

Diversity of diapsid fifth metatarsals from the Lower Triassic karst deposits of Czatkowice, southern Poland—functional and phylogenetic implications
MAGDALENA BORSUK-BIAŁYNICKA
Borsuk-Białynicka, M. 2018. Diversity of diapsid fifth metatarsals from the Lower Triassic karst deposits of Czatkowice, southern Poland—functional and phylogenetic implications. Acta Palaeontologica Polonica 63 (3): 417–434.
Three morphotypes of the fifth metatarsal (MttV), one of the most informative bones of the postcranium, have been described herein from the Early Triassic karst deposits of the Czatkowice locality (Southern Poland). Two of them have been assigned to a basal archosauriform Osmolskina czatkowicensis and a basal lepidosauromorph, Sophineta cracoviensis, respectively, while one is undetermined saurian. Two morphological categories of the hooked fifth metatarsals recognized from this assemblage account for two different solutions to the problem of improvement of locomotion. A strongly inflected (sensu Robinson 1975) MttV shaft consists of two parts, a distal one lying on the ground in a plantigrade manner and a proximal one bent at an angle to get align with the ventral surface of the crus and proximal tarsus. In contrast, a straight shaft of the hooked MttV, suggests its subvertical life position and thus a digitigrade foot stance. The hooking of the fifth metatarsal, that is currently accepted saurian synapomorphy, appeared in phylogeny in a primitive state referred to herein as a neckless type: with neither a neck-shaped articular protrusion for the fourth distal tarsal nor a directly medial orientation of the articular facet. A derived long-necked type with protruding arricular part and more directly medial orientation of the articular facet appeared, at various stages of further phylogeny. A strong plantar–dorsal inflexion of the fifth metatarsal associated with a protrusion of lateral plantar tubercle, dates from a directly pre-lepidosaurian stage of evolution.
Key words: Reptilia, Diapsida, fifth metatarsal, functional morphology, Triassic, Poland.
Magdalena Borsuk-Białynicka [borsuk.b@twarda.pan.pl], Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818 Warszawa, Poland.
Received 16 November 2017, accepted 16 May 2018, available online 16 June 2018.
Copyright © 2018 M. Borsuk-Białynicka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The fifth metatarsal is one of the most informative elements, among the postcranial bones in reptiles. For a long time, it evolved the interest of researchers, because of its unusual morphology characteristic of such groups of reptiles as modern diapsids (Sauria sensu Gauthier 1984) and turtles (possibly convergently evolved, but see Rieppel 1995 for contradictory opinion), different from the remaining metatarsals. The consensus now exists that the uniform rod-like metatarsals, with the fifth one articulated terminally with the fifth distal tarsal is plesiomorphic in reptiles, whereas the medially bend fifth metatarsal, that articulates with the fourth, instead of the fifth distal tarsal, which disappears, is derived (Schaeffer 1941; Lee 1997). Goodrich (1916: 264) was the first to discuss the taxonomic significance of the medially bent fifth metatarsal, for which he introduced the term “hooked fifth metatarsal”. Detailed studies from the last century (Romer 1922, 1956; Schaeffer 1941; Robinson 1975) on the structure and function of the pes in tetrapods demonstrated the role of the fifth metatarsal in amniote locomotion. Along with more recent studies on myology and arthrology of the pes in extant lizards (Rewcastle 1980; Brinkman 1980a, b; Russell and Bauer 2008; Sullivan 2010), they allow a more detailed functional approach to this bone.
The collection of about 30 complete or fragmentary hooked fifth metartarsals, on which this study has been based comes from the Lower Triassic karst deposits from the locality Czatkowice (southern Poland). The Czatkowice material, composed of the disarticulated skeletons of about ten small vertebrate taxa, has been attributed to particular taxa on the basis of reconstructed skull (Borsuk-Białynicka and Evans 2009a, b; Evans 2009; Evans and Borsuk-Białynicka 2009a, b) or tooth (Borsuk-Białynicka and Lubka 2009) structure. Isolated postcranial bones are not always readily assignable, but metatarsal V, elaborated for functional purposes, has more potential in this respect. It also provides much information on the function of the foot as a whole, and is thus the main concern of this paper. Unfortunately, relatively few Permo-Triassic reptile specimens preserve articulated feet with adequately described fifth metatarsals, so as to afford a basis for comparison with the isolated bones. These include the articulated pes of Saurosternon (Carroll 1975), considered a non-saurian basal diapsid by Ezcurra et al. (2014) and that of Protorosaurus, currently considered the earliest unambigous archosauromorph (Ezcurra et al. 2014) both from the late Permian.
The present paper refers to the early stages of phylogeny of Sauria (sensu Gauthier et al. 1988b), including the dichotomy between archosauromorphs and lepidosauromorphs.
The goal of the present paper is to present the observed morphological variation of the fifth metatarsal in both functional and phylogenetic terms.
Institutional abbreviations.—PIMZ, Paläontologisches Institut und Museum der Universität, Zurich, Switzerland, ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland; ZPE, Department of Paleobiology and Evolution, Warsaw University, Poland.
Other abbreviations.—dTIV, distal tarsal IV; MttIV, MttV, the fourth and fifth metatarsal, respectively.
Material and methods
This analysis is based on a collection of 10 complete and almost 20 fragmentary fifth metatarsals, identified as saurian bones on the basis of hooking. All of them come from the Early Triassic vertebrate assemblage from Czatkowice (southern Poland). Czatkowice locality, situated in Southern Poland near Kraków, is a Lower Triassic karst formation that has yielded a rich small vertebrate assemblage of reptiles (Borsuk-Białynicka and Evans 1998, 2003, 2009a, b; Borsuk-Białynicka and Lubka 2009; Borsuk-Białynicka and Sennikov 2009; Evans 2009; Evans and Borsuk-Białynicka 2009a), and amphibians (Evans and Borsuk-Białynicka 2009b; Shishkin and Sulej 2009). An account of the geology and taphonomy of the locality has been presented by Paszkowski (2009) and Cook and Trueman (2009), respectively. The age of the assemblage has been discussed by Borsuk-Białynicka et al. (1999, 2003), and subsequently defined as early late Olenekian by Shishkin and Sulej (2009).
For the sake of brevity the fifth metatarsal has been referred to as MttV, the fourth one as MttIV and distal tarsal IV as dTIV, where necessary. The term “outer process” has been used for “the proximal lateral extension” of the fifth metatarsal.
The problem of the proper orientation of MttV was faced by Robinson (1975). She recommended three terms to refer to their 3D structure (Figs. 1, 2). Hooking has been currently understood as a medial protrusion of the proximal head of the MttV. I suggest that the medial slope of the articular facet for the contact with the dTIV is more essential for defining the hooked MttV than the medial protrusion of the proximal head. Some MttV morphotypes considered hooked, because of the proximo-medial orientation of the articular facet for the dTIV, have the proximal end evenly expanded in medio-lateral axis. This is here referred to as a neckless structure (Fig. 1D, F1). The medial protrusion of articular part of MttV associated with more directly medial orientation of the facet for dTIV, is here referred to as a long-necked type (Fig. 1A1–C1, F2). The hooking angle (Robinson 1975) may be determined as an angle between the long axis of the MttV (more or less parallel to the lateral border of the shaft) and a line perpendicular to the articular facet for the dTIV (Fig. 1), measured in the dorsal plane. However, this measure is far from precision, particularly so if the distal part of the bone is out of the straight (Fig. 1C1) or the articular facet is reoriented dorsally (Fig. 1A1). The hooking angle is thus only a rough measure of the amount of bending of the MttV.
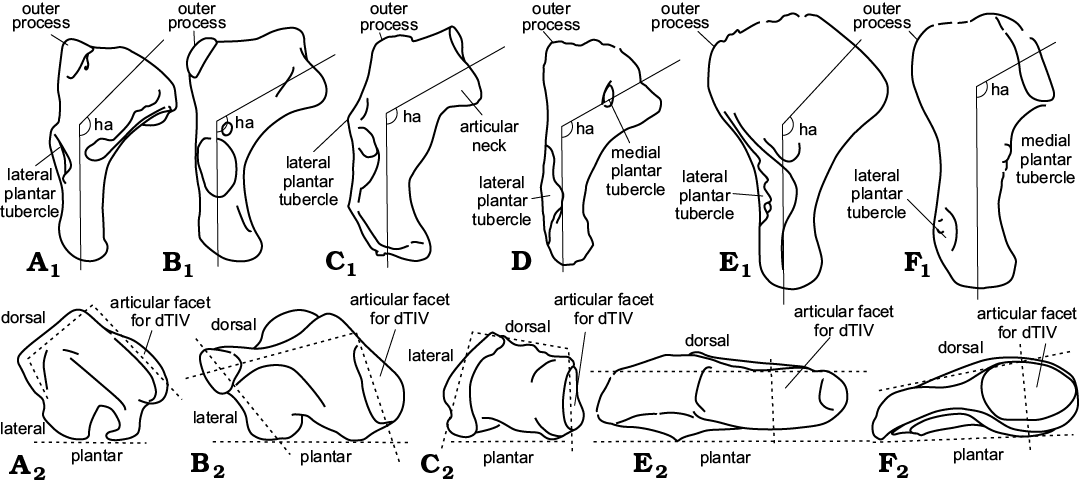
Fig. 1. Comparison of the fifth metatarsal architecture. Left bones in plantar view demonstrate hooking and articular facet orientation (A1–C1, D, E1, F1) and in proximal view (plantar surface down) (A2–C2, F2). A. Iguana iguana (Linnaeus, 1758), Iguanidae, Recent. B. Sphenodon punctatus (Gray, 1842), Rhynchocephalia, Recent. C. Sophineta cracoviensis Evans and Borsuk-Białynicka, 2009, Lepidosauromorpha, Early Triassic, Poland. D. Gephyrosaurus bridensis Evans, 1980, Rhynchocephalia, Early Jurassic, South Wales, UK (right MttV reversed). E. Osmolskina czatkowicensis Borsuk-Białynicka and Evans, 2003, Archosauriformes, Early Triassic, Poland. F. Morphotype X, Early Triassic, Poland. The dashed lines approximate the outlines of the fifth metatarsals as seen in proximal view (for additional explanation see Material and methods). ha, hooking angle. Not to scale. A, B1, C, after Robinson (1975); D, after Evans (1981).
Inflection is a bending of the long axis in plantar–dorsal plane (Fig. 2); and angulation is “the right-angle bending of the proximal end of the hooked fifth metatarsal, as seen in proximal view” (Robinson 1975). The angulation is here understood as a projection on a horizontal plane of the articular facet for the fourth tarsal, dorsal face of the bone, and plantar and lateral limitations of the bone as seen in proximal view (Fig. 1A2–F2). Angulation is not any precise term, but the proximal aspect of the bone is useful for demonstration of differences between morphotypes.
It should be kept in mind that the orientation of the joint facet in an isolated bone may not always be the same as the orientation of the joint, as that depends on both articulated facets. A description should be clear as to what is meant. The functional interpretation of disarticulated bones has important limitations with regard to the angulation of the limb as a whole, and thus the angle of the articular facets and their detailed morphology and subdivision should be considered.
The location of muscle insertions relative to the joints, i.e., estimation of the moment arms of the muscles, is important for a functional interpretation of morphological features (Robinson 1975; Brinkman 1980a; Rewcastle 1980; Sullivan 2010). Again, this interpretation should be in reference to a possible life position (Brinkman 1980a, b), but the designation of the surfaces of particular bones must be anatomical. Dorsal and plantar will be used instead of anterior and posterior to avoid misunderstanding.
Missing data made a phylogenetic analysis impractical in the present case. A phylogenetic scheme derived from currently accepted cladograms (mostly Evans 1988; Dilkes 1998; Ezcurra et al. 2014) was used to plot the discussed morphological features.
A Nikon SMZ 800 microscope with drawing apparatus was used for anatomical studies and a SEM Philips XL 20 was used for observation and scanning. Stereo-pairs have been used to demonstrate 3D morphology.
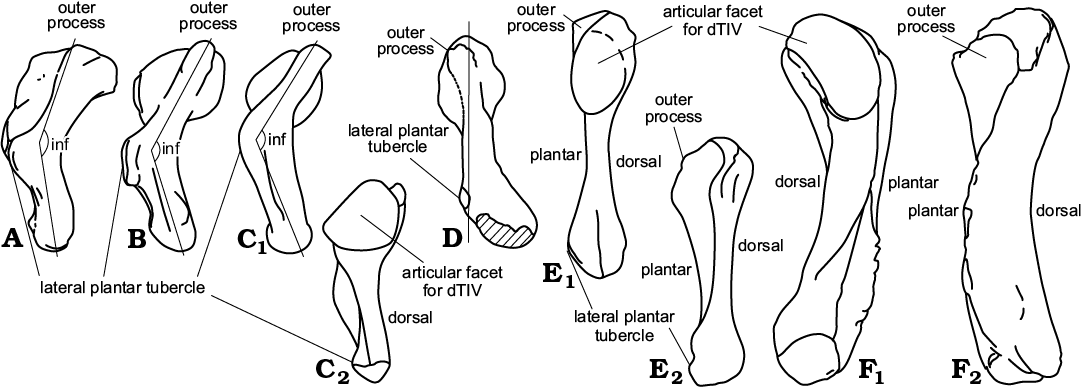
Fig. 2. Comparison of fifth metatarsal architecture. Right bones in lateral view (A, B , C1, D, E2, F2) and in medial view (C2, E1, F1). A. Iguana iguana (Linnaeus, 1758), Iguanidae, Recent. B. Sphenodon punctatus (Gray, 1842), Rhynchocephalia, Recent. C. Sophineta cracoviensis Evans and Borsuk-Białynicka, 2009, Lepidosauromorpha, Early Triassic, Poland. D. Gephyrosaurus bridensis Evans, 1980, Rhynchocephalia, Early Jurassic, South Wales, UK; straight line approximates not-inflacted shape of the MttV. E. Morphotype X, Early Triassic, Poland (left MttV reversed). F. Osmolskina czatkowicensis Borsuk-Białynicka and Evans, 2003, Archosauriformes, Early Triassic, Poland. inf, inflection angle. Not to scale. A, B after Robinson (1975); D, after Evans (1981).
The number of taxa considered herein on the basis of literature data, has been limited to those with a relevant fossil record. They are as follows:
Boreopricea funerea Tatarinov, 1978—Archosauromorpha, Early Triassic, Northern Russia, almost complete, articulated skeleton with both feet, but one MttV only (after Tatarinov 1978; Benton and Allen 1997).
Captorhinus aguti Case, 1911—basal Amniota, numerous specimens; Early Permian, North America (after Holmes 2003).
Gephyrosaurus bridensis Evans, 1980—Rhynchocephalia, disarticulated material, MttV preserved; Early Jurassic, South Wales fissure fillings, UK (after Evans 1981)
Macrocnemus bassani (Nopcsa, 1930)—Archosauromorpha, six articulated fragmentary pedes; Anisian–Ladinian boundary (Middle Triassic), Monte San Gorgio, Italy (after Rieppel 1989).
Mesosuchus browni Watson, 1912—Rhynchosauridae, some fragmentary, partly articulated feet; late Early Triassic to early Middle Triassic, South Africa (after Dilkes 1998).
Pamelaria dolichotrachela Sen, 2003—Archosauromorpha, disarticulated material belonging to several individuals and a complete skeleton including a complete hind limb; Middle Triassic, India (after Sen 2003).
Petrolacosaurus kansensis Lane, 1945—basal Diapsida, abundant articulated material including several pedes, some of them complete; late Carboniferous, North America (after Reisz 1981).
Prolacerta broomi Parrington, 1935—Archosauromorpha, articulated skeleton and hind leg, including foot; Early Triassic, South Africa (after Gow 1975).
Protorosaurus speneri Meyer, 1832—Archosauromorpha, four almost complete pes specimens; late Permian, Germany and England (after Gottmann-Quesada and Sander 2009).
Saurosternon baini Huxley, 1868—basal Diapsida, articulated skull-less skeleton; late Permian, South Africa (after Carroll 1975).
Archosauriformes:
Erythrosuchus africanus Broom, 1905—Erythrosuchidae, two foot specimens; Early Triassic, South Africa (after Gower 1996).
Euparkeria capensis, Broom, 1913—Euparkeriidae, two foot specimens; Middle Triassic, South Africa (after Ewer 1965).
Marasuchus illoensis (Romer, 1972)—Dinosauriformes; Middle Triassic, South America (after Sereno 1991).
Riojasuchus tenuiceps Bonaparte, 1969—Crurotarsi; Late Triassic, South America (after Sereno 1991).
Rhamphorhynchus sp.—Pterosauria; Late Jurassic, Europe, Africa (pes figured by Wellnhofer 1991: 57).
Pteranodon longipes Marsh, 1876—Pterosauria; Late Cretaceous, North America (after Bennett 2001).
Scleromochlus taylori Woodward, 1907—Ornithodira, disarticulated bones with incomplete articulated right foot; Late Triassic, England (after Benton 1999).
Articulated pes specimens of extant lizards:
Ctenosaura sp. ZPAL z.p. RI/8 and Varanus niloticus (Linnaeus, 1766), ZPAL z.p. RI/31 and an articulated skeleton of Sphenodon punctatus Gray, 1831 ZPE z.m. 483, were also examined.
Systematic palaeontology
Clade Sauria Gauthier, 1984
Clade Archosauromorpha Huene, 1946
Clade Archosauriformes Gauthier, 1986
Family Euparkeriidae Huene, 1920
Genus Osmolskina Borsuk-Białynicka and Evans, 2003
Type species: Osmolskina czatkowicensis Borsuk-Białynicka and Evans, 2003; early late Olenekian, Czatkowice I, southern Poland.
Osmolskina czatkowicensis Borsuk-Białynicka and Evans, 2003
Figs. 1E, 2F, 3.
Material.—MttV: two more or less complete specimens, ZPAL RV/1346, left; ZPAL RV/1347, right; (Fig. 3A, B), and 12 fragmentary specimens, ZPAL RV/1976–1988, with roughly estimated lengths (approximately 11 mm).
Description.—The fifth metatarsal attributed to Osmolskina (Figs. 1F1, 2F, 3) ranges in roughly estimated length from 10–12 mm, but most are 11 mm, there being no definitely juvenile bones. The Osmolskina MttV morphotype is distinguished from other Czatkowice MttV morphotypes by a conspicuous medio-lateral broadening of the proximal half, symmetrical with respect to the long axis of the shaft with no individualization of the articular part (a neckless type) and a moderate inflection (sensu Robinson’s 1975). The hooking angle is roughly 135°. The articular facet faces proximo-medially, and is slightly inclined onto the dorsal side. The facet is oval, much shorter in its plantar–dorsal axis than in the medio-lateral one (the ratio ~1:2). Proximally the bone ends in a point, where the articular facet sloping away medio-distally meets its proximo-lateral face sloping outwords to the outer process (Fig. 1E1). The lateral margin displays a crest extending from the outer process down to the distal end (Figs. 1E1, 3), and wrapping onto the plantar surface, thus twisting the distal end of the bone with respect to the proximal one. About one third of the length from the distal end, the crest turns into a tuberosity corresponding to the lateral plantar tubercle. Further distally, the shaft of the bone narrows substantially. A barely developed rugosity on the medial side corresponds to the medial plantar tubercle (Fig. 3A), placed proximal to the lateral one. The distal end bears an articular facet for the first phalanx of digit V on its medial corner. The facet is damaged in all specimens studied.
Remarks.—An additional right MttV, ZPAL RV/1993, from the same age and locality, exceeding the size range of O. czatkowicensis, considered an undetermined archosauriform by Borsuk-Białynicka and Sennikov (2009), has been illustrated (Fig. 3C) for comparison.
Stratigraphic and geographic range.—Early late Olenekian, Poland.
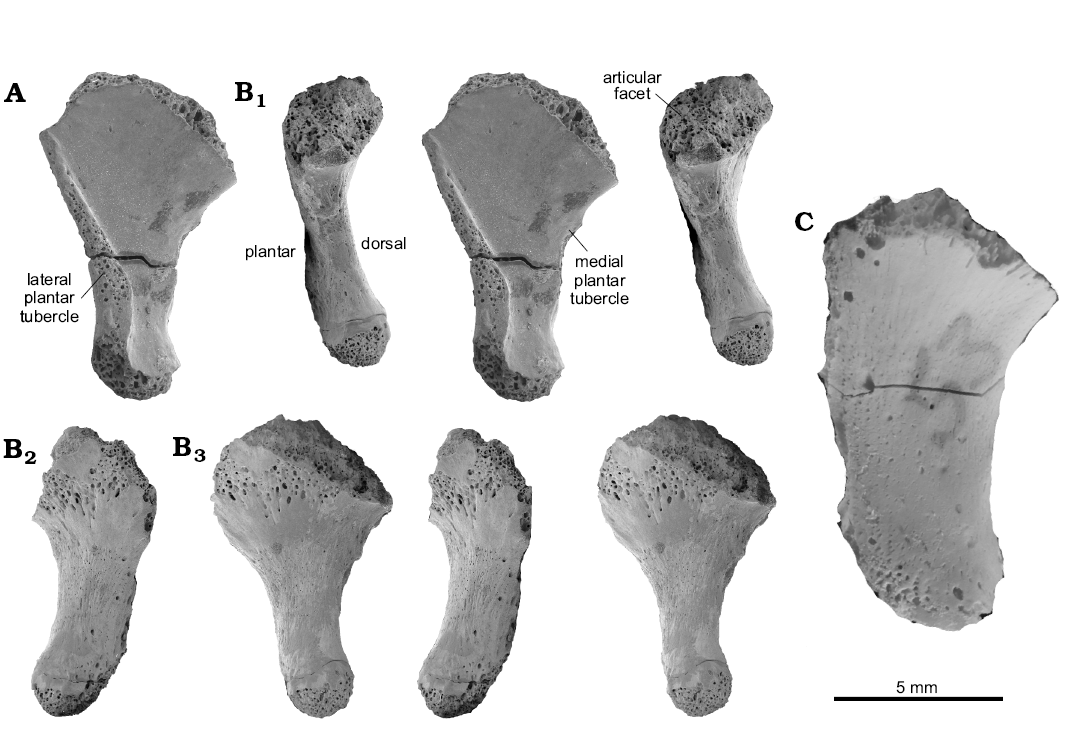
Fig. 3. Archosauriform Osmolskina czatkowicensis Borsuk-Białynicka and Evans, 2003 (A, B) and Archosauriformes gen. et sp. indet (C) all from the Lower Triassic Czatkowice locality, Poland. A. ZPAL RV/1347, adult, left MttV in plantar view. B. PAL RV/1346, young adult, right MttV in medial (B1), lateral (B2), and dorsal (B3) views. B1, B2, reversed. C. ZPAL RV/1993, right MttV in dorsal view. SEM photographs; A, B, stereo-pairs.
Clade Sauria Gauthier, 1984
Clade Lepidosauromorpha Gauthier, 1984
Family uncertain
Genus Sophineta Evans and Borsuk-Białynicka, 2009
Type species: Sophineta cracoviensis Evans and Borsuk-Białynicka, 2009; early late Olenekian, Czatkowice I, southern Poland.
Sophineta cracoviensis Evans and Borsuk-Białynicka, 2009
Figs. 1C, 2C, 4A, B.
Material.—MttV: four complete specimens, ZPAL RV/1353, right (Fig. 4B); ZPAL RV/1990, left (Fig. 4A); ZPAL RV/1418, right; ZPAL RV/1999, right. Seven less complete unnumbered specimens. All from early late Olenekian, Czatkowice I, southern Poland.
Description.—The Sophineta cracoviensis MttV morphotype is the smallest of all, ranging from 3.5 to 5.0 mm (7 specimens), mostly about 4 mm in length, and very lepidosaurian in its strong inflection and hooking. The hooking angle is about 110°, but the orientation of the long axis of the shaft is difficult to determine in view of irregular shape of the latter (Figs. 1C1, 4). The proximal articular facet is situated at the top of a well developed medial process, referred to as “articular neck” (Fig. 1C1) that protrudes mediad from the stem. The protrusion results in a strongly concave medial border and substantial asymmetry of the bone in dorsal aspect. The plantar surface is distinctly folded in transverse axis, A plantar convexity of the articular neck, that extends disto-laterally to about the midlength of the bone (a sort of roll-like eminence) and a proximo-distal furrow (Fig. 4B1, C) that borders it laterally, both contribute to this folding. The facet for dTIV is oval, but closer to circular (the ratio ~1:1.4) than that in Osmolskina or morphotype X. The facet faces directly mediad and probably only contacted dTIV, allowing some flexion-extension movement. The inflection of the bone is strong (Fig. 4B2), which makes the dorsal face concave. This concavity is enhanced by a dorsal inclination of the outer process. Among the Czatkowice taxa, Sophineta is distinguished by a laterally convex lateral border. The lateral plantar tubercle covers the entire middle one third of the length of this border. It lies within a crest that invades the plantar surface and contributes to the medial twist of the bone shaft. The medial plantar tubercle is poorly developed and situated above the mid-length of the medial border. The distal end of the bone is twisted medio-plantar with respect to the proximal part and bears an articular facet for the first phalanx of the digit at its medial corner. The facet forms a short oval, its long axis oriented medio-lateral.
Stratigraphic and geographic range.—Early late Olenekian, Poland.
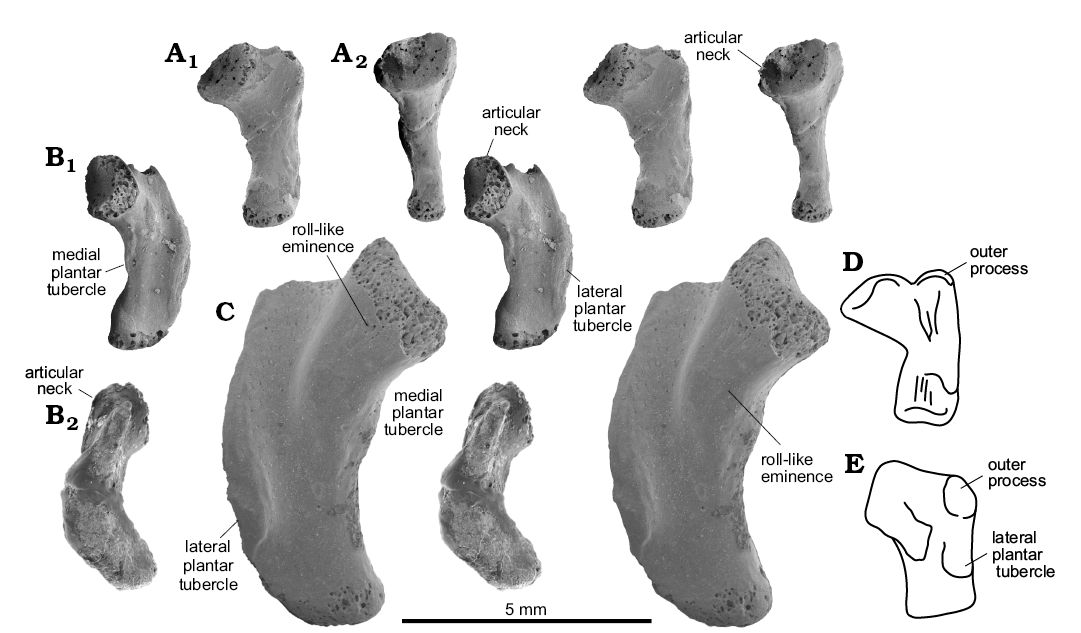
Fig. 4. Lepidosauromorph saurian Sophineta cracoviensis Evans and Borsuk-Białynicka, 2009 (A, B) and morphotype Y (C) from the Lower Triassic of Czatkowice, southern Poland and Macrocnemus bassani (Nopcsa, 1930) (D, E) from the Middle Triassic of Switzerland. A. ZPAL RV/1990, left MttV in dorsal (A1) and medial (A2) views. B. ZPAL RV/1353, right MttV in plantar (B1) and lateral (B2) views. C. ZPAL RV/1989, left MttV in plantar view. D. PIMZ T 2816, left MttV (reversed) in plantar view. E. PIMZ T 2472, right MttV in plantar view. A–C, SEM stereo-pairs. D, E not to scale, after Rieppel (1989: fig. 8B, E).
Sophineta cf. cracoviensis Evans and Borsuk-Białynicka, 2009
Morphotype Y
Fig. 4C.
Material.—MttV: ZPAL RV/1989, left bone (Fig. 4C); ZPAL RV/1343, proximal half of the left MttV. All from early late Olenekian, Czatkowice I, southern Poland.
Description.—Morphologically the morphotype Y of MttV falls within the variability range of Sophineta but is twice as large. Specimen ZPAL RV/1989 is 8.6 mm in length, the estimated length of ZPAL RV/1343 is about the same.
Clade Sauria Gauthier, 1984
Family indet.
Genus et species indet.
Morphotype X
Figs. 1I, K, 2E, 5.
Material.—MttV: three complete specimens, ZPAL RV/1991, adult; ZPAL RV/1992, adult (Fig. 5A, C); ZPAL RV/1354, juvenile (Fig. 5B), all from the left side. All from early late Olenekian, Czatkowice I, southern Poland.
Description.—MttV morphotype X elements roughly 5.5 and 6.0 in length plus one juvenile, 2 mm in length. This morphotype is distinguished by its straight, i.e., not inflected, shaft (Fig. 2E2). The hooking is roughly 120°. The articular part of the proximal end does not protrude as much as it does in Sophineta, and the asymmetry of the bone in dorsal view is lesser. The articular facet for dTIV is oval (extended medio-laterally, much shorter along the plantar–dorsal axis), more or less as in Osmolskina. The lateral plantar tubercle is situated near the distal end of the bone and is poorly developed. Situated on the medial border of the bone, at over half its length, there is a slight tuberosity that might correspond to the medial plantar tubercle. Alternatively it may be a mark of insertion of the flexor digitorum brevis. The distal end of the bone is twisted medio-plantar with respect to the proximal part and bears a damaged articular facet at its medial corner.
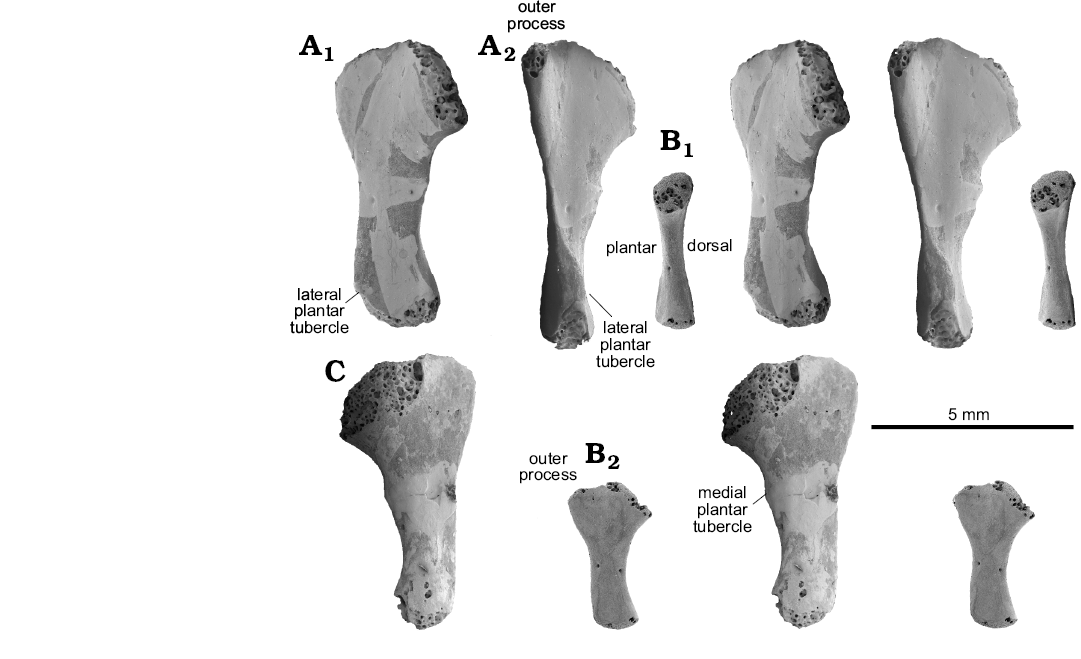
Fig. 5. Sauria indet. morphotype X from the Lower Triassic of Czatkowice, southern Poland. A. ZPAL RV/1991, adult left MttV in plantar (A1) and lateral-slightly plantar (A2) views. B. ZPAL RV/1354, juvenile left MttV in medial (B1) and plantar (B2) views. C. ZPAL RV/1992, adult left MttV in dorsal view. SEM stereo-pairs.
Discussion
Taxonomy
Four diapsid taxa Osmolskina, Czatkowiella, Pamelina, and Sophineta from the Lower Triassic of Czatkowice (southern Poland) assemblage have been recognized as an euparkeriid, a protorosaurid, a kuehneosaur, and a stem-lepidosaur, respectively, on the basis of skull characters (Borsuk-Białynicka and Evans 2009a, b; Evans 2009; Evans and Borsuk-Białynicka 2009a, b). Reconstructed skull lengths are 67, 40, 40, and 11 mm, respectively. The proportions of MttV length to skull length are not taxonomically conclusive, but may be helpful to discriminate between taxa within a certain assemblage. Two MttV morphotypes, the largest and the smallest, roughly correspond to the skull proportions of Osmolskina and Sophineta, respectively. The largest morphotype, ranging from 10 to 12 mm in length, is considered to be archosauriform and can be attributed to O. czatkowicensis. One larger specimen of the same morphology (ZPAL R/1993; 15 mm in length Fig. 3C) might belong to the Czatkowice Archosauriformes gen. indet. (Borsuk-Białynicka and Sennikov 2009). This MttV morphotype closely corresponds to that of Euparkeria (Ewer 1965) in the proximo-medial orientation of the articular facet for the dTIV, and the pointed shape of the proximal end (Fig. 1E1) that results from a relatively distal position of the outer process (about the level of the distal limit of the articular facet). The same overall shape of the MttV has been shown in Erythrosuchus (Gower 1996). A morphology of the plantar tubercle is unknown in both Euparkeria and Erythrosuchus. It is neither figured nor described (Ewer 1965: 414, pl. 34 and Gower 1996: figs. 5, 6). A small element evident in the uncatalogued specimen of Euparkeria figured by both Broom (1913: pl. 75) and Ewer (1965: pl. 34), proximal to the outer process) is here understood to be a piece of matrix filling the gap between the fifth metatarsal and calcaneum, and it has no bearing on the shape of the former.
The smallest MttV morphotype from Czatkowice, ranging from 3.5 to 5.0 mm, displays a lepidosaurian 3D structure, being both hooked (in terms of medial protrusion) and inflected. It is therefore attributed to Sophineta cracoviensis. The inflection mostly affects the lateral side (Fig. 2C). It results from a strong protrusion of the lateral plantar tubercle that changes the orientation of the proximal one third of the plantar surface exactly as in squamates. Therefore both size and morphology support the assignment of the smallest morphotype to Sophineta. Sophineta differs from lizards in the weak development of the medial plantar tubercle, and from Sphenodon in having a lateral instead of central position for the existing plantar tubercle. Sophineta has a circular shape and medial orientation of the facet for dTIV, in contrast to some extant lizards (e.g., Iguana, Agama, Xantusia) which have the facet oval and more dorsal in orientation (Brinkman 1980a).
In view of the attribution of a large number of bones from the Czatkowice assemblage to two other taxa: a protorosaurid, Czatkowiella harae, and a kuehneosaurid, Pamelina polonica, MttV morphotype X would be expected to belong to one of them. Size does not discriminate between these taxa (reconstructed skull length 40 mm in both). The relative frequency of these taxa in the Czatkowice assemblage suggests that it may belong to Czatkowiella, which is better represented than Pamelina, both in number of specimens and presence of different ontogenetic stages, exactly as morphotype X. However, this argument is not decisive. Among basal archosauromorphs, morphotype X resembles Protorosaurus in having an almost featureless straight shaft. However, its proximal articular part protrudes medially in a sort of neck, and the articular facet faces more directly medially than proximo-medially as it does in Protorosaurus. The outer process, level with the proximal limit of the articular facet, and is thus slightly more proximal than in Protorosaurus. The lateral plantar tubercle, very distal in MttV morphotype X, is absent in Protorosaurus, at least from the more proximal part of the shaft (Gottmann-Quesada and Sander 2009: fig. 25C). If morphotype X belongs to Czatkowiella, it would support the protorosaurid relationships of the latter, as suggested by Borsuk-Białynicka and Evans (2009a). Its attribution to the kuehneosaurid genus Pamelina would be more problematic. Kuehneosauridae, currently regarded as lepidosauromorphs (Evans 2009; see Müller 2004 for different opinion), were small, specialised Triassic reptilian gliders recorded from the Olenekian of Poland (Evans 2009) to the Carnian–Rhaetian of England (Robinson 1962; 1967) and North America (Colbert 1970). However, no information on the structure of the MttV in this group is currently available. Moreover, their gliding adaptations (elongated ribs) provide no implications concerning pes structure. A final taxonomic decision concerning the affiliation of MttV morphotype X is thus not possible.
Two specimens, designated as MttV morphotype Y, correspond in overall morphology to the MttV attributed to Sophineta but are more than twice its size. However, the inflexion is apparently slightly lesser and the outline of the articular facet for dTIV is slightly more ovoid. As Sophineta specimens do not show any juvenile features, ontogenetic variation seems improbable. MttV morphotype Y probably belongs to Sophineta, but may differ from S. cracoviensis at a specific level. However, the sample is too small to be conclusive.
Functional aspects.—As evidenced by numerous dissections (Perrin 1892; Schaeffer 1941; Brinkman 1980a; Sullivan 2010), the fifth metatarsal of lepidosaurs receives the insertions of important flexors of the pes, notably gastrocnemius and peroneus muscles. As demonstrated by Robinson (1975) and currently accepted, MttV exerts a lever action at the ankle joint, first to rotate the pes and then to lift the distal tarsus and metatarsus, so that the foot becomes digitigrade in gait and the body is lifted clear off the ground. The assumption that the relationships between the muscles and the MttV in the ancestral saurians was the same as it is in the modern lepidosaurs, and that their function was similar, seems reasonable.
In the Sauria (comprising Lepidosauromorpha, Archosauromorpha, their most recent common ancestor, and all their extinct descendants (Gauthier 1984; Gauthier et al 1988b) the proximal end of the fifth metatarsal is much larger in transverse axis than the distal end. Two factors, hooking and lateral expansion of the proximal end, contribute to this shape. Both features are associated with the action of the peronei muscle (Fig. 6C). The peroneus brevis attaches to the outer process (Robinson 1975) and rotates the pes prior to its plantarflexion (Brinkman 1980a). Peroneus longus wraps around the lateral border of MttV on its passage from its plantar to dorsal surface (Fig. 6C) and flexes the joint, but also contributes to transverse arching of the pes (plantar concave). The main pedal flexor, m. gastrocnemius, attaches onto the plantar tubercles. A full integration of the pes, distal to the mesotarsal joint, seems to be critical for the moment arm of the gastrocnemius with respect to the pes, as already stressed by Lee (1997). Still, the variability of the proximal articulation of the fifth metatarsals in the Czatkowice assemblage and elsewhere, suggests differences in the position and mobility of this bone, as discussed below.
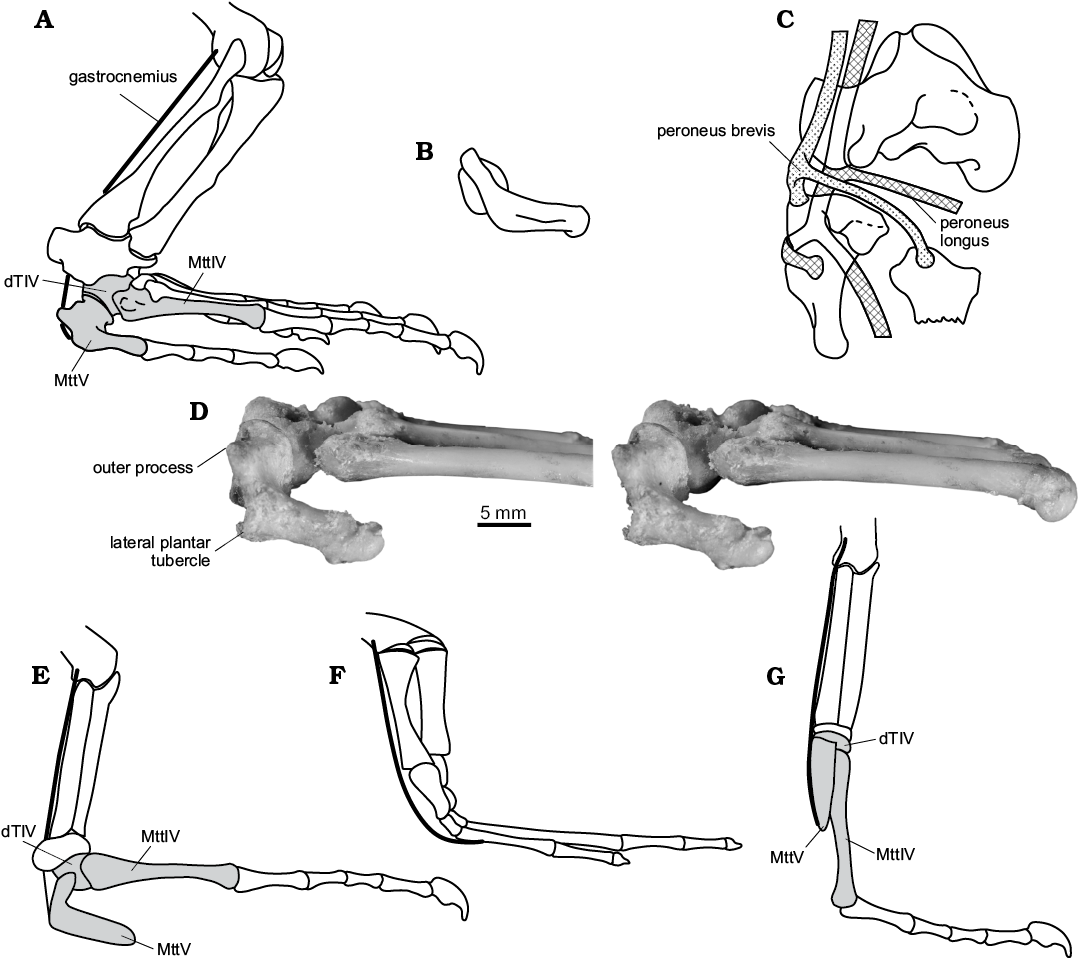
Fig. 6. General outline of muscle equipment of the lower leg and life position of the foot in Sauria. A. Varanus sp., left crus and pes in right lateral view (from Robinson 1975: fig. 8, reversed). B. Sophineta cracoviensis Evans and Borsuk-Białynicka, 2009, Lepidosauromorpha, Early Triassic, Poland, right MttV in lateral view; approximated life position. C. General courses and insertions of the peronei muscles in lizards, right pes in dorsal view (from Robinson 1975: fig. 11F, reversed). D. Varanus niloticus (Linnaeus, 1766), Recent, ZPAL z.p. RI/31, stereo-pairs of right foot: tarsus and proximal metatarsus in lateral view. E–G. Schematic drawings of foot showing proposed differences in levering conditions. E, G. Sauria. E. Plantigrade style with inflected MttV, as shown in A and D in squamates. F. Early tetrapods. G. Digitigrade style with straight MttV.
Position of MttV relative to the remaining metatarsals.—In Sophineta, a substantial protrusion of the articular part results in separation of the shaft of MttV from the remaining metatarsus. Nonetheless, the long axis of the MttV shaft turns medio-distally to become subparallel to the main axis of the remaining metatarsus. A possible contact with MttIV cannot be confirmed. Both, the long articular “neck” and the sub-circular proximal articular facet without any trace of the contact with MttIV show that MttV does not share the intimate association of metatarsals I–IV with each other. It suggests a possibility of some mobility within the joint between MttV and the remaining pes.
In Osmolskina, the neckless structure of the MttV, and the proximo-medial inclination of the articular facet, both suggest that the fifth metatarsal must have diverged laterad (assuming a more or less vertical joint facet on dTIV) from the remaining metatarsus rather stiffly (no mobility in medio-lateral axis). The condition is similar to that in Euparkeria, as figured by Ewer (1965: pl. 34), which suggests no more than limited mobility of MttV (except for some flexibility in the dorso-plantar plane, more dorsal than ventral; Fig. 3D). The ovoid, very short along the dorso-plantar axis, and rather flat facet for the dTIV seems consistent with this suggestion.
MttV morphotype X displays a short articular neck and the hooking angle (120°) intermediate between that of Sophineta (110°) and that of Osmolskina (135°) (Fig. 1F2, C1 and F1, respectively). This indicates that MttV morphotype X may have been positioned slightly more parallel to the remaining metatarsus in life than it was in Osmolskina. The facet for dTIV is similar in morphology to that in Osmolskina in that it is oval, short in dorso-plantar axis, more extensive in dorsal than in ventral view (Fig. 5C).
Important for the present discussion is a probable correlation between the plane of action of the pedal flexors and the angulation of the pes with respect to the parasagittal body plane at a plantarflexing phase of gait. In the fully erect extremities of mammals, where the action of pedal plantarflexion occurs in a parasagittal plane (Hildebrandt 1974), the calcaneal tuber extends posteriad. It is also posterior in extant crocodiles that use two different gaits, one of which is erect, and is characterized by a nearly parasagittal position of the femur (Brinkman 1980b). A lateral calcaneal flange of basal archosaurs (e.g., Sereno 1991; Gower 1996), a posterolateral tuber of extinct crurotarsans (Parish 1986; Sereno 1991), and a morphologically and functionally analogous lateral calcaneal flange of the extant Varanus (Sullivan 2010), all correspond to more abducted position of the femur and a sprawling pattern of gait. In lizards, the lateral orientation of the metatarsus at the time of pedal plantarflexion is a rule (Brinkman 1980a), though a degree of inclination, of the hind foot relative to parasagittal plane varies widely (50–90° of the angle between MttIV and parasagittal plane; Brinkman 1980a: 279, fig. 8). Consequently, the lateral margin of the MttV is more or less posterior at the initial phase of the propulsive action. Lateral means posterior is a common rule with respect to the hind extremity, from the beginning of tetrapod history (Romer 1956; Schaeffer 1941; Sukhanov 1968). Probably, the functional advantage of the hooking was not only to shift the insertions of the muscles further from the joint they operated, but also to shift them posteriad, to orient a propulsive stroke more directly anterior. The shift of the joint axis of the fifth metatarsal from transverse (MttV extending parallel to the more medial metatarsals) to vertical (the proximal end of MttV extending laterad) may have involved the peronei muscles in the first place.
Next example of the same functional association (lateral side of posterior extremity faces more or less posterior) is a distally divergent position of the fifth metatarsal in Osmolskina, which shifts the attachment of the flexor muscle, gastrocnemius this time, more lateral i.e more posteriad of the metatarsus axis and the fulcrum within the mesotarsal joint. Osmolskina shares this character not only with Euparkeria (Ewer 1965), but also with other immediate outgroups of Archosauria (Fig. 7A, B), such as Erythrosuchus (Gower 1996) and chanaresuchids (Sereno 1991). In the Dinosauromorpha, that achieved the erect gait, the MttV became subparallel to the metatarsus axis or deflected slightly behind Fig. 7F, G), if it was present at all (Serreno 1991). The lateral parts of the pes has got eventually reduced with stabilization of the erect position of the hind leg.
In this context, the asymmetry of the plantar tubercles in Czatkowice morphotypes may be interpreted in terms of limb orientation. Sophineta differs from all lizards in high asymmetry of the tubercles, the lateral one being decidedly stronger than the medial one. The lateral tubercle is well developed at about the middle one third of the lateral margin of the plantar surface of MttV, whereas the medial one is hardly visible distal of the articular neck of this bone (Fig. 4B1, C). Probably, the medial flexors, and notably the medial slip of the femoral gastrocnemius, were much less important than the lateral slip that occupied a posterior position in the abducted extremity. In the less abducted extremity of Iguana, MttIV axis to anteroposterior axis of the body angle just 50° versus 90° in Cnemidophorus (Brinkman 1980a), the plantar side, instead of lateral faces more or less posteriad and is more symmetric in terms of development of the plantar tubercles. In some lizards they join across the plantar surface into an annulus. Unique for Sphenodon is the only one plantar tubercle situated in the middle of the width of the MttV plantar surface.
Interestingly, the asymmetry in the development of plantar tubercles, described above in Sophineta, is displayed also in Osmolskina in spite of differences in overall morphology of the bone. Consequently, the position of the extremity is supposed to have been similar to that in basal lepidosauromorphs. This corresponds to the lateral orientation of the calcaneal tuber in Osmolskina (Borsuk-Białynicka and Sennikov 2009), less so with the supposed facultative bipedality of this animal suggested by Borsuk-Białynicka (2008) and supported by analysis of Euparkeria structure presented by Ewer (1965). However, all the lizards, both quadrupedal and facultatatively bipedal are basically sprawlers (Snyder 1954), and such must have been biomechanic constitution of euparkeriids.
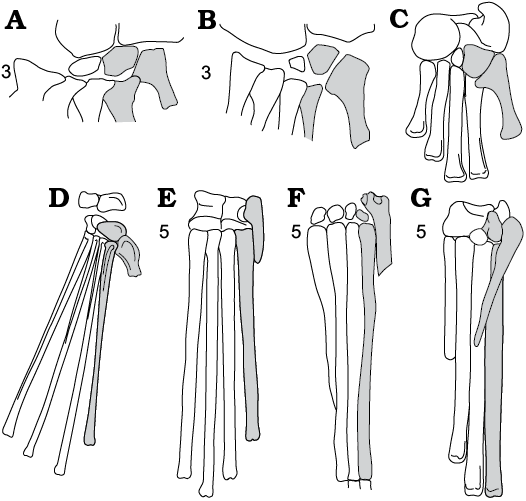
Fig. 7. Schematic relations between distal tarsals and metatarsals in Archosauriformes. A. Erythrosuchus, Erythrosuchidae, Early Triassic, Africa (after Gower 1996). B. Euparkeria, Euparkeriidae, Middle Triassic South Africa (after Ewer 1965). C. Riojasuchus, Crurotarsi, Late Triassic South America (after Sereno 1991). D. Rhamphorhynchus, Pterosauria, Late Jurassic, Europe, Africa (after Wellnhofer 1991). E. Pteranodon, Pterosauria, Late Cretaceous, North America (after Bennett 2001). F. Scleromochlus, Ornithodira, Late Triassic, England (after Benton 1999). G. Marasuchus, Dinosauriformes, Middle Triassic, South America (after Sereno and Arcucci 1994). Dorsal (A, B) and plantar (C, G) views. dTIV, MttIV and MttV shaded in grey. 3, saurian synapomorphy: neckless hooked MttV; 5, ornithodiran synapomorphy: MttV straight, reduced in size and importance. Not to scale.
Angulation sensu Robinson (1975). Plantigrady versus digitygrady.—The outline of fifth metatarsal in a proximal view, referred to as angulation by Robinson (1975; see Material and methods for details) is informative as concerns its position and possible mobility with respect to the remaining pes. The outline differs dramatically in the iguanids (exemplified by Iguana; Fig. 1A2) in being subpentagonal, and is intermediate in Sphenodon (Fig. 1B2). In Iguana, the facet is turned obliquely dorsal and occupies part of the dorsal surface of the shaft (Fig. 1A2), the position of the MttV being latero-plantar with respect to the rest of the metatarsus, which is thus transversally arched, ventrally concave. The position of the MttV is variable within the squamates. According to Brinkman (1980a: fig. 6) in Xantusia it is similar to that in the iguanids, but in Varanus it is more like in Sophineta, in Agama similar to Sphenodon, As shown by Robinson (1975), Sphenodon displays a medial protrusion of the articular part (Fig. 1B), the circular outline of the facet (Fig. 2B) and its generally medial orientation (the 3D structure of the facet is unknown. The plane of the articulation slopes slightly dorsally (personal observation on Sphenodon punctatus Gray, 1831 ZPE z.m. 483).
Some differences that exist between the proximal aspects of the fifth metatarsals in Osmolskina and morphotype X (Fig. 1E2, F2) on one side and in squamates, Sophineta (Fig. 1C2), and morphotype Y on the other, consist in a dorso-plantar depth being much shorter in Osmolskina and morphotype X. These differences should be understood in terms of the degree of inflection of the bone along with the development of the plantar tubercles. The inflection is minor in morphotype X (Fig. 1F2) and strongest in Sophineta, Sphenodon, and extant lizards, in which the proximal portion of the shaft produces a proximally turned “platform” that is extensive in its dorso-plantar diameter (Fig. 1C2). The proximal orientation of this “platform” is in reference to the isolated bone, but the orientation is posterior and sub-vertical (Fig. 6A, B, F) in the articulated plantigrade extremity, especially during plantarflexion of the ankle joint. In this position the plantar tubercles are situated on the back surface of the lower leg and at a certain distance below the axis of the mesotarsal joint (Fig. 6A), around which the flexion-extension movements occur. This distance is a lever arm of the gastrocnemius. Schaeffer (1941) and Robinson (1975), both discussed a so-called pulley problem of the foot architecture, that appears, when the line of action of the gastrocnemius changes from vertical (crus position) to horizontal (plantigrade foot position), over the arcuate back side of the of the foot (Fig. 6F). The problem appears when the foot skeleton is not consolidated as is the case in the lower tetrapods. Shifting the attachment site of gastrocnemius from the horizontal to the subvertical, posterior side of the leg (Fig. 6) as is the case in Sauria, in therapsids and mammals (Robinson 1975), is a method to avoid this problem. Bending the fifth metatarsal into two parts, the proximal part tending to align the crus and the distal one remaining horizontal, is one solution to the problem (Fig. 6E). Another solution may rely upon rotation (in an evolutionary sense) of the whole bone to a vertical position (Fig. 6G), so that it tends to align with the crus along its entire length, with the effect that the attachment point for the gastrocnemius becomes posterior to the plantar profile of the leg, and posterior to mesotarsal joint. All the above observations suggest that a hooked fifth metatarsal with a straight shaft was sub-vertical in life, and the whole foot was digitigrade (Fig. 6G).
Limited as they are, the arguments for digitigrady of some Early Triassic diapsids, based on the MttV structure, support the opinion that a departure from the sprawling gait associated with a facultataive bipedality appeared early in diapsid phylogeny. The digitigrade foot posture could logically have been a preadaptation to this type of gait. This has been hypothesized for a number of fossil taxa beginning with an early Permian bipedal reptile, Eodibamus cursorius (Berman et al. 2000; see also Santi 1993) on the basis of different postcranial characters (limb disparity, pelvis and sacral features, caudals). Snyder’s (1954, 1962) papers provided an inspiration for studies of bipedality. Snyder (1962) realized that skeletal adaptations required for facultative bipedality in lizards were not specific. Different quantitative features contribute to the ability to raise the anterior body in swift locomotion. Shortened forelimbs and a shortened body led Ewer (1965) to conclude that Euparkeria was capable of running bipedally. The same was hypothesized for Osmolskina (Borsuk-Białynicka 2008). The latter author based on co-positioning of the ilio-sacral joint (on the medial side of the ilium) and the acetabulum, as in iguanids capable of bipedal locomotion (Snyder 1954) but not lizards that lack this ability. Disparity between forlimb and hindlimb in lengths has been considered as indicative of facultative bipedality in Prolacerta (Gow 1975), whereas Rieppel (1989) used further arguments for this mode of locomotion in Macrocnemus, namely the important volume of the caudifemoralis (powerful femur retractor), and the sacrum (much expanded and distally overlapping pleurapophyses of the two sacral vertebrae) and basin structure related to trunk musculature that assists in raising the trunk (Rieppel 1989: 384). According to Sereno (1991), a semi-erect position was ancestral for archosaurs, whereas basal ornithodirans were fully erect, digitigrade and bipedal. It concerns, among others, Scleromochlus (Benton 1999) and pterosaurs (Padian 2008, 2017), although some authors (Wellnhofer 1991 and references therein) advocated a plantigrade stance and gait in pterosaurs. The basic structure of the pterosaurian foot (Fig. 7D–F) is consistent with a digitigrade posture, in having a mesotarsal joint, and elongate subparallel metatarsals I–IV, of which II–IV are subequal in length and contribute, along with the digits, to a symmetrical foot. A small degree of disparity between the digits indicates an orientation of the foot with the dorsal side anteriad and mobility in a parasagittal plane (Brinkmann 1980a). In Scleromochlus (Fig. 7F), the MttV is straight, reduced in length, obviously not very important for levering the foot. In contrast to Bennett (2001: 131), lifting a long plantigrade foot off the substrate without the levering function of the MttV seems hardly possible. In basal Dinosauromorpha, the MttV becomes subparallel to the MttIV, deflected behind the pes and tapering distally, with the articular facet for dTIV oriented parallel to the shaft axis, as demonstrated by Marasuchus (Fig. 7G) and Silesaurus (Dzik 2003). As widely known, the MttV disappeared in later archosaur phylogeny. In the erect, digitigrade foot of the ornithodirans, the attachment of the gastrocnemius must have moved down the metatarsus, but ornithodiran locomotion is beyond the scope of the present paper.
Fulcrum of the foot leverage system. Rotation and arching of the pes.—As already stressed by Lee (1997), the integration of the MttV, that bears the muscle insertions, with the foot, that is to be moved, is critical to the levering of the pes on the crus. However, the levering system did function in the early tetrapods in spite of their uniform metatarsals operated by flexor muscles extending to the plantar surface of the foot, and using the convex plantar surface of the pes as a pulley (Fig. 6F). Although quite ineffective, it was sufficient, because of the limited importance of the foot in propulsion (Schaeffer 1941). Some amount of mobility within the pes (between metatarsal I–IV) does occur even in extant crocodiles (Brinkman 1980b: fig. 10). This is reminiscent of the “internal deformation” and “differential dorsiflexion” of the foot in the early tetrapods (Holmes 2003).
In the extant lizards the mesotarsal joint, between the proximal tarsals and the fourth distal tarsal, is the main or the only fulcrum of the system (Rewcastle 1980; Brinkman 1980a). In extant lizards the rotation (sensu Brinkman 1980a: 279) of the pes, just prior to pedal plantarflexion, occurs around the long axis of the metatarsus in such a way that “its extensor surface eventually faces anterior” (Brinkman 1980a: 279). The action is activated by the peroneus brevis muscle (Fig. 6C) that first rotates and then plantarflexes the pes (Brinkman 1980a: 285). The metatarsus is involved as a unit. This is to say that, by this movement, the MttV is integrated with a part of the pes distal of the mesotarsal jont. This integration is also of importance for the activity of the main pedal flexor, the gastrocnemius.
The degree of integration of MttV with the remaining foot was the subject of contention between Robinson (1975) and Brinkman (1981a). Robinson (1975) assumed some mobility in the MttV–dTIV joint in lizards, consisting in the medio-plantar shift of the MttV with respect to the rest of the metatarsus. This movement would direct the plantar surface of the fifth metatarsal towards the plantar surface of the first metatarsal resulting in a transverse arching of the pes. A grasping ability was the assumed adaptive sense of these movements.
Brinkman (1980a) based on detailed studies (mainly cineradiography) of the movements of crus and pes during locomotion cycle in Iguana. According to this author, based on Iguana, the articular surfaces of the MttV–dTIV joint are closely approximated and are bound together by four ligaments. It should be added that in iguanids the plane of the joint is oblique instead of parallel to the flexion-extension plane (Fig. 1A). Addtionally, the distal ends of metatarsals IV and V are fastened together by the intermetatarsal tendon (see also the relevant description in Varanus bengalensis; Rewcastle 1980). Other arguments are equivocal. The characteristic overlap of the proximal ends of metatarsals I–IV (in dorsal aspect each metatarsal overlaps the one immediately lateral to it; see e.g., Robinson 1975: fig. 9) integrates the metatarsus, but it rarely, if ever, involves MttV. According to Brinkman (1980a), metatarsals I–III form a functional unit in extant lizards, whereas according to Robinson (1975: 478), the first four metatarsals “form a long and narrow bar confined in a common bag of skin and unable to diverge”. In any case the fifth metatarsal is not included.
Robinson’s (1975) studies concerned the 3D construction of the fifth metatarsal and of the pes as a whole, in Iguana and Varanus, along with observations on living animals. Her studies resulted in a model of pes function provided with a hypothesis on the adaptive sense behind transformations of the pes during phylogeny. Both these authors referred to extant lizards, but their results may shed some light on the function of the pes at the early stages of saurian phylogeny.
The differences between long-necked and neckless structure of MttV revealed by Czatkowice diapsids, suggest differences in the degree of possible movements within the MttV–dTIV joint. It should be stressed, that in those early saurians that had the fifth metatarsal of the long-necked type (Fig. 8F–L), the overlap between MttV and dTIV was probably feeble, apparently less than in the neckless types (Fig. 8D).
Assuming that at the early stage of saurian phylogeny the mesotarsal joint was not yet elaborated for foot rotation (as it is in extant lepidosaurs see Rewcastle 1980), some amount of mobility of the MttV may have been helpful for locomotion. Probably the MttV alone rotated around its long axis, by the action of the peroneus brevis muscle that pulled upward on the lateral, posterior edge of this bone, which resulted in its turning the extensor surface anterior. This movement could have forced the rotation of the remaining pes. I suppose that this was the initial movement at the early stage of evolution of propulsive ability of the foot, and probably not only in lepidosaurs (see below).
The directly medial orientation of the facet for the dTIV in Sophineta suggests the foot less arcuate in transverse section (assuming the lateral orientation of corresponding facet on the dTIV), Possibly, its arcuation resulted from actual medio-plantar movement of the fifth metatarsal. Most likely, it was accomplished by the activity of the peroneus longus that is assumed to pass from ventral to dorsal side by wrapping around the lateral side of the metatarsal as it does in extant lizards (Robinson 1975 and Fig. 6C herein). If this is correct, the arcuation of the pes was functional rather than structural (in contrast to extant lizards), but became structural later in squamate phylogeny, as evidenced by the extant iguanids (Brinkman 1980a). Moreover, a subsequent decrease in mobility of the MttV may have paralleled the elaboration of the mesotarsal joint (Rewcastle 1980) which eventually became the only fulcrum of the foot leverage system and allowed a rotation of the foot as a whole.
The rotation of the pes at the beginning of the propulsive phase of gait must have occurred not only in lizards, but by necessity, in all sprawlers in order to combine the more or less lateral position of the limb with the forward movement of the body. The differences between early lepidosaurian and archosauriform structure of the MttV–dTIV joint (long-necked versus neckless MttV morphology) suggest different solution to this problem. In lepidosaurians the locomotion probably based more on pes mobility, whereas in archosauriform the whole hind leg movements may have been more important.
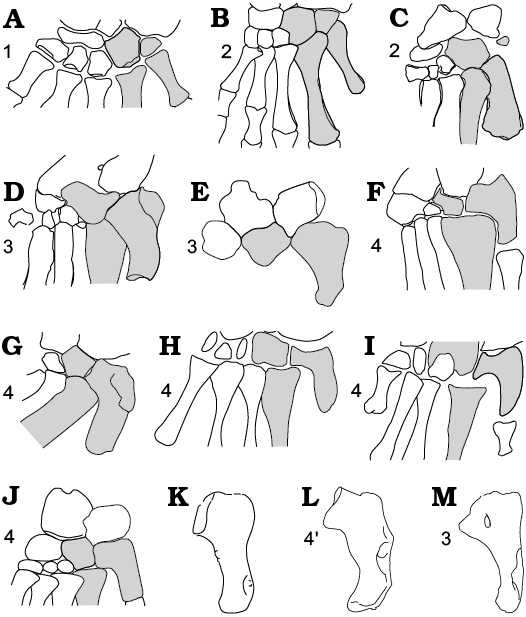
Fig. 8. Schematic relations between distal tarsals and metatarsals in diapsids and their out-group. A. Captorhinus, basal Amniota, Early Permian, North America (after Heaton and Reisz 1982). B. Petrolacosaurus, basal Diapsida, Late Carboniferous, North America (after Reisz (1981). C. Saurosternon, basal Diapsida, Late Permian South Africa (after Carroll 1975). D. Protorosaurus, Archosauromorpha, Late Permian, Germany and England (after Gottmann-Quesada and Sander 2009). E. Boreopricea, Early Triassic, Northern Russia (after Benton and Allen 1997). F, G. Macrocnemus, Middle Triassic, Italy (after Rieppel 1989: fig. 8D, F, respectively). H. Prolacerta, Archosauromorpha, Early Triassic, South Africa (after Gow 1975). I. Pamelaria, Archosauromorpha, Middle Triassic, India (after Sen 2003). J. Mesosuchus, Rhynchosauridae, Early–Middle Triassic, South Africa (after Dilkes 1998). K. MttV morphotype X, Early Triassic. Poland. L. MttV of Sophineta, Early Triassic. Poland. M. MttV of Gephyrosaurus, Lepidosauromorpha, Early Jurassic, UK (after Evans 1981). A–M all in plantar view. dTIV, MttIV and MttV. dTIV, MttIV, and MttV shaded in grey. 1, plesiomorphic state; 2, diapsid synapomorphy: foot integration; 3, saurian synapomorphy: neckless hooked MttV; 4, long-necked hooked MttV; 4’, lepidosaurian synapomorphy: dorso-ventral inflexion of the long-necked hooked MttV. Not to scale.
Phylogeny
Review of relevant taxa.—The following considerations on the transformations of the fifth metatarsal in diapsid evolution (Fig. 9) follow the recent phylogenetic schemes (Dilkes 1998; Sereno 1991; Ezcurra et al. 2014) developed from comprehensive phylogenetic analyses of earlier decades (including Benton 1985; Evans 1988; Gauthier 1984; Gauthier et al 1988a) and supplemented by new data. The number of discussed taxa has been reduced to those with a relevant fossil record (see Material and methods).
Two taxa, represented by more or less fully preserved pedes, Petrolacosaurus (late Carboniferous, North America; Reisz 1981) and Saurosternon (late Permian, South Africa; Carroll 1975), represent the early stage of diapsid phylogeny. Petrolacosaurus is widely accepted as a sister taxon of the Neodiapsida (sensu Benton 1985). The exact position of Saurosternon has been changing through time. Considered as a basal lizard by Carroll (1975), then a basal lepidosauromorph (Evans 1988), eventually as a non-saurian basal diapsid (Ezcurra et al. 2014).
Both Petrolacosaurus and Saurosternon present a basically plesiomorphic type of metatarsus as exemplified by Captorhinus (Figs. 8A, 9: 1). The unhooked fifth metatarsal contacts the tarsus proximally, by means of a facet displaying only an incipient medial inclination. Petrolacosaurus (Fig. 8B) is the more plesiomorphic than Saurosternon in having a complete set of distal tarsals and uniform metatarsalia, including the fifth one. In Saurosternon the fifth tarsal is rudimentary and the fifth metatarsal, probably autapomorphic for this taxon, is much thicker than the remaining four (Fig. 8C). However, both these genera display a dorsal overlap of the metatarsals (Lee 199), including the MttV (Fig. 9: 2, direction from I to V in dorsal view).

Fig. 9. A phylogeny of diapsids with the main transitions of the fifth metatarsal mapped onto it (based on Benton 1985; Evans 1988; Gauthier et al. 1988a; Sereno 1991; Dilkes 1998; Ezcurra et al. 2014). 1, plesiomorphic state; 2, diapsid synapomorphy: foot integration; 3, saurian synapomorphy: neckless hooked MttV; 4, long-necked hooked MttV; 4’, lepidosaurian synapomorphy: dorso-ventral inflexion of the long-necked hooked MttV; 5, ornithodiran synapomorphy: MttV straight, reduced in size and importance.
The morphology of MttV in Petrolacosaurus and Saurosternon implies a plesiomorphic, uniform flexor musculature of the foot in both of them. A modern morphology with individualized flexors must have appeared only in Sauria.
Three next taxa, Protorosaurus speneri (late Permian, Germany and England; Gottmann-Quesada and Sander 2009), Sophineta cracoviensis (Early Triassic, Southern Poland; Evans and Borsuk-Białynicka 2009a), and Gephyrosaurus bridensis (Early Jurassic, South Wales fissure fillings; Evans 1981), document the early stages in the evolution of the pes after the bifurcation of the saurian stem into the Lepidosauromorpha and Archosauromorpha. Protorosaurus is one of the most ancient Archosauromorpha, and is represented by four almost complete pes specimens (two of them, left and right juvenile pedes of the same individual). Sophineta is a basal lepidosaurian. Gephyrosaurus is currently regarded as a basal rhynchocephalian (Evans and Jones 2010; Ezcurra et al. 2014). The MttV of Protorosaurus (Fig. 8D) has a rather robust shaft with a broadened proximal part that bears a proximo-medially oriented articular facet for both the dTIV and the calcaneum, and no medially projecting neck-shaped articular region. A somewhat distal placement of the outer process results in a triangular, proximally pointed shape of the proximal end of the bone.
In Protorosaurus the articular facet for dTIV seems to be oval, with the shortest diameter in the dorso-plantar axis, as it is in Osmolskina. The shaft curves medio-distally to maintain the original orientation of the metatarsus, which is also true of Gephyrosaurus and Sophineta. In Protorosaurus the dorsal overlap of the proximal part of the MttV by MttIV, was substantial (Gottmann-Quesada and Sander 2009: fig. 25C), as it also was in Saurosternon (Carroll 1975), and so, the metatarsus was integrated as a whole.
The MttV of Gephyrosaurus shares the same neckless shape of the proximal end, broadened in transverse axis, proximally pointed and lacking a projecting articular part (Fig. 8M). It differs from both Protorosaurus and Saurosternon in the sub-circular outline of its articular facet (Evans 1981: fig. 30C), which is reminiscent of that of Sophineta and Sphenodon. As a whole the bone is more slender than that of Protorosaurus, particularly in its distal part (Fig. 1D). It is substantially concave medially but not inflected (Figs. 2D, 8M). The lateral plantar tubercle is distal in position. A flat ovoid scar located in the proximal part of the plantar surface of the MttV about the middle of its width, was interpreted by Evans (1981: fig. 30B and Fig. 1D herein) as the medial plantar tubercle, and is thus more lateral than usual. Interestingly it is reminiscent of the position of the only plantar tubercle in Sphenodon (Fig. 1B), but is substantially less developed. A possible homology of the medial plantar tubercle in Gephyrosaurus with the only plantar tubercle of Sphenodon is doubtful, in view of the normal medial position of this tubercle in the Late Triassic Planocephalosaurus (Frazer and Walden 1984). Apart from this, the MttV of Planocephalosaurus is exactly like that of Gephyrosaurus in its neckless structure, proximo-medial slope of the facet for dTIV, and very low position of the lateral plantar tubercle. In contrast, the Late Triassic Clevosaurus (Frazer 1988) and the Late Jurassic rhynchocephalians, Homoeosaurus, Kallimodin, Sapheosaurus described by Cocude Michel (1963) all display the long-necked structure of the MttV, but the evidence concerning the inflection is lacking. Other information on foot structure in Rhynchocephalia and MttV in particular, is scarce, in spite of rich record of this group in the Mesosoic (Frazer 1994; Frazer and Benton 1989; Evans and Jones 2010 and references therein).
Compared to Gephyrosaurus (Fig. 1D), Sophineta (Figs. 1C1, 2B) appears more derived in several features of the MttV, such as the long-necked articular part, capped by a medially directed sub-circular facet for the dTIV, and a shaft that is significantly inflexed (Figs. 1C1 versus 2D) and ventrally convex, enhanced by a ventral protrusion of the lateral plantar tubercle. The discrepancy between the MttV structure in Gephyrosaurus and Planocephalosaurus from one side and Sophineta from another, and the current phylogenetic position attributed to these taxa is an interesting problem, which may not be solved herein.
The basic type of MttV referred to as neckless type has been retained not only by Protorosaurus and Gephyrosaurus but also by some early Archosauromorpha (Boreopricea; Figs. 8E, 9: 3), as well as by most Triassic Archosauriformes.
Among the basal Archosauromorpha, Boreopricea funerea Tatarinov, 1978 from the Early Triassic of northern Russia, known from the whole skeleton, seems rather primitive in its decidedly sprawling posture. Recorded is a nearly complete right foot, and an incomplete left one, but the bones have been mounted incorrectly, as already noted by Rieppel et al. (2003). The drawings of the fifth metatarsal by Benton and Allen (1997) suggest that what is labelled dorsal (Benton and Allen 1997: figs. 14G, 15B) is plantar in fact, and vice versa (Benton and Allen 1997: figs. 14H, 15A). A spindle-shaped eminence figured just below half the length of the lateral border of the bone in Benton and Allen (1997: fig. 14G) is the lateral plantar tubercle in my opinion, and thus the aspect is plantar. A convexity of the articular portion of the MttV in this figure (Benton and Allen 1997: fig. 14G), supports my opinion that the presented view is plantar.
What is evident about the fifth metatarsal of Boreopricea is that it is stout and displays the neckless structure (Figs. 8E, 9: 3). According to Benton and Allen’s (1997) reconstruction, the calcaneum and the fourth tarsal contributed to the oblique joint for the fifth metatarsal, as in Protorosaurus (see above). The articular facet for the MttIV may have allowed the MttV to bend more up than down (Benton and Allen 1997: fig 15A). The outer process of MttV seems to be directed proximally, more than in Osmolskina and Euparkeria (Fig. 7B), and is less protruding than in Prolacerta (Fig. 8H) or Macrocnemus (Fig. 8F, G). The lateral plantar tubercle is located within the distal half of the lateral margin, more or less as in Osmolskina. Nothing is known about the inflection. Rieppel et al. (2003) considered Boreopricea to be a basal archosauromorph. Its MttV structure is consistent with this statement.
Basal archosauromorphs, other than Boreopricea, such as the Early Triassic Prolacerta, and the Middle Triassic Pamelaria and Macrocnemus, were studied by Gow (1975), Sen (2003), and Rieppel (1989), respectively and pes structure was reconstructed in each case. Gow’s (1975) material included two and that of Sen one specimen of articulated pes (Fig. 8H, I). Rieppel’s (1989) study, based on six fully preserved pes specimens (four of them in plantar view), demonstrated not only individual and ontogenetic variability, but also biases derived from dislocation of bones and their exposure from different sides (Rieppel 1989: fig. 8). The details of tarsal structure, including the elaboration of the mesotarsal joint, cannot be discussed on the basis of those specimens, but the shape of the MttV may be.
Prolacerta, Pamelaria, and Macrocnemus, are derived in the strong medial protrusion of the articular part of the MttV referred to herein as “long-necked structure” (Fig. 8). In Prolacerta (Gow 1975) and Pamelaria (Sen 2003) the hooking angle is apparently close to 100° (Fig. 8H, I, respectively), more or less as in Sophineta. It is similar in Macrocnemus (Rieppel 1989: fig. 8C, D, F), but is apparently more open (Rieppel 1989: fig. 8B and E), perhaps due to oblique exposure of the specimens. In both, Prolacerta (Fig. 8H) and Pamelaria (Fig. 8I) the articular facet for dTIV seems to be generally medial in orientation. It is similar in Macrocnemus (Rieppel 1989: fig. 8C–F, except for specimen PIMZ T 286, fig. 8B). All three taxa (Fig. 8F, H, I) share a prominent outer process that projects proximally. In Macrocnemus, the lateral plantar tubercle is visible in one specimen (Rieppel 1989: fig. 8E) at about the mid-length of the lateral side of the plantar surface (Fig. 4E). The presence of a medial plantar tubercle is questionable. No information on the degree of inflection is available. The Early Triassic basal rhynchosaur, Mesosuchus, known from some specimens of articulated pedes and only one reconstructed tarsus (Dilkes 1998) shares the long-necked structure with the above discussed basal archosauromorphs, but also with lepidosaurs, which is suggestive of homoplasy, and is here interpreted as evidence of some amount of mobility related to the initial stages of foot rotation.
The mutual relationships of basal archosauromorphs (so called “prolacertiforms”) are not fully resolved (Rieppel et al. 2003), except for the position of Prolacerta as a sister taxon of archosauriforms (Dilkes 1998), that is widely accepted. Pamelaria is considered closely related to Prolacerta according to Sen (2003). Rhynchosaurs are a clade of basal archosauromorphs, that appear as a sister group of the clade of archosauriforms plus Prolacerta (Dilkes 1998; Müller 2004).
Compared to basal Archosauromorpha, MttV morphotype X (Figs. 1F, 5) appears relatively straight and more gracile, its proximal end amounting to half the total length of the bone (Fig. 5C) instead of two thirds in Boreopricea. The medial protrusion of the articular part is just incipient, the articular neck short and thick (Figs. 1F2 and 5), but the overall structure of the bone looks derived. Its articular facet slopes proximo-medially, more medially than in Boreopricea and Protorosaurus. With Macrocnemus, Pamelaria, and Mesosuchus, MttV morphotype X shares a proximally protruding outer process (Fig. 5A2, B2), a character related to the function of the m. peroneus brevis, but difficult to interpret in strict biomechanical terms. The morphology of MttV morphotype X seems to be autapomorphic for an unknown group. Either it falls within the variability range of basal archosauromorphs, or it represents an early stage of locomotor adaptation in kuehneosaurids. The problem must await future studies.
The neckless type occurs in basal archosauriforms, such as Erythrosuchus (Fig. 7A), Euparkeria (Fig. 7B), and Osmolskina (Figs. 1H and 3), and in most basal and earliest archosauromorph Protorosaurs, but differs in Proterosuchus vanhoepeni, regarded as the sister taxon to archosauriforms. The latter taxon has the proximal articular part of MttV more individualized from the shaft and the articular facet for dTIV medial rather than proximo-medial in orientation (Cruickshank 1972), much as in most basal archosauromorphs (Fig. 8F–J) rather than in basal archosauriforms (Fig. 7A, B). If not biased by the reconstruction, it demonstrates the range of random variability at this stage of phylogeny.
The overlap of the proximal ends of the metatarsals that appeared early in diapsid phylogeny persisted in many saurian lines and particularly in lepidosaurs, but mostly between metatarsals I to IV. The overlap between the MttIV and MttV (if correctly reconstructed) has been probably reduced in many Sauria, as shown in the literature (Benton and Allen 1997; Dilkes 1998; Ewer 1965; Gower 1996; Sen 2003; less so in Rieppel 1989), but was significant in Protorosaurus (Figs. 8D, 9). Characteristic of basal Archosauriformes (Sereno 1991) the distally divergent position of the MttV shaft in relation to MttIV (Fig. 7A, B) improves the levering of the pes (see above). Along with a predominance of the lateral plantar tubercle over the medial one (e.g., in Osmolskina) it is regarded here (see Functional aspects) as an indication of a much abducted position of the hind leg and consequently a posterior orientation of the lateral side of the extremity in rest position.
A semi-erect position was ancestral for archosaurs, while basal ornithodirans became fully erect, digitigrade and bipedal (Sereno 1991). The MttV became subparallel to the MttIV and tapered distally (Fig. 7E–G). The same was true of Scleromochlus (Benton 1999) and most basal pterosaurs (Padian 2008), but some of them displayed a more plesiomorphic aspect (Fig. 7D). In basal Dinosauromorpha, the MttV became deflected behind the pes, and had the articular facet for dTIV oriented parallel to the shaft axis (Sereno 1991), as demonstrated by Marasuchus (Fig. 7G) and Silesaurus (Dzik 2003). The importance of the MttV in locomotion gradually diminished as this bone disappeared in later phylogeny of Dinosauromorpha, while the attachment of the gastrocnemius probably moved down the metatarsus.
Summary of evolution of MttV characters in Sauria.—Uniform metatarsals I–V, articulated proximally with distal tarsals I–V are characteristic of the early tertrapods, including earliest diapsids such as late Permian Petrolacosaurus (Reisz 1981) and Saurosternon (Carroll 1975), and are currently considered plesiomorphic in Sauria (Fig. 9: 1).
However, Petrolacosaurus and Saurosternon share a proximal overlap of the metatarsals I–V (dorsally from I to V), equal to integration of the metatarsus, which is important for the development of the levering system of the pes and considered synapomorphic of the Diapsida (Fig. 9: 2) (Lee 1997, Gauthier et al. 1988b). According to Lee (1997) it first appeared at the base of the protorothyridid-diapsid clade (slightly crownwards of Captorhinus). This overlap is persistent in Sauria, but mostly between MttI and IV. It may be significantly reduced between MttV and MttIV in subsequent phylogeny. The large overlap in Protorosaurus (Figs. 8D, 9) may show that the tendency to its reduction appeared after the split of the Sauria into Archosauromorpha and Lepidosauromorpha.
Synapomorphic of Sauria is a presence of the hooked MttV along with a loss of the dTV (Lee 1997). More exactly, synapomorphic of Sauria is a primitively hooked, neckless structure of the hooked MttV (Fig. 9: 3), characterized by a proximal end evenly expanded in medio-lateral axis, without any neck-like protrusion of the articular part, and by a proximo-medial orientation of the articular facet for the dTIV. From the morphological point of view, this structure requires only a small evolutionary reorientation of the articular facet for the dTIV, and is more readily derivable from that of the non-saurian diapsids, than the long-necked structure characterized by the elaborated articular part.
Both, the neckless and the long-necked MttV morphology are scattered within the Sauria. The neckless morphology occurs in archosauromorphans (Protorosaurus, Fig. 8D, and Boreopricea, Fig. 8E, from the late Permian and Early Traissic, respectively). It is present in a basal Late Jurassic rhynchocephalian Gephyrosaurus (Fig. 8M), a Triassic rhynchocephalian Planocephalosaurus, and basal archosauriforms Erythrosuchus (Fig. 7A), Osmolskina, and Euparkeria (Fig. 7B). The long-necked MttV, with medially oriented subcircular facet for dTIV occurs in archosauromorphans Macrocnemus, Prolacerta, Pamelaria, in rhynchosaurian Mesosuchus (Figs. 8F–J, 9: 4) and in lepidosaurians, beginning with Sophineta. (Figs 8L, 9: 4’). It is derived and considered homoplastic in Sauria. The long-necked morphology implies some degree of mobility within the MttV–dTIV joint. The apparent reduction of the contact between MttIV and MttV (Fig. 8H, I) is consistent with this mobility.
A disto-lateral divergence of the MttV in relation to the MttIV (Figs. 7A, B, 9: 3’) is characteristic of basal Archosauriformes (Sereno 1991), including Osmolskina.
A straight, narrow MttV parallel and posterolateral to the metatarsus, but becoming reduced in phylogeny (Figs. 7E, F, 9: 5) is synapomorphic for the Ornithodira (Sereno 1991). Attachment of m. gastrocnemius probably extended distad over the plantar surface of the pes.
A set of characters (Fig. 9: 4’), including a long-necked MttV, inflection of the MttV shaft, ventral protrusion of the lateral plantar tubercle, and sub-circular articular facet for dTIV oriented medially is probably synapomorphic (Fig. 9: 9) for Lepidosauria. Associated with these characters is attenuation or reduction of the MttIV overlap on MttV related to initial stages of development of pes rotation and arcaution. The trend was reversed later in phylogeny by various types of union between MttV and dTIV (Brinkman 1980a) most likely parallel to elaboration of the mesotarsal joint that allowed a rotation of the foot as a unit.
Conclusions
A degree of medial inclination of the proximal end of the hooked fifth metatarsal, varies in Sauria from proximo-medial to directly medial. Although variability of this character is most likely continuous, two types of the hooked MttV may be recognized based on a degree of isolation of the articular part from the stem of this bone. Referred to as a neckless type, evenly expanded proximal end of MttV associated with proximo-medial, rather than medial, orientation of the articular facet for the dTIV, is a state of the hooked fifth metatarsal synapomorphic of Sauria. Derived in Sauria, but developed in parallel in different subgroups, is a long-necked type, in which the articular part of MttV is distinct from its stem and bears a medially oriented facet for the dTIV.
Individualization of the articular part of MttV associated with a possible withdrawal of the overlap of the MttIV on MttV is here considered suggestive of a development of some mobility of MttV in relation to the rest of the pes. This mobility was disadvantageous for the levering system of the pes, but was probably of some advantage for the development of pes rotation. A hypothesized reason for that was that in the early Sauria the rotation of the pes began by lifting the MttV first, while the remaining metatarsus only followed it. A subsequent reintegration of the pes in squamates probably paralleled the elaboration of the mesotarsal joint. The differences between early lepidosaurian and early archosauriform structure of the MttV/dTIV joint relate to the activity of the peroneus brevis muscle, and the rotatory abilities of the pes. On the other hand, the two types of the shaft of MttV, that occur in the Czatkowice assemblage, represent two different solutions form the point of view of the gastrocnemius muscle. The lepidosaurian solution consists of bending the MttV shaft (inflection sensu Robinson 1975) into two parts, a distal one lying on the ground in a plantigrade manner and a proximal one aligned with the ventral surface of the crus and proximal tarsus, and enhancing the lever arm of the pes flexor (Fig. 6E). A straight shaft of the hooked MttV is considered here suggestive of a digitigrade foot position (Fig. 6G). The main adaptive reason lying behind these solutions is the elimination of the pulley problem (first discussed by Schaeffer 1941 in early tetrapods) in the levering system of the foot (Fig. 6F), by straightening of the action line of the gastrocnemius muscle. The propulsion that involved pes rotation in lepidosaurians, in archosauriforms probably relied mostly upon movements of the hidnlimb as a whole. Most likely, these differences were size dependent.
Acknowledgements
I am indebted to the referees Susan E. Evans (University College of London, UK) and Corvin Sullivan (Key Laboratory of Evolutionry Systematics of Vertebrates, Institute of Vertebrate Paleontology and Paleoantropology, Beijing, China) for the huge work they did to correct the manuscript of this paper. I highly appreciate the discussions with Corvin Sullivan that helped me to improve the manusctipt. Thanks are due to Jerzy Dzik (Institute of Paleobiology Polish Academy of Sciences, Warsaw, Poland) for his editorial advises, Andrei G. Sennikov (Paleontological Institute RAN, Moscow, Russia) and Mateusz Tałanda (Faculty of Biology, Warsaw University, Poland) for their comments on the reconstruction of the skeleton of Boreopricea funerea Tatarinov, 1978 from the Paleontological Institute RAN Moscow. I am also grateful to those members of the staff of the Institute of Paleobiology. Polish Academy of Sciences that kindly helped me in technical problems: Ewa Hara for preparation and repair of broken specimens, Cyprian Kulicki for SEM, Marian Dziewński for light photographs and Aleksandra Hołda-Michalska for computer arrangement of the figures.
References
Bennett, C.S. 2001. The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon. Palaeontographica A 260: 1–153.
Benton, M.J. 1985. Classification and phylogeny of the diapsid reptiles. Zoological Journal of the Linnean Society 84: 97–164. Crossref
Benton, M.J. 1999. Scleromochlus taylori and the origin of dinosaurs and pterosaurs. Philosophical Transactions of the Royal Society of London 354: 1423–1446. Crossref
Benton, M.J. and Allen, J.L. 1997. Boreopricea from the Lower Triassic of Russia, and the relationships of the prolacertiform reptiles. Palaeontology 40: 931–953.
Berman, D.S., Reisz, R.R., Scott, D., Henrici, A.C., Sumida, S.S., and Martens, T. 2000. Early Permian bipedal reptile. Science 290: 969–972. Crossref
Borsuk-Białynicka, M. 2008. Evolution of the iliosacral joint in diapsid phylogeny. Neues Jahrbuch für Geologie und Paläontologie 249: 297–311. Crossref
Borsuk-Białynicka, M. and Evans, S.E. 1998. A basal archosauriform from the Early Triassic of Poland. Acta Paleontologica Polonica 48, 649–652.
Borsuk-Białynicka, M. and Evans, S.E. 2003. A basal archosauriform from the Early Triassic of Poland. Acta Palaeontologica Polonica 48: 649–652.
Borsuk-Białynicka, M. and Evans, S.E. 2009a. A new long-necked reptile (Diapsida: Archosauromorpha) from the Early Triassic of Poland. Palaeontologia Polonica 65: 203–234.
Borsuk-Białynicka, M. and Evans, S.E. 2009b. Cranial and mandibular osteology of the early Triassic archosauriform Osmolskina czatkowicensis from Poland. Palaeontologia Polonica 65: 235–282.
Borsuk-Białynicka, M. and Lubka, M. 2009. Procolophonids from the Early Triassic of Poland. Palaeontologia Polonica 65: 107–144.
Borsuk-Białynicka, M. and Sennikov, A.G. 2009. Archosauriform postcranial remains from the Early Triassic karst deposits of Southern Poland. Palaeontologia Polonica 65: 283–326.
Borsuk-Białynicka, M., Cook, E., Evans, S.E., and Maryańska, T. 1999. A microvertebrate assemblage from the Early Triassic of Poland. Acta Palaeontologica Polonica 44: 167–188.
Borsuk-Białynicka, M., Maryańska, T., and Shishkin, M.A. 2003. New data on the age of the bone breccia from the locality Czatkowice 1 (Cracow Upland, Poland). Acta Palaeontologica Polonica 44: 167–188.
Brinkman, D. 1980a. Structural correlates of tarsal and metatarsal functioning in Iguana (Lacertilia: Iguanidae) and other lizards. Canadian Journal of Zoology 58: 277–289. Crossref
Brinkman, D. 1980b. The hind limb step cycle of Caiman sclerops and the mechanics of the crocodile tarsus and metatarsus. Canadian Journal of Zoology 58: 2187–2200. Crossref
Broom, R. 1913. On the South African pseudosuchian Euparkeria and allied genera. Proceedings of the Zoological Society of London 1913: 619–633. Crossref
Carroll, R.L. 1975. Permo-Triassic “lizards” from the Karroo. Palaeontologia Africana 18: 71–87.
Cocude-Michel, M. 1963. Les rhynchocephales et les sauriens des calcaires lithographiques (Jurassique superieur) d’Europe Occidentale. 9–187. Theses Faculte des Sciences, Université de Nancy, Nancy.
Colbert, E.H. 1970. The Triassic gliding reptile Icarosaurus. Bulletin of the American Museum of Natural History 143: 89–142.
Cook, E. and Trueman, C.N. 2009. Taphonomy and geochemistry of a vertebrate macroremains assemblage from the Early Triassic karst deposits at Czatkowice 1, southern Poland. Palaeontologia Polonica 65: 17–30.
Cruickshank, A.R.I. 1972. The proterosuchian thecodonts. In: K.A. Joysey and T.S. Kemp (eds.), Studies in Vertebrate Evolution, 89–119. Oliver and Boyd, Edinburgh.
Dilkes, D.W. 1998. The Early Triassic rhynchosaur Mesosuchus browni and the relationships of basal archosauromorph reptiles. Philosophical Transactions of the Royal Society of London B 353: 501–541. Crossref
Dzik, J. 2003. A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology 22: 556–574. Crossref
Ewer, R.F. 1965. The anatomy of the thecodont reptile Euparkeria capensis Broom. Philosophical Transactions of the Royal Society of London B 248: 379–445. Crossref
Evans, S.E. 1981. The postcranial skeleton of the Lower Jurassic eosuchian Gephyrosaurus bridensis. Zoological Journal of the Linnean Society 73: 81–116. Crossref
Evans, S.E. 1988. The early history and relationships of the Diapsida. In: M.J. Benton (ed.), The Phylogeny and Classification of the Tetrapods, 221–260. Oxford University Press, Oxford.
Evans, S.E. 2009. An early kuehneosaurid reptile from the Early Triassic of Poland. Palaeontologia Polonica 65: 145–178.
Evans, S.E. and Borsuk-Białynicka, M. 2009a. A small lepidosauromorph reptile from the Lower Triassic of Poland. Palaeontologia Polonica 65: 179–202.
Evans, S.E. and Borsuk-Białynicka, M. 2009b. The Early Triassic stem-frog Czatkobatrachus from Poland. Palaeontologia Polonica 65: 79–106.
Evans, S.E. and Jones, M.E.H. 2010. The origin, early history and diversification of lepidosauromorph reptiles. In: S. Bandyopadhyay (ed.), New Aspects of Mesozoic Biodiversity. Lecture Notes in Earth Sciences 132, 27–44. Springer Verlag, Berlin. Crossref
Ezcurra, M.D., Scheyer, T.M., and Butler, R.J. 2014. The origin and early evolution of Sauria: reassessing the Permian saurian fossil record and the timing of the crocodile-lizard divergence. Revisiting the Permian saurian record. PLOS ONE 9 (2): e89165. Crossref
Frazer, N.C. 1988. The osteology and relationships of Clevosaurus Philosophical Transactions of the Royal Society Biological Sciences 321: 125–178.
Frazer, N.C. 1994. Assemblages of small tetrapods from British Late Triassic fissure deposits. In: N.C. Frazer and H.-D. Sues (eds.), In the Shadow of the Dinosaurs: Early Mesosoic Tetrapods, 214–226. Cambridge University Press, Cambridge.
Frazer, N.C. and Benton, M.J. 1989. The Triassic reptiles Brachyrhinodon and Polysphenodon and the relationships of the sphenodontids. Zoological Journal of the Linnean Society 96: 413–445. Crossref
Frazer, N.C. and Walden, G.M. 1984. The postcranial skeleton of the Upper Triassic sphenodontid Planocephalosaurus robinsonae. Palaeontology 27: 575–595.
Gauthier, J.A. 1984. A Cladistic Analysis of the Higher Taxonomic Categories of the Diapsida. 564 pp. Unpublished Ph.D. Dissertation, University of California, Berkeley.
Gauthier, J.A., Estes, R., and de Queiroz, K. 1988a. A phylogenetic analysis of the Lepidosauromorpha. In: R. Estes and G. Pregill (eds.), Phylogenetic Relationships of the Lizard Families, 15–98. Stanford University Press, Stanford.
Gauthier, J.A., Kluge, A.G., and Rowe, T. 1988b. Amniote phylogeny and the importance of cladistics. Cladistics 4: 105–209. Crossref
Goodrich, E.S. 1916. On the classification of the Reptilia. Proceedings of the Royal Society of London B 89: 261–276. Crossref
Gottmann-Quesada, A. and Sander, P.M. 2009. A description of the early archosauromorph Protorosaurus speneri Meyer, 1832, and its phylogenetic relationships. Palaeontographica A 287: 123–220. Crossref
Gow, C.E. 1975. The morphology and relationships of Youngina capensis Broom and Prolacerta broomi Parrington. Palaeontologia Africana 18: 89–131.
Gower, D. 1996. The tarsus of erythrosuchid archosaurs, and implications for early diapsid phylogeny. Zoological Journal of the Linnean Society 116: 347–375. Crossref
Heaton, M.J. and Reisz, R.R. 1982. Phylogenetic relationships of captorhinomorph reptiles. Canadian Journal of Earth Sciences 23: 402–418. Crossref
Hildebrandt, M. 1974. Analysis of Vertebrate Structure. 709 pp. John Wiley & Sons, New York.
Holmes, R.B. 2003. The hind limb of Captorhinus aguti and the step cycle of basal amniotes. Canadian Journal of Earth Sciences 40: 515–526. Crossref
Lee, M.S.Y. 1997. The evolution of the reptilian hindfoot and the homology of the hooked fifth metatarsal. Journal of Evolutionary Biology 10: 53–263. Crossref
Müller, J. 2004. The relationships among diapsid reptiles and the influence of taxon selectio. In: G. Arratia, M.V.H. Wilson, and R. Cloutier (eds.), Recent Advances in the Origin and Early Radiation of Vertebrates, 379–408. Verlag Dr. Friedrich Pfeil, Munchen.
Padian, K. 2008. Were pterosaur ancestors bipedal or quadrupedal?: Morphometric, functional, and phylogenetic considerations. Zitteliana B 28: 21–33.
Padian, K. 2017. Structure and evolution of the ankle bones in pterosaurs and othwer ornithodirans. Journal of Vertebrate Paleontology 37 (5) [published online; doi: 10.1080/02724634.2017.1364651]. Crossref
Parish, J.M. 1986. Locomotor evolution in the hindlimb and pelvis of the Thecodontia (Reptilia: Archosauria). Hunteria 1: 1–35.
Paszkowski, M. 2009. The early Triassic karst of Czatkowice 1, Southern Poland. Palaeontologia Polonica 65: 7–16.
Perrin, A. 1892. Contributions sur l’étude de la myologie comparé: membre posterieur chez un certain nombre de batraciens et de sauriens. Bulletin scientifique de la France et de la Belgique 24: 372–552.
Reisz, R.R. 1981. A diapsid reptile from the Pennsylvanian of Kansas. Museum of Natural History, University of Kansas Publications 7: 1–74.
Rewcastle, S.C. 1980. Form and function in lacertilian knee and mesotarsal joints: a contribution to the analysis of sprawling locomotion. Journal of Zoology 191: 147–170. Crossref
Rieppel, O. 1989. The hind limb of Macrocnemus bassani (Nopcsa) (Reptilia, Diapsida):development and functional anatomy. Journal of Vertebrate Paleontology 9: 736–387. Crossref
Rieppel, O., Fraser, N.C., and Nosotti, S. 2003. The monophyly of Protorosauria (Reptilia, Archosauromorpha): a preliminary analysis. Atti della Società italiana di scienze naturali e del Museo civico di storia naturale di Milano 144: 359–382.
Robinson, P.L. 1962. Gliding lizards from the Upper Keuper of Great Britain. Proceedings of the Geological Society of London 1601: 137–146.
Robinson, P.L. 1975. The functions of the hooked fifth metatarsal in lepidosaurian reptiles. In: M.J.-P. Lehman (ed.), Problèmes Actuelles de Paleontologies – Evolution des Vertebrès. Colloques International du C.N.R.S. 218, 461–483. Paris.
Robinson, P.L. 1967. Triassic vertebrates from upland and lowland. Science and Culture 33: 169–173.
Romer, A.S. 1922 The locomotor apparatus of certain primitive and mammal-like reptiles. Bulletin of the American Museum of Natural History 48: 517–606.
Romer, A.S. 1956. Osteology of Reptiles. 772 pp. University of Chicago Press, Chicago.
Russell, A.P. and Bauer, A.M. 2008. The appendicular locomotor apparatus of Sphenodon and normal-limbed Squamates. In: C. Gans, A.S. Gaunt, and A. Kraig (eds.), Biology of the Reptilia. Volume 21. Morphology I. The Skull and Appendicular Locomotor Apparatus of Lepidosauria, 1–466. Society for the Study of Amphibians and Reptiles, Ithaca.
Santi, G. 1993. Functional bipedalism in some Permo-Triassic reptiles: a possible scale of the degree of efficiency. Studi Trentini di Scienze Naturali Acta Biologica 68: 267–280.
Schaeffer, B. 1941. The morphological and functional evolution of the tarsus in amphibians and reptiles. Bulletin of the American Museum of Natural History 70: 395–472.
Sen, K. 2003. Pamelaria dolichotrachela, a new prolacertid reptile from the Middle Triassic of India. Journal of Asian Earth Sciences 21: 663–681. Crossref
Sereno, P.C. 1991. Basal archosaurs: phyletic relationships and functional implications. Journal of Vertebrate Paleontology 11: 1–49. Crossref
Sereno, P.C. and Arcucci, A. 1994. Dinosaurian precursors from the middle Triassic of Argentina: Marasuchus lilloensis gen. nov. Journal of Vertebrate Paleontology 14: 53–73. Crossref
Snyder, R.C. 1954. The anatomy and function of the pelvic girdle and hind-limb in lizard locomotion. American Journal of Anatomy 95: 1–46. Crossref
Snyder, R.C. 1962. Adaptations of bipedal locomotion of lizards. American Zoologist 2: 191–203.
Sukhanov, V.B. [Suhanov, V.B.] 1968. Obŝaâ sistema lokomocii nazemnyh pozvonočnyh i osobiennosti perdiviženiȃ nižših tetrapod. 227 pp. Paleontologičeskij Institut Akademii Nauk SSSR, Nauka, Leningrad.
Sullivan, C. 2010 The role of the calcaneal “heel” as a propulsive lever in basal archosaurs and extant monitor lizards. Journal of Vertebrate Paleontology 30: 1422–1432. Crossref
Shishkin, M.A. and Sulej, T. 2009. The Early Triassic temnospondyls of the Czatkowice 1 tetrapod assemblage. Palaeontologia Polonica 65: 31–78.
Tatarinov, L.P. 1978. Triassic prolacertilians of the USSR [in Russian]. Paleontologičeskij žurnal 1978 (4): 88–100.
Wellnhofer, P. 1991. The Illustrated Encyclopedia of Pterosaurs. 192 pp. Crescent Books, New York.
Acta Palaeontol. Pol. 63 (3): 417–434, 2018
https://doi.org/10.4202/app.00444.2017