
The dentition of a eugeneodontiform shark from the Lower Pennsylvanian of Derbyshire, UK
MICHAŁ GINTER
Ginter, M. 2018. The dentition of a eugeneodontiform shark from the Lower Pennsylvanian of Derbyshire, UK. Acta Palaeontologica Polonica 63 (4): 725–735.
The anterior part of a medium-sized shark surrounded by hundreds of ostracod shells was found at the end of the last century in a dark limestone nodule from the Kinderscoutian (Bashkirian, Pennsylvanian) near Carsington, Derbyshire (England, UK). The shark is a caseodontoid eugeneodontiform, most probably belonging to Campodus agassizianus. Its dentition is of the crushing type, highly heterodont, but the symphyseal tooth whorl, typical of the most of eugeneodontiforms, was not found. The teeth are symmetrical labio-lingually which is unique in this group. The analysis of the available data leads to the conclusion that neither the tooth whorls of “Campodus variabilis” sensu Eastman, nor the mandibular dentition of “Agassizodus variabilis” sensu St. John and Worthen represent the genus Campodus and that these specimens deserve a new, probably common, name.
Key words: Chondrichthyes, Eugeneodontiformes, Campodus, teeth, Carboniferous, Bashkirian, England.
Michał Ginter [m.ginter@uw.edu.pl], Faculty of Geology, University of Warsaw, Al. Żwirki i Wigury 93, 02-089 Warszawa, Poland.
Received 30 July 2018, accepted 20 September 2018, available online 26 November 2018.
Copyright © 2018 M. Ginter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
By the end of the last century, a dark limestone nodule from the Kinderscoutian (Lower Pennsylvanian) of Derbyshire was cracked open by Nicholas Riley from the British Geological Survey, exposing dispersed remains of a medium-sized shark, surrounded by hundreds of ostracod shells. From the beginning it was evident that the ostracods must have been scavenging on the dead body of the shark. The whole specimen and the ostracods were subsequently described in two papers: Wilkinson et al. (2004) and Wilby et al. (2006), and in the latter paper an interesting taphonomic model for the shark-ostracod association was proposed. However, the fossils of the shark have not been hitherto properly identified and described. The only available taxonomic information (based on a personal communication from Peter Forey provided in 2001) was that the shark represented the euchondrocephalan genus Orodus (Wilby et al. 2006: 311).
I had a brief opportunity to study and photograph that material already in 2003, but at that time my knowledge of euchondrocephalan teeth was still too rudimentary to confirm or refute the proposed identification. Following a second examination of the specimen in January 2018, I am convinced that the shark is not Orodus, despite the orodont general outline of its teeth, but a caseodontoid eugeneodontiform. It is also very likely that it belongs to the same species as the partly articulated specimen from the Chokierian (Upper Mississippian) of Belgium described by Lohest (1884) under the name of Campodus agassizianus De Koninck, 1844, and a tooth illustrated from the Serpukhovian of the vicinities of Tula, Russia, by Romanovsky (1864) as Orodus elegans. The main aim of this paper is to describe in detail and illustrate the remnants of the shark which was scavenged by ostracods, and particularly its interesting dentition.
In addition to the description of the shark from Derbyshire, there is a serious nomenclatural problem surrounding its identity. Lohest’s (1884) identification of his specimen as Campodus agassizianus is at least disputable. This question will be presented in the last part of the paper, based on the recent examination of a half of the type specimen of C. agassizianus housed at The Natural History Museum in London, UK. Also, I hope that the analysis of the new specimen from Derbyshire will shed new light on the 150-years old discussion on the validity of the genus Agassizodus St. John and Worthen, 1875, and the relation between the latter genus and Campodus.
Institutional abbreviations.—GSM, British Geological Survey, Keyworth, England, UK; NHMUK, The Natural History Museum, London, England, UK; UNSM, University of Nebraska State Museum, Lincoln, Nebraska, USA.
Geological setting and taphonomy
The detailed information concerning the locality and its geology was provided in Wilkinson et al. (2004) and Wilby et al. (2006). Here I present a summary of their data. The Lower Hays Farm borrow pit (now flooded) (SK 2478 5033 to 2496 5042) near Carsington, Derbyshire (Fig. 1), formerly provided one of the best sections in central England of the generally poorly exposed Bowland Shale Formation (Bashkirian, Pennsylvanian). In 1986 the pit exposed about 87 m of strata extending from near the base of the Chokierian (H1a) stage to near the top of the Kinderscoutian (R1c) stage. The sequence consists largely of unfossiliferous, grey, calcareous mudstone and silty mudstone, but includes numerous discrete, organic-rich, dark grey to black, fissile mudstone, or so-called marine-bands. Many of these contain large carbonate concretions and yield fully marine faunas dominated by goniatites and bivalves.
The material forming the subject of this paper is derived from a large, discoidal, dark grey, fine-grained concretion that was collected loose from the borrow pit in 1986. The precise marine band from which it came is not known but, based on the presence of the goniatite Reticuloceras, it has been assigned to the Kinderscoutian (R1a–c) part of the sequence. Since all of the exposed R1c marine bands appear to lack this type of concretion, it is assumed to have been derived from either an R1a or R1b marine band.
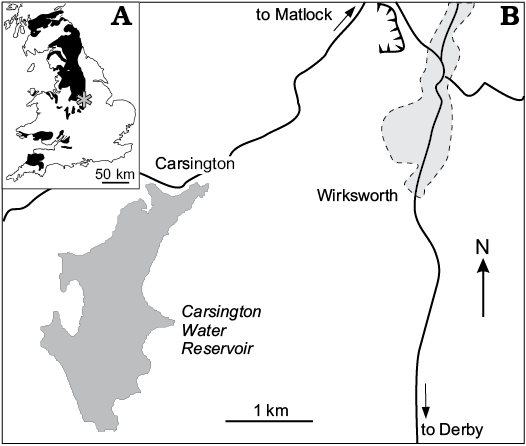
Fig. 1. A. Sketch map of Carboniferous outcrops (black) in Britain with the position of study area in Derbyshire (asterisk); after Dineley and Metcalf (1999). B. The position of the studied outcrop, currently submerged by the Carsington Water Reservoir.
The concretion contains an unusual association between a moderately-sized shark and a large number of individuals of the myodocopid ostracod Eocypridina carsingtonensis Wilkinson, Williams, Siveter, and Wilby, 2004. Only the anterior end of the shark is present, the rest presumably lay in the sediment beyond the margins of the concretion and was not recovered. It is preserved parallel to the bedding and, based on its degree of articulation, does not appear to have become buoyant post-mortem or to have been excessively disturbed by macro-scavengers. Its teeth remain closely associated in the centre of the concretion and indicate that the specimen may have been originally approximately 2 m long. Posterior to the teeth are numerous fragments of cartilage (up to 30–70 mm), some of which have become displaced to positions just above the majority of the rest of the fossil.
Wilby et al. (2006) proposed the following sequence of events after the death of the shark (Fig. 2): (i) The carcass arrived at the bottom and sank part-way into the soft sediment. (ii) The shark’s upper surface protruded above the sediment and formed a short-lived necrobenthic island, attracting the ostracods. (iii) A decay-induced collapse of the carcass resulted in anoxic sediment caving-in from the walls of the shark and large numbers of the firmly anchored ostracods being smothered and asphyxiated. (iv) Further releases of putrefaction gases led to the partial disassociation of many of the ostracod valves and the shark skeleton. (v) Minor sedimentary compaction caused the breakage of numerous valves and was halted by the development of the concretion centred on the shark.
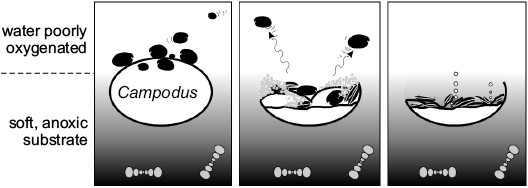
Fig. 2. Schematic taphonomic model for the shark-ostracod association illustrating the key stages in its development; after Wilby et al. (2006).
Material and methods
Currently the nodule is broken in many pieces, each bearing a separate collection number. Most of the pieces contain a mess of broken shark teeth and/or fragments of calcified cartilage, ostracods, bivalves, and goniatite shells (Fig. 3). The teeth are in various positions and states of preservation. Sometimes large parts of the crowns can be observed, with the minutest details of surface ornamentation; in the other cases internal microstructure of the coronal or basal tissues is displayed. Usually the crowns of larger teeth are intensely white and shiny, and therefore clearly visible on the dark background; the spongey bases and basal internal parts of the crowns are brown.

Fig. 3. A part of the dark limestone nodule (GSM 105524) from the Kinderscoutian of Derbyshire, UK, showing the diversity of fossils.
It seems that many teeth were broken before the fossilisation, but the greatest damage was done to them during the fragmentation of the nodule. No chemical preparation was applied to the fossils.
Two series of digital photographs of the material have been made, one in 2003 and another in 2018. In both cases the same camera, a Nikon Coolpix 4500, was used. During the first, informal session, the measurements were not made and the collection numbers have not been noted. This was corrected in 2018, but one tooth photographed in 2003 has not been located amongst the material. Because of its importance, it is figured here (Fig. 6E) with no number and scale bar.
Systematic palaeontology
Class Chondrichthyes Huxley, 1880
Subclass Euchondrocephali Lund and Grogan, 1997
Order Eugeneodontiformes Zangerl, 1981
Superfamily Caseodontoidea Zangerl, 1981
Family indet.
Genus Campodus De Koninck, 1844
Type species: Campodus agassizianus De Koninck, 1844; Carboniferous, Pennsylvanian, Chokierian; Belgium, near Liège.
Campodus agassizianus De Koninck, 1844
Figs. 3–8.
1844 Campodus agassizianus gen. et sp. nov.; De Koninck 1844: 617–618, pl. 55: 1.
1864 Orodus elegans sp. nov.; Romanovsky 1864: 157–158, pl. 3: 1.
1884 Campodus agassizianus De Koninck, 1844; Lohest 1884: 295–305, pl. 3: 1–3, pl. 4: 1, 4–6.
2004 Orodus sp.; Wilkinson et al. 2004: pl. 2.
2006 Orodus sp.; Wilby et al. 2006: 311, fig. 2.
Emended diagnosis.—A eugeneodontiform with a crushing, heterodont dentition in which the largest and most characteristic teeth display the following combination of features. The teeth are elongated mesio-distally. The crown, built entirely of tubular dentine, is symmetrical labio-lingually and slightly asymmetrical mesio-distally. The median part is elevated and the median cusp is broad, pyramidal, with a rounded tip. There are three similar cusps on each side of the median cusp. All the cusps are connected with a median crest and bear strong transverse ridges. The median crest and the transverse ridges are ornamented with secondary ridges, transverse or oblique to the major forms. All the ridges are serrated. The base is of euselachian type. The smallest teeth, probably from the anterior and posterior parts of the jaw, are bar-like, with no elevation in the crown. Numerous minute transverse ridges with serrated edges are situated symmetrically on labial and lingual sides.
This diagnosis is based almost exclusively on the most characteristic teeth, probably situated in the middle of the jaw ramus, because only such teeth are really diagnostic. Teeth of this type are present in three of the reference materials: Romanovsky (1864), Lohest (1884) (Figs. 4, 7A–C), and the collection from Derbyshire described herein (Figs. 3, 5, 6), but unfortunately do not occur in the holotype (Figs. 7D, 8), of which only the small, bar-like teeth are known (see the discussion below).
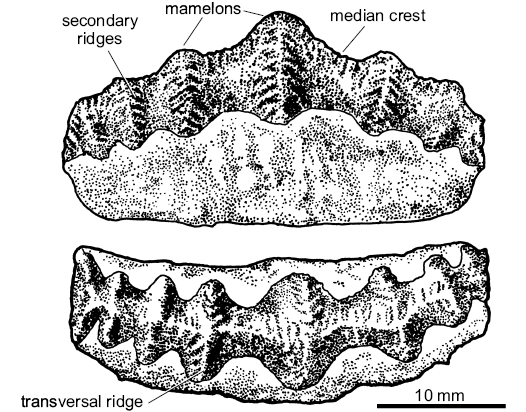
Fig. 4. Morphological terminology of the teeth of Campodus agassizianus De Koninck, 1844. The drawing based on Lohest’s (1884) Belgian material re-illustrated in Ginter et al. (2010).

Fig. 5. The largest and best preserved tooth (GSM 105459) of the eugeneodontiform shark Campodus agassizianus De Koninck, 1844 from the Kinderscoutian of Derbyshire, UK. Note the tubular dentine in the broken median cusp (black arrow) and a probable bar-like tooth at the bottom of the photograph (white arrow).

Fig. 6. Teeth of the eugeneodontiform shark Campodus agassizianus De Koninck, 1844, in the fragments of nodule from the Kinderscoutian of Derbyshire, UK. A. A half of horizontal section of a large tooth (GSM 105471a). B. Lingual view of a half of a medium-sized tooth with well preserved surface (GSM 105471b). C. Close-up of long teeth (GSM 105524). D. Two cusps (mamelons) of a large tooth with abraded surface and uncovered tubular dentine canals; at the bottom part (arrows): aboral views of bases of two teeth (GSM 105471c). E. A half of a section of a large tooth (specimen not located in the collection during the second investigation). F. The upper part of a crown with a relatively high median cusp (GSM 105471d). Scale bars 5 mm apart from E where it is unknown.
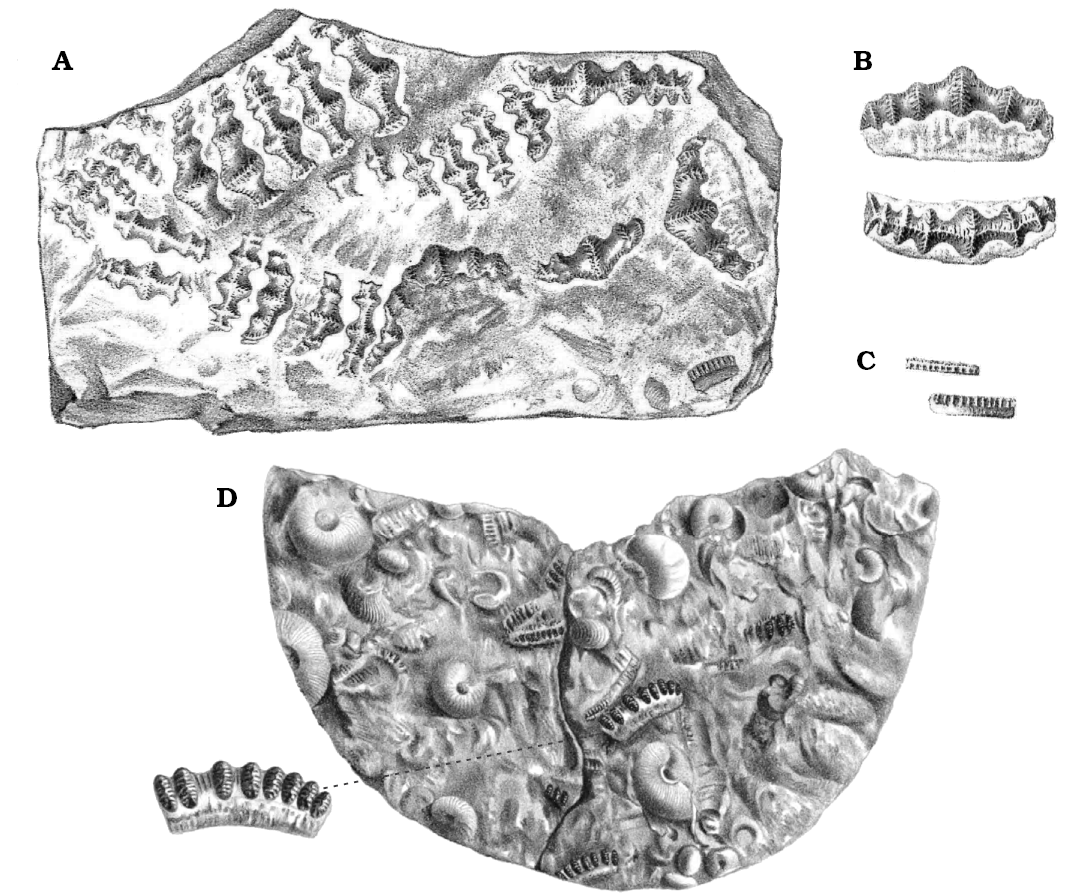
Fig. 7. Teeth of Campodus agassizianus De Koninck, 1844, from the Chokierian of Belgium, near Liège. A–C. Specimens whereabouts unknown, from Lohest (1884). D. Holotype (left part NHMUK P.28754, see Fig. 8 herein; right part whereabouts uncertain), from De Koninck (1844). Not to scale.

Fig. 8. Left part of the holotype, NHMUK P.28754 (right part, see Fig. 7D, whereabouts uncertain, probably in France) of the eugeneodontiform shark Campodus agassizianus De Koninck, 1844, from the Chokierian of Belgium, near Liège. A. General view. B. Close-up of a group of teeth.
Description.—The following text is based mostly on the characteristics of the fragments of the anterior part of a shark dispersed in a limestone nodule (GSM 105458–105522) from Lower Hays Farm borrow pit (now flooded) near Carsington, Derbyshire, England, UK (Kinderscoutian, Bashkirian, Pennsylvanian), and the description provided by Lohest (1884) of his specimen from the Chokierian of Belgium. The whereabouts of the latter specimen are unknown; a cast of it is allegedly present at the University of Liège (Valentin Fischer, personal communication 2018), but the description and drawings are so good that they are sufficient for comparison. The word “mamelon” adopted here from Lohest’s text means in this case a broad cusp.
General features: The teeth of Campodus agassizianus are elongated mesio-distally, straight or gently curved. The height of the crown is approximately equal to that of the root; the middle region, in general, is more elevated than the extremities. The teeth are formed of five to twelve conical mamelons, slightly elongated transversally to the length of a tooth and connected with each other by a concave area. In the middle of a tooth the mamelons are larger and the spaces between them are wider than in the lateral areas.
Along the tooth crown there runs a high median crest. The crest is ornamented by low, transverse or oblique ridges which may be delicately serrated. From time to time such transverse ridges are larger and in that case they form median crests of the mamelons. These crests are also ornamented by small, sharp ridges, similar to those ornamenting the main, mesio-distal crest. Therefore, there are four orders of ornamentation: the median crest of a tooth (mesio-distal) with its own low, serrated ridges; the median crests of mamelons (labio-lingual); the ridges ornamenting the mamelons’ crests; the serrations on the latter ridges.
The base (root) is of a classic euselachian type sensu Ginter et al. (2010). There are numerous openings of basal vascular canals in the orolingual and aboral-labial areas and a flat surface devoid of foramina in the aboral-lingual part (Fig. 6D, arrows). Due to the overlapping of bases, when the teeth are in place in a tooth-family, these flat surfaces form together a continuous aboral surface with no canal openings. The tooth families observed in the Belgian material consist of a revolver of at least five teeth.
The dentition is characterised by a strong monognathic heterodonty. The largest teeth are generally symmetrical with the median part highly elevated (Figs. 4, 5, 6A, E). The smaller the tooth, the lower the elevation of the crown. There also occur very small, bar-like teeth with no median elevation at all in which the mamelons are replaced by numerous transverse (labio-lingual), serrated ridges (Figs. 7C, D, 8).
In the concretion from Derbyshire there occur extremely long teeth (Figs. 3, 6C), absent from the Belgian material. They apparently have no more than three mamelons in the median part and the rest is reduced to a long median crest with labial and lingual transverse ridges. Due to the fragmentary character of the material it is difficult to say whether such teeth are symmetrical or not. Usually only one lateral part of a tooth is completely preserved and this can measure up to 20 mm in its mesio-distal dimension.
The symphyseal tooth whorl, typical of the eugeneodontiforms, has not been found in any of the studied materials.
Microstructure: The crowns of the teeth are composed entirely of tubular dentine (orthotrabeculine sensu Zangerl et al. 1993), a tissue typical of all the Euchondrocephali, and which was highly resistant to compressive stress. In Mesozoic and modern holocephalans it forms the tritors on the tooth-plates, in petalodonts it covers the biting parts of the teeth with a relatively thin layer, but in orodonts, Palaeozoic holocephalans and eugeneodontiforms it is the major component of the crown. Here, in Campodus, the tubules, surrounded by hypermineralised orthodentine, run from the base-crown interface upwards, almost to the surface of the crown (Figs. 5, 6E; Lohest 1884: pl. 4: 4–6; compare Ginter et al. 2010: fig. 10D and Stahl 1999: fig. 20). However, their upper ends are closed by a thin layer of compact dentine, so the crown surface is smooth and shiny (Figs. 5, 6B, F). Lohest (1884: 297–298), in the section dedicated to the tooth microstructure of his Belgian specimen, noted that the surface is somewhat rough, shagreen-like, due to the nodes formed by the ends of the tubules. However, although I can confirm this observation on a few surfaces of the teeth from Derbyshire, I suppose that this is the result of initial abrasion (compare Stahl 1999: fig. 21.2). Stronger abrasion leads to uncovering of the tubules and a perforated surface (Fig. 6D).
The base is built of trabecular dentine, with a spongey network of canals. Because the preservation of the bases is, in most cases, much worse than that of the crowns, I could not trace any special system of basal canals. Lohest (1884) did not analyse the microstructure of the base.
Remarks.—The diversity of teeth in the specimens from Derbyshire and Belgium corresponds to the heterodonty observed on a jaw of “Agassizodus variabilis” from Osage, Kansas (St. John and Worthen 1875: pl. 8: 1; for the validity of this name, see the Discussion below). In that splendid specimen, the bar-like, minute teeth occur in the anterior and posterior parts of the jaw and the largest teeth with the most elevated median part in the middle region of the jaw (Fig. 9). The teeth of intermediate size and shape occupy the intermediate positions. The only teeth from Derbyshire which have no equivalents in the specimen from Osage are those which are extremely long, with reduced mamelons (Fig. 6C). They are also absent from the Lohest’s Belgian specimen. However, they are very similar to an isolated tooth also referred to by St. John and Worthen (1875: pl. 8: 2) as “A. variabilis”, but from the Mills County, Iowa. There was no firm ground for the latter authors’ decision to place the bunch of loose teeth from Iowa (St. John and Worthen 1875: pl. 8: 2–21) in the same species as the jaw from Kansas, but their intuition that they represent upper and lower jaws, respectively, might have been correct.
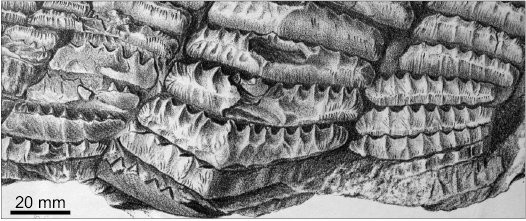
Fig. 9. Fragment of the mandibular dentition of “Agassizodus variabilis” from the Coal Measures (Pennsylvanian) of Osage County, Kansas, USA; from St. John and Worthen (1875: pl. 8: 1).
The potential similarity of the dentition model between “Agassizodus variabilis” and Campodus agassizianus does not mean that the individual teeth of these two species are identical. Indeed, this is not the case: the teeth of Campodus are virtually symmetrical across the mesio-distal midline (see particularly Figs. 6A, E, 7A) while those of “Agassizodus” have strong transverse ridges on the labial side of the crown and the lingual side is almost flat. In the words of St. John and Worthen (1875: 315) “[The teeth of Agassizodus are] distinguished by the prevailing prominence of the buttressed condition of the anterior [= labial] coronal borders, and the relative uniformity or evenness of the posterior [= lingual] face.” This difference between the labial and lingual side is typical of all later Pennsylvanian to Triassic eugeneodontiforms, whereas the Orodus-like labio-lingual symmetry of the teeth of Campodus situates this genus at the basal position of the group (see the Discussion below).
The single tooth of Campodus illustrated by Romanovsky (1864: pl. 3: 1) as Orodus elegans probably represents the middle part of a jaw ramus. It is mesio-distally symmetrical, relatively short, preserving the elevated middle part and only three mamelons on each side. Unfortunately the Romanovsky collection has been lost (Alexander Ivanov and Oleg Lebedev, personal communication 2018) and only the illustration is available.
Stratigraphic and geographic range.—Serpukhovian–Bashkirian boundary beds, Carboniferous. Serpukhovian, Mississippian, Moscow Syneclise, Russia (Romanovsky 1864); Chokierian, Bashkirian, Pennsylvanian, Liège, Belgium (De Koninck 1844; Lohest 1884); Kinderscoutian, Bashkirian, Pennsylvanian, Derbyshire, England, UK (this paper).
Discussion
The name of the shark from Derbyshire.—Judging from the available material and the literature, there is no doubt that the shark from the Kinderscoutian of Derbyshire represents the same species as the tooth described as Orodus elegans by Romanovsky (1864) and the partial dentition described as Campodus agassizianus by Lohest (1884). Because the name Campodus agassizianus was originally given to a group of teeth from the Chokierian of Belgium by De Koninck in 1844, this name has the priority. However, there are serious doubts as to whether the specimens described by De Koninck (1844) and Lohest (1884) should be treated as conspecific. The 20th-century authors writing about Campodus have either not touched upon the problem and in their comparisons used Lohest’s (1884), much more complete specimen as if it was the type (Eastman 1902a, b, 1903) or have entirely agreed with Lohest (1884) and De Koninck (1844) (who authorised Lohest’s 1884 work) in their identification of Lohest’s specimen (Eaton 1962). Zangerl in his volume of the Handbook of Paleoichthyology (1981: 77) expressed certain doubts, but finally also agreed with that view, and Hampe in Ginter et al. (2010) followed his reasoning.
At the beginning of my study I also have adopted this way of thinking, until my personal observation of the half of De Koninck’s (1844) holotype of C. agassizianus housed at The Natural History Museum in London (NHMUK 28754; Figs. 7D, 8; the other, more interesting part is probably in France). I realised that all the teeth illustrated by De Koninck (1844) are those bar-like, minute elements from the anterior or posterior part of a jaw (by comparison with “Agassizodus” from Osage). Such teeth I consider highly non-diagnostic, especially when only one side can be seen, and this is the case as far as De Koninck’s (1844) specimen is concerned. Bar-like teeth of “Agassizodus variabilis” (St. John and Worthen 1875: pl. 8: 1) or Caseodus eatoni (Zangerl 1981: fig. 86A, B; Ginter et al. 2010: fig. 109) are very similar and probably only a careful examination from both sides can distinguish them.
Thus, I was determined to restrict the name Campodus agassizianus to the specimen described by De Koninck (1844) and translate the specimen described by Lohest (1884), together with the material from Derbyshire and Orodus elegans Romanovsky, 1864, to a new genus. However, I realised that such a taxonomic decision would probably cause more harm than good and would blur the discussion on the dentition of early eugeneodontiforms instead of leading to reasonable solutions. Moreover, De Koninck and Lohest might have had some reasons unknown to me (the proximity of the findings?) to believe that their specimens were conspecific. Therefore, I finally decided to rely on their authority and refer all the mentioned materials to as C. agassizianus.
The Campodus–Agassizodus problem.—If we accept that the shark from Derbyshire really belongs to Campodus, we enter the complicated discussion concerning the relationships between Campodus and Agassizodus and the identity of various dental elements historically (and in most cases incorrectly) referred to as Campodus. The most important specimens allegedly belonging to Campodus are as follows:
– two incomplete teeth from the Meramecian (Viséan) Salem Formation of Missouri illustrated by Zangerl (1981: fig. 87) as Campodus sp.;
– a spectacular symphyseal tooth whorl (UNSM 1000), referred to as Campodus variabilis by Eastman (1902a: pl. 8: 1; 1902b: pl. 1; 1903: pl. 1, top), the cast of which is figured here (Fig. 10), from the Pennsylvanian (Coal Measures) of Cedar Creek, Nebraska; the original is in the collections of University of Nebraska State Museum, Lincoln, Nebraska, USA, specimen number UNSM 1000 (Wayne Itano, personal communication 2018);
– a less perfectly preserved symphyseal tooth whorl, also referred to as C. variabilis by Eastman (1902b: pl. 2, pl. 3: 1), with a few attached lateral tooth families, from the Pennsylvanian (Coal Measures) of Osage County, Kansas.
The first two teeth resemble Campodus by its general shape, but they are partly damaged and the ornamentation of the crown is apparently partly abraded. From the drawings provided by Zangerl (1981) it is difficult to judge to what extent the ornamentation was similar to that of C. agassizianus from Derbyshire. I suppose that originally it was not much richer than it is now and particularly the secondary ridges, branching from the transverse ridges of the mamelons, were absent from the beginning. The teeth do, indeed, look like intermediate forms between orodonts and eugeneodontiforms, and their low stratigraphic position (Viséan) confirms that. I agree with Zangerl (1981) that the teeth represent the genus Campodus, but probably not C. agassizianus.
Both symphyseal tooth whorls, very similar to each other and most probably conspecific, illustrated by Eastman (1902a, b, 1903) are composed of several roof-shaped, separate teeth, with the strong transverse ridges only on the labial side of the crown. This makes them look somewhat similar to the largest teeth of St. John and Worthen’s (1875) “Agassizodus”, but not to the teeth of Campodus sensu stricto, because the latter have equal ridges on both sides. Ignoring this difference and supposing that the tooth whorls and the mandible of Agassizodus variabilis (sensu St. John and Worthen 1875: pl. 8: 1) represent the same species, Eastman (1902a) created a new taxonomic combination, viz. Campodus variabilis (see particularly the attempted reconstruction of the whole dentition in Eastman 1903: pl. 1). From that date the name Campodus in connection with the spectacular tooth whorls migrated into the textbooks and labels accompanying museum specimens, e.g., in The Natural History Museum of London (see Fig. 10).

Fig. 10. Cast of a symphyseal tooth whorl (NHMUK P.9673) of the eugeneodontiform shark “Campodus variabilis” sensu Eastman (1902–1903) from the Coal Measures (Pennsylvanian) of Cedar Creek, Nebraska, USA. A. Lateral view. B. Oral view.
Following Zangerl (1981: 77) and Hampe in Ginter et al. (2010: 123) and contrary to Eastman (1902a, b, 1903) and Eaton (1962) I consider the Eastman’s understanding of Campodus to be too inclusive and the labio-lingual symmetry/asymmetry of the crowns as a sufficiently robust character to allow generic distinction. Therefore, the genus Campodus should be restricted to C. agassizianus and, possibly, Campodus sp. sensu Zangerl (1981: fig. 87) and Eastman’s (1902a, b, 1903) tooth whorls and the mandible illustrated by St. John and Worthen’s (1875: pl. 8: 1) should be excluded from this genus. This, however, does not solve the problem of the identity of the latter specimens.
The history of this problem is perfectly summarised by Hampe in Ginter et al. (2010: 125) but my conclusions partly differ from those presented in that text. Analysing all the available data I consider that St. John and Worthen (1875) incorrectly connected the mandible from Osage, Kansas, with the type specimens of Lophodus (changed to Agassizodus) variabilis Newberry and Worthen, 1870. On the other hand, I think that Eastman’s (1903) intuition on connecting his symphyseal tooth whorls with the mandible from Osage was correct, although in fact only the bar-like anterior teeth could be compared directly. The characters of the individual teeth in the tooth whorls (Fig. 10) have much in common with the largest mandibular teeth in St. John and Worthen’s specimen (Fig. 9). Therefore, it is necessary to erect a common new genus and species for all these three remarkable specimens. That new taxon should be placed among the Caseodontidae.
Campodus as a basal eugeneodontiform.—The taxonomic analysis in the previous section leads indirectly to a conclusion that Campodus is the earliest known eugeneodontiform and, even if Campodus sp. sensu Zangerl (1981) is included, its stratigraphic range is relatively short: Meramecian (middle Viséan)–Kinderscoutian (early Bashkirian). The general outline of the major lateral teeth is similar to those of the orodonts, typical Mississippian euchondrocephalans with a crushing dentition. However, the heterodonty in the orodonts is less pronounced, and in particular they do not have bar-like anterior and posterior lateral teeth. As stated above, symphyseal teeth or tooth whorls, typical of the Eugeneodontiformes, have not been found in the materials of Campodus. This does not mean, though, that they did not exist. There have been suggestions that Chiastodus obvallatus Trautschold, 1879, a single tooth from the Mississippian or Pennsylvanian of Myachkova, Russia (Hampe in Ginter et al. 2010: fig. 117) could belong to a symphyseal tooth family of Campodus, but this idea still requires confirmation. If such a symphyseal tooth family was found, I would rather suppose it to be similar to “Campodus” sensu Eastman (1902a) than to a cutting whorl of Edestus Leidy, 1855.
Ginter et al. (2010) placed Campodus within the Caseodontoidea, but without designation of a family. The similarity of the lateral teeth to those of Caseodus and the mandible of “Agassizodus” sensu St. John and Worthen (1875) on the one hand, but the presence of the labio-lingual symmetry on the other suggests two alternative solutions: placement of Campodus in the Caseodontidae or erection a new family for that genus.
The scenario of the early evolution of caseodontoid dentition could have been as follows: an orodont ancestor, (i) diversification of the mandibular teeth, emergence of small bar-like teeth, the major teeth still with rather simple ornamentation (as in Campodus sp. sensu Zangerl 1981), (ii) creation of elaborate ornamentation, further diversification of the teeth, emergence of specialised symphyseal tooth families (?), the labio-lingual symmetry still retained (Campodus agassizianus), (iii) enlargement of symphyseal tooth families and whorls, enlargement of transverse ridges on the labial side and reduction on the lingual side (Pennsylvanian and later caseodontids, but it also concerns all late eugeneodontiforms). It is difficult to say how and when the edestid cutting tooth whorls were formed, but it must have happened relatively early: the famous tooth whorl of Lestrodus (originally Edestus) newtoni (Woodward, 1917) from Yorkshire (see Ginter et al. 2010: fig. 126) was dated as early as Bashkirian.
Conclusions
The remains of a chondrichthyan devoured by scavenging ostracods, found in the limestone nodule from the Kinderscoutian (Bashkirian, Lower Pennsylvanian) of Derbyshire most probably belong to a basal caseodontoid eugeneodontiform, Campodus agassizianus De Koninck, 1844. Its major distinction from all the other Eugeneodontiformes is the labio-lingual symmetry of lateral teeth.
The arrangement of teeth on a jaw was probably similar to the famous mandible of “Agassizodus variabilis” (sensu St. John and Worthen 1875: pl. 8: 1; a new name for this specimen is required) from the Coal Measures of Osage, Kansas. Eastman’s (1902a, b, 1903) idea that the symphyseal tooth whorls named by him as “Campodus variabilis” are conspecific with “Agassizodus variabilis” may be correct, but these tooth whorls definitely do not belong to Campodus. It is still unknown whether Campodus had a specialised symphyseal tooth family at all. Future students of this genus should decide whether to place it in the family Caseodontidae or in a new, monotypic family. Both solutions I consider equally justified.
Acknowledgements
My warm thanks are due to several people affiliated to the British Geological Survey at Keyworth, UK: Mark Williams who drew my attention to the specimen in 2003, Ian P. Wilkinson who provided literature on the subject, Nick Riley (currently Carboniferous Limited, Keyworth, UK) who hosted me in his house in 2018 and told me the history of the nodule, and Paul J. Shepherd who organised my study at the British Geological Survey. I am also grateful to Emma Bernard who organised my study at The Natural History Museum in London, UK and to the colleagues who informed me about the situation of the other specimens of Campodus: Alexander Ivanov (St. Petersburg University, Russia), Oleg Lebedev (Palaeontological Institute of Russian Academy of Sciences, Moscow, Russia), Sebastien Olive (Royal Belgian Institute of Natural Sciences, Brussels, Belgium) and Valentin Fischer (University of Liège, Belgium). The insightful reviews by Christopher J. Duffin (Sutton, UK) and Wayne Itano (University of Colorado, Boulder, USA) greatly improved the manuscript. My research in the UK in 2003 was funded from the Royal Society scholarship to Richard J. Aldridge and in 2018 from the University of Warsaw Rector’s Microgrant.
References
De Koninck, L.G. 1844. Description des animaux fossiles qui se trouvent dans le terrain Carbonifère de la Belgique. 650 pp. Dessain, Liège.
Dineley, D.L. and Metcalf, S.J. 1999. Fossil Fishes of Great Britain. xxi + 675 pp. Joint Nature Conservation Committee, Peterborough.
Eastman, C.R. 1902a. On Campyloprion, a new form of Edestus-like dentition. Geological Magazine, decade 4 9: 148–152.
Eastman, C.R. 1902b. Some Carboniferous cestraciont and acanthodian sharks. Bulletin of the Museum of Comparative Zoology 39: 53–99.
Eastman, C.R. 1903. Carboniferous fishes from the central western States. Bulletin of the Museum of Comparative Zoology 39: 163–226.
Eaton, T.H. 1962. Teeth of edestid sharks. University of Kansas Publications 12: 347–362.
Ginter, M., Hampe, O., and Duffin, C.J. 2010. Chondrichthyes. Paleozoic Elasmobranchii: teeth. In: H.-P. Schultze (ed.), Handbook of Paleoichthyology 3D. 168 pp. Friedrich Pfeil, München.
Huxley, T.H. 1880. A Manual of the Anatomy of Vertebrated Animals. 431 pp. D. Appleton, New York.
Leidy, J. 1855. Indications of five species, with two new genera, of extinct fishes. Proceedings of the Academy of Natural Sciences of Philadelphia 7: 414.
Lohest, M. 1884. Rechèrches sur les poissons des terrains Paléozoïques de Belgique. Premiere Partie. Poissons de l’amplelite alunifère, des genres Campodus, Petrodus, et Xystracanthus. Annales de la Société Géologique de Belgique 11: 295–325.
Lund, R. and Grogan, E.D. 1997. Relationships of the Chimaeriformes and the basal radiation of the Chondrichthyes. Reviews in Fish Biology and Fisheries 7: 65–123. Crossref
Newberry, J.S. and Worthen, A.H. 1870. Descriptions of Vertebrates. Geological Survey of Illinois 4: 343–374.
Romanovsky, G. 1864. Description de quelques restes de poissons fossiles trouvés dans le calcaire carbonifère du gouvernement de Toula. Bulletin de la Société impériale des Naturalistes de Moscou 37: 157–170.
Stahl, B.J. 1999. Chondrichthyes III. Holocephali. In: H.-P. Schultze (ed.), Handbook of Paleoichthyology 4. 164 pp. Friedrich Pfeil, München.
St. John, O. and Worthen, A.H. 1875. Geology and palaeontology. Part II. Palaeontology of Illinois. Section I. Descriptions of fossil fishes. Geological Survey of Illinois 6: 245–488.
Trautschold, H. 1879. Die Kalkbrüche von Mjatschkowa. Eine Monographie des oberen Bergkalks. Schluss. Nouveau Mémoires de la Société Impériale des Naturalistes de Moscou 14: 3–82.
Wilby, P.R., Wilkinson, I.P., and Riley, N.J. 2006. Late Carboniferous scavenging ostracods: feeding strategies and taphonomy. Transactions of the Royal Society of Edinburgh: Earth Sciences 96: 309–316. Crossref
Wilkinson, I.P., Williams, M., Siveter, D.J., and Wilby, P.R. 2004. A Carboniferous necrophagous myodocopid ostracod from Derbyshire, England. Revista Española de Micropaleontología 36: 195–206.
Woodward, A.S. 1917. On a new species of Edestus from the Upper Carboniferous of Yorkshire. Quarterly Journal of the Geological Society of London 72: 1–6. Crossref
Zangerl, R. 1981. Chondrichthyes I: Paleozoic Elasmobranchii. In: H.-P. Schultze (ed.), Handbook of Paleoichthyology 3A. 115 pp. Gustav Fischer, Stuttgart.
Zangerl, R., Winter, H.F., and Hansen, M.C. 1993. Comparative microscopic dental anatomy in the Petalodontida (Chondrichthyes, Elasmobranchii). Fieldiana: Geology, N.S. 26: 1–43.
Acta Palaeontol. Pol. 63 (4): 725–735, 2018
https://doi.org/10.4202/app.00533.2018