
A new species of the metriorhynchid crocodylomorph Cricosaurus from the Upper Jurassic of southern Germany
SVEN SACHS, MARK T. YOUNG, PASCAL ABEL, and HEINRICH MALLISON
Sachs, S., Young, M.T., Abel, P., and Mallison, H. 2019. A new species of metriorhynchid crocodylomorph Cricosaurus from the Upper Jurassic of southern Germany. Acta Palaeontologica Polonica 64 (2): 343–356.
Here we describe a new species of the metriorhynchid thalattosuchian Cricosaurus, C. bambergensis sp. nov., from the Upper Jurassic Torleite Formation of Wattendorf near Bamberg, Bavaria (southern Germany). The holotype and only known specimen is a nearly complete skeleton that shows a number of diagnostic traits including a bicarinate dentition formed by labiolingually compressed tooth crowns that lack a conspicuous enamel ornamentation and the presence of a distinct midline ridge with paired depressions on the palatines. Our phylogenetic analysis recovers a grouping of Cricosaurus bambergensis sp. nov. with C. elegans and C. suevicus. The implications of the new Cricosaurus species to the species complex from the late Kimmeridgian–early Tithonian of southern Germany is discussed. Our description of C. bambergensis demonstrates that the specific, and morphological, diversity of Cricosaurus in southern Germany was higher than previously thought. This coincides with the recent trend of re-evaluating the species-complexes of extant taxa, and the identification of new “cryptic species”. As such, the crocodylomorph fossil record will need to be re-examined to ensure there is not an underestimation of their biodiversity.
Key words: Crocodylomorpha, Metriorhynchidae, Cricosaurus, Upper Jurassic, Torleite Formation, Germany.
Sven Sachs [Sachs.Pal@gmail.com], Naturkunde-Museum Bielefeld, Abteilung Geowissenschaften, Adenauerplatz 2, 33602 Bielefeld, Germany; Im Hof 9, 51766 Engelskirchen, Germany (correspondence address).
Mark T. Young [mark.young@ed.ac.uk], School of GeoSciences, Grant Institute, University of Edinburgh, James Hutton Road, Edinburgh, EH9 3FE, UK.
Pascal Abel [pascal.abel@ifg.uni-tuebingen.de], Senckenberg Centre for Human Evolution and Palaeoenvironment, Eberhard-Karls Universität Tübingen, Sigwartstraße 10, 72076 Tübingen, Germany.
Heinrich Mallison [mallison@palaeo3d.com], Palaeo3D, Talstr. 14, 86554 Pöttmes, Germany.
Received 30 August 2018, accepted 17 January 2019, available online 4 April 2019.
Copyright © 2019 S. Sachs et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The metriorhynchid genus Cricosaurus was established by Wagner (1858) based upon material from the Altmühl Valley in Bavaria (southern Germany), and is one of the first described thalattosuchian taxa. Later, Fraas (1902) considered Cricosaurus to be a subjective junior synonym of Geosaurus Cuvier, 1824, but a reassessment by Young and Andrade (2009) clarified its status as a distinct, valid genus. Cricosaurus currently has six valid species: C. araucanensis (Gasparini and Dellapé, 1976), C. elegans (Wagner, 1852), C. lithographicus Herrera, Gasparini, and Fernández, 2013b, C. schroederi (Kuhn, 1936), C. suevicus (Fraas, 1901), and C. vignaudi (Frey, Buchy, Stinnesbeck, and López Oliva, 2002), which are known from localities in Europe and America, and cover a geological time span reaching from the Late Jurassic to the Early Cretaceous. “Cricosaurus” saltillensis (Buchy, Vignaud, Frey, Stinnesbeck, and González, 2006) and “C.” macrospondylus (Koken, 1883) have not been recovered in the same clade as other Cricosaurus species in the most recent phylogenetic analyses of Metriorhynchidae (e.g., Ősi et al. 2018). Furthermore, the validity of the Late Jurassic species Cricosaurus medius Wagner, 1858 is unclear.
During an excavation led by the Naturkunde-Museum Bamberg on 5 June 2014, a new Cricosaurus skeleton (NKMB-P-Watt14/274; Fig. 1) was found in a quarry in Wattendorf near Bamberg (Mäuser 2015). At this locality lithographic limestone of the late Kimmeridgian Wattendorf Member of the Torleite Formation, one of the oldest such occurrences in southern Germany, are exposed. The Wattendorf quarry yielded numerous plant and invertebrate fossils, but also exquisitely preserved fishes, rhynchocephalians, turtles, pterosaurs, and thalattosuchians (Mäuser 2014). The latter record includes, next to the Cricosaurus skeleton, some isolated tooth crowns, a fragmentary rostrum of a teleosaurid and the posterior portion of another metriorhynchid skeleton (Mäuser 2014).
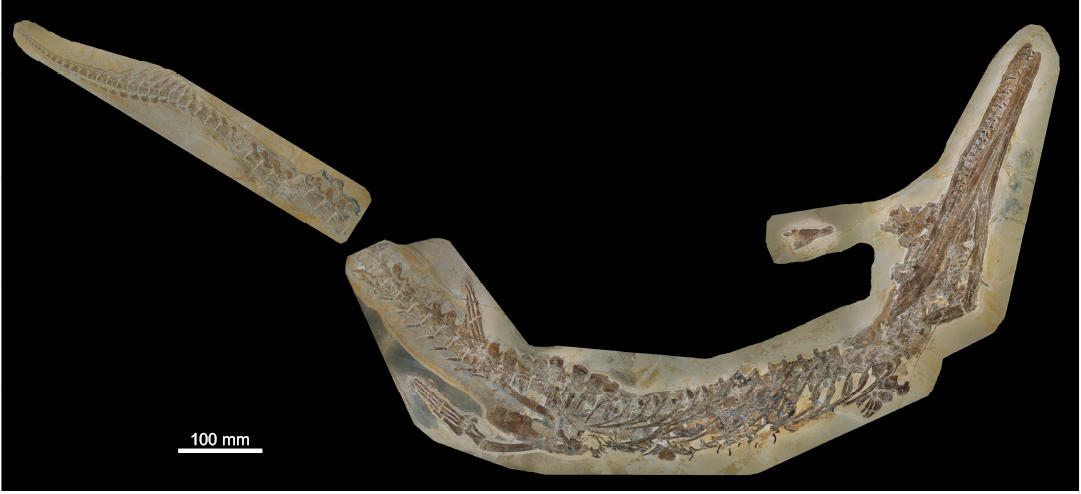
Fig. 1. Metriorhynchid thalattosuchian Cricosaurus bambergensis sp. nov., NKMB-P-Watt14/274, holotype, late Kimmeridgian of Wattendorf (southern Germany). Skeleton in ventrolateral view.
Here, we provide an in-depth description of the Wattendorf Cricosaurus skeleton (NKMB-P-Watt14/274). The specimen is largely articulated and almost complete, but some parts such as the skull roof and the posterior region of the right mandibular ramus are poorly preserved. Furthermore, most of the tail is displaced and was found next to the remainder of the skeleton. This preservational damage has probably been caused by a larger predator (Mäuser 2015).
Most Cricosaurus skeletons are incomplete or partially disarticulated, such as those known for C. araucanensis (Herrera et al. 2009, 2013a, 2017). Until now, only one species has been known from fully, or nearly fully, articulated specimens, Cricosaurus suevicus (see Fraas 1902). However, NKMB-P-Watt14/274 shows some unique traits in the skull and at the tail fluke. These help to distinguish it from all known Cricosaurus species and justify the erection of a new species for the specimen from Wattendorf.
Institutional abbreviations.—BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany; GPIT, Institut und Museum für Geologie und Paläontologie der Universität Tübingen, Tübingen, Germany; NKMB, Naturkunde-Museum Bamberg, Bamberg, Germany; NHMUK, Natural History Museum, London, UK; SMNS, Staatliches Museum für Naturkunde Stuttgart, Germany.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:3B83FBAE-D826-4CC8-834E-D426960AB99A
Geological setting
NKMB-P-Watt14/274 is preserved in carbonate rocks of the Torleite Formation (late Kimmeridgian), which in Bavaria forms the lowermost formation of the Upper Weißjura Group of the Franconian Jura (Fig. 2). The Torleite Formation occurs in most parts of the Franconian Jura and can be subdivided into several geographically separated members (Niebuhr and Pürner 2014). The matrix of NKMB-P-Watt14/274 is a light ochre, micritic lithographic limestone of the Wattendorf Member, the northernmost member of the Torleite Formation (Fürsich et al. 2007; Niebuhr and Pürner 2014). The member exhibits one of the oldest known occurrences of lithographic limestone in southern Germany (upper Aulacostephanus pseudomutabilis Tethys Ammonite Zone; Schweigert 2007; Niebuhr and Pürner 2014). It differs from most other lithographic limestone of southern Germany by a higher abundance of coarse-grained facies and a lack of coccoliths and calcispheres (Fürsich et al. 2007).
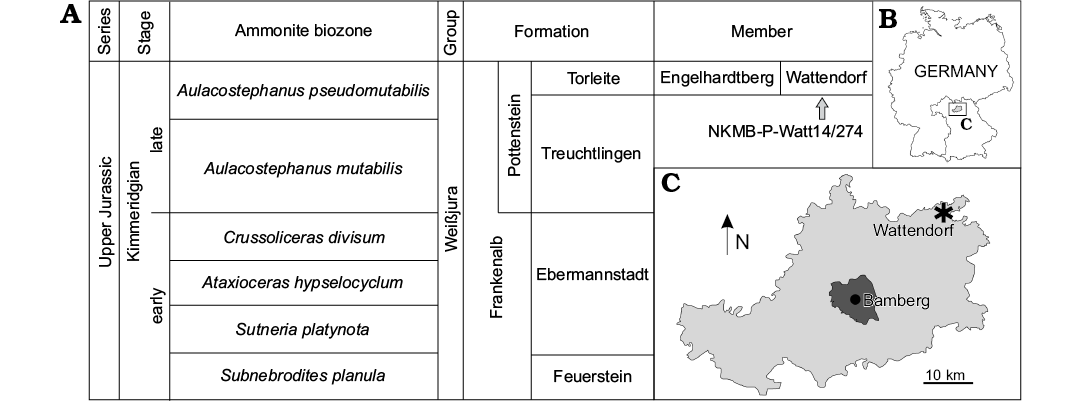
Fig. 2. A. Reduced stratigraphic table of the Kimmeridgian rock units of the northern Franconian Jurassic, based on Niebuhr and Pürner (2014). B, C. Maps showing the locality of NKMB-P-Watt14/274 at Wattendorf (asterisk) within the Bamberg district and its placement in Germany.
Palaeoenvironment.—As for all units of the Weißjura Group, the rocks of the Torleite Formation formed on a subtropical carbonate shelf in the northern part of the Tethys Sea (Schmid et al. 2005). At the base of the Torleite Formation, a transgression led to the formation of small basins on top and within the former sponge-microbial reefs of the Frankenalb Formation (Röper 2005; Niebuhr and Pürner 2014). These so called “Wannen” basins were lagoonal environments with repeatedly dysoxic and anoxic conditions at the seafloor, benefiting an often-articulated preservation of the deposited organisms. Still, the upper water layers remained habitable as evident by a rich diversity of nektonic and planktonic species (Fürsich et al. 2007).
The Wattendorf Member was formed in a laterally more restricted basin further north of the Solnhofen archipelago and its associated lagoons. Fossils of land plants and terrestrial vertebrates demonstrate a placement in the proximal realm, which is also evident by the intercalations of coarse-grained facies (Fürsich et al. 2007). Within the Solnhofen archipelago the Wattendorf “Wanne” is most similar to the Pfraundorf locality, both showing a similar faunal assemblage consisting of diverse osteichthyes and crustaceans as well as the specialized bivalve Aulacomyella (Fürsich et al. 2007). Other thalattosuchian records from the Wattendorf member comprise so-far undescribed skeletal remains and isolated teeth, indicating the presence of Dakosaurus and Geosaurus.
Systematic palaeontology
Crocodylomorpha Hay, 1930 (sensu Nesbitt 2011)
Thalattosuchia Fraas, 1901 (sensu Young and Andrade 2009)
Metriorhynchidae Fitzinger, 1843 (sensu Young and Andrade 2009)
Genus Cricosaurus Wagner, 1858
Type species: Stenosaurus elegans Wagner, 1852; Daiting, Bavaria, Germany; Mörnsheim Formation, Hybonoticeras hybonotum Tethyian Ammonite Zone, lower Tithonian, Upper Jurassic. Now referred to as Cricosaurus elegans (Wagner, 1852) Wagner, 1858 [following recommendation 67B of the International Commission on Zoological Nomenclature (ICZN) code].
Emended diagnosis.—Metriorhynchid crocodylomorph with the following unique combination of characters: dentition has variable labiolingual compression, with some species having teeth with a sub-circular cross-section and others with a sub-rectangular one; dentition is bicarinate, but the carinae are faint (i.e., a low carinal keel); external surface of the dermatocranium is largely smooth, lacking conspicuous ornamentation; acute angle formed by the posterolateral and posteromedial processes of the frontal; dorsal margin of the supratemporal arcade is lower than the medial process of the frontal; orbits as large as the supratemporal fenestrae (at least in morphologically mature individuals); surangular and angular well-developed, anterolaterally exceeding beyond the anterior margin of the orbits; symphysial part of the mandible low; lateral margin of the prefrontal rounded; external nares separated by a bony premaxillary septum, with the nares terminating posterior to the second maxillary alveoli; calcaneum tuber absent or vestigial; proximal end of metatarsal I greatly enlarged (modified from Young and Andrade 2009 and Herrera et al. 2013b).
Cricosaurus bambergensis sp. nov.
Figs. 1, 3–7.
ZooBank LCID: urn:lsid:zoobank.org:act:AAAD9169-1411-451C-BB B5-F069E938575F
Etymology: Named after the city of Bamberg, where the specimen is housed and that is the namesake of the Bamberg district, which encompasses the type locality at Wattendorf.
Holotype: NKMB-P-Watt14/274, almost complete skeleton (Figs. 1, 3–7).
Type locality: Kalk- und Dolomitbruch der Andreas Schorr GmbH, Wattendorf, Bamberg district, Bavaria, southern Germany (Fig. 2).
Type horizon: Aulacostephanus pseudomutabilis Tethys Ammonite Zone, Wattendorf Member, Torleite Formation, late Kimmeridgian, Upper Jurassic (Fig. 2).
Diagnosis.—A member of Cricosaurus with the following unique combination of characters (proposed autapomorphic characters are indicated by an asterisk): bicarinate dentition, lacking conspicuous enamel ornamentation; tooth crowns in the premaxilla, maxilla and dentary show distinct labiolingual compression (shared with C. suevicus and C. elegans); the maxillary tooth row continues posteriorly past the anterior margin of the orbit (thalattosuchian symplesiomorphy); presence of pronounced reception pits for dentary tooth crowns on the lateral margins of the anterior and mid-portions of the maxilla; palatines have a distinct midline ridge with paired depressions on either side*; morphology of the tail displacement unit: the distal-most three preflexural vertebrae have neural spines that are oriented very strongly posteriorly, their posterior margin lies on top of the prezygapophyses of the adjacent vertebra; these neural spines appear almost rod-like in lateral view; the three vertebrae anterior to them have a similar morphology, but the neural spines look more like a “regular” neural spine, albeit strongly oriented posteriorly*.
Cricosaurus bambergensis retains the following four metriorhynchid tail fluke symplesiomorphies (unlike C. suevicus): (i) the distal-most preflexural caudal vertebra has a rod-like hemapophysis; (ii) the proximal-most five or six flexural vertebrae have anteriorly oriented neural spines, but not strongly oriented anteriorly; (iii) the flexural hemapophyses contact one another along their anterior–posterior margins; and (iv) the distal-most postflexural hemapophysis return to a rod-like morphology.
Description.—Supplementary Online Material available at http://app.pan.pl/SOM/app64-Sachs_etal_SOM.pdf illustrates the specimen in detail and contains further information: SOM 1, a text file with information on 3D model and orthophoto generation, 3D file viewing information, and further information on the phylogenetic analysis; SOM 2, 3D models of the fossil slabs; SOM 3, a very high resolution orthophoto of the 3D models; SOM 4, *.emf files for bootstrap values and Bremer support values and nexus file.
General remarks on the skull: The cranium and mandible of Cricosaurus bambergensis are preserved in articulation and are exposed in ventral view (Fig. 3). The skull is largely damaged, but part of the palate and the mandible are well-preserved and bear traceable sutures. Unidentifiable skull roof elements are preserved adjacent to the skull.
Premaxillae: The premaxillae are exposed in ventrolateral view (Fig. 3). Their lateral sides are undulated and convex at the alveoli. The exposed ventral side is nearly flat and the premaxillae are separated posteroventrally by the pointed anteroventral portions of the conjoint maxillae. The premaxillary–maxillary suture is visible ventrally and terminates at the position of the third premaxillary tooth. The conjoint premaxillae form a rounded tip of the rostrum.
Maxillae: The maxillae are exposed in ventral and ventrolateral view (Fig. 3). As at the premaxillae the alveolar edges are undulated, forming pronounced reception pits for dentary tooth crowns. The right maxilla bears 23 teeth. The exposed ventral surface shows that the conjoined maxillae form a pointed tip that separates the premaxillae posteroventrally. This tip extends to the level of the third premaxillary tooth. Posterolaterally a jugal contact is established. A ventral midline suture is clearly visible and the maxillae are slightly dorsally inclined throughout the ventral midline.
Jugal: Part of the strap-shaped jugal is preserved posterior to the maxilla on the right side of the cranium (Fig. 3).
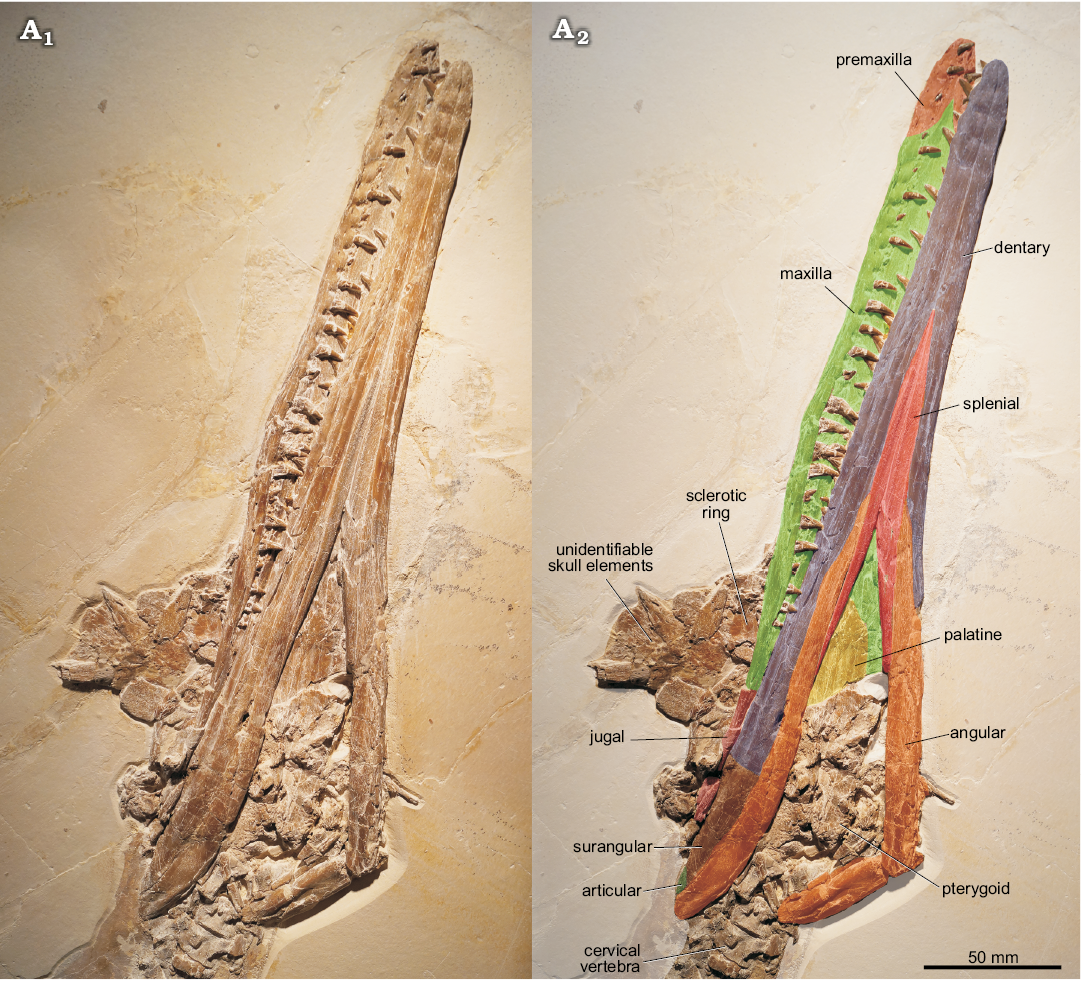
Fig. 3. Metriorhynchid thalattosuchian Cricosaurus bambergensis sp. nov. (NKMB-P-Watt14/274) holotype, late Kimmeridgian of Wattendorf (southern Germany). A1, skull in ventrolateral view; A2,illustrates the preserved elements which can be unambiguously identified (please note that the dentary-surangular contact cannot be traced and is based upon the position in C. suevicus).
Palatines: The almost complete palatines are well preserved and exposed in ventral view (Figs. 3, 4B). The anterior portion of the conjoined palatines is obscured by the mandibular rami, and the posterior part, where the palatines contact the pterygoids, is damaged. Interestingly, the conjoined palatines form a prominent ridge at the midline suture that flares anteriorly and demarcates two elongate depressions that are placed adjacent to the ridge (Figs. 4B).
Pterygoids: Damaged remnants of the pterygoids are present, but the poor preservation does not provide any information (Fig. 3).
Sclerotic ring: Fragments from the sclerotic ring are preserved along with some unidentifiable skull elements near the posterior portion of the right maxilla (Fig. 3).
Unidentifiable skull elements: A number of skull roof elements, most likely from the nasal–prefrontal–frontal complex and probably also from the postorbital and squamosal are placed dorsally in the posterior part of the skull (Fig. 3), but they cannot be identified more precisely.
Dentaries: The dentaries are visible in ventral view, but no clear surangular suture can be traced (Fig. 3). The straight ventral midline contact is well visible and forms more than 50% of the symphysis. The later continues in the conjoined splenials that separate the dentaries posteriorly. The surangulodentary groove starts in about the middle of the mandible and becomes more prominent posteriorly. The exact tooth count cannot be provided as some tooth positions are obscured.
Surangulars: The surangular cannot be separated from the dentary. The surangulodentary groove is visible in lateral view, on the posterior region of the ramus (Fig. 3). The surangular forms the posterodorsal part of the mandible and is demarcated ventrally by the angular. Posteriorly the surangular forms part of the dorsal portion of the retroarticular process and, at the posterior end, contacts the articular.
Angulars: The elongate, strap-like angular is well visible and starts at about mid-length of the mandible and forms the posteroventral demarcation of the latter (Fig. 3). It terminates at the posterior end of the retroarticular process of which it forms the ventral portion. The latter is broken off, displaying a bulged midline part. Anteriorly, a splenial contact is established on the medial side of each angular and a long dentary suture is formed laterally. Posterodorsally the angular contacts the articular.
Articular: Only a small portion of the articular is exposed posterodorsally at the retroarticular process (Fig. 3).
Hyoids: Remnants of the hyoids are preserved adjacent to the atlas-axis (Fig. 4C). They are curved, strap-shaped elements, bearing a flaring, transversely flat posterior end.
Dentition: Each premaxilla bears three and the right maxilla 23 tooth positions. The number of dentary teeth is uncertain as some are obscured. All well preserved tooth crowns are slender, labiolingually compressed and have an elliptical cross-section (Fig. 4A). The enamel surface is smooth, but a faint carina with a low keel extends along the mesial and distal margins of each crown. Some smaller premaxillary and maxillary teeth are replacement teeth.
Atlas-axis vertebrae and ribs: An elongate element adjacent to the first post-axial cervical vertebra most likely represents part of the atlas-axis (Fig. 4C). The bone is exposed in ventral view, but the preservation is poor and it is partly obscured by a rib. It is thus not possible to determine whether the elongate portion represents the axis only or both elements conjoined. On the right side, adjacent to the elongate bone, there is a curved element that might represent the atlantal or axial rib.

Fig. 4. Metriorhynchid thalattosuchian Cricosaurus bambergensis sp. nov. (NKMB-P-Watt14/274) holotype, late Kimmeridgian of Wattendorf (southern Germany). A. Dentition. B. Palatines showing the midline keel seperating symmetrical depressions (dashed outline). C. First cervical vertebrae. Abbreviations: aac, atlas-axis vertebrae; cv, cervical vertebra; hy, hyoid.
Postaxial cervical vertebrae and ribs: An articulated series of five postaxial cervical vertebrae is preserved (Figs. 4C, 5). The dimensions of the centra cannot be provided as they are largely obscured by matrix and bone fragments. The first and second postaxial cervicals are exposed in ventrolateral view (Fig. 4C). The posterior articulation surface of the first centrum is partly visible, indented and surrounded by a thickened rim. The dorsolaterally placed diapophysis can be seen in the first and second vertebrae (SOM 3). It is transversely compressed and less prominent than the ventrolaterally placed parapophysis. The latter has an oval shape and the surface between each dia- and parapophysis is slightly indented. The cervical ribs are only fragmentary preserved.
Dorsal vertebrae and ribs: An articulated series of 18 dorsal vertebrae is present (Fig. 5, SOM 3). Most dorsals are visible in ventral view and are partly obscured by ribs. Only the posterior-most dorsals are exposed in lateral view. The centra are longer than wide/high and have concave ventral sides. Adjacent to the articular faces there are some longitudinal striations. In ventral view, the transverse processes are only slightly posteriorly inclined. Their oval diapophyses are dorsoventrally compressed. The parapophyses are only well preserved in the posterior-most dorsals. They are placed anteriorly on the transverse processes and are smaller than the diapophyses. The zygapophyses on the posterior dorsals exceed the margins of the centra longitudinally with about half of their length. The neural spines in these vertebrae are about as high as the corresponding centra and have slightly concave anterior and posterior edges and a convex dorsal side when seen in lateral view.
The dorsal ribs are preserved adjacent to the centra but the rib heads are largely obscured. The ribs are slender and slightly offset proximally. The distal portion of each rib is transversely compressed.
Gastralia: The disarticulated gastral basket is represented by some curved, strap-shaped elements ventral to the dorsal ribs (Fig. 1, SOM 3).
Sacral vertebrae and ribs: Two articulated sacral vertebrae are preserved and exposed in lateral view (Fig. 1, SOM 3). The centra are longer than high and show longitudinal striations adjacent to the articular faces. The ventral sides of the centra are concave, but they are largely obscured by the sacral ribs. The morphology of the neural arches and neural spines resembles that of the dorsal vertebrae. The sacral ribs are still attached to the centra. The first sacral rib is gently posteriorly inclined and flares distally. The second sacral rib is slightly larger and likewise flares distally. The distal portions of both ribs are obscured by the femur.
Caudal vertebrae and ribs: All caudal vertebrae are preserved in articulation, but the tail is interrupted after the 13th vertebra. The preserved portion incorporates 28 preflexural, six flexural and 20 postflexural caudal vertebrae (Figs. 1, 6; SOM 3). All caudals are preserved in lateral view and have concave ventral sides. The preflexural centra are longer than high but become gradually shorter posteriorly. The articular faces have slightly expanded lateral sides and are gently angled ventrally for the articulation with the hemapophyses. The sub-triangular transverse processes are preserved in the proximal-most caudals and are placed at about the dorsoventral midsection of the vertebrae. They are, however, broken off and pushed downwards, displaying their dorsal aspect. The zygapophyses are reduced in the proximal part of the tail. The proximal caudal hemaphophyses are strap-shaped and first occur between the third and fourth vertebrae. They become rapidly shorter and are missing in the distal-most preflexural caudals. The neural spines in the three proximal-most caudals resemble the morphology of the posterior dorsal and sacral neural spines. In the fourth caudal a prominent anterodorsal process is formed (Fig. 1). The margin between this process and the remainder of the neural spine is excavated. The neural spines of the fifth and sixth preflexural caudals are obscured by the overlying right hindlimb. The anterodorsal process is placed more ventrally in the 7th–9th preflexural caudals, and the remaining neural spines are anteroposteriorly narrower and more posteriorly inclined, leading to a larger excavation between the two parts. The anterior process is also present in the first caudals behind the cut, but here it is reduced and more dorsally oriented. The inclination of the neural spines continues after the cut, but they become gradually lower, more lobate and posteriorly inclined in all of the preflexural vertebrae. In the proximal caudals the neural spines only barely exceed the level of the centra with their posterior portions.
In the post-cut caudals, in the mid-tail section, about 1/3 of neural spine length exceeds the level of each centrum whereas in the distal-most preflexural caudals about half of their length exceeds the centrum posteriorly. The neural spines are here also more posteriorly inclined. The last preflexural centrum is stout, compared with the other preflexural centra, and leaned anteriorly with a slightly higher and more ventrally protruding posterior articular face. In the first flexural centrum the anterior and posterior articular faces are both gently ventromedially inclined and the posterior face again reaches further ventrally than the anterior one. In the remaining caudals the anterior and posterior faces are not noticeably inclined and the posterior one always protrudes slightly more ventrally. The neural spine in the last preflexural caudal is only gently posteriorly inclined and more erected than that of the preceding vertebrae and its prezygapophysis faces anteriorly, whereas the neural spines in all flexural caudals are facing anteriorly (Fig. 6). In the first flexural vertebra the neural spine meets the apex of the shorter preceding neural spine and here the prezygapophyses are likewise anteriorly oriented. The following neural spine is more elongate and the prezygapophysis faces dorsally. The neural spine of the third flexural caudal is about as long as that of the third one but broader, more anteriorly inclined and the prezygapophysis faces dorsally. The neural spines of the 4th–6th flexural caudals become rapidly shorter and the reduced prezygapophyses are facing anterodorsally. Postzygapophyses cannot be seen in any of these vertebrae. In the first and second postflexural caudal vertebrae the neural spines are very reduced and prominently posteriorly inclined, and only remnants of the prezygapophyses are present as small tips. In the remaining postflexural caudals the neural spines are reduced to low straps and no neural spines are present in the last seven vertebrae. The hemapophyses of the last preflexural caudal are formed as a rectangular bar that exceeds the preceding centrum completely. Here a short, posteroventrally inclined hemapophyseal dorsal process is formed. In the 1st–3rd flexural caudals the dorsal processes are longer and more posteriorly inclined and the ventral hemapophyseal parts are lobe-shaped. The same morphology continues in the 4th–6th flexural caudals but the hemapophyseal dorsal processes are less posteriorly inclined and the lobate part becomes smaller. Hemapophyses are visible to the 10th postflexural caudal but the shape changes from lobate to low-rectangular at the intersection between the flexural to the postflexural caudals. There are no hemapophyses visible in the distal-most caudals.
Scapulae: The scapulae are obscured by the coracoids (Fig. 5) and can therefore not be described.
Coracoids: Both coracoids are preserved in ventral view (Fig. 5). They are flat elements with concave anterior and posterior sides. The anterior edge is more strongly concave and almost semi-circular, whereas the posterior one is transversely wider. The mid-section of each coracoid is constricted. The proximal and distal coracoid portions are expanded and have convex edges. A small, circular, distally placed coracoid foramen is present.
Humerus: A fragment preserved adjacent to the right coracoid (Fig. 5) might represent part of a humerus.
Radius and ulna: The right radius and ulna are preserved and overlap each other (Fig. 5). They are flat, circular elements. It is uncertain which of these elements represents the radius and which the ulna.
Carpal elements: The carpal elements are preserved (Fig. 5), but due to disarticulation an exact identification is not possible.
Metacarpal elements: The metacarpal elements are preserved in original arrangement (Fig. 5). The first metacarpal is a crescent-shaped element that is the largest in the row. It has a strongly convex anterior side and a slightly concave posterior one. The second to fourth metacarpals are rod-shaped elements of about equal size. The fifth metacarpal is largely obscured by a rib.
Phalanges (forelimb): Several hourglass-shaped phalanges are preserved (Fig. 5). They are still in original position showing that five digits with two phalanges each are formed. The proximal phalanges are clearly larger and more elongate than the distal ones. The anterior-most proximal phalanx is placed distal to the first metacarpal and is wider than the other phalanges of the forelimb. The distal phalanges have a sub-circular shape.

Fig. 5. Metriorhynchid thalattosuchian Cricosaurus bambergensis sp. nov. (NKMB-P-Watt14/274) holotype, late Kimmeridgian of Wattendorf (southern Germany). Pectoral girdle and forelimb elements. Abbreviations: ca, carpal elements; co, coracoid; cv, cervical vertebra; dv, dorsal vertebra; ?h, questionable humerus; mc, metacarpal elements; ph, phalanges; ru, radius and ulna; sc, scapula.

Fig. 6. Metriorhynchid thalattosuchian Cricosaurus bambergensis sp. nov. (NKMB-P-Watt14/274) holotype, late Kimmeridgian of Wattendorf (southern Germany). Distal part of the tail showing the flexural and postflexural region.
Ilium: The ilium is largely obscured, preventing a description (SOM 3).
Pubes: The pubes are complete, but the right pubis partly overlies the left one (Fig. 7). They are gracile elements having fan-shaped distal portions with a convex distal edge. The anterior and posterior sides are concave, but the pubes are massively constricted at mid-length, having only about ¼ of the expansion of the distal end. The proximal head is only slightly wider than the midshaft in lateral view.
Ischia: The ischia are largely obscured or damaged (SOM 3). The dorsal portion of the right ischium is in original articulation with the femur that largely obscures it. A slightly convex edge is formed posterodorsally, whereas the anterior side is deeply concave from the dorsal acetabular end to the anteroventral portion of the ischium. The latter protrudes slightly more anteriorly than the acetabular head. The ventral side of the left ischium is almost flat.
Femora: The femora are elongate, sigmoidally curved elements (Fig. 7). They are narrowest at about mid-length and lack a fourth trochanter. The proximal end of each femur is obscured by other bones. The distal ends articulating with the tibiae and fibulae are gently convex.
Tibiae: Both tibiae are preserved (Fig. 7). They are stout, long-rectangular elements that are much wider than the fibulae. Proximally each tibia forms most of the contact with the femur. The anterior edges of the tibiae are straight, and the posterior ones are slightly concave. Their distal ends are slightly narrower than the proximal ones.
Fibulae: Both fibulae are preserved in original alignment with the tibiae (Fig. 7). They are narrow, rod-shaped elements with a slightly constricted shaft due to concave anterior and posterior edges. The proximal and distal ends are slightly convex. Tibiae and fibulae are about equal in length.
Tarsal elements: The tarsal elements are sufficiently preserved in the right hindlimb (Fig. 7). Astragalus and calcaneum are slightly displaced. The astragalus, adjacent to the fibula, is kidney-shaped and the largest tarsal element. The calcaneum which contacts the tibia has an oval shape and is slightly smaller than the astragalus. Distal tarsal 3 is placed proximal to the metatarsals 2 and 3 and is smaller than the posteriorly placed distal tarsal 4 that distally contacts the fourth metatarsal.
Metatarsal elements: The metatarsals are preserved in original alignment, along with the phalanges, in both limbs (Fig. 7). The first metatarsal is the widest and shortest element in the row. It is placed slightly more proximally than the other metatarsals and contacts the distal tarsal 3 posteriorly. The proximal end of the first metatarsal is fan like expanded and more than twice as wide as the distal one. The first metatarsal terminates at the level of the distal fourth of the other metatarsal elements, but may be slightly displaced. The second metatarsal is slightly shorter than the third and fourth ones. All are rod-shaped elements with slightly expanded distal and ventral portions. The fifth metatarsal is reduced into a small triangular bone.
Phalanges (hindlimb): The phalanges are preserved in original alignment in the first to fourth digits of both limbs (Fig. 7). The first digit consists of two phalanges, the proximal one being rod-shaped and placed half way between the second metatarsal and first phalanx of the second digit. The distal phalanx of the first digit is pointed and less than half as long as the corresponding proximal phalanx. The second digit has three phalanges. The proximal one is rod-shaped and slightly longer than the according one of the first digit. It is placed at level with the proximal phalanx of the third digit but is slightly shorter. The second phalanx of the second digit is about half as long as the proximal one, but likewise rod-shaped. The terminal phalanx of the second digit again is pointed and ends at level of the distal end of the second phalanx of the third digit. The third digit bears four phalanges, of which the first to third are rod shaped. The proximal one is the largest in the hindlimb. It terminates further distally than the corresponding phalanx of the second digit and so does the shorter third phalanx of the third digit. The fourth phalanx of the third digit is about 1/3 the length of the proximal one, whereas the pointed fourth phalanx is formed as a small tip only. The fourth digit is the longest and formed the trailing edge of the limb. It consists of five phalanges of which the first to third are rod-shaped. The proximal phalanx starts slightly distal to the according one of the third digit. It also terminates slightly further proximal to the later. The same applies to the second and third phalanx of the fourth digit. The short, rectangular fourth phalanx is placed half way between the third and fourth phalanges of the third digit. The pointed fifth phalanx of the fourth digit forms the distal-most part of the entire limb.
Stratigraphic and geographic range.—Wattendorf Member, Torleite Formation, late Kimmeridgian, Wattendorf, Bavaria, southern Germany.
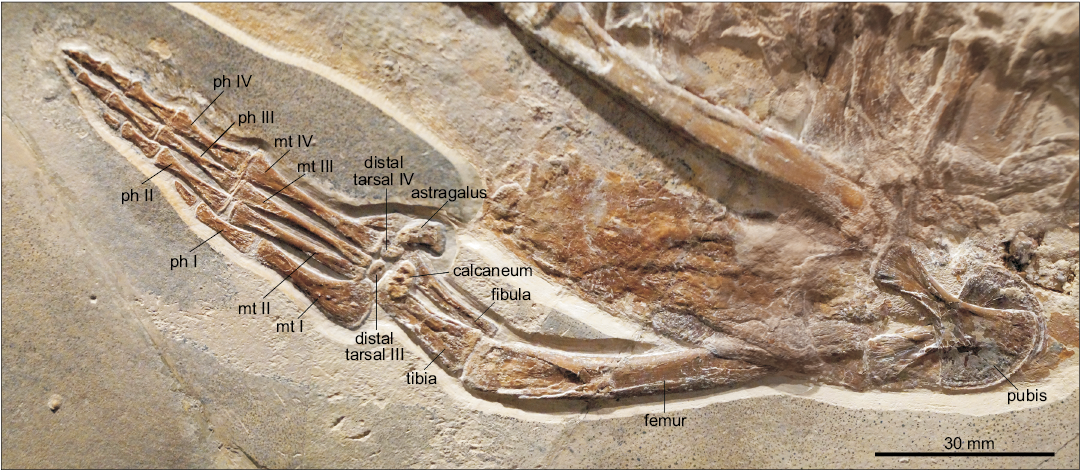
Fig. 7. Metriorhynchid thalattosuchian Cricosaurus bambergensis sp. nov. (NKMB-P-Watt14/274) holotype, late Kimmeridgian of Wattendorf (southern Germany). Hindlimb and pelvic girdle. Abbreviations: mt, metatarsals; ph, phalanges.
Phylogenetic analysis
SOM 1 and SOM 4 include further information and files on the phylogenetic analysis.
A phylogenetic analysis was conducted to assess the evolutionary relationships of Cricosaurus bambergensis sp. nov. within Thalattosuchia. The dataset employed forms part of the ongoing Crocodylomorph SuperMatrix Project. This dataset (referred to as the Hastings + Young matrix, or H + Y matrix by Ristevski et al. 2018 and Ősi et al. 2018) was herein revised to include six new characters and one new operational taxonomic unit (OTU) (C. bambergensis) (SOM 1). Furthermore, we revised the dental characters relating to the carinae and position of the end of the maxillary tooth-row for C. suevicus and C. elegans, and dermatocranial ornamentation characters for C. elegans.
This resulted in the current iteration of the H + Y matrix containing a total of 141 OTUs scored for 460 characters. A total of 26 characters representing morphoclines were treated as ordered (7, 23, 35, 43, 56, 64, 105, 171, 173, 182, 212, 230, 238, 268, 278, 334, 359, 362, 367, 378, 413, 416, 426, 427, 429, 441). Postosuchus kirkpatricki Chatterjee, 1985 was used as the outgroup.
The phylogenetic analyses were conducted following the methodology implemented using freeware TNT v1.5-beta, Willi Hennig Society Edition (Goloboff et al. 2008, Goloboff and Catalano 2016; programme has been made available with the sponsorship of the Willi Hennig Society). Memory settings were increased with General RAM set to 900 Mb and the maximum number of trees to be held set to 99 999.
Cladogram-space was first searched using the advanced search methods in TNT. A driven search was performed with 100 initial addition sequences, and the minimum score set to be found 10 times. The default settings for the advanced search methods were changed to increase the iterations of each method per replicate (except for tree fusion, which was kept at 3 rounds).
Sectorial search: 10 sectorial search drifting cycles for selections of above 75, and below 75 it used 10 starts and fuse trees 10 times; 10 rounds of consensus sectorial searches (CSS); and 10 rounds of exclusive sectorial searches (XSS).
Ratchet: 50 ratchet iterations, and stops perturbation phase when 50 substitutions are made or 99% of swapping is completed.
Drift: 50 drift cycles, and stops perturbation phase when 50 substitutions are made or 99% of swapping is completed.
The resultant cladograms from the driven advanced methods search were the basis for the final run. A traditional methods tree bisection and reconnection (TBR) search was performed on the most parsimonious cladograms (MPCs) saved in the RAM. This gave the final set of MPCs. Nodal support was assessed by performing Bremer support and bootstrap analyses (for bootstrap values, Bremer support values and nexus file see SOM 4), with the latter set to 1000 random replicates of advanced methods searching (default settings and 10 addition replicates).
The phylogenetic analysis recovered 2592 MPCs with 1508 steps (ensemble consistency index CI = 0.417; ensemble retention index RI = 0.843; rescaled consistency index RC = 0.352; ensemble homoplasy index HI = 0.583). Overall, the strict consensus topology recovered here (Fig. 8) is very similar to those presented in Ristevski et al. (2018) and Ősi et al. (2018). The only differences within Thalattosuchia are: (i) the addition of Cricosaurus bambergensis, (ii) that C. suevicus is no longer being recovered within a large polytomy with the Argentinian, Mexican and Cretaceous Cricosaurus species, and (iii) the recovery of a new subclade, consisting of a polytomy of C. bambergensis, C. suevicus and C. elegans.
Previously C. elegans was found to be the basal-most species of Cricosaurus, and sister taxon to a polytomy of (C. suevicus, C. araucanensis, C. lithographicus, C. vignaudi and C. schroederi) (e.g., Ősi et al. 2018). However, now all of the late Kimmeridgian–early Tithonian Cricosaurus species from Germany form a clade, which is the sister taxon to polytomy of (C. araucanensis, C. lithographicus, C. vignaudi, and C. schroederi) (Fig. 8).
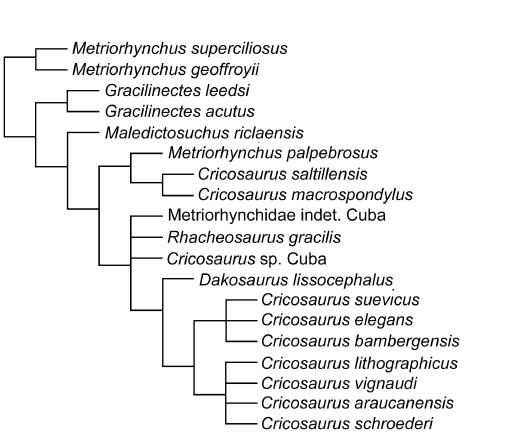
Fig. 8. Results of the phylogenetic analysis, simplified strict consensus topology of 2592 most parsimonious cladograms of Cricosaurus bambergensis sp. nov. within Metriorhynchinae (step number = 1.508; CI = 0.417; RI = 0.843; RC = 0.352; HI = 0.583).
Discussion
Southern German species of Cricosaurus.—Our description of Cricosaurus bambergensis (NKMB-P-Watt14/274), herein shows that there was at least three species of Cricosaurus present in the late Kimmeridgian–early Tithonian of southern Germany. Cranially, C. bambergensis differs from C. elegans, C. medius, and C. suevicus in that: (i) the maxillary row continues posteriorly past the anterior margin of the orbit (a thalattosuchian symplesiomorphy) and (ii) palatines have a distinct midline ridge with paired depressions either side (autapomorphy, but unknown for C. medius and C. suevicus). The tail displacement unit is also unique in C. bambergensis. The distal-most three preflexural vertebrae have neural spines that are oriented very strongly posteriorly, such that their posterior margins lie on top of the prezygapophyses of the posterior vertebra. These neural spines look almost rod-like in lateral view. The three vertebrae in front of them have a similar morphology, but the neural spines look more like a “regular” neural spine, albeit strongly oriented posteriorly. The tail fluke of C. bambergensis lacks the four apomorphies of C. suevicus, retaining the following symplesiomorphies: (i) the distal-most preflexural caudal vertebra (i.e., the vertebra with the verticalized neural spine immediately anterior to the tail bend) has a rod-like hemapophysis; (ii) the anterior-most five or six flexural caudal vertebrae have anteriorly oriented neural spines, but not strongly oriented anteriorly; (iii) the flexural hemapophyses contact one another along their anterior-posterior margins; and (iv) the distal-most postflexural hemapophysis differs, returning to a “rod-like” morphology.
Cricosaurus elegans (BSPG AS I 504), C. medius (BSPG AS VI 2), and C. suevicus (SMNS 9808) share: (i) the maxillary tooth row terminating anterior to the anterior margin of the orbits and (ii) the lack of pronounced, or regular shallow, reception pits on the lateral margins of the maxillary mid-portion and dentary. Unfortunately, there are no complete or nearly complete postcranial remains for C. elegans or C. medius. However, the vertebral column of C. suevicus (GPIT 7322, SMNS 9808, SMNS 90513) is apomorphic: (i) all the neural spines of the dorsal vertebrae are rectangular when seen in lateral view, with a distinctly flat dorsal margin being prevalent, and the neural spines of adjacent vertebrae are in close proximity to one another (autapomorphy); and (ii) the composition of the tail displacement unit is autapomorphic. In C. suevicus (GPIT 7322, SMNS 9808, SMNS 90513) the distal-most seven preflexural vertebrae have neural spines that are oriented very strongly posteriorly, with their posterior margin lying on top of the prezygapophyses of the subsequent vertebra. These neural spines look almost rod-like in lateral view. The three vertebrae immediately anterior to them have a similar morphology, but the neural spines are raised such that they do not overlap the prezygapophyses of the vertebrae immediately posterior.
Moreover, C. suevicus (GPIT 7322, SMNS 9808) has the following tail fluke apomorphies: (i) the distal-most preflexural caudal vertebra has a dorsoventrally deep hemapophysis, with a midline flange (i.e., it does not have a ”rod-like” hemapophysis as in C. bambergensis and Rhacheosaurus gracilis Von Meyer, 1831, NHMUK PV R 3948); (ii) the anterior-most five or six flexural caudal vertebrae have strongly anteriorly oriented neural spines (much more so than C. bambergensis, R. gracilis, NHMUK PV R 3948, and Metriorhynchus superciliosus de Blainville, 1853, GPIT RE 119314); (iii) the flexural hemapophyses contact one another along their posteroventral-anterodorsal margins (not their posterior–anterior margins as in C. bambergensis, R. gracilis, NHMUK PV R 3948 and M. superciliosus, GPIT RE 119314); and (iv) the distal-most postflexural hemapophyses retain their dorsoventrally deep profile (i.e., they do not return to a “rod-like” morphology, as in C. bambergensis and R. gracilis, NHMUK PV R 3948).
Characteristics that diagnose the new Cricosaurus species.—Palatal morphology: One unique morphology seen in the Bamberg specimen is the presence of a distinct midline ridge along the palatines, with paired depressions on either side. This character has not been observed in any other thalattosuchian (e.g., Andrews 1913; Parrilla-Bel et al. 2013; Herrera et al. 2013b). However, most of the Cricosaurus specimens found in Germany do not preserve the palate. The only specimen that exposes the palatines is the C. elegans holotype BSPG AS I 504, which has the thalattosuchian “normal” condition with no midline ridge or paired depressions (Fig. 9B). The Argentinian species C. araucanensis and C. lithographicus also lack this ridge-depression morphology (Gasparini and Dellapé 1976; Herrera et al. 2013b).

Fig. 9. Comparative images showing the ventral side of the skull and a close-up of the palatal region of metriorhynchid thalattosuchians Cricosaurus bambergensis sp. nov. (NKMB-P-Watt14/274, A) from the late Kimmeridgian of Wattendorf (southern Germany) and Cricosaurus elegans (BSPG AS I 504, B) from the lower Tithonian of Dating (southern Germany).
Tail displacement unit: The tails of metriorhynchids are highly regionalised, the classic example being the tail fluke. However, there is always a series of caudal vertebrae positioned intermediate between the proximal region (or tail stock sensu Lindgren et al. [2007], which have the symplesiomorphic dorsally oriented neural spines) and the tail bend itself. These vertebrae show a shift in neural spine shape and orientation (e.g., see Fraas 1902; Andrews 1913). Herein, we use the term “displacement unit” for this region (following the usage in mosasaurids, Lindgren et al. 2007). The displacement units of Cricosaurus bambergensis and Cricosaurus suevicus are structurally similar: the distal-most preflexural vertebrae have almost rod-like neural spines that are oriented very strongly posteriorly, such that their posterior margins lie on top of the prezygapophyses of the posterior vertebra. C. bambergensis has three such vertebrae, whereas C. suevicus (GPIT 7322, SMNS 9808, SMNS 90513) has seven. In front of these, are vertebrae with a similar morphology, but the neural spines look more like a “regular” neural spine and they no longer come in contact with the prezygapophyses of the posterior vertebra. For both species, there are three such vertebrae.
Tail fluke: Additional diagnostic traits are present in the tail fluke. The latter is well known for Cricosaurus suevicus (SMNS 9808, GPIT RE 7322) where the tail bend is deeper than in Cricosaurus bambergensis. This is caused by multiple morphological differences. The first concerns the distal-most preflexural caudal vertebra, in C. bambergensis, this vertebra has a rod-like hemapophysis (as with Rhacheosaurus gracilis NHMUK PV R 3948), whereas in C. suevicus these hemapophyses have the classic midline flange seen in Cricosaurus postflexural hemapophyses, which expands soft-tissue attachment area (SMNS 9808, GPIT RE 7322, Fraas, 1902).
In metriorhynchids, the proximal-most five or six flexural caudal vertebrae have anteriorly oriented neural spines. These neural spines help support the soft tissue upper lobe (e.g., see NHMUK PV R 3948, Young et al. 2010: fig. 4a). In C. bambergensis, they are not as strongly oriented anteriorly as in C. suevicus.
In C. bambergensis the flexural hemapophyseal contact one another along their anterior–posterior margins. In C. suevicus these hemapophyses contact one another on their anterodorsal–posteroventral margins, a result of the tail bend being at a steeper angle.
Finally, shape of the distal-most postflexural hemapophyses differs between C. bambergensis and C. suevicus. In C. bambergensis these hemapophyses return to a “rod-like” morphology, whereas in C. suevicus these hemapophyses become oval-shaped, with a more elongate posterior process than anterior process, and retain some dorsoventral depth.
Dentition: Interestingly, all metriorhynchids have three alveoli per premaxilla (e.g., Andrews 1913). However, Fraas (1902) was unclear whether the lectotype of Cricosaurus suevicus (SMNS 9808) had four. Unfortunately, the premaxillary lateral margin of SMNS 9808 is damaged, and as such it is impossible to be certain where the premaxilla–maxilla suture is. None of the other specimens referred to C. elegans or C. suevicus show any evidence of having four premaxillary teeth. The holotype of C. bambergensis also has only three premaxillary alveoli.
The lack of mesial and distal carinae on the dentition has been considered to be a diagnostic character of Rhacheosaurini (Young et al. 2011; Parrilla-Bel et al. 2013). However, the Tithonian species from Argentina (C. araucanensis and C. lithographicus) and Mexico (C. vignaudi and “C.” saltillensis) have bicarinate dentition (Frey et al. 2002; Buchy et al. 2013; Herrera et al. 2013b), as do the Valanginian species C. schroederi and “C.”/Enaliosuchus macrospondylus (SS personal observation; Hua et al. 2000). The basal rhacheosaurin Maledictosuchus nuyivijianan Barrientos-Lara, Alvarado-Ortega, and Fernández, 2018 also has carinated dentition. Based on our examination of the German Cricosaurus material, Cricosaurus bambergensis, C. elegans, C. medius, and C. suevicus do indeed have carinae, they are faint with a low keel (SMNS 9808, BSPG AS I 504, BSPG AS VI 2, NHMUK PV OR 37006). As such, the lack of carinae is not diagnostic for either Rhacheosaurini or Cricosaurus. In fact, the presence of faint carinae (a low carinal keel) appears to be diagnostic for Cricosaurus.
Species complex, chronospecies or anagenetic lineage?— The discovery of a Cricosaurus subclade from the late Kimmeridgian–early Tithonian raises the question about the nature of this lineage. Overall, C. bambergensis, C. suevicus, C. medius, and C. elegans are very similar in body size and osteology. If not for the unusual palatal and tail morphologies of C. bambergensis, we could easily envision this subclade to be an anagenetic lineage, in which the tail fluke becomes steeper and has increased the surface area for soft tissue attachment over time. While this could be the case, we plan to examine further specimens from southern Germany, and re-evaluate the type specimens of C. suevicus and C. elegans. This should help us address these issues.
Regardless, the palate morphology of C. bambergensis is both curious and unique. The presence of paired depressions on the palate suggests this is a soft-tissue structure. It possibly could be paired pneumatic diverticula, or concentration of blood vessels and/or nerves. Unfortunately, as the posterior boundary of this structure is not preserved, we cannot hypothesise its nature (or preclude it being individual variation). But, this character and the tail fluke being morphologically intermediate between more basal metriorhynchines and C. suevicus, make C. bambergensis distinct from all other known metriorhynchids.
We were recently made aware that the holotype of Cricosaurus medius, BSPG AS VI 2, had been rediscovered (Yanina Herrera, personal communication 2019). Young and Andrade (2009) had considered the holotype of C. medius to be “apparently lost”. This raises the question about the validity of C. elegans, C. medius, and C. suevicus. Cranially, all three species share the same suite of characters: the maxillary tooth row terminated prior to the orbital anterior margin, and the lack of well-developed reception pits along the dentigerous bones (both in marked contrast with C. bambergensis). Addressing this question is beyond the scope of this paper, as it will require the type specimens of all three species to be re-examined and re-described.
Conclusions
Herein we describe a new species of metriorhynchid, Cricosaurus bambergensis sp. nov., from the late Kimmeridgian (Wattendorf Member, Torleite Formation) of southern Germany. Uniquely, it has palatines with a midline ridge with paired depressions on either side, and a tail fluke no steeper than that of Rhacheosaurus gracilis. Three other species of Cricosaurus have been named from the Late Jurassic of southern Germany; however, C. bambergensis can be differentiated from all of them. The first, C. suevicus, is known from the late Kimmeridgian Nusplingen Formation, had a steeper tail fluke and longer tail displacement unit, the maxillary tooth row terminates prior to the orbits, and there is a lack of well-developed reception pits along the dentigerous bones, but the palate morphology is unknown. The second, C. elegans (known from the early Tithonian Altmühltal and Mörnsheim formations), had the “normal” thalattosuchian palatine condition, the maxillary tooth row terminates prior to the orbits, and there is a lack of well-developed reception pits along the dentigerous bones, but the tail is unknown. The final named species, C. medius, is also known from the early Tithonian Mörnsheim Formation, the maxillary tooth row terminates prior to the orbits, and there is a lack of well-developed reception pits along the dentigerous bones; but the palate and tail are unknown. Determining whether C. suevicus, C. medius, and C. elegans are conspecific is beyond the scope of this paper, but our phylogenetic analysis does recover the southern Germany species in an unresolved clade at the base of Cricosaurus.
Acknowledgements
We are grateful to Matthias Mäuser and Thomas Bechmann (both NKMB) and Oliver Rauhut (BSGP) for providing access to the specimen under his care. We also thank the reviewers Yanina Herrera (National University of La Plata, Buenos Aires, Argentina), Christopher Brochu (University of Iowa, Iowa City, USA), and an anonymous referee for their helpful comments which improved our manuscript. MTY is financially supported by a Leverhulme Trust Research Project grant (RPG-2017-167).
References
Andrews, C.W. 1913. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay, Part II. 206 pp. British Museum (Natural History), London.
Barrientos-Lara, J.I., Alvarado-Ortega, J., and Fernández, M.S. 2018. The marine crocodile Maledictosuchus (Thalattosuchia, Metriorhynchidae) from the Kimmeridgian deposits of Tlaxiaco, Oaxaca, southern Mexico. Journal of Vertebrate Paleontology [published online 10.1080/02724634.2018.1478419]. Crossref
Buchy, M.-C., Vignaud, P., Frey, E., Stinnesbeck, W., and González, A.H.G. 2006. A new thalattosuchian crocodyliform from the Tithonian (Upper Jurassic) of northeastern Mexico. Comptes Rendus Palevol 5: 785–794. Crossref
Buchy, M.-C., Young, M.T., and Andrade, M.B. 2013. A new specimen of Cricosaurus saltillensis (Crocodylomorpha: Metriorhynchidae) from the Upper Jurassic of Mexico: evidence for craniofacial convergence within Metriorhynchidae. Oryctos 10: 9–21.
Chatterjee, S. 1985. Postosuchus, a new thecodontian reptile from the Triassic of Texas and the origin of tyrannosaurs. Philosophical Transactions of the Royal Society of London B 309: 395–460. Crossref
Cuvier, G. 1824. Sur les ossements fossiles de crocodiles. In: G. Dufour and E. D’Occagne (eds.), Recherches sur les ossements fossiles, Second édition, 143–160. Dufour and d’Occagne, Paris. Crossref
de Blainville H.D. 1853. Letter by H.D. de Blainville. In: J.A. Eudes-Deslongchamps (ed.), Lettres sur les crocodiles vivants et fossiles. Bulletin de la Société Linnéenne de Normandie (Années 1849–1853) IX: 109–120.
Fitzinger, L.J.F.J. 1843. Systema Reptilium. 136 pp. Braumüller et Seidel, Vienna.
Fraas, E. 1901. Die Meerkrokodile (Thalattosuchia n. g.) eine neue Sauriergruppe der Juraformation. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 57: 409–418.
Fraas, E. 1902. Die Meer-Krocodilier (Thalattosuchia) des oberen Jura unter specieller Berücksichtigung von Dacosaurus und Geosaurus. Palaeontographica 49: 1–72.
Frey, E., Buchy, M.-C., Stinnesbeck, W., and López Oliva, J.G. 2002. Geosaurus vignaudi n.sp. (Crocodyliformes: Thalattosuchia), first evidence of metriorhynchid crocodilians in the Late Jurassic (Tithonian) of centraleast Mexico (Puebla). Canadian Journal of Earth Sciences 39: 1467–1483. Crossref
Fürsich, F.T., Werner, W., Schneider, S., and Mäuser, M. 2007. Sedimentology, taphonomy, and palaeoecology of a laminated plattenkalk from the Kimmeridgian of the northern Franconian Alb (southern Germany). Palaeogeography, Palaeoclimatology, Palaeoecology 243: 92–117. Crossref
Gasparini, Z.B. and Dellapé, D. 1976. Un nuevo cocodrilo marino (Thalattosuchia, Metriorhynchidae) de la Formación Vaca Muerta (Jurasico, Tithoniano) de la Provincia de Neuquén (República Argentina). Congreso Geológico Chileno, Actas 1: c1–c21.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Hay, O.P. 1930. Second Bibliography and Catalogue of the Fossil Vertebrata of North America. 1074 pp. Carnegie Institute Washington, Washington DC.
Herrera, Y., Fernández, M.S., and Gasparini, Z. 2013a. Postcranial skeleton of Cricosaurus araucanensis (Crocodyliformes: Thalattosuchia): morphology and palaeobiological insights. Alcheringa 37: 285–298. Crossref
Herrera, Y., Fernández, M.S., Lamas, S.G., Campos, L., Talevi, M., and Gasparini, Z. 2017. Morphology of the sacral region and reproductive strategies of Metriorhynchidae: A counter-inductive approach. Transactions of the Royal Society of Edinburgh, Earth and Environmental Science 106: 247–255. Crossref
Herrera, Y., Fernández, M.S., and Varela, J.A. 2009. Morfología del miembro anterior de Geosaurus araucanensis Gasparini y Dellapé, 1976 (Crocodyliformes: Thalattosuchia). Ameghiniana 46: 657–667.
Herrera, Y., Gasparini, Z., and Fernandez, M.S. 2013b. A new Patagonian species of Cricosaurus (Crocodyliformes, Thalattosuchia): first evidence of Cricosaurus in Middle–Upper Tithonian lithographic limestones from Gondwana. Palaeontology 56: 663–678. Crossref
Hua, S., Vignaud, P., Atrops, F., and Clément, A. 2000. Enaliosuchus macrospondylus Koken, 1883 (Crocodylia, Metriorhynchidae) du Valanginien de Barret-le-Bas (Hautes Alpes, France): un cas unique de remontée des narines externs parmi les crocodiliens. Géobios 33: 467–474. Crossref
Koken, E. 1883. Die Reptilien der norddeutschen unteren Kreide. Zeitschrift der deutschen Geologischen Gesellschaft 35: 735–827.
Kuhn, O. 1936. Crocodilia. Fossilium Catalogus I: Animalia. 114 pp. Junk, Gracenhage.
Lindgren, J., Jagt, J.W.M., and Caldwell, M.W. 2007. A fishy mosasaur: the axial skeleton of Plotosaurus (Reptilia, Squamata) reassessed. Lethaia 40: 153–160. Crossref
Mäuser, M. 2014. Frankenland am Jurastrand. 60 pp. Pfeil Verlag, Munich.
Mäuser, M. 2015. Das Wattendorfer Etappenkrokodil. Fossilien 6: 31–37.
Nesbitt, S. J. 2011. The early evolution of Archosauria: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Niebuhr, B. and Pürner, T. 2014. Plattenkalk und Frankendolomit—Lithostratigraphie der Weißjura-Gruppe der Frankenalb (außeralpiner Oberjura, Bayern). Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 83: 5–72.
Ősi, M., Young, M.T., Galácz, A., and Rabi, M. 2018. A new large-bodied thalattosuchian crocodyliform from the Lower Jurassic (Toarcian) of Hungary, with further evidence of the mosaic acquisition of marine adaptations in Metriorhynchoidea. PeerJ 6: e4668. Crossref
Parrilla-Bel, J., Young, M.T., Moreno-Azanza, M., and Canudo, J.I. 2013. The first metriorhynchid crocodylomorph from the Middle Jurassic of Spain, with implications for evolution of the subclade Rhacheosaurini. PLoS ONE 8: e54275 Crossref
Ristevski, J., Young, M.T., Andrade, M.B., and Hastings, A.K. 2018. A new species of Anteophthalmosuchus (Crocodylomorpha, Goniopholididae) from the Lower Cretaceous of the Isle of Wight, United Kingdom, and a review of the genus. Cretaceous Research 84: 340–383. Crossref
Röper, M. 2005. Field Trip B: East Bavarian Plattenkalk-different types of Upper Kimmeridgian to Lower Tithonian Plattenkalk deposits and facies. Zitteliana B26: 57–70.
Schmid, D.U., Leinfelder, R.R., and Schweigert, G. 2005. Stratigraphy and palaeoenvironments of the Upper Jurassic of Southern Germany—a review. Zitteliana B26: 31–41.
Schweigert, G. 2007. Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany—first results and open questions. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 245: 117–125. Crossref
Von Meyer, C.E.H. 1831. Neue fossile Reptilien aus der Ordung der Saurier. Nova Acta Academiae Leopoldino-Carolinae Curios 15: 173–184.
Wagner, A. 1852. Neu-aufgefundene Saurier-Ueberreste aus den lithographischer Schiefern und den obern Jurakalke. Abhandlungen der Mathemathisch-Physikalischen Classe der Königlich Bayerischen Akademie der Wissenschaften 6: 661–710.
Wagner, A. 1858. Zur Kenntniss der Saurier aus den lithographischen Schiefern. Abhandlungen der Mathemathisch-Physikalischen Classe der Königlich Bayerischen Akademie der Wissenschaften 8: 415–528.
Young, M.T. and Andrade, M.B. 2009. What is Geosaurus? Redescription of Geosaurus giganteus (Thalattosuchia: Metriorhynchidae) from the Upper Jurassic of Bayern, Germany. Zoological Journal of the Linnean Society 157: 551–585. Crossref
Young, M.T., Bell, M.A., and Brusatte, S.L. 2011. Craniofacial form and function in Metriorhynchidae (Crocodylomorpha: Thalattosuchia): modelling phenotypic evolution with maximum-likelihood methods. Biology letters 7: 913–916. Crossref
Young, M.T., Brusatte, S.L., Ruta, M., and Andrade, M.B. 2010. The evolution of Metriorhynchoidea (Mesoeucrocodylia, Thalattosuchia): an integrated approach using geometric morphometrics, analysis of disparity and biomechanics. Zoological Journal of the Linnean Society 158: 801–859. Crossref
Acta Palaeontol. Pol. 64 (2): 343–356, 2019
https://doi.org/10.4202/app.00541.2018