
Hippopotamid dispersal across the Mediterranean in the latest Miocene: a re-evaluation of the Gravitelli record from Sicily, Italy
ROBERTA MARTINO, JOHANNES PIGNATTI, LORENZO ROOK, and LUCA PANDOLFI
Martino, R., Pignatti, J., Rook, L., and Pandolfi, L. 2021. Hippopotamid dispersal across the Mediterranean in the latest Miocene: a re-evaluation of the Gravitelli record from Sicily, Italy. Acta Palaeontologica Polonica 66 (Supplement to 3): 67–78.
The first dispersal of Hippopotamidae out of Africa is recorded around 6 Ma, but this event is documented only in a few European localities. Among them, the uppermost Miocene deposits of Gravitelli in Sicily yielded particularly abundant hippopotamid remains. These specimens, published at the beginning of the 20th century, went lost during the 1908 earthquake that destroyed the city of Messina. The specimens from Gravitelli were ascribed to a new species, Hippopotamus siculus; their generic attribution was not questioned during the first half of the past century and they have not been revised in recent decades. The remains of the Gravitelli hippopotamid were mainly represented by isolated teeth and a few postcranial remains. Morphological and dimensional characters of the specimens, such as long lower premolars, low-crowned molars, a lower canine with longitudinal ridges and a groove on the lateral surface and the overall dimensions suggest that the Sicilian hippopotamid was characterized by plesiomorphic features. The morphology of the specimens collected from Gravitelli is similar to that of Hexaprotodon? crusafonti, Archaeopotamus harvardi, Hexaprotodon sivalensis and Hexaprotodon garyam. Hexaprotodon? siculus is also morphometrically similar to Hexaprotodon sivalensis, but the lower premolars in the former are longer and wider than in the latter. Accordingly, we provisionally refer the Gravitelli hippopotamid to the genus Hexaprotodon. Hexaprotodon? siculus is dimensionally different from the Spanish latest Miocene hippopotamid, herein referred to as Archaeopotamus crusafonti, and the two species are considered as valid taxa. The paleobiogeography of the latest Miocene hippopotamids from the Mediterranean Basin is discussed.
Key words: Mammalia, Hippopotamidae, dental morphology, Miocene, Gravitelli, Italy.
Roberta Martino [roberta.aska@gmail.com], via Ninfa, 9, Latina, Italy.
Johannes Pignatti [johannes.pignatti@uniroma1.it], Dipartimento di Scienze della Terra, “Sapienza” Università di Roma, Piazzale Aldo Moro 5, 00185 Rome, Italy.
Lorenzo Rook [lorenzo.rook@unifi.it] and Luca Pandolfi [luca.pandolfi@unifi.it], Dipartimento di Scienze della Terra, Università degli Studi di Firenze, Viale Giorgio La Pira 4, 50121 Florence, Italy.
Received 14 October 2020, accepted 26 March 2021, available online 10 August 2021.
Copyright © 2021 R. Martino et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The first dispersal of Hippopotamidae out of Africa took place around 6 Ma (Boisserie 2007). In Europe, hippopotamid remains occur in the uppermost Miocene deposits of Spain and Italy and they were ascribed to different species. In Spain, the scanty hippopotamid remains were referred to Hexaprotodon? crusafonti (Aguirre, 1963) (Fig. 1), whilst in Italy two different species were recorded: Hexaprotodon? pantanellii (Joleaud, 1920) from the Casino basin, Tuscany (Pantanelli 1879; Boisserie 2005) and Hexaprotodon? siculus (Hooijer, 1946) from Gravitelli, Sicily (Seguenza 1902, 1907; Hooijer 1946; Boisserie 2005; Fig. 1). In contrast to the other circum-Mediterranean records, the hippopotamid material collected at the beginning of 19th century from Gravitelli was particularly abundant. Seguenza (1902, 1907) described and figured part of the collected remains and ascribed them to Hippopotamus sivalensis Falconer and Cautley, 1836. The faunal list of Gravitelli, now attributed to the Mammal Neogene Zone 13 (MN 13), includes several mammal taxa such as Mesopithecus sp. (aff. ?Mesopithecus monspessulanus), Metailurus parvulus, Viverridae indet., Thalassictis hyaenoides, Zygolophodon borsoni (recte Mammut borsoni), Zygolophodon turicensis, Diceros cf. D. pachygnathus (recte Ceratotherium sp.), Reduncini indet., ?Gazella deperdita, ?Parabos sp., and Microstonyx major erymanthius (recte Propotamochoerus sp.) (Rook 1992; Kotsakis et al. 1997; Rook 1999; Van der Made 1999; Gallai and Rook 2006; Rook et al. 2006; Pandolfi and Rook 2017; Pandolfi et al. 2021). The faunal assemblage of Gravitelli is considered close to that from Cessaniti (Calabria) and As Sahabi (Libya) and has been used for paleobiogeographic considerations on dispersal events of latest Miocene mammals from Africa to Europe (Bernor and Rook 2008; Marra et al. 2017). Unfortunately, all the specimens described by Seguenza (1902, 1907) went lost during the 1908 earthquake that destroyed the city of Messina, and all subsequent considerations on the mammal remains from Gravitelli are based only on the published figures. Hooijer (1946) revised the works by Seguenza (1902, 1907) and, based on morphological traits, erected the new hippopotamid species Hippopotamus siculus, which was later provisionally assigned to Hexaprotodon (Boisserie 2005). Nevertheless, the hippopotamid from Gravitelli has not been revised in the last decades.
The hippopotamid remains published by Seguenza (1902, 1907) are here revised in order to clarify and update their systematic position and their paleobiogeographic implications.
Institutional abbreviations.—MSNAF, Museo di Storia Naturale dell’Accademia dei Fisiocritici, Siena, Italy; RMCA, Royal Museum of Central Africa, Tervuren, Belgium.
Other abbreviations.—C/c, canine; DP/dp, deciduous upper/lower premolar; MN, Mammal Neogene Zone; M/m, upper/lower molar; P/p, upper/lower premolar.
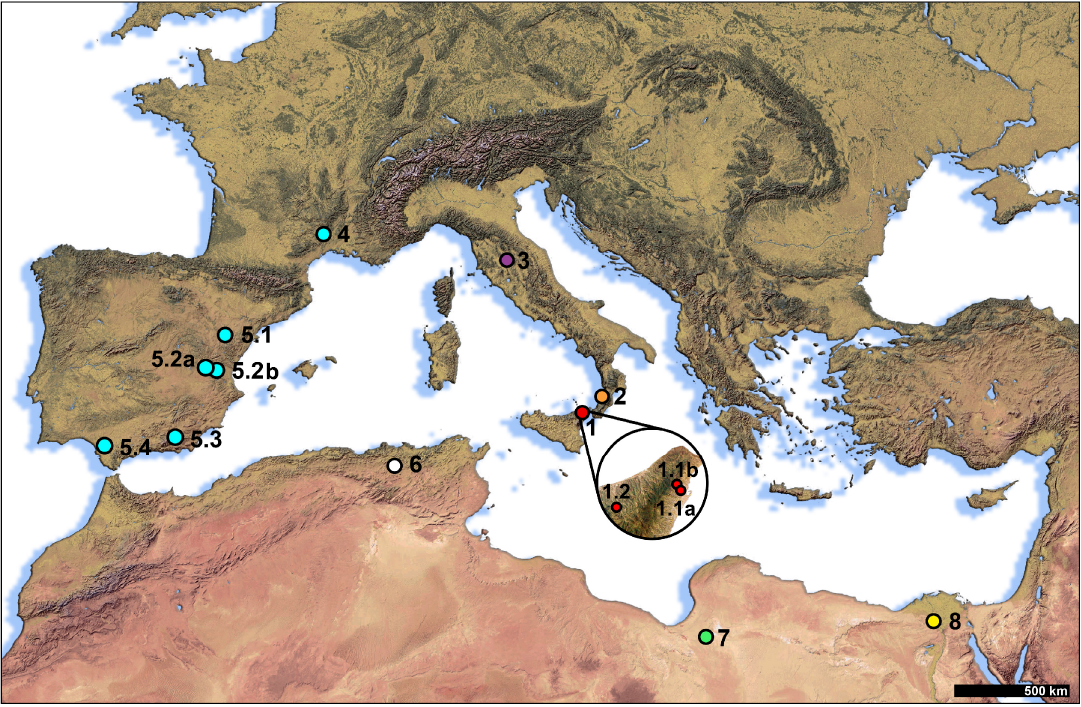
Fig. 1. Selected records of circum-Mediterranean hippopotamids during the latest Miocene–earliest Pliocene. 1, Hexaprotodon? siculus (Hooijer, 1946) (1.1a, Gravitelli and 1.1b Scirpi; San Pier Niceto) Messina, Sicily, late Miocene; 2, ?Hippopotamidae indet., Cessaniti, Calabria, late Miocene; 3, Hexaprotodon? pantanellii (Joleaud, 1920), Casino Basin, Tuscany, late Miocene; 4, Archaeopotamus crusafonti (Aguirre, 1963), La Mosson (Montpellier), France, early Pliocene; 5, Archaeopotamus crusafonti (Aguirre, 1963) (5.1, Las Casiones, Teruel; 5.2a, Venta del Moro and 5.2b, La Portera, Valencia; 5.3, Arenas del Rey, Granada; 5.4, El Arquillo, Siviglia), Spain, late Miocene; 6, Hexaprotodon? hipponensis (Gaudry, 1876), Pont-de-Duvivier, Algeria, early Pliocene; 7, Hexaprotodon? sahabiensis (Gaziry, 1987), As Sahabi, Libya, late Miocene; 8, Hexaprotodon protamphibius andrewsi (Arambourg, 1947), Egypt, early Pliocene.
Material and methods
The morphological terminology for the teeth follows Thenius (1989) and Boisserie et al. (2010) (Fig. 2). The morphological terminology for the postcranial remains follows Mazza (1995). The material collected from Gravitelli was published by Seguenza (1902, 1907). Descriptions and figures reported by Seguenza (1902, 1907) are scarce and most of the remains are represented only in part (Figs. 3, 4). The revised remains are morphologically and morphometrically compared with late Miocene and early Pliocene hippopotamids: Hexaprotodon garyam Boisserie, Likius, Vignaud, and Brunet, 2005, Hexaprotodon sivalensis (Falconer and Cautley, 1836), Saotherium mingoz Boisserie, 2005, Archaeopotamus harvardi (Coryndon, 1977), A. lothagamensis (Weston, 2000), A. qeshta Boisserie in Boisserie et al., 2017, Hexaprotodon? crusafonti (Aguirre, 1963), Hexaprotodon? sahabiensis (Gaziry, 1987), Hexaprotodon? hipponensis (Gaudry, 1876), Hexaprotodon protamphibius (Arambourg, 1944), and Hexaprotodon protamphibius andrewsi (Arambourg, 1947). Hexaprotodon? siculus remains are also compared with the living species Hippopotamus amphibius Linnaeus, 1758; and Choeropsis liberiensis (Morton, 1844). The measurements of Hexaprotodon? siculus were published by Seguenza (1902, 1907).
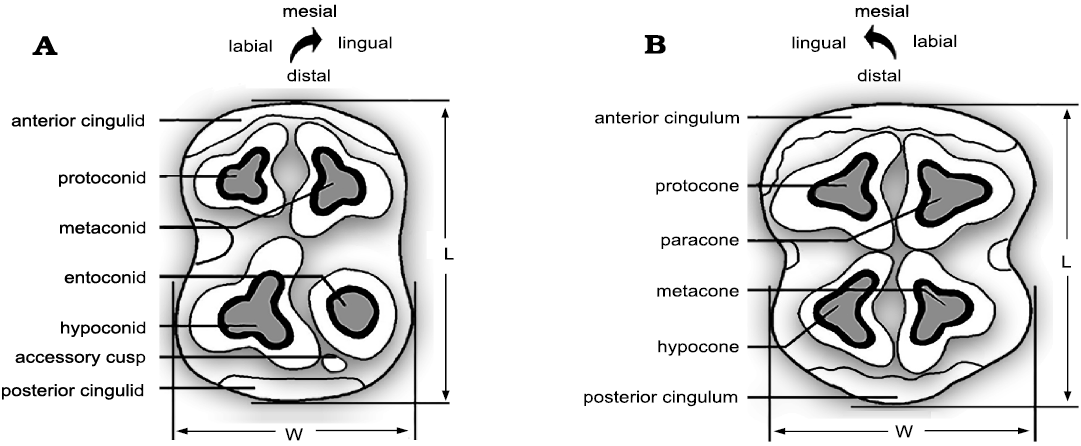
Fig. 2. Hippopotamid cheek teeth nomenclature based on the left m2 and M2 in occlusal view. Accessory cusps may be present or absent on upper and lower teeth. The m3 is characterized by a fifth distal cusp called hypocolunid. Modified from Htike (2012).
Systematic palaeontology
Cetartiodactyla Montgelard, Catzeflis, and Douzery, 1997
Cetancodonta Arnason, Gulerg, Solweig, Ursing, and Janke, 2000
Hippopotamoidea Gray, 1821 (sensu Gentry and Hooker 1988)
Hippopotamidae Gray, 1821
Hippopotaminae Gray, 1821
Genus Hexaprotodon Falconer and Cautley, 1836
Type species: Hexaprotodon sivalensis Falconer and Cautley, 1836, from Mio-Pliocene strata of the Siwalik Hills, India/Pakistan.
Hexaprotodon? siculus (Hooijer, 1946)
Figs. 3–5, 6B1, B2.
1902 Hippopotamus (Hexaprotodon) sivalensis Falconer and Cautley, 1836; Seguenza 1902: 115–175, pl. 7: 1–14, 20, 22, 23.
1907 Hippopotamus (Hexaprotodon) sivalensis Falconer and Cautley, 1836; Seguenza 1907: 89–122, pl. 6: 1–22, pl. 7: 1–16.
1946 Hippopotamus siculus sp. nov.; Hooijer 1946: 301–319.
2005 Hexaprotodon? siculus (Hooijer, 1946); Boisserie 2005: 143.
Material.—Material from the late Miocene of Sicily, listed in Seguenza (1902: pl. 7; here Fig. 3): a maxillary fragment with DP3, DP4, M1 (pl. 7: 1–3) and a fragment of a DP? (pl. 7: 20) from San Pier Niceto; left astragalus (pl. 7: 4–7, 8, 9) from Scirpi or Gravitelli; a partial M (pl. 7: 10, 11), an C (pl. 7: 13, 14), and an unciform (pl. 7: 22, 23) from Gravitelli. Material from late Miocene of Sicily (Gravitelli site), listed in Seguenza (1907: pls. 6 and 7; here Figs. 4 and 5 respectively): mandible fragment with a m1 (pl. 6: 1–3), a cervical vertebra (pl. 6: 4–6), a M3 (pl. 6: 7, 8), a M1 or M2 (pl. 6: 9, 10), a C fragment (pl. 6: 11), two dp3 (pl. 6: 14–16), a P2 (pl. 6: 17, 18), a dp (pl. 6: 19, 20), m2–m3 (pl. 7: 1, 2), a m1 (pl. 7: 3, 4), a p2 (pl. 7: 5, 6), a p3 (pl. 7: 7, 8), a p4 (pl. 7: 9, 10), a c (pl. 7: 11, 12), and some incisor fragments (pl. 7: 13–16), a fragment of a radius (pl. 5: 49, 50).
Seguenza (1907) attributed to Hippopotamus a portion of a distal radius (pl. 5: 49, 50) a proximal part of a radius (pl. 5: 51, 52), a scapula fragment (pl. 6: 12, 13), and a distal part of a metacarpal (pl. 6: 21, 22) from late Miocene of Sicily (Gravitelli site). Hooijer (1946) attributed the distal part and the proximal part of the radius to Parabos? and the scapula fragment together with the distal part of a metacarpal to a rhinoceros. We do not agree with the attribution of the radius to a bovid, due to both morphological and morphometric traits of the figured bone, and we include it within the Hippopotamidae material.
Description.—Dental characters cannot be easily recognized due to the impossibility to observe the original specimens lost in 1908. Photos and descriptions in Seguenza (1902, 1907) are therefore the only documentation of the presence of a hippopotamid in Sicily during the late Miocene (Figs. 3, 4, 5). Deciduous premolars do not display useful diagnostic features. Four fragmentary incisors were figured by Seguenza (1907; Fig. 5). According to the available text and figures, two grooves, one on each side, are present on the lower incisors. Seguenza (1907: 116–117) reported the presence of six different partial incisors that were all collected from a single mandible, completely destroyed during the excavation. This statement, anyway, testifies to the hexaprotodonty (presence of six incisors, while the extant Hippopotamus amphibius is characterized by four incisors, the tetraprotodont condition) of the Sicilian species. The upper canines are characterized by a deep posterior groove and by two less defined lateral grooves, one on the medial side and one on the lateral side (Fig. 3). A lower canine fragment described by Seguenza (1907) displays a rough enamel, longitudinal striae and transversal growth striae (Fig. 4). The lower canine has longitudinal ridges and a groove on the lateral surface (Fig. 5). The P2 is mostly triangular and simply built (Fig. 4). The lower premolars (p2 and p3) are mainly triangular and partially incomplete (Fig. 5). The p2 has a distolingual cusp. The p3 is characterized by a distolingual cusp surrounded by a crenulated cingulid and more developed than one of the p2. The p4 of the Sicilian species is broken and worn. A single cusp is visible, but a second lower and less developed cusp was probably present distally. In lateral view the cingulid is strongly elevated whilst, in occlusal view, some well-developed cristae are visible in the distal part of the tooth. All molars from Gravitelli are low crowned. The M1 in the maxillary fragment displays a thick crenulated cingulum and a finely striated enamel (Fig. 3). M1–M2 (Fig. 4) is characterized by a trefoil wear pattern not completely developed. The enamel is relatively thick on the protocone and the cingulum is crenulated on the lingual side. An upper molar, referred to M2 by Hooijer (1946), is partially broken, and only the metacone and paracone are present (Fig. 3). The cingulum is crenulated on these cusps and the enamel is finely striated. The posterior cusps of M3 (Fig. 4) are narrower than the anterior cups (Hooijer 1946). This tooth is unworn and the crown is not particularly high; the cusps are simple with not particularly well-developed grooves. The cingulum is crenulated, and it is higher and more complex on the posterior cusps compared to the anterior cups.
The lower molars from Gravitelli are low-crowned. The m1 displays a longitudinal valley between metaconid and protoconid, located labial to the valley between entoconid and hypoconid (Hooijer 1946; Fig. 4). The unworn cusps are particularly slender, the cingulid is crenulated and only visible on the posterior cusps. The m3 and m2 were partially figured by Seguenza (1907), who did not provide their occlusal view (Fig. 5). The m2 does not display a posterior lobe on the entoconid. The m3 is characterized by slender cusps and a large hypoconulid. The cingulid on this tooth is crenulated and higher on the labial side of the hypoconulid. The cusps are almost unworn, and the enamel is finely striated.
The vertebra cervicalis is severely damaged (Fig. 4). It displays a ventral spine that is not particularly prominent. The posterior face of the vertebra is larger than the anterior. The radius is broken and displays on the distal epiphysis an antero-posteriorly oriented crest between the articular surfaces for the scaphoid and with the lunar (Seguenza 1907: pl. 5: 49, 50). The unciform displays a concave articulation with the cuneiform and a narrow articulation with the lunar. Two astragali were figured by Seguenza (1902), one collected from Gravitelli and the other one from Scirpi (Fig. 3). Unfortunately, one astragalus lacks a part of the proximal throclea and the other astragalus was only figured in plantar view by Seguenza (1902). These bones display a comparable size of the oblique articular surface for the cuboid and of the articular surface for the navicular. The complete astragalus is rather subquadrangular in plantar view; on this specimen it is also possible to observe a stop facet that could limit the degree of flexion of the tarsus.
Stratigraphic and geographic range.—Upper Miocene of Gravitelli, San Pier Niceto, and Scirpi (Messina, Sicily, South Italy), European Mammal Neogene Zone 13.

Fig. 3. Late Miocene hippopotamid Hexaprotodon? siculus (Hooijer, 1946) from Sicily, Italy; original plate from Seguenza (1902: pl. 7). Maxillary fragment with DP3, DP4, and M1 from San Pier Niceto, in lingual (1), occlusal (2), and labial (3) views. Fragment of a upper deciduous? tooth from San Pier Niceto (20), two left astragali one from Gravitelli and one from Scirpi (4, anterior view; 5 and 8, medial view; 6, lateral view; 7 and 9, posterior view), partial left M from Gravitelli (10, labial view; 11, occlusal view), C section from Gravitelli (12, view of the transverse section), C and its section from Gravitelli (13, labial view; 14, view of the transverse section), unciform from Gravitelli (22, lateral? view; 23, anterior view).

Fig. 4. Late Miocene hippopotamid Hexaprotodon? siculus (Hooijer, 1946) from Gravitelli, Sicily, Italy; original plate from Seguenza (1907: pl. 6). Mandible fragment with M1, in labial (1), occlusal (2), and lingual (3) views, cervical vertebra in ventral (4), caudal (5), and cranial (6) views, left M3 (7, labial view; 8, occlusal view), right M1–M2 (9, labial view; 10, occlusal view), C fragment (11, labial? view), right dp3 (14, labial view; 15, occlusal view), left dp3 (16, lingual view), P2 (17, lingual view; 18, occlusal view), dp (19, lingual view; 20, occlusal view).
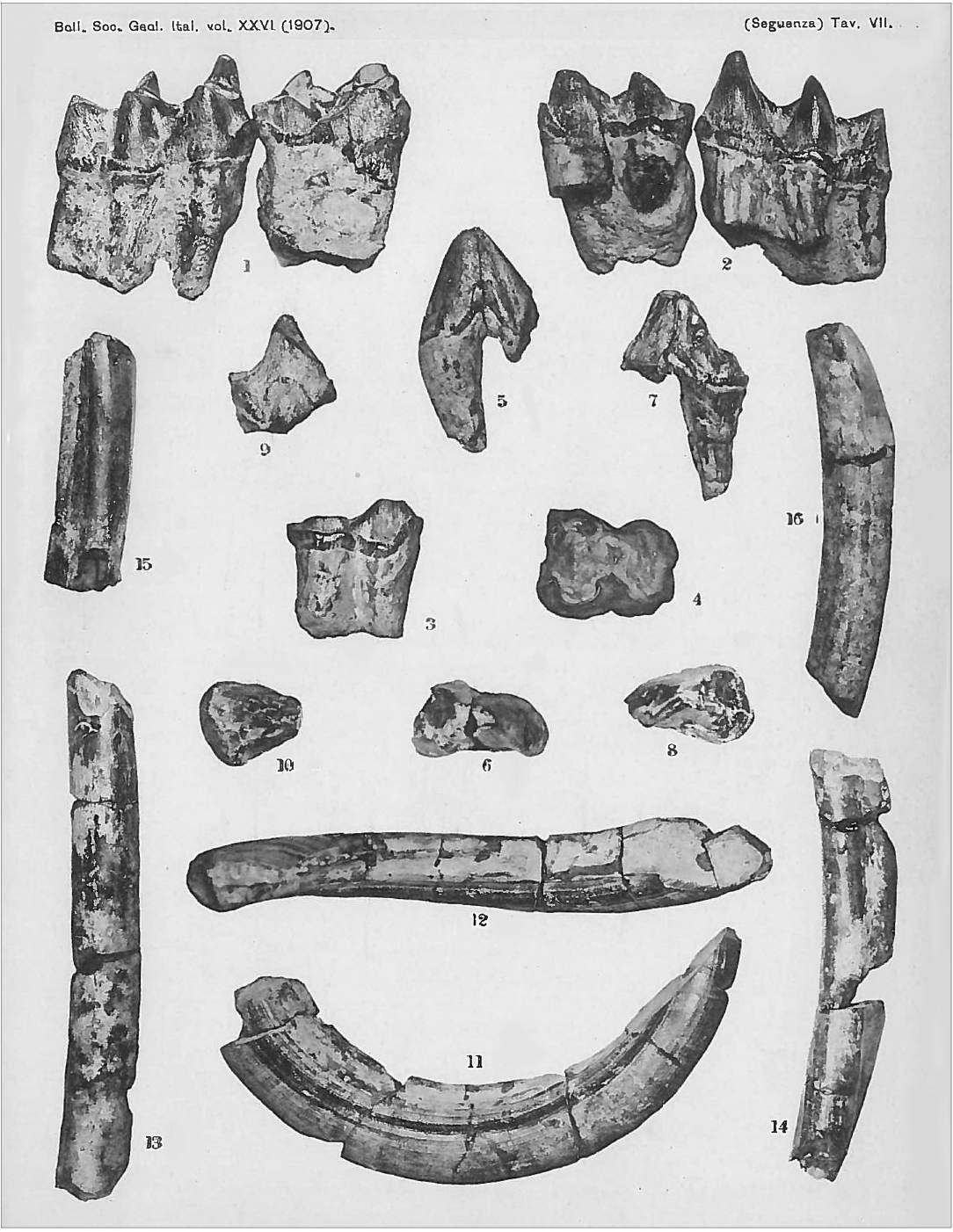
Fig. 5. Late Miocene hippopotamid Hexaprotodon? siculus (Hooijer, 1946) remains from Gravitelli, Sicily, Italy; original plate from Seguenza (1907: pl. 7). Left m2 and m3 in labial (1) and lingual (2) views, right m1 (3, labial view; 4, occlusal view); left p2r (r, refuse) (5, labial view; 6, occlusal view), right p3 (7, lingual view; 8, occlusal view), right p4 (9, labial view; 10, occlusal view), lower canine (11, lingual view; 12, dorsal view), and some incisor fragments (13–16, lateral view).
Discussion
Comparison with late Miocene–early Pliocene peri-Mediterranean hippopotamids.—The morphological comparison highlights an affinity of the Sicilian hippopotamus with Hexaprotodon? crusafonti. The upper canine section typical of the Gravitelli Hexaprotodon? siculus specimens is also shared by He.? crusafonti (Aguirre 1963; Fig. 6). Nevertheless, a deep posterior groove is also displayed in species confidently attributed to the Hexaprotodon (Boisserie 2005), and in the Saotherium (Boisserie et al. 2003) and Archaeopotamus (Weston 2000, 2003). This character can therefore be considered as a plesiomorphic feature within the Hippopotamidae and is thus not diagnostic at the genus level. The lower canine from Gravitelli displays longitudinal ridges and a groove on the lateral surface as in Archaeopotamus harvardi (see Weston 2003). In Hexaprotodon sivalensis several specimens do not display the groove on the lateral side, but this character is variable, being documented in some fossil remains (De Visser 2008). However, a shallow groove on the lateral side of the lower canine is more or less developed in several hippopotamid species and usually well-expressed in later ontogenetic stages (Boisserie 2005). The enamel of Saotherium mingoz is slightly ridged or smooth and without a well-defined groove (Boisserie et al. 2003). P2 from Gravitelli is wide, mostly triangular in shape, with a main singular cusp as in Hexaprotodon sivalensis, Archaeopotamus harvardi, Hexaprotodon garyam, and Saotherium mingoz (Hooijer 1950; Boisserie et al. 2003, 2005; Weston 2003). The p2 from Gravitelli is wider and longer than in He. sivalensis (Table 1). The distolingual cusp displayed by the studied p3 also characterizes He. sivalensis, S. mingoz, A. harvardi, He.? crusafonti, and He. garyam (Hooijer 1950; Lacomba et al. 1986; Boisserie et al. 2003, 2005; Weston 2003); this tooth is distally wide as in A. harvardi and He.? crusafonti (Aguirre 1963; Lacomba et al. 1986; Weston 2000, 2003). The p3 in Hexaprotodon? hipponensis shows more tubercles and a lingual cusp more developed than in He.? siculus (Gaudry 1876). The length of p3 from Gravitelli is similar to that of A. harvardi, whereas it is smaller in He. sivalensis (Table 1). The fourth lower premolar in He.? siculus is distinctly wider in its distal part; this character is also shared by He.? crusafonti, He. garyam, and occasionally, A. harvardi (Lacomba et al. 1986; Weston 2003; Boisserie et al. 2005). Dimensionally, the p4 from Gravitelli is closer to A. harvardi and He. garyam, whereas the dimensions of He. sivalensis are considerably smaller (Table 1, Fig. 7). The lower premolars of He.? siculus are morphometrically closer to A. harvardi and He. garyam, whereas the dimensions of He. sivalensis are always smaller than in the Sicilian specimens. The grooves on M3 from Gravitelli are less developed than in He. sivalensis and He.? hipponensis (Hooijer 1946). The crown on M3 from Gravitelli is more brachyodont than in He. sivalensis. Low-crowned teeth are typical of African species such as A. harvardi and He. garyam, whereas Asian hippos display higher crowned teeth (Boisserie 2005). Low-crowned teeth are also typical of the late Miocene species He.? crusafonti (Aguirre 1963; Lacomba et al. 1986). In general, the observed trend in the Hippopotamidae is towards higher crowned molars (Boisserie 2005). In the third lower molar of A. harvardi the cingulid is not continuous and may be separated from the mesial and distal cusps by a ridge (Weston 2003), as in the Sicilian species. In the Gravitelli hippopotamid, the cusps on the m3 are more slender and the hypoconulid is more developed than in He. sivalensis. The morphology of the third lower molar resembles that of He.? crusafonti in lingual and labial views. Dimensionally, the m3 from Gravitelli is close to A. harvardi, He. sivalensis, He.? sahabiensis, and He. garyam (Hooijer 1950; Gaziry 1987; Weston 2003; Boisserie et al. 2005; Table 1). The stop facet observed on the astragalus of the Sicilian species is also documented on some specimens from Lothagam, attributed to A. harvardi (Weston 2003). The height difference between the articular surface for the cuboid and the articular surface for the navicular in A. harvardi is similar to that displayed by the Sicilian specimens, whereas in He. sivalensis the height difference is greater. A single astragalus attributed to Hexaprotodon? crusafonti was described by Alcalá (1994) from Las Casiones (Spain). Similarly, to the specimens from Sicily, the height between the articulations of the navicular and cuboid is comparable. A single He. protamphibius andrewsi astragalus is figured by Stromer (1914). This bone is less quadrangular, smaller and more slender than in the Sicilian specimens and it possibly lacks the stop facet. In addition, the astragalus of He. protamphibius andrewsi displays a well-developed tuberosity below the medial ridge, similarly to He. sivalensis. The astragalus of the Sicilian species is morphologically more similar to that of Hippopotamus amphibius than to Choeropsis liberiensis (Fig. 8). Some studies highlight that astragalus morphology in bovids is a useful tool for paleohabitat reconstructions (DeGusta and Vrba 2003 and references therein). A study based on hippopotamuses’ astragali has not yet been performed, but it could probably test terrestrial versus aquatic adaptations in the different fossil hippopotamid species. The astragalus index ([max. width = 84 mm/max. length = 109 mm] × 100 = 77; measured by Seguenza 1902) in the Gravitelli hippopotamid is close to the values of He. protamphibius andrewsi, A. harvardi, He. sivalensis, and He. protamphibius (Stromer 1914; Hooijer 1946; Harrison 1997). The radius of He. sivalensis displays a distal crest less directed antero-posteriorly than in the material from Gravitelli (Hooijer 1946). In addition, the articulation between cuneiform and unciform is less concave and the articulation between unciform and lunar is narrower in He. sivalensis than in the Sicilian specimens (Hooijer 1946).
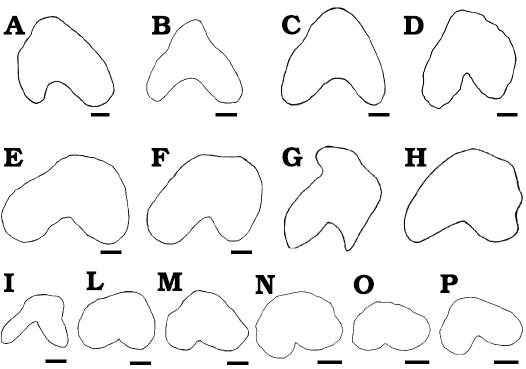
Fig. 6. Upper canine of various hippopotamids in cross section. A. Archaeopotamus crusafonti (Aguirre, 1963), Arenas del Rey (Spain), late Miocene (Aguirre 1963). B. Hexaprotodon? siculus (Hooijer, 1946), Gravitelli (Sicily, Italy), late Miocene (Seguenza 1902; Hooijer 1946). C. Hexaprotodon? siculus (Hooijer, 1946), Gravitelli (Sicily, Italy), late Miocene (Seguenza 1902; Hooijer 1946). D. Hexaprotodon garyam Boisserie, Likius, Vignaud, and Brunet, 2005, Toros-Menalla (Chad), late Miocene (Boisserie et al. 2005). E. Hexaprotodon sivalensis (Falconer and Cautley, 1836), Siwalik Hills (India/Pakistan), late Miocene–early Pliocene (Hooijer 1946). F. Hexaprotodon sivalensis (Falconer and Cautley, 1836), Siwalik Hills (India/Pakistan), late Miocene–early Pliocene (Hooijer 1946). G. Subadult Archaeopotamus harvardi (Coryndon, 1977), Lothagam (Kenya), late Miocene (Weston 2003). H. Adult Archaeopotamus harvardi (Coryndon, 1977), Lothagam (Kenya), late Miocene (Weston 2003). I. Choeropsis liberiensis (Morton, 1844), Leiden Museum collection (Hooijer 1946). L. Hexaprotodon? protamphibius andrewsi (Arambourg, 1947), Wadi-Natrum (Egypt), early Pliocene (Stromer 1914). M. Hippopotamus amphibius Linnaeus, 1758, Leiden Museum collection (after Hooijer 1946). N–P. Subadult Hexaprotodon amphibius Linnaeus, 1758, Zaire, Holocene; original data sampled by LP at the Royal Museum of Central Africa, Tervuren, Belgium. RMCA n. A6035M127 (N), RMCA n. A6035M194 (O), RMCA n. A6035M190 (P). Scale bars 10 mm. A, scale redrawn form Aguirre (1963); B,C, E, F, I, L, M, scales redrawn from Hooijer (1946); D, scale redrawn from Boisserie et al. (2005); G, H, scale not provided in Weston (2003).
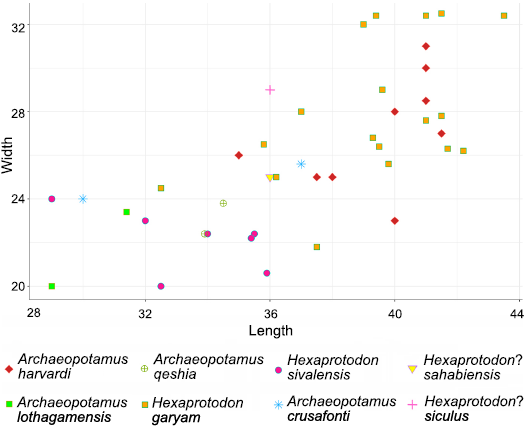
Fig. 7. Length-width diagram (in mm) of p4 in different hippopotamid species. Measurements from the literature (see Table 1).
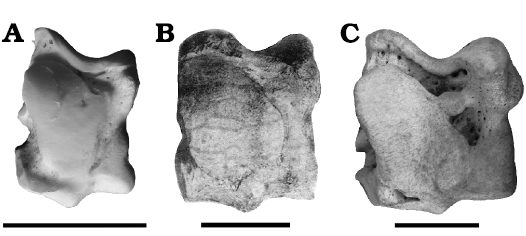
Fig. 8. Astragali of hippopotamids in plantar view. A. Choeropsis liberiensis (Morton, 1844) (RMCA n. RG35716), Holocene, specimen from zoo, Recent. B. Hexaprotodon? siculus (Hooijer, 1946) (Seguenza 1902: pl. 7: 9), late Miocene, Gravitelli or Scirpi, Sicily, Italy. C. Hippopotamus amphibius Linnaeus, 1758 (RMCA n. RG508), Holocene, Zaire. Scale bar in B estimated according to the maximum height of the astragalus (97 mm) indicated by Seguenza (1902). Scale bars 50 mm.
Table 1. Comparative dimensions of upper and lower teeth of Miocene hippopotamids (minimum–maximum dimension in mm; mean; number of specimens). Data for Hexaprotodon? siculus from Seguenza (1902, 1907), Hooijer (1946, 1950); Archaeopotamus crusafonti from Aguirre (1963); Archaeopotamus crusafonti from Alcalá et al. (1986); Hexaprotodon? sahabiensis from Gaziry (1987), Pavlakis (2008); Archaeopotamus harvardi from Coryndon (1977), Archaeopotamus lothagamensis from Weston (2000, 2003); Hexaprotodon garyam from Boisserie et al. (2005); Hexaprotodon sivalensis from De Visser (2008); Archaeopotamus qeshta from Boisserie et al. (2017). Values of Hexaprotodon? pantanelli were measured at MSNAF (n.2828). The second lower premolar reported of Hexaprotodon? pantanellii is partially broken and it was reported in Pantanelli (1879) as 32 mm but the specimen measured at MSNAF is 29.56 mm; this tooth was probably broken after Pantanelli (1879). Abbreviations: L, length, W, width.
|
Species/ tooth |
He.? siculus |
He.? |
A. crusafonti |
He.? |
He.? |
A. |
A. lothagamensis |
A. |
He. |
He. |
|
|
p2 |
L |
>41.7 |
>29.56–32(?) |
41 |
|
|
31.3–41; 36.27; 6 |
|
29.2–40.2; 34.7; 2 |
29.8–47.1; 36; 17 |
31–34; 32.5; 2 |
|
W |
27 |
19.69 |
? |
|
|
21.4–24.7; 22.85; 5 |
|
18.2–19.7; 18.95; 2 |
19–27.6; 22.12; 17 |
19.5; 19.5; 2 |
|
|
p3 |
L |
>40.6 |
|
37.5 |
|
35 |
36.5–45.1; 40.04; 6 |
|
35.7–39.1; 37.4; 2 |
31.7–48; 39.42; 20 |
31.3–39; 35.15; 2 |
|
W |
26 |
|
21 |
|
|
22.5–26.6; 24.62; 6 |
|
19.6–23.1; 21.35; 2 |
18–32.9; 24.17; 19 |
19.2 |
|
|
p4 |
L |
36 |
|
30–37; |
36 |
|
36.4–41.6; 38.66; 8 |
30.7–33.6; 32.15; 2 |
33.9–34.5; 34.2;2 |
33.4– 43.1; 39.1; 19 |
29.3–35.9; 32.25; 8 |
|
W |
29 |
|
24–25.6; 24.8;2 |
25 |
|
24.9–30.6; 27.41; 8 |
20.5–23.8; 22.15; 2 |
22.4–23.8; 23.1; 2 |
21.7– 32.5; 27.81; 18 |
20.6–22.4; 22.33; 7 |
|
|
m1 |
L |
40.5–41; 40.75; 2 |
|
36–40.5; 38.25; 2 |
36–36.42; 36.21; 2 |
|
35.5–41; 38.25; 8 |
36.5–40; 38.25; 2 |
|
23–28; 25.5; 20 |
31–41; 36.67; 3 |
|
W |
28–32; 30; 2 |
|
25–26.5; 25.75; 2 |
25.78–30; 27.89; 2 |
|
26.1–35; 31.34; 9 |
24–30; 28; 2 |
25.2–25.7; 25.45; 2 |
19–23; 21; 20 |
24–33; 29; 4 |
|
|
m2 |
L |
46–50; 48; 2 |
44.6 |
37–40.4; 37.43; 3 |
39 |
|
41.2–51.3; 47.44; 11 |
41.1–42.7; 41.90; 2 |
41.5–44.8; 43.15; 2 |
41.6–54; 49.28; 31 |
43–52; 48.17; 9 |
|
W |
35–39; 37; 2 |
33 |
28–29; |
34 |
|
33–38.4; 36.16; 10 |
27.8–34.0; 30.90; 2 |
31.8–33.7; 32.75; 2 |
30.8–45.2; 37.53; 26 |
31–40; 36.5; 10 |
|
|
m3 |
L |
63 |
|
>53–55; |
59 |
|
58.3–68; 63.1; 16 |
49.8–51.1; 50.45; 2 |
54.9–62.9; 58.9; 2 |
59.1–70.7; 63.26; 31 |
56–72; 65.55; 12 |
|
W |
35 |
|
28.2–>36.1; 29.1; 3 |
33 |
|
32–40.2; 35.9; 15 |
28.3–29.2; 28.75; 2 |
31.5 |
31.9–45.2; 38.12; 31 |
33–42; 38.55 ;12 |
|
|
M1 |
L |
40 |
|
|
34 |
|
35–46; 40.5; 11 |
|
|
36–49; 42.5; 21 |
31–46; 41.86; 10 |
|
W |
37 |
|
|
33 |
|
38–46; |
|
|
31–43; 37; 21 |
36–41; 38.5; 10 |
|
|
M3 |
L |
44 |
|
|
39 |
|
44.0–51.7; 47.17; 13 |
|
|
37.8–57.4; 47.74; 49 |
36.8–51; 47; 10 |
|
W |
44 |
|
|
42 |
|
41.4–56.2; 46.48; 13 |
|
|
40.8–52.2; 46.54; 48 |
35.15–52; 48.4; 10 |
|
An overview of late Miocene–early Pliocene hippopotamids from the circum-Mediterranean area.—The morphology of the specimens collected from Gravitelli and assigned as Hexaprotodon? siculus is similar to that of He.? crusafonti, Archaeopotamus harvardi, He. sivalensis, and He. garyam. The dimensions of the cranial remains from Gravitelli fall within the variability of A. harvardi and He. garyam. Hexaprotodon? siculus is also morphometrically similar to He. sivalensis but the lower premolars in the former are always longer and larger than in the latter. Accordingly, and considering its hexaprotodont condition, we tentatively refer the Gravitelli hippopotamid to the genus Hexaprotodon in agreement with Boisserie (2005).
Due to the tetraprotodont condition and the clear morphometrical differences with Hexaprotodon? siculus, the Spanish Miocene hippopotamid He.? crusafonti is considered as a valid species. Hexaprotodon? crusafonti was firstly described from the upper Miocene deposit of Arenas del Rey by Aguirre (1963). This Spanish species was later identified at Venta del Moro (Aguirre et al. 1973, Morales 1984), El Arquillo (Crusafont et al. 1964; Alcalá and Montoya 1998), Las Casiones (Alcalá 1994) and La Portera (Lacomba et al. 1986). Hexaprotodon? crusafonti was also collected from the lower Pliocene deposit of La Mosson, France (Faure and Méon 1984). The material from La Portera (Lacomba et al. 1986) includes a left semi-complete hemimandible with the mandibular symphysis, two broken first incisors and two second incisor alveoli, suggesting the tetraprotodont condition of this species. Two important features characterize this taxon: a premolar row longer than the molar row and a mandibular symphysis that is long and narrow. These characters are both diagnostic of the Archaeopotamus (Boisserie 2005). The tetraprotodont condition displayed by the Spanish species could suggest an evolutionary stage more advanced than the African coeval forms, such has A. harvardi, A. lothagamensis, and He. garyam. Hexaprotodon protamphibius collected from different geological sequences from Shungura (Ethiopia) testifies to a well-defined evolutionary trend in hippopotamids: from an archaic hexaprotodont condition towards a more advanced tetraprotodont condition (Coryndon 1978). Hexaprotodon? crusafonti is therefore closer to Archaeopotamus than to species confidently attributed to Hexaprotodon (as previously suggested by Weston 2000, 2003), and it could thus be more properly referred to as Archaeopotamus crusafonti. The re-attribution of these specimens to Archaeopotamus suggests that this genus originally recorded in Africa (Kenya and possibly Tanzania) and Arabia (Abu Dhabi) was also present in Southern Europe (Harrison 1997; Weston 2003; Boisserie 2005; Boisserie et al. 2017). The peri-Mediterranean area was intensively colonized by hippopotamids during the latest Miocene. In addition to the Italian species (He.? siculus and He.? pantanellii) and the Spanish species (A. crusafonti), some other forms were collected from Libya (He.? sahabiensis, Gaziry 1987), Algeria (He.? hipponensis, Gaudry 1876) and Egypt (He. protamphibius andrewsi; Andrews 1902; Stromer 1914; Arambourg 1947) (Fig. 1). Since these remains are scarce and mainly fragmentary, a proper and exhaustive study is problematic. Remains attributed to He.? pantanellii and ascribed to the late Miocene (late MN13) were collected during the 19th century from the Casino Basin, Siena, Italy (Pantanelli 1879; Joleaud 1920; Boisserie 2005; Gallai 2005). These remains include a mandibular symphysis fragment with four broken incisors, some isolated incisors, a broken second lower premolar, a second lower molar and a fragment of a lower canine. Unfortunately, the original second lower molar described and figured by Pantanelli (1879) was lost. The scarce and fragmentary record from Casino does not support the validity of He.? pantanellii as a separate species, and these specimens should be, more cautiously, assigned to Hippopotamidae indet. This taxon probably arrived through the Iberian Peninsula and colonized Tuscany during the Messinian (Martino et al. 2020). The remains from Gravitelli, along with some fragmentary elements from the Casino Basin in Tuscany (Martino et al. 2020), are the only evidence of the presence of hippopotamids in Italy during the late Miocene. Hippopotamid remains were also doubtfully reported from Cessaniti, Calabria (Ferretti et al. 2003; Marra et al. 2011, 2017), and attributed to a time span between 8.1 and 7.2 Ma (Marra et al. 2017; Fig. 1). Recently, this scarce material, an incisor and a femur previously assigned to a hexaprotodontid hippopotamid, has been attributed to an anthracotheriid (Marra 2019). Pending further evidence, the remains from Gravitelli indicate that around 6 Ma African hippopotamids dispersed in Southern Europe. Hexaprotodon? sahabiensis was collected from As Sahabi, Libya, a site dated around 6.5 Ma (Bernor and Rook 2008). The teeth described by Gaziry (1987) show several archaic features, such as the sub-selenodont or selenodont aspect of the molar cusps. Weston (2003) noticed that some features displayed by He.? sahabiensis, such as the premolar row as long as the molar row, upper incisors with a mesial-lingual groove and molars dimensions, are similar to those of Archaeopotamus. A mandibular portion from Sahabi was later described by Pavlakis (2008). This fragment confirmed the hexaprotodont condition of the Sahabi species. The sagittal cross section of the mandible of He.? sahabiensis is peculiar among the Hippopotamidae (Boisserie 2005; Pavlakis 2008) but the mandibular portion was probably deformed by taphonomic processes that affected the mandibular symphysis. Pavlakis (2008) highlighted a strong morphologic and morphometric affinity between He.? sahabiensis and A. lothagamensis. Following Pavlakis (2008), the relationship between these two species should be reinvestigated. Hexaprotodon? hipponensis was collected from the lower Pliocene of Pont-de-Duvivier, Algeria (Gaudry 1876), and is represented by very scarce remains: four semi-complete incisors, two broken incisors, two lower premolars, a broken molar and two canine fragments. The third premolar is characterized by an additional cusp and by tubercles that originated from the cingulid, which is highly crenulated. The incisors are long and straight. The wear surface is delimitated by a tracked margin, a characteristic that is typical of this species. Some other remains were later collected from Wadi Natrum, Egypt, and initially attributed to He.? hipponensis by Andrews (1902) and Stromer (1914). Arambourg (1947), in contrast with previous authors, noticed a closer affinity with He. protamphibius (Arambourg 1944) and attributed the lower Pliocene material from Egypt to the subspecies He. protamphibius andrewsi. Two upper molars were illustrated by Andrews (1902), in particular an unworn M3 and a M2 with a weak wear on the lingual cusps. These molars are both quadrangular with a crenulated high cingulum and striated enamel. In the transverse valleys of the M3 there are also several tubercles probably originating from the cingulum (Andrews 1902). The remains from Egypt, subsequently partially described and figured by Stromer (1914), are quite abundant but they have not been revised recently and their location is unclear, preventing any useful considerations on these specimens.
To sum up, during the latest Miocene–early Pliocene, the circum-Mediterranean area was characterized by the presence of at least five different hippopotamid taxa: He.? siculus in Sicily (late Miocene), A. crusafonti in Spain (late Miocene) and in France (early Pliocene), He.? hipponensis in Algeria (early Pliocene), He.? sahabiensis in Libya (late Miocene), and He. protamphibius andrewsi in Egypt (early Pliocene). The last three taxa need to be revised in detail, being based on fragmentary and poorly preserved remains. Furthermore, He.? pantanellii cannot be considered as a valid species because the material collected from the Casino Basin is too scarce and fragmentary for a generic and specific determination and it should be assigned as Hippopotamidae indet.
Conclusions
The morphological and morphometric characters of the hippopotamid remains from the uppermost Miocene deposits of Gravitelli are difficult to assess because all the original material was lost in 1908 due to the catastrophic earthquake that destroyed the city of Messina. The specimens were described and partially figured by Seguenza (1902, 1907). Seguenza (1902) also reported that some material (a maxillary fragment, a partial premolar, and a astragalus) were collected from San Pier Niceto and Scirpi. This information indicates that during the late Miocene at least three different Sicilian localities were inhabited by hippopotamids. The hippopotamid specimens from Gravitelli are mainly represented by teeth and a few poorly preserved postcranial remains. Some characters of the teeth, such as the length of the lower premolars, the low-crowned molars with crenulated cingulids, the weak trefoil wear pattern and hexaprotodonty suggest that He.? siculus displays archaic features. The characters displayed by the Sicilian hippopotamid differ from those of other species collected from the peri-Mediterranean area. In particular, A. crusafonti is morphometrically smaller than He.? siculus and is characterised by a tetraprotodont condition. Hexaprotodon? hipponensis has incisors characterised by a tracked margin and lower premolars with a well-developed additional cusp, whilst He.? sahabiensis has sub-selenodont or selenodont molar cusps. Hexaprotodon? siculus is morphologically closer to Archaeopotamus harvardi, Hexaprotodont sivalensis and He. garyam, and it is morphometrically similar to He. sivalensis. Accordingly, we provisionally refer the Gravitelli hippopotamid to the genus Hexaprotodon. Hexaprotodon? siculus arrived in Sicily during the Messinian, probably from the North African coast, but its ancestor is still unknown. The hippopotamids that colonized the Mediterranean area around the Mio-Pliocene transition were probably closely related, but the scant collected material, which is generally poorly figured and described, does not permit a detailed study. Our analysis of the published figures and descriptions reveals that some of them are probably related to Archaeopotamus, thus suggesting a dispersal of this genus from East Africa to North Africa, and later to Southern Europe. A re-analysis of all the late Miocene hippopotamid remains from the circum-Mediterranean area is necessary to shed light on the evolutionary history of these species as well as to resolve their phylogenetic relationships.
Acknowledgements
Thanks to the reviewers: Athanassios Athanassiou (Ministry of Culture, Ephorate of Palaeoanthropology and Speleology, Athens, Greece) and an anonymous reviewer, the editor Olivier Lambert (Terre et Histoire de la Vie, Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium) for their constructive and useful advices. This work originates within a project on late Neogene vertebrate evolution developed at the University of Florence (coordinator LR). LP prepared this paper within the research project “Ecomorphology of fossil and extant Hippopotamids and Rhinocerotids” of the University of Florence (“Progetto Giovani Ricercatori Protagonisti” initiative). LP thanks the European Commission’s Research Infrastructure Action, EU-SYNTHESYS+ project BE-TAF-1367; part of this research received support from the SYNTHESYS+ Project http://www.synthesys.info/. This work is part of the “Augusto Azzaroli (1921–2015) centennial papers” by the Vertebrate Paleontology Group at the Earth Sciences Department, University of Florence.
References
Aguirre, E.D. 1963. Hippopotamus crusafonti n. sp. del Plioceno inferior de Arenas del Rey (Granada). Notas y comunicaciones del Instituto Geológico y Minero de España 69: 215–230.
Aguirre, E.D., Robles, F., Thaler, L., López Martínez, N., Alberdi, M.T., and Fuentes, C. 1973. Venta del Moro, nueva fauna finimiocena de Moluscos y Vertebrados. Estudios geológicos 29: 569–578.
Alcalá, L. 1994. Macromamíferos neógenos de la fosa de Alfambra-Teruel. 554 pp. Instituto de Estudios Turolenses-Museo Nacional de Ciencias Naturales-CSIC, Teruel.
Alcalá, L. and Montoya, P. 1998. Hexaprotodon crusafonti (Hippopotamidae, Mammalia) del Mioceno superior de El Arquillo (Fosa de Teruel). Boletín de la Real Sociedad Española de Historia Natural (Sección Geológica) 94: 93–100.
Andrews, C.W. 1902. Note on a Pliocene vertebrate fauna from the Wadi-Natrun, Egypt. Geological Magazine 9: 433–439. Crossref
Arambourg, C. 1944. Au sujet de l’Hippopotamus hipponensis Gaudry, 1876. Bulletin de la Société géologique de France (Série 5) 14: 147–152. Crossref
Arambourg, C. 1947. Contribution a l’étude géologique et paléontologique du bassin du lac Rudolphe et de la basse vallée de l’Omo. Mission scientifique de l’Omo 1932–1933. Géologie-Anthropologie 1: 232–562.
Arnason, U., Gulerg, A., Solweig, G., Ursing, B., and Janke, A. 2000. The mitochondrial genome of the sperm whale and a new molecular reference for estimating eutherian divergence rate. Journal of Molecular Evolution 50: 569–578. Crossref
Bernor, R.L. and Rook, L. 2008. A current view of As Sahabi large mammal biogeographic relationships. Garyounis Scientific Bulletin Special Issue 5: 283–290.
Boisserie, J.-R. 2005. The phylogeny and taxonomy of Hippopotamidae (Mammalia: Artiodactyla): a review based on morphology and cladistic analysis. Zoological Journal of the Linnean Society, 143: 1–26. Crossref
Boisserie, J.-R. 2007. Family Hippopotamidae. In: D.R. Prothero and S.E. Foss (eds.), The Evolution of Artiodactyls, 106–119. Johns Hopkins University Press, Baltimore.
Boisserie, J.-R., Brunet, M., Andossa, L., and Vignaud P. 2003. Hippopotamids from the Djurab Pliocene faunas, Chad, Central Africa. Journal of African Earth Sciences 36: 15–27. Crossref
Boisserie, J.-R., Lihoreau, F., Orliac, M., Fisher, R.E., Weston, E.M., and Ducrocq, S. 2010. Morphology and phylogenetic relationships of the earliest known hippopotamids (Cetartiodactyla, Hippopotamidae, Kenyapotaminae). Zoological Journal of the Linnean Society 158: 325–366. Crossref
Boisserie, J.-R., Likius, A., Vignaud, P., and Brunet, M. 2005. A new late Miocene hippopotamid from Toros-Ménalla, Chad. Journal of Vertebrate Paleontology 25: 665–673. Crossref
Boisserie, J.-R., Schuster, M., Beech, M.J., Hill, A., and Bibi, F. 2017. A new species of hippopotamine (Cetartiodactyla, Hippopotamidae) from the late Miocene Baynunah Formation, Abu Dhabi, United Arab Emirates. Palaeovertebrata 41: e2. Crossref
Coryndon, S.C. 1977. The taxonomy and nomenclature of the Hippopotamidae (Mammalia, Artiodactyla) and a description of two new fossil species. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen 80: 61–88.
Coryndon, S.C. 1978. Hippopotamidae. In: V.J. Maglio and H.B.S. Cooke (eds.), Evolution of African Mammals, 483–495. Harvard University Press, Cambridge. Crossref
Crusafont, M., Adrover, R., and Golpe, J.M. 1964. Découverte dans le Pikermien d’Espagne du plus primitif des hippopotames: Hippopotamus (Hexaprotodon) primaevus n. sp. Comptes rendus hebdomadaires des séances de l’Académie des sciences 258: 1572–1575.
De Visser, J.A. 2008. The Extinct Genus Hexaprotodon Falconer & Cautley, 1836 (Mammalia, Artiodactyla, Hippopotamidae) in Asia: Paleoecology and Taxonomy. 390 pp. Ph.D. Thesis, Universiteit Utrecht, Utrecht.
DeGusta, D. and Vrba, E. 2003. A method for inferring paleohabitats from the functional morphology of bovid astragali. Journal of Archaeological Science 30: 1009–1022. Crossref
Falconer, H. and Cautley, P.T. 1836. Note on the fossil Hippopotamus of the Siwalik hills. Asiatic Researches 19: 39–53.
Faure, M. and Méon, H. 1984. L’Hippopotamus crusafonti de La Mosson (près Montpellier). Première reconnaissance d’un Hippopotame néogène en France. Comptes-rendus des séances de l’Académie des sciences. Série 2, Mécanique-physique, chimie, sciences de l’univers, sciences de la Terre 298 (3): 93–98.
Ferretti, M., Rook, L., and Torre, D. 2003. Stegotetrabelodon (Proboscidea, Elephantidae ) from the late Miocene of Southern Italy. Journal of Vertebrate Paleontology 23: 659–666. Crossref
Gallai, G. 2005. Tafonomia e paleobiologia della fauna mammaliana turoliana (Miocene superiore) del bacino del Casino (Siena): risultati preliminari. Rendiconti Società Paleontologica Italiana 2: 119–125.
Gallai, G. and Rook, L. 2006. Propotamochoerus sp (Suidae, Mammalia) from the late Miocene of Gravitelli, (Messina, Sicily, Italy) rediscovered. Rivista Italiana di Paleontologia e Stratigrafia 112: 317–321.
Gaudry, A. 1876. Sur un Hippopotame fossile découvert à Bone (Algérie). Bulletin de la Société géologique de France (Série 3) 4: 502–504.
Gaziry, A.W. 1987. Hexaprotodon sahabiensis (Artiodactyla, Mammalia): a new hippopotamus from Libya. In: N.T. Boaz, A. El-Arnauti, A.W. Gaziry, J. de Heinzelin, and D. Boaz (eds.), Neogene Paleontology and Geology of Sahabi, 303–315. Alan R. Liss, New York.
Gentry, A. and Hooker, J. 1988. The phylogeny of the Artiodactyla. In: M. Benton (ed.), The Phylogeny and Classification of the Tetrapods, Vol. 2, 235–272. Clarendon Press, Oxford.
Gray, J.E. 1821. On the natural arrangement of vertebrate animals. London Medical Repository 15: 296–310.
Harrison, T. 1997. The anatomy, paleobiology, and phylogenetic relationships of the Hippopotamidae (Mammalia, Artiodactyla) from the Manonga Valley, Tanzania. In: T. Harrison (ed.). Neogene Paleontology of the Manonga Valley, Tanzania, 137–190. Plenum Press, New York. Crossref
Hooijer, D.A. 1946. Notes on some Pontian mammals from Sicily figured by Seguenza. Archives néerlandaises de Zoologie 7: 301–333. Crossref
Hooijer, D.A. 1950. The fossil Hippopotamidae of Asia, with notes on the Recent species. Zoologische Verhandelingen 8: 1–124.
Htike, T. 2012. Review on the taxonomic status of Hexaprotodon iravaticus (Mammalia, Artiodactyla, Hippopotamidae) from the Neogene of Myanmar. Shwebo University Research Journal 3: 94–110.
Joleaud, L. 1920. Contribution à l’étude des hippopotames fossiles. Bulletin de la Société géologique de France (Série 4) 20: 13–26.
Kotsakis, T., Barisone, G., and Rook, L. 1997. Mammalian biochronology in an insular domain: the Italian Tertiary faunas. Mémoires et travaux de l’Institut de Montpellier 21: 431–441.
Lacomba, J.I., Morales, J., Robles, F., Santisteban, C., and Alberdi, M.T. 1986. Sedimentologia y paleontologia del yacimiento finimioceno de La Portera (Valencia). Estudios geologicos Instituto de Investigaciones deológicas Lucas Mallada 42: 167–180. Crossref
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. 824 pp. Laurentii Salvii, Stockholm. Crossref
Marra, A.C. 2019. Contribution of the late Miocene mammals from Calabria and Sicily to the palaeogeography of the central Mediterranean. Atti della Accademia Peloritana dei Pericolanti-Classe di Scienze Fisiche, Matematiche e Naturali 97 (S2): A29-1–12.
Marra, A.C., Carone, G., Agnini, C., Ghinassi, M., Oms, O., and Rook, L. 2017. Stratigraphic and chronologic framework of the upper Miocene Cessaniti succession (Vibo Valentia, Calabria, Italy). Rivista Italiana di Paleontologia e Stratigrafia (Research In Paleontology and Stratigraphy) 123: 379–393.
Marra, A.C., Solounias, N., Carone, G., and Rook, L. 2011. Palaeogeographic significance of the giraffid remains (Mammalia, Arctiodactyla) from Cessaniti (Late Miocene, Southern Italy). Geobios 44: 189–197. Crossref
Martino, R., Pignatti, J., Rook, L., and Pandolfi, L. 2020. Systematic revision of hippopotamid remains from the Casino basin, Tuscany, Italy. Fossilia 2020: 29–31. Crossref
Mazza, P. 1995. New evidence on the Pleistocene hippopotamuses of Western Europe. Geologica Romana 31: 61–241.
Montgelard, C., Catzeflis, F. M., and Douzery, E. 1997. Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S rRNA mitochondrial sequences. Molecular Biology and Evolution 14 (5): 550–559. Crossref
Morales, J. 1984. Venta del Moro: su Macrofauna de mamíferos y biostratigrafía continental del Mioceno terminal mediterráneo. 340 pp. Ph.D. Thesis, Universidad Complutense de Madrid, Madrid.
Morton, S.G. 1844. On a supposed new species of Hippopotamus. Proceedings of the National Academy of Sciences, Philadelphia 2: 4–27.
Pandolfi, L. and Rook, L. 2017. Rhinocerotidae (Mammalia, Perissodactyla) from the latest Turolian localities (MN 13; late Miocene) of central and northern Italy. Bollettino della Società Paleontologica Italiana 56: 45–56.
Pandolfi, L., Marra, A.C., Carone, G., Maiorino, L., and Rook, L. 2021. A new rhinocerotid (Mammalia, Rhinocerotidae) from the latest Miocene of Southern Italy. Historical Biology 33: 194–208. Crossref
Pantanelli, D. 1879. Sugli strati miocenici del Casino (Siena) e considerazioni sul Miocene superiore. Atti della Reale Accademia dei Lincei, Memorie della classe di scienze fisiche, matematiche e naturali (Ser. 3) 3: 309–327.
Pavlakis, P. 2008. Rediscovered hippopotamid remains from As Sahabi. Circum-Mediterranean geology and biotic evolution during the Neogene period: The perspective from Libya. Garyounis Scientific Bulletin, Special Issue 5: 179–187.
Rook, L. 1992. Italian Messinian localities with vertebrate faunas. Paleontologia y Evolució 24–25: 141–147.
Rook, L. 1999. Late Turolian Mesopithecus (Mammalia, Primates, Colobinae) from Italy. Journal of Human Evolution 36: 535–547. Crossref
Rook, L., Gallai, G., and Torre, D. 2006. Lands and endemic mammals in the late Miocene of Italy: constrains for paleogeographic outlines of Tyrrhenian area. Palaeogeography, Palaeoclimatology, Palaeoecology 238: 263–269. Crossref
Seguenza, G. 1902. I vertebrati fossili della Provincia di Messina. Parte seconda. Mammiferi e geologia del Piano Pontico. Bollettino della Società Geologica Italiana 21: 115–175.
Seguenza, G. 1907. Nuovi resti di mammiferi pontici di Gravitelli presso Messina. Bollettino della Società Geologica Italiana 26: 89–122.
Stromer, E. 1914. Mitteilungen über Wirbeltierreste aus dem Mittelpliocän des Natrontales (Ägypten), 3. Artiodactyla: A. Bunodontia: Flußpferd. Zeitschrift der Deutschen Geologischen Gesellschaft 66: 1–33.
Thenius, E. 1989. Zähne und Gebiß der Säugetiere. Handbuch der Zoologie VIII (56). 513 pp. De Gruyter, Berlin. Crossref
Van der Made, J. 1999. Biogeography and stratigraphy of the Mio-Pleistocene mammals of Sardinia and the description of some fossils. Deinsea 7 (1): 337–360.
Weston, E.M. 2000. A new species of hippopotamus Hexaprotodon lothagamensis (Mammalia: Hippopotamidae) from the late Miocene of Kenya. Journal of Vertebrate Paleontology 20: 177–185. Crossref
Weston, E.M. 2003. Fossil Hippopotamidae from Lothagam. In: M.G. Leakey and J.M. Harris (eds.), Lothagam. The Dawn of Humanity in Eastern Africa. 441–483. Columbia University Press, New York. Crossref
Acta Palaeontol. Pol. 66 (Supplement to 3): 67–78, 2021
https://doi.org/10.4202/app.00838.2020