
![]()
An anhanguerian pterodactyloid mandible from the lower Valanginian of Northern Germany, and the German record of Cretaceous pterosaurs
PASCAL ABEL, JAHN J. HORNUNG, BENJAMIN P. KEAR, and SVEN SACHS
Abel, P., Hornung, J.J., Kear, B.P., and Sachs, S. 2021. An anhanguerian pterodactyloid mandible from the lower Valanginian of Northern Germany, and the German record of Cretaceous pterosaurs. Acta Palaeontologica Polonica 66 (Supplement to 3): 5–12.
The record of Cretaceous pterosaur remains from Germany is sparse. The material recovered to date includes the fragmentary holotypes of Targaryendraco wiedenrothi and Ctenochasma roemeri, as well as a few isolated pterodactyloid teeth and some indeterminate skeletal elements, together with a plaster cast of a large Purbeckopus manus imprint. Here, we report the discovery of a pterodactyloid pterosaur mandible from lower Valanginian strata of the Stadthagen Formation in the Lower Saxony Basin of Northern Germany. Based on the size and spacing of its alveoli, this fossil is attributable to the cosmopolitan Early Cretaceous pteranodontoid clade Anhangueria. Moreover, it represents the first and only known pterosaur from the Valanginian of Germany and is one of only a handful Valanganian pterosaur occurrences presently recognized worldwide. In addition to the approximately coeval Coloborhynchus clavirostris from the Hastings Bed Group of southern England, the Stadthagen Formation pterosaur mandible is among the stratigraphically oldest identifiable anhanguerians.
Key words: Pterosauria, Pterodactyloidea, Anhangueria, Cretaceous, Stadthagen Formation, Lower Saxony.
Pascal Abel [pascal.abel94@web.de], Senckenberg Centre for Human Evolution and Palaeoenvironment, Eberhard-Karls-University Tübingen, Sigwartstraße 28, 72076 Tübingen, Germany.
Jahn J. Hornung [jahn.hornung@yahoo.de], Niedersächsisches Landesmuseum Hannover, Willy-Brandt-Allee 5, 30169 Hannover, Germany.
Benjamin P. Kear [benjamin.kear@em.uu.se], Museum of Evolution, Uppsala University, Norbyvägen 22, 75236 Uppsala, Sweden.
Sven Sachs [sachs.pal@gmail.com], Naturkunde-Museum Bielefeld, Abteilung Geowissenschaften, Adenauerplatz 2, 33602 Bielefeld, Germany.
Received 16 September 2020, accepted 17 May 2021, available online 24 August 2021.
Copyright © 2021 P. Abel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Pterosaurs were the first vertebrates to evolve an active volant lifestyle and were globally widespread from the Middle Triassic until their extinction at the end of the Cretaceous (e.g., Butler et al. 2009; Longrich et al. 2018). Among the paraphyletic succession of early pterosaur lineages, only the “short-tailed” pterodactyloids and anurognathids survived beyond the Jurassic–Cretaceous boundary, with pterodactyloids subsequently diverging into the Azhdarchoidea and Pteranodontoidea (Kellner 2003; Hone 2020). One of the most successful of these Cretaceous pterodactyloid clades were the Anhangueria, a cosmopolitan radiation of toothed and often large pteranodontoids, best known from the Aptian–Albian strata of Great Britain and Brazil (Rodrigues and Kellner 2013), but ranging up into the Turonian elsewhere (Rodrigues and Kellner 2013; Pentland et al. 2019, Frey et al. 2020). To date, the stratigraphically oldest known anhanguerian is Coloborhynchus clavirostris Owen, 1874 from the upper Berriasian–lower Valanginian of England (Rodrigues and Kellner 2013; Pentland et al. 2019), which suggests possible origination of the group shortly after the Jurassic–Cretaceous transition.
Here we describe a new fragmentary mandible (RE 551.763.120 A 0333/1) of an anhanguerian pterodactyloid from the lower Valanginian of Northern Germany. This specimen was found by fossil collector and preparator Karl-Heinz Hilpert (formerly at the Geologisch-Paläontologisches Museum Münster) in the abandoned Sachsenhagen clay pit in Lower Saxony, and subsequently donated to the Ruhr Museum Essen in 1998. RE 551.763.120 A 0333/1 is important, because together with C. clavirostris it represents one of the oldest known anhanguerian occurrences. In addition, it is one of only very few pterosaur specimens currently identified from the German Cretaceous (von Meyer 1851; Wild 1990; Hornung and Reich 2013; Lanser 2015) and is also the first documented pterosaur fossil found in Germany that is of Valanginian age.
Institutional abbreviations.—GZG.IF, Ichnofossil collection, Geoscience Centre, University of Göttingen, Göttingen, Germany; LWL, LWL-Museum für Naturkunde, Münster, Germany; RE, Ruhr Museum, Essen, Germany; RPMH, Römer-Pelizaeus-Museum Hildesheim, Hildesheim, Germany; SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany.
Geological setting
The Sachsenhagen clay pit is located approximately 30 km west of Hannover (Fig. 1A) and was operated as brickworks from 1904 to 1986. Most of the quarry area has since been backfilled and used as a waste dump. The exposed profile spans a lithofacially mostly homogenous, pelitic succession of the upper Berriasian upper Isterberg Formation and lower Valanginian lower Stadthagen Formation (Fig. 1B). The transition between these units correlates with a shift from euryhaline to stenohaline conditions following a marine transgression (Kaufmann et al. 1980; Mutterlose 2017). Paleogeographically, the deposits were formed in a shallow gulf that inundated the Lower Saxony Basin as a southward extension of the North Sea Basin (Fig. 1C; see Mutterlose 2017).
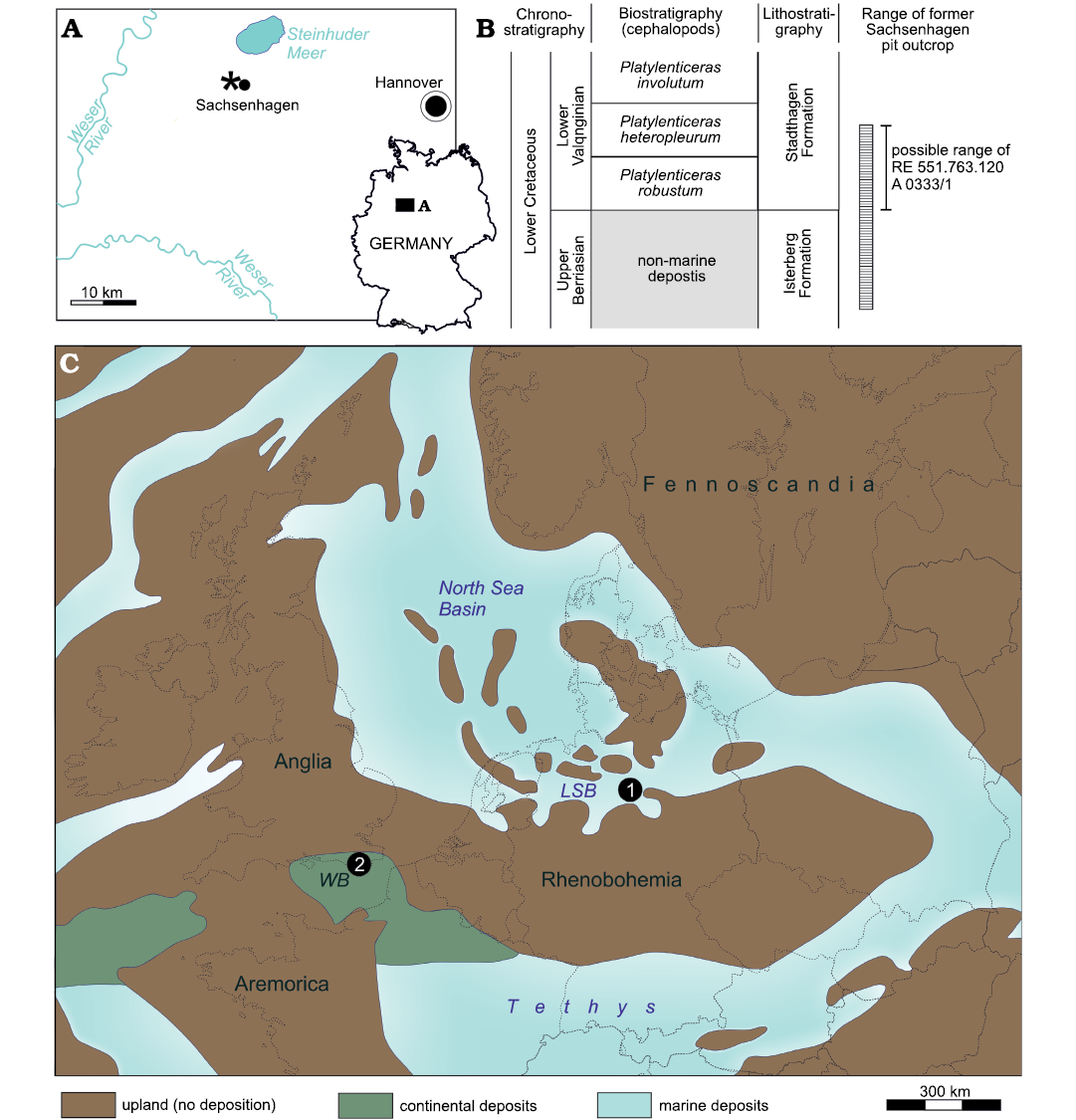
Fig. 1. A. Geographic location (asterisk) of the former Sachsenhagen clay-pit. B. Stratigraphical framework of the Lower Cretaceous in the former Sachsenhagen clay-pit. Modified after Mutterlose et al. (1997), Mutterlose (2017), and Erbacher et al. (2014a, b). C. Palaeogeographic reconstruction of Central Europe during the Valanginian–Hauterivian interval showing the position of Sachsenhagen (1) in the Lower Saxony Basin (LSB) and St. Leonard-on-Sea (2), the locus typicus of the approximately coeval Coloborhynchus clavirostris, in the Wessex Basin (WB). Modified after Mutterlose et al. (1997).
Locality information indicates that RE 551.763.120 A 0333/1 derived from the lower Valanginian lower Stadthagen Formation spanning the Platylenticeras robustum and P. heteropleurum (Mutterlose 1984, 2017) ammonite zones. Zawischa (1992) and Frerichs and Girod (2017) documented a rich vertebrate fossil assemblage, including actinopterygians (Frerichs 2017a), plesiosaurs (Frerichs 2017b), the thalattosuchian crocodylomorph “Enaliosuchus schroederi” Kuhn, 1936 (see Schroeder 1923; Sickenberg 1961; Sachs et al. 2020), and additional indeterminate crocodyliform remains (Frerichs 2017b). Most of the material is currently located in private collections.
Systematic palaeontology
Pterosauria Kaup, 1834
Pterodactyloidea Plieninger, 1901
Pteranodontoidea Kellner, 1996
Anhangueria Rodrigues and Kellner, 2013
Anhangueria indet.
Fig. 2.
Material.—RE 551.763.120 A 0333/1, an incomplete mandible from the abandoned Sachsenhagen clay pit, Sachsenhagen, Lower Saxony, Germany (approximate coordinates 52°23’51” N, 09°15’18” E); Platylenticeras robustum or Platylenticeras heteropleurum ammonite biozones (NW-European ammonite province of the Sub-Boreal Realm: sensu Kemper et al. 1981, Marek and Shulgina 1996), lower Valanginian, Lower Cretaceous, Stadthagen Formation.
Description.—The preserved mandible includes the posterior section of the symphysis and parts of both dentaries and angulars. The splenials have been disarticulated and are now lost. Thus, the Meckelian groove is exposed on the medial surfaces of both rami (Fig. 2). The mandible has a sub-circular cross-section anterior to the symphyseal contact. There is no evidence of a sagittal crest. The external ventral face of the mandibular rami is shallowly convex. The dorsal side of the symphyseal portion is strongly concave and has an irregular midline suture (Fig. 2A). A conspicuous lateromedial furrow is visible along the length of the rami ventral to the alveolar row. The post-symphyseal space is filled with sediment. However, both rami diverge at an angle of about 23° relative to the longitudinal axis.

Fig. 2. Mandible of anhanguerian pterosaur RE 551.763.120 A 0333/1 from the lower Valanginian of Sachsenhagen (Northern Germany), in dorsal (A), left lateral (B), ventral (C), and right lateral (D) views. Photographs (A1–D1), explanatory drawings (A2–D2).
The mandible has suffered some diagenetic crushing, resulting in a prominent longitudinal crack on the lateral surface of the left ramus (Fig. 2B). Especially the lateral surface of the right ramus forms two longitudinal bulges, resulting in an undulating surface. The postsymphyseal portions of both rami were previously broken und glued back to the symphysis. Finally, the entire surface area of bone and exposed matrix on RE 551.763.120 A 0333/1 has been varnished and painted at some stage of preparation. This has created a pattern of irregular grooves and ridges along the ventral face of the symphysis. We consider these to be artificial, yet some bulges and rugosities on the median ventral face of the symphysis and posterior parts of the rami could possibly be pathological (Fig. 2C).
Dentary: The dentaries form the mandibular sulcus, alveolar row, and the base of the Meckelian groove. The dorsal trace of the splenial suture is located ventrally adjacent to the alveoli. The ventral suture is not visible. The angular contacts are exposed on the lateral surfaces of the mandibular rami (Fig. 2B, D), however, there is no evidence of the coronoid contacts. On the right ramus, thin ridges extending parallel to the alveolar row and posterodorsally across the lateral surface posterior to the fifth alveolus (Fig. 2D) might border nerve and/or blood vessel channels.
Angular: The angulars form about the ventrolateral third of the external surfaces of the rami, and taper to the level of the posterior-most symphyseal alveoli. Medially, the angulars border the Meckelian groove and presumably sutured to the splenials dorsally.
Alveoli: Five alveoli are preserved on the left ramus, and six on the right; only two alveoli are placed adjacent to the symphysis on either side. They are about 1.5–1.9 times longer than wide, and each alveolus is slightly anteromedially directed (Table 1). The alveolar margins are flush with the surrounding bone. The interalveolar spaces are 2.5–4 times the length of the alveoli and increase in length posteriorly to the second preserved alveolus, but then decreases again further posteriorly (Table 1).
Table 1. Linear measurements (in mm) of RE 551.763.120 A 0333/1. Alveoli measured in posterior direction. Asterisks indicate incomplete alveoli and/or alveoli filled with sediment.
|
Diastemae, ramus |
left |
14.25 |
16.84 |
12.42 |
12.65 |
|
|
|
right |
15.94 |
16.68 |
12.76 |
12.14 |
13.58 |
|
|
|
Alveolar length, ramus |
left |
4.56* |
4.95 |
4.37* |
4.27 |
4.54 |
|
|
right |
4.42 |
4.17 |
5.20* |
4.83 |
5.31* |
5.20 |
|
|
Alveolar width, ramus |
left |
3.12* |
2.99 |
2.87* |
2.70 |
2.96 |
|
|
right |
2.86 |
2.43 |
2.75 |
2.58 |
2.56* |
2.81 |
|
Maximum preserved length |
117.63 |
|
Preserved symphysis, dorsal length |
28.27 |
|
Preserved symphysis, ventral length |
40.00 |
|
Dentary (left), maximum height |
18.73 |
|
Dentary (left), maximum width |
5.86 |
Discussion
Comparisons and classification.—Among Early Cretaceous toothed pterodactyloids, RE 551.763.120 A 0333/1 differs from ctenochasmatids and boreopterids, as these latter possess large numbers of teeth in a “comb-like” arrangement (Kellner 2003; Lü and Ji 2010; Jiang et al. 2014). Lonchodraco Rodrigues and Kellner, 2013 and other putative “lonchodectids”, such as Lonchodectes compressirostris Owen, 1851, Serradraco sagitirostris Owen, 1874, Prejanopterus curvirostris Vidarte and Calvo, 2010, or Targaryendraco wiedenrothi Wild, 1990, are distinguished from the specimen described herein by their “lanceolate” jaw outline, inter-alveolar spacing that is sub-equal to the corresponding alveolar length (Wild 1990; Pereda-Suberbiola et al. 2012; Rodrigues and Kellner 2013; Rigal et al. 2017), and alveolar margins raised to a pedestal (Unwin 2001; Rodrigues and Kellner 2013). Likewise, dsungaripterids have closely spaced alveoli, where the dental bone bulges to envelope the base of the tooth crowns (Martill et al. 2000; Witton 2013; Chen et al. 2020). In istiodactyliforms, the alveoli are even more densely packed (Lü et al. 2008; Witton 2013; Kellner et al. 2019).
By contrast, the alveolar morphology of RE 551.763.120 A 0333/1 is very similar to known anhanguerians, which typically have well-spaced alveoli along the mid- and posterior sections of the mandible (e.g., Wellnhofer 1987; Elgin and Frey 2011; Rodrigues and Kellner 2013). A taxonomic assignment of RE 551.763.120 A 0333/1 within Anhangueria, however, proves to be difficult. Especially the taxa of the specimen-rich Cambridge Greensand and other Lower Cretaceous strata of Great Britain are often only based on anterior rostral fragments, prohibiting a direct comparison with RE 551.763.120 A 0333/1 (Rodrigues and Kellner 2013), where these elements are missing. This includes most notably the roughly coeval Coloborhynchus clavirostris from the adjacent British Wealden. But even in more completely known mandibles most diagnostic traits are restricted to the anterior part of the symphysis (e.g., Steel et al. 2005).
Nevertheless, RE 551.763.120 A 0333/1 is distinct from Anhanguera piscator Kellner and Tomida, 2000, “Araripesaurus santanae” Wellnhofer, 1985, Mythunga Molnar and Thulburn, 2007, and “Tropeognathus robustus” Wellnhofer, 1987, because it lacks inter-alveolar spaces that decrease in length towards the back of the jaw (Wellnhofer 1985, 1987; Veldmeijer 2003; Pentland and Poropat 2019). RE 551.763.120 A 0333/1 further differs from Brasileodactylus araripensis Kellner, 1984, Ferrodraco lentoni Pentland, Poropat, Tischler, Sloan, R.A. Elliot, H.A. Elliot, J.A. Elliot, and D.A. Elliot, 2019, Maaradactylus spielbergi Veldmeijer, 2003, and Tropeognathus mesembrinus Wellnhofer, 1987, because these taxa have inter-alveolar spaces that successively increase in length (Wellnhofer 1987; Veldmeijer 2003; Veldmeijer et al. 2009; Pentland et al. 2019). Lastly, unlike Anhanguera piscator, “Araripesaurus santanae”, and “Santanadactylus araripensis” Wellnhofer, 1985, RE 551.763.120 A 0333/1 possesses proportionately large alveoli compared to the transverse width of the mandibular ramus (Wellnhofer 1985; Kellner and Tomida 2000). Besides the alveolar pattern, RE 551.763.120 A 0333/1 differs further from Maaradactylus spielbergi by its more pronounced sulcus (Veldmeijer 2003).
The number of symphyseal teeth is variable among anhanguerians, ranging from six in Cearadactylus atrox Leonardi and Borgomanero 1985 (Unwin 2002; Vila Nova et al. 2014) or Tropeognathus mesembrinus to at least eleven in other far-related species, such as Maaradactylus spielbergi, Brasileodactylus araripensis, or Aetodactylus halli Myers, 2010 (Veldmeijer 2003; Veldmeijer et al. 2009; Myers 2010; Pinheiro and Rodridgues 2017). This makes it impossible to identify the position of the alveoli preserved in our specimen. Nevertheless, in the majority of anhanguerians preserving the mandible, the mandibular rami bear only two to five post-symphyseal teeth (Wellnhofer 1985, 1987; Kellner and Tomida 2000; Veldmeijer 2003; Vila Nova et al. 2014; Pinheiro and Rodrigues 2017), indicating the post-symphyseal alveoli preserved in RE 551.763.120 A 0333/1 may also represent the posterior-most alveoli of the mandible. A notable exception is Aetodactylus halli, which has 13 post-symphyseal alveoli (Myers 2010). However, Aetodactylus halli, together with the probably closely related Cimoliopterus cuvieri Bowerbank, 1851, differ from RE 551.763.120 A 0333/1 and other anhanguerians in their more densely packed teeth (Myers 2010, 2015). Furthermore, they were found to nest outside Anhangueria by some recent phylogenetic studies (Pêgas et al. 2019).
Unlike RE 551.763.120 A 0333/1 and “Santanadactylus araripensis” (Wellnhofer 1985), the angulars in Cearadactylus atrox and Anhanguera piscator do not extend as far as the posterior end of the tooth row (Leonardi and Borgomanero 1985; Kellner and Tomida 2000). The condition of RE 551.763.120 A 0333/1 is closer to “Santanadactylus araripensis”, where the lateral portion of angular reaches almost the level of the posterior-most alveolus, which is in this taxon the fourth post-symphyseal tooth (Wellnhofer 1985). The same author interpreted the angular in “Araripesaurus santanae” to extend distinctly beyond the tooth bearing portion of the mandible, reaching at the least the level of the in anterior direction fifth alveolus. However, Kellner and Tomida (2000) argued that the angular may be significantly shorter in both “Santanadactylus araripensis” and “Araripesaurus santanae” but based this on the assumption of a congeneric relationship with Anhanguera piscator. This cannot be confirmed, because “Santanadactylus araripensis” and “Araripesaurus santanae” are considered to be nomina dubia (Pinheiro and Rodrigues 2017). Irrespectively, the angular of RE 551.763.120 A 0333/1 is most closely comparable to “Santanadactylus araripensis” as described by Wellnhofer (1985), although the anterior extremity is less anteriorly tapered in RE 551.763.120 A 0333/1.
Overall, complete mandibles are only barely known in anhanguerians, and often the mandibular bones are fused beyond distinction, suggesting these specimens may represent adult individuals (Kellner and Tomida 2000; Vila Nova et al. 2014). Evidently, RE 551.763.120 A 0333/1 likely derived from an osteologically immature animal.
Cretaceous pterosaurs from Germany.—Germany is world famous for its record of articulated Upper Jurassic pterosaur remains (e.g., Wellnhofer 1970, 1978; Bennett 2002, 2007; Tischlinger 2010; Hone et al. 2013; Tischlinger and Frey 2013). Conversely, pterosaur fossils of Cretaceous age are extremely rare despite the earliest documented occurrences dating back to the 19th century. This is also in stark contrast to other Cretaceous pterosaur-bearing sites in the world with a rich pterosaur diversity like the northeastern Brazilian Santana Group (e.g., Wellnhofer 1985, 1987; Kellner and Tomida 2000; Unwin 2002), or the British Wealden and Cambridge Greensand (e.g., Unwin 2001; Rodrigues and Kellner 2013; Rigal et al. 2017). The German record is currently restricted to the Lower Cretaceous. However, only the Valanginian and Hauterivian pterosaur specimens derive from strata that were deposited in some distance from the paleo-coastline. All other remains originated from non-marine, lagoonal to fully limnic and fluviatile environments.
The first Cretaceous pterosaur find described from Germany has been the holotype of Ctenochasma roemeri (non-catalogued, von Meyer 1851) from the lower Berriasian Münder Formation of the Deister region of Lower Saxony. It consisted only of a fragmentary mandible, but unfortunately the original specimen in the collection of Clausthal is not traceable (SS personal observation).
Similarly, Hornung and Reich (2013) described a right manus track (GZG.IF.00102), assignable to the large pterosaur ichnotaxon Purbeckopus cf. pentadactylus Delair, 1963. This positive plaster cast derived from an imprint found in upper Berriasian silicilastics of the Deister Formation near Bückeburg and was made sometime around 1935. However, the original may have never been collected, and the cast was likely produced in situ.
A fragmentary wing phalanx and jaw elements (GZG.STR.50291, GZG.STR.50292, RPMH “Wd. 025”) from the upper Berriasian Fuhse Formation near Sehnde in Lower Saxony have yet to be described in detail (Hornung 2013). Additional isolated remains have been recovered from upper Valanginian–lower Hauterivian deposits in the Leine Uplands of Lower Saxony and include what was identified as a metacarpal IV and holotype of “Ornithocheirus hilsensis” (Koken, 1883). However, a pterosaurian affinity was disputed (e.g., von Meyer 1884; Williston 1885, 1886; but see also Koken 1885, 1886), and the bone has recently been recognized as the pedal phalanx of a theropod dinosaur (Hornung 2020). Today “O. hilsensis” is considered a nomen dubium and the whereabouts of the specimen are unknown (Hornung 2020).
The comparatively best-preserved pterosaur from the Cretaceous of Germany is the holotype of Targaryendraco wiedenrothi (SMNS 56628) from the lowermost Hauterivian of the Stadthagen Formation near Hannover in Lower Saxony. Wild (1990) initially named it Ornithocheirus wiedenrothi, although its assignment to the “wastebasket” genus Ornithocheirus Seeley, 1869, is widely debated (Fletcher and Salisbury 2010; Ford 2013; Rodrigues and Kellner 2013; Abel et al. 2019; Pêgas et al. 2019). Fletcher and Salisbury (2010) grouped it in Ornithocheiridae close to Aussiedraco molnari Kellner, Rodrigues, and Costa, 2011, and another indeterminate specimen. Ford (2013) considered the species to be a “lonchodectid” and recombined it as Lonchodectes wiedenrothi. However, Rodrigues and Kellner (2013) argued against the validity of Lonchodectes and an affinity of the specimen as Ornithocheirus to be indeterminable, whereas Abel et al. (2019) reiterated a “lonchodectid” affinity. In a subsequent publication, Pêgas et al. (2019) redescribed the material as Targaryendraco wiedenrothi and found it to nest outside of Anhangueria, close to Aussiedraco molnari and Barbosania gracilirostris Elgin and Frey, 2011.
Lastly, Lanser (2015) reported several isolated teeth of putative ornithocheirids from various middle Barremian–lower Aptian karst-fillings in the Sauerland, North Rhine-Westphalia.
RE 551.763.120 A 0333/1 adds to this sparse record as the currently only recognized Valanginian pterosaur. Furthermore, it is one of only a handful Valanginian pterosaur remains in the world (Barrett et al. 2008; Cadena et al. 2020). Of these, only the probable Valanginian C. clavirostris can be confidently assigned to Anhangueria, although, other fragmentary finds may represent anhanguerians as well (Cadena et al. 2020). Finally, RE 551.763.120 A 0333/1 is, next to the teeth described by Lanser (2015), the only clear evidence for anhanguerian pterosaurs found in Germany.
Pterosaurs most frequently occur in limnic, lagoonal, and pelagic paleoenvironments (e.g., Wellnhofer 1970; Wang and Zhonghe 2006; Hone et al. 2018). Given that such depositional settings are typical of the German Cretaceous, the observed scarcity of pterosaur fossils is likely not reflecting their paleoecology, but rather the preservational biases of their fragile bones, and the reduction of Valanginian carbonate platforms in conjunction with a global cooling (Gréselle and Pittet 2010), which might have hindered the formation of Konservatlagerstätten (Butler et al. 2009). This may also explain the globally sparse record of Valanginian pterosaurs. Indeed, sampling is another evident limiting factor relative to other European Early Cretaceous pterosaur assemblages (e.g., the Cambridge Greensand, Unwin 2001; Ford and O’Connor 2004), and suggests that future exploration will likely yield new discoveries.
Conclusions
RE 551.763.120 A 0333/1 is important because it represents the only unambigious pterosaur fossil of Valanginian age recovered from Germany to date.
RE 551.763.120 A 0333/1 is among only a handful of Valanginian pterosaur occurrences yet identified worldwide.
The British Coloborhynchus clavirostris is the only other confidently assigned anhanguerian taxon from the respective time interval, representing together with RE 551.763.120 A 0333/1 the oldest known records of this group.
The sparseness of the German Cretaceous pterosaur record may be best explainable by sampling and preservation biases and does not reflect a paleobiological signal.
Acknowledgements
Achim Reisdorf (Ruhr Museum Essen, Germany) and Udo Scheer (previously Ruhr Museum Essen) generously assisted with access to specimen and historical information. We thank the editor Daniel Barta (Tahlequah, USA) and the Editorial Team from Acta Palaeontologica Polonica for handling of the manuscript, as well as Eberhard Frey (Staatliches Museum für Naturkunde Karlsruhe, Germany) and a second anonymous reviewer for their constructive comments that helped to enhance our manuscript.
References
Abel, P., Hornung, J., Kear, B., and Sachs, S. 2019. Reassessment of the enigmatic pterosaur ‘Ornithocheirus’ wiedenrothi from the Lower Cretaceous of Northern Germany. In: J.M. Stark and A. Huysseune (eds.), International Congress of Vertebrate Morphology (ICVM) Abstract Issue. Journal of Morphology Supplement 280 (S1): S73–S74.
Barrett, P.M., Butler, R.J., Edwards, N.P., and Milner, A.R. 2008. Pterosaur distribution in time and space: an atlas. Zitteliana B28: 61–107.
Bennett, S.C. 2002. Soft tissue preservation of the cranial crest of the pterosaur Germanodactylus from Solnhofen. Journal of Vertebrate Paleontology 22 (1): 43–48. Crossref
Bennett, S.C. 2007. A review of the pterosaur Ctenochasma: taxonomy and ontogeny. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 245: 23–31. Crossref
Bowerbank, J.S. 1851. On the pterodactyles of the Chalk Formation. Proceedings of the Zoological Society of London 19: 14–20. Crossref
Butler, R.J., Barrett, P.M., Nowbath, S., and Upchurch, P. 2009. Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement. Paleobiology 35: 432–446. Crossref
Cadena, E.A., Unwin, D.M., and Martill, D.M. 2020. Lower Cretaceous pterosaurs from Colombia. Cretaceous Research 114: 104526. Crossref
Chen, H., Jiang, S., Kellner, A.W.A., Cheng, X., Zhang, X., Qiu, R., Li, Y., and Wang, X. 2020. New anatomical information on Dsungaripterus weii Young, 1964 with focus on the palatal region. PeerJ 8: e8741. Crossref
Delair, J.B. 1963. Notes on Purbeck fossil footprints, with description of two hitherto unknown forms from Dorset. Proceedings of the Dorset Natural History and Archaeological Society 84: 92–100.
Elgin, R.A. and Frey, E. 2011. A new ornithocheirid, Barbosania gracilirostris gen. et sp. nov.(Pterosauria, Pterodactyloidea) from the Santana Formation (Cretaceous) of NE Brazil. Swiss Journal of Palaeontology 130: 259–275. Crossref
Erbacher, J., Hiss, M., Luppold, F.W., and Mutterlose, J. 2014a. Minden-Braunschweig-Gruppe. Litholex Online Database, Record No. 2008168. https://litholex.bgr.de/pages/Einheit.aspx?ID=2008168.
Erbacher, J., Hiss, M., Luppold, F.W., and Mutterlose, J. 2014b. Stadthagen-Formation. Litholex Online Database, Record No.: 2008147. https://litholex.bgr.de/pages/Einheit.aspx?ID=2008147.
Fletcher, T.L. and Salisbury, S.W. 2010. New pterosaur fossils from the Early Cretaceous (Albian) of Queensland, Australia. Journal of Vertebrate Paleontology 30: 1747–1759. Crossref
Ford, T.L. 2013. Will the real Lonchodectes fly in? In: J.M. Sayão, F.R. Costa, R.A.M. Bantim, and A.W.A. Kellner (eds.), Short Communications/International Symposium on Pterosaurs, Rio Ptero 2013, 65–67. Taylor & Francis, Rio de Janeiro.
Ford, T.D. and O’Connor, B. 2002. Coprolite mining in England. Geology Today 18 (5): 178–181. Crossref
1rerichs, U. 2017a. Fische. Arbeitskreis Paläontologie Hannover 45: 59–62.
Frerichs, U. 2017b. Reptilien. Arbeitskreis Paläontologie Hannover 45: 63–68.
Frerichs, U. and Girod, P. 2017. Überblick über die Fossilien aus der ehemaligen Ziegeleitongrube Sachsenhagen. Arbeitskreis Paläontologie Hannover 45: 8–14.
Frey, E.D., Stinnesbeck, W., Martill, D.M., Rivera-Sylva, H.E., and Múzquiz, H.P. 2020. The geologically youngest remains of an ornithocheirid pterosaur from the late Cenomanian (Late Cretaceous) of northeastern Mexico with implications on the paleogeography and extinction of Late Cretaceous ornithocheirids. Palaeovertebrata 43: 1–12. Crossref
Gréselle, B. and Pittet, B. 2010. Sea-level reconstructions from the Peri-Vocontian Zone (South-east France) point to Valanginian glacio-eustasy. Sedimentology 57: 1640–1684. Crossref
Hone, D.W. 2020. A review of the taxonomy and palaeoecology of the Anurognathidae (Reptilia, Pterosauria). Acta Geologica Sinica 94: 1676–1692. Crossref
Hone, D.W., Habib, M.B., and Lamanna, M.C. 2013. An annotated and illustrated catalogue of Solnhofen (Upper Jurassic, Germany) pterosaur specimens at Carnegie Museum of Natural History. Annals of Carnegie Museum 82: 165–191. Crossref
Hone, D.W., Witton, M.P., and Habib, M.B. 2018. Evidence for the Cretaceous shark Cretoxyrhina mantelli feeding on the pterosaur Pteranodon from the Niobrara Formation. PeerJ 6: e6031. Crossref
Hornung, J.J. 2013. Contributions to the Palaeobiology of the Archosaurs (Reptilia: Diapsida) from the Bückeberg Formation (“Northwest German Wealden”—Berriasian–Valanginian, Lower Cretaceous) of northern Germany. 400 pp. Unpublished Thesis, Georg August University, Göttingen.
Hornung, J.J. 2020. Comments on “Ornithocheirus hilsens” Koken, 1883 —one of the earliest dinosaur discoveries in Germany. PalArch’s Journal of Vertebrate Palaeontology 17: 1–12.
Hornung, J.J. and Reich, M. 2013. The first record of the pterosaur ichnogenus Purbeckopus in the late Berriasian (Early Cretaceous) of Northwest Germany. Ichnos 20: 164–172. Crossref
Jiang, S.-X., Wang, X.-L., Meng, X., and Cheng, X. 2014. A new boreopterid pterosaur from the Lower Cretaceous of western Liaoning, China, with a reassessment of the phylogenetic relationships of the Boreopteridae. Journal of Paleontology 88: 823–828. Crossref
Kaufmann, R., Opfermann, H.-U., and Petsch, K. 1980. Zur Entwicklungsgeschichte der tiefen Unterkreide (Berrias/Valangin) im Süden des Rehburger Sattels unter besonderer Berücksichtigung der Tongrube Sachsenhagen. Ballerstedtiana – Beitrag zur naturwissenschaftlichen Erforschung Schaumburg-Lippes und angrenzender Gebiete 3: 5–27.
Kaup, J.J. 1834. Versuch einer Eintheilung der Säugethiere in 6 Stämme und der Amphibien in 6 Ordnungen. Isis 27: 311–315.
Kellner, A.W.A. 1984. Occurrência de uma mandíbula de Pterosauria (Brasileodactylus araripensis, nov. gen, nov sp.) na Formação Santana, Cretáceo da Chapada do Araripe, Ceará, Brasil. In: 33 Anais Congresso Brasileiro de Geologia 2, 578–590. Sociedade Brasileira de Geologia, Rio de Janeiro.
Kellner, A.W.A. 1996. Pterosaur phylogeny. Journal of Vertebrate Paleontology 16 (3): 45A. Crossref
Kellner, A.W.A. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geological Society, London, Special Publications 217: 105–137. Crossref
Kellner, A.W.A. and Tomida, Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian–Albian), northeastern Brazil. National Science Museum Monographs 17: 9–137.
Kellner, A.W.A., Caldwell, M.W., Holgado, B., Dalla Vecchia, F.M., Nohra, R., Sayão, J.M., and Currie, P.J. 2019. First complete pterosaur from the Afro-Arabian continent: insight into pterodactyloid diversity. Scientific Reports 9 (1): 1–9. Crossref
Kellner, A.W.A., Rodrigues, T., and Costa, F.R. 2011. Short note on a pteranodontoid pterosaur (Pterodactyloidea) from western Queensland, Australia. Anais Da Academia Brasileira de Ciências 83 (1): 301–308. Crossref
Kemper, E., Rawson. P.F., and Thieuloy, J.-P. 1981. Ammonites of Tethyan ancestry in the early Lower Cretaceous of north-western Europe. Palaeontology 24: 251–311.
Koken, E. 1883. Die Reptilien der norddeutschen unteren Kreide. Zeitschrift Der Deutschen Geologischen Gesellschaft 35: 735–827.
Koken, E. 1885. Ueber Ornithocheirus hilsensis Koken. Zeitschrift Der Deutschen Geologischen Gesellschaft 32: 214–215.
Koken, E. 1886. Über Ornithocheirus hilsensis Koken. Zoologischer Anzeiger 9: 21–23.
Kuhn, O. 1936. Crocodilia. In: W. Quenstedt (ed.). Fossilium Catalogus I: Animalia, 75. W. Junk, Berlin.
Lanser, K.P. 2015. Nachweise von Pterosauriern aus einer unterkreidezeitlichen Karstfüllung im nördlichen Sauerland (Rheinisches Schiefergebirge, Deutschland). Geologie Und Paläontologie in Westfalen 87: 93–117.
Leonardi, G. and Borgomanero, G. 1985. Cearadactylus atrox nov. gen., nov. sp. novo Pterosauria (Pterodactyloidea) da Chapada do Araripe, Ceará, Brasil. In: D.A. Campos, C.S. Ferreira, I.M. Brito, and C.F. Viana (eds.), Trabalhos Apresentados No VIII Congresso Brasileiro De Paleontologia 1983, 75–80. Ministérios das Minas e Energia, Departamento Nacional da Produção Mineral, Brasília.
Longrich, N.R., Martill, D.M., and Andres, B. 2018. Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous–Paleogene boundary. PLOS Biology 16 (3): e2001663. Crossref
Lü, J. and Ji, Q. 2005. A new ornithocheirid from the Early Cretaceous of Liaoning Province, China. Acta Geologica Sinica (English Edition) 79: 157–163. Crossref
Marek, S. and Shulgina, N. 1996. Biostratigraphic correlation between Lower Cretaceous deposits in the central region of East-European Platform and the Polish Lowlands. Geological Quarterly 40: 129–140.
Martill, D.M., Frey, E., Diaz, G.C., and Bell, C.M. 2000. Reinterpretation of a Chilean pterosaur and the occurrence of Dsungaripteridae in South America. Geological Magazine 137: 19–25. Crossref
Meyer, O. von 1884. Ueber Ornithocheirus hilsensis Koken und über Zirkonzwillinge. Zeitschrift Der Deutschen Geologischen Gesellschaft 36: 411–416.
Meyer, H. von 1851. Ctenochasma Römeri. Palaeontographica – Beiträge zur Naturgeschichte der Vorwelt 2 (3): 82–84.
Molnar, R.E. and Thulburn, R.A. 2007. An incomplete pterosaur skull from the Cretaceous of north-central Queensland, Australia. Arquivos do Museo Nacional, Rio de Janeiro 65 (4): 461–470.
Mutterlose, J. 1984. Die Unterkreide-Aufschlüsse (Valangin–Alb) im Raum Hannover-Braunschweig. Mitteilungen aus dem Geologischen Institut der Universität Hannover 24: 1–61.
Mutterlose, J. 2017. Die ehemalige Tongrube Sachsenhagen. Arbeitskreis Paläontologie Hannover 45: 2–7.
Mutterlose, J., Wippich, M., and Geisen, M. 1997. Cretaceous depositional environments of NW Germany. Bochumer Geologische und Geotechnische Arbeiten 46: 1–134.
Myers, T.S. 2010. A new ornithocheirid pterosaur from the Upper Cretaceous (Cenomanian–Turonian) Eagle Ford Group of Texas. Journal of Vertebrate Paleontology 30: 280–287. Crossref
Myers, T.S. 2015. First North American occurrence of the toothed pteranodontoid pterosaur Cimoliopterus. Journal of Vertebrate Paleontology 35(6): e1014904. Crossref
Owen, R. 1851. A Monograph of the Fossil Reptilia of the Cretaceous Formations. Part 1, 1–118. The Palaeontographical Society, London. Crossref
Owen, R. 1874. The fossil reptilia of the Mesozoic formations. Part 1. Pterosauria. Palaeontographical Society Monographs 27: 1–14. Crossref
Pêgas, R.V., Holgado, B., and Leal, M.E.C. 2019. On Targaryendraco wiedenrothi gen. nov. (Pterodactyloidea, Pteranodontoidea, Lanceodontia) and recognition of a new cosmopolitan lineage of Cretaceous toothed pterodactyloids. Historical Biology 33 (8) [available online, https://doi.org/10.1080/08912963.2019.1690482]. Crossref
Plieninger, F. 1901. Beiträge zur Kenntnis der Flugsaurier. Palaeontographica 48: 65–90
Pentland, A.H. and Poropat, S.F. 2019. Reappraisal of Mythunga camara Molnar & Thulborn, 2007 (Pterosauria, Pterodactyloidea, Anhangueria) from the upper Albian Toolebuc Formation of Queensland, Australia. Cretaceous Research 93: 151–169. Crossref
Pentland, A.H., Poropat, S.F., Tischler, T.R., Sloan, T., Elliott, R.A., Elliott, H.A., Elliott, J.A., and Elliott, D.A. 2019. Ferrodraco lentoni gen. et sp. nov., a new ornithocheirid pterosaur from the Winton Formation (Cenomanian–lower Turonian) of Queensland, Australia. Scientific Reports 9 (1): 13454. Crossref
Pereda-Suberbiola, X., Knoll, F., Ruiz-Omeñaca, J.I., Company, J., and Fernández-Baldor, F.T. 2012. Reassessment of Prejanopterus curvirostris, a basal pterodactyloid pterosaur from the Early Cretaceous of Spain. Acta Geologica Sinica (English Edition) 86: 1389–1401. Crossref
Pinheiro, F.L. and Rodrigues, T. 2017. Anhanguera taxonomy revisited: Is our understanding of Santana Group pterosaur diversity biased by poor biological and stratigraphic control? PeerJ 5: e3285. Crossref
Rigal, S., Martill, D.M., and Sweetman, S.C. 2017. A new pterosaur specimen from the Upper Tunbridge Wells Sand Formation (Cretaceous, Valanginian) of southern England and a review of Lonchodectes sagittirostris (Owen, 1874). Geological Society, London, Special Publications 455: 221–232. Crossref
Rodrigues, T. and Kellner, A.W.A. 2013. Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England. ZooKeys 308: 1–112. Crossref
Sachs, S., Young, M.T., and Hornung, J.J. 2020. Reassessment of Enaliosuchus schroederi, a metriorhynchid crocodylomorph from the Lower Cretaceous of Northern Germany In: E. Vlachos, E. Manzanares, V.D. Crespo, C. Martínez-Pérez, H.G. Ferrón, J.L. Herráiz, A. Gamonal, F.A.M. Arnal, F. Gascó, and P. Citton (eds.), 2nd Palaeontological Virtual Congress, May 1–15th, 2020—Book of Abstracts, 141.
Schroeder, H. 1923. Ein Meereskrokodilier aus der Unteren Kreide Norddeutschlands. Jahrbuch Der Preußischen Geologischen Landesanstalt Zu Berlin 42: 352–364.
Seeley, H.G. 1869. Index to the Fossil Remains of Aves, Ornithosauria, and Reptilia, from the Secondary System of Strata, Arranged in the Woodwardian Museum of the University of Cambridge. 143 pp. Deighton, Bell, and Co., Cambridge.
Sickenberg, O. 1961. Das wiedergefundene Typusexemplar vom Meereskrokodil aus Sachsenhagen. Bericht Der Naturhistorischen Gesellschaft Hannover 105: 5–6.
Steel, L., Martill, D.M., Unwin, D.M., and Winch, J.D. 2005. A new pterodactyloid pterosaur from the Wessex Formation (Lower Cretaceous) of the Isle of Wight, England. Cretaceous Research 26: 686–698. Crossref
Tischlinger, H. 2010. Pterosaurs of the “Solnhofen” Limestone: new discoveries and the impact of changing quarrying practices. Acta Geoscientica Sinica 31: 62–63.
Tischlinger, H. and Frey, E. 2013. A new pterosaur with mosaic characters of basal and pterodactyloid Pterosauria from the upper Kimmeridgian of Painten (Upper Palatinate, Germany). Archaeopteryx 31: 1–13.
Unwin, D.M. 2001. An overview of the pterosaur assemblage from the Cambridge Greensand (Cretaceous) of Eastern England. Fossil Record 4: 189–221. Crossref
Unwin, D.M. 2002. On the systematic relationships of Cearadactylus atrox, an enigmatic Early Cretaceous pterosaur from the Santana Formation of Brazil. Fossil Record 5: 239–263. Crossref
Veldmeijer, A.J. 2003. Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil. Scripta Geologica 125 (35): e139.
Veldmeijer, A.J., Meijer, H.J., and Signore, M. 2009. Description of pterosaurian (Pterodactyloidea: Anhangueridae, Brasileodactylus) remains from the Lower Cretaceous of Brazil. Deinsea 13: 9–40.
Vidarte, C.F. and Calvo, M.M. 2010. Un nuevo pterosaurio (Pterodactyloidea) en el Cretácico Inferior de La Rioja (España). Boletín Geológico y Minero 121: 311–328.
Vila Nova, B.C., Sayão, J.M., Neumann, V.H., and Kellner, A.W.A. 2014. Redescription of Cearadactylus atrox (Pterosauria, Pterodactyloidea) from the Early Cretaceous Romualdo Formation (Santana Group) of the Araripe Basin, Brazil. Journal of Vertebrate Paleontology 34: 126–134. Crossref
Wang, X. and Zhonghe, Z. 2006. Pterosaur assemblages of the Jehol Biota and their implication for the Early Cretaceous pterosaur radiation. Geological Journal 41: 405–418. Crossref
Wellnhofer, P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Bayerische Akademie der Wissenschaften, Mathematisch-Wissenschaftliche Klasse, Abhandlungen 141: 1–133.
Wellnhofer, P. 1978. Pterosauria. Handbuch der Palaeoherpetologie, Teil 19. 82 pp. Gustav-Fischer Verlag, Lutherstadt Wittenberg.
Wellnhofer, P. 1985. Neue Pterosaurier aus der Santana-Formation (Apt) der Chapada do Araripe, Brasilien. Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie 187: 105–182.
Wellnhofer, P. 1987. New crested pterosaurs from the Lower Cretaceous of Brazil. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie 27: 175–186.
Wild, R. 1990. Ein Flugsaurierrest (Reptilia, Pterosauria) aus der Unterkreide (Hauterive) von Hannover (Niedersachsen). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 181: 241–254. Crossref
Williston, S.W. 1885. Über Ornithocheirus hilsensis Koken. Zoologischer Anzeiger 8: 628–629.
Williston, S.W. 1886. Über Ornithocheirus hilsensis Koken. Zoologischer Anzeiger 9: 282–283.
Witton, M.P. 2013. Pterosaurs: Natural History, Evolution, Anatomy. 304 pp. Princeton University Press, Oxfordshire. Crossref
Zawischa, D. 1992. Fossilien aus der Tongrube Sachsenhagen. Arbeitskreis Paläontologie Hannover 20 (2): 33–50.
Acta Palaeontol. Pol. 66 (Supplement to 3): 5–12, 2021
https://doi.org/10.4202/app.00818.2020