

Three new bivalve genera from Triassic hydrocarbon seep deposits in southern Turkey
STEFFEN KIEL
Kiel, S. 2018. Three new bivalve genera from Triassic hydrocarbon seep deposits in southern Turkey. Acta Palaeontologica Polonica 63 (2): 221–234.
Three new bivalve genera and species are described from Upper Triassic hydrocarbon seep deposits from the Kasımlar shales in the Taurus Mountains in southern Turkey. Terzileria gregaria and Kasimlara kosuni belong to the carditiid family Kalenteridae, and Aksumya krystyni belongs to the anomalodesmatan superfamily Pholadomyoidae. A single specimen is described in open nomenclature as Kasimlara sp. due to its significantly more angular shell profile compared to K. kosuni. The kalenterids Terzileria and Kasimlara narrow the stratigraphic gap between two seep-inhabiting “modiomorphid” clades: the Silurian–Devonian Ataviaconcha and the Late Jurassic–Cretaceous Caspiconcha. This raises the questions whether these four genera are members of a single phylogenetic lineage that continuously inhabited deep-sea hydrocarbon seeps from the Silurian to the Cretaceous (the “single lineage hypothesis”), or are repeated offshoots of various lineages that convergently developed similar morphological adaptations to this habitat (the “repeated colonization hypothesis”). Aksumya represents the first anomalodesmatan genus that appears to be restricted to the seep environment, considering that all previous claims of seep-inhabiting anomalodesmatans are questionable.
Key words: Bivalvia, Pholadomyoidae, Kalenteridae, hydrocarbon seeps, Triassic, Taurus Mountains, Turkey.
Steffen Kiel [steffen.kiel@nrm.se], Swedish Museum of Natural History, Department of Palaeobiology, Box 50007, 10405 Stockholm, Sweden.
Received 8 February 2018, accepted 16 March 2018, available online 23 April 2018.
Copyright © 2018 S. Kiel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Deep-sea hydrothermal vents and methane seeps harbor unique faunal associations relying on chemosynthetic primary production (Sibuet and Olu 1998; Van Dover 2000). These animals appear to have an evolutionary history largely independent of that of photosynthesis-based food chains and may be buffered from extinction events affecting shallow-water ecosystems (Tunnicliffe 1992; Little and Vrijenhoek 2003; Kiel 2015). Mollusks are a major constituent of this fauna at present-day vents and seeps, and have become a model group for studies on their physiology, adaptations, and biogeography (Childress et al. 1986; Peek et al. 1998; Distel et al. 2000; Lorion et al. 2013; Kiel 2016; Johnson et al. 2017). Due to the strong link of these faunas to geological features, Earth’s history and the fossil record play a major role in understanding their evolution (Campbell and Bottjer 1995; Tunnicliffe et al. 1996; Kiel and Little 2006; Johnson et al. 2010; Vrijenhoek 2013).
Paramount to linking Recent taxa to their fossil counterparts, and hence to understand their evolutionary history and phylogenetic relationships, is a proper documentation and identification of the fossil species. The Cenozoic fossil record of mollusks especially in hydrocarbon seeps is relatively well known (Squires and Goedert 1991; Amano and Kiel 2007; Saether et al. 2010; Amano and Jenkins 2011; Kiel and Amano 2013; Kiel and Taviani 2017; and references therein) and also the number of Late Jurassic to Cretaceous seep faunas is increasing (Kaim et al. 2008, 2014, 2017; Kiel 2013; Hryniewicz et al. 2017a; and references therein). Beyond the Late Jurassic, however, the fossil record of seeps and especially of properly described seep-inhabiting mollusks, is sparse (Jakubowicz et al. 2017; Hryniewicz et al. 2017b). The purpose of the present contribution is to describe three new bivalve genera from recently reported Upper Triassic seep deposits in southern Turkey, and to briefly discuss their evolutionary implications.
Institutional abbreviations.—NRM, Swedish Museum of Natural History, Stockholm, Sweden.
Material and methods
The sources of the present material are limestone deposits from the Kasımlar Basin in the Anamas Akseki Autochthon in the Taurus Mountains in southern Turkey (Fig. 1; Kiel et al. 2017). These sites were first mentioned in the literature in a report on their brachiopods (Ager et al. 1978). The Kasımlar Basin is interpreted as a small ocean basin partially bordered by carbonate platforms and filled by “deeper water” (cf. Lukeneder et al. 2012), siliciclastic sediments known as Kasımlar Formation or Kasımlar shales (Gutnic et al. 1979). Within these siliciclastics, three seep deposits were found on a slope on the north side of the road just West of the village of Yakaafşar, at the turnoff to Terziler, referred to as Terziler I–III, and two were found about six km NW of the village of Dumanlı, referred to as Dumanlı I and II, during field work in 2015. Further material was collected by Leopold Krystyn (Vienna, Austria) in 1973 from the same area as the Terziler I–III localities; this site is here referred to as “Leo’s Terziler seep deposit”. The seep deposits were dated by conodonts that were etched from the carbonates; the Dumanlı sites are dated as middle to late early Norian based on Epigondolella triangularis, the Terziler sites are dated as late Carnian based on Quadralella polygnathiformis (Tekin 1999; Chen et al. 2015; Kiel et al. 2017). A more detailed description of these sites and their fauna can be found in Kiel et al. (2017).
The specimens were extracted from the rock matrix using hammer and chisel in the field, and a pneumatic micro-hammer in the lab. The material is housed in the paleontological collection of the Swedish Museum of Natural History in Stockholm (NRM); figured specimens are numbered Mo184051–184074. All specimens except polished surfaces were coated with ammonium chloride prior to photography.
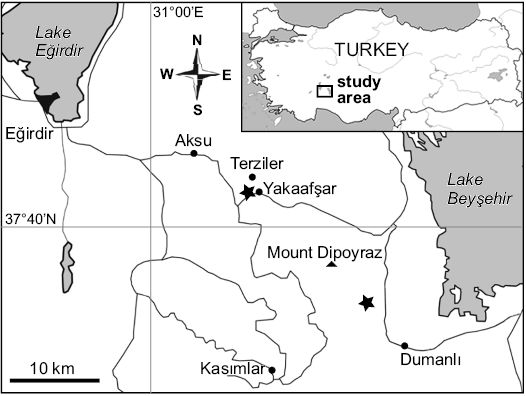
Fig. 1. Locality map of the Late Triassic seep deposits (stars), in southern Turkey, where the here described bivalves were found (from Kiel et al. 2017).
Systematic paleontology
Class Bivalvia Linnaeus, 1758
Clade Archiheterodonta Giribet in Taylor et al. 2007
Order Carditida Dall, 1889
Family Kalenteridae Marwick, 1953
Genus Terzileria nov.
Etymology: For the village of Terziler, near the type locality.
Type species: Terzileria gregaria sp. nov.; see below.
Species included: The type species and possibly also the “permophorid” reported from a Norian, Late Triassic seep deposit in eastern Oregon, USA (Peckmann et al. 2011).
Diagnosis.—Modioliform shell, subterminal umbones, elongate-lenticular escutcheon, shell surface with growth lines and faint posterodorsal groove; hinge plate edentulous, ligament nymph short and close to shell margin, elongate posterior lateral tooth quite distinct in left valve but less so in right valve; anterior adductor muscle scar deep, oval to almost round, positioned at anterior end of shell; anterior pedal retractor scar not fused with adductor muscle scar; pallial line entire and running close to ventral shell margin, no sinus; posterior adductor muscle scar round with projection (probably fused posterior pedal retractor scar) on anteroventral side.
Remarks.—Most similar to Terzileria is the Permian Netschajewia Likharev, 1925, with its modioliform outline, edentulous hinge plate, and the elongate posterior lateral tooth being stronger in the left valve. Terzileria differs from Netschajewia by having an elongate-lenticular escutcheon (lacking in Netschajewia) and a more-or-less evenly convex globosity of the shell, whereas Netschajewia has a distinct broad ridge running from the umbo to the posteroventral margin (Murchison et al. 1845). The available descriptions and illustrations of the type species of Netschajewia (i.e., Pleurophorus modioliformis King in de Verneuil and Murchison, 1844 = Mytilus pallasi de Verneuil in Murchison et al., 1845) did not discuss or show the anterior pedal retractor muscle scar, the posterior adductor muscle scar and the possible presence of a pallial sinus (Murchison et al. 1845; King 1850; Newell 1957; Chavan 1969).
Also very similar to Terzileria is the Jurassic–Cretaceous seep-restricted genus Caspiconcha Kelly in Kelly et al. 2000. They differ by the following features: (i) in Caspiconcha the anterior adductor muscle scar is in a more dorsal position than in Terzileria; (ii) in Caspiconcha the anterior adductor muscle scar and the pedal retractor scar are fused, whereas in Terzileria they are separated; (iii) in Terzileria the pallial line is closer to the shell margin than in Caspiconcha; (iv) secondary pallial attachment scars tend to be found in the anterior half of the shell in Caspiconcha, whereas in Terzileria they tend to occur more posteriorly (Kelly et al. 2000; Kiel et al. 2010; Jenkins et al. 2013, 2018).
The type species of Kalentera Marwick, 1953 is similar in shell outline, the lateral tooth in the left valve and the shape of the anterior adductor muscle scar (Marwick 1953; Chavan 1969) but is clearly distinct from Terzileria by having cardinal teeth. Other species of Kalentera are more trapezoidal in outline and have a distinctive angular ridge running from umbo to the posteroventral corner (Grant-Mackie 1979; Covacevich et al. 1991; Damborenea 2004).
The genus Healeya Hautmann, 2001 has a similarly shaped pallial line, anterior adductor muscle scars and a separated pedal retractor scars, but clearly differs from Terzileria by its strong and sharp diagonal keel running from the umbo to the posteroventral margin, and by its thicker hinge plate (Hautmann 2001). Similar to Terzileria regarding modioliform shell and subterminal umbo is the Late Triassic Myoconcha (Pseudomyoconcha?) auriculata Broili, 1903 as figured in Hautmann (2001), but it differs from Terzileria by having fine radial ribs on the shell surface. Myoconcha Sowerby, 1824 has a narrower and more pointed anterior margin than Terzileria, and possesses cardinal teeth, whereas Terzileria is edentulous (Chavan 1969). The Late Triassic Ouamouia grantmackie Campbell, 1984 from New Caledonia and New Zealand is more bean-shaped than Terzileria (by having a broader anterior margin and a less wide posterior margin), has a much stronger hinge plate that features cardinal teeth, a smaller anterior adductor muscle scar, and fine radial striation on the anteroventral shell surface (Campbell 1984). The type species of Triaphorus Marwick, 1953, Triaphorus zealandicus (Trechmann, 1917), also has a broader anterior margin and a narrower posterior margin compared to Terzileria, and possesses cardinal teeth (Chavan 1969; Damborenea 2004). Most other kalenterid genera differ from Terzileria by possessing cardinal teeth (i.e., Permophorus, Celtoides, Pseudopermophorus, Curionia, Myoconcha, and Pseudomyoconcha; see Chavan 1969).
The new genus Kazimlara described below from the same seep deposits differs by being much more inflated, having a much smaller anterior adductor muscle scar, and by showing a lateral curvature of the shell that is not seen in Terzileria.
Stratigraphic and geographic range.—Late Triassic hydrocarbon seep deposits in Turkey and possibly also in Oregon.
Terzileria gregaria sp. nov.
Figs. 2, 3.
2017 Caspiconcha-like bivalve; Kiel et al. 2017: fig. 4B, C.
Etymology: For its gregarious occurrence at the type locality.
Type material: Holotype: Mo184051, an internal mold showing features of the pallial line and muscle attachment scars. Paratypes: Mo184052–184060 from type locality.
Type locality: The Terziler III seep deposit, southern Turkey.
Type horizon: Late Carnian, Late Triassic seep deposits of the Kasımlar shales.
Material.—The type material and 75 specimens from Terziler III; five specimens from Terziler II; one specimen from Leo’s Terziler seep deposit; see Table 1 for measurements.
Table 1. Measurements (in mm) of Terzileria gregaria sp. nov.
|
Specimen number |
Locality |
Length |
Height |
Width |
|
Mo184051 |
Terziler III |
21.5 |
12 |
8 |
|
Mo184052 |
Terziler III |
28 |
13 |
11 |
|
Mo184053 |
Terziler III |
16.5 |
10 |
6.5 |
|
Mo184055 |
Terziler III |
25 |
13 |
9 |
|
Mo184056 |
Terziler III |
25 |
14 |
9 |
|
Mo184057 |
Terziler III |
20 |
11 |
8.2 |
|
Mo184058 |
Terziler III |
23.3 |
12.5 |
9 |
|
Mo184060 |
Terziler III |
25 |
14 |
9.5 |
|
Mo184075 |
Terziler III |
28 |
15.7 |
11.2 |
|
Mo184076 |
Terziler III |
25 |
13 |
10.5 |
|
Mo184077 |
Terziler III |
24 |
11.5 |
8.5 |
|
Mo184078 |
Terziler III |
19 |
10 |
7.2 |
|
Mo184079 |
Terziler III |
19 |
10 |
7.8 |
|
Mo184080 |
Leo’s Terziler site |
33 |
15 |
13 |
Diagnosis.—As for genus.
Description.—Shell elongate modioliform, largest specimen 33 mm long; length:height ratio about 1.7–2.0; moderately inflated, width:length ratio about 0.38–0.40; beak subterminal, prosogyrate; shell surface smooth except for fine growth increments; escutcheon elongate-lenticular and marked by a sharp ridge; ligament nymph short, slightly less than 1/3 of shell length, close to shell margin, indicating external ligament; short, heart-shaped lunule. Posterodorsal margin very gently convex, posterior end well rounded, ventral margin gently convex, anterior margin truncate and pointing anteroventrally. Anterior adductor muscle scars D-shaped, steeply deepening toward the anterior, terminating anteriorly at a triangular wall perpendicular to the anterior-posterior shell axis; concentric growth increments. Anterior pedal retractor scars elongate-oval, nearly parallel to anterior-posterior shell axis, positioned below and just anterior to the umbones. Pallial line close and subparallel to ventral and posterior shell margin with a distinct bulge just posterior to the anterior adductor muscle scar; reaches the posterior adductor scar at the center of its posterior margin; some specimens show strong secondary pallial attachment scars radiating toward the shell’s interior, especially along the ventral and posterior margins. Posterior adductor scar positioned in the middle of the posterior half of the shell, roundish, with a sinuous indentation underneath the onset of the posterior pedal retractor scar, which projects spine-like from the anterior side of the dorsal margin of the adductor scar. The hinge plate is edentulous, the posterior lateral tooth is elongate, and is well-developed in the left valve but inconspicuous in the right valve.
Remarks.—The “possible permophorid” reported from a Late Triassic seep deposit in Oregon, USA (Peckmann et al. 2011), might also belongs to Terzilieria, and differs from T. gregaria by being more elongate and having a broader anterior margin. The Early Jurassic Myoconcha neuquena Leanza, 1940 was recently discussed as an early member or possible ancestor of Caspiconcha (Jenkins et al. 2018). It could therefore, at least stratigraphically speaking, be a link between Terzileria and Caspiconcha. Myoconcha neuquena differs from T. gregaria mainly by having the anterior pedal retractor and adductor muscle scars fused, by its oblique posterior adductor muscle scar with a much broader ventral extension, and by its thicker hinge plate (as shown in Griffin and Pastorino 2006: figs. 4-1, 4-2).
Stratigraphic and geographic range.—Late Carnian, Late Triassic seep deposits in the Kasımlar shales in southern Turkey.
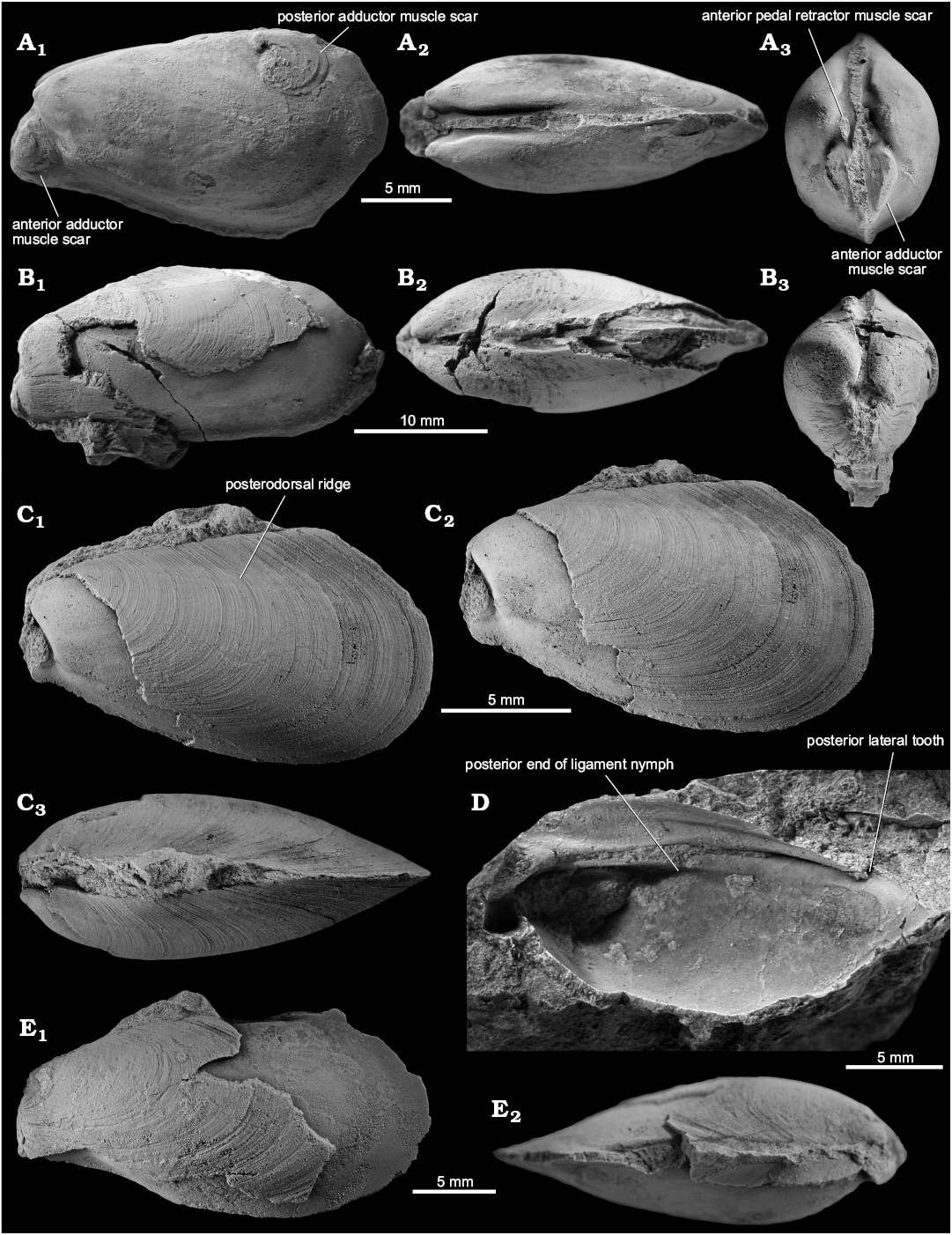
Fig. 2. The kalenterid bivalve Terzileria gregaria gen. et sp. nov. from the late Carnian, Late Triassic Terziler III seep deposit in southern Turkey. A. Mo184051, holotype, internal mold, left valve (A1), dorsal (A2) and anterior (A3) views. B. Mo184052, paratype, specimen with shell partially preserved, left valve (B1), dorsal (B2) and anterior (B3) views. C. Mo184053, paratype, small specimen with shell preserved on most of posterior side, left valve (C1), the same at slightly different angle (C2), dorsal view (C3). D. Mo184054, fragmentary but articulated specimen, interior of the dorsal side, showing the edentulous hinge. E. Mo184055, paratype, large specimen showing outer shell surface of the anterior half of the shell; left valve (E1) and dorsal view (E2).
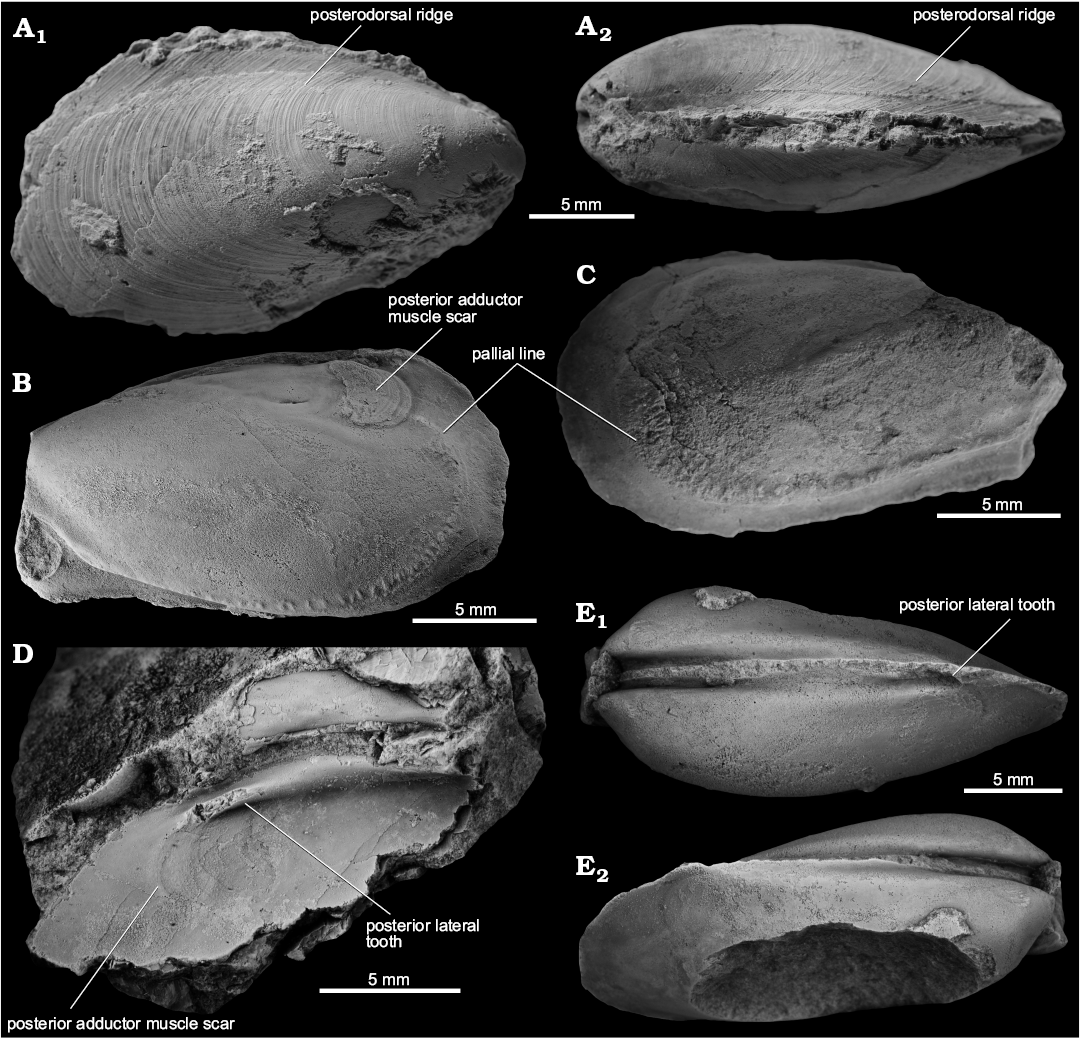
Fig. 3. The kalenterid bivalve Terzileria gregaria gen. et sp. nov. from the late Carnian, Late Triassic Terziler III seep deposit in southern Turkey. A. Mo184056, specimen with preserved shell showing irregular growth increments; right valve (A1) and dorsal view (A2). B. Mo184057, left valve of an internal mold, note mantle muscle scars along the pallial line. C. Mo184058, right valve of a specimen. D. Mo184059, shell fragment preserving the posterodorsal part of the inner shell surface of a left valve. E. Mo184060, internal mold showing the impression of the posterior lateral tooth in the left valve (E1) and the lack of this feature in the right valve (E2).
Genus Kasimlara nov.
Etymology: For its occurrence in the Kasımlar shales in southern Turkey.
Type species: Kasimlara kosuni sp. nov.; see below.
Species included: The type species and Kasimlara sp. (described below).
Diagnosis.—Strongly inflated, laterally curved, modioliform shell with subterminal umbo, small but deep anterior adductor muscle scar, pallial line with very small, pointed sinus.
Remarks.—Kasimlara kosuni sp. nov. resembles the type species of Healeya, H. gonoides (Healey, 1908) regarding shell outline, inflation, and the strong diagonal ridge (Hautmann 2001). But H. gonoides has a distinct anterior pedal retractor scar that is not apparent in Kasimlara kosuni, and although K. kosuni is only known from internal molds, it is obvious that its hinge plate is not as strong as in H. gonoides. Furthermore, the strong diagonal ridge across the shell surface has two strong keels in H. gonoides whereas Kasimlara sp. shows only one keel.
The main differences between Kasimlara and Terzileria described above are (i) the much smaller anterior adductor muscle scar in Kasimlara, (ii) the much stronger inflation of Kasimlara, (iii) the lack of lateral shell curvature in Terzileria, and (iv) the presence of a small pallial sinus in Kasimlara, which Terzileria lacks.
Stratigraphic and geographic range.—Late Triassic hydrocarbon seep deposits in Turkey.
Kasimlara kosuni sp. nov.
Figs. 4, 5.
2017 new genus of bivalve clade Modiomorphida; Kiel et al. 2017: fig. 4D.
Etymology: For Erdal Koşun (Antalya, Turkey).
Type material: Holotype: Mo184061, an internal mold of a large left valve. Paratypes: Mo184062–184064) from the type locality.
Type locality: The Terziler III seep deposit (southern Turkey).
Type horizon: Late Carnian, Late Triassic seep deposits of the Kasımlar shales.
Material.—The type material and three further specimens from the type locality; see Table 2 for measurements.
Table 2. Measurements (in mm) of Kasimlara kozuni sp. nov., * incomplete.
|
Specimen number |
Locality |
Length |
Height |
Width |
|
Mo184061 |
Terziler III |
57 |
26 |
11 (single) |
|
Mo184063 |
Terziler III |
20 |
9 |
10 |
|
Mo184064 |
Terziler III |
21* |
10 |
12.2 |
|
Mo184081 |
Terziler III |
36 |
15 |
18 |
|
Mo184082 |
Terziler III |
32* |
21 |
19.5 |
|
Mo184083 |
Terziler III |
29.5 |
13 |
8 (single) |
Diagnosis.—As for genus.
Description.—Strongly inflated, modioliform shell that is laterally curved to some degree, with the direction of curving differing between specimens; umbo subterminal, strong ridge running from umbo to posteroventral margin; anterior adductor muscle scar small but deep, cone-shaped pointing toward the anterior; onset of pallial line posterior to anterior adductor muscle scar, distant from ventral margin and running subparallel to it; secondary pallial attachment scars occur as pits at the anterior side of the shell, along the ventral margin they are fine but numerous and strongly inclined anteriorly; very small pallial sinus, posterior adductor muscle scar indistinct, round; inner shell surface with fine radial striations close to the dorsal and ventral margins, and pustules in the area between. The largest specimen reaches 57 mm in length, 26 mm in height, and the single valve is 11 mm thick.
Remarks.—The lateral curvature of the shell is either to the left, as in the holotype (meaning that the commissure of the right valve is concave, that of the left valve is convex), whereas the two paratypes illustrated in Fig. 5 are curved to the right. I have not seen any specimen without lateral curvature.
Stratigraphic and geographic range.—Late Triassic hydrocarbon seep depossit in Turkey.
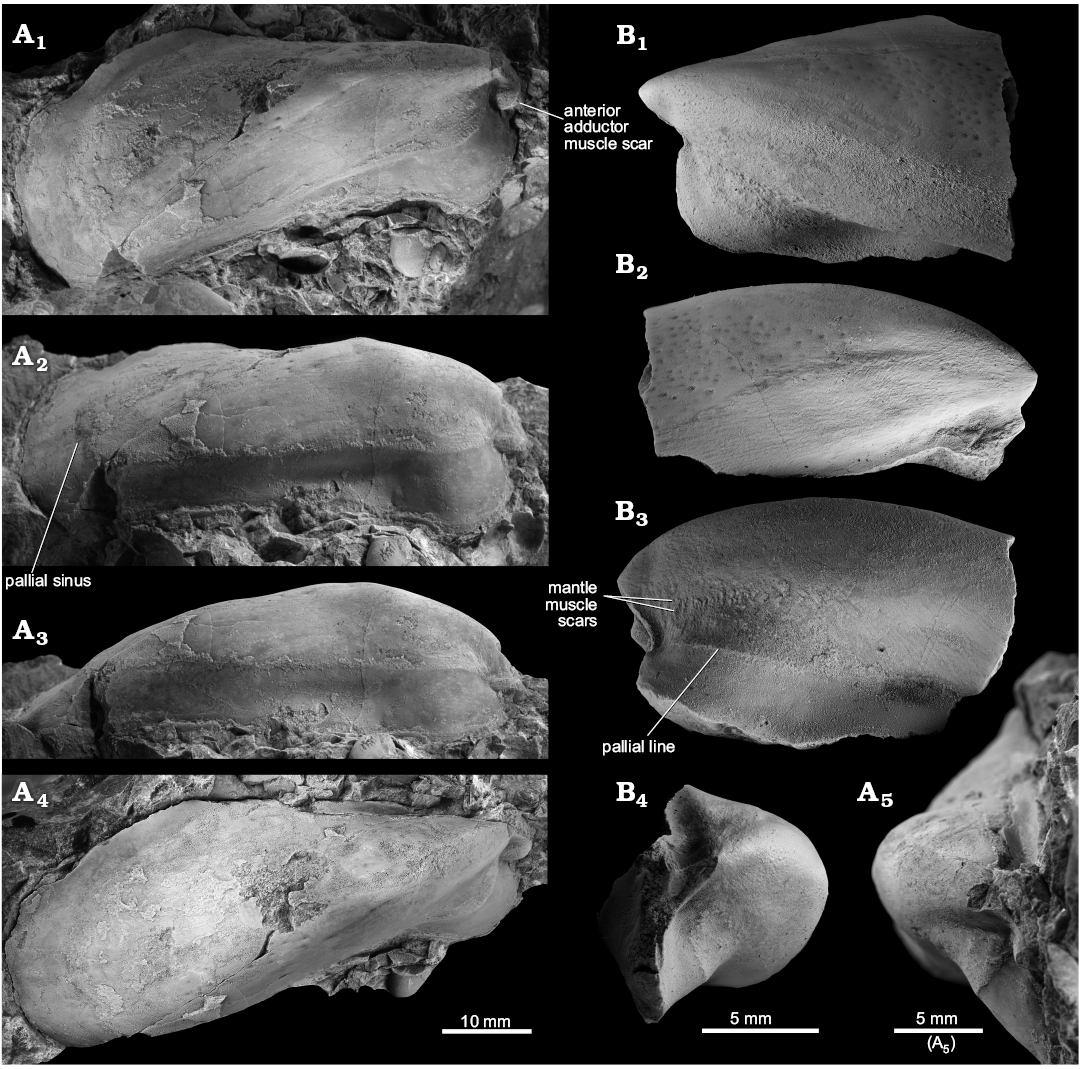
Fig. 4. The kalenterid bivalve Kasimlara kosuni gen. et sp. nov. from the late Carnian, Late Triassic Terziler III seep deposit in southern Turkey. A. Mo184061, holotype, large internal mold of right valve; lateral (A1), lateral-ventral (A2), ventral (A3), lateral-dorsal (A4), and anterior (A5) views. B. Mo184062, internal mold of anterior part of left valve; showing tubercles (as pits in the mold) on the lateral side of the shell’s interior (B1), fine radial striation on the dorsal inner side of the shell (B2), ventral view on the dorsal inner side of the shell (B3), anterior side, note the strong inflation of the shell (B4).
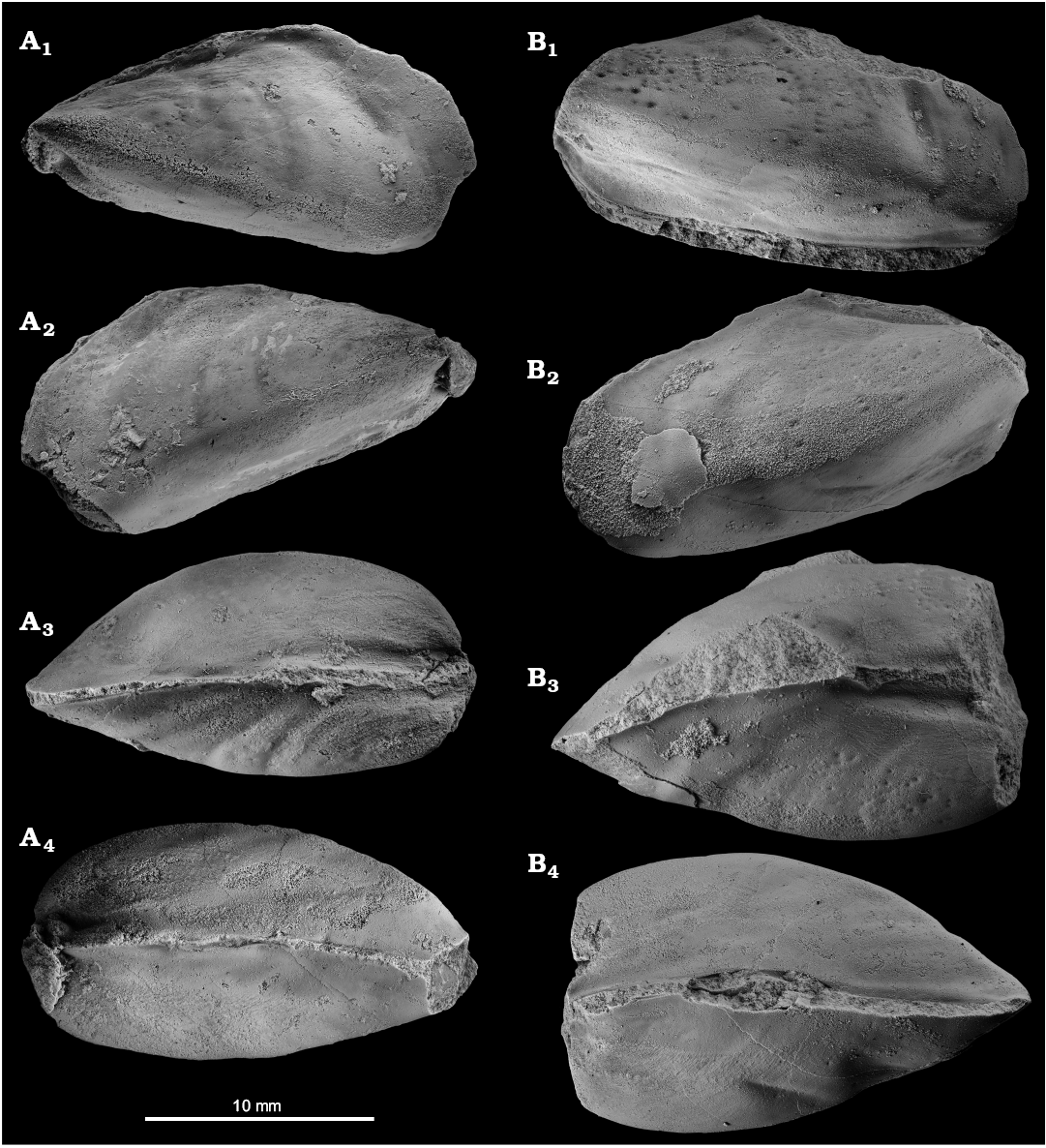
Fig. 5. The kalenterid bivalve Kasimlara kosuni gen. et sp. nov. from the late Carnian, Late Triassic Terziler III seep deposit in southern Turkey. A. Mo184063, internal mold of an articulated specimen; left (A1) and right (A2) valves, dorsal (A3) and ventral (A4) sides, showing the curvature of the shell. B. Mo184064, internal mold of articulated specimen; left (B1) and right (B2) valves, dorsal (B3) and ventral (B4) sides, showing the curvature of the shell; note the strong, healed predation scars on the posterior side of the left valve (B1) and on the posteroventral side of the right valve (B4).
Kasimlara sp.
Fig. 6.
Material.—Mo184065, a single left valve from the Terziler II seep deposit (southern Turkey), late Carnian, Late Triassic of the Kasımlar shales.
Description.—Strongly inflated, mytiliform to modioliform shell that is laterally slightly curved, 35 mm long, 15 mm high, single valve 7 mm thick; posterodorsal margin straight, anterior margin evenly curved, reaching the ventral margin at the posterior end of the shell; ventral margin nearly straight; umbo subterminal, strong, angular ridge running from umbo to posteroventral margin; shell surface smooth except for commarginal growth increments.
Remarks.—Details of the anterior margin and of the internal features of the shell (hinge, pallial line, muscle scars) are unknown. The anterior side of the shell is more pointed than in Kasimlara kosuni and the ridge running from the umbo to the posteroventral margin has an angular profile, which differs from the more curved profile of this ridge in Kasimlara kosuni. Whether these differences fall within the phenotypic variation of Kasimlara kosuni, or if this specimen belongs to a separate species cannot be decided with the limited available material; hence the specimen is left in open nomenclature.
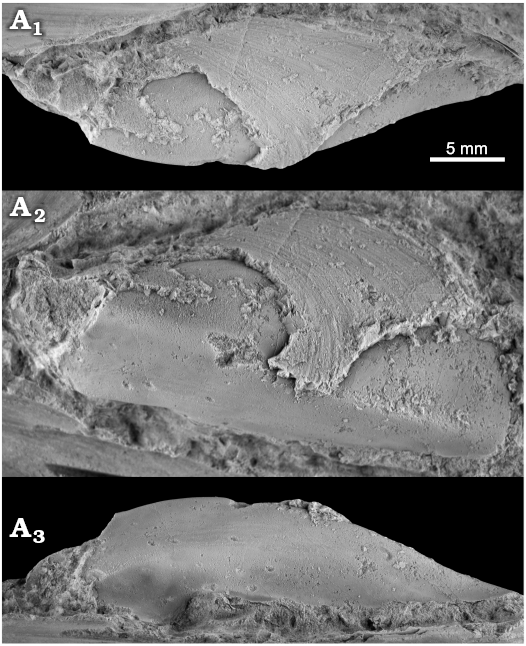
Fig. 6. The kalenterid bivalve Kasimlara sp. from the late Carnian, Late Triassic Terziler II seep deposit in southern Turkey. Mo184065, single left valve; view on the dorsal (A1) and the ventral (A3) sides, lateral view (A2).
Clade Anomalodesmata Dall, 1889
Superfamily Pholadomyoidea Gray, 1847
Family uncertain
Genus Aksumya nov.
Etymology: Combined from the town Aksu near the type locality, and the bivalve genus Mya.
Type species: Aksumya krystyni sp. nov; see below.
Species included: The type species and possibly also the “nuculanid” from a Norian, Late Triassic seep deposit in Oregon (Peckmann et al. 2011).
Diagnosis.—Shell nuculaniform with elongated, tapering posterior side, broad umbonal sulcus and sinuous ventral margin, surface sculpture of commarginal growth lines and fine pustules; hinge plate with two blunt teeth in each valve and a short and strong ligament nymph holding an calcified external ligament; shell interior with radial striation; anterior adductor muscle scar roundish to quadrate, fused dorsally with anterior pedal retractor scar; pallial line entire and close to ventral margin, no pallial sinus.
Remarks.—Regarding its overall shell shape, muscle scar and hinge characters, and the fine pustules on the shell surface, Aksumya can safely be placed among the Pholadomyoidea. But assigning it to any of the currently recognized families is rather difficult. With some Paleozoic Grammysiidae (= Sanguinulitidae) it shares the broad umbonal sulcus and sinuous ventral margin, and possibly its hinge dentition (i.e., Tellinomorpha, cf. Morris and Dickins 1991). However, similar shell shapes are also found among the Mesozoic–Recent Pholadomyidae (Pachymya, Machomya), and the hinge features shown here on polished surfaces and interpreted as teeth also resemble the pits for internal ligaments documented for Pholadomya (Newell 1969).
Almost identical regarding shell shape, external surface and hinge characters is the Viséan, early Carboniferous genus Tellinomorpha de Koninck, 1885 (type T. cuneiformis de Koninck, 1885). However, there are several reasons why I hesitate placing Aksumya krystyni in this genus: (i) Tellinomorpha is based on a single right valve with unknown internal features; (ii) the posterior siphonal area is broken in that single specimen, hence its length is unknown, and furthermore, it is demarcated by the upturn of the growth lines (cf. Morris and Dickins 1991) and this upturn is angular in T. cuneiformis but gently rounded in Aksumya krystyni; (iii) the only known specimen of T. cuneiformis is about 100 million years older than Aksumya krystyni and I find it unlikely that such decent-sized bivalves (exceeding 70 mm in length) should have remained unnoticed during such a long geologic time interval, spanning most of the Carboniferous, the well-sampled Permian, and half of the Triassic.
Among the Pholadomyidae, species of Arcomya Roemer, 1839 are less inflated, have much narrower gapes anterior and posterior of the umbones, the pallial line crosses the anteroventral margin obliquely, whereas it largely follows the ventral shell margin in Aksumya, and pallial line and posterior adductor muscle scar are more distant from the posterior margin than in Aksumya. In Pachymya Sowerby, 1826 the shell is more inflated posteriorly and also has pallial line and posterior adductor muscle scar more distant from the posterior margin than Aksumya. Machomya de Loriol in de Loriol and Cotteau, 1868 has an anterior internal rib, whereas Aksumya has such a rib on the posterior side. Externally, shells of Homomya Agassiz, 1843 and of Pleuromya Agassiz, 1842 are quite similar, but both genera have a deep pallial sinus, which Aksumya lacks.
Stratigraphic and geographic range.—Late Triassic hydrocarbon seep deposits in Turkey and possibly also in Oregon.
Aksumya krystyni gen. et sp. nov.
Figs. 7–9.
2017 new genus of bivalve clade Anomalodesmata; Kiel et al. 2017: fig. 4A.
Etymology: For Leopold Krystyn (Vienna, Austria).
Type material: Holotype: Mo184067, a specimen with preserved shell surface. Paratypes: Mo184066, Mo184068–184074 from the type locality.
Type locality: Leo’s Terziler seep deposit, southern Turkey.
Type horizon: Late Carnian, Late Triassic seep deposits of the Kasımlar shales.
Material.—Ten specimens from the type locality, two from Terziler I, six specimens and many fragments from Terziler II, 14 specimens from Terziler III, 18 specimens and many fragments from Dumanlı I, 12 specimens and many fragments from Dumanlı II; southern Turkey, late Carnian, Late Triassic of the Kasımlar shales. See Table 3 for measurements.
Table 3. Measurements (in mm) of Aksumya krystyni sp. nov., * incomplete.
|
Specimen number |
Locality |
Length |
Height |
Width |
|
Mo184066 |
Leo’s Terziler site |
48 |
22 |
14.5 |
|
Mo184067 |
Leo’s Terziler site |
56 |
29 |
19.5 |
|
Mo184068 |
Leo’s Terziler site |
61 |
29 |
19 |
|
Mo184069 |
Leo’s Terziler site |
73 |
ca. 34 |
23 |
|
Mo184070 |
Leo’s Terziler site |
53 |
26 |
7 (single) |
|
Mo184071 |
Leo’s Terziler site |
58.5 |
29 |
18 |
|
Mo184072 |
Leo’s Terziler site |
49* |
28 |
18 |
|
Mo184084 |
Leo’s Terziler site |
64 |
35 |
22 |
|
Mo184085 |
Leo’s Terziler site |
52* |
28 |
18 |
|
Mo184086 |
Leo’s Terziler site |
46 |
24 |
14 |
|
Mo184074 |
Terziler III |
36* |
18 |
15 |
|
Mo184087 |
Terziler III |
38 |
19 |
12 |
|
Mo184088 |
Terziler III |
22* |
13 |
4 (single) |
|
Mo184089 |
Terziler III |
24 |
12 |
7 |
|
Mo184090 |
Terziler III |
16 |
9 |
6 |
|
Mo184073 |
Dumanlı I |
56* |
35* |
21 |
|
Mo184091 |
Dumanlı II |
71 |
36 |
14 |
|
Mo184092 |
Dumanlı II |
65* |
41 |
21 |
|
Mo184093 |
Dumanlı II |
79 |
36* |
|
|
Mo184094 |
Dumanlı II |
65* |
38 |
20 |
|
Mo184095 |
Dumanlı II |
66.5 |
31 |
9 (single) |
Diagnosis.—As for genus.
Description.—Elongate nuculaniform shell, umbo slightly prosogyrate, positioned in the anterior half of the shell; surface sculptured by commarginal growth lines and tiny, irregularly distributed spines; ligament at least partially external and calcified; anterodorsal margin running more-or-less straight down to about 2/5 of the shell height, anterior margin below rounded or truncate; ventral margin straight to slightly concave; anterior variable, ranging from broad and blunt, to tapering into a blunt, pointed, posteroventral end, to acute-rounded and pointing slightly upward. Shell moderately inflated, with broad and shallow sulcus running from the umbo to the ventral margin; escutcheon lanceolate, deep, bordered by a sharp ridge; largest specimen ca. 74 mm long. Entire inner shell surface covered by fine radial grooves; ventral margin of inner shell surface crenulated; broad groove running from umbo to anterodorsal margin, bordering the anterior adductor muscle scar, which is indistinct, situated mostly below the midline of the shell, roundish to quadrate, and fused dorsally with the anterior pedal retractor scar; pallial line indistinct, onset apparently at the posteroventral margin of the anterior adductor muscle scar, running close to the ventral shell margin, no pallial sinus; posterior adductor muscle scar indistinct, roundish, situated in the posterodorsal corner of the shell, above a broad and shallow groove that radiates from about the middle part of the shell (anterior half of escutcheon) toward the posterior margin. Hinge plate narrow and short, with two blunt teeth in each valve; ligament nymph short and strong holding an external and calcified ligament
Remarks.—The “Nuculanid” from the early Norian, Late Triassic Greylock Butte seep deposits in Oregon, USA (Peckmann et al. 2011: fig. 3G) almost certainly also belongs to Aksumya. That species reaches 50 mm in length and is the most common mollusk (n = 15) at this Halorella-dominated seep deposit. It has a shorter and taller shell with a steeper and more angular anterior margin than Aksumya krystyni sp. nov.
Stratigraphic and geographic range.—Late Carnian to late early Norian (Late Triassic) seep deposits in the Kasımlar shales in southern Turkey.
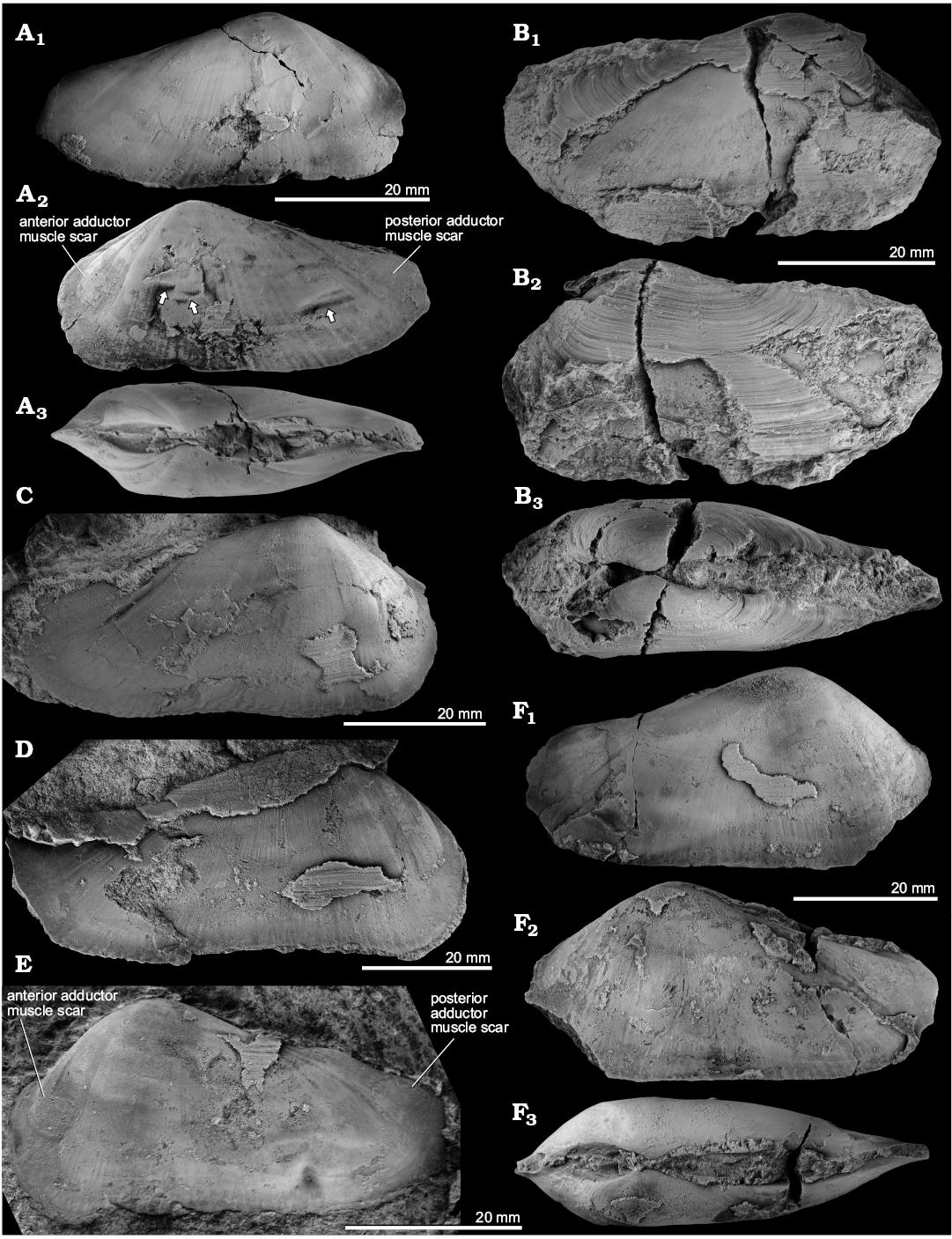
Fig. 7. The pholadomyoid bivalve Aksumya krystyni gen. et sp. nov. from the late Carnian, Late Triassic, Leo’s Terziler seep deposit in southern Turkey. A. Mo184066, paratype, internal mold of an articulated specimen with interior radial striations, right (A1) and left (A2) valves, note healed predation scars (arrows), dorsal view (A3). B. Mo184067, holotype, specimen with preserved external surface, showing the overall outline of the shell; right (B1) and left (B2) valves, dorsal view (B3). C. Mo184068, paratype, right valve with particularly strong ridge running from the middle of the dorsal margin to the anteroventral corner of the posterior adductor muscle scar. D. Mo184069, paratype, right valve of a large specimen showing the projection on the anterodorsal side of the anterior adductor muscle scar. E. Mo184070, paratype, left valve, and predation marks on the posteroventral side. F. Mo184071, paratype, internal mold of articulated specimen, right (F1) and left (F2) valves, dorsal view (F3).

Fig. 8. The pholadomyoid bivalve Aksumya krystyni gen. et sp. nov. from the late Carnian, Late Triassic Leo’s Terziler seep deposit in southern Turkey. A. Mo184067, holotype, close-up on the small shell spines. B. Mo184072, specimen with preserved ligament.

Fig. 9. The pholadomyoid bivalve Aksumya krystyni gen. et sp. nov. from the late Carnian, Late Triassic in southern Turkey; polished surfaces of cross sections showing hinge and nymph details. A. Mo184073 from Dumanlı I; cross section cut horizontally through the umbones, showing hinge (white arrows) and ligament nymph (black arrows). B. Mo184074 from Terziler III, vertical cross section showing hinge area just below umbo, seen from the anterior; white dotted line in B2 indicates the hinge area, black dotted line indicates the inferred shell material of the umbones. Abbreviations: LV, left valve; RV, right valve.
Discussion
The presence of two distinct and new modiomorphid genera at the Upper Triassic seep deposits in the Kasımlar shales in southern Turkey had already been indicated in the initial report of these sites (Kiel et al. 2017) and they are here described as Terzileria and Kasimlara. They significantly increase the diversity of modiomorphid bivalves at seeps and shorten the stratigraphic gap between the Silurian–Devonian Ataviaconcha (Jakubowicz et al. 2017; Hryniewicz et al. 2017b) and the Late Jurassic–Late Cretaceous Caspiconcha (Kelly et al. 2000; Kiel et al. 2010; Jenkins et al. 2013, 2018).
The phylogenetic relationships between members of the “modiomorphid” families Kalenteridae, Myoconchidae, and Modiomorphidae remain unclear (Hryniewicz et al. 2017b), and the distinctions between these groupings are indeed based on a rather limited set of characters (Chavan 1969). There are two general possibilities for the relationships among the seep-inhabiting genera Ataviaconcha, Terzileria, Kasimlara, and Caspiconcha. The first possibility, coined here the “single lineage hypothesis”, is that these four genera are members of a single lineage that has continuously inhabited hydrocarbon seeps from the Silurian to the Cretaceous. An analog from the more recent geologic record is the Bathymodiolinae, whose members have been living almost exclusively in deep-water reducing habitats such as hydrothermal vents, methane seeps, and large accumulations of organic matter since the Eocene (Kenk and Wilson 1985; Distel et al. 2000; Lorion et al. 2013; Kiel and Amano 2013). An interesting implication of this hypothesis is that this putative deep-water lineage would have survived the end-Permian mass extinction event, as did certain shallow-water representatives of the kalenterids (Damborenea 2004).
The second possibility, here coined the “repeated colonization hypothesis”, is that the four genera represent individual offshoots of one or several “modiomorphid” lineages that repeatedly developed similar morphological adaptations (cf. Hryniewicz et al. 2017b) to the methane-seep environments. This hypothesis raises interesting questions of its own: was there a single “source habitat” from these repeated colonization started, and why did this group that successfully colonized reducing habitats over the course of 350 million years, fail to colonize them again after the Cretaceous?
Chemosymbiosis is unknown from extant Anomalodesmata (Harper et al. 2006; Taylor and Glover 2010) and the alleged thraciid Nipponothracia from fossil seep deposits in Japan proved to be a lucinid (Kase et al. 2007; Kiel 2013). Recently, members of three genera of presumed anomalodesmatans (Anhembia, Tambaquyra, and Maackia) were reported from concretionary limestone deposits from the Permian of Brazil, which were interpreted as cold seep deposits (Matos et al. 2017). However, (i) the published field images and rock samples look unlike other, confirmed, seep deposits, (ii) the carbonates lack banded and botryoidal rim cements characteristic for seep carbonates (cf., Peckmann and Thiel 2004), and (iii) their carbon isotope signature is not very negative and is well within the range of septarian and other concretions resulting from the oxidation of organic matter (Irwin et al. 1977). Thus the interpretation of these “anomalodesmatans” as seep-inhabiting bivalves is not followed here. This makes Aksumya the first seep-inhabiting and probably seep-restricted genus of the Anomalodesmata.
Acknowledgments
I thank Jörn Peckmann (University of Hamburg, Germany), Leopold Krystyn (University of Vienna, Austria), and Erdal Koşun and Ferdi Demirtaş (both Akdeniz University, Antalya, Turkey) for their company and assistance with logistics and field work, Leopold Krystyn for making his earlier collections from Turkey available, and Liz Harper (University of Cambridge, UK) for insights to anomalodesmatan systematics. I also thank the reviewers Susana Damborenea (Museo de Ciencias Naturales, La Plata, Argentina), Krzysztof Hryniewicz (Institute of Paleobiology PAS, Warsaw, Poland), and Christopher McRoberts (State University of New York at Cortland, USA) for their insightful comments that helped to improve this manuscript. Financial support was provided by the FWF – Der Wissenschaftsfonts, through grant M1779-N29.
References
Agassiz, L. 1842–1845. Études critiques sur les Molluscques Fossiles. Monographie des Myes. 287 pp. H. Wolprath, Neuchâtel.
Ager, D.V., Gutnic, M., Juteau, T., and Monod, O. 1978. New early Mesozoic brachiopods from southern Turkey. Bulletin of the Mineral Research and Exploration Institute of Turkey 91: 59–75.
Amano, K. and Jenkins, R.G. 2011. Fossil record of extant vesicomyid species from Japan. Venus 69: 163–176.
Amano, K. and Kiel, S. 2007. Fossil vesicomyid bivalves from the North Pacific region. The Veliger 49: 270–293.
Broili, F. 1903. Die Fauna der Pachycardientuffe der Seiser Alp. Palaeontographica 50: 145–227.
Campbell, H.J. 1984. New records and taxa of Permian and Triassic fossils from New Caledonia and New Zealand. Alcheringa 8: 151–167. Crossref
Campbell, K.A. and Bottjer, D.J. 1995. Brachiopods and chemosymbiotic bivalves in Phanerozoic hydrothermal vent and cold seep environments. Geology 23: 321–324. Crossref
Chavan, A. 1969. Superfamily Carditacea Fleming, 1820. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology Part N, Mollusca 6, Bivalvia Volume 2, N543–N561. The Geological Society of America, Boulder and The University of Kansas, Lawrence.
Chen, Y., Krystyn, L., Orchard, M.J., Lai, X.-L., and Richoz, S. 2015. A review of the evolution, biogstratigraphy, provincialism and diversity of Middle and early Late Triassic conodonts. Papers in Palaeontology 2: 235–263. Crossref
Childress, J.J., Fisher, C.R., Brooks, J.M., Kennicutt, M.C.I., Bidigare, R.R., and Anderson, A.E. 1986. A methanotrophic marine molluscan (Bivalvia, Mytilidae) symbiosis: mussels fueled by gas. Science 233: 1306–1308. Crossref
Covacevich, V., Pérez, E., and Escobar, F. 1991. Presencia del género Kalentera Marwick, 1953 (Mollusca: Bivalvia) en el Sinemuriano al Sur de Taltal, Chile. Sexto Congreso Geológico Chileno (Viña del Mar), Actas 1: 68–71.
Dall, W.H. 1889. Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Gulf of Mexico (1877–1878) and in the Caribbean Sea (1879–1880), by the U.S. Coast Survey steamer “Blake” XXIX. Report on the Mollusca. Part 2, Gastropoda and Scaphopoda. Bulletin of the Museum of Comparative Zoology, Harvard University 18: 1–492.
Damborenea, S.E. 2004. Early Jurassic Kalentera (Bivalvia) from Argentina and its palaeobiologeographical significance. Ameghiniana 41: 185–198.
Distel, D.L., Baco, A.R., Chuang, E., Morrill, W., Cavanaugh, C.M., and Smith, C.R. 2000. Do mussels take wooden steps to deep-sea vents? Nature 403: 725–726. Crossref
Grant-Mackie, J.A. 1979. On a new Kalentera (Pelecypoda: Cypricardiacea) from the Upper Triassic of New Zealand. New Zealand Journal of Geology and Geophysics 3: 74–80. Crossref
Gray, J.E. 1847. A list of the genera of Recent Mollusca, their synonyma and types. Proceedings of the Zoological Record of London 15: 129–219.
Griffin, M. and Pastorino, G. 2006. Madrynomya bruneti n. gen. and sp. (Bivalvia: ?Modiomorphoidae): a Mesozoic survivor in the Tertiary of Patagonia? Journal of Paleontology 80: 272–282. Crossref
Gutnic, M., Monod, O., Poisson, A., and Dumont, J.F. 1979. Géologie des Taurides occidentales (Turquie). Mémoir de la Société géologique de France (nouveau series) 137: 1–109.
Harper, E.M., Dreyer, H., and Steiner, G. 2006. Reconstructing the Anomalodesmata (Mollusca: Bivalvia): morphology and molecules. Zoological Journal of the Linnean Society 148: 395–420. Crossref
Hautmann, M. 2001. Die Muschelfauna der Nayband-Formation (Obertrias, Nor-Rhät) des östlichen Zentraliran. Beringeria 29: 3–181.
Healey, M. 1908. The fauna of the Napeng-beds or the Rhaetic beds of upper Burma. Palaeontologia Indica, New Series II (4): 1–88.
Hryniewicz, K., Amano, K., Jenkins, R.G., and Kiel, S. 2017a. Thyasirid bivalves from Cretaceous and Paleogene cold seeps. Acta Palaeontologica Polonica 62: 705–728. Crossref
Hryniewicz, K., Jakubowicz, M., Belka, Z., Dopieralska, J., and Kaim, A. 2017b. New bivalves from a Middle Devonian methane seep in Morocco: the oldest record of repetitive shell morphologies among some seep bivalve molluscs. Journal of Systematic Palaeontology 15: 19–41. Crossref
Irwin, H., Curtis, C., and Coleman, M. 1977. Isotopic evidence for source of diagenetic carbonates formed during burial of organic-rich sediments. Nature 269: 209–213. Crossref
Jakubowicz, M., Hryniewicz, K., and Belka, Z. 2017. Mass occurrence of seep-specific bivalves in the oldest-known cold seep metazoan community. Scientific Reports 7: 14292. Crossref
Jenkins, R.G., Kaim, A., Hikida, Y., and Kiel, S. 2018. Four new species of the Jurassic to Cretaceous seep-restricted bivalve Caspiconcha and implications for the history of chemosynthetic communities. Journal of Paleontology [published online, doi: 10.1017/jpa.2018.7] Crossref
Jenkins, R.G., Kaim, A., Little, C.T.S., Iba, Y., Tanabe, K., and Campbell, K.A. 2013. Worldwide distribution of modiomorphid bivalve genus Caspiconcha in late Mesozoic hydrocarbon seeps. Acta Palaeontologica Polonica 58: 357–382.
Johnson, S.B., Krylova, E.M., Audzijonyte, A., Sahling, H., and Vrijenhoek, R.C. 2017. Phylogeny and origins of chemosynthetic vesicomyid clams. Systematics and Biodiversity 15: 346–360. Crossref
Johnson, S.B., Warén, A., Lee, R.W., Kano, Y., Kaim, A., Davis, A., Strong, E.E., and Vrijenhoek, R.C. 2010. Rubyspira, new genus and two new species of bone-eating deep-sea snails with ancient habits. Biological Bulletin 219: 166–177. Crossref
Kaim, A., Hryniewicz, K., Little, C.T.S., and Nakrem, H.A. 2017. Gastropods from the Late Jurassic–Early Cretaceous seep deposits in Spitsbergen, Svalbard. Zootaxa 4329: 351–374. Crossref
Kaim, A., Jenkins, R.G., and Warén, A. 2008. Provannid and provannid-like gastropods from Late Cretaceous cold seeps of Hokkaido (Japan) and the fossil record of the Provannidae (Gastropoda: Abyssochrysoidea). Zoological Journal of the Linnean Society 154: 421–436. Crossref
Kaim, A., Jenkins, R.G., Tanabe, K., and Kiel, S. 2014. Mollusks from late Mesozoic seep deposits, chiefly in California. Zootaxa 3861: 401–440. Crossref
Kase, T., Kurihara, Y., and Hagino, K. 2007. Middle Miocene chemosynthetic thraciid Nipponothracia gigantea (Shikama, 1968) from central Japan is a large lucinid bivalve (Lucinoidea; Mollusca). The Veliger 49: 294–302.
Kelly, S.R.A., Blanc, E., Price, S.P., and Withham, A.G. 2000. Early Cretaceous giant bivalves from seep-related limestone mounds, Wollaston Forland, Northeast Greenland. In: E.M. Harper, J.D. Taylor, and J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia. Geological Society of London, Special Publication 177: 227–246. Crossref
Kenk, V.C. and Wilson, B.R. 1985. A new mussel (Bivalvia: Mytilidae) from hydrothermal vents in the Galapagos Rift Zone. Malacologia 26: 253–271.
Kiel, S. 2013. Lucinid bivalves from ancient methane seeps. Journal of Molluscan Studies 79: 346–363. Crossref
Kiel, S. 2015. Did shifting seawater sulfate concentrations drive the evolution of deep-sea vent and seep ecosystems? Proceedings of the Royal Society B 282: 20142908. Crossref
Kiel, S. 2016. A biogeographic network reveals evolutionary links between deep sea hydrothermal vent and methane seep faunas. Proceedings of the Royal Society B 283: 20162337. Crossref
Kiel, S. and Amano, K. 2013. The earliest bathymodiolin mussels: Evaluation of Eocene and Oligocene taxa from deep-sea methane seep deposits in western Washington State, USA. Journal of Paleontology 87: 589–602. Crossref
Kiel, S. and Little, C.T.S. 2006. Cold seep mollusks are older than the general marine mollusk fauna. Science 313: 1429–1431. Crossref
Kiel, S. and Taviani, M. 2017. Chemosymbiotic bivalves from Miocene methane-seep carbonates in Italy. Journal of Paleontology 91: 444–466. Crossref
Kiel, S., Campbell, K.A., and Gaillard, C. 2010. New and little known mollusks from ancient chemosynthetic environments. Zootaxa 2390: 26–48.
Kiel, S., Krystyn, L., Demirtaş, F., Koşun, E., and Peckmann, J. 2017. Late Triassic mollusk-dominated hydrocarbon-seep deposits from Turkey. Geology 45: 751–754. Crossref
King, W. 1850. A monograph of the Permian fossils of England. Monograph of the Palaeontographical Society 3: xxxvii + 258. Crossref
Koninck, L.G. de 1885. Faune du calcaire carbonifère de la Belgique. 5e partie: Lamellibranches. Annales du Musée royal d’Histoire naturelle de Belgique, Série Paléontologique 11: 1–283.
Leanza, A.F. 1940. Myoconcha neuquena n. sp. del Lias de Piedra Pintada en el Neuquén. Notas del Museo de La Plata, Paleontología 5: 123–131.
Likharev, B.K. 1925. Zur Frage über das Alter der Perm-Kalksteine der Onega-Dwina Wasserscheide. Mémoires de la Société Russe de Minéralogie 54: 109–152.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis,locis. Vol. 1. Editio decima. 824 pp. Laurentius Salvius, Holmiae.
Little, C.T.S. and Vrijenhoek, R.C. 2003. Are hydrothermal vent animals living fossils? Trends in Ecology and Evolution 18: 582–588. Crossref
De Loriol, P. and Cotteau, G. 1868 Monographie paléontologique et géologique de l’étage portlandien du département de l’Yonne. Bulletin de la Société des Sciences historiques et naturelles le l’Yonne 1: 1–260.
Lorion, J., Kiel, S., Faure, B.M., Masaru, K., Ho, S.Y.W., Marshall, B.A., Tsuchida, S., Miyazaki, J.-I., and Fujiwara, Y. 2013. Adaptive radiation of chemosymbiotic deep-sea mussels. Proceedings of the Royal Society B 280: 20131243. Crossref
Lukeneder, S., Lukeneder, A., Harzhauser, M., İslamoğlu, Y., Krystyn, L., and Lein, R. 2012. A delayed carbonate factory breakdown during the Tethyan-wide Carnian Pluvial Episode along the Cimmerian terranes (Taurus, Turkey). Facies 58: 279–296. Crossref
Marwick, J. 1953. Divisions and faunas of the Hokonui System (Triassic and Jurassic). New Zealand Geological Survey Palaeontological Bulletin 21: 1–141.
Matos, S.A., Warren, L.V., Fürsich, F.T., Alessandretti, L., Assine, M.L., Riccomini, C., and Simoes, M.G. 2017. Paleoecology and paleoenvironments of Permian bivalves of the Serra Alta Formation, Brazil: Ordinary suspension feeders or Late Paleozoic Gondwana seep organisms? Journal of South American Earth Sciences 77: 21–41. Crossref
Morris, N.J. and Dickins, J.M. 1991. Upper Palaeozoic anomalodesmatan Bivalvia. Bulletin of the British Museum (Natural History) Geology 47: 51–100.
Murchison, R.I., Keyserling, A.F.M.L.N.A., Graf von, and de Verneuil, E. 1845. Géologie de la Russie d’Europe et des montagnes de l’Oural. 511 pp. J. Murray, London.
Newell, N.D. 1957. Notes on certain primitive heterodont pelecypods. American Museum Novitates 1857: 1–14.
Newell, N.D. 1969. Superfamily Pholadomyacea Gray, 1847. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology Part N, Mollusca 6, Bivalvia Vol. 2, N818–N843. The Geological Society of America and The University of Kansas, Lawrence.
Peckmann, J. and Thiel, V. 2004. Carbon cycling at ancient methane-seeps. Chemical Geology 205: 443–467. Crossref
Peckmann, J., Kiel, S., Sandy, M.R., Taylor, D.G., and Goedert, J.L. 2011. Mass occurrences of the brachiopod Halorella in Late Triassic methane-seep deposits, Eastern Oregon. Journal of Geology 119: 207–220. Crossref
Peek, A.S., Feldman, R.A., Lutz, R.A., and Vrijenhoek, R.C. 1998. Cospeciation of chemoautotrophic bacteria and deep sea clams. Proceedings of the National Academy of Sciences of the USA 95: 9962–9966. Crossref
Roemer, F.A. 1836–1839. Die Versteinerungen des norddeutschen Oolithen-Gebirges. 218 pp. Hahn, Hannover.
Saether, K.P., Little, C.T.S., Campbell, K.A., Marshall, B.A., Collins, M., and Alfaro, A.C. 2010. New fossil mussels (Bivalvia: Mytilidae) from Miocene hydrocarbon seep deposits, North Island, New Zealand, with general remarks on vent and seep mussels. Zootaxa 2577: 1–45.
Sibuet, M. and Olu, K. 1998. Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins. Deep-Sea Research II 45: 517–567. Crossref
Sowerby, J. de C. 1824. The Mineral Conchology of Great Britain 5 (81): 99–114. Sherwood and Co., London.
Sowerby, J. de C 1826. The Mineral Conchology of Great Britain 6 (87): 1–12. Sherwood and Co., London.
Squires, R.L. and Goedert, J.L. 1991. New Late Eocene mollusks from localized limestone deposits formed by subduction-related methane seeps, southwestern Washington. Journal of Paleontology 65: 412–416. Crossref
Taylor, J.D. and Glover, E.A. 2010. Chemosymbiotic bivalves. In: S. Kiel (ed.), The Vent and Seep Biota, 107–136. Springer, Heidelberg. Crossref
Taylor, J.D., Williams, S.T., Glover, E.A., and Dyal, P. 2007. A molecular phylogeny of heterodont bivalves (Mollusca: Bivalvia: Heterodonta): new analyses of 18S and 28S rRNA genes. Zoologica Scripta 36: 587–606. Crossref
Tekin, U.K. 1999. Biostratigraphy and systematics of late Middle to Late Triassic radiolarians from the Taurus Mountains and Ankara region, Turkey. Geologisch-Paläontologische Mitteilungen Innsbruck (Special Issue) 5: 1–296.
Trechmann, C.T. 1917. The Trias of New Zealand. Quarterly Journal of the Geological Society of London 73: 165–246. Crossref
Tunnicliffe, V. 1992. The nature and origin of the modern hydrothermal vent fauna. Palaios 7: 338–350. Crossref
Tunnicliffe, V., Fowler, M.R., and McArthur, A.G. 1996. Plate tectonic history and hot vent biogeography. In: J.C. MacLeod, P.A. Tyler, and C.L. Walker (eds.), Tectonic, Magmatic, Hydrothermal and Biological Segmentation of Mid-Oceanic Ridges. Geological Society, Special Publication 118: 225–238. Crossref
Van Dover, C.L. 2000. The Ecology of Deep-Sea Hydrothermal Vents. 424 pp. Princeton University Press, Princeton.
de Verneuil, E. and Murchison, R.I. 1844. Note sur les équivalents du système permien en Europe, suivie d’un coup d’œil général sur l’ensemble de ses fossiles, et d’un tableau des espèces. Bulletin de la Société géologique de France, Série 2 1: 475–517.
Vrijenhoek, R.C. 2013. On the instability and evolutionary age of deep-sea chemosynthetic communities. Deep-Sea Research II 92: 189–200. Crossref
Acta Palaeontol. Pol. 63 (2): 221–234, 2018
https://doi.org/10.4202/app.00466.2018