

Osteology of the dorsal vertebrae of the giant titanosaurian sauropod dinosaur Dreadnoughtus schrani from the Late Cretaceous of Argentina
KRISTYN K. VOEGELE, MATTHEW C. LAMANNA, and KENNETH J. LACOVARA
Voegele, K.K., Lamanna, M.C., and Lacovara K.J. 2017. Osteology of the dorsal vertebrae of the giant titanosaurian sauropod dinosaur Dreadnoughtus schrani from the Late Cretaceous of Argentina. Acta Palaeontologica Polonica 62 (4): 667–681.
Many titanosaurian dinosaurs are known only from fragmentary remains, making comparisons between taxa difficult because they often lack overlapping skeletal elements. This problem is particularly pronounced for the exceptionally large-bodied members of this sauropod clade. Dreadnoughtus schrani is a well-preserved giant titanosaurian from the Upper Cretaceous (Campanian–Maastrichtian) Cerro Fortaleza Formation of southern Patagonia, Argentina. Numerous skeletal elements are known for Dreadnoughtus, including seven nearly complete dorsal vertebrae and a partial dorsal neural arch that collectively represent most of the dorsal sequence. Here we build on our previous preliminary description of these skeletal elements by providing a detailed assessment of their serial positional assignments, as well as comparisons of the dorsal vertebrae of Dreadnoughtus with those of other exceptionally large-bodied titanosaurians. Although the dorsal elements of Dreadnoughtus probably belong to two individuals, they exhibit substantial morphological variation that suggests that there is minimal, if any, positional overlap among them. Dreadnoughtus therefore preserves the second-most complete dorsal vertebral series known for a giant titanosaurian that has been described in detail, behind only that of Futalognkosaurus. The dorsal sequence of Dreadnoughtus provides valuable insight into serial variation along the vertebral column of these enormous sauropods. Such variation includes the variable presence of divided spinodiapophyseal laminae and associated spinodiapophyseal fossae. Given that dorsal vertebrae are the only elements that overlap between known remains of most giant titanosaurian taxa, the dorsal series of Dreadnoughtus provides a means to directly compare the morphologies of these sauropods. The dorsal vertebrae of Dreadnoughtus and Futalognkosaurus have dorsoventrally narrow transverse processes, unlike the condition in Puertasaurus. Further, Dreadnoughtus and Argentinosaurus have ventromedially inclined prezygapophyses, whereas Futalognkosaurus has almost horizontal prezygapophyses. The continued inclusion of new, well-represented skeletons of titanosaurians such as Dreadnoughtus in phylogenetic and functional morphological studies will aid in deciphering the interrelationships and paleobiology of Titanosauria.
Key words: Sauropoda, Titanosauria, Dreadnoughtus, dorsal vertebrae, Cretaceous, Cerro Fortaleza Formation, Argentina.
Kristyn K. Voegele [voegele@rowan.edu] and Kenneth J. Lacovara [lacovara@rowan.edu] (corresponding author), Department of Geology, Rowan University, 201 Mullica Hill Road, Glassboro, NJ 08028, USA.
Matthew C. Lamanna [lamannam@carnegiemnh.org], Section of Vertebrate Paleontology, Carnegie Museum of Natural History, 4400 Forbes Avenue, Pittsburgh, PA 15213, USA.
Received 26 May 2017, accepted 18 October 2017, available online 16 November 2017.
Copyright © 2017 K.K. Voegele et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Titanosauria is a diverse clade that includes what are arguably the largest-bodied and the smallest known sauropods (Wilson 2006). Unfortunately, however, many titanosaurians are known only from very incomplete remains (Upchurch et al. 2004). As a consequence of the large body sizes of many representatives of this group, preservational biases—such as the amount of sediment needed to fully bury a large sauropod—result in their frequently fragmentary preservation (González Riga and Astini 2007; Casal et al. 2014). The holotype of Dreadnoughtus schrani (MPM-PV 1156) is an exception, being roughly twice as skeletally complete (Lacovara et al. 2014; Ullmann and Lacovara 2016) as the next most completely represented giant titanosaurian, Futalognkosaurus (Calvo et al. 2007a, b; Calvo 2014). Dreadnoughtus is consequently an important taxon for understanding the anatomy of these dinosaurs.
Dreadnoughtus was discovered in the Upper Cretaceous (Campanian–Maastrichtian) Cerro Fortaleza Formation of Santa Cruz Province in southern Patagonia, Argentina. A total of 145 bones were recovered from the partially articulated holotype and the somewhat smaller paratype (MPM-PV 3546), including seven mostly complete dorsal vertebrae and one dorsal neural arch fragment (Lacovara et al. 2014). Of what are frequently regarded (by, e.g., Benson et al. 2014; Lacovara et al. 2014; González Riga et al. 2016; Ullmann and Lacovara 2016; Carballido et al. 2017) as the largest titanosaurian taxa yet described (i.e., “Antarctosaurus” giganteus, Argentinosaurus, Dreadnoughtus, Elaltitan, Futalognkosaurus, Notocolossus, Paralititan, Patagotitan, and Puertasaurus), only Futalognkosaurus (Calvo et al. 2007a, b) and Dreadnoughtus have more than 70% of their dorsal series described in detail. The only bones that overlap between all of these largest taxa are axial elements. Dorsal vertebrae representing up to nine serial positions may be preserved, collectively, between the holotype and the most complete paratype of the recently described giant titanosaur Patagotitan (Carballido et al. 2017), but these await detailed description. Six dorsal vertebrae are described for Argentinosaurus (Bonaparte and Coria 1993; a seventh undescribed posterior dorsal is reported by Salgado and Powell 2010), whereas Elaltitan (Powell 2003; Mannion and Otero 2012) and Paralititan (Smith et al. 2001; MCL personal observation) each include three dorsal vertebrae, and Notocolossus and Puertasaurus each include only one dorsal vertebra (Novas et al. 2005; González Riga et al. 2016). The preserved axial remains of “Antarctosaurus” giganteus include only two incomplete caudal vertebrae, making it the only giant titanosaurian to be preserved without dorsal vertebral remains (Huene 1929). Despite the limited information available on the dorsal vertebral series in most gigantic titanosaurian taxa, as well as the positional differences that exist between many of the preserved vertebrae, these elements provide the only means to directly compare the morphologies of eight of the nine most massive titanosaurians. Among other skeletal elements, caudal vertebrae exhibit the next greatest degree of preservation, with all giant titanosaurians except Argentinosaurus preserving at least one of these elements (Huene 1929; Bonaparte and Coria 1993; Smith et al. 2001; Powell 2003; Novas et al. 2005; Calvo et al. 2007a, b; Mannion and Otero 2012; Lacovara et al. 2014; González Riga et al. 2016). The appendicular skeletons of these sauropods preserve even less overlap, with the humerus (i.e., those of Dreadnoughtus, Elaltitan, Futalognkosaurus, Notocolossus, Paralititan, and Patagotitan) being the most commonly represented element (Smith et al. 2001; Powell 2003; Mannion and Otero 2012; Calvo 2014; Lacovara et al. 2014; González Riga et al. 2016; Ullmann and Lacovara 2016; Carballido et al. 2017).
Sauropod dorsal vertebrae are thought to exhibit phylogenetically significant characters as well as variation at the individual, species, and higher taxonomic levels (Wilson 2012). Because a broad consensus on the phylogenetic relationships of most titanosaurian taxa has not yet been reached (Upchurch et al. 2004; Curry Rogers 2005; Carballido and Sander 2014; Salgado et al. 2015; Poropat et al. 2015, 2016; González Riga et al. 2016; Gorscak and O’Connor 2016), well-preserved dorsal vertebral series such as that of Dreadnoughtus can provide data important for resolving titanosaurian relationships (Wilson et al. 2011). Here, we provide a detailed description of the dorsal vertebrae of Dreadnoughtus schrani, building upon their initial description by Lacovara et al. (2014). We also discuss the probable serial positions of these vertebrae and compare these elements to those of other titanosaurian taxa, especially other gigantic forms.
Institutional abbreviations.—MPM-PV, Museo Padre Molina, Paleontología de Vertebrados, Río Gallegos, Argentina.
Other abbreviations.—ACDL, anterior centrodiapophyseal lamina; ACPL, anterior centroparapophyseal lamina; aPCDL, accessory posterior centrodiapophyseal lamina; a-SPDL, anterior ramus of spinodiapophyseal lamina; CDF, centrodiapophyseal fossa; CPAF, centroparapophyseal fossa; CPOF, centropostzygapophyseal fossa; CPOL, centropostzygapophyseal lamina; CPRF, centroprezygapophyseal fossa; CPRL, centroprezygapophyseal lamina; PACDF, parapophyseal centrodiapophyseal fossa; PADF, paradiapophyseal fossa; PCDL, posterior centrodiapophyseal lamina; PCDL-F, posterior centrodiapophyseal lamina fossa; PCPL, posterior centroparapophyseal lamina; POCDF, postzygapophyseal centrodiapophyseal fossa; PODL, postzygodiapophyseal lamina; POSDF, postzygapophyseal spinodiapophyseal fossa; POSL, postspinal lamina; PPDL, paradiapophyseal lamina; PRSDF, prezygapophyseal spinodiapophyseal fossa; PRSL, prespinal lamina; p-SPDL, posterior ramus of spinodiapophyseal lamina; SPDL, spinodiapophyseal lamina; SPDL-F, spinodiapophyseal lamina fossa; SPOF, spinopostzygapophyseal fossa; SPOL, spinopostzygapophyseal lamina; SPRF, spinoprezygapophyseal fossa; SPRL, spinoprezygapophyseal lamina; TPOL, intrapostzygapophyseal lamina; TPRL, intraprezygapophyseal lamina.
Material and methods
Below, the dorsal vertebrae of Dreadnoughtus (MPM-PV 1156?-4 through MPM-PV 1156-11) are described in their hypothesized serial sequence, from anterior to posterior. The descriptive terminology used here follows Wilson (1999, 2012) for vertebral laminae and Wilson et al. (2011) for fossae. Our assignments of the serial position of each vertebra were designated based on comparisons to taxa known from complete, articulated dorsal series (e.g., Futalognkosaurus, Calvo et al. 2007a, b; Opisthocoelicaudia, Borsuk-Białynicka 1977; Overosaurus, Coria et al. 2013; Trigonosaurus, Powell 1987, Campos et al. 2005). The disarticulated, scattered disposition of the dorsal vertebrae in the Dreadnoughtus quarry obscures the assignment of individual vertebrae to the holotype or paratype specimens, except in the cases of MPM-PV 1156-6 and MPM-PV 1156-11, which can be definitively assigned to the holotype. This results in a hypothetical anteroposterior sequence of dorsals, similar to that proposed for the African titanosaurian Malawisaurus, in which dorsal vertebrae of multiple individuals were also discovered in disarticulated condition (Gomani 2005). If a particular vertebra cannot be definitively assigned to a specific individual, it is herein designated as “MPM-PV 1156?-#” following Lacovara et al. (2014). The last number of the specimen number refers to the sequential anteroposterior order of skeletal elements of the holotype of Dreadnoughtus, beginning with the skull, and hence does not directly correspond to hypothesized serial position in the axial column. Although titanosaurians reportedly might have had up to 11 dorsal vertebrae (Borsuk-Białynicka 1977; see also discussion in Lacovara et al. 2014), Dreadnoughtus is predicted to have had ten, as in most other titanosaurians (Powell 1987; Campos et al. 2005; Calvo et al. 2007a, b; Curry Rogers 2009; Coria et al. 2013).
Systematic palaeontology
Sauropoda Marsh, 1878
Neosauropoda Bonaparte, 1986
Titanosauria Bonaparte and Coria, 1993
Genus Dreadnoughtus Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014
Type species: Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014; Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian.
Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014
Type material: Holotype (MPM-PV 1156) includes a fragment of maxilla, one tooth, a posterior cervical vertebra (~9th), up to eight dorsal vertebrae, the partial sacrum, the nearly complete caudal series, both sternal plates, the left scapula, coracoid, humerus, ulna, and radius, the complete pelvis, the left femur, both tibiae, the left fibula and astragalus, the right metatarsals I and II, and the ungual of right pedal digit I. Paratype (MPM-PV 3546) includes a partial anterior cervical vertebra (~4th), six anterior caudal vertebrae, both ilia, pubes, and ischia, and the deformed left femur.
Type horizon: A crevasse splay deposit in an exposure of the Cerro Fortaleza Formation, regarded as middle–late Campanian to early Maastrichtian in age, based on the ages of underlying and overlying strata in the region (Egerton 2011).
Type locality: Northern part of the valley of the Río La Leona between Lago Argentino and Lago Viedma, Santa Cruz Province, southern Patagonia, Argentina (Lacovara et al. 2014).
Material.—Type material only.
Description.—General Characteristics: Unless otherwise noted, the dorsal vertebral structures of Dreadnoughtus generally exhibit bilateral (i.e., left-right) symmetry. We provide a thorough description of each vertebra, only omitting select common structures that exhibit no noteworthy features beyond merely being present. If present, any such structures are labeled in the figures. Dorsal vertebral centra are strongly opisthocoelous and camellate, with well-developed, undivided lateral pneumatic fossae that are generally more anteriorly placed in more posteriorly-positioned vertebrae (Fig. 1, SOM: fig. S1 in the Supplementary Online Material available at http://app.pan.pl/SOM/app62-Voegele_etal_SOM.pdf). In some vertebrae, taphonomic deformation may have accentuated the depth of the lateral pneumatic fossae. Most dorsal vertebral centra (as preserved) are noticeably wider than tall (Figs. 2, 3 and SOM: figs. S2, S3); nevertheless, taphonomic distortion has altered the width of several vertebrae. In general, the centra exhibit no other conspicuous features, such as a ventral keel or ventrolateral ridges (e.g., Fig. 4B). The ventral surfaces range from flat to transversely convex due to differing degrees of taphonomic deformation. Despite being, in some cases, distorted, the transverse processes were clearly directed laterally or slightly anterolaterally (Fig. 1). The seven mostly complete dorsal vertebrae have neural arch pedicels that are situated at or near the anterior margins of their respective centra but that are inset from the posterior margins (Fig. 1). The distance of the neural arch pedicel from the posterior margin of the centrum is obscured by the lack of preservation of this margin in some vertebrae. The heights of the neural arch pedicels (sensu Lacovara et al. 2014) are nearly equal to those of the neural spines (with the neural spines occasionally being taller; Fig. 1); this appears to have been only minimally affected by taphonomic deformation. Hyposphene-hypantrum articulations are not present in the dorsal vertebrae of Dreadnoughtus. Most of the neural spines are oriented posterodorsally. Measurements of the dorsal vertebrae of Dreadnoughtus are provided in Lacovara et al. (2014: supplementary table 1).
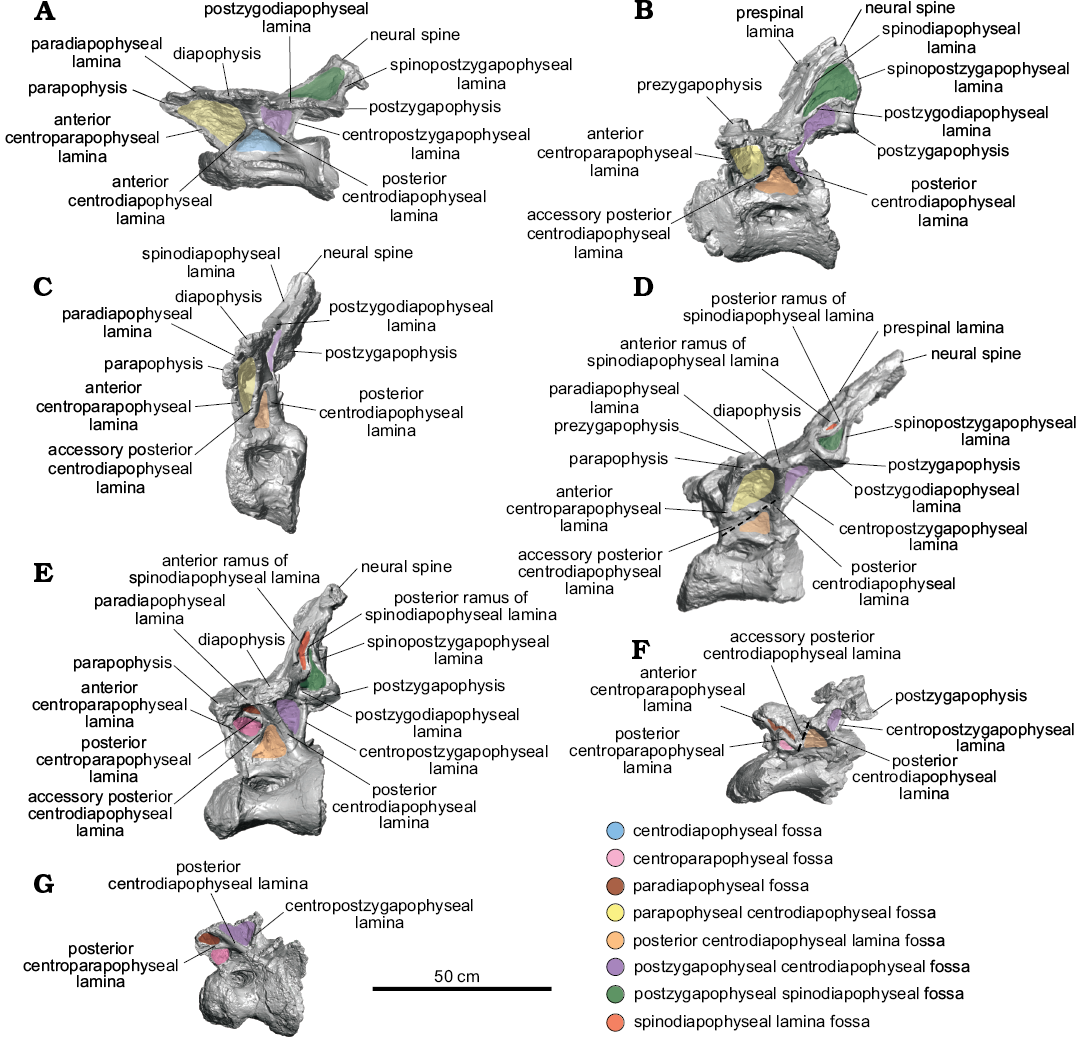
Fig. 1. Nearly complete dorsal vertebrae of titanosaurian sauropod Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014, from Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian; in left lateral view. A. MPM-PV 1156?-4, estimated as the ~4th dorsal vertebra. B. MPM-PV 1156?-5, ~5th. C. MPM-PV 1156-6, ~6th. D. MPM-PV 1156?-8, ~7th. E. MPM-PV 1156?-9, ~8th. F. MPM-PV 1156?-10, ~9th. G. MPM-PV 1156-11, ~10th. Dashed lines indicate fractured neural arch laminae. Colours indicate neural arch fossae.
Dorsal vertebra MPM-PV 1156?-4: This vertebra is essentially complete (Figs. 1A, 2A, 3A, 4, 5A). Of the centrum, only the posteroventral edge and the posterior end of the right lateral margin are incomplete, and of the neural arch, only the anterolateral portion of the right prezygapophysis and the edge of the left ACPL are not preserved. Taphonomic deformation has resulted in significant dorsoventral compression of both the centrum and neural arch, obscuring the neural canal. To a lesser extent, there has also been shear in the left lateral direction, as the neural arch is no longer transversely centered over the centrum. There has been no apparent anteroposterior compression and no observable twisting about the dorsoventral axis (i.e., the transverse processes are still mediolaterally oriented; Figs. 2A, 3A, 4).
Of the dorsal vertebrae preserved, MPM-PV 1156?-4 is considered the anterior-most in serial position, approximately the fourth dorsal vertebra (Lacovara et al. 2014). The parapophyses are located on the neural arch and, as preserved, are level with the diapophyses (Figs. 1A, 2A, 4A). This situation, which is unusual for an anterior dorsal vertebra, is potentially a consequence of uneven amounts of dorsoventral compression acting on the anterior and posterior regions of the vertebra. Even accounting for compression, the centrum appears longer anteroposteriorly than it is tall dorsoventrally, and the transverse processes are wide and flat (Figs. 1A, 2A, 3A, 4). As preserved, the lateral pneumatic fossae of MPM-PV 1156?-4 extend almost the entire length of the centrum (Fig. 1A).
The CPRLs are observed as single (i.e., undivided) broad ridges, and, due to deformation, are difficult to distinguish from the thin ACPLs. Taphonomic shear has rendered the TPRL asymmetrical, with the right side being slightly shorter than the left (Fig. 2A). On the posterior face of the neural arch, the CPOLs overlap, presumably due to compression (Fig. 3A). The TPOL is faint and angled anteroventrally. The CPRF, which is bounded by the CPRLs and TPRL, and the CPOF, which is bounded by the CPOLs and TPOL, are difficult to distinguish because of compression, but both are probably present. Both the ACDLs and PCDLs are prominent, and each shows evidence of dorsoventral compression in the form of partly overlapping broken segments. These laminae are parallel for the middle third of their lengths but are not fused. On the ventral side of the transverse process, they diverge and contact the diapophysis at separate locations. The CDFs are triangular and smaller than the PACDFs and POCDFs. The POCDFs are deep and approximate a nearly right triangle in shape (Fig. 1A). In contrast to the situation in more posterior dorsal vertebrae of Dreadnoughtus, in MPM-PV 1156?-4 the PACDFs are present because PCPLs are not present. The current shape of the PACDFs is a result of dorsoventral compression. Overall, the PACDFs are rectangular and shallow, although they are deeper anteriorly. The PODLs form the posterolateral edges of the transverse processes, extending posteromedially and then posterolaterally to the postzygapophyses, and defining an embayment in dorsal view (Figs. 4A, 5A).
The dorsal portion of the neural spine is posteroventrally compressed, as indicated by the presence of external, “finished” bone (as opposed to trabecular bone) on the preserved dorsal surface of the spine (Fig. 4A). It is possible that the dorsal-most tip of the neural spine is not preserved, but this is difficult to ascertain. As preserved, the strongly posterodorsal orientation of the neural spine places the postzygapophyses posterior to the centrum. This was probably the condition prior to deformation, but the degree of compression and the presence of both plastic and brittle deformation imply that slight posterior displacement of the neural spine and postzygapophyses cannot be ruled out. Unlike the condition in all other preserved vertebrae of Dreadnoughtus, thin left and right SPRLs are present and join the prominent PRSL about 4 cm dorsal to the base of the neural spine (Fig. 4A). Although it is prominent along most of its length, the PRSL is less distinct near the preserved end of the neural spine. The SPRLs bound a small SPRF. Both SPDLs are most prominent in the middle of their extent (Fig. 4A). Taphonomic deformation renders it difficult to determine if the SPDLs and/or PRSL extend to the tip of the neural spine. The PRSDF and POSDF are shallow near the diapophysis but become deeper near the transverse midline of the vertebra. The shallow, ventrally concave SPOFs are divided by the robust POSL.
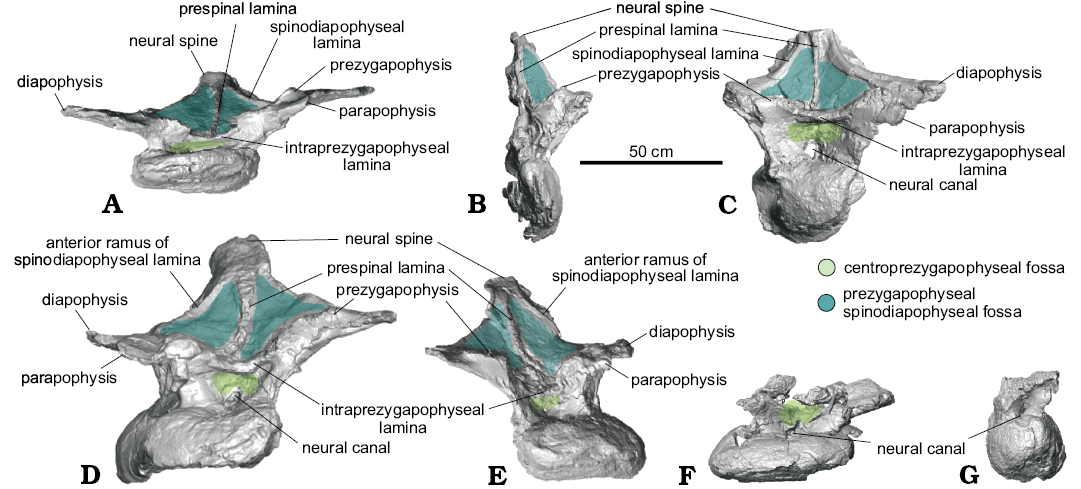
Fig. 2. Nearly complete dorsal vertebrae of titanosaurian sauropod Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014, from Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian; in anterior view. A. MPM-PV 1156?-4, estimated as the ~4th dorsal vertebra. B. MPM-PV 1156?-5, ~5th. C. MPM-PV 1156-6, ~6th. D. MPM-PV 1156?-8, ~7th. E. MPM-PV 1156?-9, ~8th. F. MPM-PV 1156?-10, ~9th. G. MPM-PV 1156-11, ~10th. Colours indicate neural arch fossae.
Dorsal vertebra MPM-PV 1156?-5: MPM-PV 1156?-5 has been taphonomically crushed transversely (Figs. 1B, 2B, 3B, 5B). Although a significant portion is missing from the right side of the neural arch, the majority of the right side of the centrum is preserved, with the exception of the ventral portion, albeit compressed medially. The right anterolateral face of the neural arch is not preserved, including the PRSDF and the right prezygapophysis, making the presence of a CPRF difficult to determine. The left side of the vertebra is mostly complete, but the lateral portions of the parapophysis and diapophysis, including associated parts of the ACPL, PODL, and PCDL are not preserved. Furthermore, the center of the deep, cone-shaped left PACDF is not preserved. Surrounding this fossa, the ACPL and aPCDL are missing where they would intersect the centrum. In addition, the ACPL is missing portions of its lateral edge along its preserved length. As best observed in posterior view, the right side of both postzygapophyses and the majority of the left CPOL are not preserved. On the right side of the vertebra only two laminae are preserved: the CPOL and a portion of the undivided SPOL. The dorsal part of the neural spine, dorsal to the postzygapophyses, is mostly complete but severely folded posteromedially such that the postzygapophyses are nearly in contact with one another (Fig. 3B).
Both the aPCDL and the PCDL are clearly visible and they intersect at the base of the ventral side of the transverse process (Fig. 1B). Between these laminae is a shallow, triangular PCDL-F. Transverse compression has resulted in breakage and offset of the left ACPL, and only the dorsal portion (with slight medial twisting due to compression) of the left CPRL is preserved. Unlike in MPM-PV 1156?-4, the right CPOL is thin. This lamina exhibits a mediolateral twist such that, in posterior view, its lateral surface is visible in its dorsal extent and its medial surface is visible in its ventral extent (Fig. 3B). The left PODL arcs posterodorsally above the postzygapophysis before merging with it (Figs. 3B, 5B). Between this lamina and the SPOL, just dorsal to where they contact the postzygapophysis, is a presumed soft tissue attachment surface (Fig. 1B). This feature can only be identified on the left side of the vertebra owing to lack of preservation on the right side. There is no indication of a TPOL, but the distortion of the neural spine makes this difficult to confirm. If the TPOL is truly absent, then the CPOF and SPOFs would be confluent.
The POSL is clearly visible at almost the center of the folded neural spine (Fig. 3B). The PRSL appears prominently on the anterior face of the neural spine, but its prominence may have been exaggerated by taphonomic distortion. Lack of preservation prohibits determination of the presence of the SPRLs, and if they are present, whether they join the PRSL (Fig. 2B). Thus, the presence of a SPRF is unknown. Due to the aforementioned distortion, the left SPDL appears as a ridge of bone instead of a distinct lamina (Figs. 1B, 5B).
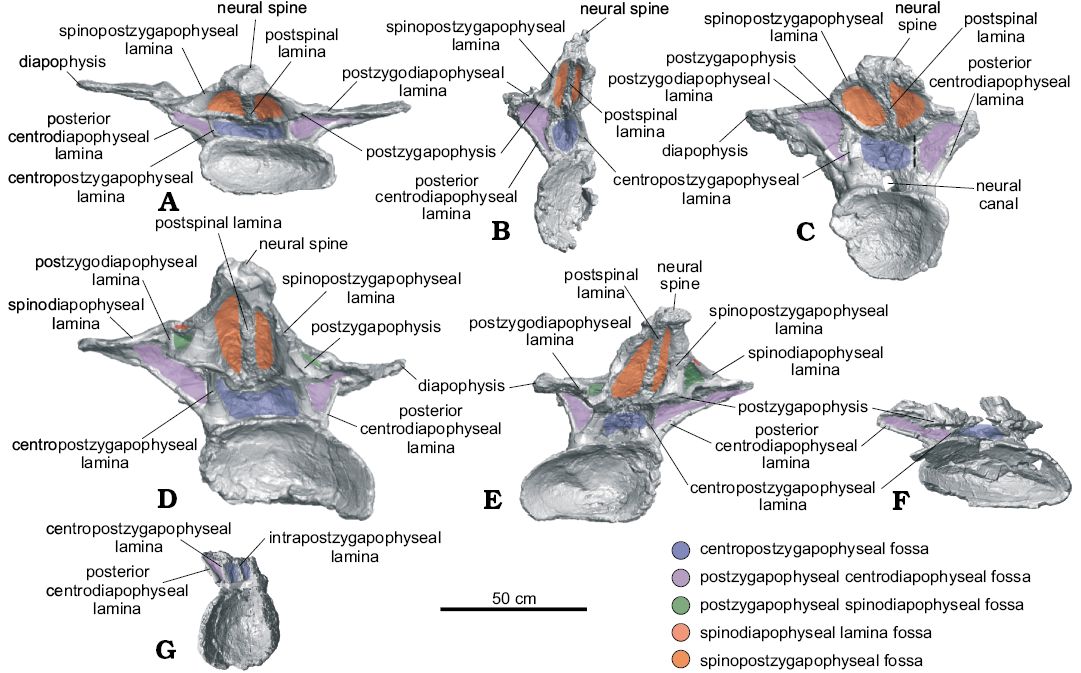
Fig. 3. Nearly complete dorsal vertebrae of titanosaurian sauropod Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014, from Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian; in posterior view. A. MPM-PV 1156?-4, estimated as the ~4th dorsal vertebra. B. MPM-PV 1156?-5, ~5th. C. MPM-PV 1156-6, ~6th. D. MPM-PV 1156?-8, ~7th. E. MPM-PV 1156?-9, ~8th. F. MPM-PV 1156?-10, ~9th. G. MPM-PV 1156-11, ~10th. Dashed lines indicate fractured neural arch laminae. Colours indicate neural arch fossae.
Dorsal vertebra MPM-PV 1156-6: MPM-PV 1156-6 is mostly complete but strongly anteroposteriorly compressed (Figs. 1C, 2C, 3C, 5C). The right transverse process is incomplete, missing the parapophysis, diapophysis, and PPDL. Also, the lateral margins and the tip of the neural spine, a portion of the left side of the posterior part of the centrum, and a ventral section of the right CPOL are not preserved. There is evidence of minor dorsoventral compaction; for instance, the left CPOL is broken into overlapping segments (Fig. 3C). This deformation prohibits the determination of the placement of the zygapophyses in relation to the centrum.
Although there is a broken ridge of bone in the location of the PRDL on the left side of the vertebra, this feature is most likely a result of compression of the prezygapophysis as it lacks any indication of a smooth external surface and does not extend the entire length from the prezygapophysis to the diapophysis (Fig. 2C). The presence of a left PPDL is probable, though it has been compressed into a mostly obscured cleft between the parapophysis and diapophysis (Fig. 1C). The aPCDLs become thinner as they extend dorsally, contrasting with the PCDLs which are thick along their entire length. These two laminae bound the deep PCDL-F and appear to merge before contacting the diapophysis (Fig. 1C). The PACDF and POCDF are large and deep.
The left diapophysis is angled anteriorly, a condition that is most likely due to anteroposterior compression (Fig. 1C). Although taphonomic deformation may have accentuated the curvature of the PODLs, these laminae likely had the same curved morphology described above in MPM-PV 1156?-5, in which they extend posterodorsally onto the neural spine dorsal to the postzygapophyses before changing direction to intersect the postzygapophyses (Fig. 5C). The SPDLs are thicker on this vertebra than in any of the other preserved vertebrae (Figs. 1C, 2C, 5C). The CPRLs and CPOLs are undivided and wide, and they respectively delimit a shallow CPRF and CPOF, which are difficult to observe due to compression. The TPRL is well defined as a thick bar, whereas the TPOL is thin and descends ventromedially from the postzygapophyses (Fig. 3C).
An oval, textured area (partly obscured by putty applied during preparation) is present on the left side at the junction of the neural spine and transverse process, just dorsal to the postzygapophysis. This feature might represent a soft tissue attachment site similar to that described on the preceding MPM-PV 1156?-5. This region is not preserved on the right side. A second, faint, possible soft tissue attachment site is present on the dorsal surface of the left transverse process, directly dorsal to the diapophysis. This potential scar is semicircular in outline and exhibits mediolaterally-oriented striations. A scar is readily apparent at this location in more posteriorly-positioned dorsal vertebrae (see below; Fig. 6B). As with the potential attachment site on the lateral aspect of the neural spine, this scar is only visible on the left side of the vertebra, since these structures are not preserved on the right.
This vertebra exhibits small, presumably pneumatic cavities on the posterior face of the neural spine on both sides of the robust POSL (Fig. 3C). These cavities are also visible on the dorsal surface of the right prezygapophysis (Fig. 2C). It is likely that these pneumatic cavities were exposed by abrasion prior to burial or during the preparation process. The PRSL extends the entire preserved length of the anterior surface of the neural spine (Fig. 2C); it is unknown how far it would have extended onto the missing dorsal-most portion. Judging from its consistent prominence over the preserved length of the neural spine, it is plausible that the PRSL extended to the tip of the neural spine, or nearly so. The SPOFs, PRSDFs, and POSDFs are shallow, potentially as a result of taphonomic deformation. This possibility is supported by the fact that the PRSDFs are relatively deeper near the TPRL.
Dorsal vertebra MPM-PV 1156?-7: MPM-PV 1156?-7 consists of a fragment of neural arch (Fig. 7). It preserves two articular surfaces that are connected by laminae, forming triangular fossae. The most reasonable identifications for these articular surfaces are the left diapophysis and postzygapophysis. Assuming these identifications are correct, the CPOL, PCDL, and PODL are preserved, as is a portion of the SPDL. The SPDL might show the beginning of a bifurcation into a- and p-SPDLs (Fig. 7A). Based on the preserved geometry of the laminae in this fragment, it likely is from a middle or posterior dorsal vertebra. Overlap precludes it from belonging to any of the other preserved vertebrae of Dreadnoughtus.
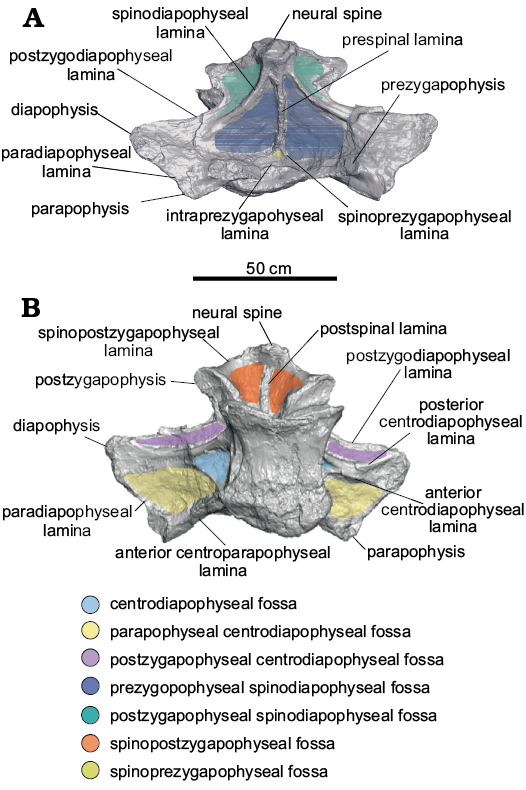
Fig. 4. Most anterior dorsal vertebra preserved of titanosaurian sauropod Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014, MPM-PV 1156?-4 (estimated as the ~4th dorsal vertebra), from Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian; in dorsal (A) and ventral (B) views. Colours indicate neural arch fossae.
Dorsal vertebra MPM-PV 1156?-8: In general, the left side of this vertebra is more complete than the right, but both parapophyses and diapophyses are lacking their lateral-most margins (Figs. 1D, 2D, 3D, 5D, 6B). The majority of the posterior border of the centrum is incomplete. Several segments of the PRSL, a lateral portion of the left CPRL immediately ventral to the left prezygapophysis, and the tip of the neural spine are not preserved. Although the right aPCDL is distinct, the left aPCDL is not preserved.
Overall, MPM-PV 1156?-8 exhibits left lateral shear of the centrum and neural arch in relation to the sagittal axis (Figs. 2D, 3D). The sinuous shape of the left SPDL (Fig. 1D) and breakage of the right ACPL suggest the vertebra might also have been subject to dorsoventral compression in addition to lateral shearing. Shearing has also displaced the neural arch such that the left zygapophyses are dorsolateral to the centrum. The transverse processes have been offset such that the right is positioned more anteroventrally than the left (Figs. 2D, 3D). Deformation and perhaps a lack of preservation have resulted in a rough knob of bone being situated in the location of the right prezygapophyseal articular surface (Fig. 2D).
The parapophysis is at essentially the same dorsoventral level as the diapophysis (Fig. 1D). The left transverse process has a well-defined attachment scar medial to the diapophysis (Fig. 6B). This scar is similar in placement and shape to those on MPM-PV 1156-6 and MPM-PV 1156?-9.
As in the two preceding vertebrae, the PODLs of this vertebra also extend posterodorsally just above the postzygapophyses before curving sharply ventrally to meet these latter structures (Figs. 1D, 5D). The degree of this curvature increases from MPM-PV 1156?-5 to MPM-PV 1156-6, then, in MPM-PV 1156?-8, the PODLs are nearly straight before abruptly curving ventrally to intersect the postzygapophyses (Fig. 5B–D). Taphonomic deformation cannot fully account for the shape of the PODLs in any of these vertebrae; therefore, their curvature is likely a genuine feature. As preserved, the CPRFs are small and shallow, with the left CPRF being deeper. The PPDLs are short and better defined on the left side (Fig. 1D). As in MPM-PV 1156-6, the TPOL is thin, faint, and directed ventromedially. The CPOFs are shallow and wide. The PACDFs are deep, and the left appears to be confluent with the PCDL-F because the left aPCDL is not preserved (Fig. 1D). On the right side, the PCDL-F is shallower than the PACDF and POCDF, and forms a ventrally pointing triangle in lateral view.
Both SPDLs are divided into a- and p-SPDLs, creating small SPDL-Fs (which are shallow and difficult to distinguish except near the split) that are located dorsal to the deep POSDFs (Figs. 1D, 5D). The SPDLs are better preserved near the transverse processes and on the left side of the vertebra. The left p-SPDL has been compressed into the POSDF. There is an articulation or attachment surface where each p-SPDL merges with each single, broad SPOL on the ventrolateral edges of the neural spine. This scar is more visible on the left side (Fig. 1D). Deformation makes it difficult to ascertain, but the SPDLs and PODLs might merge for a short distance along the posterior edge of the transverse process; however, these laminae are clearly separate ventral to the ventral margin of the SPDL-F. Since the neural spine is mostly complete, it is probable that the PRSL extended to, or very nearly to, the apex of the spine. The large, poorly defined PRSDFs surround the PRSL and are deepest where that lamina meets the TPRL (Fig. 2D). The SPOFs are shallow and dorsoventrally elongate. Although the POSL is not well preserved, it appears to extend dorsally from the TPOL and to become prominent near the level of the postzygapophyses. The POSL is missing the majority of its external surface but seems to have been thick.
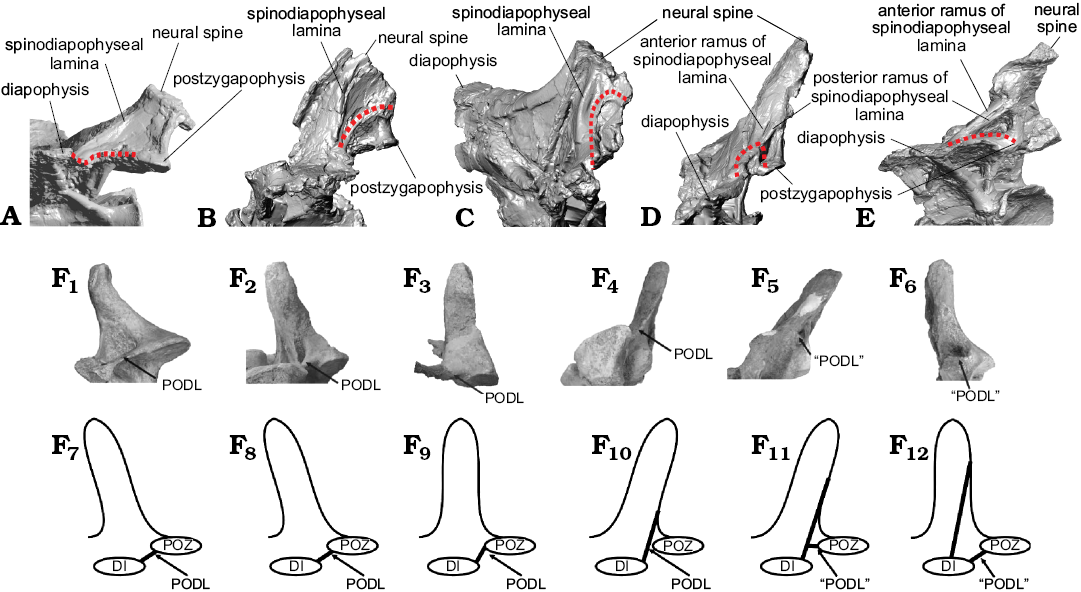
Fig. 5. Comparison of lamina capture in dorsal vertebrae of titanosaurian sauropod Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014, from Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian (A–E, shown in non-standard views for best visualization) and “disconnection” (sensu Gallina 2011) in presacral vertebrae of Bonitasaura salgadoi Apesteguía, 2004 from Río Negro Province, Argentina; Santonian (F). A. Left side of MPM-PV 1156?-4, estimated as the ~4th dorsal vertebra. B. Left side of MPM-PV 1156?-5, ~5th. C. Right side of MPM-PV 1156-6 (mirrored), ~6th. D. Right side of MPM-PV 1156?-8 (mirrored), ~7th. E. Right side of MPM-PV 1156?-9 (mirrored), ~8th. Changes in the PODLs through the sequence are denoted by a dotted line. F. Presacral vertebrae reproduced from Gallina (2011) under a Creative Commons Attribution License (CC BY 4.0). F1, F7, estimated13th? cervical vertebra; F2, F8, ~1st dorsal vertebra; F3, F9, ~2nd dorsal vertebra; F4, F10, ~3rd dorsal vertebra; F5, F11, 6th? dorsal vertebra; F6, F12, ~10th? dorsal vertebra. Photographs (F1–F6) and explanatory drawings (F7–F12). Abbreviations: DI, diapophysis; PODL, postzygodiapophyseal lamina; “PODL”, “new incipient horizontal lamina arises from the postzygapophysis pointing towards the diapophysis”, as per Gallina (2011: fig. 6A); POZ, postzygapophysis.
Dorsal vertebra MPM-PV 1156?-9: This vertebra is nearly complete (Figs. 1E, 2E, 3E, 5E, 6A). A few fragments are missing from the anterior and posterior margins of the centrum, as well as from the ventral portion of the PRSL and the middle of the right PODL. As seen in right anterolateral view, the ventral margins of the right CPRL and ACPL are also missing; furthermore, the summit of the neural spine, the posterolateral end of the right diapophysis, and the medial side of the left postzygapophysis are not preserved. MPM-PV 1156?-9 has been sheared in a right lateral direction, contrasting the direction of shear in the preceding MPM-PV 1156?-8 (Figs. 2E, 3E). This distortion results in many of the fossae of MPM-PV 1156?-9 being deeper on the left side. The neural spine has been compressed anteriorly, as evidenced by the offset of the right SPDL (Fig. 5E) and the nonlinear nature of the left SPDL (Figs. 1E, 5E).
The centrum is wider than tall, possibly as a result of taphonomic deformation (Figs. 2E, 3E). Even though it is compressed and offset, the majority of the neural spine is fairly vertical, but with a minor posterior inclination. The parapophysis is at almost the same height as the diapophysis (Fig. 1E). A semicircular area of parallel grooves is present on the dorsal surface of the diapophysis, as in MPM-PV 1156-6 and MPM-PV 1156?-8 (Fig. 6B). This area can be identified on both sides of the vertebra but is better preserved on the right.
The undivided CPRLs and ACPLs are contiguous on both sides, yet they appear divergent on the left side as a result of deformation. MPM-PV 1156?-9 is the anterior-most dorsal vertebra of Dreadnoughtus known to have had PCPLs, CPAFs, and PADFs (Fig. 1E). The PCPL and aPCDL intersect the PCDL at its approximate midpoint, roughly halfway between the centrum and diapophysis. These laminae are present on both sides but better defined on the left. The PCDL-Fs are shallow, especially on the right side due to shear. In contrast, the POCDFs are deep and posteriorly facing; that on the right side is more expansive.
The PRSL is faint near the ventral end of the neural spine, more prominent along the middle of its extent, and narrow toward the incompletely preserved tip of the spine (Fig. 2E). The original length of the neural spine is unknown, thus the PRSL might or might not have reached the tip. The PRSL intersects the faint TPRL, which is more distinct on the right side. The PPDLs curve medially; the right PPDL is more distinct (Fig. 1E). Unlike the condition in the preceding three vertebrae, in MPM-PV 1156?-9 the PODLs are essentially straight (Fig. 5E). The CPOLs are broad in posterior view. Deformation of the neural spine has resulted in shallow PRSDFs. The SPOFs, CPOFs, and CPRFs are also shallow.
The SPDLs are clearly split into a- and p-SPDLs, with SPDL-fs dorsal to the POSDFs (Figs. 1E, 5E, 6A). Curvature and breakage of the SPDLs are a result of taphonomic distortion. The POSDFs have the shape of dorsally pointing right triangles that are bound anteriorly by the p-SPDLs. On the left side, this lamina has been compressed into the POSDF (Figs. 1E, 6A). There is a distinct soft tissue attachment scar on the left side where the p-SPDL and the undivided SPOL meet (Fig. 6A), as in MPM-PV 1156?-8 and MPM-PV 1156-6. On the right side, the neural arch is fractured at this location, possibly obstructing the identification of this feature. The right SPOL is broader and better preserved than the left. Taphonomic alteration of the posterior side of the neural arch has obscured identification of the TPOL and rendered the POSL poorly defined. The latter is best developed on the dorsal portion of the neural spine (Fig. 3E).
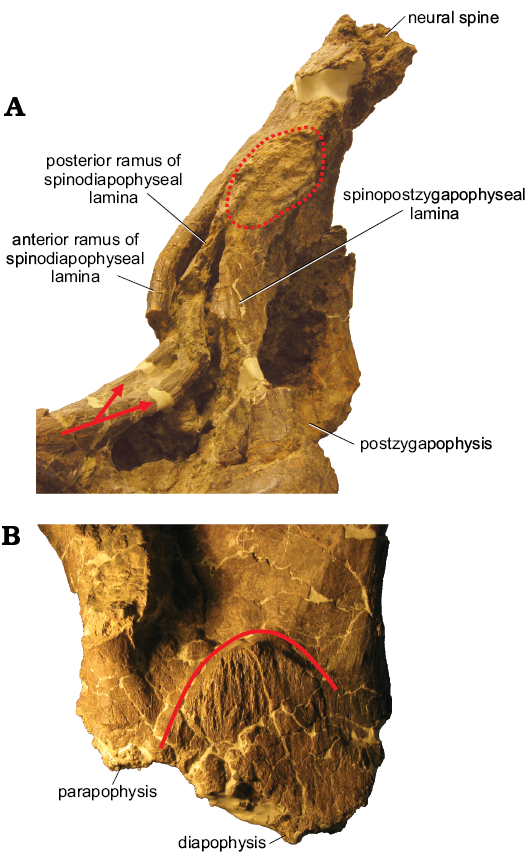
Fig. 6. Examples of osteological correlates of soft tissue attachment on dorsal vertebrae of titanosaurian sauropod Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014, from Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian. A. Neural spine of MPM-PV 1156?-9 in left lateral view. Arrows mark the initial split of the a- and p-SPDL. Dotted line encircles the osteological correlate on the neural spine. B. Left transverse process of MPM-PV 1156?-8 in dorsal view. Solid line marks the semicircular margin of the striations on the transverse process. Not to scale.
Dorsal vertebra MPM-PV 1156?-10: Much of the dorsal part of the neural arch of MPM-PV 1156?-10 is missing, and the whole vertebra has been severely dorsoventrally compressed (Figs. 1F, 2F, 3F). The prezygapophyses, transverse processes, and neural spine are not preserved, nor are portions of the left ventral, anterior, and posterior margins of the centrum. The boundaries between the lateral pneumatic fossae and the posterior face of the centrum are incomplete on both sides. The right PCDL is not present, and only the most ventral portion of the left is preserved, though it is missing its lateral margin. Only the most ventral segments of the aPCDLs are present, and the left is also missing its lateral margin. Moreover, as preserved, the CPOLs are severely dorsoventrally crushed.
Dorsoventral pressure has compressed all of the fossae spanning the neural arch and centrum. Some of the laminae that form the boundaries of these fossae are fractured (Fig. 1F). The preserved portion of the TPRL is compressed. The CPRLs appear single and broad, although their widths have been accentuated by dorsoventral compaction. As best observed in lateral view, branched or parallel laminae at both anterodorsal corners of the centrum suggest the presence of a distinct ACPL that is separate from the CPRL, but taphonomic distortion precludes confirmation of this. On the left side, the PCDL would likely have been the most robust lamina. Only the left PCPL is preserved (Fig. 1F), and it is better developed than in the preceding MPM-PV 1156?-9. It is offset due to dorsoventral compression but would likely have been connected to the aPCDL as in MPM-PV 1156?-9. The right PCPL might be present, but poor preservation of the appropriate region of the vertebra renders this difficult to determine. Deformation and lack of preservation prevent assessment of the original condition of any fossae.
Dorsal vertebra MPM-PV 1156-11: Although this vertebra exhibits little distortion, it is comprised of only the centrum and the anterior, left, and posterior bases of the neural arch (Figs. 1G, 2G, 3G). The zygapophyses, transverse processes, and neural spine are not preserved. The posterior margin of the centrum is mostly damaged, except on the left side. The only evidence of distortion in this vertebra is a small offset on the posterior half of the left side of the centrum.
Though the parapophyses and diapophyses are not preserved, their locations can be predicted based on the geometry of the preserved laminae, which suggests that the parapophyses and diapophyses were at about the same dorsoventral level. Despite the lack of preservation of a horizontal TPOL, there is a thin vertical lamina in the center of the ventral portion of the posterior face of the neural arch, dorsal to the neural canal; this is likely a ventral projection of the TPOL (Fig. 3G). Three distinct laminae are preserved on the left side of the neural arch (Fig. 1G). The PCDL is the most robust, although the CPOL is also thickened. The thin PCPL arises from the PCDL instead of from the aPCDL (as seen in the preceding two vertebrae), as there is no indication of the aPCDL in this vertebra. The PCPL of MPM-PV 1156-11, although thin, is more strongly developed than in any other dorsal vertebra of Dreadnoughtus. The ACPL is missing its exterior surface. Portions of the left PADF, CPAF, and POCDF are preserved, but are too incomplete to allow comment on their shape or depth. The CPRF and CPOF are shallow, and the former is broad whereas the latter is taller than wide.
Stratigraphic and geographic range.—Type locality and horizon only.
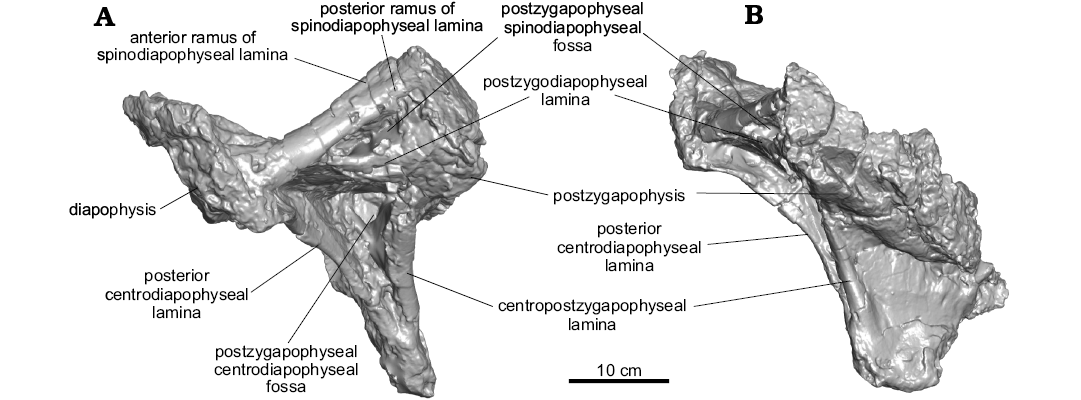
Fig. 7. Partial neural arch of titanosaurian sauropod Dreadnoughtus schrani Lacovara, Lamanna, Ibiricu, Poole, Schroeter, Ullmann, Voegele, Boles, Carter, Fowler, Egerton, Moyer, Coughenour, Schein, Harris, Martínez, and Novas, 2014 (MPM-PV 1156?-7), from Santa Cruz Province, Argentina; middle–late Campanian to early Maastrichtian; in left lateral (A) and posterior (B) views.
Discussion
Comparisons.—Along with caudal vertebrae, the only skeletal elements that overlap between almost all of the largest known titanosaurians (listed in the Introduction) are dorsal vertebrae. Comparisons between the dorsal vertebrae of giant titanosaurians are limited because the serial positions of the preserved vertebrae vary considerably (e.g., the only dorsal vertebrae preserved in Notocolossus, González Riga et al. 2016 and Puertasaurus, Novas et al. 2005 are more anterior than any known for Dreadnoughtus). Among these giant titanosaurians, Calvo et al. (2007a: 519) described Futalognkosaurus as having dorsoventrally narrow transverse processes throughout the dorsal series, unlike the “deep” condition in Puertasaurus. The transverse processes in the preserved dorsal vertebral series of Dreadnoughtus are also dorsoventrally narrow. However, Dreadnoughtus and Argentinosaurus have ventromedially inclined prezygapophyses, whereas Futalognkosaurus has almost horizontal prezygapophyses (Calvo et al. 2007a). The dorsal vertebrae of Argentinosaurus and Puertasaurus are reported as having broader and less-defined laminae compared to those of Patagotitan, in which the dorsal vertebrae possess comparatively thin, well-defined laminae (Carballido et al. 2017); Dreadnoughtus also exhibits thin, well-defined laminae.
Additionally, there are notable similarities and differences among the dorsal vertebrae of Dreadnoughtus and other, smaller-bodied macronarians. For example, the presence of the PCPLs within dorsal vertebral regions (anterior, middle, and posterior) varies among titanosauriforms. In Dreadnoughtus, Malawisaurus (Gomani 2005), and Phuwiangosaurus (Suteethorn et al. 2009), the PCPL is present only in posterior dorsal vertebrae, as opposed to Epachthosaurus in which this lamina (which is one of the laminae reported with uncertain homology by Martínez et al. 2004) is present only in middle dorsal vertebrae (no anterior dorsal vertebrae are known for this genus; Powell 1990; Martínez et al. 2004). In Euhelopus, Opisthocoelicaudia, Overosaurus, and Trigonosaurus, by contrast, the PCPL is present in middle and posterior dorsal vertebrae (Borsuk-Białynicka 1977: text-figs. 2, 3, pl. 3; Wilson and Upchurch 2009; Campos et al. 2005; Coria et al. 2013). The PCPL is also present throughout the dorsal vertebral series of Rapetosaurus (Curry Rogers 2009). As in Rapetosaurus and Saltasaurus, the PCPL of Dreadnoughtus is single and undivided (Mannion et al. 2013). This contrasts the divided condition in Andesaurus, Malawisaurus, and Opisthocoelicaudia (D’Emic 2012; Mannion et al. 2013).
Unlike the condition in some macronarians (e.g., Camarasaurus, Malawisaurus, Opisthocoelicaudia, Tastavinsaurus; Carballido and Sander 2014), the lateral end of the dorsal vertebral transverse process of Dreadnoughtus does not curve smoothly into the dorsal surface of the process. The morphology in Dreadnoughtus is instead similar to that in many somphospondylans, including Euhelopus and the titanosaurians Epachthosaurus, Lirainosaurus, Neuquensaurus, Saltasaurus, and Trigonosaurus (as coded in the phylogenetic data matrices of Sanz et al. 1999; D’Emic 2012; Mannion et al. 2013; and Carballido and Sander 2014). The neural spines of Dreadnoughtus possess subparallel lateral margins (Figs. 2, 3), as in a minority of titanosauriforms (e.g., Mendozasaurus, Rapetosaurus, Sauroposeidon; Carballido and Sander 2014). In Dreadnoughtus, the dorsoventral height of the posterior dorsal neural spines is more than 20% the total height of their respective vertebrae (Fig. 1). This is also seen in, for example, the macronarians Andesaurus, Brachiosaurus, Camarasaurus, Ligabuesaurus, and Malawisaurus (Salgado et al. 2015).
Division of the SPDLs in middle and posterior dorsal vertebrae is also seen in the middle and Late Cretaceous titanosaurians Epachthosaurus, Muyelensaurus, Overosaurus, Patagotitan, Rapetosaurus, Saltasaurus, and Trigonosaurus (Powell 1992; Martínez et al. 2004; Campos et al. 2005; Calvo et al. 2007c; Curry Rogers 2009; Coria et al. 2013; Poropat et al. 2016; Carballido et al. 2017), and possibly Opisthocoelicaudia (see Salgado et al. 1997: 22–23). Of these taxa, only Saltasaurus and Rapetosaurus also preserve this condition in the anterior dorsal vertebrae (Powell 1992). It is possible that the so-called “diapophyseal lamina” in the middle dorsal vertebrae of Tapuiasaurus (Zaher et al. 2011: fig. 4a, b) might also represent a divided SPDL. Because it appears to connect the diapophysis to the neural spine, it is likely part of the SPDL according to the terminology of Wilson (1999, 2012).
The dorsal vertebrae of Dreadnoughtus preserve two different potential osteological correlates for soft tissue attachment, and both of these occur on multiple vertebrae (e.g., Fig. 6). As in Epachthosaurus, Lirainosaurus, Opisthocoelicaudia, Saltasaurus, Trigonosaurus, and some other somphospondylans (Borsuk-Białynicka 1977; Sanz et al. 1999; Campos et al. 2005), the transverse processes of several vertebrae of Dreadnoughtus preserve a well-defined, semicircular area with mediolaterally-oriented striations located dorsomedial to the diapophyseal facet. This area has previously been suggested to indicate a soft tissue attachment site (Sanz et al. 1999; D’Emic 2012). A second attachment scar occurs where the p-SPDL and SPOL intersect in the posterior-most middle and posterior dorsal vertebrae of Dreadnoughtus. To our knowledge, the only similar soft tissue correlate reported in the dorsal series of a titanosauriform is a “roughened, flattened region” on the dorsal surface of the neural spine in the anterior dorsal vertebrae of Euhelopus (see Wilson and Upchurch 2009: 220). Although, in Dreadnoughtus, the possible attachment area occurs in more posterior dorsal vertebrae and is located further laterally, these osteological correlates may correspond to the attachment of related soft tissues.
Serial variation.—Although not all of the preserved dorsal vertebrae of Dreadnoughtus can be definitively differentiated between the holotype and paratype, there is minimal, if any, positional overlap of these elements between the two individuals. Variation is clearly identifiable within this mixed series. Taphonomic deformation has changed the shape of almost every preserved vertebra, resulting in altered forms of some features that are frequently used for the assignment of serial position (e.g., the position of the parapophyses relative to the diapophyses, the orientation of the neural spine). However, neither of these factors is significant enough to preclude the identification of the likely serial progression of these vertebrae. The presence and absence of specific laminae and fossae adds additional information that is useful for positional assignment.
Despite being the most anteriorly preserved vertebra in the series, MPM-PV 1156?-4 is unlikely to be any more anterior than the fourth dorsal vertebra (Lacovara et al. 2014). The parapophyses are situated on the neural arch, anterior to and slightly below the dorsoventral level of the diapophyses (although this condition is likely influenced by taphonomic deformation; see above). The position of the parapophyses on the neural arch demonstrates that MPM-PV 1156?-4 cannot be one of the first one or two vertebrae in the dorsal sequence. Nevertheless, the specimen exhibits CDFs (Fig. 1A), which, in titanosauriforms, are most often present in anterior dorsal vertebrae (e.g., Euhelopus, Wilson and Upchurch 2009, Huabeisaurus, D’Emic et al. 2013, Lirainosaurus, Díez Díaz et al. 2013, Malawisaurus, Gomani 2005, Rapetosaurus, Curry Rogers 2009). Consequently, we conclude that MPM-PV 1156?-4 is an anterior dorsal vertebra, but that it was not positioned at the very beginning of the series; hence, we tentatively identify the specimen as the fourth dorsal. Unlike the next vertebra in the progression, MPM-PV 1156?-5, MPM-PV 1156?-4 has linear PODLs that extend directly from the diapophyses to the postzygapophyses (Fig. 5A, B). This feature distinguishes MPM-PV 1156?-4 from the following series of middle dorsal vertebrae.
In MPM-PV 1156?-5, the preserved PODLs exhibit an arced or “hooked” shape. The degree of curvature of these laminae can be seen to increase and then decrease through the middle dorsal series of Dreadnoughtus, as MPM-PV 1156?-5 has moderately curved PODLs, MPM-PV 1156-6 and MPM-PV 1156?-8 have strongly curved PODLs, and MPM-PV 1156?-9 has PODLs that are straight along the majority of their length before turning to meet the postzygapophyses (Fig. 5A–E). This is similar in form to the “disconnection” of the PODLs seen in the northern Patagonian titanosaur Bonitasaura (Gallina 2011; Fig. 5F). As reported by Gallina (2011), as one moves from anterior to posterior through the dorsal sequence of Bonitasaura, the PODLs migrate to a position dorsal to the postzygapophyses along the neural spine (thereby becoming SPDLs according to the nomenclature of Wilson 2012); then, in the posterior dorsals, novel “PODLs” arise that connect the diapophysis and postzygapophysis along a short, straight path. In contrast to the morphology in Bonitasaura (Gallina 2011), in Dreadnoughtus, the SPDLs are not reduced during this transition of the PODLs. Wilson (2012) termed this type of transition of vertebral laminae “lamina capture”, and argued that topology should always dictate the names of laminae in sauropod vertebrae. Thus, in the present paper, laminae that extend from the diapophyses to the neural spine are referred to as SPDLs regardless of whether or not they reach those landmarks via lamina capture.
The structure of the p-SPDLs provides the best criterion to place the next four vertebrae that follow MPM-PV 1156?-4 in a specific anteroposterior progression (Figs. 1, 5A–E). In MPM-PV 1156?-5, the SPDLs are of average thickness for a Dreadnoughtus dorsal vertebra. However, in MPM-PV 1156-6, the SPDLs are among the thickest laminae on the vertebra. MPM-PV 1156?-8 and MPM-PV 1156?-9 have SPDLs that bifurcate into anterior and posterior branches (of average thickness) that enclose SPDL-Fs. The extent of bifurcation is greater in MPM-PV 1156?-9, resulting in larger SPDL-Fs. This pattern occurs on both the left and right sides of these vertebrae. Unfortunately, this region is not preserved in MPM-PV 1156?-10 and MPM-PV 1156-11, and it is therefore impossible to determine if bifid SPDLs were present in the most posterior dorsal vertebrae of Dreadnoughtus as well.
Because MPM-PV 1156?-5 has a hook-shaped PODL and a SPDL of average thickness, it is likely the 5th dorsal vertebra (Fig. 5B). MPM-PV 1156-6 would follow (therefore, it would be the 6th dorsal; Lacovara et al. 2014) based on the hook-shaped PODLs and hypertrophied SPDLs (Fig. 5C). This vertebra was also found near the scapula (MPM-PV 1156-48) in the torso region of the partially-articulated holotype skeleton, supporting the conclusion that it is unlikely to be a posterior dorsal vertebra. MPM-PV 1156?-8 is predicted to be the 7th dorsal vertebra (Lacovara et al. 2014) because it has bifurcated SPDLs with SPDL-F (Fig. 5D). MPM-PV 1156?-9 would follow (8th; Lacovara et al. 2014) as it has a wider and longer bifurcation of the SPDLs (Fig. 5E). The neural arch fragment MPM-PV 1156?-7 would belong to a vertebra located somewhere in this region of the sequence, as suggested by the bifurcation of the SPDL (Fig. 7A), but lack of preservation prevents a more precise interpretation of its serial position. If there is any sequential overlap preserved in the dorsal vertebrae of the holotype and paratype specimens of Dreadnoughtus, it would most likely be in this region of the sequence. However, considering the preservational state of the vertebrae and the taphonomic data collected (see Lacovara et al. 2014), this determination cannot be made with certainty. Since there are significant morphological differences between the vertebrae, it is reasonable to assume that there is little to no positional overlap between them, and that the sequence proposed herein is likely a serial progression, or nearly so (even though it is probably an amalgamation of two individuals).
MPM-PV 1156?-9 exhibits the most anterior occurrence of PCPLs, which are also seen in MPM-PV 1156?-10 and MPM-PV 1156-11 (Fig. 1E–G). In MPM-PV 1156?-9, the PCPLs are short, whereas in MPM-PV 1156?-10 and MPM-PV 1156-11 they increase in length and robustness. This feature aids significantly in the identification of the sequential positions of these vertebrae, and thus, their serial assignments. Based on the aforementioned aspects of the PCPLs, MPM-PV 1156?-10 is predicted to be the ~9th dorsal vertebra, and MPM-PV 1156-11 the ~10th. The identification of MPM-PV 1156-11 as the ~10th dorsal is supported by the presence of a ventral extension of the TPOL, as in Bonitasaura (Gallina 2011). Although this feature also occurs in the anterior dorsal vertebrae of Bonitasaura, Malawisaurus, Mendozasaurus, Muyelensaurus, and Rapetosaurus (González Riga 2003; Curry Rogers 2009; Gallina 2011; Poropat et al. 2016), MPM-PV 1156-11 cannot be an anterior dorsal vertebra of Dreadnoughtus because of the elevated location of the parapophyses. This assignment is additionally supported by the presence of a PCPL and the lack of an ACDL, which within the morphological variation of the other dorsal vertebrae of Dreadnoughtus is only seen in the posterior half of the sequence. Additionally, MPM-PV 1156-11 was discovered near the second caudal vertebra (MPM-PV 1156-14) in the pelvic region of the partially-articulated holotype skeleton, further supporting the identification of the former vertebra as a posterior dorsal.
Conclusions
Dreadnoughtus schrani is important in being the most completely preserved giant titanosaurian sauropod dinosaur yet described (Lacovara et al. 2014). Seven nearly complete dorsal vertebrae and an additional fragment of an eighth are preserved, making the dorsal series of Dreadnoughtus one of the most completely known among giant titanosaurians (the dorsal series of the comparably-sized Futalognkosaurus is complete, and Patagotitan preserves up to nine dorsal vertebral positions, but the dorsal vertebrae of these taxa are, respectively, incompletely prepared [Futalognkosaurus, Calvo et al. 2007a, b; MCL personal observation] or have only been briefly described [Patagotitan, Carballido et al. 2017]). Despite variation in the regions and positions of the dorsal vertebrae preserved in giant titanosaurian taxa, some noteworthy comparisons between them can be made. For example, the articular surfaces of the dorsal vertebral prezygapophyses are almost horizontal in Futalognkosaurus (Calvo et al. 2007a) and only slightly ventromedially inclined in Notocolossus and Puertasaurus (Novas et al. 2005; González Riga et al. 2016), whereas in Argentinosaurus (Bonaparte and Coria 1993) and Dreadnoughtus they are more strongly ventromedially inclined.
Most dorsal vertebrae of Dreadnoughtus exhibit significant taphonomic deformation, yet still preserve delicate laminae and rugose and striated areas that probably correspond to soft tissue attachment sites. This excellent preservation allows the sequential progression and approximate serial positions of these vertebrae to be confidently determined. Although the preserved dorsal vertebrae of Dreadnoughtus probably pertain to both the holotype and paratype specimens, only two of them can be definitively assigned to the holotype (MPM-PV 1156). Even so, the morphologies of these vertebrae indicate that there is minimal, if any, positional overlap between them. The criteria used to determine serial progression in Dreadnoughtus might be useful in estimating the positions of dorsal vertebrae of less completely represented titanosaurians.
Although no dorsal vertebral autapomorphies of Dreadnoughtus have yet been identified, either by Lacovara et al. (2014) or in the present study, these vertebrae exhibit distinctive morphologies of the neural arch laminae within specific regions of the axial column. Bifurcation of the SPDLs is useful in identifying middle to posterior dorsal vertebrae, whereas the presence of PCPLs aids in the identification of posterior dorsal vertebrae. The completeness of the postcranial skeleton of Dreadnoughtus renders it an important taxon for understanding the anatomy of titanosaurians, especially very large-bodied members of the clade. Future investigations into the paleobiology and phylogenetic relationships of these remarkable creatures will benefit from continued discoveries and detailed descriptions of similarly complete skeletons.
Acknowledgements
Portions of the research occurred at Rowan University (Glassboro, USA), Drexel University (Philadelphia, USA), The Academy of Natural Sciences of Drexel University (Philadelphia, USA), and Carnegie Museum of Natural History (Pittsburgh, USA). We thank Rubén Martínez (Universidad Nacional de la Patagonia San Juan Bosco, Comodoro Rivadavia, Argentina), Fernando Novas (Museo Argentino de Ciencias Naturales, Buenos Aires, Argentina), the Museo Argentino de Ciencias Naturales, the Museo Padre Molina, and the Province of Santa Cruz for permitting, customs, and insurance. We are grateful to members of the paleontology laboratories of the above institutions for their assistance, especially Zachary Boles, Aja Carter, George Keighton, Emma Fowler, Elena Schroeter, and Paul Ullmann (all Drexel University), Russell Christoforetti and Daniel Pickering (both Carnegie Museum of Natural History), and Jason Poole (The Academy of Natural Sciences of Drexel University). This work was supported by the National Science Foundation (EAR Award 0603805 to KJL and DGE Award 1002809 to KKV), Drexel and Rowan university post-doctoral fellowships (KKV), and the generosity of Adam Schran (Philadelphia, USA). We thank Stephen Brusatte (Univeristy of Edinburgh, UK) for editorial assistance, and Phil Mannion (Imperial College London, UK) and Stephen Poropat (Swinburne University of Technology, Melbourne, Australia) for helpful reviews which improved the manuscript. This research formed part of the doctoral thesis of KKV at Drexel University, which was supervised by KJL, Aleister Saunders, Sorin Siegler, and James Spotila (all Drexel University), MCL (Carnegie Museum of Natural History), and Mary Schweitzer (North Carolina State University, Raleigh, USA).
References
Apesteguía, S. 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. Naturwissenschaften 91 (10): 493–497. Crossref
Benson, R.B.J., Campione, N.E., Carrano, M.T., Mannion, P.D., Sullivan, C., Upchurch, P., and Evans, D.C. 2014. Rates of dinosaur body mass evolution indicate 170 million years of sustained ecological innovation on the avian stem lineage. PLoS Biology 12: e1001853. Crossref
Bonaparte, J.F. and Coria, R.A. 1993. Un nuevo y gigantesco saurópodo titanosaurio de la Formación Río Limay (Albiano–Cenomaniano) de la provincia del Neuquén, Argentina. Ameghiniana 30: 271–282.
Borsuk-Białynicka, M. 1977. A new camarasaurid sauropod Opisthocoelicaudia skarzynskii gen. n., sp. n. from the Upper Cretaceous of Mongolia. Palaeontologia Polonica 37: 1–64.
Calvo, J.O. 2014. New fossil remains of Futalognkosaurus dukei (Sauropoda, Titanosauria) from the Late Cretaceous of Neuquén, Argentina. In: E. Cerdeño (ed.), 4th International Palaeontological Congress, “The History of Life: A View from the Southern Hemisphere”, Abstract Volume, 325. International Palaeontological Association, Mendoza.
Calvo, J.O., González-Riga, B.J., and Porfiri, J.D. 2007c. A new titanosaur sauropod from the Late Cretaceous of Neuquén, Patagonia, Argentina. Arquivos do Museo Nacional, Rio de Janeiro 65: 485–504.
Calvo, J.O., Porfiri, J.D., González Riga, B.J., and Kellner, A.W.A. 2007a. Anatomy of Futalognkosaurus dukei Calvo, Porfiri, González Riga & Kellner, 2007 (Dinosauria, Titanosauridae) from the Neuquén Group (Late Cretaceous), Patagonia, Argentina. Arquivos do Museo Nacional, Rio de Janeiro 65: 511–526.
Calvo, J.O., Porfiri, J.D., González-Riga, B.J., and Kellner, A.W.A. 2007b. A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur. Anais da Academia Brasileira de Ciências 79: 529–541. Crossref
Campos, D., Kellner, A.W.A., Bertini, R.J., and Santucci, R.M. 2005. On a titanosaurid (Dinosauria, Sauropoda) vertebral column from the Bauru Group, Late Cretaceous of Brazil. Arquivos do Museo Nacional, Rio de Janeiro 63: 565–593.
Carballido, J.L. and Sander, P.M. 2014. Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from the Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria. Journal of Systematic Palaeontology 12: 335–387. Crossref
Carballido, J.L., Pol, D., Otero, A., Cerda, I.A., Salgado, L., Garrido, A.C., Ramezani, J., Cúneo, N.R., and Krause, J.M. 2017. A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs. Proceedings of the Royal Society B 284: 20171219. Crossref
Casal, G.A., Ibiricu, L.M., Allard, J.O., Martínez, R.D., Luna, M., and González Riga, B.J. 2014. Tafonomía del titanosaurio Aeolosaurus colhuehuapensis, Cretácico Superior, Patagonia central, Argentina: un ejemplo de preservación en facies fluviales de desbordamiento. Revista Mexicana de Ciencias Geológicas 31: 163–173.
Coria, R.A., Filippi, L.S., Chiappe, L.M., García, R., and Arcucci, A.B. 2013. Overosaurus paradasorum gen. et sp. nov., a new sauropod dinosaur (Titanosauria: Lithostrotia) from the Late Cretaceous of Neuquén, Patagonia, Argentina. Zootaxa 3683: 357–375. Crossref
Curry Rogers, K. 2005. Titanosauria: a phylogenetic overview. In: K. Curry Rogers and J.A. Wilson (eds.), The Sauropods: Evolution and Paleobiology, 50–103. University of California Press, Berkeley.
Curry Rogers, K. 2009. The postcranial osteology of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 29: 1046–1086. Crossref
D’Emic, M.D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society 166: 624–671. Crossref
D’Emic, M.D., Mannion, P.D., Upchurch, P., Benson, R.B.J., Pang, Q., and Cheng, Z. 2013. Osteology of Huabeisaurus allocotus (Sauropoda: Titanosauriformes) from the Upper Cretaceous of China. PLoS ONE 8: e69375. Crossref
Díez Díaz, V., Pereda Suberbiola, X., and Sanz, J.L. 2013. The axial skeleton of the titanosaur Lirainosaurus astibiae (Dinosauria: Sauropoda) from the latest Cretaceous of Spain. Cretaceous Research 43: 145–160. Crossref
Egerton, V.M. 2011. The Geology, Paleontology and Paleoecology of the Cerro Fortaleza Formation, Patagonia (Argentina). 275 pp. Ph.D. dissertation, Drexel University, Philadelphia.
Gallina, P.A. 2011. Notes on the axial skeleton of the titanosaur Bonitasaura salgadoi (Dinosauria-Sauropoda). Anais da Academia Brasileira de Ciências 83: 235–245. Crossref
Gomani, E.M. 2005. Sauropod dinosaurs from the Early Cretaceous of Malawi, Africa. Palaeontologia Electronica 8: 27A.
González Riga, B.J. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina. Ameghiniana 40: 155–172.
González Riga, B.J. and Astini, R.A. 2007. Preservation of large titanosaur sauropods in overbank fluvial facies: a case study in the Cretaceous of Argentina. Journal of South American Earth Sciences 23: 290–303. Crossref
González Riga, B.J., Lamanna, M.C., Ortiz David, L.D., Calvo, J.O., and Coria, J.P. 2016. A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot. Scientific Reports 6: 19165. Crossref
Gorscak, E. and O’Connor, P.M. 2016. Time-calibrated models support congruency between Cretaceous continental rifting and titanosaurian evolutionary history. Biology Letters 12: 20151047. Crossref
Huene, F. von 1929. Los saurisquios y ornitisquios del Cretacéo Argentino. Anales del Museo de la Plata 3: 1–196.
Lacovara, K.J., Lamanna, M.C., Ibiricu, L.M., Poole, J.C., Schroeter, E.R., Ullmann, P.V., Voegele, K.K., Boles, Z.M., Carter, A.M., Fowler, E.K., Egerton, V.M., Moyer, A.E., Coughenour, C.L., Schein, J.P., Harris, J.D., Martínez, R.D., and Novas, F.E. 2014. A gigantic, exceptionally complete titanosaurian sauropod dinosaur from southern Patagonia, Argentina. Scientific Reports 4: 6196. Crossref
Mannion, P.D. and Otero, A. 2012. A reappraisal of the Late Cretaceous Argentinean sauropod dinosaur Argyrosaurus superbus, with a description of a new titanosaur genus. Journal of Vertebrate Paleontology 32: 614–638. Crossref
Mannion, P.D., Upchurch, P., Barnes R.N., and Mateus, O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoological Journal of the Linnean Society 168: 98–206. Crossref
Martínez, R.D., Giménez, O., Rodríguez, J., Luna, M., and Lamanna, M.C. 2004. An articulated specimen of the basal titanosaurian (Dinosauria: Sauropoda) Epachthosaurus sciuttoi from the early Late Cretaceous Bajo Barreal Formation of Chubut Province, Argentina. Journal of Vertebrate Paleontology 24: 107–120. Crossref
Novas, F.E., Salgado, L., Calvo, J., and Agnolin, F. 2005. Giant titanosaur (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia. Revista del Museo Argentino de Ciencias Naturales 7: 37–41. Crossref
Poropat, S.F., Mannion, P.D., Upchurch, P., Hocknull, S.A., Kear, B.P., Kundrát, M., Tischler, T.R., Sloan, T., Sinapius, G.H., Elliott, J.A., and Elliott, D.A. 2016. New Australian sauropods shed light on Cretaceous dinosaur palaeobiogeography. Scientific Reports 6: 34467. Crossref
Poropat, S.F., Upchurch, P., Mannion, P.D., Hocknull, S.A., Kear, B.P., Sloan, T., Sinapius, G.H.K., and Elliott, D.A. 2015. Revision of the sauropod dinosaur Diamantinasaurus matildae Hocknull et al. 2009 from the middle Cretaceous of Australia: implications for Gondwanan titanosauriform dispersal. Gondwana Research 27: 995–1033. Crossref
Powell, J.E. 1987. Morfología del esqueleto axial de los dinosaurios titanosauridos (Saurischia, Sauropoda) del Estado de Minas Gerais, Brasil. Anais do X Congresso Brasileiro de Paleontología, Río de Janeiro 1: 155–171.
Powell, J.E. 1990. Epachthosaurus sciuttoi (gen. et sp. nov.) un dinosaurio saurópodo del Cretácico de Patagonia (Provincia de Chubut, Argentina). Actas V Congreso Argentino de Paleontología y Bioestratigrafía, Tucumán 1: 123–128.
Powell, J.E. 1992. Osteología de Saltasaurus loricatus (Sauropoda, Titanosauridae) del Cretácico Superior del noroeste Argentino. In: J.L. Sanz and A.D. Buscalioni (eds.), Los Dinosaurios y su Entorno Biotico, Actas del Segundo Curso de Paleontología en Cuenca, 165–230. Instituto “Juan de Valdes,” Cuenca.
Powell, J.E. 2003. Revision of South American titanosaurid dinosaurs: palaeobiological, palaeobiogeographical and phylogenetic aspects. Records of the Queen Victoria Museum 111: 1–173.
Salgado, L. and Powell, J.E. 2010. Reassessment of the vertebral laminae in some South American titanosaurian sauropods. Journal of Vertebrate Paleontology 30: 1760–1772. Crossref
Salgado, L., Coria R.A., and Calvo, J.O. 1997. Evolution of titanosaurid sauropods. I: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 34: 3–32.
Salgado, L., Gallina, P.A., and Paulina Carabajal, A. 2015. Redescription of Bonatitan reigi (Sauropoda: Titanosauria), from the Campanian–Maastrichtian of the Río Negro Province (Argentina). Historical Biology 27: 525–548. Crossref
Sanz, J.L., Powell, J.E., Le Loeuff, J., Martínez, R., and Pereda Suberbiola, X. 1999. Sauropod remains from the Upper Cretaceous of Laño (northcentral Spain). Titanosaur phylogenetic relationships. Estudios del Museo de Ciencias Naturales de Álava 14: 235–255.
Smith, J.B., Lamanna, M.C., Lacovara, K.J., Dodson, P., Smith, J.R., Poole, J.C., Giegengack, R., and Attia, Y. 2001. A giant sauropod dinosaur from an Upper Cretaceous mangrove deposit in Egypt. Science 292: 1704–1706. Crossref
Suteethorn, S., Le Loeuff, J., Buffetaut, E., Suteethorn, V., Talubmook, C., and Chonglakmani, C. 2009. A new skeleton of Phuwiangosaurus sirindhornae (Dinosauria, Sauropoda) from NE Thailand. Geological Society, London, Special Publications 315: 189–215. Crossef
Ullmann, P.V. and Lacovara, K.J. 2016. Appendicular osteology of Dreadnoughtus schrani, a giant titanosaurian (Sauropoda, Titanosauria) from the Late Cretaceous of Patagonia, Argentina. Journal of Vertebrate Paleontology 36: e1225303. Crossref
Upchurch, P., Barrett, P.M., and Dodson, P. 2004. Sauropoda. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria. Second Edition, 259–322. University of California Press, Berkeley. Crossref
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639–653. Crossref
Wilson, J.A. 2006. Overview of titanosaur evolution and phylogeny. In: Colectivo Arqueologico-Paleontológico de Salas de los Infantes (eds.), Actas de las III Jornadas sobre Dinosaurios y su Entorno, 169–190. Salas de los Infantes, Burgos.
Wilson, J.A. 2012. New vertebral laminae and patterns of serial variation in vertebral laminae of sauropod dinosaurs. Contributions from the Museum of Paleontology, University of Michigan 32: 91–110.
Wilson, J.A. and Upchurch, P. 2009. Redescription and reassessment of the phylogenetic affinities of Euhelopus zdanskyi (Dinosauria: Sauropoda) from the Early Cretaceous of China. Journal of Systematic Palaeontology 7: 199–239. Crossref
Wilson, J.A., D’Emic, M.D., Ikejiri, T., Moacdieh, E.M., and Whitlock, J.A. 2011. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. PLoS ONE 6: e17114. Crossref
Zaher, H., Pol, D., Carvalho, A.B., Nascimento, P.M., Riccomini, C., Larson, P., Juarez-Valieri, R., Pires-Domingues, R., da Silva, N.J. Jr., and Campos, D. 2011. A complete skull of an Early Cretaceous sauropod and the evolution of advanced titanosaurians. PLoS ONE 6: e16663. Crossref
Acta Palaeontol. Pol. 62 (4): 667–681, 2017
https://doi.org/10.4202/app.00391.2017