
Early ontogenetic growth stages of Middle Ordovician orthoceratoid cephalopods from Bohemia
MARTINA AUBRECHTOVÁ, VOJTĚCH TUREK, and ONDŘEJ ZICHA
Aubrechtová, M., Turek, V., and Zicha, O. 2020. Early ontogenetic growth stages of Middle Ordovician orthoceratoid cephalopods from Bohemia. Acta Palaeontologica Polonica 65 (3): 575–588.
This study describes 18 specimens representing the earliest growth parts of orthoceratoid cephalopods, most of which have been recently obtained from the Middle Ordovician strata of the Prague Basin, central Bohemia. Only about 20 specimens of Ordovician orthoceratoids displaying embryonic shell are known globally and the Bohemian material thus significantly expands their current record. The studied collection includes twelve specimens assigned to the species Bactroceras sandbergeri and shows that the morphological variability in early ontogenetic growth stages of this species was rather low. The absence of a cicatrix was definitely confirmed in this phylogenetically important cephalopod. Six other specimens investigated herein are indeterminate orthoceratids. Initial chambers, where preserved, are generally large compared to known, stratigraphically younger specimens. This is interpreted here as supporting the previously postulated trend of decreasing the egg- and hatchling sizes in orthoceratoid cephalopods from mid-Ordovician times onwards. Alternatively, the large sizes of these Bohemian orthoceratids may be explained by the position of the Prague Basin in relatively cool-water high palaeolatitudes during the Middle Ordovician.
Key words: Cephalopoda, Orthoceratida, early ontogeny, embryonic shell, Paleozoic, Darriwilian, Bohemia, Czech Republic.
Martina Aubrechtová [aubrechm@gmail.com], Institute of Geology and Palaeontology, Faculty of Science, Charles University in Prague, Albertov 6, 12843 Praha 2, Czech Republic; Institute of Geology of the Czech Academy of Sciences, Rozvojová 269, 165 00, Prague 6-Lysolaje, Czech Republic.
Vojtěch Turek [vojtech.turek@nm.cz], National Museum Prague, Václavské náměstí 68, 115 79 Praha 1, Czech Republic.
Ondřej Zicha [ondrej.zicha@gmail.com], BioLib, z. s., Jugoslávských partyzánů 34, 160 00, Praha 6, Czech Republic.
Received 2 January 2020, accepted 23 March 2020, available online 14 May 2020.
Copyright © 2020 M. Aubrechtová et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.F
Introduction
Knowledge of the morphology of initial chambers and early ontogenetic characters is of key importance for the understanding of phylogenetic pathways and affinities of individual cephalopod groups (see Engeser 1996; de Baets et al. 2012; Klug et al. 2015). However, the embryonic development is insufficiently known in some important groups. This is particularly the case in the phylogenetically earliest (Early and Middle Ordovician) orthoceratoids, a group from which the bactritoids, ammonoids, and coleoids originated (Kröger et al. 2011b; Klug et al. 2015 and references therein). In fact, only a handful of specimens representing apicalmost parts of orthoceratoid phragmocones are known (Table 1). Stratigraphically earliest are Bactroceras angustisiphonatum (Rüdiger, 1889) figured by Evans (2005, uppermost Floian–lowermost Dapingian stages of Svalbard) and Arionoceras? sp. studied and illustrated by Evans (2007, Floian? Stage of Peru). Stratigraphically younger are the Darriwilian Bactroceras sandbergeri (Barrande, 1868) from the Prague Basin, Central Bohemia (originally described as Tretoceras parvulum Barrande, 1874), Orthoceras sp. discussed by Balashov (1957) from the Kunda region of Estonia and Archigeisonoceras sp. described by Kröger (2006) from the Swedish island of Öland.
From the Late Ordovician strata, embryonic shells of orthoceratids have been found more frequently (Table 1). However, the number of recognised species is comparable to that in the Middle Ordovician. Ruedemann (1912) described “half a dozen” juvenile individuals of Orthoceras hudsonicum with preserved embryonic shells (Alplan’s creek, New York, USA). Nine orthoceratid apical parts assigned to three species were reported by Kröger (2007, Hirnantian Stage of Estonia); two years later Kröger et al. (2009) illustrated late Katian embryonic shells of ?Isorthoceras sp. (Indiana Castle Shale, New York, USA) and Orthocerida indet. (Fjäcka Shale, Dalarna, Sweden). And finally, Kröger et al. (2011a) figured apical growth parts in two specimens of the species Transorthoceras osmundsbergense (Hirnantian Stage, Siljan District, Sweden); the authors assumed the species is probably conspecific with Orthoceratidae gen. et sp. indet. B of Kröger (2007).
Table 1. Number of specimens representing earliest growth stages of orthoceratoid cephalopods currently known from the Ordovician strata of Bohemia and other regions. Data after Balashov (1957), Evans (2005, 2007), Kröger (2006, 2007), Kröger et al. (2009, 2011a) and Ruedemann (1912); note that the three specimens figured by Ruedemann (1912) do not constitute the total number of specimens available (asterisked).
|
Ordovician stratigraphy |
Number of specimens with initial chambers per region |
Total |
|||||||
|
Series |
Stages |
Bohemia |
Peru |
Estonia |
Sweden (Öland) |
Sweden (Siljan) |
New York |
Svalbard |
|
|
Upper |
Hirnantian |
|
|
4 |
|
2 |
|
|
6 |
|
Katian |
|
|
|
|
1 |
1+3* |
|
5* |
|
|
Sandbian |
|
|
|
|
|
|
|
0 |
|
|
Middle |
Darriwilian |
19 |
|
1 |
3 |
|
|
|
23 |
|
Dapingian |
|
1 |
|
|
|
|
1 |
2 |
|
|
Lower |
Floian |
|
|
|
|
|
|||
|
Tremadocian |
|
|
|
|
|
|
|
0 |
|
In this paper, we significantly expand the above records by describing 18 more apices of Middle Ordovician orthoceratoids. Two specimens were identified in the historical collections of the National Museum in Prague; the remainder of the material was recently collected by one of the authors (OZ), from an area around the city of Rokycany (Plzeň Region, Western Bohemia) and the territories of Beroun and Prague (Fig. 1).
In the text below, we compare the morphology of the Bohemian specimens with that of other known Ordovician orthoceratoid apices from elsewhere and discuss their relevance for reconstructing reproductive strategies in early Paleozoic orthoceratoids.
Institutional abbreviations.—MWB S, Museum of West Bohemia, Plzeň, Czech Republic; NM L, National Museum, Prague, Czech Republic.
Geological setting, material,
and methods
The specimens studied herein originate from the early–mid Darriwilian Šárka Formation and the late Darriwilian part of the Dobrotivá Formation of the Prague Basin (Bohemia, Fig. 1). The lithology and depositional conditions of both formations, including palaeogeographic position of the Prague Basin during the time of its deposition, is discussed in detail by Aubrechtová and Turek (2018) and references therein.
Eighteen specimens were investigated in this study, sixteen of which originate from the Šárka Formation (Fig. 1). NM L 46566 was collected by František Hanuš in the 1930’s at the locality of Praha-Šárka cihelna (brickyard). NM L 46565 originates from the collection of Celda Klouček from the locality of Praha-Šárka. The remaining specimens were recently collected by OZ in the area of the city of Rokycany at the following localities: Mýto (Svatoštěpánský rybník-Teletník; MWB S 06761, 06765), Mýto (Svatoštěpánský rybník-Kařízek; MWB S 06762), Rokycany-Díly (MWB S 06759, 06763), Těškov (MWB S 06760, 06764), and Volduchy (MWB S 06823–06828); one specimen comes from the locality of Praha-Hanspaulka (NM L 59577).
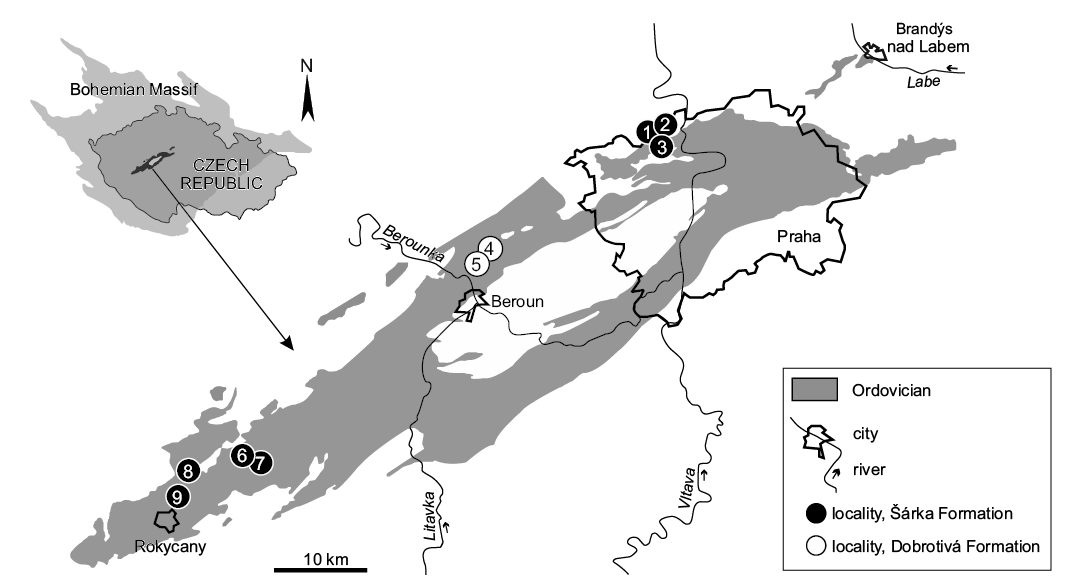
Fig. 1. Position of Ordovician rocks of the Prague Basin within the Bohemian Massif and the localities from which the herein studied specimens originate (see the text below). 1, Praha-Šárka cihelna (brickyard); 2, Praha-Šárka; 3, Praha-Hanspaulka; 4, Malé Přílepy; 5, Lhotka u Berouna; 6, Těškov; 7, Mýto-Svatoštěpánský rybník; 8, Rokycany-Díly; 9, Volduchy. Adapted after Manda (2008).
Two specimens from the Dobrotivá Formation (NM L 59578, 59872) were found by OZ at localities of Lhotka u Berouna and Malé Přílepy (Fig. 1).
All specimens from the Šárka Formation are preserved in 3D in siliceous, originally carbonaceous, nodules (Kukal 1962). The specimens are mostly preserved as counterparts but internal moulds or their fragments are also present. The specimens are not deformed or recrystallized with the exception of NM L 46566, which is entirely in-filled with brown-orange carbonate crystals. NM L 59578 from the Dobrotivá Formation is preserved 3D in a grey-coloured sandy nodule; NM L 59872 is embedded in a dark-grey silicified nodule.
Measurements were performed using either a handheld calliper or an Olympus SZH10 microscope with an integrated scale (angles of expansion of the shells were not precisely measurable in short fragments). Latex casts were prepared from selected counterparts of specimens. Specimens were coated in ammonium chloride (NH4Cl) using the equipment illustrated by Parsley et al. (2018: text-fig. 4), and photographed with Canon EOS 6D and SONY ILCE-7RM3 digital cameras. The specimen of Bactroceras sandbergeri (Barrande, 1868) (= Tretoceras parvulum Barrande, 1874), NM L 10331, with preserved initial chamber was studied for comparisons (see also Dzik 1981 and Aubrechtová 2015).
Systematic palaeontology
Class Cephalopoda Cuvier, 1797
Order Rioceratida King and Evans, 2019
Family Bactroceratidae King and Evans, 2019
Genus Bactroceras Holm, 1898
Type species: Bactroceras avus Holm, 1898, by original designation, junior synonym of B. sandbergeri (Barrande, 1867) according to Aubrechtová (2015); Middle Ordovician, upper Darriwilian Stage, Seby Limestone Formation, Öland, Sweden.
Bactroceras sandbergeri (Barrande, 1867)
Figs. 2–5, 6A.
Lectotype: NM L 6584, figured by Barrande (1867, pl. 245, fig. 18), designated and re-figured by Aubrechtová (2015, p. 202, fig. 2A).
Type locality: Osek u Rokycan, central Bohemia, Czech Republic
Type horizon: Middle Ordovician, lower–middle Darriwilian Stage, Šárka Formation, Corymbograptus retroflexus Biozone.
Material.—Thirteen internal moulds and counterparts of fragments of juvenile shell parts from the localities Mýto (Svatoštěpánský rybník-Teletník, 49.7863217 N, 13.7601539 E; MWB S 06761), Mýto (Svatoštěpánský rybník-Kařízek, 49.7959761N 13.7655044E; MWB S 06762), Praha-Hanspaulka (50.1055886 N, 14.3824408 E; NM L 59577), Rokycany-Díly (49.7569444 N, 13.6013889 E; MWB S 06759, 06763), Těškov (49.7945714 N, 13.6958797 E; MWB S 06760, 06764) and Volduchy (49.77091 N, 13.626269 E; MWB S 06823–06828); Šárka Formation, lower and middle Darriwilian Stage, Middle Ordovician; Czech Republic.
Description.—MWB S 06759 (Fig. 2A) is a part and counterpart of an embryonic portion of the shell with the initial chamber. The total length of the specimen is 2.7 mm. The subspherical initial chamber (= first phragmocone chamber) is bean-shaped in median section, 2.5 mm in diameter and 0.9 mm in length (shape ratio 0.36). The second phragmocone chamber is 2.1 mm in basal diameter and 1.8 mm in length. The diameter of the initial chamber exceeds that of the second phragmocone chamber (constriction ratio 0.84). The phragmocone is circular in cross-section. The marginal (ventral) siphuncle is weathered out and a cavity is left. The cavity appears at about the mid-length of the initial chamber. The caecum is cylindrical, 0.3 mm in diameter; its apical end clearly bends towards the inside of the initial chamber at about its mid-length. The external surface as shown by the latex cast of the counterpart is smooth. The specimen is accompanied by another orthoceratoid shell ornamented with transverse lirae, an indeterminate hyolith, the trilobite Ormathops and the brachiopod Euorthisina sp.
MWB S 06760 (Fig. 2C) is a counterpart of a shell and the third septum of the phragmocone showing the position of the siphuncle. The specimen shows an imprint of the initial chamber. The total length of the specimen is 5.2 mm. The initial chamber (= first phragmocone chamber) was bean-shaped in median section, 2.5 mm in diameter and 1.2 mm in length (shape ratio 0.48). The diameter of the initial chamber exceeds that of the second phragmocone chamber (basal diameter of the second phragmocone chamber was 2.2 mm, constriction ratio 0.88). The phragmocone is circular in cross-section. The specimen is accompanied by the hyolith Bactrotheca teres and other indeterminate invertebrate fragments.
MWB S 06761 (Fig. 3A) is a fragment of a counterpart of a juvenile shell. The total length of the specimen is 13.1 mm. The initial chamber (= first phragmocone chamber) is bean-shaped in median section, 2.2 mm in diameter and 1.1 mm in length (shape ratio 0.5). The diameter of the initial chamber exceeds that of the second phragmocone chamber (basal diameter of the second phragmocone chamber was 2 mm, constriction ratio 0.91). The phragmocone has a maximum diameter of 2.5 mm and expands very slowly (angle of expansion 2°). The phragmocone is circular in cross-section. The external surface is smooth. The specimen is accompanied by a hyolith and other indeterminate invertebrate fragments.
MWB S 06762 (Fig. 3B) is a fragment of a counterpart of a juvenile shell. The total length of the specimen is 20 mm. The initial chamber is bean-shaped in median section, 2.5 mm in diameter and 1.6 mm in length (shape ratio 0.64). The diameter of the initial chamber exceeds that of the second phragmocone chamber (basal diameter of the second phragmocone chamber was 2.3 mm, constriction ratio 0.92). The phragmocone expands very slowly. The phragmocone is circular in cross-section. The external surface is smooth.
MWB S 06763 (Fig. 3C) is a fragment of a counterpart of a juvenile shell with one phragmocone chamber preserved adorally. The total length of the specimen is 10.4 mm. The initial chamber is bean-shaped in median section, 2 mm in dorso-ventral diameter and 1.3 mm in length (shape ratio 0.65). The diameter of the initial chamber exceeds that of the second phragmocone chamber (basal diameter of the second phragmocone chamber is 1.8 mm, constriction ratio 0.9). The phragmocone has a maximum dorso-ventral diameter of 2.3 mm and expands very slowly. The phragmocone is circular in cross-section. The septum of the adoralmost phragmocone chamber displays the submarginal (subventral) siphuncle, which has a circular cross-section and a diameter of 0.25 mm. The external shell surface is smooth. The specimen is accompanied by the bellerophontid Tropidodiscus pusillus, the trilobite Pricyclopyge cf. binodosa and an indeterminate bivalve.
MWB S 06764 (Fig. 2B) is a fragment of a counterpart of a juvenile shell with one phragmocone chamber preserved adorally. The total length of the specimen is 3.5 mm. The initial chamber is bean-shaped in median section, very short, 2.6 mm in dorso-ventral diameter and 1.4 mm in length (shape ratio 0.54). The diameter of the initial chamber exceeds that of the second phragmocone chamber (basal diameter of the second phragmocone chamber was 2.2 mm, constriction ratio 0.85). The phragmocone has a maximum dorso-ventral diameter of 2.2 mm and expands very slowly; it is circular in cross-section. The length of the adoralmost phragmocone chamber is 1.2 mm. The siphuncle is narrow, submarginal (ventral) with a circular cross-section. The external shell surface is smooth. The specimen is accompanied by the bellerophontid Tropidodiscus pusillus.
NM L 59577 (Fig. 2D) is a counterpart of an embryonic shell with the initial chamber. The adoral part of the shell is very slightly bent. The total length of the specimen 4.5 mm. The initial chamber (= first phragmocone chamber) is bean-shaped in median section, 2.0 mm in diameter and 1.2 mm in length (shape ratio 0.6). The diameter of the initial chamber exceeds that of the second phragmocone chamber (basal diameter of the second phragmocone chamber is 1.7 mm, constriction ratio 0.85). The phragmocone is circular in cross-section. No shell ornamentation is discernible. The nodule containing this specimen also yielded the rare graptolite Dichograptus? leptotheca (Petr Kraft, personal communication 2019).
MWB S 06823 (Fig. 6A) represents the third phragmocone chamber (estimated based on the shell diameter and length of this chamber) and the adapical part of the living chamber. The phragmocone has a minimum dorso-ventral diameter of 1.8 mm, a minimum lateral diameter of 1.6 mm and expands very slowly; it is slightly compressed in cross-section. The length of the third phragmocone chamber is 2.7 mm. The siphuncle is narrow, submarginal (ventral) with a circular cross-section; its diameter at the foramen of the adapical septum is 0.25 mm. The specimen is accompanied by undetermined hyoliths and the bellerophontid Tropidodiscus pusillus.
The three remaining specimens MWB S 06824–06826 from the locality Volduchy are juvenile shell fragments that morphologically correspond to the above described specimens. However, their preservation hinders precise measurements.
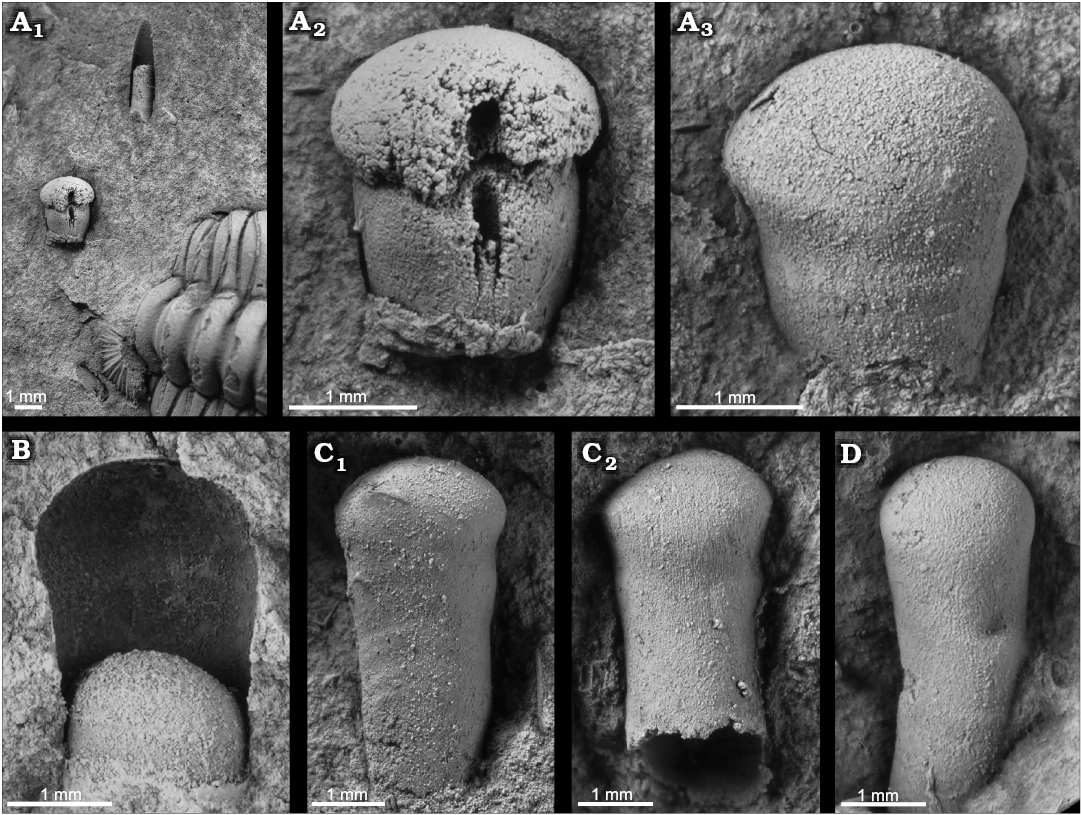
Fig. 2. Apical shell parts of the orthoceratoid Bactroceras sandbergeri (Barrande, 1867), Middle Ordovician, lower Darriwilian Stage, Šárka Formation, Czech Republic. A. MWB S 06759, Rokycany-Díly, internal mould in ventral view (A1), with accompanying trilobite Ormatops sp., brachiopod Euorthisina sp., and indeterminate hyolith; detail (A2), note the decrease of apical angle at second phragmocone chamber; latex cast of counterpart in ventral view (A3). B. MWB S 06764, Těškov, internal mould in lateral view, venter on right hand-side (initial chamber bent towards dorsum), initial and second phragmocone chamber preserved as counterpart. C. MWB S 06760, Těškov, latex casts of both sides of counterparts (C1, C2), note suspicious surface structures. D. NM L 59577, Praha-Hanspaulka, latex cast of counterpart.

Fig. 3. Apical shell of the orthoceratoid Bactroceras sandbergeri (Barrande, 1867); Middle Ordovician, lower Darriwilian Stage, Šárka Formation, Czech Republic. A. MWB S 06761, Mýto-Svatoštěpánský rybník, latex cast of one of counterparts in lateral view, note fine, obliquely transverse surface ornamentation. B. MWB S 06762, Mýto-Svatoštěpánský rybník, counterpart (B1) and its latex cast (B2), in lateral view (initial chamber bent towards dorsum). C. MWB S 06763, Rokycany-Díly, internal mould in lateral view, single (adoralmost) phragmocone chamber showing ventrally situated siphuncle (C1), latex cast of the second counterpart (C2).
Remarks.—MWB S 06759, 06763, and 06764 are shell fragments of larger individuals as indicated by the absence of living chambers. By contrast, specimen MWB S 06823 probably represents the shell of an early juvenile individual. The rest of the specimens (counterparts) do not show any internal structures and therefore can represent both complete shells of freshly hatched individuals, or adapical parts of later growth stages.
The specimens unequivocally belong to Bactroceras sandbergeri (Barrande, 1867) because they exhibit short, bean-shaped in median section, rather large and constricted initial chambers. These characters correspond in all aspects to the only apex previously known from the species (Barrande 1868: pl. 247: 26–28; Barrande 1874: 801; see also Dzik 1981 and Aubrechtová 2015). The only difference concerns the caecum, which is slightly more curved in MWB S 06759 than in Barrande’s specimen mentioned above.
In Bactroceras the apicalmost chamber is known also in B. angustisiphonatum (Rüdiger, 1889) (see Evans 2005), where it is, however, much smaller in size (1.3 mm in height, 1 mm in diameter, shape ratio 1.3), less constricted (constriction ratio 0.95) and more ovoid in shape (Table 2 and Figs. 4, 5).
Of all known Ordovician orthoceratoids, the shape of the initial chamber of Bactroceras sandbergeri is most similar to that of Orthoceras sp. (Balashov 1957; Figs. 4, 5 herein). The latter is, however, much smaller in diameter (see Table 2), the caecum is much shorter and the siphuncle has a ventro-central position. Compared to B. sandbergeri, the initial chamber of Archigeisonoceras sp. (Kröger 2006; Figs. 4, 5 herein) is smaller and only very slightly constricted; the caecum is bent inwards and relatively long but expands inside the initial chamber.
Table 2. Measurements (in mm) and dimensions of specimens of orthoceratoid apical shells parts currently known from the Ordovician strata of Bohemia and other regions. Abbreviations: B., Bactroceras; D, diameter of initial chamber; D1, diameter at 1st septum; L, length of initial chamber; D1/D, constriction ratio (diameter at first septum/diameter of initial chamber); L/D, shape ratio of initial chamber (length of initial chamber/diameter of initial chamber).
|
Stratigraphy |
Locality |
Species (specimen ID) |
References |
D |
L |
L/D |
D1 |
D1/D |
|
|
Lower–Middle |
Floian? Stage, |
Rio Yuscamayo, Cuzco Province, Peru |
Arionoceras? sp. (OUM BU185a) |
1.5 |
1.7 |
1.13 |
1.15 |
0.77 |
|
|
uppermost Floian–lowermost
Dapingian stages, |
North Ny Friesland, Spitsbergen, Norway |
Bactroceras angustisiphonatum (NMW2001.7G.50) |
1.1 |
1.25 |
1.14 |
1.05 |
0.95 |
||
|
Middle Ordovician |
Darriwilian Stage |
Kunda, Estonia |
Orthoceras sp. |
0.95 |
0.55 |
0.58 |
0.8 |
0.84 |
|
|
Darriwilian Stage, |
Island of Öland, Sweden |
Archigeisonoceras sp. (Mo 160705) |
1.5 |
0.9 |
0.6 |
|
0.97 |
||
|
Archigeisonoceras sp. (Mo 160676) |
1.7 |
1 |
0.59 |
|
|
||||
|
Archigeisonoceras sp. (Mo 160678) |
1.6 |
1 |
0.63 |
|
|
||||
|
Darriwilian Stage, |
Central Bohemia, Czech Republic |
orthoceratid sp. 1 (NM L 46566) |
this paper |
3 |
3.1 |
1 |
2.6 |
0.87 |
|
|
orthoceratid sp. 2 (MWB S 06765) |
this paper |
2 |
2 |
1 |
1.9 |
0.95 |
|||
|
orthoceratid sp. 4 (MWB S 06827) |
this paper |
1.8 |
1.3 |
0.72 |
1.5 |
0.83 |
|||
|
Darriwilian Stage, |
orthoceratid sp. 5 (NM L 59872) |
this paper |
1.8 |
|
|
1.8 |
1 |
||
|
Darriwilian Stage, |
B. sandbergeri (NM L 10331) |
2.5 |
1.5 |
0.6 |
2 |
0.8 |
|||
|
B. sandbergeri (MWB S 06759) |
this paper |
2.5 |
0.9 |
0.36 |
2.1 |
0.84 |
|||
|
B. sandbergeri (MWB S 06760) |
this paper |
2.5 |
1.2 |
0.48 |
2.2 |
0.88 |
|||
|
B. sandbergeri (MWB S 06761) |
this paper |
2.2 |
1.1 |
0.5 |
2 |
0.91 |
|||
|
B. sandbergeri (MWB S 06762) |
this paper |
2.5 |
1.6 |
0.64 |
2.3 |
0.92 |
|||
|
B. sandbergeri (MWB S 06763) |
this paper |
2 |
1.3 |
0.65 |
1.8 |
0.9 |
|||
|
B. sandbergeri (MWB S 06764) |
this paper |
2.6 |
1.4 |
0.54 |
2.2 |
0.85 |
|||
|
B. sandbergeri (NM L 59577) |
this paper |
2 |
1.2 |
0.6 |
1.7 |
0.85 |
|||
|
Upper Ordovician |
Katian Stage |
Alplan’s creek, Saratoga County, New York, USA |
Orthoceras
hudsonicum |
Ruedemann 1912: pl. 8: 4 |
1.6 |
1.6 |
1 |
1.28 |
0.8 |
|
Orthoceras
hudsonicum |
Ruedemann 1912: pl. 8: 5 |
1.4 |
1.46 |
1.04 |
1.26 |
0.9 |
|||
|
Orthoceras
hudsonicum |
Ruedemann 1912: pl. 8: 6 |
1.4 |
1.2 |
0.86 |
1.24 |
0.89 |
|||
|
upper Katian Stage, |
New York, USA |
?Isorthoceras sp. (NYSM 17619) |
1.2 |
|
|
|
1 |
||
|
Dalarna, Sweden |
Orthocerida indet (NRM PZ 8874) |
0.7 |
|
|
|
1 |
|||
|
Hirnantian Stage, |
Porkuni quarry, Porkuni, Estonia |
Orthoceratidae indet A (TUG 1227/1) |
1.3 |
0.8 |
0.62 |
|
|
||
|
Orthoceratidae indet B (TUG 1227/2) |
1 |
0.66 |
0.66 |
0.95 |
0.95 |
||||
|
Eriditidae indet (TUG 1227/44) |
0.8 |
0.7 |
0.88 |
0.9 |
1.13 |
||||
|
Eriditidae indet (TUG 1227/47) |
0.9 |
1 |
1.11 |
|
|
||||
Stratigraphic and geographic range.—Šárka Formation, lower and middle Darriwilian, Middle Ordovician of central Bohemia, Czech Republic; middle Darriwilian, Middle Ordovician of Wales, United Kingdom; middle and upper Darriwilian, Middle Ordovician, Oslo Region, Norway; upper Darriwilian, Middle Ordovician, Seby Limestone Formation of Öland, Sweden; Grey Lituites Limestone, Folkeslunda Limestone, upper Darriwilian, Middle Ordovician of Mecklenburg, Germany (Pleistocene glacial erratic boulders).
Bactroceras sp.
Material.—One specimen (NM L 59578), part of a juvenile phragmocone, from the locality Lhotka u Berouna (49.9977650 N, 14.0993506 E); Dobrotivá Formation, upper Darriwilian Stage, Middle Ordovician; Czech Republic.
Description.—The specimen is a fragment of an adapical part of a juvenile phragmocone with visible concave side of one septum. The length of the specimen is 3.5 mm. The phragmocone has a maximum diameter of 2.5 mm and expands at a very low angle (which cannot be precisely measured). The phragmocone is circular in cross-section. The septal foramina of the single visible septum is circular in cross-section, c. 0.1 mm in diameter and submarginal in position. The external surface of the shell is smooth.
Remarks.—The narrow, submarginal siphuncle indicates the specimen should be assigned to the genus Bactroceras. However, the preservation of the specimen does not permit determination to the species level. The specimen represents the first record of Bactroceras from the late Darriwilian Dobrotivá Formation of the Prague Basin.
Order Orthoceratida Kuhn, 1940
Family undetermined
Orthoceratida indet. sp. 1
Fig. 7A.
Material.—One specimen (NM L 46566), phragmocone, from the locality Praha-Šárka cihelna (brickyard); Šárka Formation, lower and middle Darriwilian Stage, Middle Ordovician; Czech Republic.
Description.—The specimen is a part and a counterpart of a phragmocone with the initial chamber. Adapical third of the phragmocone is longitudinally broken off and in-filled with a brownish carbonate mineral; the broken portion was reconstructed using a latex cast prepared from the counterpart. The total length of the specimen is 46.8 mm. A weakly sinuous line along the contact of the fossil and the surrounding rock indicates the presence of a low annulation on the surface of the shell. The surface is, however, slightly corroded and does not show any other ornamentation. The initial chamber is egg-shaped, very slightly asymmetric from one side (Figs. 5, 7A), 3 mm in diameter and 3.1 mm in length (shape ratio c. 1). The diameter of the initial chamber exceeds that of the adjacent phragmocone chamber (basal diameter of the second phragmocone chamber is 2.6 mm, constriction ratio 0.87). The first, straight and directly transverse, suture is faintly indicated. The phragmocone is circular in cross-section with a maximum diameter of c. 7 mm and expands with an angle of 6°.
Remarks.—The initial chamber of the specimen stands out among other apical shell parts known from Ordovician orthoceratids in its large size reaching 3 mm in diameter and 3.1 mm in length. It may resemble the initial chamber described by Evans (2007, Arionoceras?), which is ovoid in shape; the latter is, however, smaller in dimensions, more distinctly constricted (see Table 2 and Figs. 4, 5) and has a greater angle of expansion (13°).
The Bohemian specimen more likely belongs to one of the Middle Ordovician orthoceratid species described by Joachim Barrande, in which early ontogenetic stages are not yet known. Unfortunately, the collection of Joachim Barrande contains either fragmentary material, or specimens representing only late ontogenetic stages. Moreover, the preservation of the specimen studied here does not show the character of the siphuncle and ornamentation is only slightly indicated, which prevents the specimen from being more precisely identified.
Orthoceratida indet. sp. 2
Figs. 5, 7C.
Material.—One specimen (MWB S 06765), phragmocone, from the locality Mýto-Svatoštěpánský rybník-Teletník (49.7863217 N, 13.7601539 E); Šárka Formation, lower and middle Darriwilian Stage, Middle Ordovician; Czech Republic.
Description.—The specimen consists of two counterparts of a phragmocone composed of two dislocated fragments. In each of the two counterparts there is one septum preserved. The rest of the shell was reconstructed from a latex cast. Total combined length is 15.3 mm (apical fragment 9.1 mm and adoral fragment 6.2 mm). The initial chamber is egg-shaped, 2 mm in dorso-ventral diameter and 2 mm in length (shape ratio 1). Its diameter very slightly exceeds that of the adjacent phragmocone chamber (basal diameter of the second phragmocone chamber is 1.9 mm, constriction 0.95). The phragmocone is circular in cross-section. The siphuncle is 0.25 mm in diameter (corresponding shell diameter 23 mm) and sub-central in position (shifted slightly towards the venter, situated 10 mm from the ventral side of the shell wall, 13 mm from the dorsal). The phragmocone has a maximum diameter of 25 mm and expands slowly with an angle of 3°. The convex side of the adoralmost preserved septum shows massive hyposeptal deposits surrounding the siphuncle (see Figs. 5, 7C1). In outline, these bilaterally symmetrical deposits resemble a figure “8”; they are more massive ventrally than dorsally (Figs. 5, 7C1). The external surface of the shell is smooth. The adoral phragmocone fragment shows two straight and slightly oblique suture lines spaced 1.8 mm apart (at a corresponding shell diameter 25 mm). The specimen is accompanied by a small lingulate brachiopod and the bellerophontid Tropidodiscus pusillus.
Remarks.—The initial chamber of MWB S 06765 classified here as Orthoceratida indet. sp. 2 differs from Orthoceratida indet. sp. 1 in being smaller in dimensions, less ovoid and less constricted (Figs. 4, 5). Hyposeptal cameral deposits are present. The only known cephalopod from the Ordovician of Bohemia comparable in shape and position of deposits is Orthoceras bonum Barrande, 1867. However, this species was previously speculated to represent an actinoceratid (Marek 1999; Manda 2008; Evans et al. 2013) or alternatively a pseudorthoceratid (unpublished personal observation of MA and VT), none of which exhibit the initial chamber morphology described above in MWB S 06765.
Orthoceratida indet. sp. 3
Fig. 7B.
Material.—One specimen (NM L 46565), juvenile shell, from the locality Praha-Šárka; Šárka Formation, lower and middle Darriwilian Stage, Middle Ordovician; Czech Republic.
Description.—The specimen is a counterpart of a juvenile shell with only the adoralmost septum preserved; the initial chamber is broken off. Total length is 25 mm. Minimum preserved diameter of the phragmocone is 1.7 mm, the maximum diameter is 4.5 mm; the phragmocone is circular in cross-section. The siphuncle is narrow, 0.7 mm in diameter (at maximum shell diameter) and central in position. The phragmocone expands with an angle of 7°. The external shell surface shows irregular, fine and straight transverse ornamentation; in the adapical and middle parts of the specimen the ornamentation is accentuated and regularly developed.
Remarks.—NM L 46565 has an angle of expansion comparable to Orthoceratida indet. sp. 1 but the former must have had a much smaller initial chamber. In its small size, NM L 46565 resembles Orthoceratida indet. sp. 2 but the latter has a lower angle of expansion. Based on the comparisons to Orthoceratida indet. sp. 2, the initial chamber in NM L 46565 probably did not exceed 1.9 mm in diameter.
Orthoceratida indet. sp. 4
Fig. 6B.
Material.—Two specimens (MWB S 06827, 06828), counterparts of juvenile shell parts from the locality Volduchy; Šárka Formation, lower and middle Darriwilian Stage, Middle Ordovician; Czech Republic.
Description.—MWB S 06827 is a counterpart of the embryonic shell including the initial chamber. The length of the specimen is 3.2 mm. The initial chamber (= first phragmocone chamber) is bean-shaped in median section, 1.8 mm in diameter. The phragmocone is constricted at a distance of 1.3 mm from the tip of the initial chamber and at 1.5 mm of shell diameter. This constriction is assumed to correspond to the position of the first phragmocone septum; the shape ratio of the initial chamber is then 0.72 and constriction ratio 0.83. The phragmocone is circular in cross-section. Longitudinal, faintly sinuous shell ornamentation; it disappears at the adoral part of the specimen where it is replaced by slightly oblique transverse sculpture.
Orthoceratida indet. sp. 5
Fig. 6C.
Material.—One specimen (NM L 59872), embryonic shell, from the locality Malé Přílepy (50.0124397 N, 14.1148514 E); Dobrotivá Formation, upper Darriwilian Stage, Middle Ordovician; Czech Republic.
Description.—The specimen is a counterpart of an embryonic shell. Total length is 3.3 mm. Shell diameter is 1.8 mm along the specimen length; no marked constriction is discernible on the phragmocone. The phragmocone is circular in cross-section. The external shell surface at its apical part shows fine longitudinal structures, while in the adoral part, transverse lirae are observed.
Remarks.—The size and shape of the initial chamber in NM L 59872 is very similar to that in MWB S 06827 assigned above to Orthoceratida indet. sp. 4. The preservation of both specimens is, however, too fragmentary to allow more precise comparisons and unequivocal taxonomic classification.
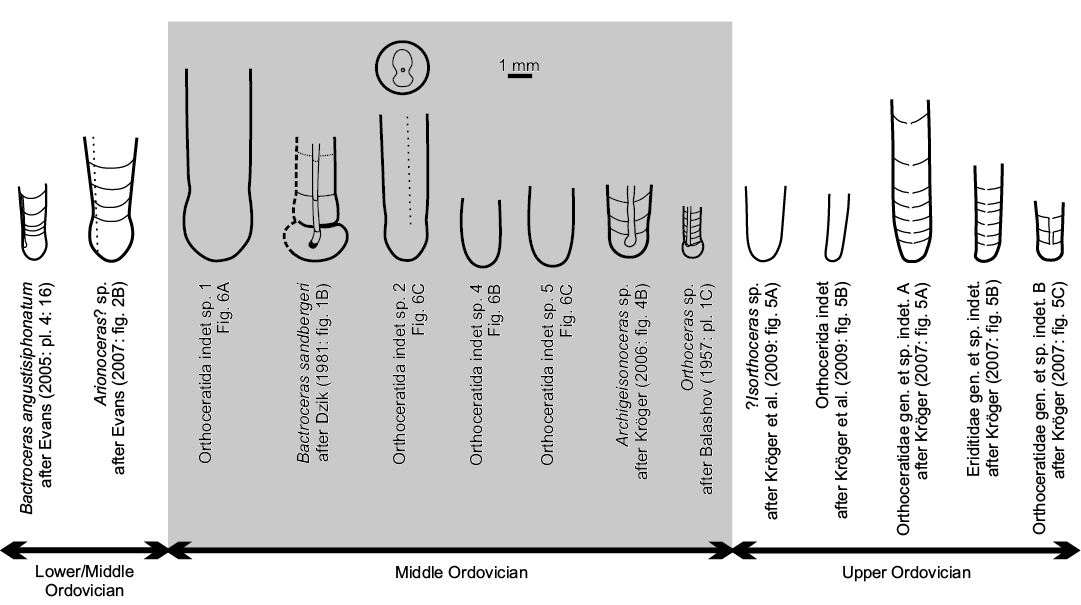
Fig. 5. Drawings of known embryonic shells with initial chambers of Ordovician orthoceratoids (cf. Kröger and Mapes 2007: fig. 4 and Klug et al. 2015: fig. 1.3.). Dotted lines indicate the assumed position of the siphuncle in Arionoceras? sp. and Orthoceratida indet. sp. 2. In Orthoceratida indet. sp. 2, note a sketch of hyposeptal cameral deposits (see the text below and Fig. 7C1). The stratigraphic position of Bactroceras angustisiphonatum and Arionoceras? sp. is either uppermost Lower Ordovician, or lowermost Middle Ordovician (David Evans, personal communication 2019; see also Evans 2005 and 2007). Orthoceratidae gen. et sp. indet B (Kröger 2007) is likely to be conspecific with Transorthoceras osmundsbergense according to Kröger et al. (2011a).
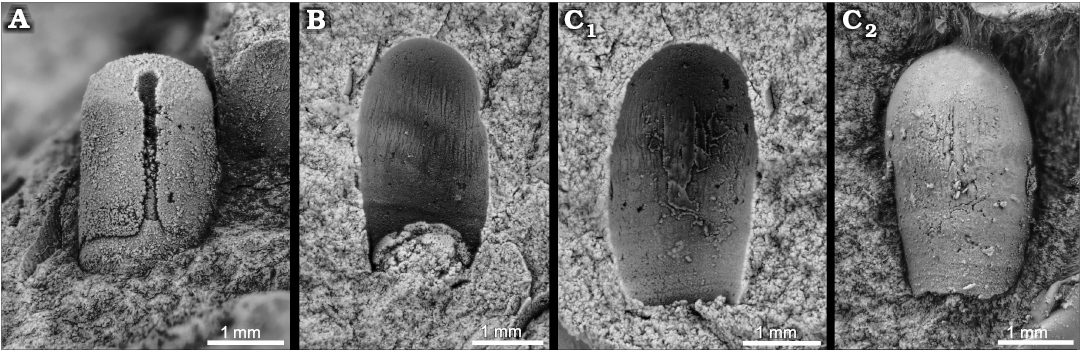
Fig. 6. Apical and juvenile shell parts of orthoceratid cephalopods; Middle Ordovician, Darriwilian Stage, Šárka and Dobrotivá formations, Czech Republic. A. Bactroceras sandbergeri (Barrande, 1867), MWB S 06823, Volduchy, third phragmocone chamber (estimated based on shell diameter) and adjacent part of body chamber. B. Orthoceratida indet. sp. 4, MWB S 06827, Volduchy, counterpart of initial chamber and adjacent part of phragmocone and one corroded phragmocone septum; note longitudinal striae. C. Orthoceratida indet sp. 5, NM L 59872, Malé Přílepy, counterpart (C1), latext cast (C2); note longitudinal striae.
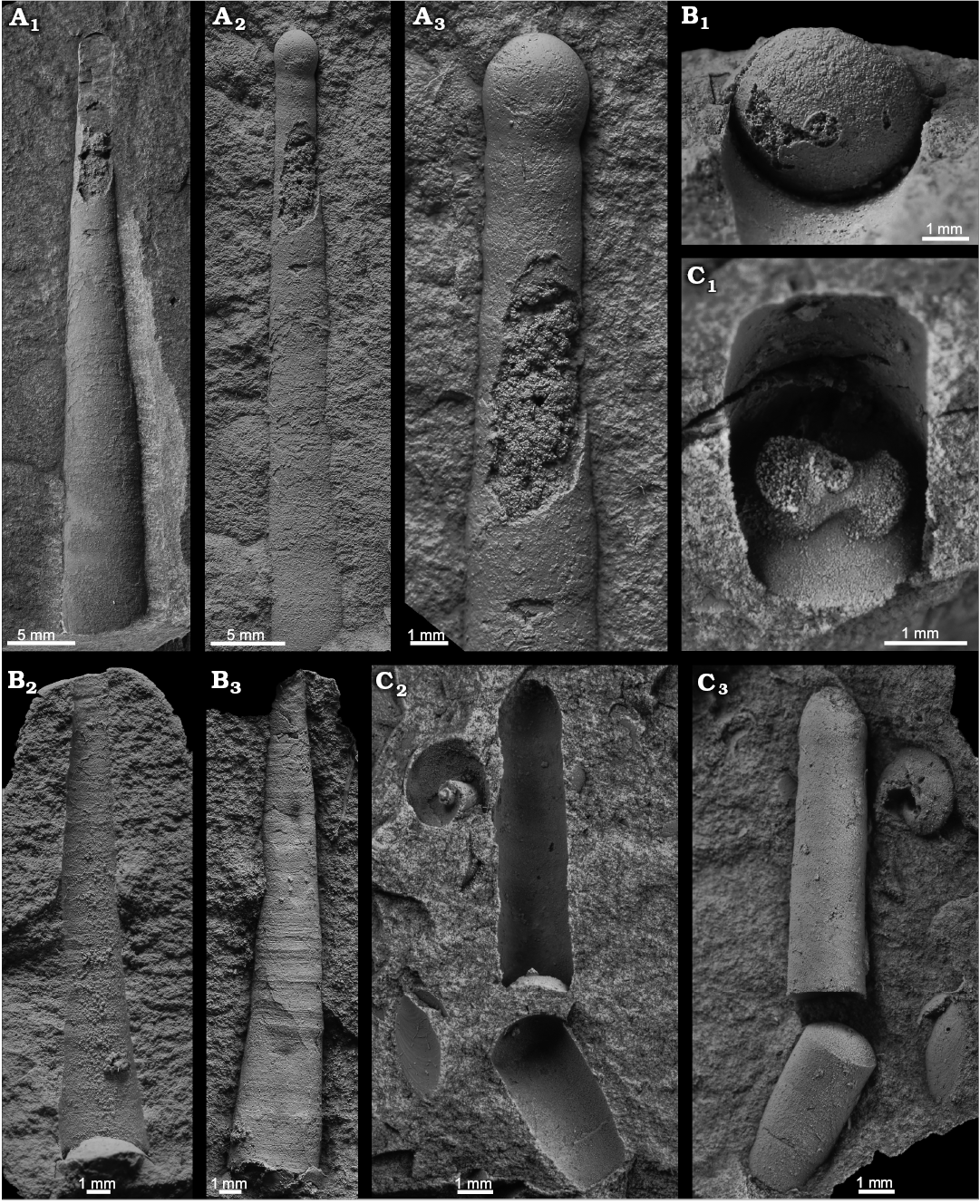
Fig. 7. Apical shell parts of three indeterminate orthoceratid cephalopods; Middle Ordovician, lower Darriwilian Stage, Šárka Formation, Czech Republic. A. Orthoceratida indet. sp. 1, NM L 46566, Praha-Šárka cihelna (brickyard), part (A1) and latex cast of the counterpart (A2), detail of latex cast (A3), note large size of egg-shaped initial chamber. B. Orthoceratida indet. sp. 3, NM L 46565, Praha-Šárka, single (adoralmost) phragmocone chamber showing centrally situated siphuncle (B1), counterpart with one adoral septum (B2), latex cast of counterpart showing transverse surface ornamentation (B3). C. Orthoceratida indet. sp. 2, MWB S 06765, Mýto-Svatoštěpánský rybník, detail of adoralmost septum with hyposeptal cameral deposits around the subcentral siphuncle (C1), the second counterpart (C2), note the absence of cameral deposits at septum situated more apically than in C1, latex cast of the previous associated with bellerophontid Tropidodiscus pusillus (Barrande in Perner, 1903) (C3).
Discussion
Remarks on the early ontogeny in Ordovician orthoceratoids.—Previous studies have focused on general patterns in the embryonic shell size distribution among individual groups of cephalopods and trends in the morphology of these shell parts over longer time periods. The size of embryonic shell has been used to estimate egg and hatchling size (Manda and Frýda 2010). In early Palaeozoic nautiloid cephalopods, the timing of hatching can be determined using the criteria that apply in the Recent Nautilus (except for the nepionic constriction which is not known in early Palaeozoic cephalopods with certainty; e.g., Mapes and Nützel 2009; Turek 2010; Turek and Manda 2016, 2018; Manda and Turek 2019). In contrast, there are varying opinions regarding the recognition of the hatching phase and the size of embryonic shells in orthoceratoid cephalopods. For this purpose, changes in septal spacing, apical angle and ornamentation have been used. Previous studies have concluded that orthoceratoid embryonic shells probably consisted of the initial chamber, one or two phragmocone chambers and the living chamber (Zhuravleva 1961; Kolebaba 1973, 1975, 1977; Engeser 1996). The view assuming a direct ontogenetic development of early orthoceratoids is followed by the present authors, as well. In contrast, Ristedt (1968) and Dzik (1981) supposed that orthoceratoid embryonic shell had consisted only from the first, sub-sphaerical chamber, while the adjacent cylindrical part of the shell was considered to represent the larval stage (for details dealing with this interpretation see Dzik 1981).
With the exception of Bactroceras angustisiphonatum (Evans 2005) and Orthoceras sp. (Balashov 1957), the late Early–Middle Ordovician orthoceratoid initial chambers are relatively large in dimensions (Table 2, Figs. 4, 5, 8); the morphology of adjacent parts of the shell, including the living chamber, imply the presence of relatively large embryonic shells. In contrast to that, Late Ordovician and especially younger, Silurian, orthoceratoid initial chambers and embryonic shells generally have smaller sizes (e.g., Počta 1902, 1907; Schindewolf 1933; Ristedt 1968; Kiselev 1971; Kolebaba 1973, 1975, 1977; Serpagli and Gnoli 1977; Gnoli 1998). This apparent shift in the size of initial chambers and embryonic shells in orthoceratoids from the Middle Ordovician onwards, have been recorded previously (Kröger and Mapes 2007; De Baets et al. 2012; Laptikhovsky et al. 2013, 2018; Klug et al. 2015). It has been assumed that this reflects the gradual decreasing of egg and hatchling size and to indicate changes in reproductive strategies (towards larger amounts of smaller juveniles) and life modes (towards planktotrophy and pelagic lifestyles) in orthoceratoids and their descendants, i.e., bactritoids, ammonoids, and coleoids (e.g., Kröger et al. 2009, 2011b; Laptikhovsky et al. 2013, 2018; Mapes and Nützel 2009; Manda and Frýda 2010; Klug et al. 2015; King and Evans 2019; Fuchs et al. 2020).
However, two Ordovician orthoceratoids, Bactroceras angustisiphonatum (Evans 2005) and Orthoceras sp. (Balashov 1957) contradict the above trend in being considerably smaller in dimensions than other Middle Ordovician orthoceratoids (Table 2, Figs. 4, 5, 8). In both taxa, this may reflect the shell sizes of stratigraphically early orthoceratoids (e.g., Evans 2005). Sea-water temperatures have to be taken into account, as well, and studies on fossil cephalopods have indicated that the cooling of climates/position in high palaeolatitudes induced an increase in the size of embryonic shells. In contrast, warming of climates/position in lower palaeolatitudes was reportedly accompanied by a decrease in these sizes (see Laptikhovsky et al. 2013 and references therein).
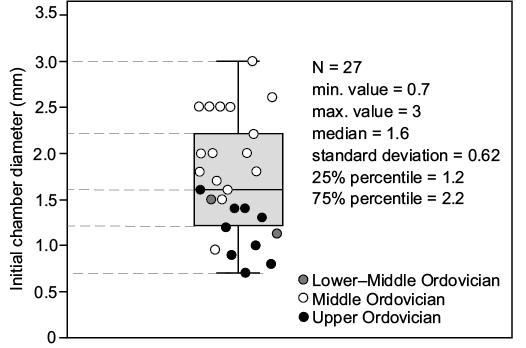
Fig. 8. Box-plot of diameter of initial chambers of Ordovician orthoceratoid cephalopods. Data from Ruedemann (1912), Balashov (1957), Evans (2005, 2007), Aubrechtová (2015), Kröger (2006, 2007), and Kröger et al. (2009).
Early ontogeny in the Bactroceras.—The only initial chamber of Bactroceras sandbergeri previously known was figured by Barrande (1868: pl. 247: 26–28) under the species Tretoceras parvulum Barrande, 1874 (see Dzik 1981; Evans 2005; Aubrechtová 2015). The initial chamber is rather different in appearance from initial chambers of other Ordovician orthoceratoids. Based on the collection of eleven additional, newly obtained specimens it can be confirmed that this is not the result of any deformation or growth anomaly: the initial chamber of B. sandbergeri is sub-sphaerical (bean-shaped in median section), relatively large and short and constricted at the first phragmocone septum. Several well preserved counterparts of initial chambers (Figs. 2, 3) definitely confirm the absence of a cicatrix, a systematically and phylogenetically important feature (Barskov 1963; Engeser 1996; Kröger 2006). These specimens also indicate that the sculpture of the embryonic and early juvenile shell in Bactroceras is smooth but can also exhibit fine longitudinal (Fig. 2C) or obliquely transverse striae (Fig. 3A, B). Unfortunately, Barrande’s original specimen (Barrande 1868) is preserved only as a corroded internal mould and therefore cannot show the character of ornamentation or prove the absence of a cicatrix with certainty.
Two specimens (NM L 10331 and MWB S 06759) preserve the caecum, which is long and tubular, moderately (MWB S 06759) to strongly (NM L 10331) bent laterally and inward. It remains close to the shell wall or in close contact with it throughout its length. The long, inward bent caecum in Bactroceras and also in Archigeisonoceras (Kröger 2006; Figs. 4, 5 herein) may have enhanced buoyancy control and migration potential of the hatchlings (Kröger et al. 2009; Kröger and Evans 2011).
The morphological variability of the initial chamber of Bactroceras is low (Fig. 4). The angle of expansion changes in earliest ontogeny—the shell diameter at the first septum is slightly larger than that at the second septum. The shell starts to expand regularly only at about the mid-way between the second and third septum (NM L 10331, MWB S 06759; Fig. 2A). In some nautiloids (Kolebaba 1973; Manda and Turek 2019) and bactritoids (Kröger and Mapes 2007) the change of juvenile shell expansion indicates the hatching phase. If the same were true for Bactroceras, then its egg would have been ovoid and c. 3 mm in length.
Another interesting fact is the pronounced difference of apex morphology between the two species of Bactroceras in which the early growth stage is known. Besides the contrasting dimensions, the initial shell of B. angustisiphonatum is ovoid in shape and less constricted than the sub-sphaerical initial chamber of B. sandbergeri (see Evans 2005 and Table 2, Figs. 4, 5 herein). Such a difference within a single genus has not yet been documented in exocochleate cephalopods. This may call into question the congenerity of the two species which are otherwise closely similar in other morphological traits of their shells.
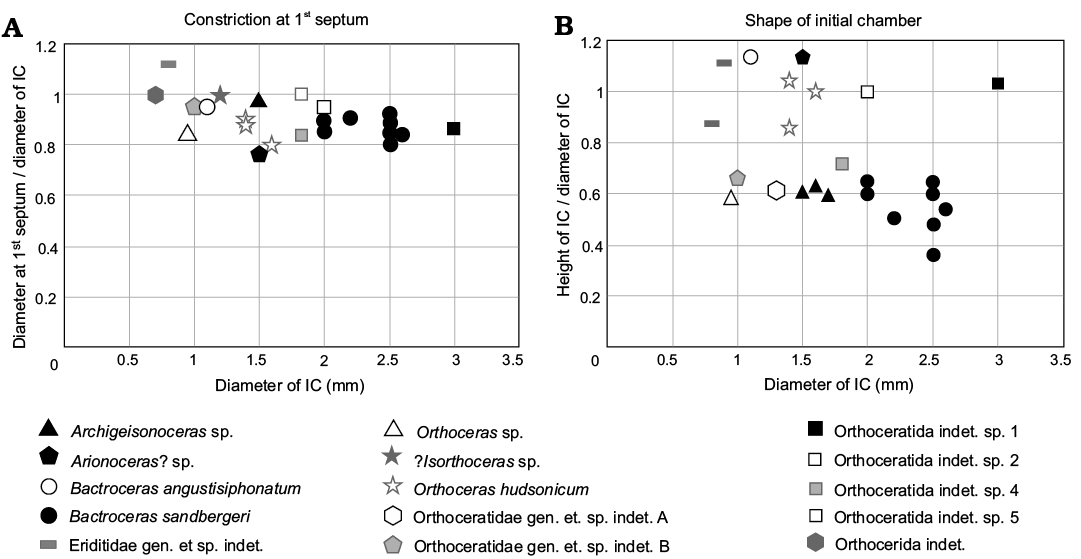
Fig. 4. Scatter plots comparing the shape of initial chambers (A) and the degree of constriction of the embryonic shell (B) in embryonic orthoceratoids known from Ordovician strata of Bohemia and globally. The constriction ratio is lower in embryonic shells that are more constricted at the first septum. The shape ratio is lower in initial shells that are wider than longer in diameter. Abbreviation: IC, initial chamber. See Table 1 for details on measured specimens and references.
Conclusions
The eighteen specimens studied in this paper significantly expand the rather sporadic record of the earliest growth stages of Ordovician orthoceratoids. The initial chamber of Bactroceras sandbergeri, one of the stratigraphically oldest known orthoceratoids, is now known in twelve specimens in total. It is large and very short, sub-sphaerical (bean-shaped in median section) and constricted at the first phragmocone septum. The absence of the cicatrix can now be definitely confirmed. The dimensions of the specimens point to a low morphological variability in both shape and size of juvenile growth stages of the species. The caecum (observed in the type specimen and one additional specimen) is long, tubular and bent inwards (dorsally). A slight decrease in the angle of expansion of the embryonic shell has been observed halfway between the second and third suture, or at the third suture. This may indicate the boundary between embryonic and post-embryonic stages.
The recently obtained specimen of Bactroceras sp. marks the first confirmed occurrence of this genus in the upper Darriwilian Dobrotivá Formation. The specimen thus fills the gap between verified Middle and Upper Ordovician occurrences of Bactroceras in the Prague Basin.
Bactroceras shows a marked intrageneric difference in the shape and size of initial chamber: in B. angustisiphonatum the initial chamber is ovoid and 2–2.5 times smaller compared to that in B. sandbergeri.
Three other orthoceratid specimens described herein are variable in their embryonic shell sizes. Orthoceratida indet. sp. 1 with the diameter of 3 mm and height 3.1 mm is the largest initial chamber known from Ordovician orthoceratids. The initial chamber in Orthoceratida indet. sp. 2 is similar in shape to the former species but is one-third smaller; an adoral septum of the phragmocone shows hyposeptal cameral deposits. Orthoceratida indet. sp. 3 differs from Orthoceratida indet. sp. 2 in the smaller dimensions of its apical shell part and from MWB S 06765 in having a higher angle of expansion. Orthoceratida indet. sp. 4 from the Šárka Formation and Orthoceratida indet. sp. 5 from the Dobrotivá Formation are very similar in the size and shape of their initial chambers; their incomplete preservation, however, hinders the possibility to determine whether the two specimens represent the same species.
The comparably large-sized cephalopod embryonic growth stages with voluminous initial chambers described here from the Darriwilian Stage of Bohemia confirm the presence of a functional phragmocone (i.e., capable of buoyancy regulation) in phylogenetically early orthoceratoids. The initial chambers and reconstructed embryonic shells are mostly larger than those known from the Late Ordovician and especially Silurian strata. This may be a reflection of previously reported shifts in reproductive strategies and modes of life in early Palaeozoic orthoceratoid cephalopods towards gradual decreasing of egg and hatchling sizes, larger amounts of smaller juveniles, juvenile planktotrophy and pelagic lifestyles. Additionally, the influence of palaeoclimatic conditions and sea-water temperatures is implied.
Acknowledgements
We would like to thank Jan Wagner (NM L) for his help with literature search and Lenka Váchová (NM L), who photographed the studied specimens. David Evans (Natural England, Bridgwater, UK) is thanked for his valuable comments and clarifications regarding some previously studied specimens of embryonic orthoceratoids. We are very grateful to Andy H. King (Geckoella, Taunton, UK), Jerzy Dzik (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland), and third anonymous reviewer for their helpful comments and constructive suggestions. This work was financially supported by the Center for Geosphere Dynamics (UNCE/SCI/006) of the Faculty of Science, Charles University, Prague, Research Plan of the Institute of Geology of the Czech Academy of Sciences (RVO67985831) (MA), the Grant Agency of the Czech Republic (GAČR) project no. 18-14575S (MA and OZ) and the Ministry of Culture of the Czech Republic (DKRVO 2020-2023/2.IV.b, National Museum, 00023272) (VT).
References
Aubrechtová, M. 2015. A revision of the Ordovician cephalopod Bactrites sandbergeri Barrande: systematic position and palaeobiogeography of Bactroceras. Geobios 48: 193–211. Crossref
Aubrechtová, M. and Turek, V. 2018. Lituitid cephalopods from the Middle Ordovician of Bohemia and their paleobiogeographic affinities. Bulletin of Geosciences 93: 401–417. Crossref
Balashov, Z.G. [Balašov, Z.G.] 1957. The protoconch of specimen of the genus Orthoceras of the Early Paleozoic [in Russian]. Doklady Akademii Nauk SSSR 116: 855–857.
Barrande, J. 1867. Systême silurien du centre de la Bohême, Iére partie: Recherches Paléontologiques, vol. II, Texte, Classe de Mollusques, Ordre des Céphalopodes. xxxvi + 712 pp. Chez l’auteur et éditeur, Prague.
Barrande, J. 1868. Systême silurien du centre de la Bohême, Iére partie: Recherches Paléontologiques, vol. II, Céphalopodes, 3me Série. xxii pp. + pls. 245–350. Chez l’auteur et éditeur, Prague.
Barrande, J. 1874. Systême silurien du centre de la Bohême, Iére partie: Recherches Paléontologiques, vol. II, Texte, Classe de Mollusques, Ordre des Céphalopodes. xxiv + 804 pp. Chez l’auteur et éditeur, Prague.
Barskov, I.S. 1963. System and phylogeny of the Pseudorthoceratidae [inRussian]. Biulleten moskovskogo obŝestva ispytatelej prirody, Otdel geologičeskij 38: 149–150. Crossref
Cuvier, G. 1797. Tableau élémentaire de l’histoire naturelle des animaux. 710 pp. Baudoin, Paris. Crossref
De Baets, K., Klug, C., Korn, D., and Landman, N.H. 2012. Early evolutionary trends in ammonoid embryonic development. Evolution 66: 1788–1806. Crossref
Dzik, J. 1981. Origin of the Cephalopoda. Acta Palaeontologica Polonica 26: 161–189.
Engeser, T. 1996. The position of the Ammonoidea within the Cephalopoda. In: N.H. Landman, K. Tanabe, and R.A. Davis (eds.), Ammonoid Paleobiology. Topics in Geobiology 13, 3–19. Plenum Press, New York. Crossref
Evans, D.H. 2005. The Lower and Middle Ordovician cephalopod faunas of England and Wales. Monograph of the Palaeontographical Society 628: 1–81.
Evans, D.H. 2007. A Middle Ordovician cephalopod fauna from Cuzco Province, southern Perú and its palaeobiogeographical significance. Geological Journal 42: 25–36. Crossref
Evans, D.H., Ghobadi Pour, M., and Popov, L.E. 2013. Review of the Early to Mid Ordovician orthoconic cephalopods from Iran. Bulletin of Geosciences 88: 21–44. Crossref
Fuchs, D., Laptikhovsky, V., Nikolaeva, S., Ippolitov, A., and Rogov, M. 2020. Evolution of reproductive strategies in coleoid mollusks. Paleobiology 46 (1): 82–103. Crossref
Gnoli, M. 1998. Some remarks and emendation of the family Arionoceratidae Dzik 1984 (Cephalopoda, Nautiloidea). Palaeontologia Electronica 1: 1–5. Crossref
Holm, G. 1898. Om ett par Bactrites-liknande Undersiluriska Orthocerformer. In: G. Holm (ed.), Palæontologiska notiser 179, ser. C, 354–366. Sveriges geologiska undersökning, Stockholm. Crossref
King, A.H. and Evans, D.H. 2019. High-level classification of the nautiloid cephalopods: a proposal for the revision of the Treatise Part K. Swiss Journal of Palaeontology 138: 65–85. Crossref
Kiselev, G.N. [Kiselev, G.N.] 1971. Embryonic shells of Silurian Michelinoceratids [in Russian]. Voprosy Paleontologii 6: 41–51.
Klug, C., Kröger, B., Vinther, J., Fuchs, D., and De Baets, K. 2015. Ancestry, origin and early evolution of ammonoids. In: N. Landman, K. Tanabe, and R.A. Davis (eds.), Ammonoid Paleobiology, 3–24. Plenum Press, New York. Crossref
Kolebaba, I. 1973. Embryonal stages of Wenlockian cephalopods from central Bohemia. Journal of the National Museum (Prague), Natural history series 142: 28–40.
Kolebaba, I. 1975. Caliceras n. gen. and ontogeny of C. capillosum (Barrande) (Nautiloidea). Časopis pro mineralogii a geologii 20: 377–392.
Kolebaba, I. 1977. New information on longitudinally sculptured orthoceroids. Časopis pro mineralogii a geologii 22: 125–138.
Kröger, B. 2006. Early growth-stages and classification of orthoceridan cephalopods of the Darriwillian (Middle Ordovician) of Baltoscandia. Lethaia 39: 129–139. Crossref
Kröger, B. 2007. Concentrations of juvenile and small adult cephalopods in the Hirnantian cherts (Late Ordovician) of Porkuni, Estonia. Acta Palaeontologica Polonica 52: 591–608.
Kröger, B. and Evans, D.H. 2011. Review and palaeoecological analysis of the late Tremadocian–early Floian (Early Ordovician) cephalopod fauna of the Montagne Noire, France. Fossil Record 14: 5–34. Crossref
Kröger, B. and Mapes, R.H. 2007. On the origin of bactritoids (Cephalopoda). Paläontologische Zeitschrift 81: 316–327. Crossref
Kröger, B., Ebbestad, J.O.R., Högström, A.E.S., Frisk, Å.M. 2011a. Mass concentration of Hirnantian cephalopods from the Siljan District, Sweden; taxonomy, palaeoecology and palaeobiogeographic relationship. Fossil Record 14: 35–53. Crossref
Kröger, B., Servais, T., and Zhang, Y. 2009. The Origin and Initial Rise of Pelagic Cephalopods in the Ordovician. PloS ONE 4: e7262. Crossref
Kröger, B., Vinther, J., and Fuchs, D. 2011b. Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules. BioEssays 33: 602–613. Crossref
Kuhn, O. 1940. Paläozoologie in Tabellen. 50 pp. Fischer Verlag, Jena.
Kukal, Z. 1962. A petrographical investigation of the Ordovician Šárka Beds in the Barrandian area. Sborník Ústředního ústavu geologického, oddíl geologický 27: 175–214.
Laptikhovsky, V.L., Nikolaeva, S., and Rogov, M. 2018. Cephalopod embryonic shells as a tool to reconstruct reproductive strategies in extinct taxa. Biological Reviews 93: 270–283. Crossref
Laptikhovsky, V.L., Rogov, M.A., Nikolaeva, S.V., and Arkhipkin, A.A. 2013. Environmental impact on ectocochleate cephalopod reproductive strategies and the evolutionary significance of cephalopod egg size. Bulletin of Geosciences 88: 83–93. Crossref
Manda, Š. 2008. Trocholites Conrad, 1838 (Nautiloidea, Tarphycerida) in the Middle Ordovician of the Prague Basin and its palaeobiogeographical significance. Bulletin of Geosciences 83: 327–334. Crossref
Manda, Š. and Frýda, J. 2010. Silurian/Devonian boundary events and their influence on cephalopod evolution: evolutionary significance of cephalopod egg size during mass extinctions. Bulletin of Geosciences 85: 513–540.
Manda, Š. and Turek, V. 2019. Embryonic and early juvenile development in the Silurian basal nautilid Peismoceras Hyatt, 1894. Swiss Journal of Palaeontology 138: 123–139. Crossref
Mapes, R.H. and Nützel, A. 2009. Late Palaeozoic mollusc reproduction: cephalopod egg-laying behavior and gastropod larval palaeobiology. Lethaia 42: 341–356. Crossref
Marek, J. 1999. Ordovician cephalopods of the Prague Basin (Barrandian area, Czech Republic): a review. Acta Universitatis Carolinae—Geologica 43: 413–416.
Parsley, R.L., Lawson, M.H., and Pojeta, J., Jr. 2018. Apractical and historical perspective on the how and why of whitening fossil specimens and casts as a precursor to their photography. Fossil Imprint 74: 237–244. Crossref
Perner, J. 1903. Systême silurien du centre de la Bohême, Iére partie: Recherches Paléontologiques, Vol. IV., Gastéropodes, Tome Ier, Texte (Patellidae et Bellerophontidae) et Planches (1 à 89). v–xi + 164 pp. R. Gerhard, Lepzig.
Počta, P. 1902. Über die Anfangskammer der Gattung Orthoceras Breyn. Sitzungsberichte der königlichen böhmischen Gesellschaft der Wissenschaften in Prag 52: 1–6.
Počta, F. 1907. Ueber die Anfangskammer von Orthoceras. Centralblatt für Mineralogie, Geologie und Paläontologie 17: 527–528.
Ristedt, H. 1968. Zur Revision der Orthoceratidae. Akademie der Wissenschaften und Literatur in Mainz, Abhandlungen der matematisch-naturwissenschaftlichen Klasse 4: 213–297.
Rüdiger, H. 1889. Über die Silur-Cephalopoden aus den mecklenburgischen Diluvialgeschieben. 86 pp. Inaugural-Dissertation, Philosophischen Fakultät der Universität Rostock, Güstrow. Crossref
Ruedemann, R. 1912. The Lower Siluric shales of the Mohawk Valley. New York State Museum Bulletin 162: 1–123.Crossref
Schindewolf, O.H. 1933. Vergleichende Morphologie und Phylogenie der Anfangskammern tetrabranchiater Cephalopoden. Eine Studie über Herkunft, Stammesentwicklung und System der niederen Ammoneen. Preußischen Geologischen Landesanstalt 148: 67–73.
Serpagli, E. and Gnoli, M. 1977. Upper Silurian cephalopods from southwestern Sardinia. Bollettino della Società Paleontologica Italiana 16: 153–196.
Turek, V. 2010. Embryonic shells in some lechritrochoceratids (Nautiloidea, Barrandeocerida). In: K. Tanabe, Y. Shigeta, T. Sasaki, and H. Hirano (eds.), Cephalopods—Present and Past, 85–92. Tokai University Press, Tokyo.
Turek, V. and Manda, Š. 2016. Early ontogeny, anomalous growth, and healed injuries in the Silurian nautiloid Ophioceras Barrande—Implications for hatching and the autecology of the Tarphycerida. Bulletin of Geosciences 91: 331–366. Crossref
Turek, V. and Manda, Š. 2018. Silurian tarphycerid Discoceras (Cephalopoda, Nautiloidea): systematics, embryonic development and paleoecology. Journal of Paleontology 92 (3): 412–431. Crossref
Zhuravleva, F.A. [Žuravleva, F.A.] 1961. Strojenie embrionalnych rakovin nautiloidej i rodstvennych im grupp [in Russian]. In: V.N. Šimanskij and F.A. Žuravleva (eds.), Osnovnye Voprosy Systematiki Nautiloidei i Rodstvennyh im Grupp. Trudy Paleontologičeskogo Instituta Akademii Nauk 90: 67–100.
Acta Palaeontol. Pol. 65 (3): 575–588, 2020
https://doi.org/10.4202/app.00721.2020