
Mollusks and a crustacean from early Oligocene methane-seep deposits in the Talara Basin, northern Peru
STEFFEN KIEL, FRIDA HYBERTSEN, MATÚŠ HYŽNÝ, and ADIËL A. KLOMPMAKER
Kiel, S., Hybertsen, F., Hyžný, M., and Klompmaker, A.A. 2020. Mollusks and a crustacean from early Oligocene methane-seep deposits in the Talara Basin, northern Peru. Acta Palaeontologica Polonica 65 (1): 109–138.
A total of 25 species of mollusks and crustaceans are reported from Oligocene seep deposits in the Talara Basin in northern Peru. Among these, 12 are identified to the species-level, including one new genus, six new species, and three new combinations. Pseudophopsis is introduced for medium-sized, elongate-oval kalenterid bivalves with a strong hinge plate and largely reduced hinge teeth, rough surface sculpture and lacking a pallial sinus. The new species include two bivalves, three gastropods, and one decapod crustacean: the protobranch bivalve Neilo altamirano and the vesicomyid bivalve Pleurophopsis talarensis; among the gastropods, the pyropeltid Pyropelta seca, the provannid Provanna pelada, and the hokkaidoconchid Ascheria salina; the new crustacean is the callianassid Eucalliax capsulasetaea. New combinations include the bivalves Conchocele tessaria, Lucinoma zapotalensis, and Pseudophopsis peruviana. Two species are shared with late Eocene to Oligocene seep faunas in Washington state, USA: Provanna antiqua and Colus sekiuensis; the Talara Basin fauna shares only genera, but no species with Oligocene seep fauna in other regions. Further noteworthy aspects of the molluscan fauna include the remarkable diversity of four limpet species, the oldest record of the cocculinid Coccopigya, and the youngest record of the largely seep-restricted genus Ascheria. Eucalliax is recorded for the first time from a seep and from the Oligocene.
Key words: Gastropoda, Bivalvia, Crustacea, Decapoda, biogeography, cold-seep, Paleogene, South America.
Steffen Kiel [steffen.kiel@nrm.se] and Frida Hybertsen [frida.hybertsen@nrm.se], Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, 10405 Stockholm, Sweden.
Matúš Hyžný [matus.hyzny@uniba.sk], Department of Geology and Paleontology, Faculty of Natural Sciences, Comenius University, Ilkovičova 6, 842 15 Bratislava, Slovakia.
Adiël A. Klompmaker [adielklompmaker@gmail.com], Department of Integrative Biology & Museum of Paleontology, University of California, Berkeley, 1005 Valley Life Sciences Building #3140, Berkeley, CA 94720, USA; The University of Alabama Museums & Alabama Museum of Natural History, The University of Alabama, Box 870340, Tuscaloosa, AL 35487, USA.
Received 2 May 2019, accepted 20 September 2019, available online 6 December 2019.
Copyright © 2020 S. Kiel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Our understanding of the fossil record of deep-water methane-seep communities, especially of the Cenozoic Era, has remarkably improved over the last decade. However, most systematic studies are restricted to a few regions in the northern Hemisphere, including western Washington state, USA (Kiel 2006, 2008; Kiel and Amano 2013; Hybertsen and Kiel 2018), Japan (Amano and Kiel 2010, 2011, 2012; Amano and Jenkins 2011a, b, 2013; Amano and Little 2014; Amano et al. 2014a; Miyajima et al. 2017), and Italy (Kiel and Taviani 2017, 2018; Kiel et al. 2018), and only a few studies have covered more than one region or had a world-wide scope (Amano and Kiel 2007; Kiel 2013; Gill and Little 2013; Kiel and Hansen 2015; Amano et al. 2015; Hryniewicz et al. 2017). For the southern Hemisphere, thorough taxonomic work has only been carried out for the Miocene seep deposits of New Zealand (Saether et al. 2010a, b, 2012, 2016; Amano et al. 2014b, 2015, 2018b), and a single Miocene site has been reported from central Chile (Contardo-Berríos et al. 2017).
Seep communities from Oligocene strata in northern Peru have long been known (Olsson 1931; Goedert and Squires 1993; Kiel and Peckmann 2007), but these species-rich faunas have never been systematically revised, except for a few lucinid bivalve specimens from Olsson’s original collection (Kiel 2013). Here we report on newly collected mollusk and crustacean specimens from these seep deposits, and describe one new genus and six new species, provide taxonomic updates on several of the known species, and provide a comparison between Olsson’s collection and the new fossils.
Institutional abbreviations.—NRM, Swedish Museum of Natural History, Stockholm, Sweden; PRI, Paleontological Research Institution, Ithaca, USA; USNM, Smithsonian Museum of Natural History, Washington, DC, USA.
Other abbreviations.—L, length; W, width; H, height; RV, right valve; LV, left valve.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:CA2D2F4A-AE41-441F-A205- B334CD055FBE
Material and methods
Details on the sampling localities in the Talara Basin in northern Peru (Fig. 1) have been presented previously (Kiel et al. 2019). In short, the material reported here originates from three areas: (i) The Belén seep site, named after a long-vanished settlement from the initial oil boom in the early 1900s, situated about 10 km SE of Negritos. (ii) The Cerro La Salina hills along the southern side of Quebrada Seca north of Mancora, where nine individual limestone concretions were sampled; these are here referred to as La Salina blocks 1 to 9. (iii) The Cerros El Pelado hills along the northern side of Quebrada Seca north of Mancora; three individual limestone concretions were sampled here, but only one of them (block 2) yielded identifiable fossils (Kiel et al. 2019). The fossils from all three areas are of early Oligocene age, with a small chance that those from Cerros El Pelado may be of late Eocene age (Kiel et al. 2019).
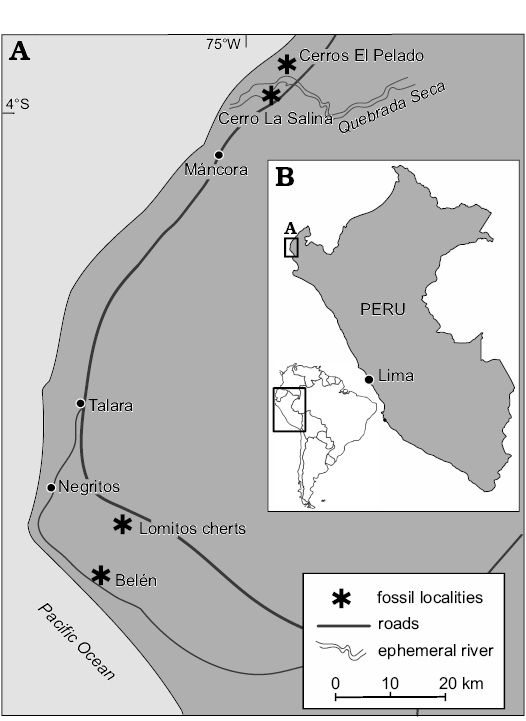
Fig. 1. Locality map of the seep deposits in the Talara Basin in northern Peru, where the here described mollusk and crustacean taxa were found (adopted from Kiel et al. 2019).
All specimens are housed at NRM, except for material collected by Axel A. Olsson, including one neritid specimen from the Lomitos cherts about 6 km east of Negritos, which was mentioned but not illustrated by Olsson (1931), housed at PRI, and the holotypes of Vesicomya tschudi Olsson, 1931, and V. ramondi Olsson, 1931, both housed at PRI. Lastly, we illustrate several specimens of “Pleurophopsis” peruviana Olsson, 1931, housed at USNM. These specimens were collected by Wendell P. Woodring and Axel A. Olsson on January 15, 1958, at USGS locality 21219, which is located on the southeastern side of Cerro La Salina and has previously been characterized petrographically and isotopically (Kiel and Peckmann 2007).
The specimens were extracted and prepared from the rock matrix using a pneumatic hammer. Small specimen were photographed using a Cognisys Stackshot 3X stacking image system and the Zerene Stacker software, combining 15 to 25 pictures for each composite image. All specimens were coated with ammonium chloride for photography, except when noted otherwise.
Systematic paleontology
Phylum Mollusca Linnaeus, 1758
Class Bivalvia Linnaeus, 1758
Subclass Protobranchia Pelseneer, 1889
Family Malletiidae Adams and Adams, 1858
Genus Neilo Adams, 1854
Type species: Neilo cumingii Adams, 1854 [= Neilo australis (Quoy and Gaimard, 1835)], by monotypy; Recent, New Zealand.
Neilo altamirano sp. nov.
Fig. 2A.
Zoobank LCID: urn:lsid:zoobank.org:act:4A36FC09-E842-43D3-9D FC-D55D9A246345
Etymology: In honour of Ali Altamirano (Lima, Peru), for his help collecting the fossils.
Holotype: NRM Mo187001, an internal and external mold of LV, with some shell material preserved.
Type locality: Cerro La Salina (block 1), northern Peru.
Type horizon: Presumably the early Oligocene part of the Heath shale.
Material.—Type material only.
Diagnosis.—Well-inflated, subrectangular shell, tallest at umbo, strongest inflation at posterior third; anterior margin truncate, straight; posterior margin straight; posterior ridge well defined; sculpture of irregularly spaced, blunt commarginal ribs, interspaces roughly twice the width of the ribs.
Dimensions (in mm).—The holotype NRM Mo187001, L = 20, H = 12, and the single valve ca. 4 mm thick.
Description.—Shell subrectangular, well-inflated, postumbonal part about 2/3 of shell length, maximum shell height at umbo; anterodorsal margin slightly convex, anterior margin truncate and straight, ventral margin slightly convex due to angulation opposite of umbo, posterior margin truncate, posterodorsal margin very slightly concave. Posterior area defined by ridge running from behind umbones to posteroventral corner; sculpture consists of irregularly spaced, blunt, commarginal ribs, with fine commarginal lines between; these ribs bent sharply upward at posterior ridge. Distinct groove internally from umbo to anteroventral margin; anterior adductor muscle scar oval and just anterior to this groove, posterior adductor muscle scar indistinct; hinge with row of taxodont teeth.

Fig. 2. Protobranch and pteriomorph bivalves from early Oligocene seep deposits from the Cerro La Salina (block 1, A; block 6, D; block 9, C) and Belén seep deposit (B), Talara Basin, northern Peru. A. Malletiid Neilo altamirano sp. nov. (NRM Mo187001, holotype), internal mold in lateral view (A1); external mold in lateral view (A2), arrows indicating the posterior ridge; view on the dorsal side (A3); close-up on the taxodont hinge dentition (A4). B. The possible nuculid Acila? sp. (NRM Mo187002), external mold of outer shell surface. C. The bathymodiolin Idas sp. (NRM Mo187003), internal mold of the entire specimen (C1), arrow indicating the taxondont teeth; close-up on taxodont teeth on posterodorsal shell margin (C2). D. Propeamussiidae indet. (NRM Mo187004), uncoated specimen showing internal radial ridges (D1); specimen coated with ammonium-chloride, highlighting external sculpture (D2).
Remarks.—No Neilo species were reported from the Eocene to Miocene deposits in Peru and Ecuador so far. Quite similar in shell outline and sculpture are Neilo ortmanni Erdmann and Morra, 1985, from the late Eocene to early Oligocene San Julián Formation in eastern Patagonia (Argentina) and Neilo ornata (Sowerby, 1846) from late Oligocene–early Miocene sediments near Puerto Deseado in eastern Patagonia (Erdmann and Morra 1985; Del Río and Camacho 1998; Griffin and Nielsen 2008). Both species differ from N. altamirano by their rostrate posterior margin, whereas N. altamirano has a vertically truncated, almost straight posterior margin. The early Miocene Neilo volckmanni (Philippi, 1887) from the Navidad Formation in Chile is quite different by having a pentagonal outline and a distinctive change in sculpture from indistinct anterior of the diagonal posterior ridge to granular-commarginal on the posterodorsal side of the shell. Neilo awamoana Finlay, 1926, a species common in the offshore siltstone facies of the early Miocene of New Zealand, is similar in outline, but has more numerous, finer, and more closely spaced collabral ribs, and a fine ridge running along the angulation from the umbo to the posteroventral margin (Beu and Maxwell 1990: pl. 16a, c). The late Oligocene to early Miocene Neilo sinangula Finlay, 1926, from New Zealand lacks this fine ridge (Finlay 1926; Marshall 1978; Eagle and Hayward 1993), but also has finer and more numerous commarginal ribs compared to N. altamirano. The early Oligocene Neilo funiculata Hutton, 1887 from New Zealand has a much longer anterior part of the shell than N. altamirano sp. nov. (Beu et al. 2012).
Stratigraphic and geographic range.—Type locality and horizon only.
Family Nuculidae Gray, 1824
Genus Acila Adams and Adams, 1858
Type species: Nucula divaricata Hinds, 1843, by subsequent designation; Recent, northern and tropical Pacific Ocean.
Acila? sp.
Fig. 2B.
Material.—One specimen (NRM Mo187002, W = 10.5 mm) from the Oligocene Belén seep site, northern Peru.
Remarks.—Judging from the divaricate ornament, this specimen appears to belong to Acila, but as other diagnostic features, such as the hinge, are missing, it is here hesitantly assigned to this genus. Olsson reported two species of Acila from Belén and Pajarabobo, Acila paita Olsson, 1931, and Acila piura Olsson, 1931. The specimen that we collected is too poorly preserved to be identified as either of these species.
Subclass Pteriomorphia Beurlen, 1944
Family Mytilidae Rafinesque, 1815
Subfamily Bathymodiolinae Kenk and Wilson, 1985
Genus Idas Jeffreys, 1876
Type species: Idas argenteus Jeffreys, 1876; Recent, North Atlantic Ocean.
Idas sp.
Fig. 2C.
Material.—One specimen (NRM Mo187003, L = 4 mm) from the Oligocene Cerro La Salina block 9, northern Peru.
Remarks.—The single specimen consists of RV mostly lacking shell material; it has straight dorsal and ventral margins, a pointed anterior margin, the umbo is positioned at about 10% of the total shell length from the anterior, and imprints of small, taxodont teeth can be seen at the posterior end of the posterodorsal margin. It is too poorly preserved to be identified, but seems to differ from the coeval Idas olympicus from Washington state, USA, by a less anteriorly positioned umbo (Kiel and Goedert 2007; Kiel and Amano 2013). Compared to geologically younger small bathymodiolins associated to whale falls in Japan, it differs by having a more modioliform shell and a more anteriorly positioned umbo (Amano and Little 2005; Amano et al. 2007; Jenkins et al. 2018a). Idas sp. reported from a whale fall at Orciano Pisano in Italy has parallel dorsal and ventral margin unlike the Peruvian specimen, and Idas reported from a Miocene seep deposit at Ca’ Fornace in northern Italy has a more curved shell compared to the Peruvian specimen.
Family Propeamussiidae Abbott, 1954
Propeamussiidae indet.
Fig. 2D.
Material.—One specimen (NRM Mo187004) from the Oligocene Cerro La Salina block 6, northern Peru.
Remarks.—The single specimen lacks ears and cannot be determined to the genus-level. Propeamussiids, also known as “mud pecten”, are common dwellers of muddy sediments in deeper water and have been reported from several fossil seep deposits (Kelly et al. 2000; Kiel et al. 2008; Kiel 2010) and wood falls (Kiel and Goedert 2006a, b; Amano et al. 2018a).
Subclass Heterodonta Neumayr, 1884
Family Kalenteridae Marwick, 1953
Genus Pseudophopsis nov.
Zoobank LCID: urn:lsid:zoobank.org:act:9FF851DC-2A55-4CC0-888D-6E91D0042CB1
Type species: Pleurophopsis peruviana Olsson, 1931; Oligocene, Peru.
Species included: The type species and Unio bitumen Cooke, 1919.
Etymology: For its superficial similarity to the vesicomyid genus Pleurophopsis.
Diagnosis.—Medium-sized, well-inflated, elongate-oval kalenterid with blunt, somewhat elevated umbo in anterior third of shell; external sculpture rough, irregular; hinge plate strong, moderately narrow, with indistinct growth lines, hinge teeth weak, sub-parallel to shell margin, often reduced in adults; anterior adductor muscle scar deep, roundish, with peg in upper half of posterior margin; anterior retractor muscle scar large, roundish, above ridge bordering adductor muscle scar; onset of pallial line at posteroventral side of anterior adductor muscle scar, no pallial sinus; ligament external, long, strong.
Remarks.—Van Winkle (1919) derived the generic name Pleurophopsis from the Paleozoic “modiomorphid” genus Pleurophorus King, 1848 (a synonym of the beetle genus Pleurophorus Mulsant, 1842 and hence replaced by Permophorus Chavan, 1954) because she considered Pleurophopsis to be a descendent of Pleurophorus based on the hinge structure and muscle scar pattern. Olsson (1931) adopted this classification when introducing Pleurophopsis peruviana, and placed Pleurophopsis in the Permophoridae. It is now firmly established that the type species of Pleurophopsis, P. unioides (see below), belongs to the Vesicomyidae (Kiel 2007; Krylova et al. 2010; Krylova and Sahling 2010). However, Olsson’s (1931) “Pleurophopsis” peruviana does indeed show “modiomorphid” hinge and muscle scar features and hence the new genus Pseudophopsis, with P. peruviana as the type species, is here placed among the Kalenteridae.
In contrast to vesicomyids, Pseudophopsis peruviana has a relatively large anterior pedal retractor scar that is clearly separated from the adductor muscle scar. The entire anterior adductor muscle scar is sunken, whereas in vesicomyids it is usually deepest on its posterior side and shallower impressed on the anterior side. Another distinguishing feature is the hinge plate, which is set distinctly inward from the umbones, whereas is situated directly underneath the umbones in vesicomyids (or in other words, the umbones are more loosely coiled than in vesicomyids). Such a distinctive hinge plate is for example seen in the kalenterid Myoconcha neuquena Leanza, 1940 from the Early Jurassic of Argentina (Leanza 1940; Griffin and Pastorino 2006). Similar fine growth lines on the hinge plate of P. peruviana (Fig. 3G1) can be seen in Myoconcha neuquena and also in the Miocene kalenterid Madrynomya bruneti Griffin and Pastorino, 2006. Madrynomya is clearly distinct from Pseudophopsis by its more coiled and more pointed umbones, and the much broader but shorter hinge plate. Myoconcha Sowerby, 1824, differs from Pseudophopsis by its bean-shaped posterior adductor muscle scar, and the anterior side of the pallial line, which is distinct from the anterior adductor muscle scar in Myoconcha, but starts at its posteroventral corner in Pseudophopsis (i.e., Leanza 1940; Newell 1957; Griffin and Pastorino 2006; Kaim and Schneider 2012). Lastly, Pseudophopsis peruviana has a coarser and much more irregular surface sculpture than any vesicomyid, and also its ligament is longer relative to shell length as in vesicomyids.
The late Mesozoic, seep-inhabiting kalenterid Caspiconcha Kelly in Kelly et al., 2000, typically has an elongated anterior adductor muscle scar with a posterodorsal projection, which is absent in Pseudophopsis, a more distinctive projection above the anterior adductor muscle scar (the so-called caspiconchid process) not seen in Pseudophopsis, and it lacks hinge dentition (Kelly et al. 2000; Kiel et al. 2010; Jenkins et al. 2013, 2018b). Further, several but not all species of Caspiconcha have an internal ridge running from the umbo to the posterior adductor scar, which is not seen in P. peruviana, and the pallial line tends to be closer to the ventral shell margin in Pseudophopsis than in Caspiconcha (see Jenkins et al. 2018b: fig. 2).
Another species included here in Pseudophopsis is the Eocene Unio bitumen Cooke, 1919, from the Elmira asphalt mine seep deposit in Cuba (Cooke 1919; Kiel and Peckmann 2007; Kiel and Hansen 2015). Pseudophopsis bitumen resembles P. peruviana by having a similar inward-set hinge plate with similarly reduced teeth, its thick ligament and very rough and irregular surface.
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru and Eocene of Cuba.
Pseudophopsis peruviana (Olsson, 1931)
Fig. 3.
1931 Pleurophopsis peruviana sp. nov.; Olsson 1931: 43–44, pl. 4: 1, 3.
Material.—One specimen (in two parts: NRM Mo187027 and NRM Mo187028) from Cerro La Salina block 1; two unnumbered specimens from Cerro La Salina block 3; several unnumbered fragments from Cerros El Pelado block 2, and several specimens (including USNM PAL 530294, 530294, 728207–728209) collected by Axel A. Olsson and Wendell P. Woodring at USGS loc. 21219. All upper Oligocene, Talara Basin, Peru. The shells reach 10 cm in length.
Remarks.—The Eocene Cuban Pseudophopsis bitumen (Cooke, 1919) has a hinge dentition that is quite similar that of small specimens of Pseudophopsis peruviana (see Kiel and Peckmann 2007: figs. 4A, B). Pseudophopsis bitumen differs from P. peruviana by having the maximum inflation in the middle, whereas it is located toward the anterior in P. peruviana.
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru.
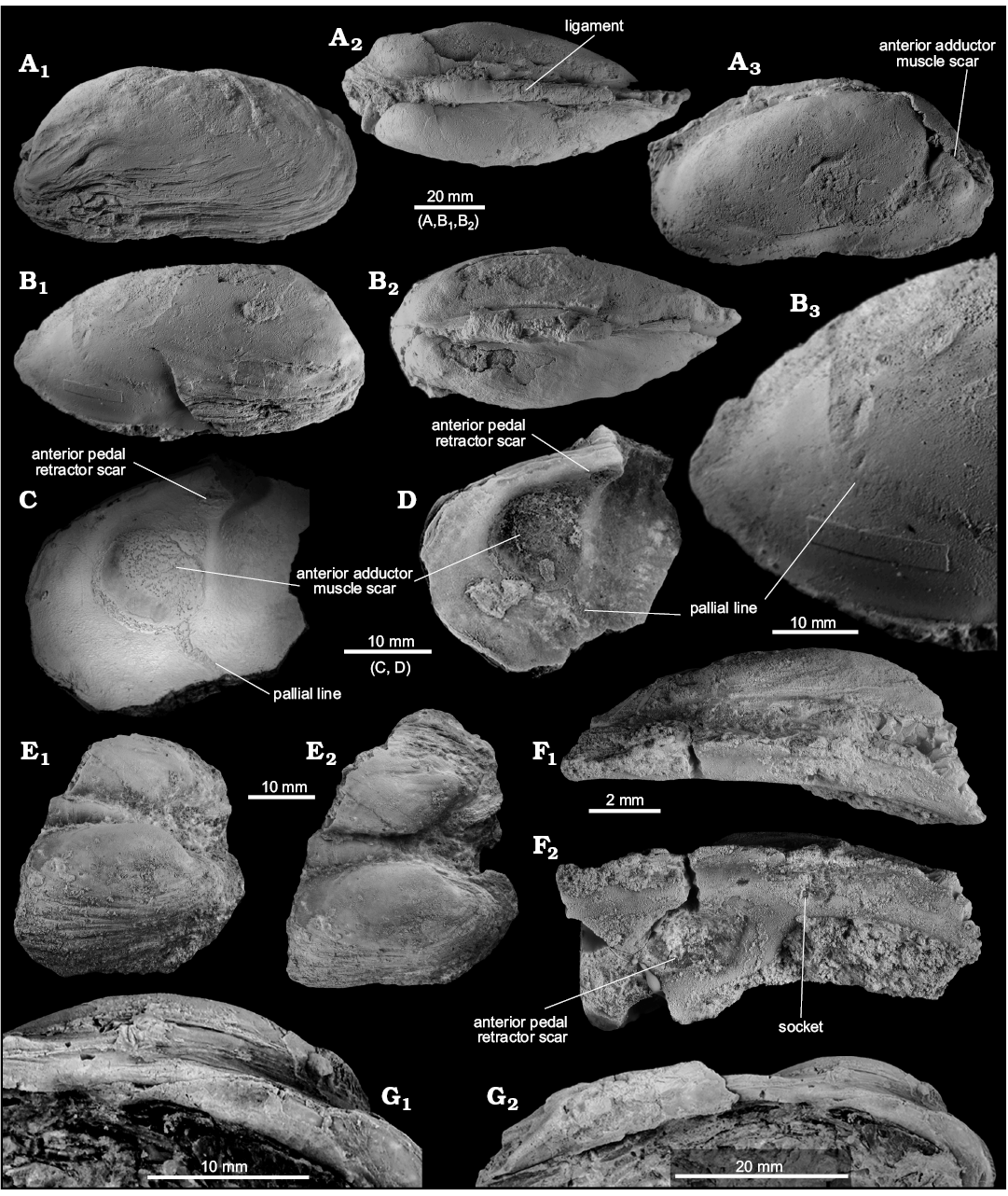
Fig. 3. The kalenterid Pseudophopsis peruviana (Olsson, 1931) from early Oligocene seep deposits at Cerro La Salina (USGS loc. 21219, A–D; block 3, E, F), Talara Basin, northern Peru; A. Large articulated specimen (USNM PAL 728207) showing rough and very irregular growth increments; outer side of left valve (A1), the long and thick ligament (A2) and deep anterior adductor muscle scar (A3). B. Large articulated specimen (USNM PAL 728208) showing posterior half of pallial line (B1, B3) and thick ligament (B2). C, D. Inner sides of two fragmentary right valves preserving anterior margin (C, USNM PAL 728209; D, USNM PAL 530294), showing anterior adductor muscle scar and pedal retractor scar, and onset of pallial line. E. Fragment (NRM Mo187027) preserving anterior half of shell and thick calcified ligament; outer side of right valve (E1) and dorsal view (E2). F. Fragment (NRM Mo187028) preserving edentulous hinge area (F1) and large anterior pedal retractor scar (F2). G. Dorsal shell margin of large left valve (USNM PAL 530295), showing edentulous hinge (G1, close-up). D and G from Kiel and Peckmann (2007: fig. 6).
Family Thyasiridae Dall, 1900
Genus Conchocele Gabb, 1866
Type species: Thyasira bisecta Conrad, 1849; Miocene, Astoria Formation, Oregon, USA.
Conchocele tessaria (Olsson, 1931)
Fig. 4.
1931 Thyasira tessaria sp. nov.; Olsson 1931: 53–54, pl. 6: 10, 14.
Material.—Six specimens (NRM Mo187005–187010, Fig. 4) and several further unnumbered specimens (often fragmentary) from Cerro La Salina blocks 1, 4, 6–9, upper Oligocene, Talara Basin, Peru.
Description.—Small to medium-sized for the genus (L max. = 35 mm), inflation moderate (L/W ratio ca. 1.9), rounded-pentagonal in outline; beak prosogyrate, elevated and pointed, umbonal angle 90–100°; posterior fold deep but not sharp, increasingly curved in larger specimens, with resulting in shallow posterior sinus; a weak, secondary sulcus also present in some specimens; anterior margin narrow, demarcated by blunt ridge; lunule symmetrical and well-demarcated; escutcheon lanceolate; external surface sculptured by irregular, commarginal growth lines only. Hinge plate narrow, edentulous, ligament nymph broad and elongate; internal surface with fine radial striations developing tubercles near ventral margin.
Remarks.—Olsson (1931) described three thyasirid species from northern Peru: T. peruviana and T. staufti from the Lomitos cherts, and T. tessaria from the Heath Formation at Pajarabobo, near Belén. Examination of Olsson’s (1931) type material at PRI showed that the specimens of all three species are more inflated than the material we collected; strongly so in the cases of T. peruviana and T. staufti, and less so in the case of T. tessaria. Based on the overall similarity of our specimens to T. tessaria, especially their pentagonal outline, the broad posterodorsal area and the blunt posterior ridge, we considered the material reported here as belonging to T. tessaria. Thyasira peruviana differs from T. tessaria by its much more prominent umbo, the much broader anterior margin, and the much smaller posterodorsal area. Thyasira staufti differs from T. tessaria by its deeper and sharper posterior sulcus, its narrower posterodorsal area, and its more oval outline. Thyasira montanita (Olsson, 1931) from allegedly coeval strata in southern Ecuador has finer growth lines, a more rounded outline, a broader anterior margin, and a broader and shorter anterior adductor muscle scar (personal observations SK).
Thyasira tessaria is here transferred to Conchocele based on its anteriorly situated beaks, the steeply sloping anterior margin, the weakly impressed lunule, and the broadly rounded posterior margin bearing a deep sulcus, characteristics of Conchocele that are not present in this combination in Thyasira. Conchocele tessaria shows the following differences to other Conchocele species: C. townsendi from Late Cretaceous strata on Seymour Island, Antarctica, has a similar pentagonal shape and narrow anterior margin, but has a deeper and sharper posterior fold (Hryniewicz et al. 2017). The Paleocene C. conradii from Svalbard is more inflated and has a longer anterior margin than C. tessaria (Hryniewicz et al. 2017). Also C. taylori Hickman, 2015, known from late Eocene to Oligocene strata of the North Pacific region, has a longer anterior margin, but its posterior fold is similar to that of C. tessaria (Hickman 2015; Hryniewicz et al. 2017). Another late Eocene to Oligocene species is Conchocele bathyaulax Hickman, 2015 from Oregon and Washington state, USA; compared to C. tessaria, it is much more inflated and has a more pointed, terminal umbo (Hickman 2015; Hryniewicz et al. 2017). The late Eocene C. kiritachiensis Hryniewicz, Amano, Jenkins, and Kiel, 2017, from a seep carbonate in Hokkaido, northern Japan, differs from Conchocele tessaria by its rhomboidal shape, very broad anterior margin, and pointed, terminal umbo (Hryniewicz et al. 2017). Finally, Conchocele adoccasa (Van Winkle, 1919) from a Miocene seep deposit in Trinidad has a broader anterior margin and a more distinct (sharper) posterior fold. This applies also to the specimens assigned to C. adoccasa from presumably Oligocene seep deposits of Colombia (Kiel and Hansen 2015). Overall, most similar to C. tessaria is the late Eocene to Oligocene C. taylori that is widespread along the North Pacific margin (Hickman 2015; Hryniewicz et al. 2017).
One specimen (NRM Mo187008) shows two indentations on LV, which might represent a failed attack by a predator such as a durophagous crustacean. Damage to fossil seep mollusks attributed crustaceans has been noted previously (e.g., Kiel et al. 2016, and references therein, Klompmaker et al. 2018).
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru.
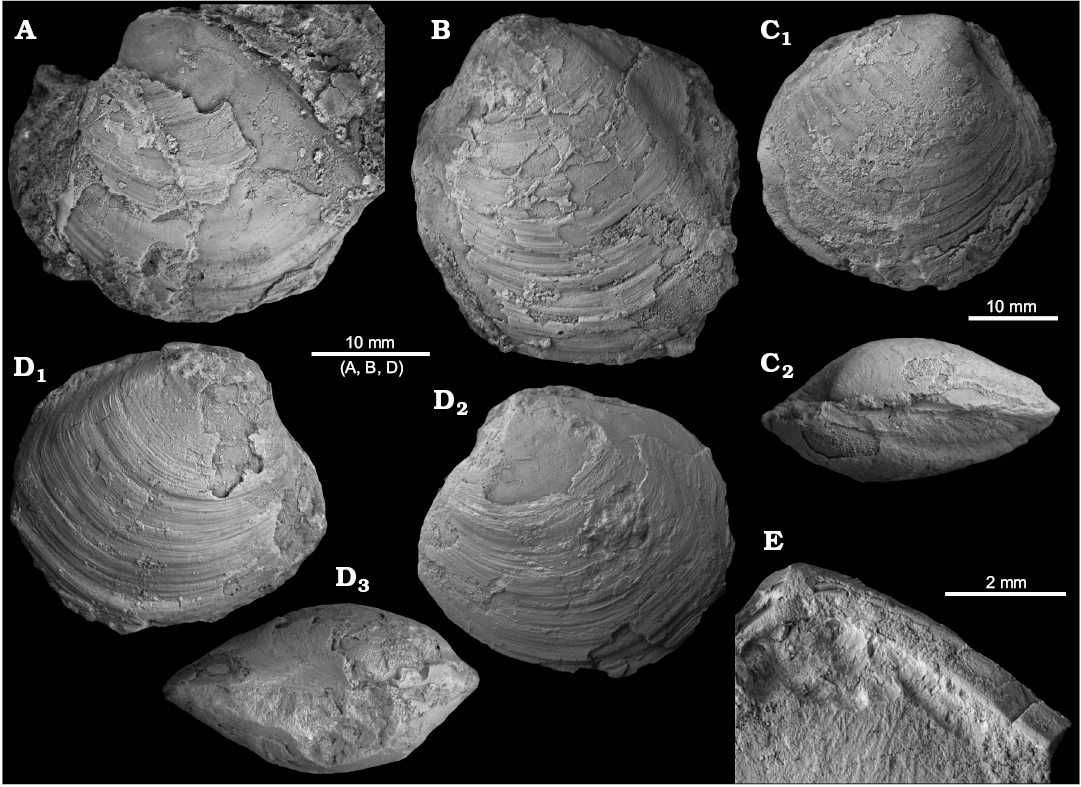
Fig. 4. The thyasirid Conchocele tessaria (Olsson, 1931) from early Oligocene seep deposits at Cerro La Salina (block 1, E; block 7, A–C; block 8, D), Talara Basin, northern Peru. A, B. Left valve of large specimen (A, NRM Mo187005; B, NRM Mo187006). C. Medium-sized specimen (NRM Mo187007), right valve showing fine growth increments (C1), dorsal view showing lunule (C2), left valve showing dorsal sulcus (C3). D. Small specimen (NRM Mo187008), left valve showing a healed shell injury (D1), right valve showing dorsal sulcus (D2) and dorsal view showing lunule posterior area (D3). E. Small specimen (NRM Mo187010) showing hinge of right valve.
Family Lucinidae Fleming, 1828
Genus Lucinoma Dall, 1901
Type species: Lucinoma filosa (Stimpson, 1851), by original designation; Recent, North Atlantic Ocean.
Lucinoma zapotalensis (Olsson, 1931)
Fig. 5.
1931 Phacoides (Lucinoma) zapotalensis sp. nov.; Olsson 1931: 49, pl. 5: 2, 5.
Material.—Three specimens (NRM Mo187011–187013) and several unnumbered, mostly fragmentary specimens from Cerro La Salina blocks 1, 3, 6–9, upper Oligocene, Talara Basin, Peru.
Remarks.—This species was reported by Olsson (1931) only from the Mambri shales near Zapotal in southern Ecuador, but not from any of the seep deposits in the Heath shale in northern Peru. According to Olsson (1931), the Mambri shales are the equivalent of the Heath shale and also of late Oligocene age. Similar is the “middle” Oligocene Lucinoma playaensis (Olsson, 1964) from the Playa Rica grits in northern Ecuador, though it differs from L. zapotalensis by having more numerous and more narrowly spaced ribs and a more rounded posterodorsal margin (Olsson 1964: pl. 6: 7). Lucinoma acutilineatum (Conrad, 1849) from the Miocene Astoria Formation in Oregon and Washington state, USA, is quite similar, but appears to differ from L. zapotalensis by being less inflated and having more widely spaced, and sharper, commarginal ribs; it also seems to have a more angular outline than L. zapotalensis (Conrad 1849; Moore 1963).
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru.
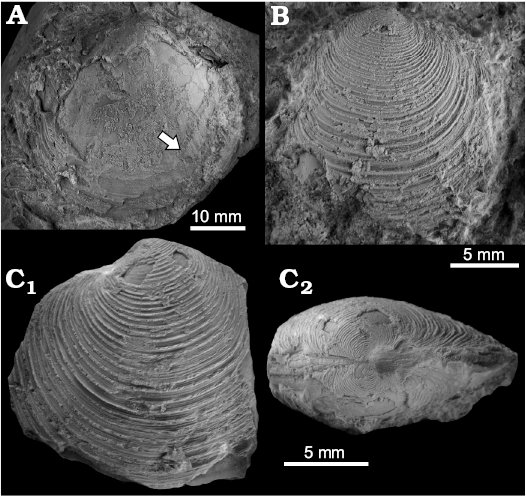
Fig. 5. The lucinid Lucinoma zapotalensis (Olsson, 1931) from early Oligocene seep deposit at Cerro La Salina (block 1, C; block 9, A) and Cerros El Pelado (block 2, B), Talara Basin, northern Peru. A. Internal mold of large left valve (NRM Mo187011), left valve showing anterior adductor muscle scar (arrow). B. NRM Mo187012, external sculpture on right valve. C. Articulated specimen (NRM Mo187013), showing external sculpture on left valve (C1) and lunule and ligament in dorsal view (C2).
Family Vesicomyidae Dall and Simpson, 1901
Genus Pleurophopsis Van Winkle, 1919
Type species: Pleurophopsis unioides Van Winkle, 1919, by monotypy; middle Miocene, Trinidad.
Pleurophopsis lithophagoides Olsson, 1931
Fig. 6.
1931 Pleurophopsis lithophagoides sp. nov.; Olsson 1931: 44, pl. 4: 2, 5, 7, 9.
Material.—Five specimens (NRM Mo187014–187018) and 63 unnumbered, often fragmentary specimens from the Belén seep site, upper Oligocene, Talara Basin, Peru.
Description.—Shell narrow (W/H ratio 0.58–0.66 of the six measured shells), elongate-oval shell (H/L ratio 0.52–0.45 of the five smallest [L = 22–27 mm] specimens, 0.33–0.40 of the five largest [L = 32–40 mm] specimens), umbones slightly elevated, prosogyrate, pointed, positioned anterior at c. 20% total shell length; anterior margin short, convex; posterodorsal margin gently sloping to acutely rounded posterior margin; ventral margin straight or slightly convex; sculpture of irregular growth increments only. Anterior adductor muscle scar kidney-shaped, somewhat impressed; anterior pedal retractor scars small, roundish and connected to adductor scar by fine groove. Narrow sinuous hinge plate; RV with two cardinals, 1 pointing anterior, 3b pointing slightly posterior; LV with three cardinals, 2a pointing antero-ventral, 2b pointing slightly posterior, 4b seen as elongated socket pointing posterior.
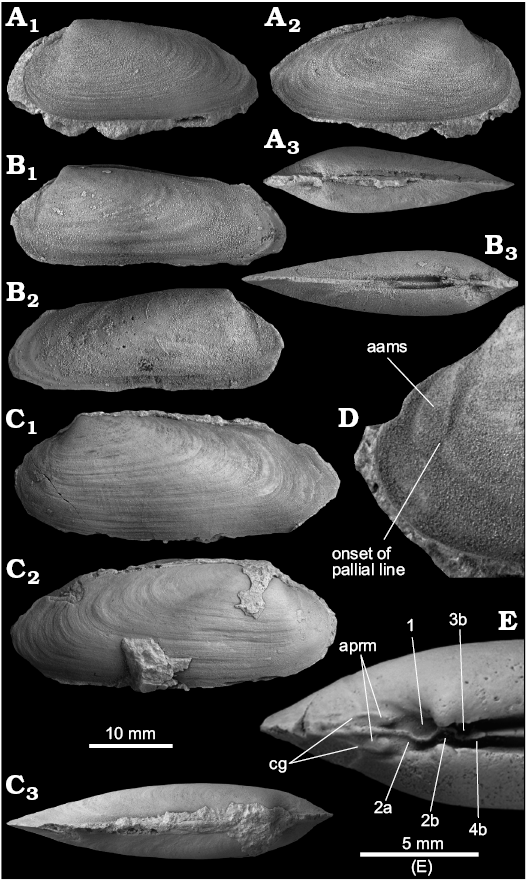
Fig. 6. The vesicomyid Pleurophopsis lithophagoides Olsson, 1931, from the early Oligocene Belén seep site, Talara Basin, northern Peru. A. Specimen with strongly sloping posterodorsal margin (NRM Mo187014), left valve (A1), right valve (A2), and dorsal view (A3). B. Specimen with rather straight posterodorsal margin (NRM Mo187015), left valve (B1), right valve (B2), and dorsal view (B3). C. Large specimen (NRM Mo187016), left valve (C1), right valve (C2), and dorsal view (C3). D. Close-up on anterior side of left valve (NRM Mo187018), showing anterior adductor muscle scar and onset of pallial line. E. Internal mold showing hinge and anterior muscle scars (NRM Mo187017). Abbreviations: aams, anterior adductor muscle scar; aprm, anterior pedal retractor scar; cg, connecting grove between aams and aprs.
Remarks.—All specimens are fragmentary, and the full extent of the hinge plate is unknown. The details on the dentition given here are derived from silicon rubber casts of parts of the hinge and observations on internal molds (Fig. 6E). Pleurophopsis lithophagoides differs from the type species P. unioides by reaching a smaller maximum size (up to 40 mm in length compared to 77 mm for P. unioides) and by being more elongated. The Peruvian Pleurophopsis talarensis sp. nov. described below differs from P. unioides mainly by being much more elongated (H/L ratio 0.26 compared to 0.33–0.4 in large P. lithophagoides). Differences to other species of Pleurophopsis are summarized in Table 1.
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru.
Table 1. Characters of the species of Pleurophopsis Van Winkle, 1919; * minimum values given only, because many Pleurophopsis species show allometric growth (i.e., they increase in length faster than in height). H, height; L, length; W, width.
|
Species |
L |
H |
H/L* |
W/H |
Ventral margin |
Pallial line posterior |
Occurrence |
Stratigraphic range |
References |
|
P. akanudaensis |
71.1 |
32.9 |
0.39 |
0.24–0.4 |
concave |
turns to anterior |
Japan Sea |
middle Miocene |
|
|
P. chinookensis |
90 |
27 |
0.3 |
0.73 |
straight to |
turns posterodorsally |
Washington state, USA |
late Eocene–Oligocene |
|
|
P. chitanii |
70.4 |
21 |
0.3 |
0.7 |
straight to |
turns to anterior |
Japan |
early to early middle Miocene |
|
|
P. hamuroi |
59.2 |
22.3 |
0.38 |
0.64 |
slightly concave |
slight turn to anterior |
Japan Sea |
early to early middle Miocene |
|
|
P. hokkaidoensis |
66.2 |
22.6 |
0.34 |
weakly inflated |
slightly concave |
turns to anterior |
Japan Sea |
early middle Miocene |
|
|
P. kuroiwaensis |
66.7 |
24.6 |
0.34 |
0.6 |
slightly convex |
turns to anterior |
Japan Sea |
latest middle Miocene |
|
|
P. lithophagoides |
40 |
15 |
0.33 |
0.58–0.66 |
straight to slightly convex |
? |
northern Peru |
early Oligocene |
Olsson 1931; and this paper |
|
P. matsumotoi |
112.5 |
39.3 |
0.35 |
0.67–0.73 |
concave |
turns to anterior |
Japanese Pacific coast |
late Oligocene–early Miocene |
|
|
P. talarensis |
37 |
9.5 |
0.26 |
0.61–0.63 |
straight |
? |
northern Peru |
early Oligocene |
this paper |
|
P. uchimuraensis |
120.5 |
27.5 |
0.17 |
0.51–0.64 |
concave |
turns to anterior |
Japan |
early to early middle Miocene |
|
|
P. unioides |
77 |
33 |
0.43 |
0.55 |
straight to |
slight turn to anterior |
Trinidad |
middle Miocene |
Pleurophopsis talarensis sp. nov.
Fig. 7.
Zoobank LCID: urn:lsid:zoobank.org:act:E668EC27-89CE-4B98-A2D0-B7A6FE856EA8
Etymology: For the Talara Basin.
Type material.—Holotype, NRM Mo187019 from block 6. Paratypes NRM Mo187022, NRM Mo187024–26 from block 6, NRM Mo187023 from block 7, and NRM Mo187020 and NRM Mo187021 from block 9.
Type locality: The Cerro La Salina seep deposits, Talara Basin, Peru.
Type horizon: Presumably the early Oligocene part of the Heath shale.
Material.—The type material and further, unnumbered fragmentary specimens from type locality and horizon, blocks 1, 2, 6–9.
Dimensions (in mm).—NRM Mo187019, L = 37, H = 9.5, W (single valve) = 3; NRM Mo187020, L = 33 (incomplete), H = 10.5, W = 6.5; NRM Mo187021, L = 36 (incomplete) and H = 10.
Diagnosis.—Small and very elongate Pleurophopsis (H/L ratio c. 0.26) with slightly curved shell with subparallel dorsal and ventral margins.
Description.—Shell small, very elongate (H/L ratio c. 0.26), and little inflated (W/H ratio c. 0.61–0.63), umbo anterior at c. 16% shell length, slightly elevated; dorsal and ventral margin subparallel, dorsal margin slightly convex, ventral margin slightly concave; no lunule or lunular incision; ligament external, about 1/3 of shell length; sculpture of irregular growth increments. Anterior adductor muscle scar subcircular, deeply impressed, pallial line starting at posteroventral corner; hinge plate narrow, hinge teeth radiating, RV with two strong cardinals, cardinal 1 short, pointing downward and somewhat anterior, cardinal 3b blunt, pointing posteroventrally; LV with three teeth, cardinal 2a very elongate, subparallel to anterodorsal margin, 2b blunt, broad, pointing downward, cardinal 4b short and thin; subumbonal pit large, elongate and deep in both valves.
Remarks.—The differences from Pleurophopsis unioides and P. lithophagoides have been outlined above. The early Miocene Japanese vesicomyid “Adulomya” uchimuraensis Kuroda, 1931, has a H/L ratio of c. 0.22 and is thus even more elongate than P. talarensis, and is also much larger (Kuroda 1931; Kanno et al. 1998; Amano and Kiel 2011). Differences to other species of Pleurophopsis are summarized in Table 1.
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru.

Fig. 7. The vesicomyid Pleurophopsis talarensis sp. nov., from early Oligocene seep deposits at Cerro La Salina (block 6, A, D, F–H; block 7, E; block 9, B, C), Talara Basin, northern Peru. A. Holotype (NRM Mo187019), right valve showing shell outline. B. Paratype (NRM Mo187022), posteriorly damaged, articulated specimen; view on dorsal side (B1) and on right valve (B2). C. Paratype (NRM Mo187020), right valve with posterior tip missing. D. Paratype (NRM Mo187023), internal mold of left valve showing anterior adductor muscle scar and pallial line (D1), arrow indicates onset of pallial; close-up on hinge area (D2. E. Paratype (NRM Mo187021), semi-articulated specimen, view on incomplete right valve). F. Paratype (NRM Mo187024), anterior part of fight valve. G. Paratype (NRM Mo187026), anterior part of right valve (G1), view on hinge area (G2). H. Paratype (NRM Mo187025), hinge area of right valve.
Genus Vesicomya Dall, 1886
Type species: Callocardia atlantica Smith, 1885, by original designation; Recent, North Atlantic Ocean.
“Vesicomya” tschudi Olsson, 1931
Fig. 8.
1931 Vesicomya tschudi sp. nov.; Olsson 1931: 54, pl. 4: 6, 8.
Material.—One small, articulated specimen (NRM Mo 187029) from Cerro La Salina block 1; one medium-sized, articulated specimen (NRM Mo187030) from Cerro La Salina block 5; one large LV (NRM Mo187031) from Cerro La Salina block 5. All upper Oligocene, Talara Basin, Peru.
Dimensions (in mm).—NRM Mo187029, L = 20, H = 14, W = 10.5; NRM Mo187030, L = 41.5, H = 27, W = 19.5; NRM Mo187031, L = 59, H = 37.5, W (single valve) = 14.
Description.—Well-inflated, oval shell with large, prosogyrate and strongly protruding umbones; post-umbonal side of shell gently tapering into a slightly truncate posterior margin; lunular incision present, no escutcheon, but posterodorsal area marked by distinct ridge; external sculpture of indistinct, irregular, commarginal growth lines; anterior adductor muscle scar bean-shaped, small, and moderately impressed; posterior adductor scar indistinct, round, of moderate size; pallial line close to ventral margin, no pallial sinus.

Fig. 8. The vesicomyids “Vesicomya” tschudi Olsson, 1931 (A–C, E) and “Vesicomya” ramondi Olsson, 1931 (D), from early Oligocene seep deposits in the Talara Basin, northern Peru. A. Small specimen (NRM Mo187029) from Cerro La Salina block 1, view on left valve (A1), dorsal view (A2), view on anterodorsal side showing lunular incision (A3, arrow). B. Medium-sized specimen (NRM Mo187030) from Cerro La Salina block 5, view on left valve (B1) and dorsal view (B2). C. Holotype (PRI 1965) from Pajarabobo, view on left valve showing posterior end of pallial line (arrow). D. Holotype (PRI 1962) from Pajarabobo, view on right valve. E. Large left valve (NRM Mo187031) from Cerro La Salina block 5, view of the outer side.
Remarks.—Along with Vesicomya tschudi, Olsson (1931) also introduced Vesicomya ramondi Olsson, 1931, from the Heath shale around Belén and Pajarabobo, but he did not comment on the differences between the two species. Squires and Gring (1996) noted that V. ramondi has a more elongated shell, as can be seen when comparing the holotypes of the two species (see Fig. 8C, D). Squires and Gring (1996) assigned Eocene specimens from seep deposits of the Wagonwheel Formation in California, USA, to V. tschudi, based on their overall shape, although the specimens showed no hinge dentition, the lunule was imperfectly preserved, and most specimens were smaller and had lower umbones than the Peruvian specimens.
Middle Eocene vesicomyid specimens from the Humptulips Formation in Washington state, USA, were identified as Archivesica cf. tschudi by Amano and Kiel (2007). Later, the same authors changed their assignment of these specimens from Archivesica to Pliocardia or a new genus (Amano and Kiel 2012). The middle Miocene Pliocardia? tanakai Miyajima, Nobuhara, and Koike, 2017, from central Japan (Miyajima et al. 2017) is similar to “V.” ramondi in terms of its outline, elongation, and absence of a pallial sinus, but is smaller, less inflated, and has smaller, more pointed, and less prosogyrate umbones than “V.” ramondi. Pliocardia? tanakai is much smaller and also more elongated than “Vesicomya” tschudi.
“Vesicomya” tschudi cannot be confidently placed in Pliocardia because it lacks a pallial sinus, a feature that is present in the type species of Pliocardia, P. bowdeniana Dall, 1903 (Krylova and Janssen 2006; personal observation SK). The presence or absence of a pallial sinus appears to be a character that distinguishes genera among the Vesicomyidae (Johnson et al. 2017). Furthermore, the genus Pliocardia Woodring, 1925 itself is in need of a taxonomic revision because the species presently assigned to it belong to at least two or three different clades (Decker et al. 2012; Valdés et al. 2013; Johnson et al. 2017). Hence the generic position of “Vesicomya” tschudi remains unresolved.
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru.
Class Gastropoda Cuvier, 1795
Limpets of uncertain affinity
Limpet indet. 1
Fig. 9A.
Material.—One specimen (NRM Mo187032) from Cerro la Salina block 9, upper Oligocene, Talara Basin, Peru.
Description.—Tall, oval shell, broader anteriorly and more pointed posteriorly, and slightly wider than high; apex in central position; anterior slope straight, posterior slope slightly convex; shell sculptured by fine, broad, and flat concentric ribs; length 5.3 mm.
Remarks.—Tall, conical shells like this are found among limpet of very disparate taxonomic affinities, including the Recent lepetellid Lepetella ionica Nordsieck, 1973, as illustrated in Dantart and Luque (1994), and the fissurellid Cornisepta antarctica (Egorova, 1982), as illustrated in (McLean and Geiger 1998). The corroded protoconch and the lack of knowledge on the shell microstructure preclude a more precise taxonomic identification.
Limpet indet. 2
Fig. 9B.
Material.—One specimen (NRM Mo187033) from Cerro la Salina block 6, upper Oligocene, Talara Basin, Peru.
Description.—Tall, narrow, elongate-oval limpet shell with pointed anterior and posterior ends and apex at about anterior third of shell; anterior slope straight, posterior margin gently convex; internal surface smooth except for faint growth increments.
Remarks.—Similar laterally compressed shells with anteriorly situated apex can be found among the pseudococculinid (Vetigastropoda) genera Pseudococculina Schepman, 1908, and Tentaoculus Moskalev, 1976 (Marshall 1986; McLean and Harasewych 1995), as well as among the pectinodontid (Patellogastropoda) genus Serradonta Dall, 1882 (Okutani et al. 1992; Jenkins et al. 2007).

Fig. 9. Limpet gastropods from early Oligocene seep deposits at Cerro La Salina (block 6, B, C; block 9, A) Talara Basin, northern Peru. A. Limpet indet. 1 (NRM Mo187032) in lateral view (A1), apical view (A2) and seen from anterior view (A3). B. Limpet indet. 2 (NRM Mo187033) in apical (B1) and lateral (B2) views. C. The cocculinid Coccopigya sp. (NRM Mo187034) in apical (C1) and lateral (C2) views.
Subclass Neomphaliones Bouchet, Rocroi, Hausdorf, Kaim, Kano, Nützel, Parkhaev, Schrödl, and Strong 2017
Order Cocculinida Haszprunar, 1987
Family Cocculinidae Dall, 1882
Genus Coccopigya Marshall, 1986
Type species: Cocculina spinigera Jeffreys, 1883, by typification of replaced name; Recent, North Atlantic Ocean.
Coccopigya sp.
Fig. 9C.
Material.—One specimen (NRM Mo187034) from Cerro la Salina block 6, upper Oligocene, Talara Basin, Peru.
Description.—Elongate-oval, moderately tall limpet shell with apex at about anterior third of shell; sculpture of indistinct but broad radial ribs, about 30 on each side of shell, more densely spaced at anterior end; fine radial growth increments visible.
Remarks.—With its narrow shell and anteriorly displaced apex, this specimen is remarkably similar to the early or middle Miocene Coccopigya compunctum (Marwick, 1931) from New Zealand (Marwick 1931; Marshall 1986).
Order Neomphalina McLean, 1981
Family Neomphalidae McLean, 1981
Genus Retiskenea Warén and Bouchet, 2001
Type species: Retiskenea diploura Warén and Bouchet, 2001, by original designation; Recent, North Pacific Ocean.
Retiskenea? sp.
Fig. 10.
Material.—One specimen (PRI 80014) from the Lomitos cherts, numerous specimens (unnumbered specimens housed at NRM) from blocks of the Cerro La Salina seep deposits. All upper Oligocene, Talara Basin, Peru.
Description.—Shell globular, 2.5 evenly convex whorls with fine, dense prosocyrt growth lines; fine spiral lines present on early whorls; spire low, aperture large.

Fig. 10. The possible neomphalid Retiskenea? sp. (PRI 80014) from the early Cenozoic Lomitos cherts seep deposits near Negritos, Talara Basin, northern Peru.
Remarks.—This species is represented by poorly preserved, small specimens only. The illustrated specimen from the Lomitos cherts shows a granular pattern on its protoconch, somewhat similar to that seen on other fossil Retiskenea species (Kiel 2006; Campbell et al. 2008; Kaim et al. 2014), but this feature might just be a preservational artifact. Hence, our assignment to Retiskenea remains tentative; they might as well belong to the Skeneidae or some other vetigastropod group.
The Eocene–Oligocene Retiskenea statura (Goedert and Benham, 1999) from seep deposits in western Washington has a taller spire than the Peruvian Retiskenea sp.? (Goedert and Benham 1999; Kiel 2006) and the two species R. kieli Campbell, Peterson, and Alfaro, 2008, and R. tuberculata Campbell, Peterson, and Alfaro, 2008, from Cretaceous seep deposits in California, USA, have a more rapidly expanding whorl profile than the Peruvian Retiskenea sp.? (Campbell et al. 2008; Kaim et al. 2014).
Subclass Vetigastropoda Salwini-Plawen, 1980
Family Pyropeltidae McLean and Haszprunar, 1987
Genus Pyropelta McLean and Haszprunar, 1987
Type species: Pyropelta musaica McLean and Haszprunar, 1987, by original designation; Recent, Juan de Fuca Ridge, NE Pacific Ocean.
Pyropelta seca sp. nov.
Fig. 11.
Zoobank LCID: urn:lsid:zoobank.org:act:005AF820-5BD6-42C1-9AE6-F1F03A3EC479
Etymology: For Quebrada Seca river valley.
Type material: Holotype: NRM Mo187036 from Cerro La Salina block 6. Paratypes: NRM Mo187035, 187037–187039 from block 6.
Type locality: Cerro La Salina block 6, Talara Basin, Peru.
Type horizon: Early Oligocene part of the Heath shale.
Material.—The type material and a few further unnumbered specimens from Cerro La Salina blocks 6 and 9, and one unnumbered specimen from Cerros El Pelado block 2. All upper Oligocene, Talara Basin, Peru.
Dimensions (in mm).—The largest specimen, L = 2.5, H = 1.4.
Diagnosis.—Tall, oval to elongate-oval shell; apex with a slight forward inclination; shell with cone-in-cone layering, surface smooth; base flat to concave.
Description.—Small limpet shell, tall, posterior slope gently and irregularly convex, anterior slope irregular, more-or-less straight; base oval to elongate-oval, flat or convex with lateral margins lower than anterior and posterior margins; apex in central or very slightly anterior position, slightly inclined towards anterior; shell surface smooth, with cone-in-cone layering.
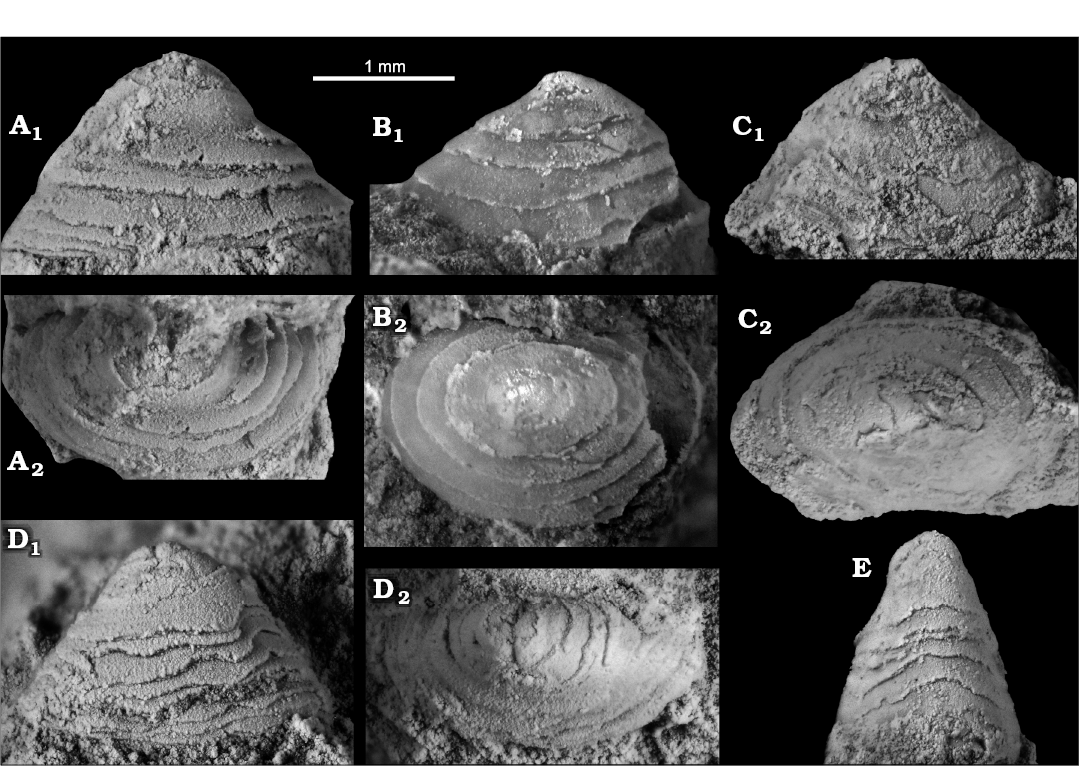
Fig. 11. The vetigastropod Pyropelta seca sp. nov. from early Oligocene seep deposits at Cerro La Salina block 6, Talara Basin, northern Peru. A. Paratype (NRM Mo187035) in lateral (A1) and apical (A2) views. B. Holotype (NRM Mo187036) in lateral (B1) and apical (B2) views. C. Paratype (NRM Mo187037) in lateral (C1) and apical (C2) views. D. Paratype (NRM Mo187038) in lateral (D1) and apical (D2) views. E. Paratype (NRM Mo187039) in anterior view.
Remarks.—Although the shell microstructure and protoconch are unknown, this species is placed in Pyropelta based on its cone-in-cone type layered shell. This type of layering is regularly seen in Pyropelta (McLean and Haszprunar 1987; McLean 1992; Sasaki et al. 2003, 2008; Zhang and Zhang 2017) but not in other cocculiniform genera, and results from alternating layers with crossed lamellar or crossed acicular structure and prismatic structure (Kiel 2004).
Compared to Pyropelta seca, Pyropelta elongata Zhang and Zhang, 2017, from methane seeps in the South China Sea is similarly elongated, but not as tall (Zhang and Zhang 2017). Pyropelta ryukyuensis Sasaki, Okutani, and Fujikura, 2008, from hydrothermal vents in the Okinawa Trough (Sasaki et al. 2008) is broader and less tall than Pyropelta seca. Even broader and lower are Pyropelta musaica and Pyropelta craigsmithi McLean, 1992; Pyropelta corymba McLean and Haszprunar, 1987, is also broader but can be as tall as Pyropelta seca (McLean and Haszprunar 1987; McLean 1992).
Stratigraphic and geographic range.—Early Oligocene of the Talara Basin, northern Peru.
Family Colloniidae Cossmann in Cossmann and Peyrot, 1917
Genus Cantrainea Jeffreys, 1883
Type species: Turbo peloritanus Cantraine, 1835, by monotypy; Recent, Mediterranean Sea.
Cantrainea sp.
Fig. 12A, B.
Material.—One specimen (NRM Mo187040) from Cerro La Salina block 1; one (NRM Mo187041) from Cerro La Salina block 9; a single poorly preserved specimen possibly also belonging to Cantrainea sp. is present on the same sample as Provanna pelada (NRM Mo187054) from Cerros El Pelado block 2. All upper Oligocene, Talara Basin, Peru.
Description.—Small, trochiform shell with angular whorl profile and moderately wide umbilicus; subsutural ramp slightly convex and only slightly sloping, smooth except for strongly prosocline growth lines, margin to whorl’s flank marked by fine groove and smooth keel; flank vertical, smooth except for fine axial growth lines; basal margin also marked by smooth keel; base smooth with inclination of about 45°; umbilicus occupying about 20 of base’s width.
Remarks.—Most similar is Cantrainea macleani Warén and Bouchet, 1993 from Recent seeps in the Gulf of Mexico, which differs by its more conical shell, resulting from a steeper subsutural ramp that is concave rather than convex as in the specimen reported here. Indeed, most fossil and Recent species of Cantrainea have a steeper subsutural ramp than the Peruvian specimens illustrated here (Marshall 1979; Vilvens 2001; Okutani 2001; Kaim et al. 2009).
Family uncertain
Trochoid incertae sedis
Fig. 12C, D.
Material.—One specimen (NRM Mo187043) from Cerro La Salina block 1, one specimen (NRM Mo187042, H = 2.6 mm) from Cerro La Salina block 2. Both upper Oligocene, Talara Basin, Peru.
Description.—Fine, subsutural row of tubercles with fine, prosocline riblets underneath; apparently two keels on whorl’s flank, and two (three) nodular spirals on base.
Remarks.—These specimens are difficult to place among the Vetigastropoda. The sculpture is similar on both specimens, but specimen NRM Mo187042 has straight sides similar to some solariellids calliotropids, or calliostomatids (Hickman and McLean 1990), whereas specimen NRM Mo187042 with its more convex whorl profile resembles certain margaritids, and also the solariellid Solariella (Pupillaria) columbiana Squires and Goedert, 1991, from late Eocene seep deposits in Washington state, USA (Squires and Goedert 1991).
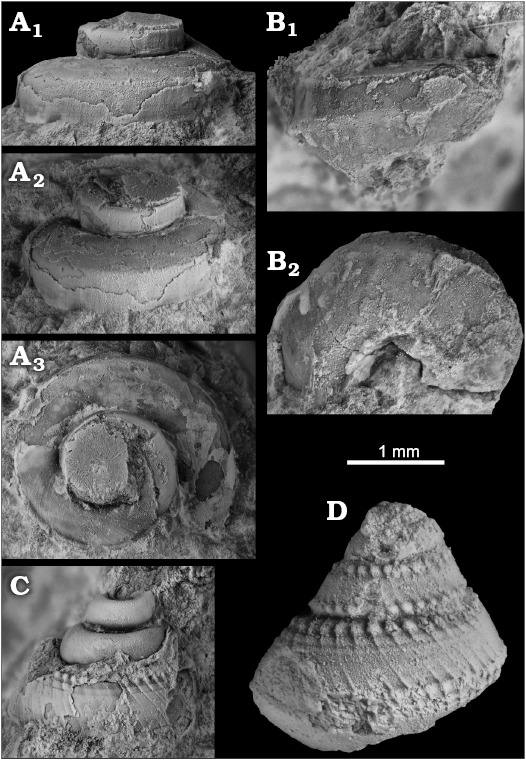
Fig. 12. Vetigastropoda from early Oligocene seep deposits at Cerro La Salina (block 1, A, D; block 2, C; block 9, B) Talara Basin, northern Peru. A, B. The colloniid Cantrainea sp. A. NRM Mo187040, specimen with base embedded in rock matrix in lateral (A1), oblique (A2) and apical (A3) views. B. NRM Mo187041, specimen with exposed base in lateral (B1) and basal (B2) views. C, D. The trochoid incertae sedis. C. NRM Mo187042. D. NRM Mo187043.
Subclass Neritimorpha Golikov and Starobogatov, 1975
Neritimorpha indet.
Fig. 13.
Material.—One specimen (PRI 80015) that is 8.5 mm across, from the Lomitos cherts, upper Oligocene, Talara Basin, Peru.
Remarks.—Olsson (1931) reported “Nerita” from the deposits near Belén and Pajarabobo and from the Lomitos cherts, but he never illustrated them. We did not find any neritids in the field, but a single specimen was found on the back side of the rock containing the holotype of Solemya lomitensis, from the Lomitos cherts. The specimen has a typical neritid shape but because neither the external surface nor the aperture is preserved, we cannot determine this specimen any further.

Fig. 13. Neritimorpha indet. (PRI 80015) from the early Cenozoic Lomitos cherts seep deposits near Negritos, Talara Basin, northern Peru.
Subclass Caenogastropoda Cox, 1960
Family Provannidae Warén and Ponder, 1991
Genus Provanna Dall, 1918
Type species: Trichotropis lomana Dall, 1918, by monotypy; Recent, NE Pacific Ocean.
Provanna antiqua Squires, 1995
Fig. 14.
1995 Provanna n. sp.; Goedert and Campbell 1995: figs. 4–7.
1995 Provanna antiqua sp. nov.; Squires 1995: 32, figs. 3–18.
2006 Provanna antiqua Squires 1995; Kiel 2006: 125, figs. 5.1–5.8.
2006 Provanna antiqua Squires 1995; Kiel and Goedert 2006a: fig. 2C.
Material.—Eight specimens (NRM Mo187044–187051) and numerous unnumbered specimens from Cerro La Salina blocks 1, 4, 6, 8, 9; one unnumbered specimen from the Belén site. All upper Oligocene, Talara Basin, Peru.
Remarks.—We cannot see any differences between the Peruvian specimens from the La Salina sites and those described from late Eocene to Oligocene seep deposits in western Washington, USA (Goedert and Campbell 1995; Squires 1995; Peckmann et al. 2002). The extant West African P. chevallieri Warén and Bouchet, 2009, is also virtually indistinguishable.
Stratigraphic and geographic range.—Late Eocene to Oligocene of western Washington state, USA; early Oligocene of the Talara Basin, northern Peru.

Fig. 14. The provannid gastropod Provanna antiqua Squires, 1995, from early Oligocene seep deposits at Cerro La Salina (block 1, A, F; block 4, H; block 6, B, C, D, G; block 8, E), Talara Basin, northern Peru. A. NRM Mo187044, specimen with distinctive axial and spiral sculpture, in abapertural view. B. NRM Mo187045, specimen with distinctive sculpture and showing the basal groove, in apertural view. C. NRM Mo187046, specimen with weak axial sculpture in the upper whorl, in apertural view. D. NRM Mo187047, fragmentary specimen with mainly spiral sculpture, in apertural view. E. NRM Mo187048, nearly smooth specimen showing slightly sinuous growth lines, in apertural view. F. NRM Mo187049, specimen with small shoulder and sculpture mainly in upper part of whorls, in apertural view. G. NRM Mo187050, specimen with faint axial and spiral sculpture, in apertural view. H. NRM Mo187051, two specimens with small shoulder and sculpture mainly in upper part of whorls..
Provanna pelada sp. nov.
Fig. 15.
Zoobank LCID: urn:lsid:zoobank.org:act:A2B18082-B13C-40BF-88DE-6C36E6AFC742
Etymology: For the type locality at Cerros El Pelado.
Type material: Holotype: NRM Mo187052. Paratypes: NRM Mo187053, NRM Mo187058.
Type locality: Cerros El Pelado block 2, Talara Basin, Peru.
Type horizon: The presumably early Oligocene part of the Heath shale.
Material.—The type material and several unnumbered specimens from the type locality.
Dimensions.—The largest specimen, H = c. 8 mm.
Diagnosis.—Slender provannid with at least two whorls and incised suture; whorls with basal constriction; shell surface nearly smooth, or with faint spiral threads or indistinct spiral cords, axial ornament consists either of fine or rough, prosocline, growth increments or indistinct prosocline ribs.
Description.—Shell small, slender, at least two whorls, suture deeply incised; whorl profile evenly convex or with slight subsutural constriction; base with constriction and bulge underneath; surface sculpture of fine, prosocline growth increments, sometimes also irregular, rough and irregular, low prosocline ribs, spiral sculpture ranging from absent to indistinct, low cords.

Fig. 15. The provannid gastropod Provanna pelada sp. nov. from the early Oligocene seep deposit at Cerros El Pelado block 3, Talara Basin, northern Peru. A. Holotype (NRM Mo187052), smooth specimen in apertural (A1) and abapertural (A2) views. B. Paratype (NRM Mo187053), specimen with faint spiral ornament, in apertural view. C. Paratype (NRM Mo187054), specimen with axial and spiral sculpture, in two lateral views (C1, C2). D. Paratype (NRM Mo187055), nearly smooth specimen with faint spirals. E. Paratype (NRM Mo187056), showing distinctive prosocline growth increments and basal constriction. F. Paratype (NRM Mo187057), smooth specimen. G. Paratype (NRM Mo187058), specimen with rough, irregular prosocline ribs and fine spiral sculpture. All in lateral view, unless otherwise indicated.
Remarks.—Compared to smooth (or nearly smooth) specimens of P. antiqua, Provanna pelada is slightly taller, its whorls are less convex, and the suture is less distinct. Also, its growth lines are prosocline, whereas those of P. antiqua are opisthocline or straight. The early Oligocene P. urahoroensis Amano and Jenkins, 2013, from a seep deposit in eastern Hokkaido, Japan, has a similar range of ornamentation, but differs from P. pelada by having lower whorls with a more convex profile, and by lacking the constriction and bulge at the base (Amano and Jenkins 2013). The two Miocene species Provanna marshalli Saether, Little, and Campbell, 2010a (from New Zealand) and Provanna hirokoae Amano and Little, 2014 (from Japan) differ from Provanna pelada by having a broader apical angle and by having both spiral and axial sculpture (Saether et al. 2010a; Amano and Little 2014). There are three smooth Recent species; among them, Provanna glabra Okutani, Fujikura, and Sasaki, 1993 is taller, P. subglabra Sasaki, Ogura, Watanabe, and Fujikura, 2016 has slightly less convex whorls, and P. laevis (Warén and Ponder 1991) has slightly more convex whorls. Provanna fortis Hybertsen and Kiel, 2018, from the middle Eocene Humptulips Formation in western Washington state, USA (Hybertsen and Kiel 2018), has stronger ornamentation and a much more angular whorl profile compared to that of Provanna pelada.
Stratigraphic and geographic range.—Type locality and horizon only.
Family Hokkaidoconchidae Kaim, Jenkins, and Warén, 2008
Genus Ascheria Kaim, Jenkins, Tanabe, and Kiel, 2014
Type species: Abyssochrysos? giganteum Kiel, Campbell, and Gaillard 2010, by original designation; Late Jurassic, California, USA.
Remarks.—Olsson (1931) introduced the genus Anconia for a large, high-spired gastropod from Punta Ancon (or Ancon Point) at Anconcito in southern Ecuador, Anconia elenensis Olsson, 1931. This species resembles members of Ascheria in every aspect (Kaim et al. 2014). Because Anconia Olsson, 1931, is a junior homonym of the grasshopper Anconia Scudder, 1876, we assign Anconia elenensis to Ascheria. Olsson (1931: 85) considered the strata at Punta Ancon as “equivalent of the Punta Bravo grits of Peru and therefore of middle Oligocene age”. Subsequent work on the radiolarians of these strata indicates a middle Eocene age instead (Ordóñez 1995). Hence, Ascheria elenensis is of roughly the same age as the large abyssochrysoid Humptulipsia raui (Goedert and Kaler, 1996) from seep deposits in the middle Eocene Humptulips Formation in Washington state, USA, and probably slightly older than the Ascheria? sp. specimens reported from seep deposits in Barbados (Kaim et al. 2014).
Ascheria salina sp. nov.
Fig. 16.
Zoobank LCID: urn:lsid:zoobank.org:act:6BDCBE13-9DA3-4AF0-9366-20014BEA1241
Etymology: For the type locality at Cerro La Salina.
Type material: Holotype: NRM Mo187061. Paratypes: NRM Mo 187059 from Cerro La Salina block 2 and NRM Mo187060 from Cerro La Salina block 1.
Type locality: Cerro La Salina block 1, Talara Basin, Peru.
Type horizon: Presumably the early Oligocene part of the Heath shale.
Material.—The type material and a few unnumbered fragments from Cerro La Salina block 1.
Dimensions.—Holotype (NRM Mo187061, H = 22 mm, W = 9 mm) with missing early whorls; fragmentary paratype (NRM Mo187060) W = 11.5 mm.
Diagnosis.—Small-sized Ascheria with blunt axial ribs that are opisthocline in lower part of whorls, a weak subsutural incision, and fine spiral incision in early whorls.
Description.—Medium-sized, high-spired cerithiform shell, at least seven slightly convex whorls. Sculpture consists of blunt axial ribs that are straight or slightly oblique in upper half of whorls and become increasingly opisthocline in lower half; ribs have subsutural constriction and blunt tubercles may develop above and below that constriction; early whorls have fine spiral incision but these disappear on whorls wider than 3 mm, later whorls with faint spiral threads; base is smooth except for opisthocyrt growth lines. Aperture appears to be elongate-oval.
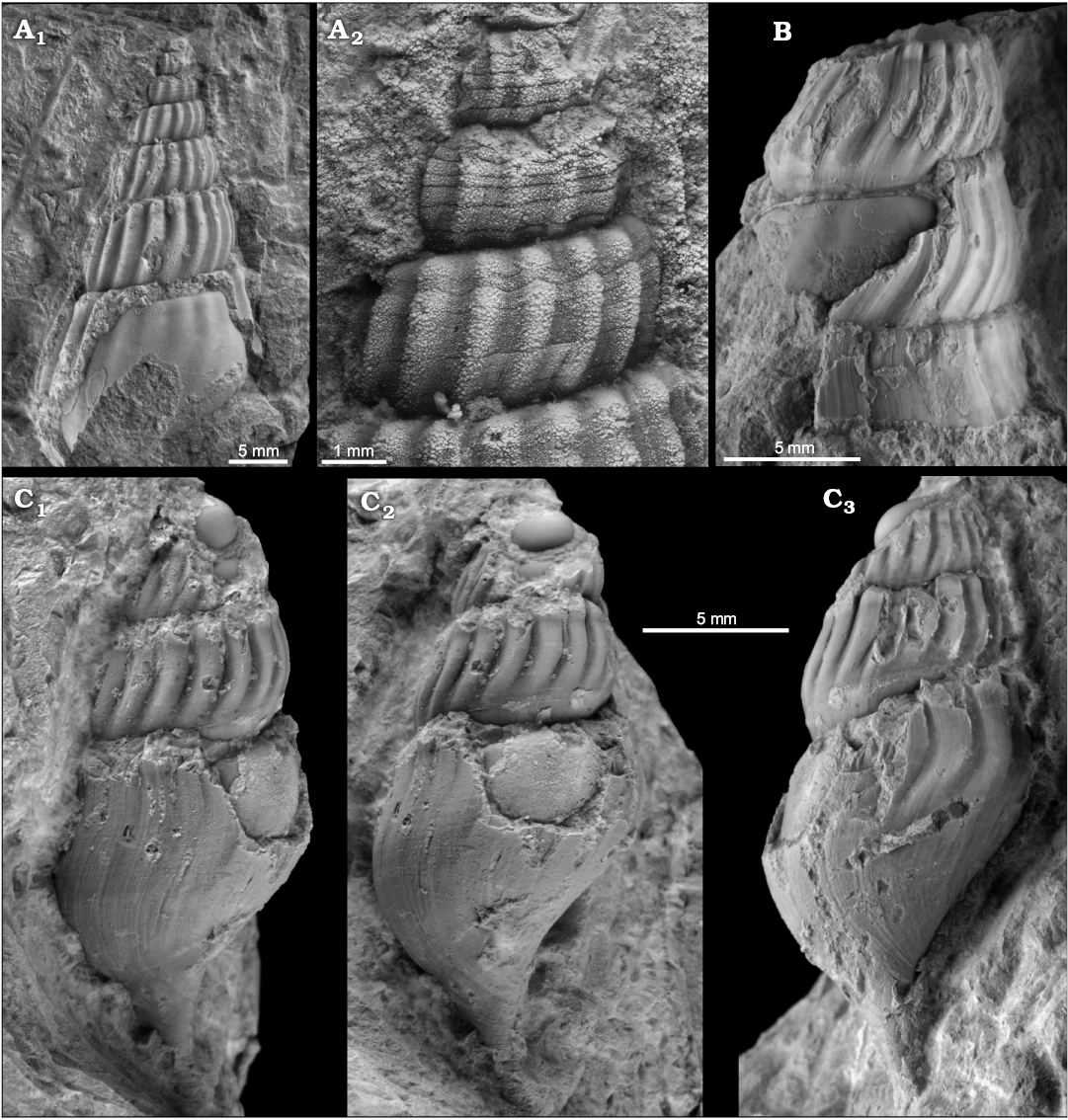
Fig. 16. The hokkaidoconchid gastropod Ascheria salina sp. nov. from early Oligocene seep deposits at Cerro La Salina (block 1, B, C; block 2, A) Talara Basin, northern Peru. A. Paratype (NRM Mo187059) showing fine spiral sculpture on early whorls (A2, close-up of the apical part). B. Paratype (NRM Mo187060) large fragmentary specimen. C. Holotype (NRM Mo187061) showing opisthocline axial ornament and faint spirals on last whorl (C1–C3, different lateral views). All in lateral view.
Remarks.—Ascheria salina sp. nov. differs from A. elenensis by the shape of the axial ornament, which is strongly opisthocline just below the suture in A. elenensis, whereas the ribs become opisthocline only in the lower half of the whorl in A. salina. Furthermore, A. elenensis grows to a much larger size and appears to be higher spired (having a narrower apical angle) and A. salina has a higher whorl profile than A. elenensis. The Late Jurassic type species A. gigantea differs from A. salina by having spiral ornament also on large whorls, whereas in A. salina such spirals are only present on very early whorls (Kaim et al. 2014). Ascheria? sp. from Barbados (including the specimens reported as “cerithid sp.” by Kugler et al. (1984: pl. 7: 6–8), and as Abyssochrysos sp. by Gill et al. (2005: fig. 5D) has more oblique or sloping axial ribs and spiral ornament is also present on large specimens, in contrast to A. salina. As Ascheria? sp. from Barbados is most likely of Eocene age (Kiel and Hansen 2015), the early Oligocene Peruvian Ascheria salina is currently the youngest member of the genus; it is also the smallest species of Ascheria.
Stratigraphic and geographic range.—Type locality and horizon only.
Family Buccinidae Rafinesque, 1815
Genus Colus Röding, 1798
Type species: Murex islandicus Mohr, 1786, by subsequent designation; Recent, North Atlantic Ocean.
Colus sekiuensis Kiel and Goedert, 2007
Fig. 17A, B.
2007 Colus sekiuensis sp. nov.; Kiel and Goedert 2007: 43, figs. 3D, E.
Material.—One specimen (NRM Mo187062; H = 21 mm) from Cerro La Salina block 2, one specimen (NRM Mo187063; H = 7 mm) from Cerro La Salina block 4. Both upper Oligocene, Talara Basin, Peru.

Fig. 17. Neogastropod and opisthobranch gastropods from early Oligocene seep deposits at Cerro La Salina (block 2, A; block 4, B; block 6, E; block 7, D) and Belén seep site (C), Talara Basin, northern Peru. A, B. The buccinid Colus sekiuensis Kiel and Goedert, 2007. A. Large specimen (NRM Mo187062), showing shape of last whorl. B. NRM Mo187063, fragment of an early whorl. C. Buccinidae indet. (NRM Mo187064), fragmentary specimen. D. NRM Mo187065, the opisthobranch “Acteon” sp. in apertural (D1) and lateral (D2) views. E. The opisthobranch Cylichna sp. (NRM Mo187066).
Remarks.—The specimens are rather imperfectly preserved; with their evenly convex whorl profile and spiral sculpture consisting of 11 low, nearly flat-topped, beaded spiral cords, the specimens are identical to those of Colus sekiuensis known from whale and wood falls from early Oligocene strata in Washington state, USA (Kiel and Goedert 2007). Cyrtochetus? chiraensis Olsson, 1931 from the Chira Formation in the Talara Basin, northern Peru, which is stratigraphically well below the Heath shale and now considered to be late Eocene in age (Higley 2004), is remarkably similar. That species differs from Colus sekiuensis by having finer and more numerous spiral cords with a more convex profile compared to the nearly flat-topped spiral of Colus sekiuensis. In general, the buccinid genus Cyrtochetus differs from Colus by having a varix at the aperture and denticles on the inner lip (Garvie 2013); however, those features cannot be recognized in the available material.
Olsson (1931) reported two species of Siphonalia? from Belén and Pajarabobo, S.? belenensis and S.? tessaria. Both differ from Colus sekiuensis by their more angular whorl profile. At least judging from the published drawing, Siphonalia noachina (Sowerby, 1846) as illustrated by Ortmann (1902: pl. 34: 5) from the late Oligocene to early Miocene of Patagonia, Argentina, has a similar overall sculpture, but a more convex whorl profile and fewer spirals per whorl.
In his faunal list for the limestone rocks near Belén and Pajarabobo, Olsson (1931: 22) lists “Austrofusus? belenensis n. sp.”, but a species of this name is neither described nor figured in his report. He also did not figure any neogastropods other than S.? belenensis and S.? tessaria from the Heath shale. Hence, Austrofusus? belenensis is a nomen dubium.
Stratigraphic and geographic range.—Oligocene to early Miocene of western Washington state, USA; early Oligocene of the Talara Basin, northern Peru.
Buccinidae indet.
Fig. 17C.
Material.—One specimen (NRM Mo187064, H = 7 mm) from the Belén seep site, upper Oligocene, Talara Basin, Peru.
Remarks.—The specimen differs from Colus sekiuensis by its distinctive opisthocline axial ribs and its seemingly more convex whorl profile. Among the buccinids reported by Olsson (1931) from northern Peru, Cyrtochetus? chiraensis Olsson, 1931, has an overall similar shape, but lacks distinctive axial ornament. Olsson’s Siphonalia species discussed above in the context of Colus sekiuensis differ from this species by their more angular whorl profile.
Subclass Heterobranchia Burmeister, 1837
Family Acteonidae d’Orbigny, 1843
Genus Acteon Montfort, 1810
Type species: Bulla tornatilis Linnaeus, 1758, by original designation; Recent, North Atlantic Ocean.
“Acteon” sp.
Fig. 17D.
Material.—One specimen (NRM Mo187065, H = 3.3 mm) from Cerro La Salina block 7, upper Oligocene, Talara Basin, Peru.
Remarks.—The single available specimen is 3.3 mm high, has a distinct shoulder, and is sculptured by numerous fine, beaded spiral cords. A similar species is Acteon annectens Meyer, 1885, from the late Eocene Moodys Branch Formation in Mississippi, USA (Dockery 1977). Acteonids have also been reported from Late Cretaceous seep deposits in Japan (Kaim et al. 2009) and Oligocene seep deposits in Washington, USA (Kiel 2006). The generic name is given in inverted commas here because genera among the Acteonidae are mainly distinguished based on anatomy and radula rather than on conchological characters (Rudman 1971).
Family Cylichnidae Adams and Adams, 1854
Genus Cylichna Lovén, 1846
Type species: Bulla cylindracea Pennant, 1777, by subsequent designation; Recent, North Atlantic Ocean.
Cylichna sp.
Fig. 17E.
Material.—One specimen (NRM Mo187066, H = 5 mm) from Cerro La Salina block 1, one unnumbered specimen from Cerro La Salina block 6. Both upper Oligocene, Talara Basin, Peru.
Remarks.—The two available specimens are 5 mm high, have an evenly convex whorl profile, and show numerous fine spiral incisions. Olsson (1931) rarely reported specimens smaller than a centimeter and, hence, such small opisthobranchs are virtually unknown from the Peruvian fossil record. A similar species is Cylichna atahualpa (Dall, 1908) from the Pacific coast of Costa Rica (Valdés and Camacho-García 2004). Cylichna-like shells have occasionally been reported from fossil seep deposits, including an Early Cretaceous site in California (Kaim et al. 2014), a Paleocene site in Spitsbergen (Hryniewicz et al. 2019), and an Oligocene site in Washington, USA (Kiel 2006).
Phylum Arthropoda Von Siebold, 1848
Class Malacostraca Latreille, 1802
Order Decapoda Latreille, 1802
Suborder Pleocyemata Burkenroad, 1963
Infraorder Axiidea Saint Laurent, 1979
Family Callianassidae Dana, 1852
Subfamily Eucalliacinae Manning and Felder, 1991
Genus Eucalliax Manning and Felder, 1991
Type species.—Callianassa quadracuta Biffar, 1970, by original designation; Recent, western Atlantic Ocean.
Eucalliax capsulasetaea sp. nov.
Fig. 18.
Zoobank LCID: urn:lsid:zoobank.org:act:3742EC6D-CCAA-443D-A113-16EA7C8C30CD
Etymology: In reference to capsulated setae, one of diagnostic features of the species.
Type material: Holotype: right propodus (NRM Ar69394) from block 2. Paratypes: right propodus (NRM Ar69376) from block 6, left carpus (NRM Ar69377) from block 7, right propodus (NRM Ar69383) from block 4, and four specimens from block 2: left propodus (NRM Ar69393), right propodus (NRM Ar69397), right propodus (NRM Ar69398), left propodus (NRM Ar69401).
Type locality: Cerro La Salina seep deposits, Talara Basin, Peru.
Type horizon: Presumably the early Oligocene part of the Heath shale.
Material.—The type material and further fragmentary specimens from Cerro La Salina blocks 1, 2–4, 6–8. Complete left carpus (NRM Ar69377), block 7. Seven near-complete left propodi: NRM Ar69378, block 7; NRM Ar69380 (counterpart NRM Ar69379), block 7; NRM Ar69381, block 7; NRM Ar69389 (counterpart NRM Ar69390), block 8; NRM Ar69393, block 2; NRM Ar69401 (counterparts NRM Ar69395, NRM Ar69400) block 1; NRM Ar69402, block 2. Five near-complete right propodi: NRM Ar69376, block 6; NRM Ar69383 (counterparts Ar0069384, NRM Ar69385), block 4; NRM Ar69394, block 2; NRM Ar69397, block 2; NRM Ar69398, block 2. Three fragmentary propodi: NRM Ar69382, block 7; NRM Ar69387, block 1; NRM Ar69396, block 2. One broken right fixed finger: NRM Ar69399, block 2. One complete right dactylus: NRM Ar69388, block 1. One fragmentary dactylus: NRM Ar69386, block 1. All upper Oligocene, Talara Basin, Peru.
Dimensions (in mm).—Measurements are given only for sufficiently preserved propodi (maximum height and length of the manus). NRM Ar69376, H = 12.6, L = 11.5; NRM Ar69378, H = 11.9, L = 12.8; NRM Ar69383, H = 16.0, L = 16.2; NRM Ar69389, H = ~10.0, L = 11.4; NRM Ar69393, H = 18.3, L = 18.1; NRM Ar69394, H = 11.6, L = 10.5; NRM Ar69397, H = 8.6, L = 8.5; NRM Ar69398, H = 12.2, L = 11.4; NRM Ar69401, H = 15.2, L = 19.1.
Diagnosis.—Major manus subrectangular; outer and inner lateral surfaces of manus densely covered with round tubercles, often with individual or serial setal pits; fixed finger with longitudinal keel.
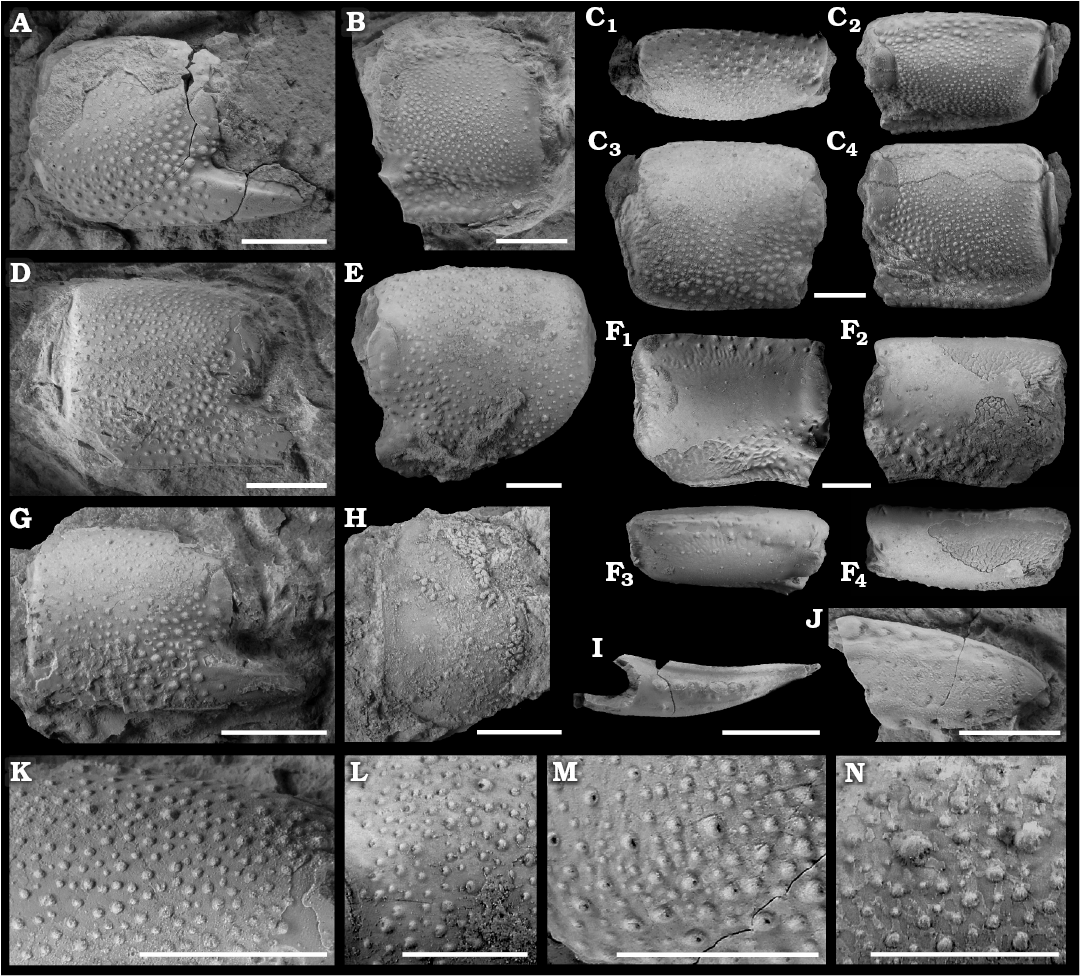
Fig. 18. The callianassid ghost shrimp Eucalliax capsulasetaea sp. nov. from early Oligocene seep deposits at Cerro La Salina, blocks 2, 4, 6, 7, in the Talara Basin, northern Peru. A. Holotype (NRM Ar69394), right propodus in outer lateral view. B. Paratype (NRM Ar69376), right propodus in inner lateral view. C. Paratype (NRM Ar69383), right propodus in dorsal (C1), dorso-lateral (C2), outer lateral (C3), and inner lateral (C4) views. D. Paratype (NRM Ar69398), right propodus in outer lateral view. E. Paratype (NRM Ar69393), left propodus in outer lateral view. F. Paratype (NRM Ar69401), left propodus in inner lateral (F1), outer lateral (F2) and dorso-lateral (F3, F4) views. G. Paratype (NRM Ar69397), right propodus in outer lateral view. H. Paratype (NRM Ar69377), left carpus in outer lateral view. I. NRM Ar69399, right fixed finger in occlusal view. J. NRM Ar69388, right dactylus in outer lateral view. K–N. Details of capsulated setae on outer lateral surfaces of major cheliped propodi. K. NRM Ar69398. L. NRM Ar69393. M. NRM Ar69394. N. NRM Ar69383. Scale bars 5 mm.
Description.—Chelipeds (pereiopod 1) robust. Major carpus taller than long, not armed; upper margin keeled; lower and proximal margins forming continuous rounded margin with an indent at articulation with merus; distal margin with flange. Major propodus rectangular, robust; upper and lower margins keeled, parallel to each other or slightly converging distally; distal margin with or without blunt tooth below articulation with dactylus; area around articulation with dactylus forming bulge; inner and outer lateral surfaces strongly convex, densely covered with tubercles, tubercles closer to margins larger than those closer to longitudinal axis; larger tubercles occur closer to margins; tubercles usually with individual or serial setal pits. Outer lateral surface of fixed finger with longitudinal keel; occlusal margin of fixed finger with serrated keel forming blunt tooth distally. Dactylus robust, unarmed, tip slightly hooked; outer lateral surface with row of round setae positioned close to occlusal margin.
Remarks.—The species is placed within Eucalliax based on the subrectangular manus, the relatively short fixed finger with a triangular outline, and the presence of a longitudinal keel on the outer lateral surface of the fixed finger. This combination of features is characteristic for the genus as discussed in detail previously (Hyžný 2012; Hyžný and Hudáčková 2012).
Eucalliax capsulasetaea sp. nov. was presumably heterochelous, as are most representatives of Callianassidae (Dworschak et al. 2012; Hyžný and Klompmaker 2015). Consequently, all the studied, rather large and massively built cheliped elements are interpreted to represent remains of major claws of pereiopod 1. The remains are nearly equally distributed between right and left chelae (five versus six propodi, respectively). Callianassid ghost shrimps in general show random asymmetry (sensu Palmer 2009) in Recent (e.g., Sakai 1969; Labadie and Palmer 1996) and fossil examples (e.g., Hyžný 2012; Hyžný and Hudáčková 2012), and Eucalliax capsulasetaea sp. nov. supports this observation.
In Eucalliax capsulasetaea sp. nov., the manus is approximately as long as tall (H/L ratio = 0.93–1.10), but one relatively large specimen is distinctly longer (H/L ratio = 0.80; see Dimensions above). NRM Ar69401 is also less tuberculate than other specimens. Nevertheless, all specimens are considered conspecific for several reasons. In extant representatives of Eucalliacinae (i.e., the species of Calliax Saint Laurent, 1973, and Eucalliax Manning and Felder, 1991), considerable variation in the manus H/L ratio is observed and distinctly elongated forms are not uncommon, especially in larger individuals (Dworschak 2006; Hyžný and Gašparič 2014). Also, the presence of setal pores on top of the tubercles covering the lateral surfaces of the elongated manus supports the conspecificity of all callianassid specimens from Cerro La Salina.
The setal pits positioned at the top of tubercles represent capsulated setae. They are not uncommon in hermit crabs (Paguroidea) and have been documented both in Recent (Komai 2003; Osawa 2012; Komai and Rahayu 2014) and fossil forms (Fraaije et al. 2011, 2015; Hyžný et al. 2016). Capsulated setae are rare in ghost shrimps. They have not been mentioned in taxonomic descriptions, but a quick survey of the published figures indicates their presence at least in fossil and extant species of Callianopsis Saint Laurent, 1973 (Karasawa 1997; Schweitzer Hopkins and Feldmann 1997; Lin et al. 2007), although they are limited to a more distal region (i.e., close to the articulation with the dactylus) and their number usually is not exceeding ten. In extant Neocallichirus karumba (Poore and Griffin, 1979) and its supposed fossil relatives, there are several (1–3) such tubercles present at the area of articulation with the dactylus (Dworschak 2008; Hyžný et al. 2016). Thus, Eucalliax capsulasetaea sp. nov. bearing numerous tubercles with capsulated setae is unique among both extant and fossil ghost shrimp species described to date, to our knowledge. The new species is morphologically closest to Eucalliax burckhardti (Böhm, 1911) from the Maastrichtian and Danian of Argentina and Mexico (Hyžný et al. 2013). Both species share densely tuberculated propodi; E. burckhardti, however, does not possess capsulated setae. Besides the presence of capsulated setae, the new species further differs from the type species E. quadracuta in the absence of diagnostic acute projections on the distodorsal and distoventral corners of the carpus and on the distodorsal corner of the propodus (Biffar 1970). Cenozoic representatives, such as E. vicetina Beschin, Busulini, De Angeli, and Tessier, 2002, from the middle Eocene of Italy; E. pseudorakosensis (Lőrenthey in Lőrenthey and Beurlen 1929) from the middle Miocene of Europe; E. yatsuoensis (Karasawa 1993) from the middle Miocene of Japan; and E. miyazakiensis Karasawa, 1993, possess smooth lateral surfaces of the major cheliped propodi. In contrast to the new species, E. yoshihiroi Karasawa, 1992, from the middle Eocene of Japan has sparse tubercles on the lateral surfaces of the major cheliped propodus, but it is not clear from the published figures alone whether setae are present at the top of the tubercles; no further details on this feature are mentioned in the description (Karasawa 1992). Newly presented material of Eucalliax reported from the late Eocene of Italy (Beschin et al. 2019) is too fragmentary and, based on the published figures, these specimens may not be congeneric with Eucalliax.
McLaughlin and Lane (1975) hypothesized that capsulated setae might act both to detect and repel predators, but more studies are needed to assess their function. The function of a limited number of capsulated setae in the ghost shrimps Neocallichirus and Callianopsis may be different than in hermit crabs and the here described new species of Eucalliax with numerous such setae. It is also worth noting that, whereas all Callianopsis species seem to possess capsulated setae, in Neocallichirus they are present only in the group of species morphologically close to N. karumba, a species differing profoundly from N. horneri Sakai, 1988, the type species of Neocallichirus. Consequently, the presence or absence of capsulated setae on ghost shrimps are not always a feature of generic importance, as currently understood.
Muscle scars are preserved in a number of specimens of Eucalliax capsulasetaea sp. nov. These are observed in specimens in which the cuticle is not fully preserved on the inner and outer sides of the manus (Fig. 18F), or, sometimes, on the cuticle. These scars represent closer (adductor) muscles (see Klompmaker et al. 2019), whereas the much smaller (abductor) opener is not seen here.
Eucalliax spp. have been found in the Late Cretaceous, Paleocene, Eocene, Miocene, Pliocene, and Recent (Hyžný and Klompmaker 2015: supplementary file 1). Thus, the new species is the first record from the Oligocene.
Stratigraphic and geographic range.—Type locality and horizon only.
Discussion
With Olsson’s (1931) and our study combined, 39 species of mollusks have been reported from the seep deposits in the Talara Basin, but only five species are shared between Olsson’s (1931) and our study (Pseudophopsis peruviana, Conchocele tessaria, Pleurophopsis unioides, P. talarensis, and “Vesicomya” tschudi; Table 2). The main reason for this difference is that Olsson did not consider small specimens: many species reported here are smaller than ca. 10 mm. Another reason might be that the sediments from which Olsson (1931) extracted his most diverse fauna (“between Belén and Pajarabobo”) appear to have been mostly eroded. This might explain why only five out of 13 of Olsson’s (1931) larger bivalve species are shared with our study.
Table 2. List of species from the lower Oligocene seep carbonates in northern Peru; * includes most likely also the “Cytherea” and “large Cyprina–Isocardia-like shells” listed by Olsson (see Kiel 2013).
|
Species |
La Salina |
El Pelado |
Olsson’s Quebrada Seca |
Belén |
Olsson’s Belén and Pajarabobo |
Lomitos |
Comment |
|
Bivalvia |
|||||||
|
Solemya lomitensis Olsson, 1931 |
|
|
|
|
|
× |
|
|
Solemya (Acharax) belenensis Olsson, 1931 |
|
|
|
|
× |
|
|
|
Nucula paboensis Olsson, 1931 |
|
|
|
|
× |
|
|
|
Acila paita Olsson, 1931 |
|
|
|
? |
× |
|
possibly same as Acila sp. in this paper |
|
Acila piura Olsson, 1931 |
|
|
|
? |
× |
|
possibly same as Acila sp. in this paper |
|
Neilo altamirano sp. nov. |
× |
|
|
|
|
|
|
|
Idas sp. |
× |
|
|
|
|
|
|
|
Propeamussiidae indet. |
× |
|
|
|
|
|
|
|
Pseudophopsis peruviana (Olsson, 1931) |
× |
× |
× |
|
× |
× |
|
|
Conchocele tessaria (Olsson, 1931) |
× |
|
|
|
× |
|
as Thyasira in Olsson (1931) |
|
Thyasira peruviana Olsson, 1931 |
|
|
|
|
|
× |
|
|
Thyasira staufti Olsson, 1931 |
|
|
|
|
|
× |
|
|
Myrtaea? cookei Olsson, 1931 |
|
|
× |
|
|
|
|
|
Myrtaea? sp. |
|
|
|
|
|
× |
|
|
Lucinoma zapotalensis (Olsson, 1931) |
× |
× |
|
|
|
|
as “Phacoides
(Lucinoma)” |
|
Nipponothracia lomitensis (Olsson, 1931) |
|
|
|
|
|
× |
as “Lucina?
lomitensis” |
|
Pleurophopsis lithophagoides Olsson, 1931 |
|
|
|
× |
× |
|
|
|
Pleurophopsis talarensis sp. nov. |
× |
|
× |
|
× |
|
reported as
P. lithophagoides |
|
“Vesicomya” tschudi Olsson, 1931 |
× |
|
|
|
× |
× |
|
|
“Vesicomya” ramondi Olsson, 1931 |
|
|
|
|
× |
|
|
|
Gastropoda |
|||||||
|
Limpet 1 |
× |
|
|
|
|
|
|
|
Limpet 2 |
× |
|
|
|
|
|
|
|
Coccopigya sp. |
× |
|
|
|
|
|
|
|
Retiskenea? sp. |
× |
|
|
|
|
|
|
|
Pyropelta seca sp. nov. |
× |
× |
|
|
|
|
|
|
Cantrainea sp. |
× |
× |
|
|
|
|
|
|
Trochoid incertae sedis |
× |
|
|
|
|
|
|
|
Neritimorpha indet. |
|
|
|
|
× |
× |
listed as “Nerita” by Olsson (1931) |
|
Ampullina or Polinices |
|
|
|
|
|
× |
internal casts |
|
Provanna antiqua Squires, 1995 |
× |
|
|
× |
|
|
|
|
Provanna pelada sp. nov. |
|
× |
|
|
|
|
|
|
Ascheria salina sp. nov. |
× |
|
|
|
|
|
|
|
Colus sekiuensis Kiel and Goedert, 2007 |
× |
|
|
|
|
|
|
|
Buccinidae indet. |
|
|
|
× |
|
|
|
|
Austrofusus? belenensis Olsson, 1931 |
|
|
|
|
× |
|
nomen dubium |
|
Siphonalia? belenensis Olsson, 1931 |
|
|
|
|
× |
|
|
|
Siphonalia? tessaria Olsson, 1931 |
|
|
|
|
× |
|
|
|
Acteon sp. |
× |
|
|
|
|
|
|
|
Cylichna sp. |
× |
|
|
|
|
|
|
|
Decapoda |
|||||||
|
Eucalliax capsulasetaea sp. nov. |
× |
|
|
|
|
|
|
Notably among the taxa we found are Coccopigya sp., which represents the oldest record of this genus, and Ascheria salina, representing the youngest and smallest member of Ascheria. Fossils are rare in the deep-water deposits of the Heath shale (Olsson 1931; Palacios Moncayo 1994) and, hence, methane seeps with their rapid in situ carbonate precipitation are more likely to preserve specimens. Consequently, the deep-water genera Neilo, Propeamussium, Coccopygia, Cantrainea, and Colus, and also the seep-restricted taxa Pyropelta, Provanna, Ascheria, and perhaps Retiskenea are here reported for the first time from the fossil record of Peru.
Among decapods, callianassid ghost shrimps are common associates of ancient seep environments (Klompmaker et al. 2018: fig. 4). Klompmaker et al. (2018) argued that they may be more commonly reported from the fossil seeps compared to modern seeps because these burrowing shrimps are difficult to catch in today’s ocean bottoms. The earliest records of callianassids from seeps thus far are mid-Cretaceous in age (Karasawa 2011; Kiel et al. 2013; Agirrezabala et al. 2013). Eucalliax had not been reported from a fossil seep thus far, unlike Callianassa, Callianopsis, Calliax, Glypturus, and Neocallichirus (see Klompmaker et al. 2018).
Conclusions
Combined with Olsson’s (1931) earlier work, our study revealed a remarkably diverse fauna of mollusks at the early Oligocene seeps in the Talara Basin in northern Peru, encompassing close to 40 species. The majority of them belong to genera known from both coeval and Recent seeps. The biogeographically closest ties are with late Eocene to Oligocene seep faunas in the northeastern Pacific region, with which they share two species and twelve genera. Notable is the absence of larger Bathymodiolus species from the seeps in the Talara basin, as these mussels are quite common in Oligocene seep deposits in the North Pacific region. A newly described ghost shrimp species of Eucalliax is the first occurrence of this genus reported from a seep.
Acknowledgments
We thank Ali Altamirano (Museo de Historia Natural, Lima, Peru) for assistance in the field, Cris Little and Jordan Bestwick (both Leeds University, UK) for letting us use their images of Pseudophopsis peruviana, Andrzej Kaim (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland) for comments on the gastropods, Jim Goedert (Wauna, USA) for comments on the provannids, Judith Nagel-Myers (St. Lawrence University, Canton, USA) and Leslie Skibinski (PRI) for loans of specimens, and the reviewers Kazutaka Amano (Joetsu University of Education, Japan) and Krzysztof Hryniewicz (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland) for their critical comments that greatly improved the manuscript. Financial support was provided by Vetenskapsrådet through grant 2016-03920 to SK, and by the Slovak Research and Development Agency through contract APVV-17-0555 to MH.
References
Abbott, R.T. 1954. American Seashells. xiv + 541 pp. Van Nostrand Reinhold, New York.
Adams, H. and Adams, A. 1853–1858. The Genera of Recent Mollusca Arranged According to Their Organization. Vol. 1: vi–xl, 1–484; Vol. 2: 1–661. John Van Voorst, London.
Agirrezabala, L.M., Kiel, S., Blumenberg, M., Schäfer, N., and Reitner, J. 2013. Outcrop analogues of pockmarks and associated methane-seep carbonates: a case study from Lower Cretaceous (Albian) of the Basque-Cantabrian Basin, western Pyrenees. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 94–115. Crossref
Amano, K. and Jenkins, R.G. 2011a. Fossil record of extant vesicomyid species from Japan. Venus 69: 163–176.
Amano, K. and Jenkins, R.G. 2011b. New fossil Bathymodiolus (s.l.) (Mytilidae, Bivalvia) from Oligocene seep-carbonates in eastern Hokkaido, Japan—with remarks on the evolution of Bathymodiolus (s.l.). The Nautilus 125: 29–35.
Amano, K. and Jenkins, R.G. 2013. A new species of Provanna (Gastropoda: Provannidae) from an Oligocene seep deposit in eastern Hokkaido, Japan. Paleontological Research 17: 325–329. Crossref
Amano, K. and Kiel, S. 2007. Fossil vesicomyid bivalves from the North Pacific region. The Veliger 49: 270–293.
Amano, K. and Kiel, S. 2010. Taxonomy and distribution of fossil Archivesica (Vesicomyidae, Bivalvia) in Japan. The Nautilus 124: 155–165.
Amano, K. and Kiel, S. 2011. Fossil Adulomya (Vesicomyidae, Bivalvia) from Japan. The Veliger 51: 76–90.
Amano, K. and Kiel, S. 2012. Two Neogene vesicomyid species (Bivalvia) from Japan and their biogeographic implications. The Nautilus 126: 79–85.
Amano, K. and Little, C.T.S. 2005. Miocene whale-fall community from Hokkaido, northern Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 215: 345–356. Crossref
Amano, K. and Little, C.T.S. 2014. Miocene abyssochrysoid gastropod Provanna from Japanese seep and whale-fall sites. Acta Palaeontologica Polonica 59: 163–172.
Amano, K., Jenkins, R.G., and Kurita, H. 2018a. New and Mesozoic-relict mollusks from Paleocene wood-fall communities in Urahoro Town, eastern Hokkaido, northern Japan. Journal of Paleontology 92: 634–647. Crossref
Amano, K., Jenkins, R.G., Ohara, M., and Kiel, S. 2014a. Miocene vesicomyid species (Bivalvia) from Wakayama in southern Honshu, Japan. The Nautilus 128: 9–17.
Amano, K., Little, C.T.S., and Campbell, K.A. 2018b. Lucinid bivalves from Miocene hydrocarbon seep sites of eastern North Island, New Zealand, with comments on Miocene New Zealand seep faunas. Acta Palaeontologica Polonica 63: 371–382. Crossref
Amano, K., Little, C.T.S., and Inoue, K. 2007. A new Miocene whale-fall community from Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 247: 236–242. Crossref
Amano, K., Little, C.T.S., Campbell, K.A., Jenkins, R.G., and Saether, K.P. 2015. Paleocene and Miocene Thyasira sensu stricto (Bivalvia: Thyasiridae) from chemosynthetic communities from Japan and New Zealand. The Nautilus 129: 43–53.
Amano, K., Saether, K.P., Little, C.T.S., and Campbell, K.A. 2014b. Fossil vesicomyid bivalves from Miocene hydrocarbon seep sites, North Island, New Zealand. Acta Palaeontologica Polonica 59: 421–428.
Beschin, C., Busulini, A., De Angeli, A., and Tessier, G. 2002. Aggiornamento ai crostacei eocenici di cava „Main“ di Arzignano (Vicenza, Italia settentrionale) (Crustacea, Decapoda). Studi e Ricerche Associazione Amici Museo Zannato Museo Civico „G. Zannato“ 2002: 7–28.
Beschin, C., Busulini, A., Fornacciari, F., Papazzoni, C.A., and Tessier, G. 2019. La fauna di crostacei associati a coralli dell’Eocene superiore di Campolongo di Val Liona (Monti Berici, Vicenza, Italia Nordorientale). Bollettino del Museo di Storia Naturale di Venezia 69: 129–215.
Beu, A.G. and Maxwell, P.A. 1990. Cenozoic Mollusca of New Zealand. New Zealand Geological Survey Paleontological Bulletin 58: 1–518.
Beu, A.G., Nolden, S., and Darragh, T.A. 2012. Revision of New Zealand Cenozoic fossil Mollusca described by Zittel (1865) based on Hochstetter’s collections from the Novara Expedition. Memoirs of the Association of Australasian Palaeontologists 43: 1–69.
Beurlen, K. 1944. Beiträge zur Stammesgeschichte der Muscheln. Bayerische Akademie der Wissenschaften, Sitzungsberichte 1–2: 133–145.
Biffar, T.A. 1970. Three new species of callianassid shrimp (Decapoda, Thalassinidea) from the western Atlantic. Proceedings of the Biological Society of Washington 83: 35–50.
Bouchet, P., Rocroi, J.-P., Hausdorf, B., Kaim, A., Kano, Y., Nützel, A., Parkhaev, P., Schrödl, M., and Strong, E.E. 2017. Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 61: 1–526. Crossref
Burkenroad, M.D. 1963. The evolution of the Eucarida, (Crustacea, Eumalacostraca), in relation to the fossil record. Tulane Studies in Geology and Paleontology 2: 2–17.
Burmeister, H. 1837. Handbuch der Naturgeschichte, Vol. 2, Zoologie. i–xii + 369–858 pp. Enslin, Berlin.
Böhm, J. 1911. Callianassa burckhardti n. sp. nebst einer Zusammenstellung der fossilen Arten der Gattung Callianassa. Zeitschrift der deutschen geologischen Gesellschaft, Monatsberichte 63: 37–46.
Campbell, K.A., Peterson, D., and Alfaro, A.C. 2008. Two new species of Retiskenea? (Gastropoda: Neomphalidae) from Lower Cretaceous hydrocarbon seep-carbonates of northern California. Journal of Paleontology 82: 140–153. Crossref
Cantraine, F.J. 1835. Diagnoses ou descriptiones succinctes de quelques espèces nouvelles de mollusques. Bulletin de l’Académie Royale des Sciences et Belles-lettres de Bruxelles 2: 380–401.
Chavan, A. 1954. Les Pleurophorus et genres voisins. Cahier Géologiques Seyssel 22: 200.
Conrad, R.A. 1849. Fossils from the northwestern America. In: J.D. Dana (ed.), U.S. Exploration Expedition, 1838–1842, under Charles Wilkes. Geology, Volume 10, 723–728 (appendix). C. Sherman, Philadelphia.
Contardo-Berríos, X., Mena-Hodges, E., and Quirago, E. 2017. Hallazgo del primer seep fósil en la costa emergida de Chile central (33°56’ S). Caracterización e implicancias. Andean Geology 44: 213–228. Crossref
Cooke, C.W. 1919. Contributions to the geology and paleontology of the West Indies IV. Tertiary mollusks from the leeward islands and Cuba. Carnegie Institution of Washington publications 291: 103–156.
Cossmann, M. and Peyrot, A. 1917. Conchologie néogénique de l’Aquitaine. Tome 3, Gastropodes, Scaphopodes et Amphineures. Actes de la Société Linnéenne de Bordeaux 69 (4), 285–365.
Cox, L.R. 1960. Gastropoda. General characteristics of Gastropoda. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Pt I. Mollusca, 1, I85–I169. Geological Society of America and University of Kansas Press, Lawrence.
Cuvier, G.L.C.F.D. 1795. Second Mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des Mollusques et de leur division en ordre, lu à la société d’Histoire Naturelle de Paris, le 11 prairial an troisième. Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts 2: 433–449.
d’Orbigny, A. 1842–43. Paléontologie Française, terrains crétacés. Vol. 2, Gastèropodes. 645 pp. Privately published, Paris.
Dall, W.H. 1882. On certain limpets and chitons from the deep waters off the eastern coast of the United States. Proceedings of the U.S. National Museum of Natural History 4: 400–414. Crossref
Dall, W.H. 1886. Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Gulf of Mexico (1877–1878) and in the Caribbean Sea (1879–1880), by the U.S. Coast Survey steamer “Blake” XXIX. Report on the Mollusca. Part 1, Brachiopoda and Pelecypoda. Bulletin of the Museum of Comparative Zoology, Harvard University 12: 171–318.
Dall, W.H. 1900. Contributions to the Tertiary fauna of Florida, with especial reference to the Miocene silex-beds of Tampa and the Pliocene beds of the Caloosahatchie River. Part 5, Teleodemacea: Solen to Diplodonta. Transactions of the Wagner free Institute of Science, Philadelphia 3: 949–1218.
Dall, W.H. 1901. Synopsis of the Lucinacea and of the American species. Proceedings of the U.S. National Museum of Natural History 23: 779–833. Crossref
Dall, W.H. 1908. The Mollusca and the Brachiopoda. Bulletin of the Museum of Comparative Zoology, Harvard University 43 (6): 205–487. Crossref
Dall, W.H. 1918. Description of new species of shells chiefly from Magdalena Bay, Lower California. Proceedings of the Biological Society of Washington 31: 5–8.
Dall, W.H. and Simpson, C.T. 1901. The Mollusca of Porto Rico. United States Fishery Commission, Bulletin 20: 351–524.
Dana, J.D. 1852. Crustacea. Part I. United States Exploring Expedition, during the years 1838, 1839, 1840, 1841, 1842, under the command of Charles Wilkes, U.S.N.13. viii + 685 pp. C. Sherman, Philadelphia.
Dantart, L. and Luque, Á.A. 1994. Cocculiniformia and Lepetidae (Gastropoda: Archaeogastropoda) from Iberian waters. Journal of Molluscan Studies 60: 277–313. Crossref
Decker, C., Olu, K., Cunha, R.L., and Arnaud-Haond, S. 2012. Phylogeny and diversification patterns among vesicomyid bivalves. PLoS ONE 7: e33359. Crossref
Del Río, C.A. and Camacho, H.H. 1998. Tertiary nuculoids and arcoids of eastern Patagonia (Argentina). Palaeontographica A 250: 47–88.
Dockery, D.T. 1977. Mollusca of the Moodys Branch Formation, Mississippi. Mississippi Office of Geology Bulletin 122: 1–212.
Dworschak, P.C. 2006. A new species of Eucalliax Manning & Felder, 1991 (Decapoda: Callianassidae) from the Philippines. The Raffles Bulletin of Zoology 54: 349–359.
Dworschak, P.C. 2008. Neocallichirus kempi Sakai, 1999, a junior synonym of Callianassa karumba Poore & Griffin, 1979 (Decapoda: Callianassidae). The Raffles Bulletin of Zoology 56: 75–84.
Dworschak, P.C., Felder, D.L., and Tudge, C.C. 2012. Infraorders Axiidea de Saint Laurent, 1979 and Gebiidea de Saint Laurent, 1979 (formerly known collectively as Thalassinidea). In: F.R. Schram, J.C. von Vaupel Klein, M. Charmantier-Daures, and J. Forest (eds.), Treatise on Zoology—Anatomy, Taxonomy, Biology—The Crustacea, Decapoda, Decapoda: Astacidea P.P. (Enoplometopoidea, Nephropoidea), Glypheidea, Axiidea, Gebiidea, and Anomura, Vol. 9B, 109–219. Brill, Leiden. Crossref
Eagle, M.K. and Hayward, B.W. 1993. Oligocene paleontology and paleoecology of Waitete Bay, northern Coromandel Peninsula. Records of the Auckland Institute and Museum 30: 13–26.
Egorova, E.N. 1982. Mollusca of the Davis Sea (the Eastern Antarctic region) [in Russian]. Rezul’taty biologičeskih issledovanij sovetskih antarktičeskih èkspedicij 7, Issledovaniâ fauny morej 26: 1–134.
Erdmann, S. and Morra, G. 1985. Nuevos Moluscos de la Formación San Julián, Provincia de Santa Cruz. Ameghiniana 22: 289–295.
Finlay, H.J. 1926. New shells from New Zealand Tertiary beds: Part 2. Transactions of the New Zealand Institute 56: 227–258.
Fleming, J. 1828. A History of British Animals, Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca and Radiata of the United Kingdom; Including the Indigenous, Extirpated, and Extinct Kinds; Together with Periodical and Occasional Visitants. xxiii + 554 pp. Bell & Bradfute, Edinburgh. Crossref
Fraaije, R.H.B., van Bakel, B.W.M., Iserbyt, A., and Jagt, J.W.M. 2011. New extinct Paguroidea (Crustacea, Decapoda, Anomura), with the first example of capsulated setae from the fossil record. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 262: 247–255. Crossref
Fraaije, R.H.B., van Bakel, B.W.M., and Jagt, J.W.M. 2015. A new Albian hermit crab (Anomura, Paguroidea) from France—another example of capsulated setae in an extinct form. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 277: 353–359. Crossref
Gabb, W.M. 1866–1869. Cretaceous and Tertiary fossils. California Geological Survey, Paleontology 2: 1–299.
Garvie, C.L. 2013. New Eocene Mollusca from the collections of the Texas Natural Science Center. Bulletins of American Paleontology 386: 163–183.
Gill, F.L. and Little, C.T.S. 2013. A new genus of lucinid bivalve from hydrocarbon seeps. Acta Palaeontologica Polonica 58: 573–578. Crossref
Gill, F.L., Harding, I.C., Little, C.T.S., and Todd, J.A. 2005. Palaeogene and Neogene cold seep communities in Barbados, Trinidad and Venezuela: An overview. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 191–209. Crossref
Goedert, J.L. and Benham, S.R. 1999. A new species of Depressigyra? (Gastropoda: Peltospiridae) from cold-seep carbonates in Eocene and Oligocene rocks of western Washington. The Veliger 42: 112–116.
Goedert, J.L. and Campbell, K.A. 1995. An Early Oligocene chemosynthetic community from the Makah Formation, northwestern Olympic Peninsula, Washington. The Veliger 38: 22–29.
Goedert, J.L. and Kaler, K.L. 1996. A new species of Abyssochrysos (Gastropoda: Loxonematoidea) from a Middle Eocene cold-seep carbonate in the Humptulips Formation, western Washington. The Veliger 39: 65–70.
Goedert, J.L. and Squires, R.L. 1993. First Oligocene record of Calyptogena (Bivalvia: Vesicomyidae). The Veliger 36: 72–77.
Golikov, A.N. and Starobogatov, Y.I. 1975. Systematics of prosobranch gastropods. Malacologia 15: 185–232.
Gray, J.E. 1824. Shells. A Supplement to the Appendix of Captain Parry’s Voyage for the Discovery of a North-west Passage, in the Years 1819–1820: Containing an Account of the Subjects of Natural History. ccxl–ccxlvi pp. Murray, London.
Griffin, M. and Nielsen, S.N. 2008. A revision of the type specimens of Tertiary molluscs from Chile and Argentina described by D’Orbigny (1842), Sowerby (1846) and Hupé (1854). Journal of Systematic Palaeontology 6: 251–316. Crossref
Griffin, M. and Pastorino, G. 2006. Madrynomya bruneti n. gen. and sp. (Bivalvia: ?Modiomorphoidae): a Mesozoic survivor in the Tertiary of Patagonia? Journal of Paleontology 80: 272–282. Crossref
Haszprunar, G. 1987. Anatomy and affinities of cocculinid limpets (Mollusca Archaeogastropoda). Zoologica Scripta 16: 305–324. Crossref
Hickman, C.S. 2015. Paleogene marine bivalves of the deep-water Keasey Formation in Oregon, part III: The heteroconchs. PaleoBios 32: 1–44.
Hickman, C.S. and McLean, J.H. 1990. Systematic revision and suprageneric classification of trochacean gastropods. Natural History Museum of Los Angeles County Science Series 35: 1–169.
Higley, D. 2004. The Talara basin province of northwestern Peru: Cretaceous–Tertiary total petroleum system. U.S. Geological Survey, e-Bulletin B-2206a: 52.
Hinds, R.B. 1843. Descriptions of new species of Nucula from the collections of Sir Edward Belcher, C.B., and Hugh Cuming, Esq. Proceedings of the Zoological Society of London 1843: 97–101.
Hryniewicz, K., Amano, K., Bitner, M.A., Hagström, J., Kiel, S., Klompmaker, A.A., Mörs, T., Robins, C.M., and Kaim, A. 2019. A late Paleocene fauna from shallow-water chemosynthesis-based ecosystems in Spitsbergen, Svalbard. Acta Palaeontologica Polonica 64: 101–141. Crossref
Hryniewicz, K., Amano, K., Jenkins, R.G., and Kiel, S. 2017. Thyasirid bivalves from Cretaceous and Paleogene cold seeps. Acta Palaeontologica Polonica 62: 705–728. Crossref
Hutton, F.W. 1887. The Mollusca of the Pareora and Oamaru Systems of New Zealand. Proceedings of the Linnean Society of New South Wales 11: 205–237.
Hybertsen, F. and Kiel, S. 2018. A middle Eocene seep deposit with silicified fauna from the Humptulips Formation in western Washington State, USA. Acta Palaeontologica Polonica 63: 751–768. Crossref
Hyžný, M. 2012. Calliaxina chalmasii (Brocchi, 1883) comb. nov. (Decapoda: Axiidea: Callianassidae: Eucalliacinae), a ghost shrimp from the Middle Miocene of Europe, with reappraisal of the fossil record of Eucalliacinae. Zootaxa 3492: 49–64.
Hyžný, M. and Gašparič, R. 2014. Ghost shrimp Calliax de Saint Laurent, 1973 (Decapoda: Axiidea: Callianassidae) in the fossil record: systematics, palaeoecology and palaeobiogeography. Zootaxa 3821: 37–57. Crossref
Hyžný, M. and Hudáčková, N. 2012. Redescription of two ghost shrimps (Decapoda: Axiidea: Callianassidae) from the Middle Miocene of the Central Paratethys: systematics, intraspecific variation, and in situ preservation. Zootaxa 3210: 1–25. Crossref
Hyžný, M. and Klompmaker, A.A. 2015. Systematics, phylogeny, and taphonomy of ghost shrimps (Decapoda): a perspective from the fossil record. Arthropod Systematics & Phylogeny 73: 401–437.
Hyžný, M., Charbonnier, S., Merle, D., Ahmed Lashari, R., Bartolini, A., and Métais, G. 2016. New Early Cenozoic ghost shrimps (Decapoda, Axiidea, Callianassidae) from Pakistan and their palaeobiogeographic implications. Geodiversitas 38: 341–353. Crossref
Hyžný, M., Fraaije, R.H.B., Martin, J.E., Perrier, V., and Sarr, R. 2016. Paracapsulapagurus poponguinensis, a new hermit crab (Decapoda: Anomura: Paguroidea) from the Maastrichtian of Senegal. Journal of Paleontology 90: 1133–1137. Crossref
Hyžný, M., Vega, F.J., and Coutiño, M.A. 2013. Ghost shrimps (Decapoda: Axiidea: Callianassidae) of the Maastrichtian (Late Cretaceous) Ocozocoautla Formation, Chiapas (Mexico). Boletín de la Sociedad Geológica Mexicana 65: 255–264. Crossref
Jeffreys, J.G. 1876. New and peculiar Mollusca of the Pecten, Mytilus and Arca families procured in the “Valorous” Expedition. Annals and Magazine of Natural History 4: 424–436. Crossref
Jeffreys, J.G. 1883. On the Mollusca procured during the Lightning and Porcupine expeditions, 6. Proceedings of the Zoological Society of London 1883: 88–115. Crossref
Jenkins, R.G., Kaim, A., Amano, K., Sakurai, K., and Matsubara, K. 2018a. A new Miocene whale-fall community dominated by bathymodiolin mussel Adipicola from Hobetsu area, Hokkaido, Japan. Paleontological Research 22: 105–111. Crossref
Jenkins, R.G., Kaim, A., and Hikida, Y. 2007. Antiquity of the substrate choice among acmaeid limpets from the Late Cretaceous chemosynthesis-based communities. Acta Palaeontologica Polonica 52: 369–373.
Jenkins, R.G., Kaim, A., Hikida, Y., and Kiel, S. 2018b. Four new species of the Jurassic to Cretaceous seep-restricted bivalve Caspiconcha and implications for the history of chemosynthetic communities. Journal of Paleontology 92: 596–610. Crossref
Jenkins, R.G., Kaim, A., Little, C.T.S., Iba, Y., Tanabe, K., and Campbell, K.A. 2013. Worldwide distribution of modiomorphid bivalve genus Caspiconcha in late Mesozoic hydrocarbon seeps. Acta Palaeontologica Polonica 58: 357–382.
Johnson, S.B., Krylova, E.M., Audzijonyte, A., Sahling, H., and Vrijenhoek, R.C. 2017. Phylogeny and origins of chemosynthetic vesicomyid clams. Systematics and Biodiversity 15: 346–360. Crossref
Kaim, A. and Schneider, S. 2012. A conch with a collar: Early ontogeny of the enigmatic fossil bivalve Myoconcha. Journal of Paleontology 86: 653–659. Crossref
Kaim, A., Jenkins, R.G., and Hikida, Y. 2009. Gastropods from Late Cretaceous hydrocarbon seep deposits in Omagari and Yasukawa, Nakagawa area, Hokkaido, Japan. Acta Palaeontologica Polonica 54: 463–690. Crossref
Kaim, A., Jenkins, R.G., and Warén, A. 2008. Provannid and provannid-like gastropods from Late Cretaceous cold seeps of Hokkaido (Japan) and the fossil record of the Provannidae (Gastropoda: Abyssochrysoidea). Zoological Journal of the Linnean Society 154: 421–436. Crossref
Kaim, A., Jenkins, R.G., Tanabe, K., and Kiel, S. 2014. Mollusks from late Mesozoic seep deposits, chiefly in California. Zootaxa 3861: 401–440. Crossref
Kanno, S., Tanaka, K., Koike, H., Narita, K., and Endo, T. 1998. Adulomya uchimuraensis Kuroda (Bivalvia) from the Miocene Bessho Formation in Shiga-mura, Nagano Prefecture, Japan. Research Reports of the Shinshushinmachi Fossil Museum 1: 17–28.
Karasawa, H. 1992. Fossil decapod crustaceans from the Manda Group (middle Eocene), Kyushu, Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series 167: 1247–1258.
Karasawa, H. 1993. Cenozoic decapod Crustacea from southwest Japan. Bulletin of the Mizunami Fossil Museum 20: 1–92.
Karasawa, H. 1997. A monograph of Cenozoic stomatopod, decapod, isopod and amphipod Crustacea from west Japan. Monograph of the Mizunami Fossil Museum 8: 1–81.
Karasawa, H. 2011. New axiidean Decapoda from the Albian (Lower Cretaceous) chemosynthetic community of Hokkaido, Japan. Bulletin of the Mizunami Fossil Museum 37: 27–29.
Kelly, S.R.A., Blanc, E., Price, S.P., and Withham, A.G. 2000. Early Cretaceous giant bivalves from seep-related limestone mounds, Wollaston Forland, Northeast Greenland. In: E.M. Harper, J.D. Taylor, and J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia Geological Society of London, Special Publication, 227–246. Geological Society of London, London. Crossref
Kenk, V.C. and Wilson, B.R. 1985. A new mussel (Bivalvia: Mytilidae) from hydrothermal vents in the Galapagos Rift Zone. Malacologia 26: 253–271.
Kiel, S. 2004. Shell structures of selected gastropods from hydrothermal vents and seeps. Malacologia 46: 169–183.
Kiel, S. 2006. New records and species of mollusks from Tertiary cold-seep carbonates in Washington State, USA. Journal of Paleontology 80: 121–137. Crossref
Kiel, S. 2007. Status of the enigmatic fossil vesicomyid bivalve Pleurophopsis. Acta Palaeontologica Polonica 52: 639–642.
Kiel, S. 2008. An unusual new gastropod genus from an Eocene hydrocarbon seep in Washington State, USA. Journal of Paleontology 82: 188–191. Crossref
Kiel, S. 2010. On the potential generality of depth-related ecologic structure in cold-seep communities: Cenozoic and Mesozoic examples. Palaeogeography, Palaeoclimatology, Palaeoecology 295: 245–257. Crossref
Kiel, S. 2013. Lucinid bivalves from ancient methane seeps. Journal of Molluscan Studies 79: 346–363. Crossref
Kiel, S., Altamirano, A.J., Birgel, D., Coxall, H.K., Hybertsen, F., and Peckmann, J. 2019. Fossiliferous methane-seep deposits from the Cenozoic Talara Basin in northern Peru. Lethaia [published online, https://doi.org/10.1111/let.12349]. Crossref
Kiel, S., Amano, K., and Jenkins, R.G. 2008. Bivalves from Cretaceous cold-seep deposits on Hokkaido, Japan. Acta Palaeontologica Polonica 53: 525–537. Crossref
Kiel, S., Amano, K., and Jenkins, R.G. 2016. Predation scar frequencies in chemosymbiotic bivalves at an Oligocene seep deposit and their potential relation to inferred sulfide tolerances. Palaeogeography, Palaeoclimatology, Palaeoecology 453: 139–145. Crossref
Kiel, S. and Amano, K. 2013. The earliest bathymodiolin mussels: Evaluation of Eocene and Oligocene taxa from deep-sea methane seep deposits in western Washington State, USA. Journal of Paleontology 87: 589–602. Crossref
Kiel, S. and Goedert, J.L. 2006a. A wood-fall association from Late Eocene deep-water sediments of Washington State, USA. Palaios 21: 548–556. Crossref
Kiel, S. and Goedert, J.L. 2006b. Deep-sea food bonanzas: Early Cenozoic whale-fall communities resemble wood-fall rather than seep communities. Proceedings of the Royal Society B 273: 2625–2631. Crossref
Kiel, S. and Goedert, J.L. 2007. Six new mollusk species associated with biogenic substrates in Cenozoic deep-water sediments in Washington State, USA. Acta Palaeontologica Polonica 52: 41–52.
Kiel, S. and Hansen, B.T. 2015. Cenozoic methane-seep faunas of the Caribbean region. PLoS ONE 10: e0140788. Crossref
Kiel, S. and Peckmann, J. 2007. Chemosymbiotic bivalves and stable carbon isotopes indicate hydrocarbon seepage at four unusual Cenozoic fossil localities. Lethaia 40: 345–357. Crossref
Kiel, S. and Taviani, M. 2017. Chemosymbiotic bivalves from Miocene methane-seep carbonates in Italy. Journal of Paleontology 91: 444–466. Crossref
Kiel, S. and Taviani, M. 2018. Chemosymbiotic bivalves from the late Pliocene Stirone River hydrocarbon seep complex in northern Italy. Acta Palaeontologica Polonica 63: 557–568. Crossref
Kiel, S., Birgel, D., Campbell, K.A., Crampton, J.S., Schiøler, P., and Peckmann, J. 2013. Cretaceous methane-seep deposits from New Zealand and their fauna. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 17–34. Crossref
Kiel, S., Campbell, K.A., and Gaillard, C. 2010. New and little known mollusks from ancient chemosynthetic environments. Zootaxa 2390: 26–48. Crossref
Kiel, S., Sami, M., and Taviani, M. 2018. A serpulid-Anodontia-dominated methane-seep deposit from the Miocene of northern Italy. Acta Palaeontologica Polonica 63: 569–577. Crossref
King, W. 1848. An account of some shells and other invertebrate forms found on the coast of Northumberland and of Durham. Annals and Magazine of Natural History 18: 233–251. Crossref
Klompmaker, A.A., Hyžný, M., Portell, R.W., Jauvion, C., Charbonnier, S., Fussell, S.S., Klier, A.T., Tejera, R., and Jakobsen, S.L. 2019. Muscles and muscle scars in fossil malacostracan crustaceans. Earth-Science Reviews 194: 306–326. Crossref
Klompmaker, A.A., Nyborg, T., Brezina, J., and Ando, Y. 2018. Crustaceans in cold seep ecosystems: fossil record, geographic distribution, taxonomic composition, and biology. PaleorXiv [preprint published online, https://doi.org/10.31233/osf.io/tws6m]. Crossref
Komai, T. 2003. Identities of Pagurus japonicus (Stimpson, 1858) and P. similis (Ortmann, 1892), with description of a new species of Pagurus. Zoosystema 25: 377–411.
Komai, T. and Rahayu, D.L. 2014. New records and new species of the hermit crab genus Pagurus Fabricius, 1775 (Crustacea: Decapoda: Anomura: Paguridae) from the Philippines. The Raffles Bulletin of Zoology 62: 620–646.
Krylova, E.M. and Janssen, R. 2006. Vesicomyidae from Edison Seamount (South West Pacific: Papua New Guinea: New Ireland fore-arc basin). Archiv für Molluskenkunde 135: 231–261. Crossref
Krylova, E.M. and Sahling, H. 2010. Vesicomyidae (Bivalvia): current taxonomy and distribution. PLoS ONE 5: e9957. Crossref
Krylova, E.M., Sahling, H., and Janssen, R. 2010. Abyssogena: a new genus of the family Vesicomyidae (Bivalvia) from deep water vents and seeps. Journal of Molluscan Studies 76: 107–132. Crossref
Kugler, H.G., Jung, P., and Saunders, J.B. 1984. The Joes River Formation of Barbados and its fauna. Eclogae Geologicae Helvetiae 77: 675–705.
Kuroda, T. 1931. Fossil Mollusca. In: F. Homma (ed.), Geology of the Central Part of Shinano, Part 4, 1–90. Kokon Shoin, Tokyo.
Labadie, L.V. and Palmer, A.R. 1996. Pronounced heterochely in the ghost shrimp, Neotrypaea californiensis (Decapoda: Thalassinidea: Callianassidae): allometry, inferred function and development. Journal of Zoology 240: 659–675. Crossref
Latreille, P.A. 1802–1803. Histoire naturelle, générale et particulière, des Crustacés et des Insectes, 3. 468 pp. F. Dufart, Paris.
Leanza, A.F. 1940. Myoconcha neuquena n. sp. del Lias de Piedra Pintada en el Neuquén. Notas del Museo de La Plata, Paleontología 5: 123–131.
Lin, F.-J., Komai, T., and Chan, T.-Y. 2007. First record of the thalassinidean genus Callianopsis de Saint Laurent, 1973 (Decapoda, Ctenochelidae) in the West Pacific, with the description of a new species from Taiwan. Crustaceana 80: 1193–1203. Crossref
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Salvius, Stockholm. Crossref
Lovén, S.L. 1846. Molluscorum litora Scandinaviae occidentalis habitantium. Öfversigt af Kongliga Vetenskaps-Akademien Förhandlingar 3: 134–160, 182–204. Crossref
Lőrenthey, E. and Beurlen, K. 1929. Die fossilen Dekapoden der Länder der Ungarischen Krone. Geologica Hungarica, Series Palaeontologica 3: 1–421.
Manning, R.B. and Felder, D.L. 1991. Revision of the American Callianassidae (Crustacea: Decapoda: Thalassinidea). Proceedings of the Biological Society of Washington 104: 764–792.
Marshall, B.A. 1978. The genus Neilo in New Zealand (Mollusca: Bivalvia). New Zealand Journal of Zoology 5: 425–436. Crossref
Marshall, B.A. 1979. The Trochidae and Turbinidae of the Kermadec Ridge (Mollusca: Gastropoda). New Zealand Journal of Zoology 6: 521–552. Crossref
Marshall, B.A. 1986. Recent and Tertiary Cocculinidae and Pseudococculinidae (Mollusca: Gastropoda) from New Zealand and New South Wales. New Zealand Journal of Zoology 12: 505–546. Crossref
Marwick, J. 1931. The Tertiary Mollusca of the Gisborne District. New Zealand Geological Survey Bulletin 113: 1–177.
Marwick, J. 1953. Divisions and faunas of the Hokonui System (Triassic and Jurassic). New Zealand Geological Survey Palaeontological Bulletin 21: 1–141.
McLaughlin, P.A. and Lane, C.E. 1975. The morphology of unique structures on the spines of a deep-water Hawaiian hermit crab (Crustacea: Decapoda: Paguridae). Journal of Zoology 176: 519–526. Crossref
McLean, J.H. 1981. The Galapagos Rift limpet Neomphalus: Relevance to understanding the evolution of a major Paleozoic–Mesozoic radiation. Malacologia 21: 291–336.
McLean, J.H. 1992. Cocculiniform limpets (Cocculinidae and Pyropeltidae) living on whale bone in the deep sea off California. Journal of Molluscan Studies 58: 401–414. Crossref
McLean, J.H. and Geiger, D.L. 1998. New genera and species having the Fissurisepta shell form, with a generic-level phylogenetic analysis (Gastropoda: Fissurellidae). Contributions in Science, Natural History Museum of Los Angeles County 475: 1–32.
McLean, J.H. and Harasewych, M.G. 1995. Review of western Atlantic species of cocculinid and pseudococculinid limpets, with descriptions of new species (Gastropoda: Cocculiniformia). Contributions in Science, Natural History Museum of Los Angeles County 453: 1–33.
McLean, J.H. and Haszprunar, G. 1987. Pyropeltidae, a new family of cocculiniform limpets from hydrothermal vents. The Veliger 30: 196–205.
Miyajima, Y., Nobuhara, T., and Koike, H. 2017. Taxonomic reexamination of three vesicomyid species (Bivalvia) from the middle Miocene Bessho Formation in Nagao Prefecture, central Japan, with notes on vesicomyid diversity. The Nautilus 131: 51–66.
Mohr, N. 1786. Forsög til en islandsk naturhistorie. 414 pp. C.F. Holm, Copenhagen.
Montfort, D.P.d. 1810. Conchyliologie systématique et classification méthodique des coquilles: Offrant leur figures, leur arrengement générique, leur description charactéristiques, leur noms; ainsi que leur synonymie en plusieurs langues. 676 pp. F. Schoell, Paris.
Moore, E.J. 1963. Miocene marine mollusks from the Astoria Formation in Oregon. U.S. Geological Survey Professional Paper 419: 1–109.Crossref
Moskalev, L.I. 1976. On the generic classification in Cocculinidae (Gastropoda, Prosobranchia) [in Russian]. Trudy Instituta Okeanologii Imeni P.P. Širšov 99: 59–70.
Mulsant, E. 1842. Lamellicornes. Histoire naturelle des Coléoptères de France. 304 pp. Maison, Paris.
Neumayr, M. 1884. Zur Morphologie des Bivalvenschlosses. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften 88: 385–419.
Newell, N.D. 1957. Notes on certain primitive heterodont pelecypods. American Museum Novitates 1857: 1–14.
Nordsieck, F. 1973. Molluschi abyssali dello Jonio. La Conchiglia 57–58: 4–7.
Okutani, T. 2001. Six new bathyal and shelf trochoidean species in Japan. Venus 60: 121–127.
Okutani, T., Fujikura, K., and Sasaki, T. 1993. New taxa and new distribution records of deepsea gastropods collected from or near the chemosynthetic communities in the Japanese waters. Bulletin of the National Science Museum Series A (Zoology) 19: 123–143.
Okutani, T., Tsuchida, S., and Fujikura, K. 1992. Five bathyal gastropods living within or near the Calyptogena-community of the Hatsushima Islet, Sagami Bay. Venus 51: 137–148.
Olsson, A.A. 1931. Contributions to the Tertiary paleontology of northern Peru: Part 4, The Peruvian Oligocene. Bulletins of American Paleontology 17: 97–264.
Olsson, A.A. 1964. Neogene Mollusks from Northwestern Ecuador. 256 pp. Paleontological Research Institution, Ithaca.
Ordóñez, M. 1995. Zonas de Radiolarios des Eoceno medio del grupo Ancon, peninsula de Santa Elena, Ecuador. Asociación Paleontológica Argentina, Publicacion Especial 3: 107–117.
Ortmann, A.E. 1902. Reports of the Princeton University Expeditions to Patagonia, 1896–1899. Vol. IV. Palaeontology. Part II. Tertiary Invertebrates. 332 pp. E. Schweizerbart’sche Verlagsbuchhandlung (E. Nägele), Stuttgart.
Osawa, M. 2012. A new species of the genus Pagurus Fabricius, 1775 (Crustacea: Decapoda: Anomura: Paguridae) from the Ryukyu Islands, southwestern Japan. In: T. Naruse, T.Y. Chan, H.H. Tan, S.T. Ahyong, and J.D. Reimer (eds.), Scientific Results of the Marine Biodiversity Expedition—KUMEJIMA 2009. Zootaxa 3367: 155–164. Crossref
Palacios Moncayo, O. 1994. Geología de los cuadrángulos de Paita, Piura, Taalra, Sullana, Lobitos, Quebrada Seca, Zorritos, Tombes y Zarumilla, Hojas 11-a, 11-b, 10-a, 10-b, 9-a, 9-b, 8-a, 8-b, 8-c y 7-c. Boletín Ingemmet. Serie A: Carta Geológica Nacional 54: 1–190.
Palmer, A.R. 2009. Animal asymmetry. Current Biology 19: R474–R477. Crossref
Peckmann, J., Goedert, J.L., Thiel, V., Michaelis, W., and Reitner, J. 2002. A comprehensive approach to the study of methane-seep deposits from the Lincoln Creek Formation, western Washington State, USA. Sedimentology 49: 855–873. Crossref
Pelseneer, P. 1889. Sur la classification phylogenetique des pélécypodes. Bulletin scientifique de la France et de la Belgique 20: 27–52.
Pennant, T. 1777. The British Zoology, 4th Edition. 154 pp. Warrington, London. Crossref
Philippi, R.A. 1887. Die tertiären und quartären Versteinerungen Chiles. 266 pp. F.A. Brockhaus, Leipzig. Crossref
Poore, G.C.B. and Griffin, D.J.G. 1979. The Thalassinidea (Crustacea: Decapoda) of Australia. Records of the Australian Museum 32: 217–321. Crossref
Quoy, J.R.C. and Gaimard, J.P. 1834–1835. Voyage de découvertes de l’Astrolabe exécuté par ordre du Roi, pendant les années 1826–1827–1828–1829, sur le commandement de M. J. Dumont d’Urville. J. Tastu, Paris. Zoologie, Mollusques. 3. 1–366 (1834), 367–954 (1835), atlas 107 pls.
Rafinesque, C.S. 1815. Analyse de la nature, ou Tableau de l’univers et des corps organisées. 224 pp. Barraveccia, Palermo. Crossref
Rudman, W.B. 1971. The family Acteonidae (Opisthobranchia, Gastropoda) in New Zealand. Journal of the Malacological Society of Australia 2: 205–214. Crossref
Röding, P.F. 1798. Museum Boltenianum. 199 pp. Johann Christian Trappi, Hamburg.
Saether, K.P., Jingeng, S., Little, C.T.S., and Campbell, K.A. 2016. New records and a new species of bivalve (Mollusca: Bivalvia) from Miocene hydrocarbon seep deposits, North Island, New Zealand. Zootaxa 4154: 1–26. Crossref
Saether, K.P., Little, C.T.S., and Campbell, K.A. 2010a. A new fossil provannid gastropod from Miocene hydrocarbon seep deposits, East Coast Basin, North Island, New Zealand. Acta Palaeontologica Polonica 55: 507–517. Crossref
Saether, K.P., Little, C.T.S., Campbell, K.A., Marshall, B.A., Collins, M., and Alfaro, A.C. 2010b. New fossil mussels (Bivalvia: Mytilidae) from Miocene hydrocarbon seep deposits, North Island, New Zealand, with general remarks on vent and seep mussels. Zootaxa 2577: 1–45.
Saether, K.P., Little, C.T.S., Marshall, B.A., and Campbell, K.A. 2012. Systematics and palaeoecology of a new fossil limpet (Patellogastropoda: Pectinodontidae) from Miocene hydrocarbon seep deposits, East Coast Basin, North Island, New Zealand with an overview of known fossil seep pectinodontids. Molluscan Research 32: 1–15. Crossref
Saint Laurent, M. de 1973. Sur la systématique et la phylogénie des Thalassinidea: définition des familles des Callianassidae et des Upogebiidae et diagnose de cinq genres nouveaux (Crustacea Decapoda). Comptes Rendus Hebdomadaires de Séances de l’Académie des Sciences, série D 277: 513–516.
Saint Laurent, M. de 1979. Sur la classification et la phylogénie des Thalassinides: définitions de la superfamille des Axioidea, de la sous-famille des Thomassiniinae et de deux genres nouveaux (Crustacea Decapoda). Comptes Rendus Hebdomadaires de Séances de l’Académie des Sciences, Paris 288: 1395–1397.
Sakai, K. 1969. Revision of Japanese callianassids based on the variations of larger cheliped in Callianassa petalura Stimpson and C. japonica Ortmann (Decapoda: Anomura). Publications of the Seto Marine Biological Laboratory 17: 209–252. Crossref
Sakai, K. 1988. A new genus and five new species of Callianassidae (Crustacea: Decapoda: Thalassinidea) from northern Australia. The Beagle, Occasional Papers of the Northern Territory Museum of Arts and Sciences 5: 51–69.
Salvini-Plawen, L. 1980. A reconsideration of systematics in the Mollusca (phylogeny and higher classification). Malacologia 19: 249–278.
Sasaki, T., Ogura, T., Watanabe, H., and Fujikura, K. 2016. Four new species of Provanna (Gastropoda: Provannidae) from vents and a seep off Nansei-shoto area, southwestern Japan. Venus 74: 1–17.
Sasaki, T., Okutani, T., and Fujikura, K. 2003. New taxa and new records of patelliform gastropods associated with chemoautosynthesis-based communities in Japanese waters. The Veliger 46: 189–210.
Sasaki, T., Okutani, T., and Fujikura, K. 2008. A new species of Pyropelta (Gastropoda: Pyropeltidae) from hydrothermal vents in the Okinawa trough, southwestern Japan. Journal of Molluscan Studies 74: 309–316. Crossref
Scudder, S.H. 1876. Appendix H9. Report on the Orthoptera collected under the direction of Lieut. G.M. Wheeler, during the season of 1875. Annual Report of the Chief of Engineer’s U.S. Geographical Surveys West of the 100th Meridian. Appendix JJ: 498–515.
Schepman, M.M. 1908–1913. The Prosobranchia of the Siboga Expedition. 352 pp. E.J. Brill, Leyden. Crossref
Schweitzer Hopkins, C. and Feldmann, R.M. 1997. Sexual dimorphism in fossil and extant species of Callianopsis de Saint Laurent. Journal of Crustacean Biology 17: 236–252. Crossref
Smith, E.A. 1885. Report on the Lamellibranchiata collected by H.M.S. Challenger during the years 1873–1876. Reports of the Scientific Results of the Challenger Expedition, Zoology 13: 1–341.
Sowerby, J. de C. 1823–1825. The Mineral Conchology of Great Britain or Coloured Figures and Descriptions of Those Remains of Testaceous Animals or Shells, Which Have Been Preserved at Various Times and Depths in the Earth. Vol. 5. 168 pp. Sherwood, London.
Sowerby, R. 1846. Descriptions of the Tertiary fossil shells from South America. In: C. Darwin (ed.), Geological Observations on the Volcanic Islands and Parts of South America Visited During the Voyage of H.M.S. “Beagle”. 548 pp. Appleton, London.
Squires, R.L. 1995. First fossil species of the chemosynthetic-community gastropod Provanna: Localized cold-seep limestones in Upper Eocene and Oligocene rocks, Washington. The Veliger 38: 30–36.
Squires, R.L. and Goedert, J.L. 1991. New Late Eocene mollusks from localized limestone deposits formed by subduction-related methane seeps, southwestern Washington. Journal of Paleontology 65: 412–416. Crossref
Squires, R.L. and Gring, M.P. 1996. Late Eocene chemosynthetic? bivalves from suspect cold seeps, Wagonwheel Mountain, central California. Journal of Paleontology 70: 63–73. Crossref
Stimpson, W. 1851. A Revision of the Synonymy of the Testaceous Mollusks of New England, with Notes on Their Structure, and Their Geographical and Bathymetrical Distribution. 58 pp. Phillips, Sampson, and company, Boston. Crossref
Valdés, Á. and Camacho-García, Y.E. 2004. “Cephalaspidean” heterobranchs (Gastropoda) from the Pacific coast of Costa Rica. Proceedings of the California Academy of Sciences 55: 459–497.
Valdés, F., Sellanes, J., and D’Elía, G. 2013. Phylogenetic position of vesicomyid clams from a methane seep off central Chile (~36°S) with a molecular timescale for the diversification of the Vesicomyidae. Zoological Studies 51: 1154–1164.
Van Winkle, K. 1919. Remarks on some new species from Trinidad. Bulletins of American Paleontology 8: 19–27.
Warén, A. and Bouchet, P. 1993. New records, species, genera, and a new family of gastropods from hydrothermal vents and hydrocarbon seeps. Zoologica Scripta 22: 1–90. Crossref
Warén, A. and Bouchet, P. 2001. Gastropoda and Monoplacophora from hydrothermal vents and seeps; new taxa and records. The Veliger 44: 116–231.
Warén, A. and Bouchet, P. 2009. New gastropods from deep-sea hydrocarbon seeps off West Africa. Deep-Sea Research I 56: 2326–2349. Crossref
Warén, A. and Ponder, W.F. 1991. New species, anatomy, and systematic position of the hydrothermal vent and hydrocarbon seep gastropod family Provannidae fam. n. (Caenogastropoda). Zoologica Scripta 20: 56–102. Crossref
Woodring, W.P. 1925. Miocene Mollusca from Bowden Jamaica, pelecypods and scaphopods. Carnegie Institution of Washington Publications 336: 1–564.
Vilvens, C. 2001. Description of a new species of Cantrainea (Gastropoda: Turbinidae: Colloniinae) from Guadeloupe. Novapex 2: 153–156.
Von Siebold, C.T.E. 1848. Lehrbuch der vergleichenden Anatomie der wirbellosen Thiere. Erster Theil. In: C.T.E. Von Siebold and H. Stannius (eds.), Lehrbuch der vergleichenden Anatomie, 679. Veit & Comp., Berlin. Crossref
Zhang, S. and Zhang, S. 2017. Description of Pyropelta elongata sp. nov. (Gastropoda, Pyropeltidae) from a methane seep area in the South China Sea. American Malacological Bulletin 35: 51–54. Crossref
Acta Palaeontol. Pol. 65 (1): 109–138, 2020
https://doi.org/10.4202/app.00631.2019