
Mississippian chondrichthyan fishes from the area of Krzeszowice, southern Poland
MICHAŁ GINTER and MICHAŁ ZŁOTNIK
Ginter, M. and Złotnik, M. 2019. Mississippian chondrichthyan fishes from the area of Krzeszowice, southern Poland. Acta Palaeontologica Polonica 64 (3): 549–564.
Two new assemblages of Mississippian pelagic chondrichthyan microremains were recovered from the pelagic limestone of the area of Krzeszowice, NW of Kraków, Poland. The older assemblage represents the upper Tournaisian of Czatkowice Quarry and the younger one the upper Viséan of the Czernka stream valley at Czerna. The teeth of symmoriiform Falcatidae are the major component of both collections. A comparison of the taxonomic composition of the assemblage from Czerna (with the falcatids and Thrinacodus as the major components) to the previously published materials from the Holy Cross Mountains (Poland), Muhua (southern China), and Grand Canyon (Northern Arizona, USA) revealed the closest similarity to the first of these, probably deposited in a deeper water environment, relatively far from submarine carbonate platforms. A short review of Mississippian falcatids shows that the late Viséan–Serpukhovian period was the time of the greatest diversity of this group.
Key words: Chondrichthyes, Falcatidae, teeth, Carboniferous, Tournaisian, Viséan, Poland, Kraków Upland.
Michał Ginter [m.ginter@uw.edu.pl] and Michał Złotnik [m.zlotnik@uw.edu.pl], Faculty of Geology, University of Warsaw, Żwirki i Wigury 93, 02-089 Warszawa, Poland.
Received 27 March 2019, accepted 30 April 2019, available online 23 August 2019.
Copyright © 2019 M. Ginter and M. Złotnik. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The outcrops of Mississippian carbonates are rare in Poland, particularly compared to Belgium or the British Isles, and our knowledge of Tournaisian and Viséan fauna from this territory, and among them chondrichthyans, is relatively poor. Only recently, the first extensive description of a rich assemblage of pelagic sharks from the upper Viséan of western Holy Cross Mountains (Todowa Grząba; Ginter et al. 2015) has been published. Here we present the results of a study on two much smaller samples from another region of southern Poland, the area of Krzeszowice, NW of Kraków (Fig. 1A). There, Mississippian limestone crops out in the valleys of small rivers near the village of Czerna and is currently mined in a huge Czatkowice Quarry, between Czerna and Krzeszowice.
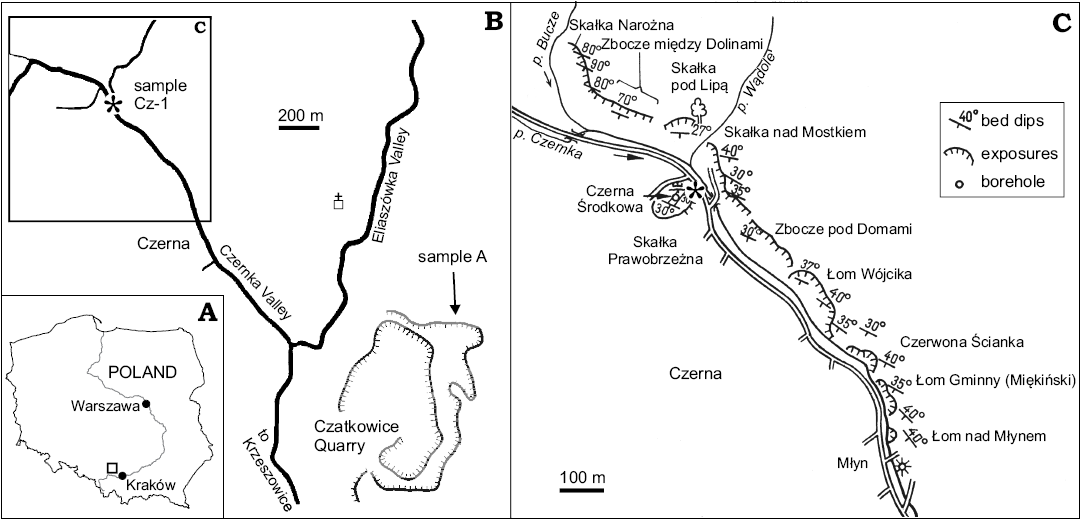
Fig. 1. A. Position of the study area (square) against the outline of Poland and Vistula River. B. Approximate positions of the studied samples: sample A from Czatkowice Quarry (arrow) and Cz-1 from Czerna (asterisk); black lines, paved roads, open square with a cross, Carmelites’ Monastery. C. Close-up of the area at NW Czerna (marked in B) showing the outcrops of Mississippian limestone accessible around 1970; asterisk, possible position of sample Cz-1; from Zajączkowski (1975), modified.
The vertebrate microfossils from the mentioned samples were loaned to MG at the beginning of 1990s as by-products of the search for conodonts. The first sample, from the Tournaisian of Czatkowice Quarry (sample A) was collected by Joanna Appelt, then an M.Sc. student at the Faculty of Geology, University of Warsaw. The second, from the Viséan of Czerna (sample Cz-1), was collected by Jerzy Dzik (Institute of Palaeobiology, Polish Academy of Sciences). The preliminary lists of vertebrate taxa from these samples were presented in unpublished dissertations (Ginter 1994; Appelt 1995) and, much later, in a conference field-trip guidebook (Skompski 2008). Illustrations of selected specimens were published twice as examples (Ginter 1995: fig. 5B, F, H; Ginter et al. 2010: fig. 33F). However, the systematic description of complete materials has not been provided thus far and to fill this gap is the main aim of this paper. At the end, a short review of Mississippian record of the symmoriiform family Falcatidae is added, placing the newly described materials in a more general context.
Institutional abbreviations.—CM, Carnegie Museum, Pittsburgh, USA; MWGUW, Museum of the Faculty of Geology, University of Warsaw, Poland; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; ZPAL, Institute of Palaeobiology, Polish Academy of Sciences, Warsaw, Poland.
Geological setting and overview of the material
Czatkowice Quarry.—This large, active quarry is situated between Czerna and Czatkowice, the northern suburb of Krzeszowice (Fig. 1B). The major part of the Tournaisian–Viséan carbonate sequence exposed in the quarry represents the carbonate ramp and platform environments. In the entire succession, there is only one lithological unit deposited in an intra-shelf basin and composed of graded, spiculitic limestone, named as the Przy Granicy Quarry Formation (Appelt 1998). This spiculitic limestone unit was perfectly exposed at the time of Appelt’s (1995, 1998) study and still in 2008 (MG personal observation) in the northern wall of the quarry exploitation level +370. Sample A, the source of vertebrate material described herein, was collected from the basal part of the unit. Its lithology was described as “peloid wackestone with bioclasts: brachiopod shells and spines, crinoids, single bryozoans, echinoid spines, foraminifers and sponge spicules” (Appelt 1995: 32) and the conodont fauna indicates the upper Tournaisian, Gnathodus cuneiformis Conodont Zone. By the time of publication of a fragment of her thesis, Appelt (1998) changed the numbering of her samples. Most probably, sample A corresponds to the new number Cz2. However, we decided to keep the original number.
In the sample, there were found 18 chondrichthyan teeth, belonging mostly to Thrinacodus, falcatids, and unidentified euselachians with protacrodont crowns (Table 1). There also occur several chondrichthyan branchial denticles, chondrichthyan, actinopterygian and acanthodian scales, and a piece of actinopterygian jaw. The state of preservation of the microfossils is rather poor.
Table 1. Vertebrate microremains in the samples from the upper Tournaisian of Czatkowice Quarry and the upper Viséan of Czerna.
|
Taxon/Sample |
Czatkowice |
Czerna |
|
Thrinacodus ferox asymmetrical |
4 |
5 |
|
Thrinacodus ferox symmetrical |
1 |
0 |
|
Jalodus sp. |
1 |
0 |
|
Denaea wangi |
0 |
3 |
|
Falcatidae indet. |
8 |
12 |
|
Ctenacanthiformes indet. |
0 |
1 |
|
Cladodontomorphi indet. |
2 |
3 |
|
Cassisodus sp. |
0 |
3 |
|
Protacrodontidae indet. |
2 |
0 |
|
Total chondrichthyan teeth |
18 |
27 |
|
ctenacanthoid scales |
– |
+ |
|
hybodontoid scales |
+? |
+ |
|
chondrichthyan branchial denticles |
+ |
– |
|
actinopterygian scales |
+ |
+ |
|
actinopterygian teeth |
+ |
+ |
|
actinopterygian vertebrae and bone fragments |
+ |
+ |
|
acanthodian scales |
+ |
– |
Czerna.—The village of Czerna extends on the slopes of the valley of the Czernka stream, running roughly in the NW-SE direction. The eastern bank of the stream is currently largely covered by forest. However, a few decades ago, there occurred numerous outcrops of Mississippian limestone (Fig. 1C), partly natural and partly due to the mining for local purposes (proposed geosite by Alexandrowicz and Alexandrowicz 1999). According to Zajączkowski (1975), the exposed rocks represent the interval from the upper Tournaisian through to the upper Viséan. Gromczakiewicz-Łomnicka (1974) reported conodonts indicating British zones Gnathodus mononodosus (= Lochriea mononodosa) and Gnathodus girtyi collinsoni, i.e., the uppermost Viséan, from the lower part of the valley. She studied mainly the outcrops called Czerwona Ścianka (= Red Wall) and Łom nad Młynem (= Over the Mill Quarry). The Viséan age of the rocks cropping out in the middle and lower part of the valley also was confirmed by Soboń-Podgórska (1975), based on foraminifers.
According to Jerzy Dzik (personal communication 2014), his sample Cz-1 was collected from a limestone layer cropping out not far from the road, at the beginning of a long escarpment, in the middle part of the valley. We interpret this description as the Czerna Środkowa (= Middle Czerna) outcrop or somewhere close to the former southern part of Łom Wójcika (= Wójcik’s Pit). The conodonts found in the sample Cz-1 indicate the Gnathodus bilineatus Conodont Zone (late Viséan, late Asbian or early Brigantian; Jerzy Dzik personal communication 1993, see also Dzik 1997) which corresponds to the age given by Zajączkowski (1975) for the rocks exposed at Łom Wójcika (Viséan V2b–V3c) and to the Czerwona Ścianka Formation sensu Paszkowski (1995: 27–28). Unfortunately, no precise information concerning the lithology of the sample is available to us and despite the intensive sampling in the area in the years 2016–2018 we did not manage to repeat Dzik’s result.
The vertebrate assemblage from the sample Cz-1 contains 27 chondrichthyan teeth, and among them asymmetrical teeth of Thrinacodus, two types of falcatids, a few other cladodonts and three crushing euselachian teeth (Table 1). In addition, there occur ten chondrichthyan scales and numerous actinopterygian microfossils: scales, teeth, vertebrae and other bone fragments. The ichthyoliths are usually slightly broken which indicates that they were deposited in a moderately energetic environment.
Systematic palaeontology
Class Chondrichthyes Huxley, 1880
Order Phoebodontiformes Ginter, Hairapetian, and Klug, 2002
Family Phoebodontidae Williams in Zangerl, 1981
Genus Thrinacodus St. John and Worthen, 1875
Type species: Thrinacodus nanus St. John and Worthen, 1875; Tournaisian, Burlington, Iowa, USA.
Thrinacodus ferox (Turner, 1982)
Fig. 2.
Material.—Ten teeth: five (ZPAL P. IV/301, 302, 321–323) from the upper Viséan, Czerna, sample Cz-1 and five (MWGUW/Ps/1/212–214, MWGUW/Ps/13/1, 18) from the upper Tournaisian, Czatkowice, sample A.
Remarks.—Most of the teeth of Thrinacodus ferox found at Czerna and Czatkowice are classical asymmetrical forms with the distal cusp much larger than the other two (Fig. 2B, C, E, F). However, in sample Cz-1 there also occurs a tooth (Fig. 2A) somewhat similar to Thrinacodus dziki (Ginter et al. 2015), with the size difference between the delicate, sigmoidal cusps less conspicuous. In sample A from Czatkowice a minute, almost symmetrical tooth was found (Fig. 2D), closely resembling that illustrated by Turner (1982: fig. 3A). Such teeth most probably represent anterior, parasymphyseal tooth families.
The teeth strongly vary in size: the length of the base in the smallest is about 0.3 mm, but in the largest it reaches 4 mm. The largest teeth are abraded and their cusps are broken.
Stratigraphic and geographic range.—Cosmopolitan in the Tournaisian, rare occurrences in the uppermost Famennian and the Viséan.
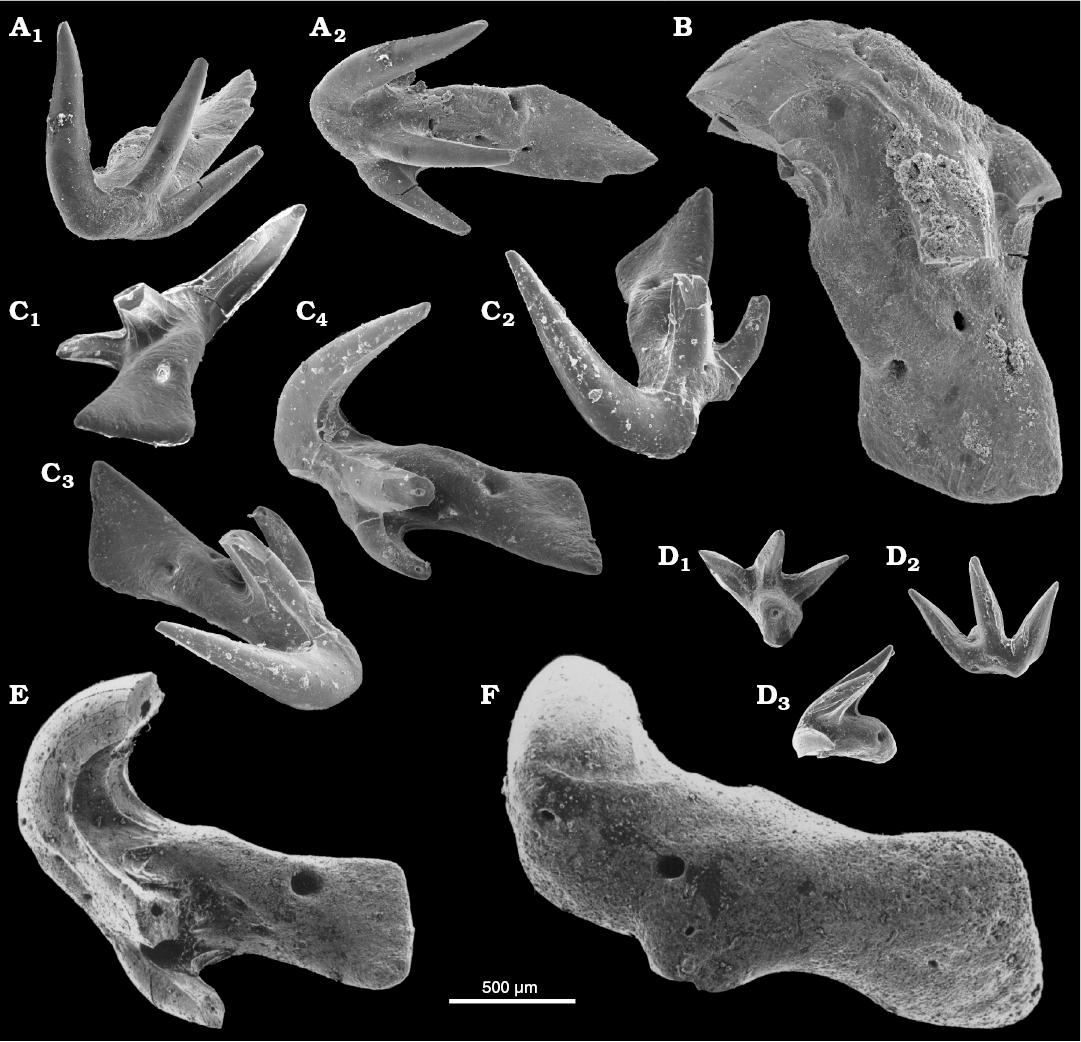
Fig. 2. Teeth of chondrichthyan fish Thrinacodus ferox (Turner, 1982) from the upper Viséan of Czerna, Poland (A, B) and the Tournaisian of Czatkowice, Poland (C–F). A. ZPAL P. IV/301, in labial (A1) and oral (A2) views. B. ZPAL P. IV/302 in oral view. C. MWGUW/Ps/13/1 in lingual (C1), labial (C2), oral (C3), and lateral (C4) views. D. MWGUW/Ps/1/212, subsymmetrical, parasymphysial (?) tooth in lingual (D1), labial (D2), and lateral (D3) views. E. MWGUW/Ps/1/214 in lateral view. F. MWGUW/Ps/1/213 in basal view.
Family Jalodontidae Ginter, Hairapetian, and Klug, 2002
Genus Jalodus Ginter, 1999
Type species: Phoebodus australiensis Long, 1990; Famennian, Mae Sam Lap, Thailand.
Jalodus sp.
Fig. 3A.
Material.—One tooth (MWGUW/Ps/13/2) from the upper Tournaisian, Czatkowice, sample A.
Description.—The tooth of Jalodus from Czatkowice has two cusps fairly well preserved, but the third cusp is broken at the base and reveals a relatively large pulp cavity. The base is slightly abraded and therefore the size and shape of the orolingual button is unknown.
Remarks.—Generally, the tooth is similar to those called by Ginter (1990: pl. 1: 7–10) Phoebodus (now Jalodus) australiensis Morphotype 2, found in a neptunian dyke at Dalnia, Holy Cross Mountains, Poland, associated to a mixed, latest Famennian to earliest Tournaisian conodont fauna. Comparable teeth are also known from the Famennian–Tournaisian boundary beds of the Dapoushang Section, China (Wang 1989: particularly pl. 29: 3, as Cladodus sp.). Because the typical specimens of Jalodus australiensis (Morphotype 1 sensu Ginter 1990) are usually found lower in the Famennian (Palmatolepis expansa–Early Palmatolepis praesulcata conodont zones), the tooth from Czatkowice may represent a younger, only slightly different, species.
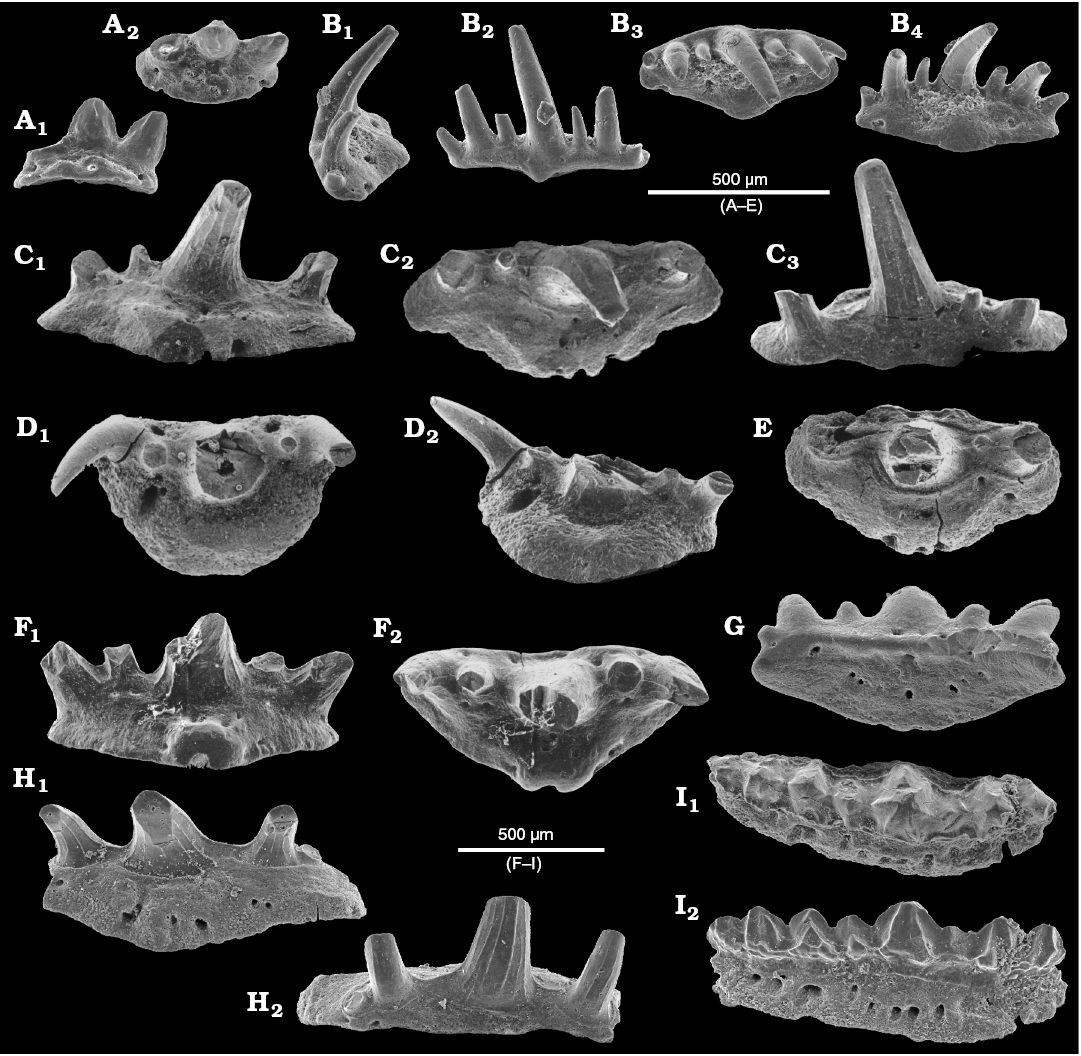
Fig. 3. Chondrichthyan fish teeth from the Tournaisian of Czatkowice, Poland. A. Jalodus sp., MWGUW/Ps/13/2, in lingual (A1) and oral (A2) views. B–H. Falcatidae indet. 1. B. MWGUW/Ps/13/3, small seven-cuspid specimen in lateral (B1), labial (B2), oral (B3), and lingual (B4) views. C–F. Large five-cuspid specimens. C. MWGUW/Ps/13/4 in lingual (C1), oral (C2), and labial (C3) views. D. MWGUW/Ps/13/5 in oral (D1) and lingual (D2) views. E. MWGUW/Ps/13/6 in oral view. F. MWGUW/Ps/1/221 in lingual (F1) and oral (F2)views. G. MWGUW/Ps/13/7, large seven-cuspid specimen in basal/labial view. H. MWGUW/Ps/13/8, large five-cuspid specimen without the intermediate cusplets in lingual (H1) and labial (H2) views. I. Protacrodontidae indet., MWGUW/Ps/13/9, in oral (I1) and lingual (I2) views.
Superorder Cladodontomorphi Ginter, Hampe, and Duffin, 2010
Order Symmoriiformes Zangerl, 1981
Family Falcatidae Zangerl, 1990
Genus Denaea Pruvost, 1922
Type species: Denaea fournieri Pruvost, 1922; early Viséan, Denée, Belgium.
Denaea wangi Wang, Jin, and Wang, 2004
Fig. 4A–C.
2004 Denaea wangi sp. nov.; Wang et al. 2004: 98–99, fig. 10A, 11.
2010 “Denaea wangi” Wang, Jin, and Wang, 2004; Ginter et al. 2010: 66–67, fig. 61.
2015 Denaea wangi Wang, Jin, and Wang, 2004; Ginter et al. 2015: 911–913, fig. 13A–D.
Material.—Three teeth (ZPAL P. IV/303–305) from the upper Viséan, Czerna, sample Cz-1.
Description.—The teeth of Denaea wangi are minute, 0.4–0.7 mm mesio-distally. The crown is cladodont, asymmetrical, with seven to eight delicate cusps inclined distally. The highest cusp is situated not exactly in the median position, but slightly displaced in the mesial direction. There is different number of lateral cusps on the mesial (2–3) and distal (3–4) sides. The difference in size between the cusps is moderate. The base is thin and flat, triangular in the oral view, with a tiny basolabial tubercle (Fig. 4A1, C1).
Remarks.—The extensive discussion on the validity of this species and its similarity to “small Damocles” from the Serpukhovian of Bear Gulch, Montana, was given by Ginter et al. (2015). Here it can be added that although the teeth of Denaea wangi usually occur together with the teeth of some larger falcatids, no specific species was observed to repeat in such associations. At Czerna, it is Falcatidae indet. 2; at Todowa Grząba (Holy Cross Mountains, Poland) it is Denaea cf. fournieri; in the type material from China it is “Stethacanthus” gansuensis; in Montana, USA: Damocles and Falcatus, etc. This supports the idea that D. wangi is indeed a small, separate species, and not only a juvenile form (as incorrectly suggested by Ginter et al. 2010).
Stratigraphic and geographic range.—Upper Viséan of northern Europe (Poland, Belgium) and China (Gansu). Perhaps also Serpukhovian of Montana, USA (articulated specimens, Ginter et al. 2010: fig. 59C).

Fig. 4. Teeth of falcatid chondrichthyan fishes from the upper Viséan of Czerna, Poland. A–C. Denaea wangi Wang, Jin, and Wang, 2004. A. ZPAL P. IV/303 in lingual (A1) and basal (A2) views. B. ZPAL P. IV/304 in labial (B1) and oral (B2) views. C. ZPAL P. IV/305 in labial (C1) and oral (C2) views. D, E. Falcatidae indet. 2. D. ZPAL P. IV/226 in lateral (D1), labial (D2), oral (D3), and lingual (D4) views. E. ZPAL P. IV/217 in labial (E1), lingual (E2), and oral (E3) views.
Falcatidae indet. 1
Fig. 3B–H.
Material.—Eight teeth (MWGUW/Ps/1/221, MWGUW/Ps/13/3–8, 19) from the Tournaisian, Czatkowice, sample A.
Description.—All the Denaea-like falcatid teeth from the Tournaisian of Czatkowice were designated here as Falcatidae indet. 1, but we are unsure whether all of them indeed represent the same species, because the diversity is quite high. There occur pentacuspid teeth, with a prominent median cusp which is vertical (Fig. 3D–F) or slightly inclined distally (Fig. 3C), but also seven-cuspid teeth with minute outer lateral cusplets (Fig. 3B, G). There is also one rather unusual, asymmetrical pentacuspid specimen, with outer lateral cusplets present, but devoid of intermediate ones (Fig. 3H). The cusps are covered (where observed) with subparallel cristae, stronger on the labial face. The bases are triangular, narrowing lingually, with an orolingual button (Fig. 3F) or at least a hump. The basolabial thickening can be in a form of a small tubercle (Fig. 3B2) to a wide shelf (Fig. 3G). The mesio-distal dimension of the bases ranges from 0.5 to 1 mm.
Remarks.—The poor preservation of most of the specimens precludes the definite identification and comparison of these teeth. None of them closely resembles typical specimens of Denaea cf. fournieri sensu Ginter et al. (2015) or other falcatids. Pentacuspid specimens of Falcatidae indet. 2 from Czerna (see below; Fig. 4D) are somewhat similar to the tooth in Fig. 3F, with its prominent, lingually placed orolingual button. However, the seven-cuspid teeth of Falcatidae indet. 2 do not have outer lateral accessory cusplets, but two pairs of intermediate cusplets instead. Falcatidae indet. 1 is probably an older, Tournaisian falcatid taxon.
Falcatidae indet. 2
Fig. 4D, E.
Material.—Twelve teeth (ZPAL P. IV/217, 226, 324–333) from the upper Viséan, Czerna, sample Cz-1.
Description.—The teeth of Falcatidae indet. 2 have wide and short bases with the orolingual button at the lingual rim. The button is perforated from the lingual side and yet another foramen opens between the button and the median cusp. The crown is cladodont, with a prominent median cusp, rounded in cross section and covered with numerous subparallel cristae. The lateral cusps are relatively low. There occur pentacuspid specimens (Fig. 4D) and, fewer and larger, seven-cuspid specimens (Fig. 4E). In the latter, the outer lateral cusps are the highest and there are two pairs of intermediate cusplets. The teeth are relatively large, 1–2 mm mesio-distally.
Remarks.—The teeth of Falcatide indet. 2 are very similar to those of Denaea cf. fournieri sensu Ginter et al. (2015), particularly as far as the form of the crown is concerned. However, it should be reminded that in the latter publication there were noted differences between the forms of the bases in two groups of specimens. Those from the Holy Cross Mountains (Poland) and Rhenish Mountains (Germany) are characterised by broad bases and vague buttons, but those from Cawdor Quarry in Derbyshire (UK), and especially from the sample Cawdor 11 (Ginter et al. 2015: fig. 7D–G) have short bases (labio-lingually) with a strong button sticking lingually, exactly like in pentacuspid specimens of Falcatidae indet. 2. Perhaps then, the material from the sample Cawdor 11 (UK) and Falcatidae indet. 2 from Czerna represent a new taxon, different from Polish and German Denaea cf. fournieri, as tentatively suggested by Ginter et al. (2015: 910).
Order Ctenacanthiformes Glikman, 1964
Family Ctenacanthidae Dean, 1909
Ctenacanthidae indet.
Fig. 5C.
Material.—One tooth (ZPAL P. IV/307) from the upper Viséan, Czerna, sample Cz-1.
Description.—The tooth is relatively small compared to most of the known ctenacanthiform species (about 0.5 mm mesio-distally). Its crown is cladodont, tricuspid, with a prominent median cusp (unfortunately the upper part is broken) and the lateral cusps considerably diverging laterally. The cusps are compressed labio-lingually, with a carina connecting their bases, and covered with strong cristae on both sides. The base bears an orolingual button and a basolabial prominence narrower than the median cusp.
Remarks.—The tooth from Czerna resembles to some extent those of Ctenacanthus terrelli (Newberry, 1889) from upper Famennian Cleveland Shale of Ohio (USA), and particularly the small specimen illustrated by Ginter (2010: fig. 3E, F), with its conspicuous cristation.

Fig. 5. Chondrichthyan fish teeth from the upper Viséan of Czerna, Poland. A, B. Cladodontomorphi indet. 1. A. ZPAL P. IV/255 in labial (A1) and basal (A2) views. B. ZPAL P. IV/306, small specimen with a broken piece lying nearby in lingual (B1) and oral (B2) views. C. Ctenacanthidae indet., ZPAL P. IV/307 in oral (C1) and labial (C2) views. D. Cladodontomorphi indet. 2, ZPAL P. IV/308 in labial (D1), lingual (D2), and oral (D3) views. E. Cassisodus sp., ZPAL P. IV/309 in oral (D1) and lingual (D2) views. F. Cassisodus sp., ZPAL P. IV/232 in lingual (E1) and oral (E2) views.
Cladodontomorphi indet. 1
Fig. 5A, B.
Material.—Two teeth (ZPAL P. IV/255, 306) from the upper Viséan, Czerna, sample Cz-1.
Description.—There are two teeth of this type, similar to each other, in the sample from Czerna. One is larger, more than 1 mm wide, and the other smaller, about 0.5 mm mesio-distally. They are very delicate and brittle, virtually symmetrical. Their crown is multicuspid, comb-like, of the cladodont design. The median cusp is very slender, covered with only a few cristae, rather strong in comparison to the width of the cusp (the smooth surface of the labial side in the smaller tooth (Fig. 5B2), is probably due to the post-mortem destruction of enameloid). There are seven lateral cusplets on each side of the median cusp in the larger tooth, of which the fourth are the highest. In the smaller tooth, there are only five cusplets on each side. The smaller cusplets often grow on a side of the larger one.
The base is triangular in the oral view and its orolingual side is somewhat wavy, with a shallow sulcus in the middle. The basolabial rim is slightly elevated, forming a kind of a parapet, at least in the larger specimen. There are numerous irregularly placed foramina in the aboral/labial region, but their presence may be an effect of abrasion.
Remarks.—Because of their size, numerous slender cusplets and the base devoid of any articulation devices, these teeth could be equivocated with Denaea wangi. However, their almost complete symmetry, the ornamentation of the larger cusps and the specific type of growth of the intermediate cusplets (on a side of the larger ones) are the features which combined preclude their belonging not only to Denaea, but also to Falcatidae and even Symmoriiformes. The closest teeth known to us are those of Tamiobatis vetustus sensu Williams (1998) from the upper Famennian of Ohio, USA, i.e., a ctenacanthiform. However, the teeth of Tamiobatis have an additional row of labial accessory cusplets, absent from the teeth from Czerna, and the median cusp in Tamiobatis is usually shorter and wider. Therefore, we temporarily leave them as Cladodontomorphi indet. 1, without determining an order.
Cladodontomorphi indet. 2
Fig. 5D.
Material.—One tooth (ZPAL P. IV/308) from the upper Viséan, Czerna, sample Cz-1.
Remarks.—At first sight the specimen looks like a large falcatid tooth, with the long and relatively slender median cusp covered with dense cristae, and the intermediate cusplets histologically separate from the main cusps. However, the trapezoidal, lingually widening base and the labio-lingual compression of the median cusp make it closer to the ctenacanthiforms. Therefore, the order cannot be determined with certainty.
Cohort Euselachii Hay, 1902
Superfamily Protacrodontoidea Zangerl, 1981
Family Protacrodontidae Cappetta, Duffin, and Zidek, 1993
Protacrodontidae indet.
Fig. 3I.
Material.—Two teeth (MWGUW/Ps/13/10, 20) from the Tournaisian, Czatkowice, sample A.
Description and remarks.—The teeth are characterised by a slightly asymmetrical, low protacrodont crown and an euselachian base. Teeth of this type are usually attributed to Protacrodontidae, but in the Carboniferous they also may represent other euselachian groups. It is notable that these two specimens are the only crushing teeth found in the material form Czatkowice.
Order Hybodontiformes Maisey, 1975
Family indet.
Genus Cassisodus Ginter and Sun, 2007
Type species: Cassisodus margaritae Ginter and Sun, 2007, Carboniferous, Mississippian, middle Tournaisian, Siphonodella crenulata Conodont Zone; Muhua, Guizhou Province, China.
Cassisodus cf. margaritae Ginter and Sun, 2007
Fig. 5E, F.
Material.—Three teeth (ZPAL P. IV/232, 309, 334) from the upper Viséan, Czerna, sample Cz-1.
Description.—The teeth are composed of a low, crushing crown and a typical euselachian base sensu Ginter et al. (2010). The crown is asymmetrical and consists of a main, pyramidal cusp, not in the median position. On the shorter side of the crown there are two lateral cusps and on the longer one there are three cusps in smaller specimens (Fig. 5E), but in the larger specimen the respective numbers are three and four (Fig. 5F). The bases of the cusps are fused. There are tubercles on both labial and lingual sides of the cusps. In the larger tooth (4 mm mesio-distally) the main part of the crown is strongly elevated, but in the smaller teeth (about 2 mm wide) the size difference between the main and the lateral cusps is almost negligible.
Remarks.—The type material of Cassisodus margaritae Ginter and Sun, 2007, was described from the middle Tournaisian of Muhua, southern China. Although the tooth designated as the holotype is compact and symmetrical (Ginter and Sun 2007: fig. 5A), the other teeth (Ginter and Sun 2007: fig. 5B–D) are very similar to those from Czerna. They are elongated mesio-distally, with smooth, fused cusps and characteristic tubercles on the lingual and labial faces. It was proposed (Ginter and Sun 2007: 714) that the holotype was an anterior tooth whereas the rest represent the lateral tooth families. The latter most probably also concerns the teeth from Czerna.
Chondrichthyan scales and branchial denticles
In the Viséan of Czerna, the predominating type of dermal cover are ctenacanthid scales sensu Reif (1978), with concave bases and numerous separate odontodes directed backwards (Fig. 6A–C). Such scales are typical of ctenacanthiforms, but also a few other Palaeozoic shark orders, such as the Phoebodontiformes (see discussion in Ginter and Skompski in press). A few minute scales (Fig. 6D) are slightly different: they have flat, palm-like crowns and spongeous bases. The form of the base resembles that of hybodontid scales sensu Reif (1978: see eg., fig. 2B–D).
The ctenacanthid and hybodontid body scales are absent from the sample from the Tournaisian of Czatkowice. However, there occur symmoriiform branchial denticles of the Stemmatias-type (Fig. 6M–O), usually with two rows of cusps. There also occur two problematic, tricuspid denticles (Fig. 6P, Q) whose function and position on the body is yet unknown to us.
Actinopterygian and acanthodian remains
Palaeoniscoid rhomboidal body scales (Fig. 6E–G, K), isolated conical teeth (Fig. 6H, J), vertebrae (Fig. 6I), and pieces of jaw (Fig. 6L) are common in both studied samples. Smooth-crowned acanthodian scales (Fig. 6R) were found only in the sample from Czatkowice.

Fig. 6. Ichthyoliths from the upper Viséan of Czerna, Poland (A–K) and the Tournaisian of Czatkowice, Poland (L–R). A–C. Ctenacanthoid scales. A. ZPAL P. IV/311 in basal view. B. ZPAL P. IV/312 in lateral view. C. ZPAL P. IV/313 in coronal view. D. Hybodontoid scale, ZPAL P. IV/314 in lateral (D1) and coronal (D2) views. E–G, K. Actinopterygian scales. E. ZPAL P. IV/316 in external view. F. ZPAL P. IV/315 in basal view. G. ZPAL P. IV/317 in external view. K. ZPAL P. IV/244 in external view. H, J. Actinopterygian teeth. H. ZPAL P. IV/318 in lingual view. J. ZPAL P. IV/319 in lateral view. I. Actinopterygian vertebra, ZPAL P. IV/320. L. Fragment of actinopterygian jaw, MWGUW/Ps/13/11 in oral view. M–O. Chondrichthyan (symmoriiform?) branchial denticles. M. MWGUW/Ps/13/12 in coronal view. N. MWGUW/Ps/13/13 in lateral view. O. MWGUW/Ps/13/14 in coronal (O1) and lateral (O2) views. P, Q. Problematic chondrichthyan? denticles. P. MWGUW/Ps/13/15 in labial view. Q. MWGUW/Ps/13/16 in lingual view. R. Acanthodian scale, MWGUW/Ps/13/17 in coronal/lateral view.
Discussion
Diversity of Mississippian Falcatidae.—The Falcatidae are small symmoriiform sharks with naked skin except for the scales along the lateral line and hook-like denticles on the skull roof, and, probably only in males, on a peculiar, anteriorly directed spine over the head. We know quite a lot about the anatomy of the early falcatids mainly thanks to two sources: lower Viséan Black Marbles of Denée in Belgium (Fournier and Pruvost 1928) and Serpukhovian Bear Gulch beds in Montana, USA (Lund 1985, 1986). The falcatid head spine can be observed on the male individual (recognisable as such due to the presence of large claspers) of Falcatus falcatus Lund, 1985, on the specimens of Damocles serratus Lund, 1986, and on a yet undescribed small shark with Denaea wangi-like teeth from Bear Gulch (CM35412, incorrectly designated as small individual of D. serratus by Ginter et al. 2010: fig. 59C). None of the specimens of Denaea fournieri Pruvost, 1922 from Belgium bears such a spine, but it is uncertain whether all of them are females or the spine is absent in both sexes.
Generally, the teeth of falcatids are of the cladodont design, with slender, separate cusps, rounded in cross section and covered with gentle, dense, subparallel cristae running from the base almost to the tip. All of the observed teeth in the articulated specimens of Falcatus and Damocles from Bear Gulch and those associated with the head of Ozarcus mapesae Pradel, Maisey, Tafforeau, Mapes, and Mallatt, 2014, from the Fayetteville Formation (upper Viséan, Arkansas, USA) are pentacuspid. The teeth of Denaea fournieri have either five or seven cusps. Unfortunately, the extraction of complete teeth associated to the articulated specimens has not been successful yet, so the comparison to the disarticulated material is difficult. And among the isolated teeth found in the pelagic facies of the Mississippian, from Illinois (USA) to northern Europe, to China, there occur various tooth forms based on the common, falcatid pattern. The following forms, differing usually by minor, secondary features, can be distinguished (for the illustrations not provided in this paper, refer to Ginter et al. 2015):
(i) Denaea cf. fournieri from Todowa Grząba (Holy Cross Mountains, Poland; Ginter et al. 2015: fig. 12; Fig. 7B), probably identical with Denaea fournieri from Denée, Belgium. The tooth crown usually consists of five or seven cusps. The median cusp is long and slender, sigmoidal in lateral view, often slightly inclined laterally, and sub-circular in cross section. The whole cusp, except for the uppermost fifth, is ornamented with dense sub-parallel cristae. The base is roughly pentagonal. The orolingual button is oval, sometimes almost split in two by the openings. In the smaller specimens the button seems to be more compact and distinct and in the larger ones its boundaries are rather vague. The basolabial projection is tubercle-like, squarish in cross-section, and not wider than the basal part of the median cusp.
(ii) Denaea cf. fournieri from Westenfeld Quarry (Rhenish Mountains, Germany; Ginter et al. 2015: fig. 10A–E). This form is probably conspecific with the former one, but differs by the larger number of cusps (up to nine) in some teeth. In such nine-cuspid specimens there is a pair of additional, small, outermost lateral cusplets.
(iii) Denaea cf. fournieri from Cawdor Quarry (Derbyshire, UK; Ginter et al. 2015: fig. 7A–F). The teeth from Derbyshire have lingually narrowing bases with a pronounced orolingual button placed at the lingual end. There are only five cusps in the crown. The angle between the median and lateral cusps is relatively high. A few central cristae on the labial face of the median cusp converge half way to the tip. These teeth are somewhat similar to Denaea williamsi Ginter and Hansen, 2010, but the ornamentation of the cusps is closer to “Stethacanthus” gansuensis Wang, Jin, and Wang, 2004.
(iv) “Stethacanthus” gansuensis from Qilian Mountain (Gansu, China; Wang et al. 2004: fig. 7, 8; Fig. 7A). The illustrated specimens of this species definitely display falcatid, and not stethacanthid characters. However, they differ from all the here listed falcatids by their wide (mesio-distally elongated) orolingual button (Fig. 7A2). The relatively strong divergence of the lateral cusps and the convergence of central labial cristae on the median cusp (Fig. 7A1) in “S.” gansuensis are the crown features similar to Denaea cf. fournieri (iii) from Derbyshire.
(v) Denaea williamsi from Cedar Grove Church (Illinois, USA; Ginter et al. 2010: fig. 62; Ginter and Hansen 2010: figs. 3, 4), Grand Canyon (Arizona, USA; Hodnett and Elliott 2018: fig. 6), Todowa Grząba (Holy Cross Mountains, Poland; Ginter et al. 2015: fig. 13E, F), and Borovichi (Novgorod Region, Russia; Savitskiy et al. 2000, pl. 16: 2–4). The teeth of D. williamsi have the specific ornament of the median cusp, composed of cristae which, on the labial side, diverge laterally at the base and then corresponding pairs join before reaching the tip, whereas on the lingual face they converge at various heights. The outermost cusps are strongly divergent, more so than in D. cf. fournieri. The base is provided with a distinct button, in the shape of horizontal figure of 8, at the lingual rim.
(vi) Denaea wangi from Qilian Mountain (Gansu, China; Wang et al. 2004: fig. 10, 11; Fig. 7C), Royseux (Belgium; Derycke et al. 2005), Todowa Grząba (Ginter et al. 2015: fig. 13A–D) and Czerna (this paper, Fig. 4A–C, probably identical with the specimen CM 35412 from the Bear Gulch, Montana, USA; Ginter et al. 2010: fig. 59C). The teeth are minute (0.4–0.7 mm mesio-distally), multicuspid and asymmetrical. The cusps (up to 11 in a row) are very delicate and smooth. The number of cusps present on either side of the median cusp differs. The base is very thin, lacking articulation devices. A shallow concavity usually occurs in place of the orolingual button.
(vii) Falcatidae indet. 1 from Czatkowice (Krzeszowice area, Poland; this paper, Fig. 3B–H). These teeth and their diversity are described in the systematic part. Due to the poor preservation the comparison with the other falcatids is rather difficult.
(viii) Falcatidae indet. 2 from Czerna (Krzeszowice area, Poland; this paper, Fig. 4D, E). The teeth of this type, described in the systematic part, have the crowns similar to Denaea cf. fournieri (i and ii), but the bases closer to those of the teeth from Derbyshire (iii) and D. williamsi.
The data presented in the Appendix 1 show that from the late Viséan through to the Serpukhovian, at least seven different falcatid species, very similar to each other, inhabited the seas of the Northern Hemisphere. Most of them lived there simultaneously. The precise number of falcatid species cannot be given at the moment, because it is highly probable that some of the tooth-based taxa in fact are conspecific with those known from articulated skeletons. One may hope that in the nearest future a few of the numerous specimens of Falcatus and Damocles from the Bear Gulch will be sacrificed and reasonably preserved teeth will be chemically extracted, suitable for the SEM observation.
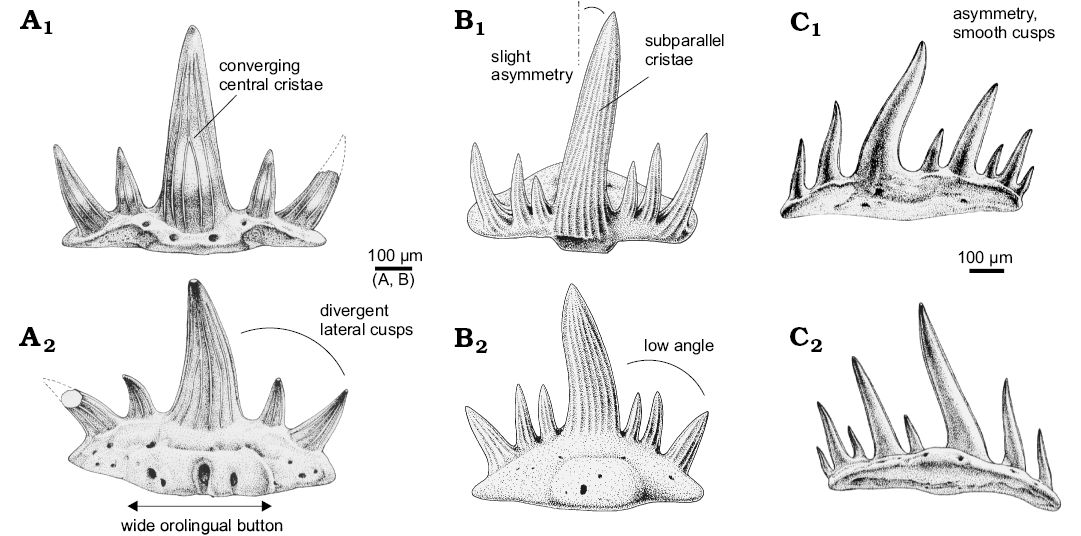
Fig. 7. Comparative drawings of selected falcatid species from the upper Mississippian with a few diagnostic morphological terms. A. The holotype of “Stethacanthus” gansuensis Wang, Jin, and Wang, 2004 (IVPP V 13666.1) from the Serpukhovian of Qilian Mountain, Gansu, China, in labial (A1) and lingual (A2) views. B. Denaea cf. fournieri (ZPAL P. IV/221) from the Viséan of Todowa Grząba, Holy Cross Mountains, Poland, in labial (B1) and lingual (B2) views. C. The holotype of Denaea wangi Wang, Jin, and Wang, 2004 (IVPP V 13668.1) from the Serpukhovian of Qilian Mountain, Gansu, China, in labial (C1) and lingual (C2) views. A, C modified from Wang et al. (2004).
The late Mississippian was definitely the best period for the Falcatidae (despite the suggestions that this is only a sampling bias; see Hodnett and Elliott 2018), but that does not mean that this group did not exist in the pre- and post-Mississippian times. There are several isolated records of teeth with falcatid morphology from the Upper Devonian (Famennian), of which probably the oldest are the teeth from the Late Palmatolepis crepida Conodont Zone of Coumiac, Montaigne Noire, France (Riemann et al. 2002). These teeth are generally symmetrical, some of them are pentacuspid (cladodonter Zahn Morphotyp N11; Riemann et al. 2002: pl. 6: 12), but the other are multicuspid with up to 11 cusps in the crown (cladodonter Zahn Morphotyp N6; Riemann et al. 2002: pl. 6: 8, 11). No species was erected based on Devonian falcatid-like teeth, they were usually designated as cladodonts, stethacanthids, or Denaea sp.
The known distribution of post-Mississippian falcatids is restricted to the areas of USA, Russia, and Central Asia (see Ginter 2018: 397–399, table 1). The flourishing Serpukhovian falcatids were replaced with Denaea saltsmani Ginter and Hansen, 2010, and at least two species of Stethacanthulus Zangerl, 1990. Judging from the available information, the falcatids did not persist into the Late Permian. The latest occurrences of Stethacanthulus meccaensis (Williams, 1985) were reported from the Capitanian (uppermost Middle Permian) of the Apache Mountains (Ivanov et al. 2013) and Guadalupe Mountains (Ivanov et al. 2015), West Texas, USA.
There are repeated records of shark teeth, externally similar to those of the Falcatidae, from the Cretaceous of Europe. Guinot et al. (2013: fig. 2a–e) presented such teeth from the deep marine deposits of the Valanginian (Lower Cretaceous) of southern France and Feichtinger et al. (2018: figs. 3, 4) from the condensed pelagic limestone, also Valanginian, of the Austrian Alps. In spite of the considerable microstructural differences, such as the triple-layered enameloid (instead of a single layer in typical falcatids) and the special vascularisation of the bases, the authors of both publications decided to include those teeth formally in the Falcatidae (see particularly the extensive discussion in Feichtinger et al. 2018: 11–12). Although the idea that the falcatids have survived in some unknown refugia between the Permian and the Cretaceous is tempting, we consider such a determined identification as premature. The differences appear to us too important and the argumentation too weak, so at this stage of knowledge we prefer to consider that the Falcatidae really went extinct before the end of the Palaeozoic.
Comparison to the other assemblages of Mississippian chondrichthyans.—Ginter et al. (2015) noted that whereas the known Mississippian assemblages of pelagic sharks differ at the species level, there are considerable similarities as far as supra-specific categories are concerned. For the sake of comparison, they proposed the following categories: (i) Falcatidae; (ii) other Cladodontomorphi; (iii) Thrinacodus; (iv) Bransonella + Jalodus; (v) Anachronistidae; (vi) Euselachii + Euchondrocephali. The assemblage from the Tournaisian of Czatkowice is too small (18 specimens) for any reasonable statistical analysis, but the assemblage from the Viséan of Czerna, with its 27 chondrichthyan teeth, is rich enough to be compared to the previously described collections.
It is visible at first sight (Table 2, Fig. 8A, B) that the proportions among the listed categories in sample Cz-1 are quite similar to those from sample TG-D from Todowa Grząba (Holy Cross Mountains, Poland), a good representative of Mississippian pelagic assemblages in Europe (Ginter et al. 2015). The predominance of falcatids and the low number of euselachians/euchondrocephalans with crushing teeth is a typical feature of chondrichthyan communities living in deeper basins, at some distance from the shallow-water carbonate platforms. The relative abundances characteristic of the latter platforms (e.g., sample MH-1, Tournaisian, Muhua, South China; Ginter and Sun 2007 or the assemblage from the Kilbride Limestone Formation, Tournaisian, Ireland; Duncan 2003, 2004, 2006; Table 2, Fig. 8D, E) are totally different: the crushing teeth predominate and the falcatids form only a tiny addition.
Table 2. Comparison of Mississippian chondrichthyan assemblages from the area of Krzeszowice (this paper), Todowa Grząba (Holy Cross Mountains, Poland; Ginter et al. 2015), Surprise Canyon Formation (middle member, Northern Arizona, USA; Hodnett and Elliott 2018), Muhua (southern China; Ginter and Sun 2007), and Kilbride Limestone Formation (Nobber, Ireland; Duncan 2003, 2004, 2006); n, number of specimens.
|
Taxon / Samples |
Krzeszowice area |
Comparative material |
||||||||
|
Czerna sample Cz-1 |
Todowa Grząba sample TG-D |
Surprise
Canyon |
Muhua sample MH-1 |
Kilbride
Limestone |
||||||
|
n |
% |
n |
% |
n |
% |
n |
% |
n |
% |
|
|
Falcatidae |
15 |
55 |
49 |
70 |
20 |
28 |
10 |
10 |
7 |
10 |
|
Other Cladodontomorphi |
4 |
15 |
3 |
4 |
6 |
8 |
25 |
25 |
0 |
0 |
|
Thrinacodus |
5 |
19 |
9 |
13 |
8 |
11 |
9 |
9 |
12 |
18 |
|
Bransonella |
0 |
0 |
4 |
6 |
2 |
3 |
0 |
0 |
0 |
0 |
|
Anachronistidae |
0 |
0 |
0 |
0 |
12 |
17 |
0 |
0 |
0 |
0 |
|
Euselachii + Euchondrocephali |
3 |
11 |
5 |
7 |
24 |
33 |
56 |
56 |
49 |
72 |
|
Total |
27 |
100 |
70 |
100 |
72 |
100 |
100 |
100 |
68 |
100 |
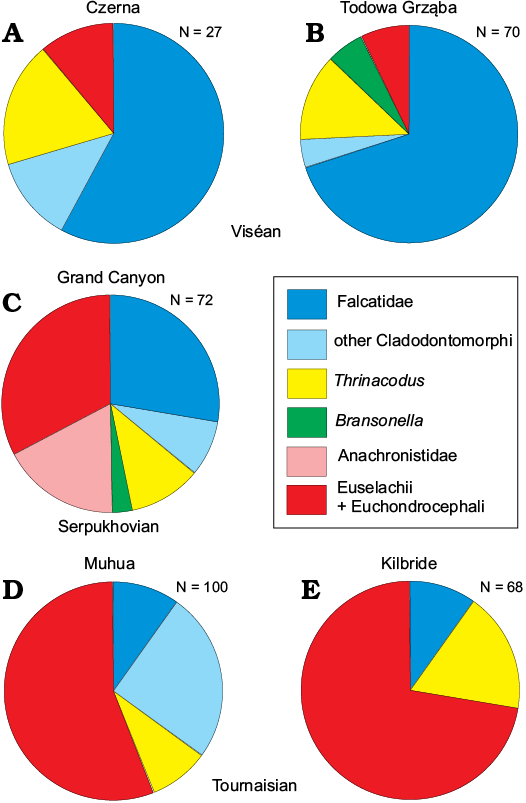
Fig. 8. Pie-diagrams illustrating relative abundances of teeth representing groups of chondrichthyan taxa in five Mississippian assemblages. A. Sample Cz-1 from the upper Viséan of Czerna near Kraków, Poland (this paper). B. Sample TG-D from the upper Viséan of Todowa Grząba, Holy Cross Mountains, Poland (Ginter et al. 2015). C. The assemblage from the Surprise Canyon Formation, middle member, Serpukhovian, Grand Canyon, Northern Arizona, USA (Hodnett and Elliott 2018). D. Sample MH-1 from the Tournaisian of Muhua, South China (Ginter and Sun 2007). E. The assemblage from the Kilbride Limestone Formation, upper Tournaisian, Nobber, Ireland (Duncan 2003, 2004, 2006). Note the predominance of falcatid teeth at Czerna and Todowa Grząba and great abundance of crushing teeth (Euselachii + Euchondrocephali) at Muhua and Kilbride. N, total number of specimens.
Whereas the Chinese assemblage from Muhua is rather well known (e.g., Ginter et al. 2010: fig. 7), the information on the chondrichthyan assemblage from the Kilbride Limestone Formation is less accessible and deserves special attention here. The material was obtained from the samples collected in the upper Tournaisian part (equivalent to the upper part of the Scaliognathus anchoralis Conodont Zone) of a disused quarry near Nobber, Co. Meath, Ireland and described by Duncan in her unpublished Ph.D. dissertation in 1999. However, formally the specimens were published in three parts, dedicated to different groups of taxa: Thrinacodus (Duncan 2003), Lissodus (Duncan 2004) and the rest (falcatids and various crushing teeth; Duncan 2006). The combined assemblage, altogether 68 teeth, consists of seven falcatids (called Denaea sp.), 12 diverse, perfectly preserved teeth of Thrinacodus ferox (Turner, 1982), 37 teeth of Lissodus spp., two protacrodonts and 10 dental elements called “Chondrichthyan Pavement Dentition” (CPD) and representing various unidentified euselachians and euchondrocephalans. According to Duncan (2006: 52), the Kilbride Limestone Formation “was deposited in moderately shallow water on a high-energy carbonate shelf”. This perfectly corresponds to the absolute predominance (72%) of crushing and grinding euselachian and euchondrocephalan teeth (Lissodus + protacrodonts + CPD) in the collection.
Quite recently, Hodnett and Elliott (2018) have published an interesting collection of chondrichthyan isolated remains, mainly teeth, from the Mississippian–Pennsylvanian boundary beds of the western Grand Canyon, Northern Arizona, USA. The fossils come from several members and several localities, so the quantitative analysis and comparison of the whole set would have no real meaning. However, we managed to separate data concerning only one, relatively uniform litostratigraphic unit: the middle member of the Surprise Canyon Formation, dated as Serpukhovian. In the type section of the Surprise Canyon Formation, Bat Tower Overlook, its middle member consists of a thinly bedded, dark reddish-brown to yellowish-gray marine limestone with brachiopods, small gastropods, crinoids, and abundant shell fragments (section 5) and a thin dark purplish-gray limestone with separations of thin silty limestone beds, containing brachiopods (section 6; see Hodnett and Elliott 2018: 3). From this member, 72 chondrichthyan teeth assigned to 22 taxa were recovered.
When grouped in the categories proposed by Ginter et al. (2015), the relative abundances are as follows: Falcatidae (with numerous Denaea williamsi) 28%, other cladodontomorphs (mainly ctenacanths) 8%, Thrinacodus 11%, Bransonella 3%, Anachronistidae 17%, Euselachii + Euchondrocephali 33%. The balanced input of cladodonts (with falcatids as a major component) vs. crushing teeth of euselachians and euchondrocephalans places this assemblage at the intermediate position between Muhua (shallow carbonate platform) and Todowa Grząba (moderately deep basin; Table 2, Fig. 8C).
Following Billingsley and Beus (1999), Hodnett and Elliott (2018) suggest that in the late Chesterian–Serpukhovian a period of subsidence (or a glacieustatic rise; Skompski 2003) allowed marine waters to gradually flood the valleys eroded earlier in the substrate, forming local estuaries. The marine middle and upper units of the Surprise Canyon Formation were deposited in such open, sometimes relatively deep, estuaries. Such an environment could have been favourable both for the taxa preferring shallow water conditions and the open marine ones. This nicely corresponds to the relative abundances in the described chondrichthyan assemblage. Similar environment was proposed for the deposition of the Indian Cave Sandstone in the south-eastern Nebraska (Pennsylvanian, Gzhelian; Fischbein et al. 2009), from which Ossian (1974) recovered a very rich and diverse assemblage of vertebrate remains, representing groups of various environmental preferences, from falcatids to tetrapods.
Conclusions
The assemblages from the Tournaisian and late Viséan of the Krzeszowice area do not add much new information to the distribution picture of Mississippian chondrichthyans. However, they confirm that the Falcatidae are a very diverse group, occurring in large numbers in the pelagic environments. Also, once again it becomes evident that euselachians and euchondrocephalans with crushing teeth are subordinate in the far offshore basins, but predominate on the shallow submarine platforms. An interesting thing is the patchy record of the Anachronistidae in the upper Viséan–Serpukhovian. Ginter et al. (2015: 920) suggested that they “are better represented in the second assemblage type”, i.e., relatively deep, but closer to submarine platforms, similar to the assemblage from Steeplehouse Quarry (Derbyshire). The abundance of anachronistid teeth in the putative estuarine environment of the Surprise Canyon Formation (Hodnett and Elliott 2018) and their total absence from the Central European basins (Rhenish Mountains and Holy Cross Mountains; Ginter et al. 2015; Czerna, Krzeszowice area, this paper) appears to confirm this suggestion.
Acknowledgements
We are most grateful to Jerzy Dzik (ZPAL) for providing the material for study. Warm thanks are also due to Stanisław Skompski (Faculty of Geology, University of Warsaw, Poland) and John-Paul Hodnett (New Mexico Museum of Natural History and Science, Albuquerque, USA) for the insightful reviews of the manuscript. The research was funded from the National Science Centre (NCN) grant 2016/23/B/ST10/03262.
References
Alexandrowicz, S.W. and Alexandrowicz, Z. 1999. Selected geosites of the Cracow Upland. Polish Geological Institute Special Papers 2: 53–60.
Appelt, J. 1995. Budowa geologiczna okolic Czatkowic koło Krakowa. 115 pp. Unpublished M.Sc. thesis, Uniwersytet Warszawski, Wydział Geologii, Warszawa.
Appelt, J. 1998. Tournaisian conodonts from the basinal carbonates of the Krzeszowice area, southern Poland. Acta Geologica Polonica 48: 135–140.
Billingsley, G.H. and Beus, S.S. 1999. Chapter A: Overview. In: G.H. Billingsley and S.S. Beus (eds.), Geology of the Surprise Canyon Formation of the Grand Canyon, Arizona. Museum of Northern Arizona, Bulletin 61: 1–8.
Cappetta, H., Duffin, C.J., and Zidek, J. 1993. Chondrichthyes. In: M.J. Benton (ed.), The Fossil Record 2, 593–609. Chapman and Hall, London.
Dean, B. 1909. Studies on fossil fishes (sharks, chimeroids and arthrodires). American Museum of Natural History, Memoir 9: 211–287.
Derycke, C., Ivanov, A., and Weber, H.M. 2005. A late Viséan vertebrate assemblage from Belgium. In: V. Hairapetian and M. Ginter (eds.), Devonian Vertebrates of the Continental Margins. IGCP 491 Yerevan Meeting. Yerevan, Armenia, May 22–27, 2005. Ichthyolith Issues Special Publication 8: 7–8.
Duncan, M. 1999. A Study of Some Irish Lower Carboniferous Fish Microvertebrates. 225 pp. Unpublished Ph.D. thesis, University of Dublin, Dublin.
Duncan, M. 2003. Early Carboniferous chondrichthyan Thrinacodus from Ireland, and a reconstruction of jaw apparatus. Acta Palaeontologica Polonica 48: 113–122.
Duncan, M. 2004. Chondrichthyan genus Lissodus from the Lower Carboniferous of Ireland. Acta Palaeontologica Polonica 49: 417–428.
Duncan, M. 2006. Various chondrichthyan microfossil faunas from the Lower Mississippian (Carboniferous) of Ireland. Irish Journal of Earth Sciences 24: 51–69. Crossref
Dzik, J. 1997. Emergence and succession of Carboniferous conodont and ammonoid communities in the Polish part of the Variscan sea. Acta Palaeontologica Polonica 42: 57–170.
Feichtinger, I., Engelbrecht, A., Lukeneder, A., and Kriwet, J. 2018. New chondrichthyans characterised by cladodont-like tooth morphologies from the Early Cretaceous of Austria, with remarks on the microstructural diversity of enameloid. Historical Biology [published online, https://doi.org/10.1080/08912963.2018.1539971] Crossref
Fischbein, S.A., Fielding, C.R., and Joeckel, R.M. 2009. Fluvial-estuarine reinterpretation of large, isolated sandstone bodies in epicontinental cyclothems, Upper Pennsylvanian, northern Midcontinent, USA, and their significance for understanding late Paleozoic sea-level fluctuations. Sedimentary Geology 216: 15–28. Crossref
Fournier, G. and Pruvost, P. 1928. Déscriptions des poissons élasmobranches du Marbre noir de Denée. Mémoires de la Société Géologique du Nord 9: 1–23.
Ginter, M. 1990. Late Famennian shark teeth from the Holy Cross Mts, Central Poland. Acta Geologica Polonica 40: 69–81.
Ginter, M. 1994. Ichtiolity dewońskie z Polski i Uralu, oraz ich znaczenie stratygraficzne. 134 pp. Unpublished Ph.D. dissertation, Uniwersytet Warszawski, Wydział Geologii, Warszawa.
Ginter, M. 1995. Ichthyoliths and Late Devonian events in Poland and Germany. Ichthyolith Issues Special Publication 1: 23–30.
Ginter, M. 1999. Famennian–Tournaisian chondrichthyan microremains from the eastern Thuringian Slate Mountains. Abhandlungen und Berichte für Naturkunde 21: 25–47.
Ginter, M. 2010. Teeth of Late Famennian ctenacanth sharks from the Cleveland Shale. In: D.K. Elliott, J.G. Maisey, X. Yu, and D. Miao (eds.), Morphology, Phylogeny and Paleobiogeography of Fossil Fishes, 145–158. Friedrich Pfeil, München.
Ginter, M. 2018. Symmoriiform sharks from the Pennsylvanian of Nebraska. Acta Geologica Polonica 68: 391–401.
Ginter, M. and Hansen, M. 2010. Teeth of the cladodont shark Denaea from the Carboniferous of central North America. In: D. Nowakowski (ed.), Morphology and Systematics of Fossil Vertebrates, 29–44. Wydawnictwo DN, Wrocław.
Ginter, M. and Skompski, S. (in press). The squamation of “Ctenacanthus” costellatus (Chondrichthyes: Ctenacanthiformes) from the Carboniferous of Lublin area, south-eastern Poland. Acta Geologica Polonica 69.
Ginter, M. and Sun, Y. 2007. Chondrichthyan remains from the Lower Carboniferous of Muhua, southern China. Acta Palaeontologica Polonica 52: 705–727.
Ginter, M., Duffin, C.J., Dean, M.T., and Korn, D. 2015. Late Viséan pelagic chondrichthyans from northern Europe. Acta Palaeontologica Polonica 60: 899–922. Crossref
Ginter, M., Hairapetian, V., and Klug, C. 2002. Famennian chondrichthyans from the shelves of North Gondwana. Acta Geologica Polonica 52: 169–215.
Ginter, M., Hampe, O., and Duffin, C.J. 2010. Chondrichthyes. Paleozoic Elasmobranchii. Teeth. In: H.-P. Schultze (ed.), Handbook of Paleoichthyology, vol. 3D, 1–168. Friedrich Pfeil, München.
Glikman, L.S. 1964. Akuly paleogena i ih stratigrafičeskoe značenie. 228 pp. Nauka, Moskva.
Gromczakiewicz-Łomnicka, A. 1974. Upper Viséan conodont fauna from the Carboniferous limestone north of Krzeszowice (environs of Cracow, Poland). Annales de la Société Géologique de Pologne 44: 475–481.
Guinot, G., Adnet, S., Cavin, L., and Cappetta, H. 2013. Cretaceous stem chondrichthyans survived the end-Permian mass extinction. Nature Communications 4: 2669 [published online, https://doi.org/10.1038/ncomms3669]. Crossref
Hay, O.P. 1902. Bibliography and catalogue of the fossil vertebrata of North America. Bulletin of the U.S. Geological Survey 179: 1–868. Crossref
Hodnett, J.-P.M. and Elliott, D.K. 2018. Carboniferous chondrichthyan assemblages from the Surprise Canyon and Watahomigi formations (latest Mississippian–Early Pennsylvanian) of the western Grand Canyon, Northern Arizona. Journal of Paleontology 92 (Memoir 77): 1–33. Crossref
Huxley, T. 1880. A Manual of the Anatomy of Vertebrated Animals. 431 pp. D. Appleton & Co., New York.
Ivanov, A.O., Nestell, G.P., and Nestell, M.K. 2013. Fish assemblage from the Capitanian (Middle Permian) of the Apache Mountains, West Texas, USA. In: S.G. Lucas, W.A. DiMichele, J.E. Barrick, J.W. Schneider, and J.A. Spielmann (eds.), The Carboniferous–Permian Transition. New Mexico Museum of Natural History and Science, Bulletin 60: 152–160.
Ivanov A.O., Nestell, M.K., and Nestell, G.P. 2015. Middle Permian fish microremains from the Early Capitanian of the Guadalupe Mountains, West Texas, USA. Micropaleontology 61: 301–312.
Long, J. 1990. Late Devonian chondrichthyans and other microvertebrate remains from northern Thailand. Journal of Vertebrate Paleontology 10: 59–71. Crossref
Lund, R. 1990. Chondrichthyan life history styles as revealed by the 320 million years old Mississippian of Montana. Environmental Biology of Fishes 27: 1–19. Crossref
Lund, R. 1985. The morphology of Falcatus falcatus (St. John and Worthen), a Mississippian stethacanthid chondrichthyan from the Bear Gulch Limestone of Montana. Journal of Vertebrate Paleontology 5: 1–19. Crossref
Lund, R. 1986. On Damocles serratus, nov. gen. et sp. (Elasmobranchii, Cladodontida) from the Upper Mississippian Bear Gulch Limestone of Montana. Journal of Vertebrate Paleontology 6: 12–19. Crossref
Maisey, J.G. 1975. The interrelationships of phalacanthous selachians. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1975 (9): 553–567.
Maisey, J.G. 2008. Some observations on Denaea fournieri (Chondrichthyes, Symmoriiformes) from the Lower Carboniferous of Belgium. Acta Geologica Polonica 58: 185–190.
Newberry, J.S. 1889. The Paleozoic fishes of North America. US Geological Survey, Monograph 16: 1–340. Crossref
Ossian, C.R. 1974. Paleontology, Paleobotany and Facies Characteristics of a Pennsylvanian Delta in Southeastern Nebraska. 20 + 393 pp. Unpublished Ph.D. dissertation, University of Texas, Austin [available at: https://www.academia.edu/1424478]
Paszkowski, M. 1995. Kraków Region. Description of stops 5–8. In: M. Szulczewski and J. Dvořák (eds.), Evolution of the Polish-Moravian carbonate platform in the Late Devonian and Early Carboniferous: Holy Cross Mts., Kraków Upland, Moravian Karst. XIII International Congress on Carboniferous–Permian, Kraków 1995, Guide to excursion B4, 23–28. Polish Geological Institute, Kraków.
Pradel, A., Maisey, J.G., Tafforeau, P., Mapes, R.H., and Mallatt, J. 2014. A Palaeozoic shark with osteichthyan-like branchial arches. Nature 509: 608–611. Crossref
Pruvost, P. 1922. Description de Denaea fournieri, sélacien nouveau du Marbre noir de Denée. Part 2 of Fournier, G. and Pruvost, P., Découverte d’un poisson nouveau dans le marbre noir de Denée. Bulletin de Academie Royal de Bruxelles, Serie 5 8: 213–218.
Reif, W.-E. 1978. Types of morphogenesis of the dermal skeleton in fossil sharks. Paläontologische Zeitschrift 52: 110–128. Crossref
Riemann, F., Schülke, I., and Thies, D. 2002. Mikrovertebratenreste aus dem basalen Famennium (triangularis-bis crepida Zone) der Montagne Noire (Frankreich). Geologica et Palaeontologica 36: 1–43.
Savitskiy, Y.V. [Savitskij, Û.V.], Ivanov, A.O., and Orlov, A.N. 2000. Atlas mikroostatkov organizmov nižnego karbona severo-zapadnogo kryla Moskovskoj sineklizy (ostrakody, konodonty, pozvonočnye). Čast’ 1, mstinskaya i putinskaya svity, 68 pp. Sankt-Peterburgskij Universitet, Sankt-Petersburg.
Skompski, S. 2003. Początek późnopaleozoicznego zlodowacenia Gondwany i jego zapis w polskich sukcesjach karbońskich. Przegląd Geologiczny 51: 658–662.
Skompski, S. 2008. Stop 5. Czatkowice Quarry. In: M. Ginter (ed.), Evolution and Diversity of Chondrichthyans. Warsaw Meeting, 2008. Ichthyolith Issues, Special Publication 11: 55–56.
Soboń-Podgórska, J. 1975. Stratygrafia karbonu dolnego w Czernej koło Krzeszowic na podstawie mikrofauny. Biuletyn Instytutu Geologicznego 282: 249–271.
St. John, O. and Worthen, A.H. 1875. Geology and Palaeontology. Part II. Palaeontology of Illinois. Section I. Descriptions of fossil fishes. Geological Survey of Illinois 6: 245–488.
Turner, S. 1982. Middle Palaeozoic elasmobranch remains from Australia. Journal of Vertebrate Paleontology 2: 117–131. Crossref
Wang, N.-Z., Jin, F., and Wang, W. 2004. Early Carboniferous fishes (acanthodian, actinopterygians and Chondrichthyes) from the east sector of north Qilian Mountain, China—Carboniferous fish sequence from the east sector of north Qilian Mountain (1) [in Chinese with English summary]. Vertebrata Palasiatica 42: 89–110.
Wang, S.-T. 1989. Biostratigraphy of vertebrate microfossils. Vertebrate microfossils. In: J. Qiang (ed.), The Dapoushang Section, 36–38, 103–108. Science Press, Beijing.
Williams, M.E. 1985. The “Cladodont level” sharks of the Pennsylvanian black shales of central North America. Palaeontographica A 190: 83–158.
Williams, M.E. 1998. A new specimen of Tamiobatis vetustus (Chondrichthyes, Ctenacanthoidea) from the Late Devonian Cleveland Shale of Ohio. Journal of Vertebrate Paleontology 18: 251–260. Crossref
Zajączkowski, W.A. 1975. Stratygrafia i litologia wapieni dinantu w Czernej koło Krzeszowic. Biuletyn Instytutu Geologicznego 282: 273–326.
Zangerl, R. 1981. Chondrichthyes I. Paleozoic Elasmobranchii. In: H.P. Schultze (ed.), Handbook of Paleoichthyology, 3A, 1–115. Gustav Fischer, Stuttgart.
Zangerl, R. 1990. Two new stethacanthid sharks (Stethacanthidae, Symmoriida) from the Pennsylvanian of Indiana, U.S.A. Palaeontographica A 213: 115–141.
Comparison of selected dental features of Mississippian Falcatidae. S, symmetrical; AS, asymmetrical.
| |
Number of cusps |
Crown |
Median cusp labial ornament |
Orolingual button |
Stratigraphic range |
Source |
|
Disarticulated specimens |
||||||
|
(i) Denaea cf. fournieri |
5–7 |
S to slightly AS |
subparallel cristae |
weak hump |
upper Viséan, |
|
|
(ii) Denaea cf. fournieri |
7–9 |
S to slightly AS |
subparallel cristae |
weak hump |
upper Viséan, |
|
|
(iii) Denaea cf. fournieri |
5 |
S |
central cristae converge |
pronounced lingual |
upper Viséan, Brigantian |
|
|
(iv) “Stethacanthus” gansuensis |
5 |
S |
central cristae converge |
wide, lingual, weak |
Serpukhovian |
|
|
(v) Denaea williamsi |
5–7 |
S |
all cristae |
pronounced lingual |
upper Viséan, Asbian– |
|
|
(vi) Denaea wangi |
5–11 |
AS |
smooth |
none |
upper Viséan, Asbian– |
|
|
(vii) Falcatidae indet. 1 |
5–7 |
S to AS |
subparallel cristae |
weak to pronounced |
upper Tournaisian |
this paper |
|
(viii) Falcatidae indet. 2 |
5–7 |
S |
subparallel cristae |
pronounced lingual |
upper Viséan, |
this paper |
|
Articulated specimens |
||||||
|
Falcatus falcatus |
5 |
S to slightly AS |
? |
? |
Serpukhovian |
|
|
Damocles serratus |
5 |
S |
? |
? |
Serpukhovian |
|
|
Denaea fournieri |
5–7 |
S to slightly AS |
subparallel cristae |
weak hump? |
lower Viséan, Moliniacian |
|
|
Ozarcus mapesae |
5 |
S |
? |
? |
upper Viséan, Brigantian |
|
|
CM 35412 |
8 |
AS |
smooth |
? |
Serpukhovian |
|
Acta Palaeontol. Pol. 64 (3): 549–564, 2019
https://doi.org/10.4202/app.00619.2019