

Two new basal coelurosaurian theropod dinosaurs from the Lower Cretaceous Sao Khua Formation of Thailand
ADUN SAMATHI, PHORNPHEN CHANTHASIT, and P. MARTIN SANDER
Samathi, A., Chanthasit, P., and Sander, P.M. 2019. Two new basal coelurosaurian theropod dinosaurs from the Lower Cretaceous Sao Khua Formation of Thailand. Acta Palaeontologica Polonica 64 (2): 239–260.
Megaraptora is a clade of mid to large-sized theropods that are long-snouted, large-clawed, highly pneumatized, and have long and gracile metatarsals. The basal member was reported from the Barremian of Japan. A more derived clade, the Megaraptoridae, is known from the Cenomanian to Santonian of Gondwana. Here two new basal coelurosaurs from the Lower Cretaceous Sao Khua Formation of Thailand are described and named as Phuwiangvenator yaemniyomi gen. et sp. nov. and Vayuraptor nongbualamphuensis gen. et sp. nov. Phuwiangvenator is a megaraptoran coelurosaur and diagnosed by the ventrally flat sacral vertebrae with sulci in the anterior and posterior region of the centra and the anterior rim of metatarsal IV sloping proximolaterally to distomedially and being much lower than that of metatarsal III anteriorly. Vayuraptor is a basal coelurosaur and diagnosed by its astragalus which has two horizontal grooves, two fossae at the base of the ascending process, the ascending process being straight laterally and straight and parallel medially with the medial rim sloping to the tip laterally, and a long and slender astragalar ascending process. Although the position of the basal coelurosaur Vayuraptor remains unclear and must await further discovery, megaraptoran affinities are likely. The Early Cretaceous megaraptoran fossil record has been recovered from the Barremian to Aptian of Asia. All Asian megaraptorans might be a monophyletic clade or a paraphyletic series relative to the Megaraptoridae. Several specimens have been reported from the Aptian to mid-Cretaceous of Australia, and one report from the Albian of South America. These fossils show a high diversity of the Early Cretaceous megaraptorans and a wide distribution during that time. The clade then became more provincial in the Late Cretaceous.
Key words: Dinosauria, Theropoda, Coelurosauria, Megaraptora, Phuwiangvenator, Vayuraptor, Cretaceous, Thailand.
Adun Samathi [asamathi@gmail.com] and P. Martin Sander [martin.sander@uni-bonn.de], Division of Paleontology, Institute of Geosciences, Rheinische Friedrich-Wilhelms Universität Bonn, Nussallee 8, Bonn, 53115, Germany.
Phornphen Chanthasit [aom025@gmail.com], Sirindhorn Museum, Department of Mineral Resources, 200 Moo 11, Kalasin, 46140, Thailand.
Received 27 August 2018, accepted 15 March 2019, available online 27 May 2019.
Copyright © 2019 A. Samathi et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Megaraptora (sensu Benson et al. 2010) is a clade of medium to large-sized highly pneumatized theropods with a long-snout, large-manual claws, and long and gracile metatarsals (Benson et al. 2010; Coria and Currie 2016). A basal member was reported from the Barremian of Japan, Fukuiraptor (Azuma and Currie 2000; Benson et al. 2010; Porfiri et al. 2014). A more derived clade, the Megaraptoridae (sensu Novas et al. 2013), is known from the Cenomanian to Santonian rocks of Gondwana (Porfiri et al. 2014; Novas et al. 2016). The clade consists of the South American taxa Megaraptor (Novas 1998), Aerosteon (Sereno et al. 2008), Murusraptor (Coria and Currie 2016), Orkoraptor (Novas et al. 2008), and Tratayenia (Porfiri et al. 2018) as well as Australovenator from Australia (Hocknull et al. 2009).
Several fragmentary specimens have been reported and referred to the Megaraptora including an astragalus (NMV P150070) from the Aptian of Australia (Agnolin et al. 2010; Benson et al. 2012b); the Lightning Ridge megaraptorid (LRF 100-106) from the Albian of Australia (Bell et al. 2015); an ulna (NMV P186076) from the Aptian–Albian of Victoria, Australia (Smith et al. 2008), and Rapator from Australia (Agnolin et al. 2010; White et al. 2013b). Some specimens have been reported from South America including caudal vertebrae from Brazil (CPPLIP 1324, Martinelli et al. 2013; and MPMA 08-003-94, Mendez et al. 2012), which were suggested to be sacral vertebrae by Motta et al. (2016) (see Sales et al. 2017); another caudal vertebra (UFRGS-PV-032-K, Sales et al. 2017); and sacral vertebrae from Brazil (SMNS 58023; Aranciaga Rolando et al. 2018). Furthermore, a left tibia with associated astragalocalcaneum (SM-NB A1-2) and postcranial material (SM-PW9B) have been recently reported from the Early Cretaceous of Thailand (Samathi and Chanthasit 2015, 2017) and are described here in detail.
Problematic taxa possibly belonging to Megaraptora are Siats from North America (Zanno and Makovicky 2013; see Novas et al. 2013 for an alternative interpretation), Aoniraptor from South America (Motta et al. 2016), and Eotyrannus from Europe (Hutt et al. 2001). This taxon was found to be a megaraptorid nested within Tyrannosauroidea by Porfiri et al. (2014). Other taxa interpreted as related with Megaraptora are Gualicho from South America (Apesteguía et al. 2016), Chilantaisaurus from China (Hu 1964; Benson and Xu 2008; see Delcourt and Grillo 2018), and Neovenator from England (Brusatte et al. 2008). Chilantaisaurus and Siats were recovered as megaraptorans more derived than Fukuiraptor by Bell et al. (2015), however, these two taxa do not exhibit megaraptoran features (Novas et al. 2013). Siats may fall among uncertain tetanurans. These results are contradictory due to the fragmentary nature of these theropods or different interpretation (see Porfiri et al. 2014).
Megaraptora phylogeny.—The phylogenetic status of Megaraptora is still debated and deserves further testing. Currently, there are three hypotheses of the phylogenetic position of Megaraptora: (i) deeply nested within Allosauroidea (Benson et al. 2010; Carrano et al. 2012); (ii) nested within Tyrannosauroidea (Novas et al. 2012a; 2013; Porfiri et al. 2014; Cau 2018); and (iii) Megaraptora are the most basal Coelurosauria (Novas et al. 2015; Apesteguía et al. 2016; Ezcurra and Novas 2016; Delcourt and Grillo 2018) (Fig. 1). The reason is the fact that they show “intermediate” or “mixed” characters in their anatomy which could be interpreted as “derived” characters for allosauroids or “basal” characters in Coelurosauria (Benson et al. 2010; Novas et al. 2013; Porfiri et al. 2014). This includes the “primitive” characters, e.g., short and broad scapula, distal tibia with vertical medial ridge, as well as the “derived” characters, e.g., elongate hindlimb, the tall ascending process of the astragalus, and pectoral and pelvic pneumaticity (Benson et al. 2010; Novas et al. 2013). The spinosaurids also show some features such as a short and broad scapula, distal tibia with vertical ridge, relatively tall and sheet-like ascending process of the astragalus, and large manual claws which arose convergently among megaraptorans. However, large manual unguals are also present in comsognathids.
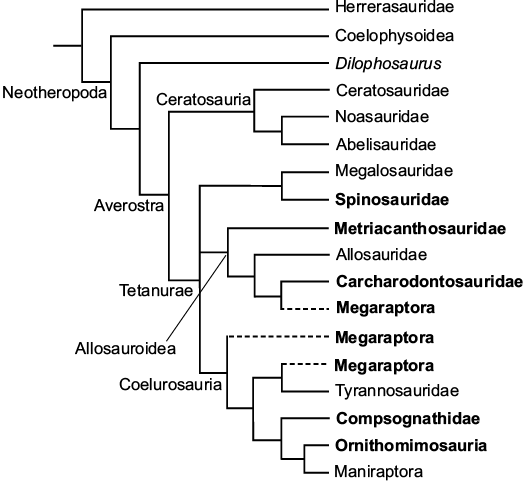
Fig. 1. Three hypotheses of the phylogenetic position of Megaraptora. Megaraptora as derived Carcharodontosauria (Benson et al. 2010; Carrano et al. 2012); basal Coelurosauria (Novas et al. 2015; Apestiguea et al. 2016; Delcourt and Grillo 2018); and basal Tyrannosauroidea (Novas et al. 2012a, 2013; Porfiri et al. 2014; Cau 2018). The tree is modified from Ezcurra and Novas (2016). Bolded are theropods found in Thailand.
New coelurosaurs, probable megaraptorans, from Thailand.—Currently, nine theropods have been reported from the Khorat Group of northeastern Thailand. Until now, five theropods have been found from the Sao Khua Formation (e.g., Buffetaut and Suteethorn 2012; see the geological setting part in this paper). There are at least two new coelurosaurian theropod specimens from the Sao Khua Formation that may belong to the Megaraptora. These are the focus of the current paper. We here describe Phuwiangvenator yaemniyomi gen. et sp. nov. based on a partial postcranial skeleton and a second, smaller coelurosaur, Vayuraptor nongbualamphuensis gen. et sp. nov., based on hindlimb elements (Samathi and Chanthasit 2017). The new Thai taxa might inform us about the phylogenetic position of Megaraptora among Avetheropoda and the origin and paleobiogeography of Megaraptora which might have originated in Asia or Southeast Asia.
Institutional abbreviations.—CPPLIP, Centro de Pesquisas Paleontológicas Llewellyn Ivor Price-Peirópolis, Uberaba County, Minas Gerais, Brazil; FPDM, Fukui Prefectural Dinosaur Museum, Katsuyama, Japan; FSAC, Faculté des Sciences Aïn Chock (University of Casablanca), Casablanca, Morocco; MB, Museum für Naturkunde, Berlin, Germany; MCNA, Museo de Ciencias Naturales y Antropológicas “Cornelio Moyano”, Mendoza, Argentina; MNBH, Musée National Boubou Hama, Niamey, Republic of Niger; MPMA, Museu de Paleontologia de Monte Alto, Brazil; MSNM, Museo di Storia Naturale di Milano, Milan, Italy; NMV, Museum Victoria, Melbourne, Australia; PRC, Paleontological Research and Education Center, Maha Sarakham University, Maha Sarakham, Thailand; SM, Sirindhorn Museum, Department of Mineral Resources, Kalasin, Thailand; SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany; UMNH, Natural History Museum of Utah, Salt Lake City, Utah, USA.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:DE9BDAD4-A657-485F-ADE2-F2DD2CCA1A7B.
Material and methods
The holotype of Phuwiangvenator yaemniyomi gen. et sp. nov. described in this paper was discovered by Preecha Sainongkham, a staff member of Phu Wiang Fossil Research Center and Dinosaur Museum in 1993, with some referred material found later. The fossils are housed at the Sirindhorn Museum, Kalasin Province under the Department of Mineral Resources, Thailand. The holotype elements of Phuwiangvenator yaemniyomi gen. et sp. nov. seem to represent a single individual, based on size (see SOM: table 1, Supplementary Online Material available at http://app.pan.pl/SOM/app64-Samathi_etal_SOM.pdf), matching articulation, and shared phylogenetic affinity. The referred elements, found about 300 m away from the holotype, seem to belong to the same animal as the holotype, based on the size, matching articulation (e.g., the right astragalocalcaneum fits perfectly to the right tibia) and shared phylogenetic affinity (i.e., the right astragalocalcaneum and the left astragalocalcaneum).
The holotype of Vayuraptor nongbualamphuensis gen. et sp. nov. consists of a left tibia with associated astragalocalcaneum with referred material (see description below). They were discovered by Paladej Srisuk (PRC) in 1988. The holotype material is housed at the Sirindhorn Museum, Kalasin Province under the Department of Mineral Resources, Thailand and the referred material is housed at the collection of the PRC, Maha Sarakham University, Maha Sarakham Province, Thailand. All the available elements of Vayuraptor nongbualamphuensis gen. et sp. nov. seem to represent a single individual, based on size and shared phylogenetic affinity.
For phylogenetic analyses, the data matrix was edited in the program Mesquite (Maddison and Maddison 2015), and the program TNT (Goloboff et al. 2008) was used and for finding the shortest tree under the parsimony and optimality criterium. We follow the analytical setting and data matrix of Apesteguía et al. (2016). This is a modified version of the Novas et al. (2013) matrix with additional characters and taxa by Apesteguía et al. (2016) and focused on Allosauroidea and basal Coelurosauria. Ceratosaurus serves as the outgroup. The 288 characters used were unordered and equally weighted with 46 theropod taxa in each analysis. We used the “New Technology” search option which included the default settings for sectorial, ratchet, tree drift, and tree fusion. Because the two Thai theropods are fragmentary, we then separated them into two different analyses for better resolution. In the first analysis, we added Phuwiangvenator and Siamotyrannus into this matrix and excluded Santanaraptor for a better resolution since this taxon is fragmentary. In the second analysis, we added Vayuraptor and Siamotyrannus into the matrix and excluded Santanaraptor and Chilantaisaurus.
Geological and geographical setting
The first dinosaur bone from Thailand was found in the 1980’s at the Phu Wiang Mountain, Khon Kaen Province. It was a fragment of a sauropod femur from the Sao Khua Formation (Buffetaut 1982). Since then, a collaboration led by the Department of Mineral Resources, Thailand and the Centre National de la Recherche Scientifique, France has led to the discovery of many other dinosaur remains including ornithischians, sauropods, and theropods (e.g., Buffetaut and Suteethorn 1992; Martin et al. 1994; Suteethorn et al. 2009).
The Sao Khua Formation, from which the two new basal coelurosaurs from Thailand are described here, is one of the most fossil-rich formations of the Khorat Group of northeastern Thailand (Fig. 2). It consists of red clay, sandstone, and conglomerate, indicating deposition in a floodplain with meandering rivers. The fauna contains freshwater hybodont sharks, actinopterygian fishes, turtles, crocodilians, and dinosaurs (Buffetaut and Suteethorn 1998). The formation was originally considered Jurassic in age, but it was later shown to be in the Early Cretaceous. Buffetaut and Suteethorn (1999) were the first to suggest an Early Cretaceous age based on the vertebrate fossils. Most recently, the age was constrained to the late Barremian based on non-marine bivalves (Tumpeesawan et al. 2010). We follow this age assignment. Until now, five distinct theropods have been reported from the Sao Khua Formation. Theses consists of a possible compsognathid (Buffetaut and Ingavat 1984), teeth of the spinosaurid Siamosaurus suteethorni (Buffetaut and Ingavat 1986), the theropod Siamotyrannus isanensis (Buffetaut et al. 1996), the ornithomimosaur Kinnareemimus khonkaenensis (Buffetaut et al. 2009), and the partial skull of a carcharodontosaurid (Buffetaut and Suteethorn 2012).
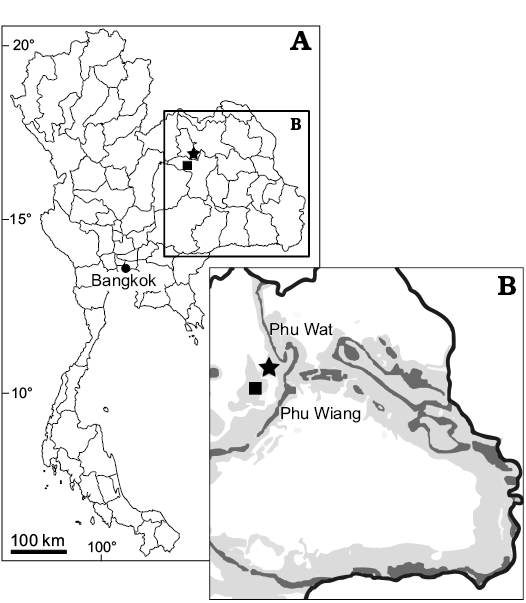
Fig. 2. Map of Thailand (A) and close-up of northeastern Thailand (B) showing the location of Phu Wiang locality, Khon Kaen Province (square) and Phu Wat locality, Nong Bua Lamphu Province (star). Dark gray, Sao Khua Formation outcrops, light gey, Phu Kradung and Khok Kruat formations.
Systematic palaeontology
Dinosauria Owen, 1842
Theropoda Marsh, 1881
Tetanurae Gauthier, 1986
Coelurosauria von Huene, 1920
Megaraptora Benson, Carrano, and Brusatte, 2010
Genus Phuwiangvenator nov.
Zoobank LCID: urn:lsid:zoobank.org:act:5FF98BC6-5B5E-45AF-8F 90-AC12DBDE57E7
Type species: Phuwiangvenator yaemniyomi gen. et sp. nov., monotypic, see below.
Etymology: Phu Wiang Mountain, Khon Kaen Province, Thailand, the place where the holotype was found, combined with Latin venator, hunter; thus “Hunter of Phu Wiang”.
Diagnosis.—Same as for the only known species.
Phuwiangvenator yaemniyomi sp. nov.
Figs. 3–14.
Zoobank LCID: urn:lsid:zoobank.org:act:AA829C5F-84C6-4287-BED 7-50DDC551A815
Etymology: In honor of Sudham Yaemniyom, former geologist of the Department of Mineral Resources, Bangkok, who found the first dinosaur bone of Thailand in 1976 at Phu Wiang Mountain.
Holotype: SM-PW9B, a partial skeleton consisting of a dorsal vertebra, three fused sacral vertebrae, right metacarpal II, right manual phalanges and unguals, right and left tibiae, left astragalocalcaneum, left metatarsal I, right metatarsals II–IV, right pedal phalanges and unguals. The bones were found in an area of about 5 square meters.
Type locality: Phu Wiang Site 9B, Phu Wiang Mountain, Khon Kaen Province, Thailand (Fig. 2).
Type horizon: Sao Khua Formation, probably upper Barremian, Lower Cretaceous.
Referred material.—SM-PW9A, an atlantal intercentrum and right astragalocalcaneum which were found together and approximately 300 m away from the holotype (Phu Wiang Site 9A).
Diagnosis.—Phuwiangvenator is a megaraptoran diagnosed by the following autapomorphies: (i) short sulci on the sacral vertebrae ventrally along the anterior and posterior part of the centrum; (ii) the anterior rim of metatarsal IV slopes from proximolaterally to distomedially. This way, the distomedial corner of the proximal articular surface of metatarsal IV in anterior view is much lower than the articular surface of metatarsal III. In addition, the distomedial corner is lower than in any other known theropod.
Description.—General morphology: Phuwiangvenator was a mid to large-sized theropod with an estimated body length of 6 m. The skeletal proportion are shared by other theropods of corresponding sized such as Australovenator.
Atlantal intercentrum: The atlantal intercentrum is well preserved, only the posterior portion is slightly eroded. The atlantal intercentrum is concave anteriorly, and high dorsoventrally. The odontoid concavity in the dorsal surface is smooth. The distinct ventrolateral process on the atlantal intercentrum is not present (Fig. 3A).
Dorsal vertebra: The dorsal vertebra (SM-PW9B-12) was recovered (Fig. 3B). The neural arch is missing and possibly was not fused to the centrum. The centrum looks similar to that of Fukuiraptor (FPDM-V-8381; AS personal observations) and Gualicho (Apesteguía et al. 2016). The centrum has no pleurocoel and no ventral and lateral rugosities in the anterior and posterior end of the centrum, the same as in Gualicho (Apesteguía et al. 2016). The centrum is longer than high with a lateral depression on the lateral sides. The anterior face of the centrum is concave, whereas the posterior is flat (platycoelous). The centrum shape is circular anteriorly and posteriorly. The vertebra of Phuwiangvenator are highly constricted, hour-glass shaped in ventral view and very concave in the ventral part in lateral view which is same as in dorsal vertebrae of Fukuiraptor, Aerosteon, Allosaurus, Tyrannosaurus, Ornitholestes, Coelurus, and Zuolong (Madsen 1976; Brochu 2003; Carpenter et al. 2005b; Choiniere et al. 2010). This differs from caudal vertebrae which normally are less concave in the ventral part in lateral view and less constricted in ventral view compared with dorsal vertebrae. The lack of a parapophysis on the centrum suggest that it belongs to the middle to posterior portion of the dorsal series. The highly constricted posterior dorsal vertebral centrum is present in Phuwiangvenator as well as in allosauroids, megaraptorids, and derived tyrannosauroids (Novas et al. 2013). The dorsal vertebra of Phuwiangvenator also differs from the sacral vertebrae in the constriction of the centrum in ventral and lateral views. There is no keel or groove on the centrum ventrally. There is no facet for the chevron, so it is clearly not a caudal vertebra. As seen on broken surfaces, the dorsal vertebra appears to have a camellate internal structure.
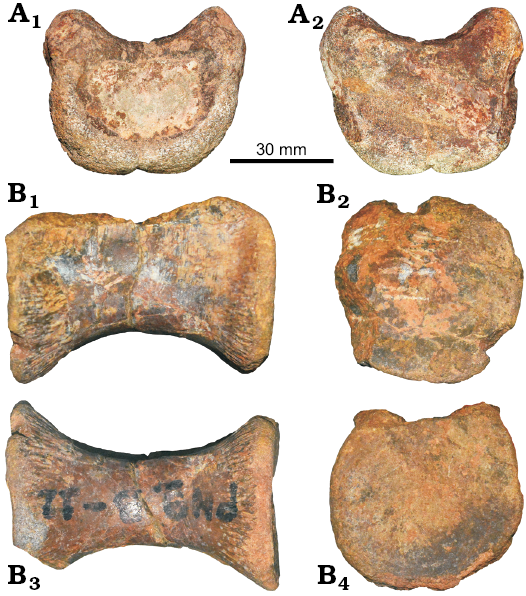
Fig. 3. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. A. SM-PW9A-1, atlantal intercentrum in anterior (A1) and posterior (A2) views. B. SM-PW9B-12, dorsal vertebra in left lateral (B1), anterior (B2), ventral (B3), and posterior (B4) views.
Sacral vertebrae: Three incomplete fused sacral vertebrae are preserved (SM-PW9B-39). The anterior face of the centrum is flat. The centra are possibly sacral 1, 2, and 3 based on the probable sacral ribs which are situated on the anterodorsal side of the centrum. The neural arches of sacral 1 and 2 are badly preserved and still covered by the matrix. The quality of preservation of the centra is generally good, especially on the right side. The left side is compressed. The centra are flat ventrally, there is no keel and no constriction on the sacrals ventrally. There are small, short sulci on the anterior and posterior part of the sacral 2 and one sulcus on the anterior part of the sacral 3 (the posterior part has no sulci) ventrally (Fig. 4). There are no pleurocoels or any foramina. The centra are longer than high. The sacral rib articulation is located low for sacral 3 anteriorly, at middle for sacral 1, and anteriorly in scaral 2.
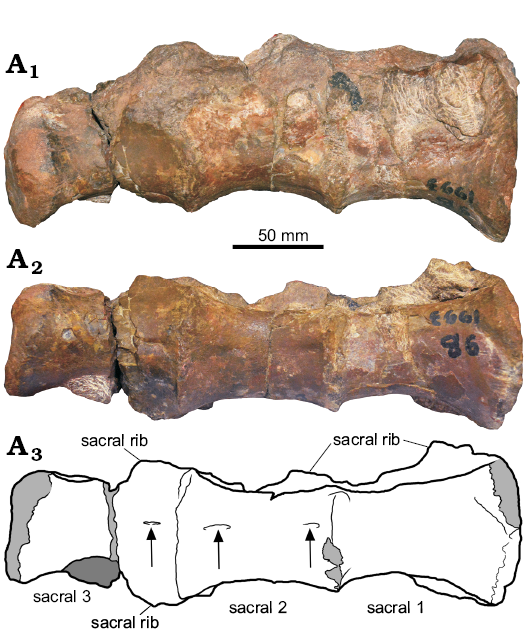
Fig. 4. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9B-39, anterior sacral vertebrae in right lateral (A1) and ventral (A2) views; line drawing in ventral view (A3). Arrows indicate ventral sulci, light grey, broken part; dark grey, matrix.
Manual phalanges and unguals: The manual phalanges and unguals consist of manual phalanx I-1 (SM-PW9B-3); manual ungual I-2 (SM-PW9B-19); metacarpal II (SM-PW9B-6, Fig. 5); manual ungual II-3 (SM-PW9B-23); manual phalanx III-1 (SM-PW9B-4); manual phalanx III-2 (SM-PW9B-7); manual phalanx III-3 (SM-PW9B-5); and manual ungual III-4 (SM-PW9B-21) (Fig. 6, 7).

Fig. 5. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9B-6, right metacarpal II in dorsal (A1), ventral (A2), distal (A3), and medial (A4) views.
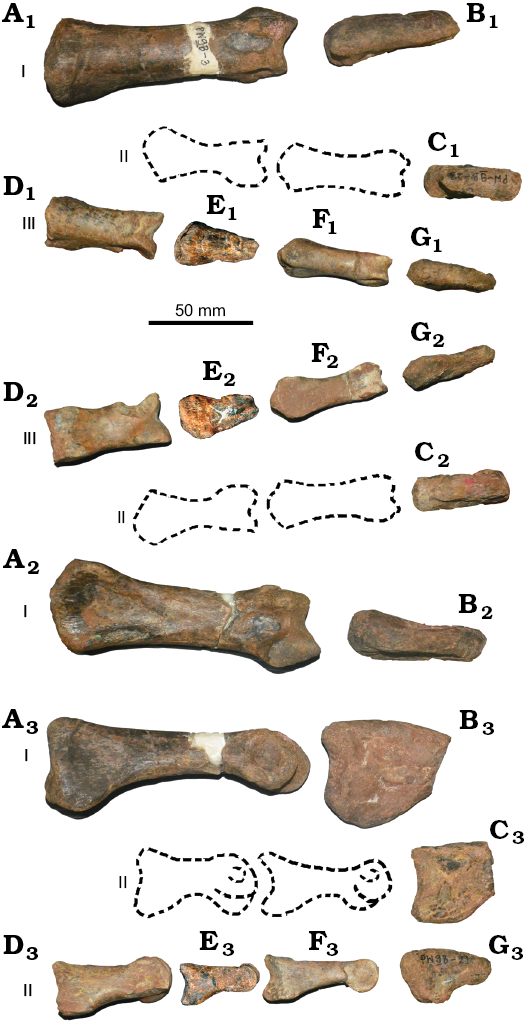
Fig. 6. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9B-3 (A), SM-PW9B-19 (B), SM-PW9B-23 (C), SM-PW9B-4 (D), SM-PW9B-7 (E), SM-PW9B-5 (F), SM-PW9B-21 (G); manual phalanges I–III (A, D, E, F) and unguals I–III (B, C, G), in dorsal (A1–G1), ventral (A2–G2), and lateral (A3–G3) views. Dashed lines are the reconstruction of manual phalanges II-1 and II-2.
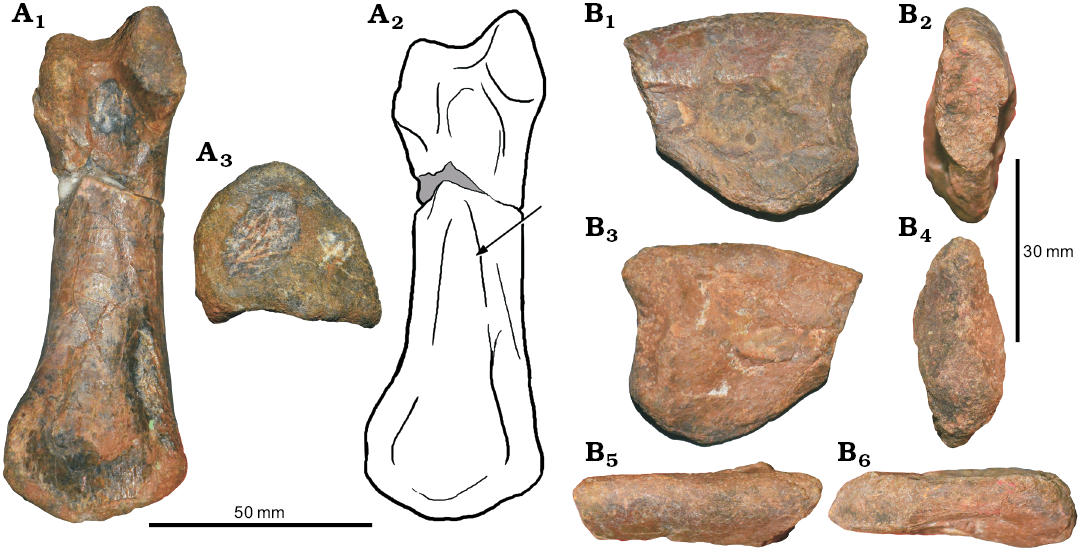
Fig. 7. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. A. SM-PW9B-3, manual phalanx I-1 in ventral (A1) and proximal (A3) views; line drawing in ventral view (A2). B. SM-PW9B-19, manual ungual I-2 in medial (B1), distal (B2), lateral (B3), proximal (B4), dorsal (B5), and ventral (B6) views. Arrow indicates the longitudinal ventral furrow.
In general, manual phalanges are referred to the Theropoda based on the presence of deep, well-defined collateral ligament pits and the presence of an extensor fossa on the dorsal surface proximal to the distal articulation (Rauhut 2003).
Only the distal part of the right metacarpal II (SM-PW9B-6) is preserved. It is long and gracile. The metacarpal is dorsoventrally flattened and resembles the metacarpal II of the basal tyrannosauroid Kileskus from Siberia (Averianov et al. 2010) and identified here to be a distal part of the right metacarpal II. The distal end of metacarpal II is asymmetrical; it is slightly expanded and divided into two distinct condyles. The distal surface of the medial condyle is larger than the lateral condyle, as in Kileskus and Australovenator (White et al. 2012). The medial condyle protrudes slightly more distally than the lateral condyle. The pits for the collateral ligaments are not well developed on both condyles. There is no distinct extensor pit on the anterior surface of the shaft proximal to the distal condyle, only a small concavity is present.
The manual phalanx I-1 (SM-PW9B-3) has deep, well-defined collateral ligament pits which are larger on the medial side than the lateral one. There is an extensor fossa on the dorsal surface proximal to the distal articulation. The shaft is proximodistally elongate, shows a high ratio of proximodistal length to transverse width and has a ginglymoid articulation which is asymmetrical and elongated mediodistally. The lateral condyle is about the same height dorsoventrally but shorter proximo-distally than the medial condyle, as in Australovenator, in which the lateral condyle is also shorter proximo-distally than the medial condyle (White et al. 2012). The lateral condyle is expanded proximally. The proximal end is asymmetrical, with the medial articular surface larger than the lateral surface. The proximal articular surface is transversely wider on its ventral margin than on its dorsal margin. A ridge on the proximal articular surface slopes slightly laterally rather than being in the middle of the articular surface. This feature is also present in Gualicho (Apesteguia et al 2016). The manual phalange I-1 of Phuwiangvenator has a longitudinal ventral furrow and longitudinal ridges, and it is convex ventrally in proximal view (Fig. 7A).
The manual phalanx III-1 (SM-PW9B-4) has well-defined collateral ligament pits. There is a shallow, poorly defined extensor fossa on the dorsal surface proximal to the distal articulation. There is no shallow furrow on the ventral surface of the phalanx. The shaft is proximodistally short and relatively robust. The ratio of proximodistal length to transverse width is approximately 2.8. The bone also has a ginglymoid articulation and is asymmetrical and elongated mediodistally. Two prominent ligament scars on the ventral surface proximal to the proximal articulation and one facet for articulation on the proximal end are present as in Allosaurus.
The manual phalanx III-2 (SM-PW9B-7) is not well preserved especially on the distal articulation. However, this bone shows deep, well-defined collateral ligament pits on the condyles and no extensor fossa on the dorsal surface proximal to the distal articulation. It is relatively shorter than the manual phalanx III-2 of other theropods.
The manual phalanx III-3 (SM-PW9B-5) has deep, well-defined collateral ligament pits on both condyles. There is no extensor fossa on the dorsal surface proximal to the distal articulation. There is no shallow furrow on the ventral surface of the phalanx. The bone is proximodistally elongate and slender, the ratio of proximodistal length to transverse width is 4. The bone has a ginglymoid articulation which is symmetrical. Two ligamental scars on the ventral surface proximal to the proximal articulation are present but not prominent or well defined. The shaft is slender, and its proximal part is extended ventrally. Two facets for articulation on the proximal end are present. Digit III is much more slender compared with Australovenator (White et al. 2012) and Allosaurus (Madsen 1976). Phalanx III-3 is as long and slender as phalanx III-3 of Australovenator (White et al. 2012).
The manual unguals consist of three unguals in this study. The manual ungual I-2 (SM-PW9B-19) is large, missing the distal portion and the surface of the lower half of the left side. There is a small depression on the right side. In proximal view, the articular surface is almost symmetrical. The medial condyle has almost the same height dorsoventrally as the lateral condyle, but it is slightly broader than the lateral condyle. Based on the distal articular surface of phalange I-1, we suggest that this ungual is an ungual I-2 and belongs to the first digit. The proximal end is slightly broader mediolaterally than the distal-most preserved portion. The prominent, sub-oval, mound-like flexor tubercle is present proximally on the ventral surface (Fig. 7). The vascular grooves are present on the lateral and medial surfaces, and are symmetrical. The proximal height/width ratio is 2.4. The manual ungual II-3 (SM-PW9B-23) is medium to large sized and missing the distal portion. The vascular groove is preserved on the medial side. It is symmetrical, the proximal end and the distal-most preserved portion are slightly equal in width mediolaterally. The sub-oval, mound-like flexor tubercle is present proximally on the ventral surface. The proximal height/width ratio is 2.67. The manual ungual III-4 (SM-PW9B-21) is small to medium sized and missing the distal portion. The vascular groove cannot be observed due to the preservation. The articular surface is symmetrical. The proximal end is slightly broader mediolaterally than the distal-most preserved portion. A prominent, sub-oval, mound-like flexor tubercle is present proximally on the ventral surface. The proximal height/width ratio is 2.27.
Tibiae: The right tibia is complete but fractured in the distal portion. The left tibia is not complete, lacking the tibial shaft, only the proximal and distal ends are preserved. The proximal part of the right tibia is better preserved than that of the left one. The tibial shaft is long and almost straight. The cnemial crest is expanded. The anterolateral process of the lateral condyle of Phuwiangvenator curves ventrally as a point process as in Neovenator (Brusatte et al. 2008), Australovenator (Hocknull et al. 2009), Allosaurus (UMNH VP 7148, 7922, 7932, 7938, 7939, 7940; AS personal observations), Lythronax (UMNH VP 20200; AS personal observations), Teratophoneus (UMNH VP 16690; AS personal observations), and Tyrannosaurus (Brochu 2003), but not in Fukuiraptor and Vayuraptor. The presence of this process may be more widespread than previously thought as suggested by Novas et al. (2013). The tibia shows derived states, the elongated and expanded medial malleolus and a distally expanded lateral malleolus (Brusatte and Sereno 2008; Figs. 8, 9).
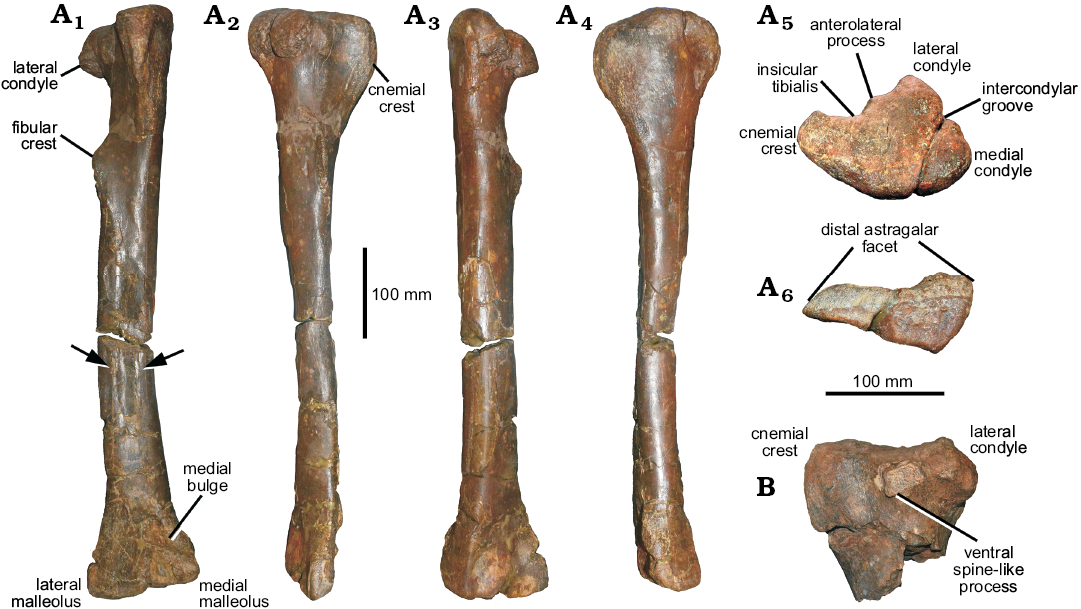
Fig. 8. Coelurosaurian theropod Phuwiangvenator
yaemniyomi gen. et sp. nov. from the Early Cretaceous
Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. A. SM-PW9B-41, right tibia in anterior (A1), lateral (A2), posterior (A3), medial (A4), proximal (A5), and distal (A6) views. B.
SM-PW9B-40, proximal left tibia in lateral view. Arrow indicates
vertical ridge on the tibia.
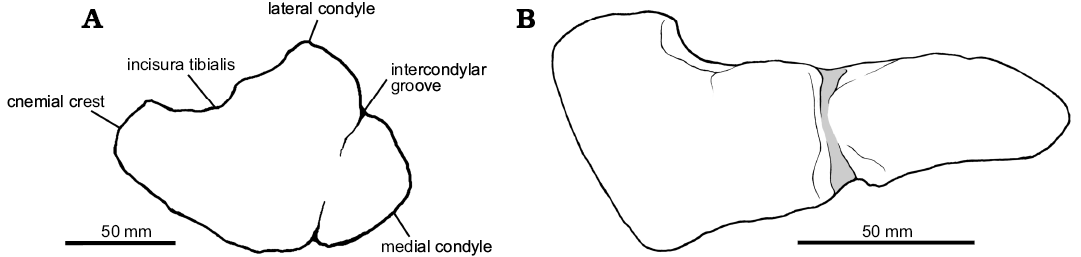
Fig. 9. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. A. SM-PW9B-41, right tibia in proximal view. B. SM-PW9B-40, left tibia in distal view. Light grey indicates broken bone.
The morphology of the distal cnemial process of the tibia is rounded. The lateral condyle is small relative to the tibial shaft. It offset from the lateral side of the proximal end of the tibia by a notch (posterior cleft) posteriorly. The fibular crest (the fibular flange) of the tibia is present on the proximal half as a pronounced longitudinal ridge. The fibular crest is clearly offset from the proximal articular facet of the tibia. It does not connect with the proximal end of the tibia (the ridge continuing from the fibular flange to the proximal articular surface of the tibia is absent). The fibular crest morphology is teardrop-shaped in lateral view. The primary nutrient foramen is present and situated posterior to the distal portion of the fibular crest. In the proximal end, the posterior rim of the lateral condyle is short and does not reach the same level as the medial condyle. The lateral malleolus of the distal tibia is overlapped by the calcaneum. The distal end of the tibia is triangular in outline, flattened anteroposteriorly, and strongly expanded laterally and medially. The lateral malleolus of the distal expansion extends laterally and extends further distally than the medial malleolus. The anteromedial buttress for the astragalus is a bluntly rounded vertical ridge on the medial side. The tibia length of Phuwiangvenator is more than 12 times its anteroposterior width at mid-length. This is a synapomorphy of megaraptorans and coelurosaurs (Porfiri et al. 2014) but also present in Spinosaurus (AS personal observations; see Discussion; SOM: table 17).
Astragalocalcaneum.—The left astragalocalcaneum (SM-PW9B-18) is nearly complete, lacking only the body of the ascending process of the astragalus (Fig. 10B). The astragalus is 103 mm wide, the calcaneum is 25 mm wide. The astragalar width/calcaneum width ratio is 4.12. The morphology of the ascending process of the astragalus is laminar. It is approximately 90 mm long estimated from the scar for the reception of the ascending process of the astragalus on the distal end of the tibia, approximately 15% of the total length of the tibia. However, this scar does not necessarily correspond to the shape and height of the ascending process of the astragalus (Rauhut and Xu 2005; Rauhut 2012). It is offset from the anterior border of the astragalar body by a shallow groove. The ascending process height relative to the depth of the astragalar body is approximately 1.7 times. The ascending process is slightly transversely narrow compare to more derived coelurosaurs, but it is wider than in Allosaurus (UMNH VP 11003; AS personal observations). The ascending process of the astragalus arises from more than half of the breath of the astragalar body. A round fossa at the base of the ascending process of the astragalus is present.
The cranioproximal process is present, but small compared to other theropods. A distinct anterior development of the lateral condyle (in ventral view) of the astragalar body is present. A proximolateral extension of the astragalus is also present as in Fukuiraptor (AS personal observations), but with some damaged on its tips.
The trapezoidal outline of the bone in distal view looks similar to Australovenator and the megaraptoran astragalus NMV P150070 (Benson et al. 2012b). In distal view, the astragalar body is concave anteriorly and straight posteriorly. Its surface is flat on the lateral half and convex on the medial half.
The astragalus condyle is significantly expanded proximally on the anterior side of the tibia and faces anterodistally. The orientation of the distal condyles of the astragalus is 30–45° anteriorly. A horizontal groove across the astragalar condyle is present anteriorly. The development of the articular surface for the distal end of the fibula in the astragalus is reduced and situated laterally. A posterolateral crest and a posteromedial crest on the astragalus are not present. The articulation between the ascending process and the fibula is not preserved. Both astragalus and calcaneum are unfused. The calcaneum bears a well-developed facet for the tibia. The calcaneum transverse development is moderate, being approximately 24% (left) and 23% (right) of the width of the astragalus.
The right astragalocalcaneum (SM-PW9A-B17) is referred to the same taxon and the same individual as the holotype of Phuwiangvenator since it perfectly matches with the right tibia of the holotype and shows the same size and characters as the left astragalocalcaneum (Fig. 10A; SOM: tables 6–9).
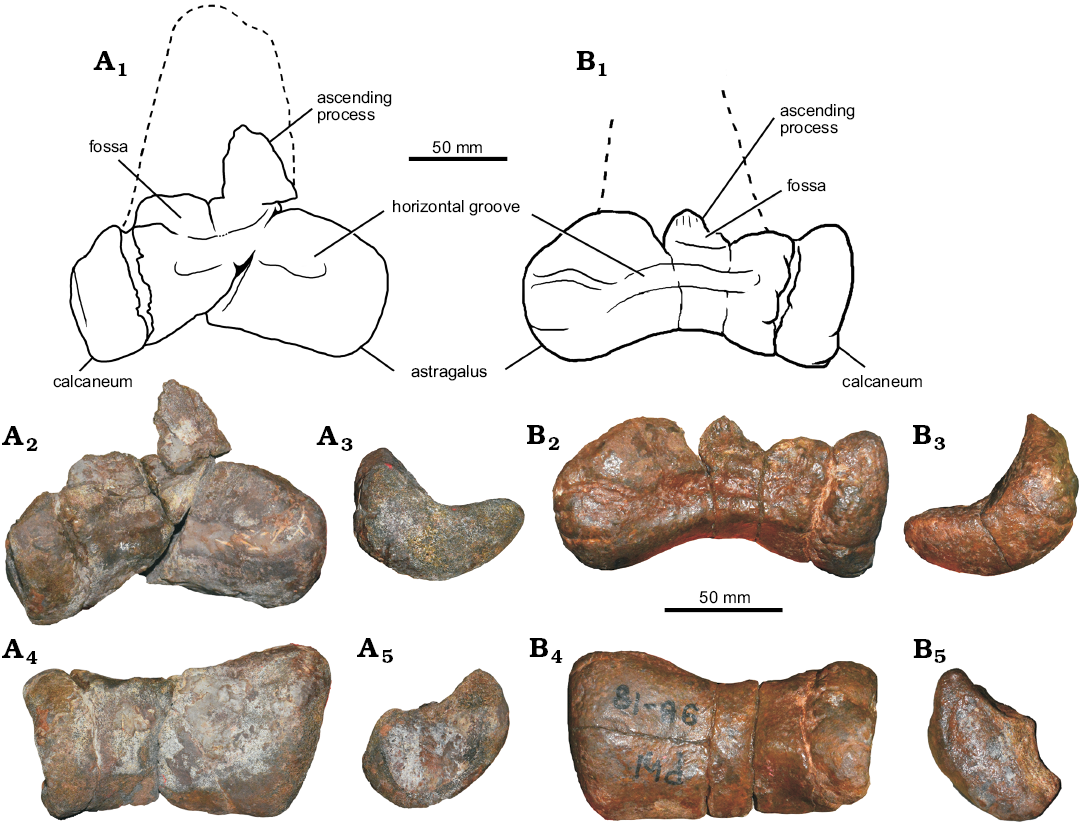
Fig. 10. Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9A-B17, right (A) and SM-PW9B-18, left (B); astragalocalcaneum in anterior (A2, B2), medial (A3, B3), distal (A4, B4), and lateral (A5, B5) views; line drawings in anterior view (A1, B1). Dashed lines are the reconstruction of the ascending processes of the astragali.
Metatarsals: The metatarsals of Phuwiangvenator comprises a left metatarsal I (SM-PW9B-1) and right metatarsals II–IV (SM-PW9B-42, 43, and 44, respectively).
The left metatarsal I (SM-PW9B-1) has well-defined collateral ligament pits which are deeper on the right side than on the left one. An extensor fossa on the dorsal surface proximal to the distal articulation is present. There is one shallow fossa on the ventral surface proximal to the distal articulation. It is proximodistally elongate. The ratio of proximodistal length to transverse width is approximately 5.2. The bone is asymmetrical and lacks a ginglymoid articulation (Fig. 11).
Fig. 11. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9B-1, left metatarsal I in lateral (A1), posterior (A2), anterior (A3), and medial (A4) views.
The metatarsal II is subequal in width to the Mt III and Mt IV proximally (see Rauhut 2003). The shape of the proximal end of the metatarsal III has a deep notch or “hourglass” shaped or “notched” outline which is most pronounced in avetheropods (Carrano et al. 2012). At the broken part, distally below the proximal end, the posterior part of the shaft of the metatarsal III is strongly pinched between the shafts of metatarsals II and IV. In proximal view, metatarsal III is anteroposteriorly oriented, not anterolateral to posteromedial. The anteroposterior orientation of the metatarsal III can be found in Australovenator, Chilantaisaurus, and Neovenator, but not in Tanycolagreus and Ornitholestes. The anterior rim of metatarsal IV slopes from proximolaterally to distomedially. This way, the distomedial corner of the proximal articular surface of metatarsal IV in anterior view is much lower than the articular surface of metatarsal III (Fig. 12).

Fig. 12. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9B-44 (A), SM-PW9B-43 (B), SM-PW9B-42 (C); metatarsals in anterior (A1–C1) and proximal (A2–C2) views; line drawings in anterior (A3–C3) and proximal (A4–C4) views; metatarsal III in medial (B5) and distal (B6) views.
Pedal phalanges and unguals: The pedal phalanges consist of phalanges II-1, II-2, III-2, III-3, IV-1, and IV4 (SM-PW9B-5–9 and SM-PW9B-45, respectively; Fig. 13). The pedal phalanges are as in normal theropods in shape.
The pedal unguals in this study consist of unguals I-2, II-3, III-4, and IV-5 (SM-PW9B-11, 22, 19, and 10, respectively). Specimens SM-PW9B-22 and SM-PW9B-19 are medium to large sized and missing the distal portion. They have a rounded cross-section that is symmetrical. SM-PW9B-10 is complete. It has flat ventral surface as in Australovenator (Fig. 14). Two vascular grooves are present on both sides. The flexor tubercle is poorly developed. It is higher dorsoventrally than broader mediolaterally and weakly curved. In general, the pedal unguals have the same shape as in other theropods. They do not show extreme dorsoventral flattening in lateral view as in Spinosaurus and some ornithomimosaurs.
Stratigraphic and geographic range.—Type locality and horizon only.
Fig. 13. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9B-11 (A), SM-PW9B-7 (B), SM-PW9B-8 (C), SM-PW9B-22 (D), SM-PW9B-5 (E), SM-PW9B-9 (F), SM-PW9B-19 (G), SM-PW9B-6 (H), SM-PW9B-45 (I), SM-PW9B-10 (J); right pedal phalanges (B, C, E, F, H, I) and unguals (A, D, G, J), in ventral (A1–J1) and dorsal (A2–J2) views; line drawings in dorsal view (A3–J3). Dashed lines are the reconstruction of pedal phalanges III-1, IV-2, and IV-3 and pedal unguals II-3 and III-4.

Fig. 14. Coelurosaurian theropod Phuwiangvenator yaemniyomi gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wiang, Khon Kaen Province, Thailand. SM-PW9B-10, right pedal ungual IV-5 in lateral (A1) and ventral (A2) views.
Genus Vayuraptor nov.
Zoobank LCID: urn:lsid:zoobank.org:act:7CCF3718-D9BE-4FE3-A 191-8754C7892CF1
Type species: Vayuraptor nongbualamphuensis gen. et sp. nov., monotypic, see below.
Etymology: From Sanskrit Vayu, God of Wind and Latin raptor, thief; “Raptor of Wind or Wind raptor” in reference to its long and slender tibia, which suggest a fast running animal.
Diagnosis.—Same as for only known species.
Vayuraptor nongbualamphuensis sp. nov.
Figs. 15–17.
Zoobank LCID: urn:lsid:zoobank.org:act:10988651-4C41-4062-A70F- 13BC6DE0D722
Etymology: From Nong Bua Lamphu Province where the specimen was recovered.
Holotype: SM-NB A1-2, left tibia with associated astragalocalcaneum (collected in August 1988 by Paladej Srisuk).
Type locality: Phu Wat Site A1 Locality, Nong Sang, Nong Bua Lamphu Province, Thailand, Fig. 2.
Type horizon: Sao Khua Formation, probably upper Barremian?, Lower Cretaceous.
Referred material.—PRC-NB A1-11, right coracoids; PRC- NB A1-4, fibula fragment; PRC-NB A1-10 rib; PRC-NB A1-3, probable pubis fragment; PRC-NB A2-20, manual phalanx, and PRC-NB A2-16, probable pedal phalanx; all from the type locality and horizon.
Diagnosis.—Vayuraptor is a basal coelurosaur and is diagnosed by the following autapomorphies: (i) astragalus has two short horizontal grooves and two foramina on the astragalar body, and two fossae at the base of the ascending process; (ii) the ascending process of the astragalus is straight laterally and straight and parallel medially at the base. In the middle of the ascending process, the medial rim slopes to the tip laterally; (iii) there is a vertical ridge starting from the tip and disappearing just above the middle of the ascending process; and (iv) extremely high and narrow ascending process of the astragalus, with a ratio of the ascending process height/ascending process width of 1.66.
Description.—General morphology: Vayuraptor was a mid to large-sized theropod with an estimated body length of 4–4.5 m. The skeletal proportions are shared by other theropods of corresponding sized such as Fukuiraptor.
Rib fragment: A rib fragment (PRC-NB A1-10) that is lacking the proximal and distal portion. There is no pneumaticity on the rib fragment.
Coracoid: The posteroventral process of the coracoid (PRC-NB A1-11) is tapering posteroventrally (the posteroventral process is broken). The ventral margin of the coracoid is expanded beyond the rim of the glenoid facet (inferred from the broken part). The coracoid is higher than long and semicircular in shape. The infraglenoid groove is absent (Fig. 15).

Fig. 15. Coelurosaurian theropod Vayuraptor nongbualamphuensis gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wat, Nong Bua Lamphu Province, Thailand. PRC-NB A1-11, right coracoid in lateral (A1) and medial (A2) views. Dashed lines are the reconstruction of the posteroventral process of the coracoid.
Phalanges: Only the distal part of manual phalanx (PRC-NB A2-20) is preserved. It lacks well-defined extensor pits on the dorsal surface proximal to the distal articulation. This was pointed out to be a coelurosaurian character by Rauhut (2003) (see Novas et al. 2016). A probable pedal phalanx (PRC-NB A2-16) is very incomplete so there is not much information to be obtained from this material.
Pubis fragment: A proximal mid-portion of a left pubis (PRC-NB A1-3) preserves a portion of the medial laminar of the pubic shaft. It looks similar to other theropods such as the middle portion of the pubis of Neovenator (see Brusatte et al. 2008).
Tibia and fibula: The left tibia (SM-NB A1-2; Fig. 16) is complete. It is long and slender. The cnemial crest is expanded. The morphology of the distal cnemial process is rounded. The anterolateral process of lateral condyle forms a horizontal projection. The tibial shaft is almost straight but the distal portion is slightly curved medially (slightly convex laterally in anterior view). The mid-shaft cross-section is sub-circular with a flattened anterior side (D-shaped cross-section at mid-shaft) as in Australovenator (Hocknull et al. 2009) and Aerosteon (cast of MCNA-PV-3139; AS personal observations) as well as other coelurosaurs. The cnemial crest is not projected proximally. The lateral condyle is small relative to the tibial shaft and offset from the lateral side of the proximal end of the tibia by a notch (posterior cleft) posteriorly. It does not reach the same level as the medial side posteriorly. The fibular crest (the fibular flange) of the tibia is present on the proximal half as a pronounced longitudinal ridge. The fibular crest is clearly offset from the proximal lateral surface of the tibia. It does not connect with the proximal end of the tibia (the ridge continuing from the fibular flange to the proximal articular surface of the tibia is absent). The ridge emerges 85 mm from the proximal end of the tibia. The fibular crest is sheet-like. The primary nutrient foramen cannot be observed, due to bad preservation.
The lateral malleolus of the distal expansion extends further distally and laterally than the medial malleolus. The lateral malleolus is overlapped by the calcaneum. The shape of the edge of lateral malleolus is a tabular notch. The distal tibia is most probably flat. The medial vertical ridge or medial bulge of the tibia anteriorly for the ascending process of the astragalus is present. This medial vertical ridge is also present in Phuwiangvenator, Australovenator, Chilantaisaurus, Aerosteon (Benson et al. 2010), and Juratyrant (Benson 2008; Rauhut 2012; Brussate and Benson 2013). The distal end of the tibia is triangular in outline, flattened anteroposteriorly, and strongly expanded laterally and medially.
The tibial length of Vayuraptor is more than 12 times its anteroposterior width at mid-length. This is a synapomorphy of megaraptorans and coelurosaurs (Benson et al. 2010) or a synapomorphy of Coelurosauria, including Megaraptora (Novas et al. 2013; Porfiri et al. 2014). In Vayuraptor this ratio is 13.9. For measurement of the tibiae see SOM: table 17. A fragment of probable fibula (PRC-NB A1-4) is preserved. It is very incomplete and does not offer much information.
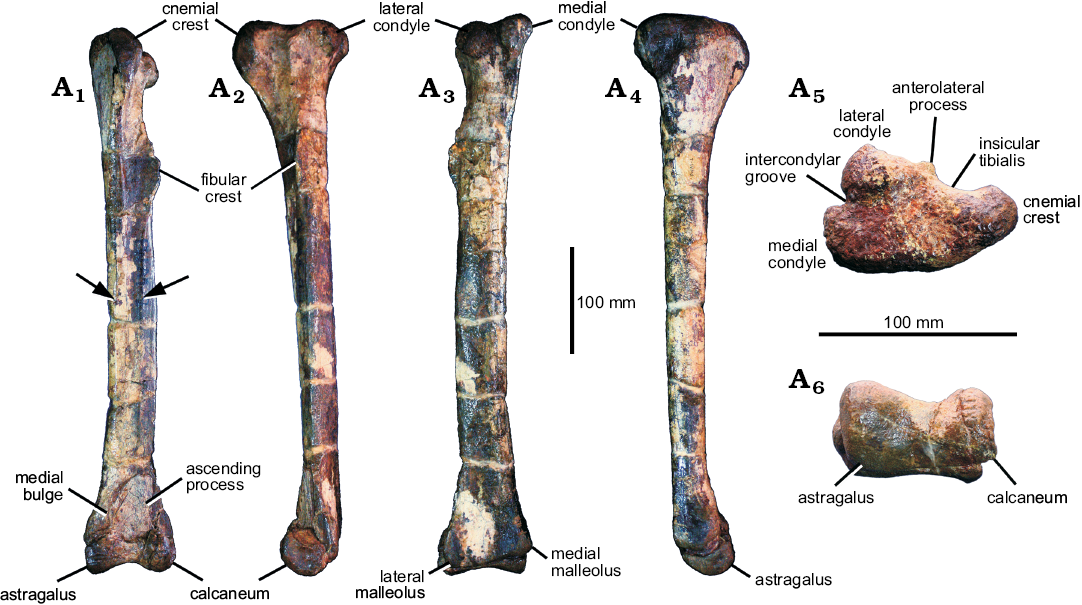
Fig. 16. Coelurosaurian theropod Vayuraptor nongbualamphuensis gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wat, Nong Bua Lamphu Province, Thailand. SM-NB A1-2, left tibia with associated astragalocalcaneum in anterior (A1), lateral (A2), posterior (A3), medial (A4), proximal (A5), and distal (A6) view. Arrow indicates vertical ridge on the tibia.
Astragalocalcaneum: The left astragalus and calcaneum (SM-NB A1-2) are well preserved, only the tip of the ascending process of the astragalus is missing (Fig. 17). The astragalus is 70 mm wide, the calcaneum is 18 mm wide. The astragalar width/calcaneum width ratio is 3.8. The ascending process of the astragalus is sheet-like, it is 70 mm long, 14% of the total length of the tibia. It is offset from the anterior border of the astragalar body by a shallow groove. Its shape is straight laterally and straight and parallel medially at the base before it slopes to the tip laterally. There is a vertical ridge on the ascending process, from the proximal end to the middle of the process. There are two horizonal grooves and two foramina at the base of the astragalar body. The distal side of the astragalus is strongly concave in anterior view. The trapezoidal outline of the bone in distal view looks similar to that of Australovenator and the megaraptoran astragalus NMV P150070 (Benson et al. 2012b). It shows an extremely high and narrow ascending process of the astragalus, with a ratio of the ascending process height/ascending process width of approximately 1.66.
The astragalar body has a concave distal surface. The ascending process of the astragalus of Fukuiraptor is 1.7 times the height of its body (Benson et al. 2010) which is the same as in Vayuraptor (= 1.7).
The ascending process of the astragalus arises more than half of the breath of the astragalus body, and it is more than twice the height of the astragalus body (7:2.5). The astragalar condyle is significantly expanded proximally on the anterior side of the tibia and faces anterodistally.
The angle of the dorsal margin of the ascending process is almost horizontal. The articulation between the ascending process and the fibula is restricted to the lateral side. The shape of the astragalar ascending process looks similar to that of Australovenator, Fukuiraptor, Aerosteon, Qianzhousaurus (Lü et al. 2014), Raptorex (Sereno et al. 2009), and Alioramus (Brusatte et al. 2012). Two fossae at the base of the ascending process are present. There is an anteroproximal (= proximolateral) expansion of the astragalar lateral condyle, as in Australovenator and Fukuiraptor (Benson et al. 2010). The development of the articular surface for the distal end of the fibula is reduced and situated on the lateral side. The posterolateral crest and posteromedial crest are absent. The calcaneum has a well-developed facet for the tibia. Both the astragalus and calcaneum are unfused. The calcaneum transverse development is moderately wide. The width of the calcaneum is approximately 25.7% of the width of the astragalus.

Fig. 17. Coelurosaurian theropod Vayuraptor nongbualamphuensis gen. et sp. nov. from the Early Cretaceous Sao Khua Formation of Phu Wat, Nong Bua Lamphu Province, Thailand. SM-NB A1-2, left astragalocalcaneum in anterior (A2), medial (A3), lateral (A4), and distal (A5) views; line drawing in anterior view (A1).
Remarks.—Atlantal intercentrum: The atlantal intercentrum of Phuwiangvenator is high in proportion and looks more similar to that of Allosaurus (UMNH VP 11289; AS personal obsevations) than to Sinraptor (Currie and Zhao 1993), Aerosteon (cast of MCNA-PV-3137; AS personal obsevations), and Orkoraptor (Novas et al. 2008) in which they are low dorsoventrally.
Dorsal vertebra: In general, the dorsal vertebra of Phuwiangvenator looks similar to that of Fukuiraptor (AS personal observations). Based on the broken bone surface, the dorsal vertebra apparently has a camellate internal structure. There are no pleurocoels, but a lateral depression and striation on the lateral and ventral side of the centrum are present as in Fukuiraptor (AS personal observations).
Sacral vertebrae: The proportion of the sacral centra of Phuwiangvenator is longer than high, the same as SMNS 58023, a megaraptoran from the Early Cretaceous of Brazil (Aranciaga Rolando et al. 2018; AS personal observations), Gualicho (Apesteguía et al. 2016), MPMA 08-003-94 (Mendez et al. 2012; AS personal observations), CPPLIP 1324 (Martinelli et al. 2013; AS personal observations), Suchomimus (MNBH GAD 500 and MNBH GAD70; AS personal observations), and Spinosaurus (FSAC-KK 11888; AS personal observations). This proportion is shorter than high in Aoniraptor (Motta et al. 2016), Datanglong (Mo et al. 2014), and Megaraptor (Porfiri et al. 2014; Aranciaga Rolando et al. 2018).
The median transverse constriction is not present in Phuwiangvenator, SMNS 58023, and Suchomimus (MNBH GAD 500 and MNBH GAD70). The constriction is present in Datanglong and Siamotyrannus.
Pleurocoels are present in SMNS 58023, Megaraptor, and Aoniraptor, but absent in Phuwiangvenator, Datanglong, and Siamotyrannus.
The flattened ventral surface of the sacrum is present in Phuwiangvenator, Aoniraptor, Falcarius (Zanno 2010), MPMA 08-003-94, CPPLIP 1324, and some derived coelurosaurs (Rauhut 2003), but convex in the SMNS 58203 (AS personal observations), Suchomimus MNBH GAD 500 (AS personal observations), Datanglong (Mo et al. 2014), and Spinosaurus.
The camellate internal structure is present in SMNS 58023, Megaraptor, Aoniraptor, and Phuwiangvenator. Generally, the vertebral pneumaticity in basal theropods is camerate, where as in ceratosaurs, carcharodontosaurs, and many coelurosaurs, a camellate internal structure is present (Benson et al. 2012a).
The sacral centra are ventrally less concave (in lateral view) in Phuwiangvenator, Megaraptor, Aoniraptor, and Datanglong, whereas they are concave in SMNS 58023, Suchomimus (MNBH GAD 500 and MNBH GAD70), and Spinosaurus (FSAC-KK 11888).
We note that the ventral groove on the sacral centra is present in some theropods such as Falcarius (Zanno 2010; AS personal observations), Neovenator (Brusatte et al. 2008), and Condorraptor (Rauhut 2005), but there is usually one longitudinal groove instead of two sulci which is longer and wider than in Phuwiangvenator (see SOM: fig. 1).
Metacarpal: An elongate metacarpal is a coelurosaurian synapomorphy (Novas et al. 2013) which is present in Phuwiangvenator. The metacarpal II of Phuwiangvenator looks similar to that of Kileskus and Falcarius, e.g., the shaft is straight, slender, and dorsoventrally flattened. The shaft of metacarpal II is slender in Phuwiangvenator, Megaraptor, Australovenator, Tanycolagreus, Coelurus, Gualicho, Kileskus, Guanlong, and Yutyrannus (Carpenter et al. 2005a, b; Xu et al. 2006, 2012; Averianov et al. 2010; Apesteguía et al. 2016; Novas et al. 2016), however, it differs from that of Megaraptor in which it is more slender and straight than in Megaraptor (Calvo et al. 2004; Novas et al. 2016). The distal end of metacarpal II is asymmetrical and divided into two condyles in Phuwiangvenator as well as in other theropods. The distal surface of the medial condyle is larger than the lateral condyle in Phuwiangvenator, Australovenator, Gualicho, and Kileskus. The medial condyle protrudes slightly more distally than the lateral condyle in Phuwiangvenator, Coelurus, and Kileskus. The pits for the collateral ligaments are not well developed on both condyles in Phuwiangvenator. There is no distinct extensor pit on the anterior surface of the shaft proximal to the distal condyle, only a small concavity is present in Phuwiangvenator and Kileskus. The shaft is straight (in dorsal view) in Phuwiangvenator, Australovenator, Tanycolagreus, Coelurus, Gualicho, Kileskus, and Guanlong.
Manual phalanges: Manual phalanx I-1 of Phuwiangvenator is concave ventrally in proximal view. This character is present in Australovenator, Megaraptor, NMV P199050 (Benson et al. 2012b), Suchomimus (MNBH GAD 500; AS personal observations), Kileskus (Averianov et al. 2010), Falcarius (Zanno 2006), and Deinocheirus (Osmólska and Roniewicz 1970).
A deep and wide furrow along the ventral surface of the manual phalanx I-1 is present in Megaraptor and Australovenator (Novas et al. 2016). A longitudinal ventral furrow and longitudinal ridges are present in Phuwiangvenator. This furrow is also present in Suchomimus and Spinosaurus, but it is restricted to the proximal part. However, the manual phalanx I-1 of Phuwiangvenator exhibits a triangular contour at the proximal end, not a square-shaped contour as present in the megaraptorids Megaraptor and Australovenator (see Novas et al. 2016).
Manual unguals: The manual ungual I-2 of Phuwiangvenator is strongly transversely compressed, being oval with a distinct dorsoventral axis in proximal view (see Novas 1998; Porfiri et al. 2014). This character is also found in Fukuiraptor, Megaraptor, Australovenator, and Chilantaisaurus. However, the manual ungual I-2 of Phuwiangvenator, in relation to its phalanx, is proportionally smaller than in Megaraptor and Australovenator (Calvo et al. 2004; White et al. 2012; Novas et al. 2016). The manual ungual I-2 of Phuwiangvenator has a curved flexor tubercle which is found in Fukuiraptor (FPDM-V43-11; AS personal observations) and Chilantaisaurus (Benson and Xu 2008), but this character is widely present among Ornithodira. The ratio of the proximal height/width of the manual ungual I-2 of Phuwiangvenator is 2.4, compared with 1.95 in Torvosaurus (Benson et al. 2010), 1.78 in Suchomimus (MNBH GAD 500; AS personal observations), 1.9 in Baryonyx (cast of BMNH R9951; AS personal observations), 1.9 in Allosaurus (UMNH VP5676; AS personal observations), 2.7 in Chilantaisaurus (Benson and Xu 2008), 2.7 in Fukuiraptor (FPDM-V43-11; AS personal observations), 2.4 in Australovenator (Benson et al. 2010), and 2.75 in Megaraptor (Benson et al. 2010). The manual ungual III-4 of Phuwiangvenator is small and much smaller than the manual unguals I-2 and II-3 as in other theropods. We note that the character of the manual ungual I-2 with an asymmetrical lateral groove and well-developed ventral ridge of Megaraptor and Australovenator (White et al. 2012; Novas et al. 2016) is not present in Phuwiangvenator and Fukuiraptor (FPDM-V43-11; AS personal observations).
Tibia: In proximal view, the lateral condyle of the tibia is level with the posterior margin of the medial condyle. This character is found in Phuwiangvenator, Vayuraptor, Australovenator (White et al. 2013a), Fukuiraptor (AS personal observations), Orkoraptor (Novas et al. 2008), Neovenator (Brusatte et al. 2008), Tyrannosaurus (Brochu 2003), Falcarius (AS personal observations), Gualicho (Apestiguea et al. 2016), and Murusraptor (Coria and Currie 2016), but not in Suchomimus (MNBH GAD 500, MNBH GAD 98, and MNBH GAD 72). In Torvosaurus (Britt 1991), Piatznitskysaurus, and Spinosaurus this character is independently evolved.
The character: “tibial lateral condyle of proximal end curves ventrally as a pointed process”; this character is present in Phuwiangvenator, Megaraptora, some coelurosaurs, and Neovenator, but not in Fukuiraptor and Vayuraptor.
The median prominence in the anterior surface of the distal end of the tibia (tibial vertical medial ridge or medial bulge) is present in Phuwiangvenator, Vayuraptor, Australovenator (White et al. 2013a), Suchomimus (MNBH GAD 500; MNBH GAD 97; and MNBH GAD 72; AS personal observations), Coelurus (Carpenter et al. 2005b), Chilantaisaurus (Benson and Xu 2008), Falcarius (Zanno 2010; AS personal observations), Spinosaurus (cast of FSAC-KK 11888; AS personal observations), Chuandongocoelurus (Rauhut 2012), Juratyrant (Benson 2008; Rauhut 2012; Brussate and Benson 2013). Masiakasaurus (Carrano et al. 2002), theropod indet. MB.R.2351 (AS personal observations), the abelisauroid MB.R.1750 (AS personal observations), the abelisauroid MB.R.1751 (AS personal observations), Aniksosaurus (Martinez and Novas 2006), possibly Bicentenaria (Novas et al. 2012b), Aerosteon, Stokesosaurus (Rauhut 2012), Ozraptor (Long and Molnar 1998), Tanycolagreus (Carpenter et al. 2005a), Segnosaurus (Zanno 2010), Erliansaurus (Xu et al. 2002), possibly NMV P150070 (Benson et al. 2012b), and possibly Fukuiraptor (AS personal observations). The posteroventral ridge of the tibia is present in Vayuraptor, Australovenator, and Tachiraptor (Langer et al. 2014). The tibia facet for the reception of the ascending process of the astragalus at the distal end which is flat anteriorly is an apomorphy shared by megaraptorans and coelurosaurians (Porfiri et al. 2014).
The character: “tibia anteriorly flat at mid-length with vertical ridges anterolaterally and anteromedially” can be found in Phuwiangvenator, Vayuraptor, Neovenator (Brusatte et al. 2008), Australovenator (White et al. 2013a), and Aerosteron (cast of MCNA-PV-3139; AS personal observations).
The character: “tibia anteriorly flat or slightly flat but no such vertical ridges” is present in Spinosaurus (cast of FSAC-KK 11888; AS personal observations), Chilantaisaurus (Benson and Xu 2008), a sinraptorid tibia from Thailand (SM 10; AS personal observations), Murusraptor (Coria and Currie 2016), and Acrocanthosaurus (Stovall and Langston 1950; see SOM: table 18).
The character: “tibia anteriorly convex or slightly convex but ridge present” can be found in Allosaurus (UMNH VP 6402, 7145, 7926, 7936, 7939) and Fukuiraptor (FPDM- V43-20).
The tibia is long and slender in the Thai theropods. The length/width ratio is more than 12 in Phuwiangvenator (13.82), Vayuraptor (13.9), Australovenator (12.53), Fukuiraptor (approximately 14.3–14.5), Aerosteon referred material (14), and Spinosaurus neotype (12.09) (Ibrahim et al. 2014) compared with Murusraptor (11.9), the Suchomimus holotype (MNBH GAD 500, 9.6), Suchomimus “subadult” (MNBH GAD97, 10.86), and Suchomimus “juvenile” (MNBH GAD72, 11.67) (see SOM: table 17).
Astragalus: The ascending process height of astragalus being more than 0.5 times the width of the astragalar body can be found in Vayuraptor, Fukuiraptor, Australovenator, possibly in Phuwiangvenator, and all other coelurosaurs. The transverse width of the ascending process of astragalus occupies the total width of the anterior surface of distal tibia (coelurosaurian character), distinct anterior development of the lateral condyle of the astragalar body and a strong inflection in the anterior margin of the astragalar body in distal view (Novas et al. 2013) (coelurosaurian characters) are present in Phuwiangvenator, Vayuraptor, Australovenator, Aerosteon, and other coelurosaurs. The presence of an astragalus with a prominent proximolateral extension is a megaraptoran synapomorphy (Novas et al. 2013). This latter feature is observed in Vayuraptor and possibly Phuwiangvenator (in the latter it is small and damaged). Transverse development of the calcaneum which is moderately wide (plesiomorphic character) is present in Phuwiangvenator and Vayuraptor. The shape of the calcaneum in lateral or medial view is strongly asymmetric, with a right angle at the posterior border (Novas et al. 2013) (coelurosaurian character). This character possibly unites the Megaraptora within Coelurosauria.
The character: “ascending process of the astragalus which is offset from distal condyles by a pronounced groove (possible coelurosaurian character)” is present in Vayuraptor and Phuwiangvenator.
The ratio of the ascending process height/astragalar body height is high in Vayuraptor (1.7), Fukuiraptor (1.7), Aerosteron (1.9), Alioramus (2.5), and Phuwiangvenator (estimated 1.73). This ratio is lower in NMV P150070 (1.43), Australovenator (1.4), Falcarius (1.3), Suchomimus (MNBH GAD97, 1.3; MNBH GAD98, 1.25), compared with Allosaurus (1.14) which has the much lower ratio (see SOM: table 19). This ratio is also low in Coelurus, Bicentenaria, and Tugulusaurus.
The ascending process of astragalus arises from the lateral side of astragalar body (not from its complete width) in Phuwiangvenator, Vayuraptor, Australovenator, Fukuiraptor, Tugurusaurus, Bicentenaria, and Coelurus. It arises from the complete width of the astragalar body in Tanycolagreus and Aerosteon. The ratio of the ascending process width at base/astragalar body width is low and the ascending process is restricted to the lateral side of the astragalar body in basal tetanurans (e.g., Novas et al. 2013) and some basal coelurosaurs. This ratio is 50% in Allosaurus, Bicentenaria (Novas et al. 2012b), Tugulusaurus (Rauhut and Xu 2005), and Coelurus (Carpenter et al. 2005b), 55% in NMV P150070, 56% in Suchomimus (MNBH GAD97), 63% in Vayuraptor, unknown in Fukuiraptor, 65% in Phuwiangvenator, 66% in Falcarius, 70% in Australovenator, 78% in Alioramus, and 83% in Aerosteon (see SOM: table 19).
The ratio of the astragalar ascending process height/ascending process width at base is high in Vayuraptor (approximately 1.66), compared with Alioramus (1.4), Phuwiangvenator (estimated 1.39), NMV P150070 (1.3), Fukuiraptor (1.1), Falcarius (1.1), Australovenator (1), Aerosteon (1), Allosaurus (UMNH VP11003, 1), and Suchomimus (MNBH GAD97, 0.88; MNBH GAD98, 0.8).
An anterior development of the lateral condyle of the astragalus in distal view is present in Phuwiangvenator, Vayuraptor, Australovenator, Aerosteon, probable Fukuiraptor, NMV P150070, Appalachiosaurus (Carr et al. 2005), Bicentenaria (Novas et al. 2012b), Coelurus, Tugulusaurus, and Alioramus. The accessory posterolateral ascending process of the astragalus is present in Fukuiraptor and NMV P150070 (see Agnolin et al. 2010; see SOM: fig. 2). The prominent proximolateral extension of the astragalus (Benson et al. 2010; Carrano et al. 2012; Porfiri et al. 2014) that projects from the anterior articular facet as a rounded triangular process is present in Australovenator, Aerosteon, Fukuiraptor (small process), Phuwiangvenator (small process), and Vayuraptor. This process forms the anterior border of the fibular facet of the astragalus and the fibular facet is shallow and bordered posteriorly by the lateral margin of the ascending process (Hocknull et al. 2009). In Fukuiraptor, the astragalus has a shallow notch for a process from the calcaneum but it is not well developed (Azuma and Currie 2000; AS personal observations). The astragalar notch for a process from the calcaneum is present in Aerosteon, Phuwiangvenator, but not Vayuraptor. The astragalus has a distinct socket that embraced the mediodistal end of the fibula (Azuma and Corrie 2000; AS personal observations), this present in Fukuiraptor, Phuwiangvenator, and Vayuraptor. The fibular contact extending up the anterolateral margin of the ascending process of the astragalus is present in Fukuiraptor (Azuma and Currie 2000; AS personal observations), Vayuraptor, and possibly Phuwiangvenator.
The long and sheet-liked ascending process of the astragalus with a slope or convex medially and straight laterally can be found in Vayuraptor, Australovenator, Fukuiraptor, Qianzhousaurus (Lü et al. 2014), Raptorex (Sereno et al. 2009), and Appalachiosaurus (Carr et al. 2005). However, the shape of the ascending process of Vayuraptor is different from the other theropods mentioned above. The ascending process with vertical ridge proximally which ends in the middle is present in Vayuraptor and Qianzhousaurus (Lü et al. 2014). Two fossae at the base of the ascending process of the astragalus can be found in Vayuraptor and Appalachiosaurus (Carr et al. 2005). One fossa is present in Qianzhousaurus, Alioramus, Raptorex, and other tyrannosauroids and ornithomimosaurs (e.g., White et al. 2013a; Lü et al. 2014; Sereno et al. 2009; Brusatte et al. 2012; Choiniere et al. 2012), whereas there is no fossa in Coelurus, Tugurusaurus, and Bicentenaria. There is a cranio-proximal process at the base of the astragalus in Vayuraptor, Australovenator, Aerosteon, and Coelurus, small in Fukuiraptor and Phuwiangvenator.
Calcaneum: In Phuwiangvenator, the facet for the fibula of the calcaneum is small and smaller than in the allosauroid Allosaurus (UMNH VP 9965). The area for astragalocalcaneum attachment in Allosaurus is small and restricted to the anteroventral part, clearly different from Phuwiangvenator. The calcaneum of Phuwiangvenator is wider anteroposteriorly than high dorsoventrally. This is in contrast to the tyrannosauroid Appalachiosaurus (Carr et al. 2005). The facet for the fibula of the calcaneum is larger and not restricted to the dorsal part as in Appalachiosaurus. In Phuwiangvenator and Vayuraptor, the calcaneum is thick and differed from Falcarius (UMNH VP 12365), which has a disk-like calcaneum.
The calcaneum of Phuwiangvenator looks more similar to that of Murusraptor than to Baryonyx and Allosaurus in the shape and ratio of the fibular facet of the calcaneum. In Baryonyx and Allosaurus, there are large fibular facets dorsal to the calcaneum, the tibia facets is situated on the medial side. In Phuwiangvenator and Murusraptor, lateral view, the fibular facet is almost the same size as the tibia facet, and the tibia facet is situated on the mediodorsal side of the calcaneum. In the evolution from basal theropods to moderately derived coelurosaurs, there is a trend of enlargement of the tibial facet of the calcaneum and a concurrent reduction of the fibular facet. As a result, the tibial facet is much larger than the fibular facet in moderately derived coelurosaurs. The calcaneum is relatively wide transversely in Phuwiangvenator, Vayuraptor, Murusraptor, Aerosteon, and Tugulusaurus. It is strongly compressed in Tanycolagreus, Alioramus, and derived coelurosaurs. The calcaneum is roughly symmetrical with a wide angle on the posterior border in Murusraptor and Aerosteon. It is asymmetrical with a right angle on the posterior border in Phuwiangvenator, Vayuraptor, and Tanycolagreus (Novas et al. 2013). The ratio of the width of the calcaneum/the width of the astragalus is 25.7% in Vayuraptor, 24% (left) and 23% (right) in Phuwiangvenator, and whereas the ratio of the width of the calcaneum/astragalus is less than 20% in coelurosaurs (Rauhut and Pol 2017).
Metatarsals: In proximal view, the lateral margin of metatarsal II is straight in Australovenator, Fukuiraptor (White et al. 2013a), and Tanycolagreus (Carpenter et al. 2005a), but the lateral margin is convex or slightly convex in Phuwiangvenator, Ornitholestes, and Allosaurus.
In proximal view, the metatarsal III is slightly concave medially in Phuwiangvenator, Torvosaurus, Ornitholestes, and possibly Fukuiraptor. It is straight in Australovenator, Afrovenator, possibly Neovenator, Tanycolagreus, Mapusaurus, and almost straight in Zuolong and Allosaurus (UMNH VP 16038). It is concave in Chilantaisaurus and Sinraptor.
In the proximal view, the shape of the medial margin of metatarsal IV is sigmoidal in Phuwiangvenator, Mapusaurus, Chilantaisaurus, Allosaurus, and Tanycolagreus. It is step-like in Australovenator, Megaraptor, Ornitholestes, and Coelurus, whereas there is a convex medial margin in Zuolong. The shape of the posterolateral margin of metatarsal IV is slightly concave in Phuwiangvenator, Australovenator, and Zuolong, whereas it is almost straight in Megaraptor, Mapusaurus, Chilantaisaurus, and Ornitholestes.
In Phuwiangvenator, metatarsal III is anteriorposteriorly oriented in proximal view, which is similar to Australovenator (White et al. 2013a), Chilantaisaurus (Benson and Xu 2008), Neovenator (Brusatte et al. 2008), Falcarius (Zanno 2010), Garudimimus (Kobayashi and Barsbold 2005), and Acrocanthosaurus (Stovall and Langston 1950; Currie and Carpenter 2000), but differs from Suchomimus (MNBH GAD70; AS personal observations), Sinraptor (cast of holotype in FPDM; AS personal observations), Tanycolagreus (Carpenter et al. 2005a), Allosaurus (UMNH VP 9892, 16038, 9877; AS personal observations), Majungasaurus (Carrano 2007), Afrovenator (Sereno et al. 1994), and Dilophosaurus (Welles 1984) in which the metatarsal III is anterolaterally-posteromedially oriented in proximal view.
In Phuwiangvenator, the low anterior rim of the metatarsal IV, which is its autapomorphy, is independently present in Pandoravenator (a basal tetanuran from the Late Jurassic of Argentina, Rauhut and Pol 2017), but in Pandoravenator the anterior rim of the metatarsal IV, which is much lower than metatarsal III, does not slope proximolaterally to distomedially.
Pedal and ungual phalanges: The pedal phalanges of Phuwiangvenator are clearly different from Suchomimus (MNBH GAD70) and Spinosaurus (FSAC-KK 11888) because in spinosaurids, their pedal phalanges are strongly dorsoventrally flattened. Ventrally flattened unguals are present in Phuwiangvenator, Australovenator, Spinosaurus, and ornithomimosaurs. However, the unguals of Spinosaurus are extremely dorsoventrally flattened and less curved than in other theropods (FSAC-KK 11888, MSNM V6897; see Maganuco and Dal Sasso 2018).
Phylogenetic analyses
The analytical method and data matrix of Apesteguía et al. (2016) with additional information for the Thai theropods was used (see Material and methods). The phylogenetic analyses were done in the TNT program (Goloboff et al. 2008) with 46 taxa (excluded Santanaraptor) and 288 characters. The first analysis recovered five most parsimonious trees (MPTs) of 923 steps with a consistency index (CI) = 0.380 and a retention index (RI) = 0.656. The second analysis with 45 taxa (excluded Santanaraptor and Chilantaisaurus) and 288 characters recovered seven MPTs of 913 steps with a CI = 0.384 and a RI = 0.662 (Fig. 18).
Synapomorphies recovered in the present analyses include:
Coelurosauria: tibia length is more than 12 times its width (character 186); astragalus ascending process height is more than 0.5 times the width of the astragalar body (character 195); astragalus ascending process occupying total width of anterior surface of distal tibia (character 196); distinct anterior development of the lateral condyle of the astragalar body in distal view is present (character 197); fibular facet of astragalus reduced (character 199); calcaneum strongly asymmetric in lateral or medial view (character 201); distal end of tibia is anteriorly flat (character 208); long and gracile metatarsal III (character 209); ascending process of astragalus is offset from distal condyle by a pronounced groove (character 211); tibia lateral condyle position is shorter than the medial condyle in proximal view (character 221); tibia lateral condyle curves ventrally as pointed process (character 246).
Megaraptora: manual phalanx I-1 with a ventral groove (character 138); strongly transversely compressed manual ungual I-2 (character 141); median prominence in the anterior surface of the tibia distal end (character 188); astragalus with proximolateral extension (character 198); sacrum ventrally flattened (character 214).
Megaraptoridae: manual ungual I lateral groove asymmetrical (character 144); manual ungual I and II with the well-developed ventral ridge (character 145). These two megaraptorid characters are not present in Phuwiangvenator.
The analyses recovered Phuwiangvenator and Vayuraptor within basal Coelurosauria, and as a non-megaraptorid Megaraptora for Phuwiangvenator.
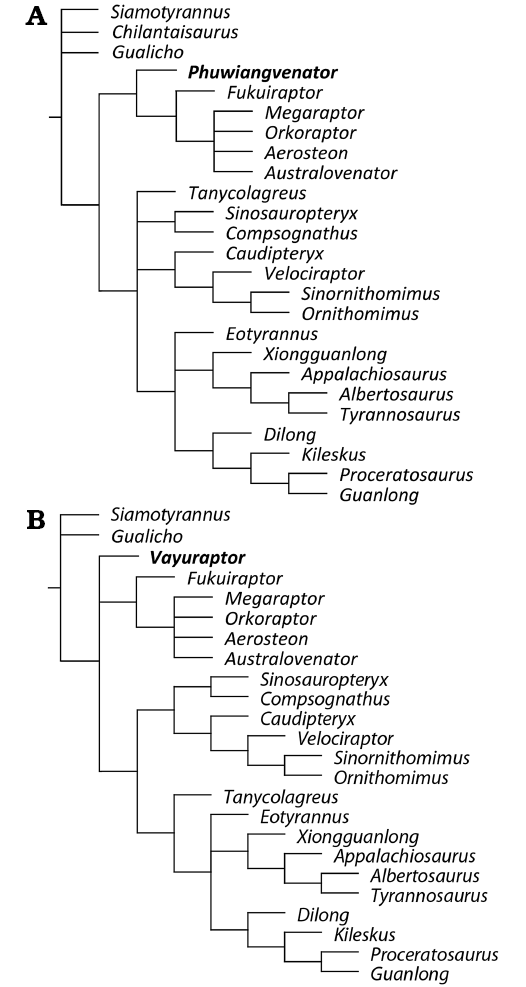
Fig. 18. Strict consensus trees of all the recovered MPTs of the phylogenetic analyses from the present study (modified from Apesteguía et al. 2016). A. Data matrix including Phuwiangvenator yaemniyomi gen. et sp. nov. B. Data matrix including Vayuraptor nongbualamphuensis gen. et sp. nov.
Discussion
Systematic and taxonomic analysis.—Phuwiangvenator yaemniyomi gen. et sp. nov. and Vayuraptor nongbualamphuensis gen. et sp. nov. show the following avetheropod and coelurosaurian characters listed below: (1) the trapezoidal or wedge-shaped, plantar surface pinched of the third metatarsal which is absent in megalosauroids and was suggested to be a possible avetheropod (= neotetanuran) synapomorphy by Benson and Xu (2008) and Holtz et al. (2004) (see Carrano et al. 2012). In fact, this character is present in Suchomimus (unknown in Vayuraptor; AS personal observations); (2) the tibia length is more than 12 times its anteroposterior width at mid-length. This is a synapomorphy of Coelurosauria, including Megaraptora (Novas et al. 2013; Porfiri et al. 2014), but can also be found in the tibia of the Spinosaurus neotype (Ibrahim et al. 2014; AS personal observations).
The following characters that place Phuwiangvenator and Vayuraptor within Coelurosauria, and Phuwiangvenator probably within Megaraptora are based on the present phylogenetic analyses (Fig. 17) and marked by an asterisk.
Phuwiangvenator yaemniyomi is identified as Coelurosauria and non-megaraptorid Megaraptora by the following characters: (1) sacrum ventral surface is flattened/without ventral keel* (Megaraptora synapomorphy) (Rauhut 2003); (2) manual phalanx I-1 is long and slender and has a furrow ventrally* (Megaraptora synapomorphy) (Novas et al. 2016); (3) the ventral margin of the proximal articular surface of the manual phalanx I-1 is concave (Novas et al. 2016); (4) manual ungual I-2 is strongly transversely compressed, being oval with a distinct dorsoventral axis in proximal view* (Megaraptora synapomorphy) (Novas 1998); (5) tibia lateral condyle position in the proximal end at its posterior margin is placed well posteriorly to the level of the posterior edge of the medial condyle* (Novas et al. 2013); (6) tibia lateral condyle of the proximal end curves ventrally as a pointed process* (Benson et al. 2010); (7) tibial median prominence in the anterior surface of the distal end is present* (Novas et al. 2013); (8) tibia length is more than 12 times its anteroposterior width at mid-length* (Coelurosauria synapomorphy) (Novas et al. 2013); (9) tibia flattened anteriorly at mid-length, usually with vertical ridges anterolaterally and anteromedially; (10) tibia facet for the reception of the ascending process of the astragalus at the distal end is more or less flat* (Coelurosauria synapomorphy) (Rauhut 2003; Benson et al. 2010; Novas et al. 2013); (11) the height of the ascending process of the astragalus is more than 0.5 times the width of the astragalar body* (Harris, 1998; Novas et al., 2013); (12) transverse width of ascending process of astragalus is occupying total width of anterior surface of distal tibia* (Rauhut 2003); (13) distinct anterior development of the lateral condyle of the astragalar body is present, strong inflexion of the anterior margin of the astragalar body in distal view* (Coelurosauria synapomorphy) (Novas et al. 2013); (14) astragalus with an anteroproximal extension is present* (Megaraptora synapomorphy) (Benson et al. 2010); (15) atragalus ascending process is offset from distal condyles by a pronounced groove* (Coelurosauria synapomorphy) (Holtz et al. 2004); (16) calcaneum, shape in lateral or medial view is strongly asymmetric, with a right angle at the posterior border* (Coelurosauria synapomorphy) (Novas et al. 2013); and (17) the pedal ungual is flattened ventrally (shared with Australovenator).
Vayuraptor nongbualamphuensis is identified as Coelurosauria by the following characters: (1) tibia lateral condyle position in the proximal end at its posterior margin is placed well posteriorly to the level of the posterior edge of the medial condyle* (Novas et al. 2013); (2) tibial median prominence in the anterior surface of the distal end is present* (Novas et al. 2013); (3) tibia length is more than 12 times its anteroposterior width at mid-length* (Novas et al. 2013); (4) tibia is flattened anteriorly at mid-length, usually with vertical ridges anterolaterally and anteromedially; (5) tibia facet for the reception of the ascending process of the astragalus at the distal end is more or less flat* (Rauhut 2003; Benson et al. 2010; Novas et al. 2013); (6) astragalus with a prominent anteroproximal extension is present* (Benson et al. 2010); (7) astragalus distinct anterior development of the lateral condyle of the astragalar body is present and there is a strong inflexion in the anterior margin of the astragalar body in distal view* (Novas et al. 2013); (8) astragalus ascending process is offset from distal condyles by a pronounced groove* (Holtz et al. 2004); (9) transverse width of the ascending process of the astragalus is occupying total width of anterior surface of distal tibia* (Rauhut 2003); (10) the height of the ascending process of the astragalus is more than 0.5 times the width of the astragalar body* (Harris 1998; Novas et al. 2013); (11) fibular facet of the ascending process of the astragalus reduced and laterally oriented (Novas et al. 2013); (12) calcaneum, shape in lateral or medial view is strongly asymmetric, with a right angle on the posterior border* (Novas et al. 2013).
Phuwiangvenator vs. Siamotyrannus: why are they not the same taxon?—In Phuwiangvenator, the dorsal vertebrae are longer than high, contrary to the dorsal vertebrae of Siamotyrannus which higher than long. The sacral vertebrae of Phuwiangvenator are also longer than high. Their centra are ventrally flattened and not constricted in ventral view. There is no median transverse constriction on the sacrum, in contrast to Siamotyrannus, where the sacral vertebrae are higher than long. The centra of this taxon are ventrally convex and also have a median transverse constriction. The ventral rims of the sacral centra of Siamotyrannus are much more convex in lateral view than those of Phuwiangvenator. Phuwiangvenator also differs from Siamotyrannus in being a smaller animal.
Vayuraptor vs. Phuwiangvenator: why are they not the same taxon?—These two new theropods belong to the basal Coelurosauria, based on their morphologies as described above. Because the fossils were recovered from the same rock formation within approximately 40 km of each other, careful comparison is needed to determine whether they belong to the same taxon or not. The overlapping materials of these two theropods are the tibia and astragalocalcaneum.
Phuwiangvenator is larger than Vayuraptor (tibial length 615 vs. 515 mm, respectively). Although the cranioproximal process of the astragalus is present in Vayuraptor and Phuwiangvenator, it is more prominent in Vayuraptor than in Phuwiangvenator. Two short horizontal grooves on the astragalar body are present in Vayuraptor, whereas only one horizontal groove is present in Phuwiangvenator. There are two fossae at the base of the ascending process of the astragalus in Vayuraptor whereas there is only one fossa in Phuwiangvenator. The calcaneum of Vayuraptor is narrower mediolaterally proximally than that of Phuwiangvenator in anterior view. A shallow notch of the astragalus for the calcaneum process is present in Phuwiangvenator. This notch is not present in Vayuraptor. The character “tibia anterolateral process of the lateral condyle curves as a pointed process” is present in Phuwiangvenator, not in Vayuraptor. This character has been compared with various ontogenetic stages of Allosaurus (UMNH VP 7148, 7922, 7932, 7938, 7939, 7940), and this process was found in all observed specimens (AS personal observations), so we conclude that the differences between Phuwiangvenator and Vayuraptor are not due to ontogeny. The shape of the fibular crest is different as well (e.g., the fibular crest is expanded proximolaterally in Phuwiangvenator vs. being expanded distolaterally in Vayuraptor). The shape of the proximal end of the tibia is different, the proximal articular surface of the medial condyle being more convex in Phuwiangvenator than Vayuraptor.
Vayuraptor vs. Siamotyrannus: why are they not the same taxon?—Vayuraptor and Siamotyrannus are represented by skeletally mature animals based on the fusion of their bones (e.g., astragalocalcaneum in Vayuraptor, and pelvis, dorsal and caudal vertebrae in Siamotyrannus). There is no overlapping material, so anatomical comparison is impossible. Vayuraptor is found here to be a basal coelurosaur. Siamotyrannus is an avetheropod, possibly a basal coelurosaur (Samathi 2013; Samathi and Chanthasit 2017; present work) or basal allosauroid (e.g., Rauhut 2003; Carrano et al. 2012). Vayuraptor differs from Siamotyrannus in being a much smaller animal. Vayuraptor is approximately 4–4.5 m in length (similar to Fukuiraptor), whereas Siamotyrannus is approximately 6.5–7 m in length (Buffetaut et al. 1996). Based on the above information, it is reasonable to conclude that Vayuraptor represents a different taxon from Siamotyrannus.
Conclusions
The present study describes the anatomy of the two new basal coelurosaurs from Southeast Asia in detail. One of them, Phuwiangvenator, is here referred to the Megaraptora. These findings extend the diversity of basal megaraptorans and support the origin of Megaraptora in Asia. The Asian megaraptorans might be a monophyletic clade or a paraphyletic series relative to the Megaraptoridae. Recently, Early Cretaceous fossils of megaraptorans have been recovered from the Barremian to Aptian of Asia. Several materials have been reported from the Aptian to mid-Cretaceous of Australia, whereas there is only one report from the Albian of South America. These show a high diversity and wide distribution during the Early Cretaceous which became more provincial in the Late Cretaceous. However, the phylogenetic position of the basal coelurosaur Vayuraptor must await further discovery of additional material and remains open to future analyses.
Acknowledgements
We would like to thank Varavudh Suteethorn and Suravech Suteethorn (both PRC) for general discussion, help, and support. Sakchai Juan-Ngam, Kamonlak Wongko, Paladej Srisuk, Preecha Sainongkham, and all staff of Sirindhorn Museum, Phu Wiang Fossil Research Center and Dinosaur Museum, and Paleontological Research and Education Center (Maha Sarakham University, Thailand) for hospitality and help during visits to the museums and the localities. Thanks to the Thai-French team and all those who took part in the excavation and preparation of the specimens. Figs. 1, 2, 4A3, 9, 10A1, 12A3, A4, B3, B4, C3, C4, and 17A1 were skillfully drawn by Wijanee Sendang (Milano, Italy) and are deeply appreciated. Thanks to Roger B.J. Benson (University of Oxford, UK) for information on theropod anatomy. Thanks to Carrie Levitt-Bussian (Natural History Museum of Utah, USA), Cristiano Dal Sasso (MSNM), Daniela Schwarz and Thomas Schossleitner (both Museum für Naturkunde, Berlin, Germany), Yoichi Azuma and Masateru Shibata (both FPDM), Paul Sereno (University of Chicago, USA), Octavio Mateus (ML, Portugal), Rainer Schoch (SMNS), and the directors and staff of MPMA and CPPLIP for access to specimens under their care. Special thanks go to the editor and reviewers, Stephen Brusatte (University of Edinburgh, UK), Thomas Holtz Jr. (University of Maryland, USA), and Fernando Novas (Museo Argentino de Ciencias Naturales, Buenos Aires, Argentina), for constructive comments that improved the quality of the manuscript. This work was done during the tenure of AS on a scholarship to study in Germany from the Ministry of Science and Technology, the Royal Thai Government (Bangkok, Thailand) and was partly supported by the Department of Mineral Resources, the Royal Thai Government via the Sirindhorn Museum (Kalasin Province, Thailand) and the Phu Wiang Fossil Research Center and Dinosaur Museum (Khon Kaen Province, Thailand). The directors of the department and the museums are greatly appreciated for permission and support for the project. Traveling was partly supported by IPID4all-DAAD 2016, 2017, the Patterson Memorial Grant SVP 2017, and AG Sander, Uni Bonn.
References
Agnolin, F.L., Ezcurra, M.D., Pais, D. F., and Salisbury, S. W. 2010. A reappraisal of the Cretaceous non-avian dinosaur faunas from Australia and New Zealand: Evidence for their Gondwanan affinities. Journal of Systematic Palaeontology 8: 257–300. Crossref
Apesteguía, S., Smith, N.D., Juárez Valieri, R., and Makovicky, P.J. 2016. An unusual new theropod with a didactyl manus from the Upper Cretaceous of Patagonia, Argentina. PLoS ONE 11(7): e0157793. Crossref
Aranciaga Rolando, A.M., Egli, F.B., Sales, M.A., Martinelli, A.G., Canale, J.I., and Ezcurra, M.D. 2018. A supposed Gondwanan oviraptorosaur from the Albian of Brazil represents the oldest South American megaraptoran. Cretaceous Research 84: 107–119. Crossref
Averianov, A.O., Krasnolutskii, S.A., and Ivantsov, S.V. 2010. A new basal coelurosaur (Dinosauria: Theropoda) from the Middle Jurassic of Siberia. Proceedings of the Zoological Institute RAS 314 (1): 42–57.
Azuma, Y. and Currie, J.P. 2000. A new carnosaur (Dinosauria: Theropoda) from the Lower Cretaceous of Japan. Canadian Journal of Earth Sciences 37: 1735–1753. Crossref
Bell, P.R., Cau, A., Fanti, F., and Smith, E.T. 2015. A large-clawed theropod (Dinosauria: Tetanurae) from the Lower Cretaceous of Australia and the Gondwanan origin of megaraptorid theropods. Gondwana Research 36: 473–487. Crossref
Benson, R.B.J. 2008. New information on Stokesosaurus, a tyrannosauroid (Dinosauria: Theropoda) from North America and the United Kingdom. Journal of Vertebrate Paleontology 28: 732–750. Crossref
Benson, R.B.J. 2010. A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods. Zoological Journal of the Linnean Society 158: 882–935. Crossref
Benson, R.B.J. and Xu, X. 2008. The anatomy and systematic position of the theropod dinosaur Chilantaisaurus tashuikouensis Hu, 1964 from the Early Cretaceous of Alanshan, People’s Republic of China. Geological Magazine 145: 778–789. Crossref
Benson, R.B.J., Butler, R.J., Carrano, M.T., and O’Connor, P.M. 2012a. Air-filled postcranial bones in theropod dinosaurs: physiological implications and the “reptile”-bird transition. Biological Reviews 87: 168–193. Crossref
Benson, R.B.J., Carrano, M.T., and Brusatte, S.L. 2010. A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic. Naturwissenschaften 97: 7–78. Crossref
Benson, R.B.J., Rich, T.H., Vickers-Rich, P., and Hall, M. 2012b. Theropod fauna from Southern Australia indicates high polar diversity and climate-driven dinosaur provinciality. PLoS ONE 7 (5): e37122. Crossref
Brochu, C.A. 2003. Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Journal of Vertebrate Paleontology 22 (Supplement 4): 1–138. Crossref
Britt, B.B. 1991. Theropods of Dry Mesa Quarry (Morrison Formation, late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri. Brigham Young University Geology Studies 37: 1–72.
Brusatte, S.L. and Benson, R.B.J. 2013. The systematics of Late Jurassic tyrannosauroid theropods from Europe and North America. Acta Palaeontologica Polonica 58: 47–54. Crossref
Brusatte, S.L. and Sereno, P.C. 2008. Phylogeny of Allosauroidea (Dinosauria: Theropoda): comparative analysis and resolution. Journal of Systematic Palaeontology 6: 155–182. Crossref
Brusatte, S.L., Benson, R.B.J., and Hutt, S. 2008. The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden Group (Barremian) of the Isle of Wight. Monograph of the Palaeontographical Society 162 (631): 1–166.
Brusatte, S.L., Carr, T.D., and Norell, M.A. 2012. The osteology of Alioramus, a gracile and long-snouted tyrannosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Mongolia. Bulletin of the American Museum of Natural History 366: 1–197. Crossref
Buffetaut, E. 1982. Mesozoic vertebrates from Thailand and their palaeobiological significance. Terra Cognita 2: 27–34.
Buffetaut, E. and Ingavat, R. 1984. A very small theropod dinosaur from the Upper Jurassic of Thailand. Comptes rendus de l’Académie des Sciences Paris II 298: 915–918.
Buffetaut, E. and Ingavat, R. 1986. Unusual theropod dinosaur teeth from the Upper Jurassic of Phu Wiang, northeastern Thailand. Revue de paléobiologie 5: 217–220.
Buffetaut, E. and Suteethorn, V. 1992. A new species of the ornithischian dinosaur Psittacosaurus from the early Cretaceous of Thailand. Palaeontology 35: 801–812.
Buffetaut, E. and Suteethorn, V. 1998. The biogeographical significance of the Mesozoic vertebrates from Thailand. In: R. Hall and J.D. Holloway (ed.), Biogeography and Geological Evolution of SE Asia, 83–90. Backhuys, Leiden.
Buffetaut, E. and Suteethorn, V. 1999. The dinosaur fauna of the Sao Khua Formation of Thailand and the beginning of the Cretaceous radiation of dinosaurs in Asia. Palaeogeography, Palaeoclimatology, Palaeoecology 150: 13–23. Crossref
Buffetaut, E. and Suteethorn, V. 2012. A carcharodontid theropod (Dinosauria,Saurischia) from the Sao Khua Formation (Early Cretaceous, Barremian) of Thailand. In: R. Royo-Torres, F. Gascó, and L. Alcalá (coord.), 10th Annual Meeting of the European Association of Vertebrate Palaeontologists. Fundamental 20: 1–290.
Buffetaut, E., Suteethorn, V., and Tong, H. 1996. The earliest known tyrannosaur from the Lower Cretaceous of Thailand. Nature 381: 689–691. Crossref
Buffetaut, E., Suteethorn, V., and Tong, H. 2009. An early “ostrich dinosaur” (Theropoda: Ornithomimosauria) from the Early Cretaceous Sao Khua Formation of NE Thailand. In: E. Buffetaut, G. Cuny, J. Le Loeuff, and V. Suteethorn (eds.), Late Palaeozoic and Mesozoic Ecosystems in SE Asia. Geological Society, London, Special Publications 315: 229–243. Crossref
Calvo, J.O., Porfiri, J.D., Veralli, C., Novas, F., and Poblete, F. 2004. Phylogenetic status of Megaraptor namunhuaiquii Novas based on a new specimen from Neuquén, Patagonia, Argentina. Ameghiniana 41: 565–575.
Carpenter, K., Miles, C., and Cloward, K. 2005a. New small theropod from the Upper Jurassic Morrison Formation of Wyoming. In: K. Carpenter (ed.), The Carnivorous Dinosaurs, 23–48. Indiana University Press, Bloomington.
Carpenter, K., Miles, C., Ostrom, J.H, and Cloward, K. 2005b. Redescription of the small maniraptoran theropods Ornitholestes and Coelurus from the Upper Jurassic Morrison Formation of Wyoming. In: K. Carpenter (ed.), The Carnivorous Dinosaurs, 49–71. Indiana University Press, Bloomington.
Carr, T.D., Williamson, T.E., and Schwimmer, D.R. 2005. A new genus and species of tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama. Journal of Vertebrate Paleontology 25: 119–143. Crossref
Carrano, M.T. 2007. The appendicular skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae). In: S.D. Sampson and D.W. Krause (eds.), Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8, Supplement to Journal of Vertebrate Paleontology 27 (2): 163–179. Crossref
Carrano, M.T., Benson, R.B., and Sampson, S.D. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology 10: 211–300. Crossref
Carrano, M.T., Sampson, S.D., and Forster, C.A. 2002. The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 22: 510–534. Crossref
Cau, A. 2018. The assembly of the avian body plan: A 160-million-year long process. Bollettino della Società Paleontologica Italiana 57: 1–25.
Choiniere, J.N., Clark, J.M., Forster, C.A., and Xu, X. 2010. A basal coelurosaur (Dinosauria: Theropoda) from the Late Jurassic (Oxfordian) of the Shishugou Formation in Wucaiwan, People’s Republic of China. Journal of Vertebrate Paleontology 30: 1773–1796. Crossref
Choiniere, J.N., Forster, C.A., and de Klerk, W.J. 2012. New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous Kirkwood Formation in South Africa. Journal of African Earth Sciences 71: 1–17. Crossref
Coria, R.A. and Currie, P.J. 2016. A new megaraptoran dinosaur (Dinosauria, Theropoda, Megaraptoridae) from the Late Cretaceous of Patagonia. PLoS ONE 11 (7): e0157973. Crossref
Currie, P.J. and Carpenter, K. 2000. A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) of Oklahoma, USA. Geodiversitas 22: 207–246.
Currie, P.J. and Zhao, X.J. 1993. A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Canadian Journal of Earth Sciences 30: 2037–2081. Crossref
Delcourt, R. and Grillo, O.N. 2018. Tyrannosauroids from the Southern Hemisphere: Implications for biogeography, evolution, and taxonomy. Palaeogeography, Palaeoclimatology, Palaeoecology 511: 379–387. Crossref
Ezcurra, M.D. and Novas, F.E. 2016. Theropod dinosaurs from Argentina. Historia Evolutiva y Paleobiogeografía de los Vertebrados de América del Sur. Contribuciones del MACN 6: 139–156.
Gauthier, J. 1986. Saurischian monophyly and the origin of birds. Memoirs of the California Academy of sciences 8: 1–55.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Harris, J.D. 1998. A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiogeographic implications, based on a new specimen from Texas. New Mexico Museum of Natural History and Science Bulletin 13: 1–75.
Hocknull, S.A., White, M.A., Tischler, T.R., Cook, A.G., Calleja, N.D., Sloan, T., and Elliott, D.A. 2009. New mid-Cretaceous (latest Albian) dinosaurs from Winton, Queensland, Australia. PLoS One 4 (7): e6190. Crossref
Holtz, Jr. T.R., Molnar, R.E., and Currie, P.J. 2004. Basal Tetanurae. In: P. Dodson, H. Osmólska, and D.B. Weishampel (eds.), The Dinosauria. Second edition, 71–110. University of California Press, Berkeley. Crossref
Hu, S.-Y. 1964. Carnosaurian remains from Alashan, inner Mongolia. Vertebrata Palasiatica 8: 42–63.
von Huene. 1920. Bemerkungen zur Systematik und Stammegeschichte einiger Reptilian. Zeitschrift f¨ur Induktive Abstammungs- und Vererbungslehre 24: 209–212. Crossref
Hutt, S., Naish, D., Martill, D.M., Barker, M.J., and Newbery, P. 2001. A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Cretaceous) of southern England. Cretaceous Research 22: 227–242. Crossref
Ibrahim, N., Sereno, P.C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D.M., Zouhri, S., Myhrvold, N., and Iurino, D.A. 2014. Semiaquatic adaptations in a giant predatory dinosaur. Science 345: 1613–1616. Crossref
Kobayashi, Y. and Barsbold, R. 2005. Reexamination of a primitive ornithomimosaur, Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda), from the Late Cretaceous of Mongolia. Canadian Journal of Earth Sciences 42: 1501–1521. Crossref
Langer, M.C., Rincón, A.D., Ramezani, J., Solórzano, A., and Rauhut, O.W. 2014. New dinosaur (Theropoda, stem-Averostra) from the earliest Jurassic of the La Quinta formation, Venezuelan Andes. Royal Society Open Science 1 (2): 140184. Crossref
Long, J.A. and Molnar, R.E. 1998. A new Jurassic theropod dinosaur from Western Australia. Records Western Australian Museum 19: 121–129.
Lü, J., Yi, L., Brusatte, S.L., Yang, L., Li, H., and Chen, L. 2014. A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. Nature Communications 5: 3788. Crossref
Maddison, W.P. and Maddison, D.R. 2015. Mesquite: a Modular System for Evolutionary Analysis. Version 3.04. available at http://www.mesquiteproject.org
Madsen, J.H., Jr. 1976. Allosaurus fragilis: a revised osteology. Utah Geological and Mineral Survey Bulletin 109: 1–163.
Maganuco, S. and Dal Sasso, C. 2018. The smallest biggest theropod dinosaur: a tiny pedal ungual of a juvenile Spinosaurus from the Cretaceous of Morocco. PeerJ 6: e4785. Crossref
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Part V. American Journal of Science 125: 417–423. Crossref
Martin, V., Buffetaut, E., and Suteethorn, V. 1994. A new genus of sauropod dinosaur from the Sao Khua Formation (Late Jurassic to Early Cretaceous) of northeastern Thailand. Comptes rendus de l’Académie des Sciences Paris II 319: 1085–1092.
Martinelli, A.G., Borges Ribeiro, L.C., Méndez, A.H., Neto, F.M., Cavellani, C.L., Felix, E., da Fonseca Ferraz, M.L., and Antunes Teixeira, V.D.P. 2013. Insight on the theropod fauna from the Uberaba Formation (Bauru Group), Minas Gerais State: new megaraptoran specimen from the Late Cretaceous of Brazil. Rivista Italiana di Paleontologia e Stratigrafia 119: 205–214.
Martínez, R. and Novas, F. 2006. Aniksosaurus darwini gen. et sp. nov., a new coelurosaurian theropod from the Early Late Cretaceous of Central Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales. Nueva Serie 8: 243–259. Crossref
Méndez, A.H., Novas, F.E., and Iori, F.V. 2012. First record of Megaraptora (Theropoda, Neovenatoridae) from Brazil. Comptes Rendus Palevol 11: 251–256. Crossref
Mo, J., Zhou, F., Li, G., Huang, Z., and Cao, C. 2014. A new Carcharodontosauria (Theropoda) from the Early Cretaceous of Guangxi, Southern China. Acta Geologica Sinica (English Edition) 88: 1051–1059. Crossref
Motta, M.J., Aranciaga Rolando, A.M., Rozadilla, S., Agnolín, F.E., Chimento, N.R., Egli, F.B., and Novas, F.E. 2016. New theropod fauna from the Upper Cretaceous (Huincul Formation) of northwestern Patagonia, Argentina. New Mexico Museum of Natural History and Science Bulletin 71: 231–253.
Novas, F.E. 1998. Megaraptor namunhuaiquii, gen. et sp. nov., a large-clawed, Late Cretaceous theropod from Patagonia. Journal of Vertebrate Paleontology 18: 4–9. Crossref
Novas, F.E., Agnolín, F.L., Ezcurra, M.D., Canale, J.I., and Porfiri, J.D. 2012a. Megaraptorans as members of an unexpected evolutionary radiation of tyrant-reptiles in Gondwana. Ameghiniana 49 (Supplement): R33.
Novas, F.E., Agnolin, F.L., Ezcurra, M.D., Porfiri, J., and Canale, J.I. 2013. Evolution of the carnivorous dinosaurs during the Cretaceous: The evidence from Patagonia. Cretaceous Research 45: 174–215. Crossref
Novas, F.E., Aranciaga Rolando, A.M., and Agnolín, F.L. 2016. Phylogenetic relationships of the Cretaceous Gondwanan theropods Megaraptor and Australovenator: the evidence afforded by their manual anatomy. Memoirs of Museum Victoria 74: 49–61. Crossref
Novas, F.E., Ezcurra, M.D., Agnolin, F.L., Pol, D., and Ortíz, R. 2012b. New Patagonian Cretaceous theropod sheds light about the early radiation of Coelurosauria. Revista del Museo Argentino de Ciencias Naturales 14: 57–81. Crossref
Novas, F.E., Ezcurra, M.D., and Lecuona, A. 2008. Orkoraptor burkei nov. gen. et sp., a large theropod from the Maastrichtian Pari Aike Formation, Southern Patagonia, Argentina. Cretaceous Research 29: 468–480. Crossref
Novas, F.E., Salgado, L., Suárez, M., Agnolín, F.L., Ezcurra, M.N.D., Chimento, N.S.R., de la Cruz, R., Isasi, M.P., Vargas, A.O., and Rubilar-Rogers, D. 2015. An enigmatic plant-eating theropod from the Late Jurassic period of Chile. Nature 522: 331–334. Crossref
Osmólska, H. and Roniewicz, E. 1970. Deinocheiridae, a new family of theropod dinosaurs. Palaeontologica Polonica 21: 5–19.
Owen, R. 1842. Report on British fossil reptiles, part II. Reports of the British Association for the Advancement of Science 11 (for 1841): 60–204.
Porfiri, J.D., Novas, F.E., Calvo, J.O., Agnolin, F.L., Ezcurra, M.D., and Cerda, I.A. 2014. Juvenile specimen of Megaraptor (Dinosauria, Theropoda) sheds light about tyrannosauroid radiation. Cretaceous Research 51: 35–55. Crossref
Porfiri, J.D., Valieri, R.D.J., Santos, D.D., and Lamanna, M.C. 2018. A new megaraptoran theropod dinosaur from the Upper Cretaceous Bajo de la Carpa Formation of northwestern Patagonia. Cretaceous Research 89: 302–319. Crossref
Rauhut, O.W.M. 2003. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontology 69: 1–215.
Rauhut, O.W.M. 2005. Osteology and relationships of a new theropod dinosaur from the Middle Jurassic of Patagonia. Palaeontology 48: 87–110. Crossref
Rauhut, O.W.M. 2012. A reappraisal of a putative record of abelisauroid theropod dinosaur from the Middle Jurassic of England. Proceedings of the Geologists’ Association 123: 779–786. Crossref
Rauhut, O.W.M. and Pol, D. 2017. A theropod dinosaur from the Late Jurassic Cañadón Calcáreo Formation of Central Patagonia, and the evolution of the theropod tarsus. Ameghiniana 54: 539–566. Crossref
Rauhut, O.W.M. and Xu, X. 2005. The small theropod dinosaurs Tugulusaurus and Phaedrolosaurus from the Early Cretaceous of Xinjiang, China. Journal of Vertebrate Paleontology 25: 107–118. Crossref
Sales, M.A., Martinelli, A.G., Francischini, H., Rubert, R.R., Marconato, L.P., Soares, M.B., and Schultz, C.L. 2017. New dinosaur remains and the tetrapod fauna from the Upper Cretaceous of Mato Grosso State, central Brazil. Historical Biology [published online, https://doi.org/10. 1080/08912963.2017.1315414]. Crossref
Samathi, A. 2013. Osteology and Phylogenetic Position of Siamotyrannus isanensis (Dinosauria; Theropoda) from the Lower Cretaceous of Thailand. 69 pp. Unpublished Master’s Thesis. Ludwig-Maximilians-Universität München, Munich.
Samathi, A. and Chanthasit, P. 2015. New Megaraptoran (Dinosauria: Theropoda) from the Early Cretaceous Sao Khua Formation of Thailand. In: 2nd International Symposium on Asian Dinosaurs, Bangkok. 69 pp. Nakhon Ratchasima Rajabhat University and Department of Mineral Resources, Bangkok.
Samathi, A. and Chanthasit, P. 2017. Two new basal Megaraptora (Dinosauria: Theropoda) from the Early Cretaceous of Thailand with comment on the phylogenetic position of Siamotyrannus and Datanglong. Journal of Vertebrate Paleontology 37: 188.
Sereno, P.C., Martinez, R.N., Wilson, J.A., Varricchio, D.J., Alcober, O.A., and Larsson, H.C.E. 2008. Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina. PLoS ONE 3 (9): e3303. Crossref
Sereno, P.C., Tan, L., Brusatte, S.L., Kriegstein, H.J., Zhao, X., and Cloward, K. 2009. Tyrannosaurid skeletal design first evolved at small body size. Science 326: 418–422. Crossref
Sereno, P.C., Wilson, J.A., Larsson, H.C., Dutheil, D.B., and Sues, H.D. 1994. Early Cretaceous dinosaurs from the Sahara. Science 266: 267–271. Crossref
Smith, N.D., Makovicky, P.J., Agnolin, F.L., Ezcurra, M.D., Pais, D.F., and Salisbury, S.W. 2008. A Megaraptor-like theropod (Dinosauria: Tetanurae) in Australia: support for faunal exchange across eastern and western Gondwana in the mid-Cretaceous. Proceedings of the Royal Society of London B 275: 2085–2093. Crossref
Stovall, J.W. and Langston, W. 1950. Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma. The American Midland Naturalist 43: 696–728. Crossref
Suteethorn, S., Le Loeuff, J., Buffetaut, E., Suteethorn, V., Talubmook, C., and Chonglakmani, C. 2009. A new skeleton of Phuwiangosaurus sirindhornae (Dinosauria, Sauropoda) from NE Thailand. In: E. Buffetaut, G. Cuny, J. Le Loeuff, and V. Suteethorn (eds.), Late Palaeozoic and Mesozoic Ecosystems in SE Asia. Geological Society of London Special Publication 315: 189–215. Crossref
Tumpeesawan, S., Sato, Y., and Nakhapadungrat, S. 2010. A new species of Pseudohyria (Matsumotoina) (Bivalvia: Trigonioidoidea) from the Early Cretaceous Sao Khua Formation, Khorat Group, northeastern Thailand. Tropical Natural History 10: 93–106.
Welles, S.P. 1984. Dilophosaurus wetherilli (Dinosauria, Theropoda). Osteology and comparisons. Palaeontographica Abteilung A 85–180.
White, M.A., Benson, R.B., Tischler, T.R., Hocknull, S.A., Cook, A.G., Barnes, D.G., Poropat, S.F., Wooldridge, S.J., Sloan, T., Sinapius, G.H., and Elliott, D.A. 2013a. New Australovenator hind limb elements pertaining to the holotype reveal the most complete neovenatorid leg. PLoS ONE 8 (7): e68649. Crossref
White, M.A., Cook, A.G., Hocknull, S.A., Sloan, T., Sinapius, G.H.K, and Elliott, D.A. 2012. New forearm elements discovered of holotype specimen Australovenator wintonensis from Winton, Queensland, Australia. PLoS ONE 7 (6): e39364. Crossref
White, M.A., Falkingham, P.L., Cook, A.G., Hocknull, S.A., and Elliott, D.A. 2013b. Morphological comparisons of metacarpal I for Australovenator wintonensisand and Rapator ornitholestoides: Implications for their taxonomic relationships. Alcheringa 37: 435–441. Crossref
Xu, X., Clark, J.M., Forster, C.A., Norell, M.A., Erickson, G.M., Eberth, D.A., Jia, C., and Zhao, Q. 2006. A basal tyrannosauroid dinosaur from the Late Jurassic of China. Nature 439: 715–718. Crossref
Xu, X., Wang, K., Zhang, K., Ma, Q., Xing, L., Sullivan, C., Hu, D., Cheng, S., and Wang, S., 2012. A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature 484: 92. Crossref
Xu, X., Zhang, X., Paul, S., Zhao, X., Kuang, X., Han, J., and Tan, L. 2002. A new therizinosauroid (Dinosauria, Theropoda) from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol. Vertebrata Pal Asiatica 40: 228–240.
Zanno, L.E. 2006. The pectoral girdle and forelimb of the primitive therizinosauroid Falcarius utahensis (Theropoda, Maniraptora): analyzing evolutionary trends within Therizinosauroidea. Journal of Vertebrate Paleontology 26: 636–650. Crossref
Zanno, L.E. 2010. Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs. Zoological Journal of the Linnean Society 158: 196–230. Crossref
Zanno, L.E. and Makovicky, P.J. 2013. Neovenatorid theropods are apex predators in the Late Cretaceous of North America. Nature Communications 4: 2827. Crossref
Acta Palaeontol. Pol. 64 (2): 239–260, 2019
https://doi.org/10.4202/app.00540.2018