph of the Ordovician trilobites of the Bala area, Merioneth. Part 4. Monograph of the Palaeontographical Society 118 (504): 98–138. Crossref
Zalasiewicz, J., Rushton, A.W.A., and Owen, A.W. 1995. Late Caradoc graptolite faunal gradients across the Iapetus Ocean. Geological Magazine 132: 611–617. Crossref
Zhou, Z. and Zhou, Z. 1982. An Ashgill (Rawtheyan) trilobite faunule from Ejin Qi, Nei Monggol (Inner Mongolia) [in Chinese with English summary]. Acta Palaeontologica Sinica 21: 659–671.
Acta Palaeontol. Pol. 69 (1): 57–72, 2024
https://doi.org/10.4202/app.01120.2023