
First record of a Late Jurassic rhamphorhynchine pterosaur from Gondwana
JHONATAN ALARCÓN-MUÑOZ, RODRIGO A. OTERO, SERGIO SOTO-ACUÑA, ALEXANDER O. VARGAS, JENNYFER ROJAS, and OSVALDO ROJAS
Alarcón-Muñoz, J., Otero, R.A., Soto-Acuña, S., Vargas, A.O., Rojas, J., and Rojas, O. 2021. First record of a Late Jurassic rhamphorhynchine pterosaur from Gondwana. Acta Palaeontologica Polonica 66 (3): 571–583.
We describe partial remains of a non-pterodactyloid pterosaur from Upper Jurassic levels of the Atacama Desert in northern Chile. The material includes a left humerus, a possible dorsal vertebra, and the shaft of a wing phalanx, all preserved in three dimensions and likely belonging to a single individual. The humerus has a hatchet-shaped deltopectoral crest, proximally positioned, and its shaft is markedly anteriorly curved, which are characteristic features of the clade Rhamphorhynchidae. In addition, the presence of a groove that runs along the caudal surface of the phalanx, being flanked by two asymmetric crests, is a distinctive feature of the clade Rhamphorhynchinae, which includes such genera as Rhamphorhynchus and Nesodactylus. Previous to this research, known records of Rhamphorhynchinae were restricted to Laurasia; thus, the specimen studied here represents the first evidence of this group found to date in Gondwana. Associated ammonoids allow us to assign the material to a middle Oxfordian age, which makes this specimen the oldest known pterosaur found in Chile, and the first of Oxfordian age in Gondwana. This discovery suggests that the clade Rhamphorhynchidae had a global distribution during the Late Jurassic.
Key words: Pterosauria, Rhamphorhynchidae, Rhamphorhynchinae, Oxfordian, Chile, Laurasia, Gondwana.
Jhonatan Alarcón-Muñoz [alarconmunoz@ug.uchile.cl], Red Paleontológica Universidad de Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile; Laboratorio de Zoología de Vertebrados, Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile; KayTreng Consultores. José Domingo Cañas 1640, DP 1502, Ñuñoa, Santiago, Chile.
Rodrigo A. Otero [otero2112@gmail.com], Red Paleontológica Universidad de Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile; Consultora Paleosuchus Ltda., Huelén 165, Oficina C, Providencia, Santiago, Chile; Museo de Historia Natural y Cultural del Desierto de Atacama. Interior Parque El Loa s/n, Calama, Región de Antofagasta, Chile.
Sergio Soto-Acuña [sesotacu@ug.uchile.cl], Red Paleontológica Universidad de Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile; KayTreng Consultores. José Domingo Cañas 1640, DP 1502, Ñuñoa, Santiago, Chile.
Alexander O. Vargas [alexvargas@uchile.cl], Red Paleontológica Universidad de Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile.
Jennyfer Rojas [jennyfer.rojas@icloud.com] and Osvaldo Rojas [orojas@calamacultural.cl], Museo de Historia Natural y Cultural del Desierto de Atacama. Interior Parque El Loa s/n, Calama, Región de Antofagasta, Chile.
Received 29 August 2020, accepted 8 March 2021, available online 6 September 2021.
Copyright © 2021 J. Alarcón-Muñoz et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Pterosaurs (Diapsida, Archosauria) have been found all over the world (Barrett et al. 2008; Upchurch et al. 2015). Despite of this, their fossil record is very biased both temporally and spatially (Dean et al. 2016). This spatial bias is particularly evident in pterosaurs that lived during the Jurassic period. Most Jurassic pterosaurs have been found in the northern hemisphere, mainly in China, Germany, France, and England (Upchurch et al. 2015), most of these being from the lithographic limestone of the Solnhofen Formation, in Germany, where numerous pterosaur remains have been recovered (Goldfuss 1831; Wellnhofer 1970, 1975a–c). During this time lapse, these territories were part of Laurasia, a supercontinent that included North America and Eurasia (Rage 2016). In contrast, Jurassic pterosaur records from Gondwana are extremely sparse (Gasparini et al. 2004; Codorniú and Gasparini 2013; Codorniú et al. 2016). Most of the Gondwanan Jurassic pterosaurs have been recovered in Argentina, being stratigraphically restricted to Toarcian–Bathonian (Rauhut et al. 2001; Unwin et al. 2004; Codorniú et al. 2010, 2016) and Tithonian (Casamiquela 1975; Codorniú and Gasparini 2013; Codorniú et al. 2016) units of Patagonia, although other records are also known from the lower Jurassic of Antarctica (Hammer and Hickerson 1994), and from the Tithonian of Tanzania (Unwin and Heinrich 1999; Costa and Kellner 2009; Costa et al. 2015). In Chile, the pterosaur record is scarce and fragmentary, with most finds restricted to Lower Cretaceous units (Casamiquela and Chong Díaz 1978; Bell and Suárez 1989; Bell and Padian 1995; Martill et al. 2000, 2006; Soto-Acuña et al. 2015; Alarcón-Muñoz et al. 2018, 2020). There are only two mentions of Jurassic pterosaurs from Chile, both from the Oxfordian Cerro Campamento Formation (Calama, northern Chile). A brief mention of an incomplete skull (MUHNCAL.20148) was referred to an indeterminate non-pterodactyloid pterosaur (Yury-Yáñez et al. 2013); however, subsequent studies and CT-scan of this material show that it belongs to the dentary of a pachycormiform fish (RAO, personal observation). The second find is represented by a partial skeleton of a pterosaur initially referred to Pterosauria indet. by Alarcón et al. (2015). Herein, we present the re-study of the latter specimen after additional preparation. Despite its fragmentary condition, the material preserves diagnostic traits which allow us to refer it to the Rhamphorhynchidae, and even more specifically, within the clade Rhamphorhynchinae, which includes Jurassic pterosaurs from Europe, Asia, and North America (Unwin and Martill 2017). This find represents the first evidence of Rhamphorhynchinae from Gondwana. Along with this, it represents the first pterosaur of Oxfordian age known from Gondwana.
Geological setting
The studied material was collected in Cerro Campamento, in the locality of Cerritos Bayos, ca. 30 km west from Calama, in the Antofagasta Region, Atacama Desert, northern Chile (Fig. 1). The fossil-bearing geological unit is the Cerro Campamento Formation (Duhart et al. 2018). The material was embedded in a concretion of yellowish-to-grey calcareous sandstone, which includes abundant ammonoid remains referable to Subvinialesphinctes profetae Gygi and Hillebrandt, 1991, and less frequent remains referable to Euaspidoceras sp. (Meléndez and Myczyński 1987; Parent 2006). Following Biese (1957, 1961), the fossiliferous concretion-bearing level is set a few meters below distinctive blackish shales that comprise the roof of the section. Biese (1957, 1961) considered these levels as belonging to upper Callovian, based on the presence of “Cosmoceras”. Contrary to this, the abundant remains of Subvinialesphinctes profetae associated to Euaspidoceras sp. and the lack of ammonoids referable to the Kosmoceras (not “Cosmoceras”) suggest a middle Oxfordian age for the concretion-bearing levels and for the material studied here.
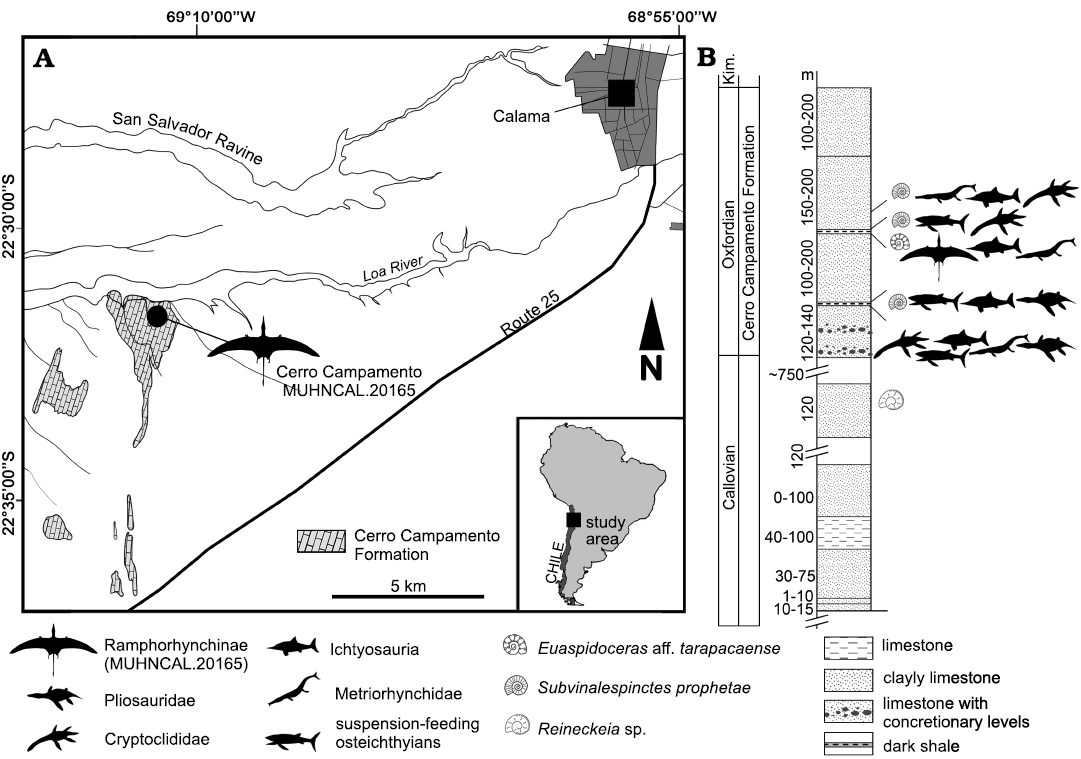
Fig. 1. A. Geographic location of the study area indicating the location of Cerro Campamento, the site where the discovery was made. B. General scheme of the stratigraphic section which includes the Cerro Campamento Formation, indicating horizons with vertebrate remains and the pterosaur materials here studied. Kim., Kimmeridgian. Modified from Biese (1961).
Material and methods
The studied specimen was found by one of the authors (ORM) during fieldwork in 2009. All the bony elements are included in a single concretion. Their occurrence in the same block, the lack of anatomically redundant elements, and the concordance in the relative proportions, suggest that all the bones belong to a single individual. The material was exposed by cracking of the concretion in to three main blocks. Each block was mechanically prepared by JAM and SSA during 2016 and 2017, initially with Dremel Engraver Tool, and during 2018, with PaleoTools ME-9100 airscribe. The bones were consolidated with cyanoacrylate.
Institutional abbreviations.―AMNH, American Museum of Natural History, New York, USA; BSP, Bayerische Staatssammlung für Paläontologie und Historische Geologie, Munich, Germany; D, Dalian Natural History Museum, Dalian, China; MB, Museum für Naturkunde der Humboldt Universität, Berlin, Germany; MJML, Museum of Jurassic Marine Life, Kimmeridge, UK; MUHNCAL, Museo de Historia Natural y Cultural del Desierto de Atacama, Calama, Antofagasta Region, Northern Chile; NHMUK, Natural History Museum, London, UK; OUM, Oxford University Museum, Oxford, UK; PRC, Palaeontological Research and Education Centre, Mahasarakham University, Maha Sarakhaml, Thailand; PSB, Petrefaktensammlung Banz, Upper Franconia, Bavaria, Germany; SMNS, StaatlichesMuseum für Naturkunde, Stuttgart, Germany.
Systematic palaeontology
Diapsida Osborn, 1903
Archosauria Cope, 1869
Pterosauria Kaup, 1834
Breviquartossa Unwin, 2003
Rhamphorhynchidae Seeley, 1870
Rhamphorhynchinae Nopcsa, 1928
Rhamphorhynchinae gen. et sp. indet.
Figs. 2‒6.
Material.―MUHNCAL.20165, a complete left humerus; two fragments of a diaphysis of a wing phalanx and a possible dorsal vertebra belonging to a single, medium sized pterosaur individual from Cerritos Bayos, west Calama, Antofagasta Region, Chile; Cerro Campamento Formation, Zone of Subvinialesphinctes prophetae and Euaspidoceras sp., middle Oxfordian.
Descriptions.—Vertebra: Among the remains there is a single incomplete vertebra (Fig. 2). This is visible in articular view and it is very eroded; it is difficult to stablish whether it is exposed in anterior, posterior, or in an approximately oblique view. The neural spine is tall (preserved height is 6.3 mm), as described for Rhamphorhynchinae (Witton 2013: 129) especially in the cervical vertebrae, although relatively tall and rectancular neural spines are also seen in the anterior dorsal vertebrae of Rhamphorhynchus (see Bonde and Christiansen 2003: fig. 5). The neural spine is laterally compressed and slightly higher than the preserved vertebral centrum. However, we cannot assure that this ratio is natural since the vertebral centrum is not complete. Only one incomplete transverse process is preserved (preserved length of ~4.5 mm). This arises from the top of the neural arch and has a dorsolateral orientation. Proximally, the height of the transverse process represents almost half of the centrum height, narrowing distally. Due to its incompleteness, its extension cannot be established. The neural canal is broad, being wider than high and representing ca. three fifths of the preserved centrum width. The poor preservation of the material makes it difficult to assess the exact shape and proportions of the neural canal, although it appears to have a dorsoventrally compressed oval shape. The centrum is poorly preserved, too. The preserved height is ~8.2 mm. Apparently, its articular contour has a semicircular shape, whereas its dorsal margin was concave, given by the ventral contour of the neural canal. The general morphology of the vertebra resembles that of a dorsal vertebra, but due its the incompleteness cannot be conclusively ascertained.

Fig. 2. Rhamphorhynchinae gen. et sp. indet. (MUHNCAL.20165) from Cerritos Bayos, west Calama, Chile; Cerro Campamento Formation, middle Oxfordian. Vertebrae of indeterminate position (A1) and schematic representation (A2).
Humerus: An almost complete left humerus is preserved. It is three-dimensionally preserved and is exposed in posterior view (Fig. 3). The humerus is 71 mm long, and the mediolateral width of the diaphysis at its midpoint is 7.7 mm, with a length/width ratio of 9.2. The humerus is assigned to a pterosaur based on the posterior inclination of the caput humeri, the presence of a well-expanded anterior deltopectoral crest, and relatively thin cortical bone (Wellnhofer 1978, 1991; Buffetaut et al. 2015). The deltopectoral crest is located at the proximal end of the humerus. The deltopectoral crest is almost complete, dorsoventrally narrow, and well extended anteriorly. It is elongated and projects approximately perpendicular from the humeral shaft. As in several non-pterodactyloid pterosaurs and in some basal pterodactyloids, the proximal and distal margins of the deltopectoral crest lie almost on the same level as the medial crest (Wellnhofer 1978: fig. 1b–d; Unwin and Martill 2017). Furthermore, the deltopectoral crest is proximodistally constricted at its base and expanded anteriorly, in a hatchet-shaped form. At its anterior end, the deltopectoral crest is partially fractured. This portion joins with a carbonate layer present in the rock, making it difficult to determine the exact morphology of the anterior edge of the deltopectoral crest. However, it is observed that the proximodistal edge curves distally, which gives the deltopectoral crest an ax-like shape.
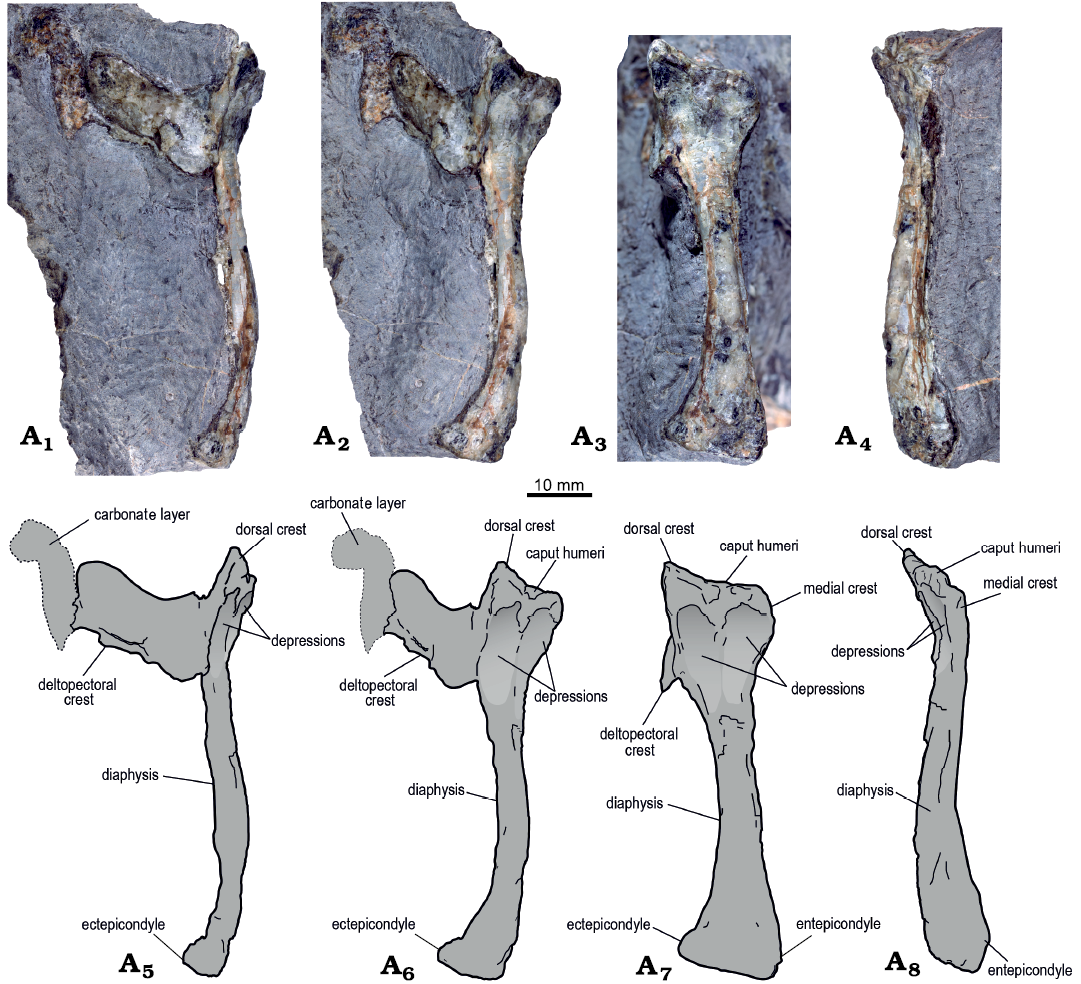
Fig. 3. Rhamphorhynchinae gen. et sp. indet. (MUHNCAL.20165) from Cerritos Bayos, west Calama, Chile; Cerro Campamento Formation, middle Oxfordian. Left humerus in dorsal (A1, A5), posterodorsal (A2, A6), posterior (A3, A7), and ventral (A4, A8) views, photographs (A1–A4) and explanatory drawings (A5–A8).
The hatchet or tongue-shaped form of the deltopectoral crest and its proximal location are features considered diagnostic of Rhamphorhynchidae by Kellner (2003) and Unwin (2003). This morphology is present in several rhamphorhynchids (Fig. 4), such as species of Rhamphorhynchus (Wellnhofer 1975a, 1978; O’Sullivan and Martill 2015), Dorygnathus (Padian 2008b), Nesodactylus (Colbert 1969), Qinglongopterus (Lü et al. 2012), and Bellubrunnus (Hone et al. 2012). The shape of the deltopectoral crest MUHNCAL.20165 especially resembles that present in the genera Rhamphorhynchus and Nesodactylus, in which the hatchet-shaped deltopectoral crest is especially evident (Colbert 1969: fig. 8; Wellnhofer 1975a: fig. 11; Fig. 4E, R). However, the deltopectoral crest of MUHNCAL.20165 is more elongated anteroposteriorly than in Nesodactylus hesperius (Fig. 4). The morphology of the deltopectoral crest of MUHNCAL.20165 is quite different of species of Campylognathoides, in which the deltopectoral crest is short, approximately triangular in shape and its proximal end is proximally curved (see O’Sullivan et al. 2013: fig. 5h, i). The proximal portion of the humerus is dorsoventrally expanded and has two well-marked depressions, limited proximally by the posterior edge of the humeral head, and separated by a medial crest. The caput humeri is partially eroded, and it is flexed posteriorly, as apparently in all pterosaurs (Wellnhofer 1978; Unwin 2003). The proximal portion of the caput humeri has a saddle-shaped articular surface, as is common in pterosaur humeri (Rauhut et al. 2017). Thus, the articular end is concave mediolaterally, and convex anteroposteriorly. The humeral head is slightly displaced posteriorly with respect to the diaphysis axis, a feature apparently present in all pterosaurs (Wellnhofer 1978; Unwin 2003; Unwin and Martill 2017). Its articular surface is partially eroded, although there are two bulges separated by a short groove. The dorsal crest of the humeral head is triangular and well projected proximodorsally, as in species of Rhamphorhynchus (Wellnhofer 1975a) and Nesodactylus (Colbert 1969). The depressions located below the humeral head extend distally until the beginning of the diaphysis. The lateral depression is wider and more excavated than the medial.
Since the humerus is exposed in posterior view, it is not possible to corroborate the presence of pneumatopores piercing the anterior surface of the proximal end of this bone. The absence of pneumatopores has been described in basal pterosaurs (Claessens et al. 2009), such as Rhamphorhynchus muensteri Goldfuss, 1831 (Bonde and Christiansen 2003). Likely, the absence of pneumatic pores in the humerus is a common character among non-pterodactyloids: in Pterodactyloidea, the humeri usually have pneumatopores (Bonde and Christiansen 2003; Claessens et al. 2009; Elgin and Hone 2013; Unwin and Martill 2017). The medial crest (also called ulnar crest) is almost squared with its more medially expanded edge slightly rounded and its distal edge forming an obtuse angle with the diaphysis. This structure corresponds to the area in which the subcoracoscapularis muscle was fixed (Colbert 1969). The medial crest is shorter than in species of Rhamphorhynchus (Wellnhofer 1975a, 1978), Dorygnathus (Padian 2008a), and Nesodactylus (Colbert 1969). The medial crest of MUHNCAL.20165 differs from that of species of Nesodactylus in that it is more extended proximo-distally and its posterior end does not form such an acute angle (Fig. 4). The medial crest of MUHNCAL.20165 also differs from that of Nesodactylus hesperius Colbert, 196; in the latter species the medial extension of the crest is much larger than in MUHNCAL.20165 (see Colbert 1969: fig. 8).
The diaphysis of MUHNCAL.20165 is robust and straight in posterior view but is more gracile than that in the humeri of rhamphorhynchids such as Rhamphorhynchus muensteri (Wellnhofer 1975a, 1978; Fig. 4D, E) and Rhamphorhynchus etchesi O’Sullivan and Martill, 2015 (O’Sullivan and Martill 2015; Fig. 4P). In dorsal and ventral views, the diaphysis of MUHNCAL.20165 is anteriorly curved, as in non-pterodactyloid pterosaurs, although the degree of curvature and extent are variable (Unwin and Martill 2017). The curvature of the shaft of the humerus of MUHNCAL.20165 is well marked (Figs. 3, 4), as in rhamphorhynchids such as species of Rhamphorhynchus (Wellnhofer 1975a, 1978), Dorygnathus (Wellnhofer 1978; Padian 2008b), and Nesodactylus (Colbert 1969), and as in some unidentified rhamphorynchid species from the Taynton Limestone Formation, Stonefield, Oxfordshire, UK (O’Sullivan and Martill 2018). The three-dimensional preservation of the humerus of MUHNCAL.20165 suggests that no relevant taphonomic distortion occurred, which indicates that the anteriorly curved shaft constitutes a distinctive feature of this taxon. In Pterodactyloidea, the shaft of the humerus tends to be straighter (Unwin and Martill 2017). The distal end is quite eroded, which makes it difficult to observe many of its features. Because the humerus is preserved in posterior view, it is not possible to observe the capitulum or the trochlea.
The distal epiphysis is lateromedially expanded, similar to the humerus of Rhamphorhynchus muensteri (Wellnhofer 1975a, 1978; Fig. 4D), of Dorygnathus banthensis Theodori, 1830 (PSB 759a, see Theodori 1852; Padian and Wild 1992; Fig. 4H), and a distal fragment of a humerus of an unidentified rhamphorynchid species from Oxford Clay Formation, England (NHMUK PV R 1995 illustrated in O’ Sullivan 2018: fig. 4). The ectepicondyle is located more proximally than the entepicondyle. The ectepicondyle is dorsally protruding and triangular in shape, whereas the entepicondyle does not protrude ventrally. The ventral half of the distal epiphysis of MUHNCAL.20165 is more distally extended than the dorsal half, whereas in the species of Rhamphorhynchus both portions of the distal epiphysis are approximately at the same level (Wellnhofer 1975a, 1978; Fig. 4D).
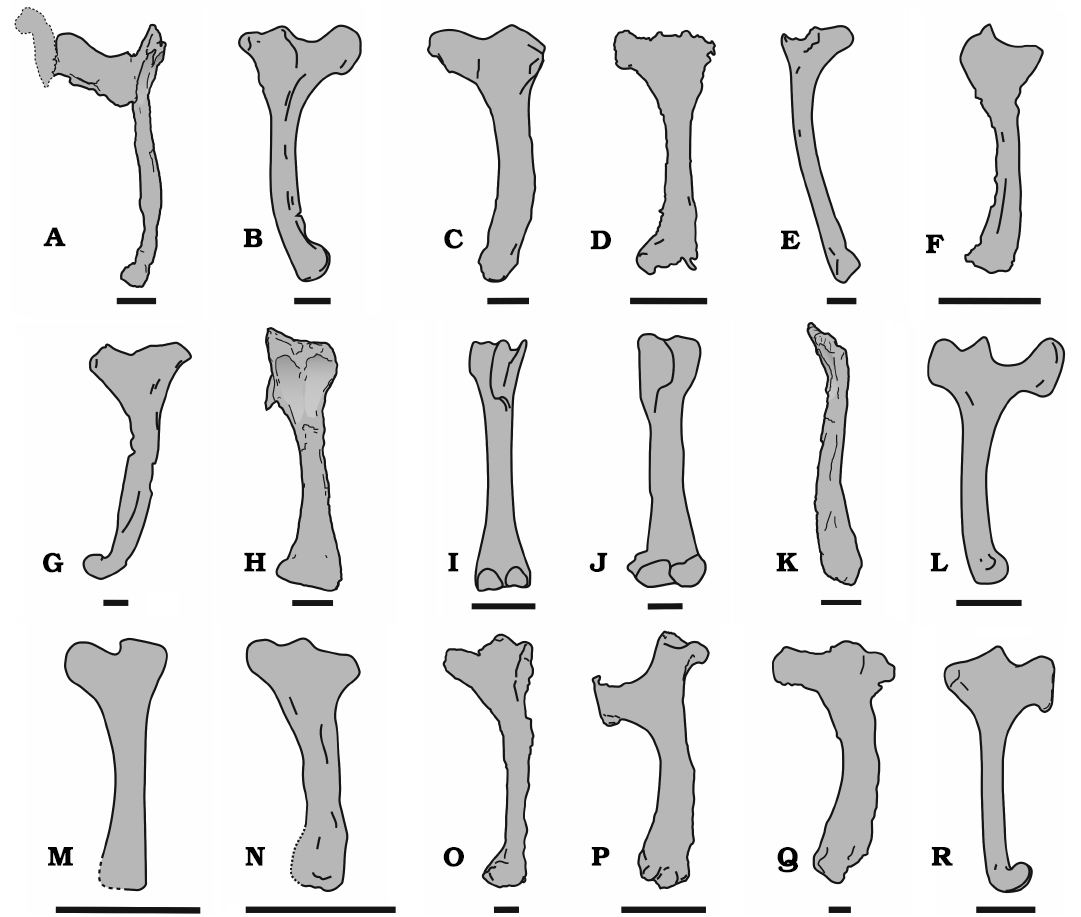
Fig. 4. Comparison between the humeri of Rhamphorhynchinae gen. et sp. indet. (MUHNCAL.20165) and other rhamphorhynchids in dorsal (A–C, E–G), ventral (D, K–R), posterior (H), and anterior (I, J) views. A. Rhamphorhynchinae gen. et sp. indet., left humerus (MUHNCAL.20165). B. Dorygnathus banthensis Theodori, 1830, right humerus (PSB 759a, from Padian and Wild 1992). C. Dorygnathus banthensis Theodori, 1830, left humerus (MB 1905.15, from Unwin and Martill 2017). D. Rhamphorhynchidae indet. right humerus (NHMUK PV R 40126b, from O’Sullivan and Martill 2018). E. Rhamphorhynchidae indet., right humerus (NHMUK PV R 40126c, from O’Sullivan and Martill 2018). F. Rhamphorhynchidae indet., left humerus (NHMUK PV R 28160a, from O’Sullivan and Martill 2018). G. Rhamphorhynchidae indet., left humerus (OUM J.23043, from O’Sullivan and Martill 2018). H. Rhamphorhynchinae gen. et sp. indet., left humerus (MUHNCAL.20165). I. Rhamphorhynchus muensteri Goldfuss, 1831, left humerus from Wellnhofer 1975a: 23, fig. 11e). J. Dorygnathus banthensis Theodori, 1830, right humerus (SMNS 50164, from Witton 2015). K. Rhamphorhynchinae gen. et sp. indet., left humerus (MUHNCAL.20165). L. Rhamphorhynchus muensteri Goldfuss, 1831, left humerus from Wellnhofer 1975a: 23, fig. 11d). M. Qinglongopterus guoi Lü, Unwin, Zhou, Gao, and Shen, 2012, left humerus (D3080/1, from Lü et al. 2012). N. Bellubrunnus rothgaengeri Hone, Tischlinger, Frey, and Röper, 2012, left humerus (BSP–1993–XVIII–2, from Hone et al. 2012). O. Sericipterus wucaiwanensis Andres, Clark, and Xing, 2010, right humerus (IVPP V14725, from Andres et al. 2010). P. Rhamphorhynchus etchesi O’Sullivan and Martill, 2015, right humerus (MJML K-1597, from O’Sullivan and Martill 2015). Q. Rhamphorhynchinae gen. et sp. indet., right humerus (PRC 64, from Buffetaut et al. 2015). R. Nesodactylus hesperius Colbert, 1969, left humerus (AMNH 2000, redraw from Colbert 1969). Scale bars: A–C, E–M, O–R, 10 mm; D, N, 5 mm.
Wing phalanx: The diaphysis of a wing phalanx is preserved in two fragments (Fig. 4). The close association of the phalanx with the left humerus suggests that both elements could correspond to the same wing, however, the incomplete preservation of the phalanx fragments makes it difficult to ascertain. The pterosaurs generally have four phalanges in the wing finger which vary in length, morphology of the epiphyses and thickness (Wellnhofer 1991; Unwin 2006).
The wing phalanx of MUHNCAL.20165 is robust, as in Rhamphorhynchus muensteri (see Wellnhofer 1975a: fig. 15). The total length of the wing phalanx (corresponding to the sum of both fragments) is 161.7 mm (85.6 and 76.1 mm, respectively). The preserved length of the wing phalanx of MUHNCAL.20165 is greater than the length of the first wing phalanx of Sericipterus wucaiwanensis Andres, Clark, and Xing, 2010, corresponding to 127.4 mm (Andres et al. 2010). The preserved length of the wing phalanx of MUHNCAL.20165 is less than that of wing phalanges I–III of Rhamphorhynchus etchesi, corresponding to 171, 175, and 163 mm, respectively (O’Sullivan and Martill 2015). However, it is possible that the complete wing phalanx of MUHNCAL.20165 was similar or even longer than that of at least one of the first three wing phalanges of R. etchesi. In addition, the preserved length of the wing phalanx of MUHNCAL.20165 is less than that of one of the longest wing phalanx described in Rhamphorhynchus muensteri, which measures about 180 mm (see Bennett 1995: fig. 1). However, as in the case of R. etchesi, it is not possible to determine with certainty whether the length of the complete phalanx of MUHNCAL.20165 was indeed less than the length of the longest wing phalanges of R. muensteri. The maximum anteroposterior width of the phalanx is 9.1 mm, and the maximum dorsoventral height is 8.3 mm. The ratio between the anteroposterior width of the mid-point of the diaphysis of the humerus and that of the wing phalanx is 0.85. This value is slightly less than in Jianchangopterus zhaoianus Lü and Bo, 2011 (which is the widest wing phalanx of this pterosaur) corresponding to 0.96 (Lü and Bo 2011). The preserved length of the phalanx in relation to the preserved length of the humerus does not support correspondence to one of the two most distal phalanges (III and IV) since these tend to be much more gracile and shorter (especially the terminal phalanx) in relation to the humerus than the two more proximal ones, as is described in Qinglongopterus guoi Lü, Unwin, Zhou, Gao, and Shen, 2012 (Lü et al. 2012). In one specimen of R. muensteri, Hone et al. (2015) mention that the wing phalanges I and II are slightly longer than the phalanges III and IV, but the latter two are more gracile. Thus, the preserved shaft probably corresponds to the first or second wing phalanx. Unfortunately, none of these fragments preserve proximal or distal ends, making it difficult to assess the complete morphology and the whole phalanx length, and therefore, precluding accurate comparisons with other pterosaurs.
In the wing phalanx of MUHNCAL.20165, there is a longitudinal furrow which is flanked by two asymmetric crests, of which the dorsal one is more prominent than the ventral one (Fig. 5). The presence of this furrow is considered a diagnostic feature of Rhamphorhynchinae (Unwin 2003). This trait was described in the rhamphorhynchines Nesodactylus hesperius of the Oxfordian of Cuba (Colbert 1969: fig. 10, modified from Gross 1937), R. muensteri from the Tithonian of Germany (Colbert 1969: fig. 10, modified from Gross 1937; see Wellnhofer 1978: fig. 13) and R. etchesi from the Tithonian of England (O’Sullivan and Martill 2015: figs. 4, 9). In R. muensteri and R. etchesi, the fourth wing phalanges present the furrow on its caudal surface, which covers practically all of its extension. In addition, this furrow can be clearly seen in the first wing phalanx of an indeterminate Rhamphorhynchinae from the Oxford Clay Formation (NHMUK PV R 4759, O’Sullivan 2018: fig. 5).

Fig. 5. Rhamphorhynchinae gen. et sp. indet. (MUHNCAL.20165) from Cerritos Bayos, west Calama, Chile; Cerro Campamento Formation, middle Oxfordian. Fragments of a wing phalanx in dorsal (A1) and posterior (A2) views.
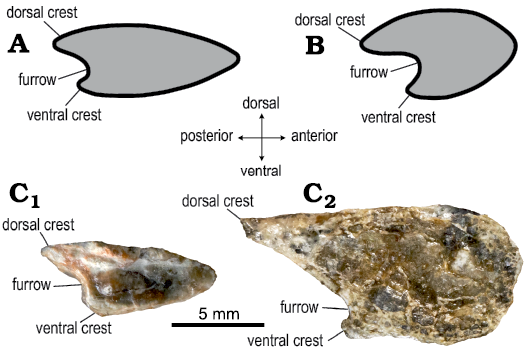
Fig. 6. Comparison of sections of the wing phalanx of rhamphorhynchinae pterosaurs. A. Nesodactylus hesperius Colbert, 1969 (modified from Colbert 1969). B. Rhamphorhynchus muensteri Goldfuss, 1831 (modified from Colbert 1969). C. Ramphorhynchinae gen. and sp. indet. MUHCAL.20165, Cerritos Bayos, west Calama, Chile; Cerro Campamento Formation, middle Oxfordian. C1 and C2, cross sections of two portions of the wing phalanx. A and B, not to scale.
The cross-section is similar to that of R. muensteri and N. hesperius, as illustrated in Colbert (1969: fig. 10) and this work (Fig. 5). Its area is variable along the diaphysis, and we interpreted the larger area as the most proximal portion of the phalanx. As in the first wing phalanx of N. hesperius (Colbert 1969: fig. 10) and R. muensteri (Gross 1937), the anterior surface of the wing phalanx of MUHNCAL.20165 is partially sharp in what we interpret as the distal portion of the phalanx, becoming more rounded in what we interpret as the proximal portion, as in the first wing phalanx section of R. muensteri illustrated in Gross (1937).
The only pterosaur outside of Rhamphorhynchinae in which the presence of a longitudinal furrow in a wing phalanx has been described is in the dimorphodontid Caelestiventus hanseni Britt, Dalla Vecchia, Chure, Engelmann, Whiting, and Scheetz, 2018, of the Upper Triassic from North America (Britt et al. 2018). This furrow was described in the terminal phalanx of this pterosaur (the only preserved postcranial element of this species). However, it is not known whether the proximal phalanges of C. hanseni had a longitudinal furrow. Furthermore, the age at which this species (and the Dimorphodontidae family) lived together along with other anatomical differences between the humeri of dimorphodontids and rhamphorhynchids do not support the inclusion of MUHNCAL.20165 in Dimorphodontidae (see discussion on this issue).
Discussion
Morphological features.―The studied material shows characters typical of pterosaurs, such as a well-developed wing-like deltopectoral crest, the posterior flexion of the humeral head with respect to the axis of the diaphysis, and extremely thin walls of the appendicular bones (Wellnhofer 1978, 1991; Kellner 2003; Unwin 2003, 2006; Buffetaut et al. 2015; Unwin and Martill 2017). Although the available bones are scarce, they have some features present in pterosaurs of the clade Rhamphorhynchidae. According to the definition of Unwin (2003), Rhamphorhynchidae includes Sordes pilosus, Rhamphorhynchus muensteri, their most recent common ancestor, and all its descendants, and is composed of the subclades Rhamphorhynchinae and Scaphognathinae. However, the phylogenetic relationships of the clade Rhamphorhynchidae are currently controversial, so the inclusion of several species in Rhamphorhynchidae is debated (Andres et al. 2014; Vidovic and Martill 2018; Baron 2020). The deltopectoral crest of the humerus of MUHNCAL.20165 suggest affinities of this specimen with Rhamphorhynchidae. In the humerus, Unwin (2003) describes the deltopectoral crest of the rhamphorhynchid pterosaurs as “tongue shaped”, while Kellner (2003) describes it as “hatchet-shaped”, since this structure is anteriorly expanded and narrowed to the base, although it should be noted that the shape of the deltopectoral crest changed along ontogeny (see Wellnhofer 1975a: fig. 11). Furthermore, both Kellner (2003) and Unwin (2003) add that the deltopectoral crest in Rhamphorhynchidae is located proximally, unlike Nyctosauridae, which present a deltopectoral crest with a similar morphology, but it is displaced distally. According to Unwin (2003), this morphology is especially evident in R. muensteri, although it is also observed in Dorygnathus banthensis and Nesodactylus hesperius. Although the deltopectoral crest in MUHNCAL.20165 is incomplete, its general shape and proximal location clearly agree with that described in Rhamphorhynchidae. Added to this is the marked anterior curvature of the diaphysis, which is a common feature in humeri of rhamphorhynchids (Wellnhofer 1975a, 1978; Unwin 2003; Kellner 2003; Hone et al. 2012; Lü et al. 2012; Unwin and Martill 2017)
Other informative features of MUHNCAL.20165 is the presence of a caudal longitudinal furrow of the wing phalanx. A caudal furrow like that present in the phalanx of MUHNCAL.20165 has been recorded exclusively in two pterosaur lineages, the Rhamphorhynchinae (Wellnhofer 1975a, b, 1978; Unwin 2003; O’Sullivan and Martill 2015; O’Sullivan 2018) and Dimorphodontidae (Britt et al. 2018). Unwin (2003) considered the presence of a furrow that runs along the caudal surface of the wing phalanges as a diagnostic feature of Rhamphorhynchinae. This feature has been described in the rhamphorhynchines as R. muensteri, R. etchesi (Wellnhofer 1978; O’Sullivan and Martill 2015) and N. hesperius (Colbert 1969). Additionally, some isolated wing phalanges with a caudal longitudinal furrow have been referred to Rhamphorhynchinae based on this trait (O’Sullivan 2018). Regarding Dimorphodontidae, this feature has been recognized in a single species, Caelestiventus hanseni, from the Upper Triassic of North America, which bears a similar caudal longitudinal furrow (Britt et al. 2018). However, the presence of a furrow in the proximal phalanges of this species is unknown. Further, the clade Dimorphodontidae so far is restricted to the Late Triassic and Early Jurassic, while Rhamphorhynchinae is restricted to the Late Jurassic (Witton 2013). In addition, other dimorphodontids such as Dimorphodon macronyx Buckland, 1829, completely lack furrows on the caudal surface of the wing phalanges (Padian 1983; Britt et al. 2018). Finally, the morphology of the humerus MUHNCAL.20165 associated with the phalanx, specially of the deltopectoral crest, does not agree with that of dimorphodontids, since the deltopectoral crest in these pterosaurs is not hatchet or tongue shaped (see Wellnhofer 1978: fig. 10). Considering this evidence, the morphology and position of the deltopectoral crest, the presence of a caudal furrow in the wing phalanx of MUHNCAL.20165 plus the Oxfordian age of this specimen, support its adscription to the clade Rhamphorhinchinae.
Paleobiogeography.—The clade Rhamphorhynchidae is well represented in Upper Jurassic rocks of former Laurasia. In contrast, specimens referred to this clade are much scarcer in the territories that were part of Gondwana (Barret et al. 2008). To date, specimens referred to Rhamphorhynchidae have been reported from the Middle and Upper Jurassic of India (Reck 1931; Rao and Shah 1963; Colbert 1969; Jain 1974; Unwin and Heinrich 1999; Barrett et al. 2008; Padian 2008a), Upper Jurassic of Tanzania (Reck 1931), and the Lower–Middle Jurassic of Argentinean Patagonia (Rauhut et al. 2001; Unwin et al. 2004; Codorniú et al. 2010, 2016). All these records were later reassigned to other groups of pterosaurs, and even to other groups of vertebrates (Table 1). However, recently, isolated teeth from the Bathonian Guelb el Ahmar in Morocco have been assigned to indeterminate rhamphorhynchids (Haddoumi et al. 2016). Additionally, an isolated tooth from Upper Jurassic–Lower Cretaceous of Ksar Metlili, Morocco, was tentatively assigned to Rhamphorhynchidae (Lasseron et al. 2020).
Table 1. Summary of findings of skeletal remains from Gondwana initially attributed to Rhamphorhynchidae, and their subsequent identifications.
|
Specimen |
Original identification |
Current taxon |
Anatomic elements |
Litostratigraphic unit |
Age |
Country |
References |
|
MB.R.2845 |
Rhamphorhynchus tendagurensis Reck, 1931 |
Pterosauria indet. |
incomplete radius and ulna |
Tendaguru Formation |
Kimmeridgian– |
Tanzania |
|
|
N° K 49/884 |
Rhamphorhynchus sp. |
Pterosauria indet. |
postcranial remains |
Kota |
Middle–Late Jurassic |
India |
|
|
I.S.I.R.48 |
Campylognathoides indicus Jain, 1974 |
Osteichthyes indet. |
left and right premaxillae |
Kota |
Middle–Late Jurassic |
India |
|
|
MPEF PV 3607, 3609, 3613, 3615 |
Rhamphorhynchidae indet. |
Allkaruen
koi |
mandible, braincase, and cervical vertebrae |
Cañadón Asfalto |
Toarcian– Bathonian |
Argentina |
The partial pterosaur skeleton from Cerro Campamento Formation constitutes the first and only representative of the clade Rhamphorhynchinae known to date from Gondwana. In addition, pterosaurs of Oxfordian age are globally scarce, and mostly restricted to Laurasia (Lydekker 1890; Sharov 1971; Colbert 1969; Buffetaut et al. 1985; Buffetaut and Guilbert 2001; Gasparini et al. 2004; Barret et al. 2008; Lü et al. 2012; O’Sullivan 2018). Therefore, the new austral record sheds light on a broader geographic distribution of rhamphorhynchines during that lapse. This may not be unexpected, given the volant capabilities of Pterosauria, and the intermittent land connectivity between Laurasia and Gondwana during the Middle and Late Jurassic.
The subsistence of most Rhamphorhynchidae was probably tied to marine and coastal ecosystems. The recovery of MUHNCAL.20165 from marine units is consistent with the fact that most known rhamphorhynchine specimens have been found in rocks formed from sediments deposited in shallow marine environments (Newton 1888; Wellnhofer 1975c; Gasparini et al. 2004; O’Sulivan and Martill 2015; although some rhamphorhynchines from China are preserved in continental rocks: He et al. 1983, Czerkas and Ji 2002; Andres et al. 2010; Lü et al. 2012). In addition, specimens of Rhamphorhynchus muensteri from Solnhofen Formation show preservation of gut contents corresponding to fish scales and a whole fish individual (Frey and Tischlinger 2012; Witton 2018), suggesting that fish may have been one of the most important food sources for these pterosaurs.
During the Oxfordian, the coasts of Gondwana and Laurasia were continuous not only through the western and eastern margins of these supercontinents, but also at the margins of internal seaways such as the Caribbean corridor and the Trans-Erythrean corridor. The fully functional Caribbean Corridor (Smith 1983; Iturralde-Vinent 2006) represents the shortest paleogeographical distance between Chile and Laurasia throughout which similar coastal ecosystems may have existed. In this regard, the Caribbean corridor has been proposed as an explanation for faunal similarities of marine deposits among Europe (UK, Germany), Cuba and South America during the Oxfordian (Fig. 7), as accounted by diapsids (Gasparini and Iturralde-Vinent 2006; Gasparini 2009; Otero et al. 2020) and bony fishes (Arratia 1994, 2008). Coeval ramphorhynchids from marine deposits of the Caribbean Corridor at Cuba (particularly Nesodactylus hesperius, Colbert 1969) are therefore of special interest for comparison to Chilean forms. Hopefully, better materials from Chile will allow a more detailed assessment of any special affinities. While discussion of such matters requires further evidence, the new material from Chile is already sufficient to document a broader distribution of Rhamphorhynchidae beyond Laurasia. They prove this clade had a global distribution and increase the knowledge about the diversity of pterosaurs that inhabited Gondwana during the Jurassic, which remains very scarce (Table 2).
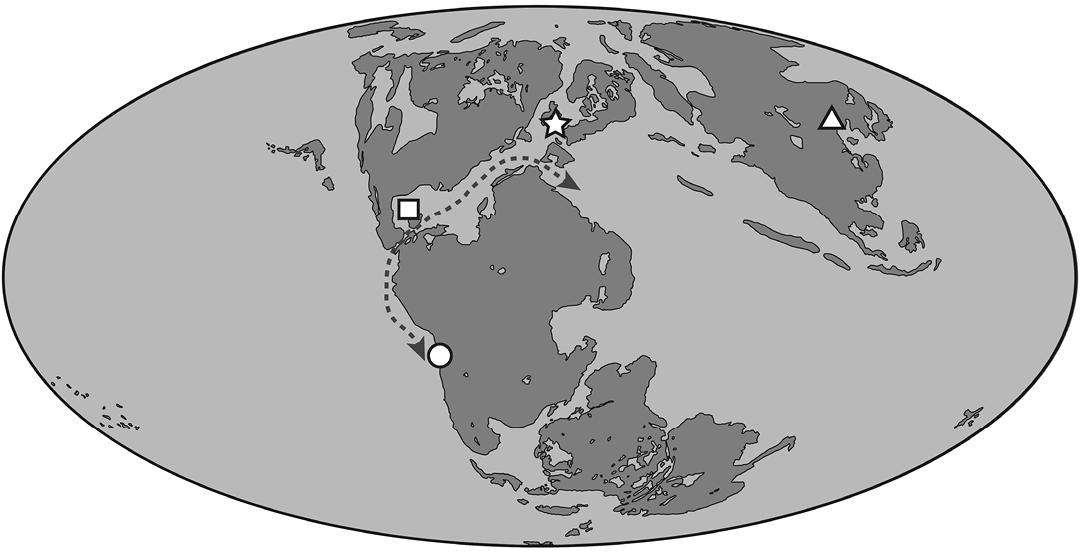
Fig. 7. Simplified map of the world during the Oxfordian. The dashed line represents the hypothetical route that would have allowed the dispersal of marine vertebrates and invertebrates between Tethys and South America during the Late Jurassic (Caribbean corridor). The finds of Oxfordian rhamphorhinchids: MUHNCAL.20165 (Rhamphorhynchinae gen. et sp. indet.), Cerro Campamento Formation, Chile (circle); Cacibupteryx caribensis (Gasparini et al. 2004) and Nesodactylus hesperius (Colbert et al. 1969), Jagua Formation, Cuba (square); Rhamphorhynchinae gen. et sp. indet. (Lydekker 1890, O’Sullivan 2018), Oxford Clay Formation, UK (star); Qinglongopterus guoi (Lü et al. 2012), Tiaojishan Formation, China (triangle). Modified map from Scotese (2014).
Table 2. Record of skeletal remains of valid Jurassic pterosaurs from Gondwana.
|
Taxon |
Age |
Chronostratigraphic unit |
Locality |
References |
|
Pterosauria indet. |
Hettangian– |
Upper Elliot Formation |
Farm Spioenkop, Senekal District, Free State, South Africa |
|
|
Pterosauria indet. |
Hettangian– |
Hanson Formation |
Mount Kirkpatrick, Central Transantarctic Mountains, Antarctica |
|
|
Allkaruen koi Codorniú Carabajal, Pol, Unwin, and Rauhut, 2016 |
Toarcian–Bathonian |
Cañadón Asfalto Formation |
Cerro Cóndor, Chubut Province, Argentina |
|
|
Pterosauria indet. |
Middle Jurassic |
Kota Formation |
Paikasigudem village, India |
|
|
Pterosauria indet. |
Middle Jurassic |
Kota Formation |
Chanda District, Deccan, India |
|
|
“Rhamphorhynchoidea” indet. |
Bathonian |
Isalo III Formation |
Faritany Majunga, Madagascar |
|
|
Rhamphorhynchidae indet. |
Bathonian |
Anoual Formation |
Guelb el Ahmar, Morocco |
|
|
Pterosauria indet. |
Bathonian |
Anoual Formation |
Guelb el Ahmar, Morocco |
|
|
Tendaguripterus recki Unwin and Heinrich, 1999 |
Kimmeridgian– |
Tendaguru |
Saurian Beds, Mtwara, Tanzania |
|
|
Dsungaripteroidea indet. |
Kimmeridgian– |
Tendaguru |
Saurian Beds, Mtwara, Tanzania |
|
|
Archaeopterodactyloidea indet. |
Kimmeridgian– |
Tendaguru |
Saurian Beds, Mtwara, Tanzania |
|
|
Wenupteryx uzi Codorniú and Gasparini, 2013 |
middle–late Tithonian |
Vaca Muerta |
Los Catutos, Neuquén Province, Argentina |
|
|
Pterodactyloidea indet. |
middle–late Tithonian |
Vaca Muerta |
Los Catutos, Neuquén Province, Argentina |
|
|
Herbstosaurus pigmaeus Casamiquela, 1975 |
late Tithonian |
Vaca Muerta |
Picún Leufú stream, Neuquén Province, Argentina |
|
|
Ctenochasmatidae |
Late Jurassic– |
Ksar Metlili |
Ksar Metlili, Morocco |
|
|
?Rhamphorhynchidae indet. |
Late Jurassic– |
Ksar Metlili |
Ksar Metlili, Morocco |
Conclusions
The specimen described here represents to date the oldest record of a pterosaur found in Chile, and the first confidently referrable to the clade Rhamphorhynchinae so far known in Gondwana. However, the absence of more complete and diagnostic material precludes a generic and specific referral for the moment.
In recent years, the findings of marine fossil vertebrates in the Cerro Campamento Formation have increased in number. Several of these finds, mainly of marine reptiles, have supported the proposals regarding the existence of close biogeographic connections between the southwestern margin of Gondwana and Europe at the end of the Jurassic. However, until now, it is not clear whether non-marine vertebrates that inhabited the same areas and at the same time, such as pterosaurs, followed similar patterns. In the studied locality, the abundance of concretions with three-dimensional preservation of delicate vertebrate remains (among which are fish and the pterosaur described here), offer good perspectives for the discovery of new well-preserved pterosaur specimens, which may help solve current questions related to their taxonomic identity, diversity, phylogenetic relationships, and biogeography.
Acknowledgements
We thank David Unwin (University of Leicester, UK) for providing photographs of pterosaur materials and suggestions that aided in the development of this work. The authors thank Michael O’Sullivan (University of Portsmouth, UK) and David Hone (Queen Mary University of London, UK) for their valuable comments and criticisms that contributed to improving the quality of this work. We thank Daniel E. Barta (Oklahoma State University Center for Health Sciences, Tahlequah, USA) for his help during the process of publishing this work. JAM, RAO, SAA, and AOV were supported by Proyecto Anillo ACT-172099 ANID-Chile and Fondecyt 1190891. All authors were supported in the field by the Corporación de Cultura y Turismo de Calama. JAM and SSA are supported by the ANID grant for Ph.D. studies in Chile.
References
Alarcón-Muñoz, J., Soto-Acuña, S., Codorniú, L., Rubilar-Rogers, D., Sallaberry, M., and Suárez, M. 2020. New ctenochasmatid pterosaur record for Gondwana: discovery in the Lower Cretaceous continental deposits of the Atacama Desert, northern Chile. Cretaceous Research 110: 104378. Crossref
Alarcón, J., Soto-Acuña, S., Otero, R., Ossa-Fuentes, L., and Rojas, O. 2015. Primer registro de pterosaurios en el Jurásico Superior (Oxfordiano) de la Formación Cerritos Bayos, Calama, Región de Antofagasta. In: R. Charrier, F. Hervé, V. Maksaev, C. Mpodozis, and J. Oyarzún (eds.), Actas del XIV Congreso Geológico Chileno, Vol. III, Área Temática 5, Bioestratigrafía y Paleontología Andina. October 4–8, 2015, 694–697. La Serena.
Alarcón-Muñoz, J., Soto-Acuña, S., Rubilar-Rogers, D., González, E., and Codorniú, L. 2018. Note on a new locality with pterosaurs (Archosauria: Pterodactyloidea) from the Atacama Region, Northern Chile. Boletín del Museo Nacional de Historia Natural, Chile 67: 145–153.
Andres, B., Clark, J.M., and Xing, X. 2010. A new rhamphorhynchid pterosaur from the Upper Jurassic of Xinjiang, China, and the phylogenetic relationships of basal pterosaurs. Journal of Vertebrate Paleontology 30: 163–187. Crossref
Andres, B., Clark, J., and Xu, X. 2014. The earliest pterodactyloid and the origin of the group. Current Biology 24: 1011–1016. Crossref
Arratia, G. 1994. Phylogenetic and paleobiogeographic relationships of the varasichthyid group (Teleostei) from the Late Jurassic of Central and South America. Revista Geológica de Chile 21: 119–165.
Arratia, G. 2008. The varasichthyid and other crossognathiforms fishes, and the break-up of Pangea. In: L. Calvin, A. Longbottom, and M. Richter (eds.), Fishes and the Break-up of Pangea. Geological Society, London, Special Publications 295: 71–92. Crossref
Baron, M.G. 2020. Testing pterosaur ingroup relationships through broader sampling of avemetatarsalian taxa and characters and a range of phylogenetic analysis techniques. PeerJ 8: e9604. Crossref
Barrett, P.M., Butler, R.J., Edwards, N.P., and Milner, A.R. 2008. Pterosaur distribution in time and space: and atlas. Zitteliana B 28: 61–107.
Bell, C.M. and Padian K. 1995. Pterosaurs fossils from the Cretaceous of Chile: Evidence for a pterosaur colony on an inland desert plain. Geological Magazine 132: 31–38. Crossref
Bell, C.M. and Suárez, M. 1989. Vertebrate fossils and trace fossils in Upper Jurassic–Lower Cretaceous red beds in the Atacama region, Chile. Journal of South American Earth Sciences 2 (4): 351–357. Crossref
Bennett, S.C. 1995. A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany: Year-classes of a single large species. Journal of Paleontology 69: 569–580. Crossref
Biese,W. 1957. Der Jura von Cerritos Bayos, Calama, República de Chile, Provinz Atacama. Geologische Jahrbuch 72: 439–493.
Biese, W. 1961. El Jurásico de Cerritos Bayos. Facultad de Ciencias Físicas y Matemáticas, Universidad de Chile, Instituto de Geología, Publicación 19: 1–61.
Blackbeard, M. and Yates, A.M. 2007. The taphonomy of an Early Jurassic dinosaur bonebed in the northern Free State (South Africa). Journal of Vertebrate Paleontology 27 (Supplement 3): 49A.
Bonde, N. and Christiansen, P. 2003. The detailed anatomy of Rhamphorhynchus: axial pneumaticity and its implications. In: E. Buffetaut and J.-M. Mazin (eds.), Evolution and Palaeobiology of Pterosaurs. Geological Society, London, Special Publications 217: 217–232. Crossref
Britt, B.B., Dalla Vecchia, F.M., Chure, D.J., Engelmann, G.F., Whiting, M.F., and Scheetz, D. 2018. Caelestiventus hanseni gen. et sp. nov. extends the desert-dwelling pterosaur record back 65 million years. Nature Ecology and Evolution 2: 1386–1392. Crossref
Buffetaut, E. and Guilbert, J.-P. 2001. An early pterodactyloid pterosaur from the Oxfordian of Normandy (northwestern France). Comptes Rendus de l’Académie des Sciences de Paris, Earth and Planetary Sciences 333: 405–409. Crossref
Buffetaut, E., Bülow, M., Gheerbrant, E., Jaeger, J., Martin, M., Manzin, J.M., Milsent, C., and Rioult, M. 1985. Zonation biostratigraphique et nouveaux restes de vertébrés dans les Sables de Glos (Oxfordien supérieur, Normandie). Comptes Rendus de l’Académie des Sciences de Paris, Earth and Planetary Sciences 300: 929–932.
Buffetaut, E., Suteethorn, V., Suteethorn, S., Deesri, U., and Tong, H. 2015. An azhdarchoid pterosaur humerus from the latest Jurassic (Phu Kradung Formation) of Phu Noi, north-eastern Thailand. Research and Knowledge 1: 43–47.
Casamiquela, R. 1975. Herbstosaurus pigmaeus (Coeluria, Compsognathidae) n. gen. n. sp. del Jurásico de Neuquén (Patagonia septentrional). Uno de los más pequeños dinosaurios conocidos. Actas del I Congreso Argentino de Paleontología y Bioestratigrafía 2: 80–103.
Casamiquela, R.M. and Chong Díaz, G.C. 1978. La presencia de Pterodaustro Bonaparte (Pterodactyloidea), del Neojurásico (?) de la Argentina, en los Andes del norte de Chile. Actas del Segundo Congreso Argentino de Paleontología y Bioestratigrafía y Primer Congreso Latinoamericano de Paleontología 1: 201–209.
Claessens, L.P.A.M., O’Connor, P.M., and Unwin, D.M. 2009. Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism. PloS ONE 4: e4497. Crossref
Codorniú, L. and Gasparini, Z. 2013. The Late Jurassic pterosaurs from northern Patagonia, Argentina. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 103: 399–408. Crossref
Codorniú, L., Carabajal, A.P., Pol, D., Unwin, D., and Rauhut, O.W.M. 2016. A Jurassic pterosaur from Patagonia and the origin of the pterodactyloid neurocranium. PeerJ 4: e2311. Crossref
Codorniú, L., Rauhut, O.W.M., and Pol, D. 2010. Osteological features of Middle Jurassic pterosaurs from Patagonia (Argentina). Acta Geoscientica Sinica 31 (Supplement 1): 12–13.
Colbert, E.H. 1969. A Jurassic Pterosaur from Cuba. American Museum Novitates 2370: 1–26.
Costa, F.R. and Kellner, A.W.A. 2009. On two pterosaur humeri from the Tendaguru beds (Upper Jurassic, Tanzania). Anais da Academia Brasileira de Ciências 81: 813–818. Crossref
Costa, F.R., Sayão, J.M., and Kellner, A.W.A. 2015. New pterosaur material from the Upper Jurassic of Tendaguru (Tanzania), Africa. Historical Biology 27: 646–655. Crossref
Czerkas, S.A. and Ji, Q. 2002. A new rhamphorhynchoid with a headcrest and complex integumentary structures. In: S.J. Czerkas (ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1: 15–41.
Dal Sasso, C. and Pasini, G. 2003. First record of pterosaurs (Diapsida, Archosauromorpha, Pterosauria) in the Middle Jurassic of Madagascar. Atti della Societa Italiana di Scienze Natuali e del Museo Civico di Storia Naturale de Milano 144: 281–296.
Dean, C.D., Mannion, P.D., and Butler, R.J. 2016. Preservational bias controls the fossil record of pterosaurs. Palaeontology 59: 225–247. Crossref
Duhart, O.P., Muñoz, B.J., Quiroz, P.D., Maestre, B.A., and Varas, M.G. 2018. Carta Sierra Gorda, Región de Antofagasta. Escala 1:100.000. Carta Geológica de Chile, Serie Geología Básica 198: 1–213.
Elgin, R.A. and Hone, D.W.E. 2013. Pneumatization of an immature azhdarchoid pterosaur. Cretaceous Research 45: 16–24. Crossref
Frey, E. and Tischlinger, H. 2012. The Late Jurassic pterosaur Rhamphorhynchus, a frequent victim of the ganoid fish Aspidorhynchus? PLoS ONE 7: e31945. Crossref
Gasparini, Z. 2009. A new Oxfordian pliosaurid (Plesiosauria, Pliosauridae) in the Caribbean Seaway. Palaeontology 52: 661–669. Crossref
Gasparini, Z. and Iturralde-Vinent, M. 2006. The Cuban Oxfordian herpetofauna in the Caribbean Seaway. Neues Jahrbuch für Geologie und Paläontologie, Abhandlunden 240: 343–371. Crossref
Gasparini, Z., Fernández, M., and De la Fuente, M. 2004. A new pterosaur from the Jurassic of Cuba. Palaeontology 47: 919–927. Crossref
Goldfuss, A. 1831, Beiträge zur Kenntnis verschiedener Reptilien der Vorwelt. Nova acta Academiae caesareae Leopoldino-Carolinae germanicae naturae curiosorum 15: 61–128.
Gross, W. 1937. Über einen neuen Rhamphorhynchus gemmingi H. v. M. des Natur-Museums Senckenberg. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 437:1–16.
Haddoumi, H., Allain, R., Meslouh, S., Metais, G., Monbaron, M., Pons, D., Rage, J-P., Vullo, R., Zouhri, S., and Gheerbrant, E. 2016. Guelb el Ahmar (Bathonian, Anoual Syncline, eastern Morocco): First continental flora and fauna including mammals from the Middle Jurassic of Africa. Gondwana Research 29: 290–319. Crossref
Hammer, W.R. and Hickerson, W.J. 1994. A crested theropod dinosaur from Antarctica. Science 264: 828–830. Crossref
He, X., Yang, D., and Su, C. 1983. A new pterosaur from the Middle Jurassic of Dshanpu, Zigong, Sichuan. Journal of Chengdu College, Geology Supplement 1: 27–33
Hone, D.W.E., Henderson, D.M. Therrien, F., and Habib, M.B. 2015. A specimen of Rhamphorhynchus with soft tissue preservation, stomach contents and a putative coprolite. PeerJ 3: e1191. Crossref
Hone, D.W.E., Tischlinger, H., Frey, E., and Röper, M. 2012. A new non-pterodactyloid pterosaur from the Late Jurassic of Southern Germany. Plos ONE 7: e39312. Crossref
Iturralde-Vinent, M.A. 2006. Origen de la biota y los ecosistemas marinos de Cuba. In: R. Claro (ed.), La Biodiversidad Marina de Cuba, 1–9. Instituto de Oceanología, CITMA, La Habana.
Jain, S.L. 1974. Jurassic pterosaur from India. Journal of the Geological Society of India 15: 330–335.
Kellner, A.W.A. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. In: E. Buffetaut and J.-M. Mazin (eds.), Evolution and Paleobiology of Pterosaurs. Geological Society, London, Special Publications 217: 105–137. Crossref
Lasseron, M., Allain, R., Gheerbrant, E., Haddoumi, H., Jalil, N-E., Métais, G., Rage, J-C., Vullo, R., and Zouhri, S. 2020. New data on the microvertebrate fauna from the Upper Jurassic or lowest Cretaceous of Ksar Metlili (Anoual Syncline, eastern Morocco). Geological Magazine 157 (3): 367–392. Crossref
Lü, J. and Bo, X. 2011. A new rhamphorhynchid pterosaur (Pterosauria) from the Middle Jurassic Tiaojishan Formation of Western Liaoning, China. Acta Geologica Sinica 85: 977–983. Crossref
Lü, J.C., Unwin, D.M., Zhou, B., Gao, C., and Shen, C. 2012. A new rhamphorhynchid (Pterosauria: Rhamphorhynchidae) from the Middle/Upper Jurassic of Qinglong, Hubei Province, China. Zootaxa 3158: 1–19. Crossref
Lydekker, R. 1890. On ornithosaurian remains from the Oxford Clay of Huntingdonshire. Quarterly Journal of the Geological Society 46: 429–431. Crossref
Martill, D.M., Frey, E., Chong Díaz, G., and Bell, C.M. 2000. Reinterpretation of a Chilean pterosaur and the occurrence of Dsungaripteridae in South America. Geological Magazine 137: 19–25. Crossref
Martill, D.M., Frey, E., Bell, C.M., and Chong Díaz, G. 2006. Ctenochasmatid pterosaur from Early Cretaceous deposits in Chile. Cretaceous Research 27: 603–610. Crossref
Meléndez, G. and Myczyński, R. 1987. Sobre la posición sistemática de los ammonites del Oxfordiense de los Andes Chilenos (Cordillera Domeyko, Chile, Provincia Andina). Geogaceta 2: 12–14.
Newton, E.T. 1888. On the skull, brain and auditory organ of a new species of pterosaurian Scaphognathus purdoni from the Upper Lias near Whitby, Yorkshire. Philosophical Transactions of the Royal Society of London 179: 503–537. Crossref
O’Sullivan, M. 2018. The pterosaur assemblage of the Oxford Clay Formation (Jurassic, Callovian–Oxfordian) from UK. In: D.W.E. Hone, M.P. Witton, and D.M. Martill (eds.), New Perspectives on Pterosaur Palaeobiology. Geological Society, London, Special Publications 455: 171–180. Crossref
O’Sullivan, M. and Martill, D.M. 2015. Evidence for the presence of Rhamphorhynchus (Pterosauria: Rhamphorhynchinae) in the Kimmeridge Clay of the UK. Proceedings of the Geologists’ Association 126: 390–401. Crossref
O’Sullivan, M., and Martill, D.M. 2018. Pterosauria of the Great Oolite Group (Bathonian, Middle Jurassic) of Oxfordshire and Gloucestershire, England. Acta Palaeontologica Polonica 63: 617–644. Crossref
O’Sullivan, M., Martill, D.M., and Groocock, D. 2013. A pterosaur humerus and scapulocoracoid from the Jurassic Whitby Mudstone Formation, and the evolution of large body size in early pterosaurs. Proceedings of the Geologists’ Association 124: 973–981. Crossref
Otero, R.A., Alarcón-Muñoz, J., Soto-Acuña, S., Rojas, J., Rojas, O., and Ortiz, H. 2020. Cryptoclidid plesiosaurs (Sauropterygia, Plesiosauria) from the Upper Jurassic of the Atacama Desert. Journal of Vertebrate Paleontology 40: e1764573. Crossref
Padian, K. 1983. Osteology and functional morphology of Dimorphodon macronyx (Buckland) (Pterosauria: Rhamphorhynchoidea) based on new material in the Yale Peabody Museum. Postilla 189: 1–44.
Padian, K. 2008a. The Early Jurassic pterosaur Campylognathoides Strand, 1928. Special Papers in Palaeontology 80: 65–107.
Padian, K. 2008b. The Early Jurassic pterosaur Dorygnathus banthensis (Theodori, 1830). Special Papers in Paleontology 80: 1–64.
Padian, K. and Wild, R. 1992. Studies of Liassic Pterosauria, I. The holotype and referred specimens of the Liassic Pterosaur Dorygnathus banthensis (Theodori) in the Petrefaktensammlung Banz, Northern Bavaria. Palaeontographica Abteilung A 225 (1–3): 59–77.
Parent, H. 2006. Oxfordian and Late Callovian ammonite faunas and biostratigraphy of the Neuquén-Mendoza and Tarapacá Basins (Jurassic, Ammonoidea, Western South-America). Boletín del Instituto de Fisiografía y Geología 76: 1–70.
Prasad, G.V.R. and Parmar, V. 2020. First ornitischian and theropod dinosaur teeth from the Middle Jurassic Kota Formation of India: Paleobiogeographic relationships. In: G.V.R. Prasad and R. Patnaik (eds.), Biological Consequences of Plate Tectonics: New Perspectives on Post-Gondwana Break-up—a Tribute to Ashok Sahni. Vertebrate Paleobiology and Paleoanthropology Series, 1–30. Springer, Cham. Crossref
Rage, J.-C. 2016. Gondwana, Tethys, and terrestrial vertebrates during the Mesozoic and Cainozoic. In: M.G. Audley-Charles and A. Hallam (eds.), Gondwana and Tethys Geological Society Special Publication 37: 255–273. Crossref
Rao, C.N. and Shah, S.C. 1963. On occurrence of Pterosaur from the Kota-Maleri beds of Chanda district. Records of the Geological Survey of India 92: 315–318.
Rauhut, O.W.M., López Arbarello, A., Puerta, P., and Martín, T. 2001. Jurassic vertebrates from Patagonia. Journal of Vertebrate Paleontology 21 (Supplement): 91A.
Rauhut, O.W.M., López-Arbarello, A., Röper, M., and Rothgaenger, M. 2017. Vertebrate fossils from the Kimmeridgian of Brunn: the oldest fauna from Solnhofen Archipielago (Late Jurassic, Bavaria, Germany). Zitteliana 89: 305–329.
Reck, H. 1931. Die deutschostafrikanischen Flugsaurier. Centralblatt für Mineralogie, Geologie und Paläontologie Abteilung B 7: 321–336.
Scotese, C.R. 2014. PALEOMAP Atlas for ArcGIS, Volume 3, The Jurassic and Triassic, Maps 32–42, Mollweide Projection. PALEOMAP Project, Evanston.
Sharov, A.G. 1971. New flying reptiles from the Mesozoic of Kazakhstan and Kirghizia [in Russian]. Trudy Paleozoologičeskogo Instituta, Akademiâ Nauk SSSR, Leningrad 130: 104–113.
Smith, P.L. 1983. The Pliensbachian ammonite Dayiceras dayiceroides and Early Jurassic paleogeography. Canadian Journal of Earth Sciences 20: 86–91. Crossref
Soto-Acuña, S., Otero, R.A., Rubilar-Rogers, D., and Vargas, A.O. 2015. Arcosaurios no avianos de Chile. In: D. Rubilar-Rogers, R. Otero, A. Vargas, and M. Sallaberry (eds.), Vertebrados Fósiles de Chile. Publicación Ocasional del Museo Nacional de Historia Natural, Chile 63: 209–263.
Theodori, C. v. 1852. Über die Pterodactylus-Knochen im Lias von Banz. Bericht der naturforschenden Gesellschaft Bamberg 1: 17–44.
Unwin, D.M. 2003. On the phylogeny and evolutionary history of pterosaurs. In: E. Buffetaut and J.-M. Mazin (eds.), Evolution and Palaeobiology of Pterosaurs. Geological Society, London, Special Publication 217: 139–190. Crossref
Unwin, D.M. 2006. The Pterosaurs From Deep Time. 347 pp. Pi Press, New York.
Unwin, D.M. and Heinrich, W.-D. 1999. On a pterosaur jaw from the Upper Jurassic of Tendaguru (Tanzania). Mitteilungen aus dem Museum fur Naturkunde Berlin, Geowissenschaftlichen Reihe 2: 121–134. Crossref
Unwin, D.M. and Martill, D.M. 2017. Systematic reassessment of the first Jurassic pterosaur from Thailand. In: D.W.E. Hone, M.P. Witton, and D.M. Martill (eds.), New Perspectives on Pterosaur Palaeobiology. Geological Society, London, Special Publications 455: 181–186. Crossref
Unwin, D.M., Rauhut, O.W.M., and Haluza, A. 2004. The first “rhamphorhynchoid” from South America and the early history of pterosaurs. Geobiologie 74: 235–237.
Upchurch, P., Andres, B., Butler, R.J., and Barrett, P.M. 2015. An analysis of pterosaurian biogeography: implications for the evolutionary history and fossil record quality of the first flying vertebrates. Historical Biology 27: 697–717. Crossref
Vidovic, S.U. and Martill, D.M. 2018. The taxonomy and phylogeny of Diopecephalus kochi (Wagner, 1837) and “Germanodactylus rhamphastinus” (Wagner, 1851). In: D.W.E. Hone, M.P. Witton, and D.M. Martill (eds.), New Perspectives on Pterosaur Palaeobiology. Geological Society, London, Special Publications 455: 125–147. Crossref
Wellnhofer, P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura Plattenkalke. Süddeutschlands. Abhandlungen der Bayerischen Akademie der Wissenschaften zu München, Mathematisch-Naturwisenschaftlichen Klasse 141: 1–133.
Wellnhoffer, P. 1975a. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. I. Allgemeine Skelettmorphologie. Palaeontographica Abteilung A 148:1–33.
Wellnhofer, P. 1975b. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. II. Systematische Beschreibung. Palaeontographica Abteilung A 148: 132–186.
Wellnhofer, P. 1975c. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. III. Palökologie und Stammesgeschichte. Palaeontographica Abteilung A 149: 1–30.
Wellnhofer, P. 1978. Pterosauria. In: P. Wellnhofer (ed.), Handbuch der Paläoherpetologie. Encyclopedia of Paleoherpetology. Teil 19, 1–88. Gustav Fischer, Stuttgart.
Wellnhofer, P. 1991. The Illustrated Encyclopedia of Pterosaurs. 192 pp. Salamander Books, London.
Witton, M.P. 2013. Pterosaurs: Natural History, Evolution, Anatomy. 340 pp. Princeton University Press, Princeton, New Jersey. Crossref
Witton, M.P. 2015. Were early pterosaurs inept terrestrial locomotors? PeerJ 3: e1018.Crossref
Witton, M. 2018. Pterosaurs in Mesozoic food webs: a review of fossil evidence. In: D.W.E. Hone, M.P. Witton, and D.M. Martill (eds.), New Perspectives on Pterosaur Palaeobiology. Geological Society, London, Special Publications 455: 7–23. Crossref
Acta Palaeontol. Pol. 66 (3): 571–583, 2021
https://doi.org/10.4202/app.00805.2020