
Diversity of chondrostean fish Coccolepis from the Late Jurassic Solnhofen Archipelago, Southern Germany
ADRIANA LÓPEZ-ARBARELLO and MARTIN EBERT
López-Arbarello, A. and Ebert, M. 2021. Diversity of chondrostean fish Coccolepis from the Late Jurassic Solnhofen Archipelago, Southern Germany. Acta Palaeontologica Polonica 66 (4): 837–846.
Late Jurassic marine vertebrates are extraordinarily well preserved in several Plattenkalk Lagerstätten in central Europe. Among them, the Solnhofen Archipelago has yielded the very rare fish Coccolepis bucklandi, which was the first fossil chondrostean to be found in sediments younger than the Triassic. The type specimen of this species was lost, but it was rediscovered recently, prompting the alpha taxonomic revision of this iconic fish. A new species Coccolepis solnhofensis has been identified among the specimens referred to C. bucklandi. The two species differ in the distinctive distribution of scutes and fringing fulcra. Based on the available evidence, C. bucklandi is restricted to the Eichstätt Basin and the Lithacoceras eigeltingense ß Horizon of the Lithacoceras riedense Subzone (Hybonoticeras hybonotum Zone), and C. solnhofensis sp. nov. is limited to the Solnhofen Basin and the slightly younger Subplanites rueppellianus Subzone (Hybonoticeras hybonotum Zone). Therefore, the two species are geographically and stratigraphically separated. The diagnosis of Coccolepis is improved with the addition of new characters, and the genus is here restricted to the two early Tithonian species from the Solnhofen Archipelago. Among the four species previously described or referred to Coccolepis, the generic assignment of “Coccolepis” australis and “Coccolepis” liassica, remains unclear. Sunolepis yumenensis is here returned to its original genus, and the new combination Condorlepis woodwardi is proposed for this Early Cretaceous coccolepidid from Australia.
Key words: Actinopterygii, Coccolepididae, Chondrostei, Mesozoic, Jurassic, Tithonian, Solnhofen, Plattenkalk, Lagerstätte.
Adriana López-Arbarello [a.lopez-arbarello@lrz.uni-muenchen.de], Department of Earth and Environmental Sciences, Ludwig Maximilian University, Richard-Wagner-Str. 10, 80333 Munich, Germany; GeoBio-Center, Ludwig-Maximilians-Universität München, Richard-Wagner-Str. 10, 80333 München, Germany.
Martin Ebert [martin.ebert@jura-museum.de], SNSB-Jura-Museum Eichstätt, Willibaldsburg, D-85072 Eichstätt, Germany.
Received 11 January 2021, accepted 29 March 2021, available online 29 October 2021.
Copyright © 2021 A. López-Arbarello and M. Ebert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
In the Late Jurassic, large carbonate platforms formed the northern shore of the Tethyan Ocean producing very rich ecosystems which are preserved in different Plattenkalk Lagerstätten in central Europe (Keupp et al. 2007; Fig. 1A). Among them are the famous lithographic limestone of the southern Franconia Alp in Bavaria. The fine-grained limestone beds of southern Bavaria were deposited within small to medium-sized depressions (Keupp et al. 2007). The Solnhofen Basin is only one among more than twelve different known depocenters (Fig. 1B), which together represent an evolving fossil archipelago spanning c. 3.5 Ma from the late Kimmeridgian to the early Tithonian (Tischlinger and Schweigert 2020). Over the last decades, the recognition of the complex spatial and temporal structure of this fossil archipelago has led researchers to refer to these deposits as the “Solnhofen Archipelago” and to emphasize the distinction of its different components (e.g., Rauhut et al. 2017). However, the early literature of the 19th century as well as the information in collection catalogues and archives of that time are usually vague concerning the precise locality for collection of the fossil specimens. Many historical specimens registered as originating from Solnhofen actually derive from depocenters other than the Solnhofen Basin (e.g., Moser et al. 2017). This lack of accuracy regarding the provenance of numerous fossils, including many type specimens collected over more than 200 years, makes it difficult to reconstruct the composition of the individual faunas corresponding to the different basins within the Solnhofen Archipelago. These difficulties become critical when trying to understand the dynamic of the Solnhofen Archipelago through space and time.
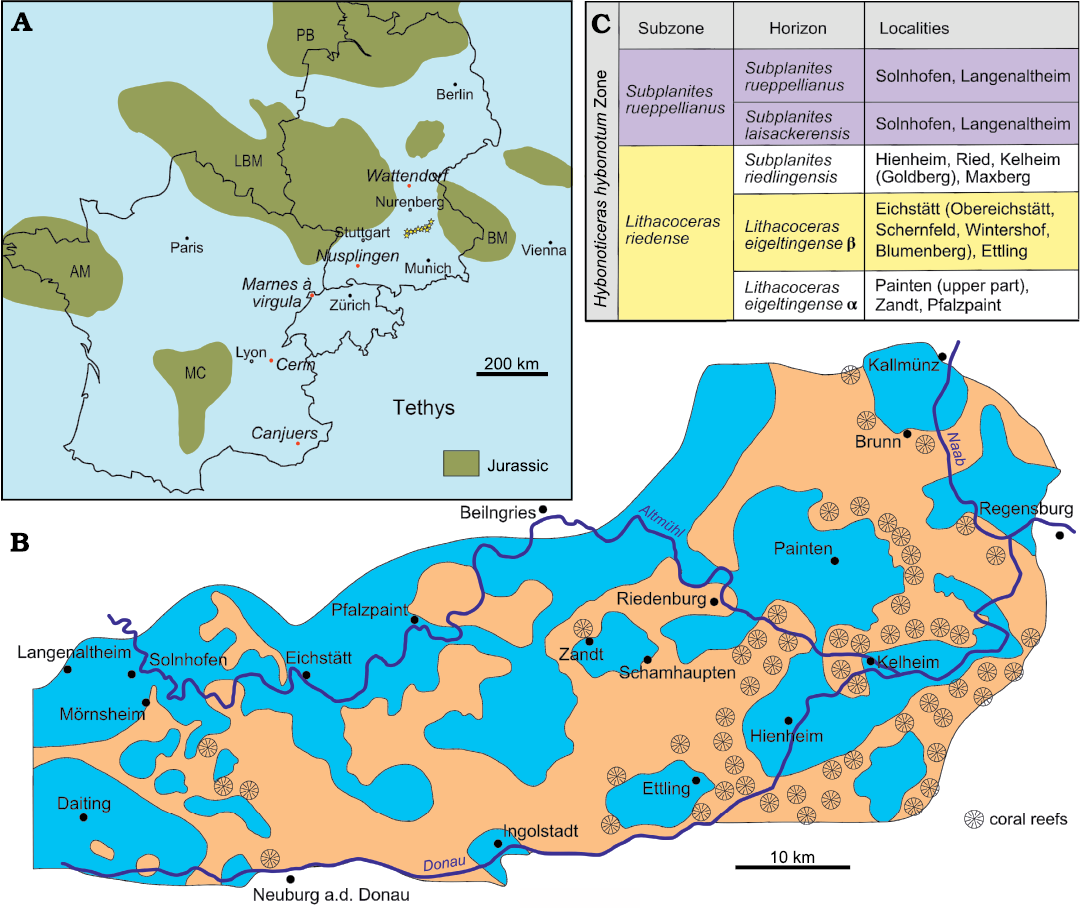
Fig. 1. Geographic and stratigraphic setting. A. Palaeogeographic reconstruction showing the location of the main Upper Jurassic Plattenkalk Lagerstätten in central Europe. The Solnhofen Archipelago is represented with several stars, other Lagerstätten are indicated with dots and italicised names. Abbreviations: AM, Armorican Massif; Bm, Bohemian Massif; LBM, London-Brabant Massif; MC, Massif Central High; PB, Pompeckjsche Block. Redrawn from Viohl (2015: fig. 85). B. Palaeogeographic reconstruction of the Solnhofen Archipelago. Orange areas represent sponge/microbial reefs and blue areas represent the basinal facies. Redrawn from Kölbl-Ebert and Cooper (2019: fig. 2). C. Biostratigraphy with ammonite stratigraphic succession of the Lithacoceras riedense and Subplanites rueppellianus subzones of the Hybonoticeras hybonotum Zone (lower Tithonian, Upper Jurassic). Redrawn from Tischlinger and Schweigert (2020: fig. 8).
Moreover, several type specimens have been lost over the centuries, which makes the alpha taxonomic work extremely problematic. As an example, the holotype of the actinopterygian Coccolepis bucklandi Agassiz, 1843, has been considered lost (Hilton et al. 2004). When Louis Agassiz described the species, the type and only known specimen was part of William Buckland’s collection in Oxford, UK. Wagner (1863) roughly mentioned that the specimen was kept in England and Woodward (1891) indicated that it was in the Oxford Museum. However, according to the head curator Eliza Howlett (personal communication, June 2019), although there is a catalogue record for the holotype of Coccolepis bucklandi, the specimen has never been found in the Earth Collections of the Oxford University Museum of Natural History. Following the comment of Agassiz “I have received in communication from Dr. Buckland a small fish (j’ai reçu en communication de la part de M. le Dr Buckland un petit poisson)” (Agassiz 1843: 300), Laure Bapst and one of the authors (ME) were able to locate the specimen in the paleontological collection of the Museum d’Histoire Naturelle in Neuchâtel, Switzerland (Fig. 2; Ebert et al. 2021).
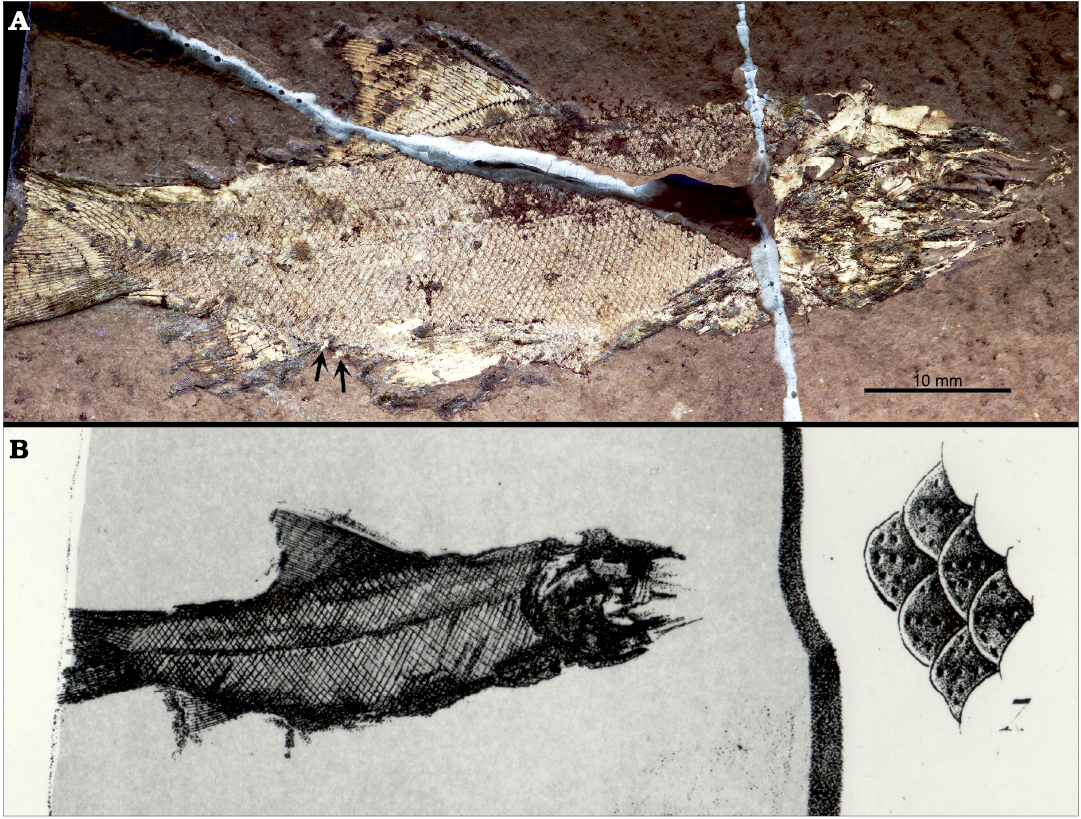
Fig. 2. Holotype of the chondrostean fish Coccolepis bucklandi Agassiz, 1843 from the Tithonian, Upper Jurassic of the Solnhofen Lagerstätte, Bavaria, Germany. A. MHNN-FOS 361 (TL ~ 75 mm), photograph taken under UV-light, courtesy of Thierry Malvesy (MHNN). Arrows point to fragmentary remains of the preanal scutes. B. Original illustration of Agassiz (1843: pl. 36: 6). Scan courtesy of Eric Hilton (Virginia Institut of Marine Science, Gloucester Point, Virginia, USA).
Coccolepis bucklandi has been an iconic taxon because it was the first discovery of a non neopterygian actinopterygian fossil in sediments younger than the Triassic (Agassiz 1843; Wagner 1863). Berg (1940) proposed the family Coccolepididae (original spelling Coccolepidae) to include this and other morphologically similar Mesozoic species. Initially classified in the unnatural palaeonisciform group, coccolepidids are currently referred to the Chondrostei (Acipenseriformes and their fossil relatives) (Hilton et al. 2004; López-Arbarello et al. 2013). Although a phylogenetic study is still necessary to confirm this referral as well as the monophyly of Coccolepididae, the strong morphological resemblance displayed by coccolepidid species between each other, and the lack of other potentially closely related lineages in the Jurassic and Cretaceous indirectly support these hypotheses.
Besides a very conservative morphology, the endo- and exoskeleton of coccolepidids are generally poorly ossified. Therefore, the preservation of coccolepidid fossils is usually poor or incomplete, making the low-level taxonomic work extremely difficult. Among the coccolepidid species currently accepted as valid, most of them were originally described as species of Coccolepis (Table 1). The relatively recent revision of several of these Coccolepis species led to their referral to the genus Morrolepis Kirkland, 1998 (Skrzycka 2014), and the erection of the genera Condorlepis López-Arbarello, Sferco, and Rauhut, 2013, and Barbalepis Olive, Taverne, and López-Arbarello, 2019. Most of the coccolepidid species have been found in freshwater environments, being C. bucklandi and “Coccolepis” liassica Woodward, 1890 (Lower Jurassic of Dorset in England), the only strictly marine members of the group (Olive et al. 2019).
Although Coccolepis bucklandi has been studied and described in detail relatively recently by Hilton et al. (2004), they were not able to examine the holotype and their work is exclusively based on five specimens referred to this species. Our detailed examination of the newly located holotype and the recognition of some significant morphological variation among the specimens referred to this species, triggered a new taxonomic study of this iconic taxon from the Solnhofen Archipelago.
Table 1. Taxonomic status of the species currently classified in Coccolepididae.
|
Original binomen |
Generic assignment herein |
|
Coccolepis bucklandi Agassiz, 1843 |
Coccolepis Agassiz, 1843 |
|
Coccolepis solnhofensis sp. nov. |
Coccolepis Agassiz, 1843 |
|
Coccolepis liassica Woodward, 1890 |
uncertain |
|
Coccolepis andrewsi Woodward, 1891 |
Morrolepis Kirkland, 1998 |
|
Coccolepis australis Woodward, 1895 |
uncertain |
|
Coccolepis macroptera Traquair, 1911 |
Barbalepis Olive, Taverne, and López-Arbarello, 2019 |
|
Coccolepis aniscowitchi Gorizdro-Kulczycka, 1926 |
Morrolepis Kirkland, 1998 |
|
Coccolepis woodwardi Waldman, 1971 |
Condorlepis López-Arbarello, Sferco, and Rauhut, 2013 |
|
Angaraichthys rohoni Sytchevskaya and Yakovlev, 1985 |
Iyalepis Sytchevskaya, 2006 |
|
Morrolepis schaefferi Kirkland, 1998 |
Morrolepis Kirkland, 1998 |
|
Oligopleurus groeberi Bordas, 1943 |
Condorlepis López-Arbarello, Sferco, and Rauhut, 2013 |
|
Plesiococcolepis hunanensis Wang, 1977 |
Plesiococcolepis Wang, 1977 |
|
Sunolepis yumenensis Liu, 1957 |
Sunolepis Liu, 1957 |
Institutional abbreviations.—JME, Jura-Museum Eichstätt, Germany; MACN, Museo Argentino de Ciencias Naturales, Ciudad Autónoma de Buenos Aires, Argentina; MCZ, Museum of Comparative Zoology (Harvard University), Cambridge, USA; MHNN, Museum d’Histoire Naturelle in Neuchâtel, Switzerland; MMG-SNSD, Museum für Mineralogie und Geologie, Senckenberg Naturkundliche Sammlungen Dresden, Germany; MPEF-PV, vertebrate paleontology collection of the Museo Paleontológico Egidio Feruglio, Trelew, Argentina; SNSB-BSPG, Bayerische Staatsammlung für Paläontologie und Geologie, Munich, Germany.
Other abbreviations.—TL, total length.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:3A157948-1BCF-4ABB-9261- 562077788582
Material and methods
The specimens were studied under stereomicroscopes Leica Wild PZO 20138 and Leica Wild M80. The line art images were made with an Intuos Pro PTH-651 Wacom tablet and the software Affinity Designer 1.8.6. Photographs were taken with a Nikon D7000 digital camera equipped with a Nikon AF-S micro 60 mm objective and a Jenoptik digital camera ProgRes C5 attached to the stereomicroscope. The fossils were examined and photographed under white and ultraviolet (UV) light (Krantz UV Lamp I 361).
Anatomical comparisons are based on published literature and high-quality photographs and notes taken by one of the authors (ALA) during direct observation of the following specimens: Condorlepis groeberi: MACN 14434 (lectotype), MACN 14433, 14432, 18630A, 18551, 18552, 18553, 18561, 18562, 18575; MPEF-PV 1496-5A, 1556-6, 1731, 1732, 1733, 1766, 1767, 3958, 10504–10507; Morrolepis schaefferi: MWC 5305, 5306, 5307.
The signs attached to the entries in the synonymy list follow Matthews (1973). The size of the specimens is indicated through their total length (TL).
Systematic palaeontology
Actinopterygii Cope, 1887
Chondrostei Müller, 1845
Coccolepididae Berg, 1940 sensu Hilton et al. 2004
Genus Coccolepis Agassiz, 1843
Type species: Coccolepis bucklandi Agassiz, 1843; Solnhofen Lagerstätte, Germany; lower Tithonian, Upper Jurassic.
Included species: Coccolepis solnhofensis sp. nov. Other species previously assigned to Coccolepis are now excluded from this genus (see Discussion).
Emended diagnosis.—Modified from Hilton et al. (2004) and López-Arbarello et al. (2013). Coccolepidid fish, with the following combination of characters: dermal bones of the skull roof, scales, and fin rays ornamented with mostly regularly arranged sharply pointed, posteriorly directed denticles; opercle is larger than subopercle; lower jaw extremely slender and short, about half the length of the maxilla; single row of large conical teeth on the lower jaw; dorsal fin origin anterior to origin of pelvic fins; large dorsal and pectoral fins, with 41–49 and more than 30 fin rays, respectively.
Coccolepis bucklandi Agassiz, 1843
Figs. 2–4A, 5A.
1843 Coccolepis bucklandi sp. nov.; Agassiz 1843: 300 (vol. 2), pl. 36, figs. 6, 7.
1881 Coccolepis bucklandi Agassiz, 1843; Vetter 1881: 37–42, pl. 1: 2.
1991 Coccolepis bucklandi Agassiz, 1843; Frickhinger 1991: figure on p. 298.
1992 Coccolepis bucklandi Agassiz, 1843; Lambers 1992: 288, pl.1: A.
1995 Coccolepis bucklandi Agassiz, 1843; Frickhinger 1995: figure on p. 298.
1999 Coccolepis bucklandi Agassiz, 1843; Lambers 1999: 270, fig. 1.
2015 Coccolepis bucklandi Agassiz, 1843; Schultze and Arratia 2015: 369–370, figs. 723, 726.
Holotype: MHNN-FOS 361 (Fig. 2). Nearly complete fish in lateral view, missing a portion of its tail and the anteriormost part of the cranium (approximately 75 mm total length). The Neuchâtel specimen has been identified with confidence as the missing type specimen (Ebert et al. 2021). The shape of the fish and the preserved bones and scales are at the finest detail consistent with Agassiz’ illustration and description of the type specimen.
Type locality: The provenance of the holotype is indicated as “Solnhofen” (written Solenhofen; see Ebert et al. 2021). However, the indication “Solnhofen” in historical labels from the 19th Century usually vaguely refers to the whole complex of basins within the Solnhofen Archipelago (e.g., Moser et al. 2017). Based on indirect evidence, MHNN FOS 361 most probably comes from one of the localities within the Eichstätt Basin (see Discussion).
Type horizon: Hybonoticeras hybonotum Zone, Altmühltal Formation, lower Tithonian, Upper Jurassic (Tischlinger and Schweigert 2020).
Material.—Type material and JME-SOS3445 from Blumenberg (Fig. 3A), JME-SOS3382 from Workerszell (Fig. 3B), and JME-SOS2340 from Schernfeld, all three localities within the Eichstätt Basin; MMG-SNSD BaJ 1845, Eichstätt. All from the Lithacoceras eigeltingense ß Horizon of the Lithacoceras riedense Subzone (Altmühltal Formation, Hybonoticeras hybonotum Zone).
Diagnosis.—Species of Coccolepis differing from the other species of the genus in the following characters: three distinct preanal scutes; fringing fulcra on pectoral, pelvic and caudal fins; numerous ventral caudal fringing fulcra.
Remarks.—According Article 73 of the International Code of Zoological Nomenclature (ICZN 1999), the single specimen described and figured by Agassiz (1943), i.e., the MHNN-FOS 361, when creating the nominal taxon Coccolepis bucklandi is the name-bearing type of this species. Considering the original type specimen lost, Hilton et al. (2004) designated the SNSB-BSPG 1904 I 19 as the neotype of C. bucklandi. After relocating the original type specimen, according to Article 75.8 of the ICZN, the rediscovered MHNN-FOS 361 becomes the name-bearing type of Coccolepis bucklandi Agassiz, 1843, and the neotype proposed by Hilton et al. (2004) is set aside.
Stratigraphic and geographic range.—Although the precise provenance of the holotype is unknown, all the other specimens referred to Coccolepis bucklandi were collected from localities within the Eichstätt Basin of the Solnhofen Lagerstätte, Bavaria, Germany. The limestone beds of the Eicshtätt Basin correspond to the Lithacoceras eigeltingense ß Horizon, Lithacoceras riedense Subzone, Hybonoticeras hybonotum Zone; Altmühltal Formation, lower Tithonian, Upper Jurassic.
Coccolepis solnhofensis sp. nov.
Figs. 4, 5.
Zoobank LSID: urn:lsid:zoobank.org:act:834F1774-D7AB-4953-B0 DE-8714E4690F99
2004 †Coccolepis bucklandi Agassiz, 1843; Hilton et al. 2004: 213–231, figs. 2–9, 11–14.
2015 Coccolepis bucklandi Agassiz, 1843; Schultze and Arratia 2015: 370, figs. 724–726.
2015 Coccolepis bucklandi Agassiz, 1843; Tischlinger and Völkl-Costantini 2015: fig. 178.
Etymology: The species name recalls the Solnhofen Lagerstätte.
Holotype: SNSB-BSPG 1904 I 19. Incompletely preserved acid-prepared specimen figured by Hilton et al. (2004: figs. 3D, 5, 7, 9, 11, 12C, D, 13, 14)
Type locality: Solnhofen Lagerstätte, Bavaria, Germany.
Type horizon: Subplanites rueppellianus Subzone, Hybonoticeras hybonotum Zone, Altmühltal Formation, Tithonian, Upper Jurassic (Tischlinger and Schweigert 2020).
Material.— Type material and MCZ VPF-5293 (Hilton et al. 2004: figs. 3A, 4, 6), SNSB-BSPG 1895 I 44 (Hilton et al. 2004: fig. 3C), SNSB-BSPG AS I 1328 (Hilton et al. 2004: figs. 8, 12A, B), and SNSB-BSPG 1986 XV 112 (Hilton et al. 2004: fig. 3B). The provenance of these specimens is discussed below.
Diagnosis.—Species of Coccolepis differing from the other species of the genus in the following characters: one predorsal scute; distinct preanal scales; fringing fulcra on pectoral and caudal fins only; ventral caudal fringing fulcra very slender and few in number.
Description.—See detailed description in Hilton et al. (2004).
Stratigraphic and geographic range.—Specimen SNSB-BSPG 1895 I 44 was collected in the locality of Hummelberg near Solnhofen, within the Solnhofen Basin. The provenance of the other specimens of C. solnhofensis, including the holotype, is only indicated as Solnhofen or lithographic limestone Solnhofen, but it is possible that the species is limited to the Solnhofen Basin (Fig. 1B; see Discussion). The limestone beds of the Solnhofen basin correspond to the Subplanites rueppellianus Subzone, upper Hybonoticeras hybonotum Zone of the Altmühltal Formation, lower Tithonian, Upper Jurassic (Fig. 1C) (Tischlinger and Schweigert 2020).
Discussion
The rediscovery of the name-bearing type specimen (MHNN-FOS 361) of Coccolepis bucklandi and the discovery of several morphological differences between JME-SOS3445 and other specimens referred to this species (Ebert et al. 2021) prompted the thorough revision of the fossils referred to this taxon. Consequently, two well-defined morphotypes were identified, which are, as far as can be determined, not only geographically, but also stratigraphically separated. One of the two morphotypes matches the morphology of MHNN-FOS 361 and, thus, corresponds to C. bucklandi Agassiz, 1843. The second morphotype is regarded as a new taxon.
Anatomical comparison
The morphology of the two species Coccolepis bucklandi and C. solnhofensis sp. nov. is very similar and mostly agree with the detailed description by Hilton et al. (2004). Therefore, we will only discuss the discovered differences between the two taxa and add a few new observations. Meristic information for all the studied specimens is provided in Ebert et al. (2021: table 1).
Skull.—Hilton et al. (2004) described the lower jaw as very slender and long, interpreting the jaw in MCZ VPF-5293 of C. bucklandi as broken. However, we disagree with their observation. The lower jaw in MCZ VPF-5293 (Hilton et al. 2004: fig. 6A) is complete or nearly complete, as it is in all the other specimens in which it is exposed (the holotype MHNN-FOS 361, JME-SOS2340, MMG-SNSD BaJ 1845 of C. bucklandi, and SNSB-BSPG 1895 I 44 of C. solnhofensis sp. nov.). Therefore, differing from other coccolepidid taxa, the lower jaw in the two species of Coccolepis is extremely slender and very short, about half the length of the maxilla.
The gular plate is not preserved in the specimens studied by Hilton et al. (2004), but it is well preserved, displaced and well exposed in internal view in JME-SOS3445 of C. bucklandi (Fig. 3A). The general shape of the median gular is oval, narrowing posteriorly, so that the anterior border is broader than the posterior, deeply convex border. The exposed surface of the gular plate is ornamented with several denticles which can be distinguished through the thin laminar bone. A V-shaped canal or groove is preserved, exiting at the anterolateral corners of the plate indicating the presence of a gular sensory canal or pit line which was possibly connected with the mandibular sensory canal.

Fig. 3. Chondrostean fish Coccolepis bucklandi Agassiz, 1843 from the Tithonian, Upper Jurassic of the Solnhofen Lagerstätte, Bavaria, Germany. A. JME-SOS3445, the skull and anterior part of the body are preserved in ventral view. The specimen is twisted at the pelvic fins and the posterior part of the body is preserved in left lateral view (TL = 120 mm). B. JME-SOS3382 (photographed under UV-light), the body is preserved in right lateral view; the skull is slightly twisted and preserved in dorsolateral view (TL = 100 mm). Arrows point to preanal scutes. A, B, Photographs courtesy of Andreas Hecker (JME).
Fins.—The pectoral fins are located lateroventrally in all coccolepidid fishes. Therefore, the total number of fin rays is difficult to assert and only possible to evaluate in ventrally or lateroventrally exposed specimens. JME-SOS3445 of C. bucklandi is preserved in ventral view and both pectoral fins are well exposed (Fig. 3A). They consist of 36 or 37 fin rays and two basal fulcra. In the same specimen, approximately 17 small fringing fulcra are fused to the basal fulcra and attached to the marginal ray. All fringing fulcra have nearly the same size and at least the more proximal ones have an elongated patch of ganoine. The right pectoral fin in the acid prepared SNSB-BSPG AS I 1328 of C. solnhofensis sp. nov. is well exposed, including 33 lepidotrichia and a similar condition of the fringing fulcra. Hilton et al. (2004) described this structure as a pectoral spine formed by the fusion of fringing fulcra (Hilton et al. 2004: figs. 7, 8, 12A). Although this pectoral spine is relatively larger in SNSB-BSPG AS I 1328 (TL ~90 mm) than in JME-SOS3445 (TL ~120 mm), the organ is formed by the fusion of basal and fringing fulcra and, thus, equivalent in both specimens. The difference in the relative size might be due to ontogeny.
According to Hilton et al. (2004) fringing fulcra are absent on the pelvic fins and we confirm this condition in C. solnhofensis sp. nov. (Hilton et al. 2004: fig. 12B). However, differing from this species, JME-SOS3445 of C. bucklandi preserves at least three fringing fulcra on the marginal rays of the right pelvic fin (Fig. 4A1).
Hilton et al. (2004) did not notice the very few and slender fringing fulcra on the ventral marginal ray of the caudal fin of the holotype SNSB-BSPG 1904 I 19 and SNSB-BSPG 1986 XV 112 and SNSB-BSPG AS I 1328 of C. solnhofensis sp. nov. These fringing fulcra are mainly formed by the terminal segments of the marginal rays or a marginal branching of these rays (Fig. 4B). The ventral caudal fringing fulcra in C. bucklandi are formed not only by those terminal segments, but there are several additional fulcra laying on the marginal rays between those terminal segments (Fig. 4A2). As a result, the ventral caudal fringing fulcra are significantly more numerous in this species; JME-SOS3445 preserves approximately 12 fringing fulcra along the ventral caudal ray.

Fig. 4. Fringing fulcra in the chondrostean fishes Coccolepis bucklandi Agassiz, 1843 (A), and Coccolepis solnhofensis sp. nov. (B) from the Tithonian, Upper Jurassic of the Solnhofen Lagerstätte, Bavaria, Germany. A. JME-SOS3445, pelvic fins and preanal scutes (A1). Arrows point to the preserved fringing fulcra; asterisks signal the three preanal scutes. Detail of the ventral margin of the caudal fin (A2). B. SNSB-BSPG AS I 1328, detail of the ventral margin of the caudal fin. Photograph taken under UV-light, courtesy of Helmut Tischlinger (Stammham, Germany).
All paired and median fin rays in the two species of Coccolepis are evenly joined, including the basal segment, the lepidotrichia are only very distally branched and they apparently divide only once.
Scales.—In both Coccolepis species, the body is covered with amioid elasmoid scales, and the axial lobe of the tail is flanked by rhomboid scales as described by Hilton et al. (2004). There is no significant difference between the species concerning these features. Differences between the taxa reside in the presence of in three preanal scutes in C. bucklandi, and a single predorsal scute C. solnhofensis sp. nov.
The anal fin in C. bucklandi is preceded by three scutes: the largest is a median scute located anterior to the vent, which is followed by a pair of smaller scutes flanking the vent (Figs. 2, 3, 4A1, 5A). These preanal scutes are well ossified and clearly distinct from the scales covering the body. Their surface is smooth and covered with a thin layer of ganoin. Their posterior border is strongly serrated with up to seven serrations. The preanal scutes are best preserved, though slightly disarticulated in JME-SOS3445. We have observed at least one preanal scute or clear remains of these scutes in all of the specimens representing this species, in which the area is preserved. In contrast, after a careful examination of the five specimens of C. solnhofensis sp. nov., we found no evidence of similar scutes. There are distinct preanal scales, however, which are preserved only in the holotype SNSB-BSPG 1904 I 19 (Fig. 5B) and in SNSB-BSPG AS I 1328. These preanal scales are identical to the adjacent amioid scales covering the body, but they are strongly mineralized. Preanal scutes or modified scales are related to gonopodial-like structures and modified anal fins and thus associated with potential sexual dimorphism in several early actinopterygians (e.g., Bürgin 1990; Lombardo 1999; Sun et al. 2012; Xu et al. 2016). Male and female individuals cannot be distinguished in the small samples representing C. bucklandi and C. solnhofensis sp. nov. because, when preserved, the presence of preanal scutes or modified preanal scales has a uniform distribution within each association, which is coincident with the distribution of the other diagnostic features. Still, sexual dimorphism can also be related to these preanal structures in C. bucklandi and C. solnhofensis sp. nov., a hypothesis that deserves further investigation.
Hilton et al. (2004) described a single basal fulcrum at the origin of the dorsal fin in the holotype of C. solnhofensis sp. nov. (SNSB-BSPG 1904 I 19). Additionally, we found a single scute preceding the dorsal fin in SNSB-BSPG 1986 XV 112 (Fig. 5C) and SNSB-BSPG 1895 I 44 of this species. No evidence of a predorsal scute have been observed in any of the specimens representing C. bucklandi.

Fig. 5. Distinct scutes and scales (indicated with the arrows) in the chondrostean fishes Coccolepis bucklandi Agassiz, 1843 (A), and Coccolepis solnhofensis sp. nov. (B, C) from the Tithonian, Upper Jurassic of the Solnhofen Lagerstätte, Bavaria, Germany. A. MHNN-FOS 361 (holotype), fragments of preanal scutes (arrows). Photograph courtesy of Thierry Malvesy (MHNN). B. SNSB-BSPG 1904 I 19 (holotype), preanal scales (arrows). C. SNSB-BSPG 1986 XV 112, predorsal scute (arrow).
Distribution
Among the specimens representing Coccolepis bucklandi, the precise provenance is only known for the three specimens in the Jura-Museum Eichstätt: Schernfeld (JME-SOS2340), Workerszell (JME-SOS3382), and Blumenberg (JME-SOS3445). These localities are within the Eichstätt Basin and dated in the Lithacoceras eigeltingense ß Horizon of the L. riedense Subzone (Altmühltal Formation, Hybonoticeras hybonotum Zone) (Tischlinger and Schweigert 2020). The locality of the specimen in Dresden (MMG-SNSD BaJ 1845) is only indicated as Eichstätt, but according to Vetter (1881) it was collected in one of the quarries around the village of Obereichstätt, which is also referred to the L. eigeltingense ß Horizon. Although the provenance of the holotype MHNN-FOS 361 is vaguely indicated as “Solnhofen”, the lithology of the rock containing the specimen matches the sediments from the quarries within the Eichstätt depocenter. Therefore, it is likely that the distribution of the species C. bucklandi is limited to the Eichstätt Basin and the L. eigeltingense ß Horizon.
On the other hand, the SNSB-BSPG 1895 I 44 of Coccolepis solnhofensis sp. nov. was collected in the locality of Hummelberg near Solnhofen, well within the Solnhofen Basin. The limestone beds of Solnhofen sensu stricto correspond to the Subplanites rueppellianus Subzone (Altmühltal Formation, Hybonoticeras hybonotum Zone) (Tischlinger and Schweigert 2020). The provenance of the other three specimens of C. solnhofensis sp. nov. is only indicated as “Solnhofen” (SNSB-BSPG AS I 1328, SNSB-BSPG 1986 XV 112) or “Lithographiche Schiefer Solnhofen” (= lithographic limestone Solnhofen, holotype SNSB-BSPG 1904 I 19), but it is possible that C. solnhofensis sp. nov. is limited to the Solnhofen Basin, which is slightly younger than the Eichstätt Basin (Tischlinger and Schweigert 2020).
Although the potential geographic and stratigraphic separation of the two species of Coccolepis should be confirm by further studies when more specimens are available and possibly through detailed lithographic analyses, their potential endemism is a valid hypothesis.
Species excluded from Coccolepis
The taxonomy of several species previously referred to Coccolepis has been revised recently resulting in their referral to the coccolepidid genus Morrolepis (Skrzycka 2014) or the erection of new genera (López-Arbarello et al. 2013; Olive et al. 2019). Other species previously referred to Coccolepis, “Coccolepis” liassica Woodward, 1890, Lower Jurassic, Dorset, England; “C.” australis Woodward, 1895, Upper Jurassic, Talbragar, New South Wales, Australia; “C.” yumenensis (Liu 1957), Upper Jurassic or Lower Cretaceous, Yumen, Gansu Province, China; and “C.” woodwardi Waldman, 1971”, Lower Cretaceous, Koonwarra, Victoria, Australia, are here excluded from the genus because they do not present the combination of features given in the emended diagnosis.
Differing from Coccolepis the skull bones of “C.” liassica are covered with ganoine and the ornamentation consists of coarse tubercles which sometimes merge producing rugae, there is a double row of teeth on the lower jaw, the dorsal and pectoral fins are significantly smaller, including only 20–25 dorsal and 18–20 pectoral fin rays, the lower jaw is notably more robust and larger, as long or longer than the maxilla (Gardiner 1960). “C.” woodwardi resembles “C.” liassica and differs from Coccolepis in the following features: opercle smaller than subopercle, double row of teeth on the lower jaw, small pectoral fins with 18 rays (Waldman 1971). The lower jaw of “C.” yumenensis is described as robust, with a double row of teeth, the opercle is smaller than the subopercle and the origin of the dorsal fin is located posterior to the insertion of the pelvic fins (Liu 1957). The more poorly known “C.” australis differs from Coccolepis in the relative position of the pelvic fins, which are closer to the pectoral fins, inserting well anterior to the origin of the dorsal fin (Woodward 1895; Waldman 1971).
Considering the differences mentioned above and pending a thorough revision of these four species, Sunolepis yumenensis Liu, 1957, most probably represents its own genus, as originally described (Table 1). As already indicated by Olive et al. (2019), “Coccolepis” woodwardi resembles Condorlepis groeberi (Bordas, 1943) very closely, including the following diagnostic features of this genus: subopercle and branchiostegals ornamented with low concentric striae; supracleithrum longer than cleithrum; small, oval postcleithrum; fringing fulcra present in caudal fin only. Accordingly, the species Coccolepis woodwardi Waldman, 1971, is here tentatively referred to Condorlepis López-Arbarello, Sferco, and Rauhut, 2013 (Table 1). “Coccolepis” australis is too poorly known to make any inference about its possible generic assignment, and “C.” liassica might represent a new genus.
Conclusions
The relocation of the holotype of Coccolepis bucklandi triggered the alpha taxonomic revision of this early Tithonian (Late Jurassic) species. As a result, the species C. solnhofensis sp. nov. has been identified among the specimens referred to C. bucklandi. The two species of Coccolepis are ostensibly geographically and stratigraphically separated. As far as we can be certain about the provenance of some of the specimens, C. bucklandi is limited to the Eichstätt Basin and the Lithacoceras eigeltingense ß Horizon of the L. riedense Subzone (Altmühltal Formation, Hybonoticeras hybonotum Zone; Tischlinger and Schweigert 2020). Similarly, C. solnhofensis sp. nov. is limited to the Solnhofen Basin and the slightly younger Subplanites rueppellianus Subzone (Altmühltal Formation, Hybonoticeras hybonotum Zone; Tischlinger and Schweigert 2020).
The distinction of the new species of Coccolepis and the comparative anatomical analysis led to the improvement of the diagnosis of this genus with the addition of new characters. Consequently, four species previously described or referred to Coccolepis are here excluded from the genus. The generic assignment of two of these species, “C.” australis and “C.” liassica, remains unclear. Sunolepis yumenensis Liu, 1957, is here returned to its original genus, and the new combination Condorlepis woodwardi (Waldman, 1971), is proposed for this Early Cretaceous coccolepidid from Australia.
Acknowledgments
We thank Thierry Malvesy, collection manager at the MHNN, and Laure Bapst (MHNN) for their assistance to our study of the relocated holotype specimen. Oliver Rauhut (SNSB-BSPG) and Markus Wilmsen (MMG-SNSD) for permission to study material housed at their institutions. We are indebted to the following colleagues for providing digital photographs: Laure Bapst, Christina Byrd (MCZ), Andreas Hecker (JME), Helmut Tischlinger (Stammham, Germany). Finally, we want to thank the editor Daniel E. Barta and reviewers Eric Hilton (Virginia Institute of Marine Science, Gloucester Point, Virginia, USA) and Lionel Cavin (Natural History Museum of Geneva, Switzerland) for their very helpful comments and suggestions. The work of ALA is financially supported by the German Research Foundation (DFG) through project LO 1405/6-1.
References
Agassiz, L. 1833–1844. Recherches sur les Poissons Fossiles. 5 vols. 1420 pp., 396 pls., with supplements. Petitpierre, Neuchâtel.
Berg, L.S. 1940. Classification of fishes both recent and fossil. Travaux de l’Institut zoologique de l’Académie des Sciences de l’URSS 5 (2): 1–517.
Bordas, A. 1943. Peces del Cretáceo del río Chubut (Patagonia). Physis 19: 313–318.
Bürgin, T. 1990. Reproduction in Middle Triassic actinopterygians; complex fin structures and evidence of viviparity in fossil fishes. Zoological Journal of the Linnean Society 100: 379–391. Crossref
Cope, E.D. 1887. Zittel’s manual of palaeontology. American Naturalist 17: 1014–1019.
Ebert, M., Bapst, L., Kölbl-Ebert, M., Malvesy T., and López-Arbarello, A. 2021. Coccolepis Agassiz, 1843 (Coccolepididae, Chondrostei) from the Upper Jurassic Solnhofen-Archipelago—rediscovery of the type specimen and open questions. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 300: 11–22. Crossref
Frickhinger, K.A. 1991. Fossilien Atlas Fische. 1088 pp. Mergus, Melle.
Frickhinger, K.A. 1995. Fossil Atlas Fishes. 1088 pp. Mergus, Melle.
Gardiner, B.G. 1960. A revision of certain actinopterygian and coelacanth fishes, chiefly from the Lower Lias. Bulletin of the British Museum [Natural History], Geology 4: 239–384. Crossref
Gorizdro-Kulczycka, F. 1926. Fishes of the Karatau shales [in Russian]. Izvestiâ Sredne-azâtskogo Komiteta 1: 184–192.
Hilton, E.J., Grande, L., and Bemis, W.E. 2004. Morphology of †Coccolepis bucklandi Agassiz, 1843 (Actinopterygii, †Coccolepidae) from the Solnhofen Lithographic Limestone deposits (Upper Jurassic, Germany. In: G. Arratia and A. Tintori (eds.), Mesozoic Fishes 3—Systematics, Paleoenvironments and Biodiversity, 209–238. Dr. Friedrich Pfeil, Munich.
ICZN 1999. International Code of Zoological Nomenclature. 306 pp. The International Trust for Zoological Nomenclature, London.
Keupp, H., Koch, R., Schweigert, G., and Viohl, G. 2007. Geological history of the Southern Franconian Alb—the area of the Solnhofen Lithographic Limestone. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 245: 3–21. Crossref
Kirkland, J.I. 1998. Morrison fishes. Modern Geology 2: 503–533.
Kölbl-Ebert, M. and Cooper, B.J. 2019. Solnhofener Plattenkalk: a heritage stone of international significance from Germany. Geological Society, London, Special Publications 486: 103–113. Crossref
Lambers, P.H. 1992. On the Ichthyofauna of the Solnhofen Lithographic Limestone Upper Jurassic, Germany). 336 pp. Rijksuniversiteit, Groningen.
Lambers, P.H. 1999. The actinopterygian fish fauna of the Late Kimmeridgian and Early Tithonian “Plattenkalk” near Solnhofen (Bavaria, Germany): state of the art. Geologie en Mijnbouw 78: 215–229. Crossref
Liu, T.S. 1957. A new Cretaceous palaeoniscid fish from Yumen of the Chiuchuan Basin, western Kansu. Vertebrata Palasiatica 1: 103–122.
Lombardo, C. 1999. Sexual dimorphism in a new species of the actinopterygian Peltopleurus from the Triassic of northern Italy. Palaeontology 42: 741–760. Crossref
López-Arbarello, A., Sferco, E., and Rauhut, O.W.M. 2013. A new genus of coccolepidid fishes (Actinopterygii, Chondrostei) from the continental Jurassic of Patagonia. Palaeontologia Electronica 16.1.7A. Crossref
Matthews, S.C. 1973. Notes on open nomenclature and on synonymy lists. Palaeontology 16: 713–719.
Moser, M, Schwarz, A., Kügel, S., Lehmann, T., Reichert, W., Weibulat, T., Weiss, M., and Triebel, D. 2017. Pilotprojekt zur Digitalisierung im Rahmen der internationalen Biodiversitätsforschung: Die fotografische und datentechnische Erfassung der fossilen Strahlenflosser (Actinopterygii) in der Bayerischen Staatssammlung für Paläontologie und Geologie. Zitteliana 89: 291–304.
Müller, J. 1845. Über den Bau und die Grenzen der Ganoiden und das natürliche System der Fische. Abhandlungen der Akademie der Wissenschaften zu Berlin (1844) 1845: 117–216.
Olive, S., Taverne, L., and López-Arbarello, A. 2019. A new genus of coccolepidid actinopterygian from the Cretaceous Iguanodon-bearing locality of Bernissart, Belgium. Cretaceous Research 95: 318–335. Crossref
Rauhut, O.W.M., López-Arbarello, A., Röper, M., and Rothgaenger, M. 2017. Vertebrate fossils from the Kimmeridgian of Brunn: the oldest fauna from the Solnhofen Archipelago (Late Jurassic, Bavaria, Germany). Zitteliana 89: 305–329.
Schultze, H.-P. and Arratia, G. 2015. Knochenfische im weiteren Sinne (Osteichthyes oder Osteognathostomata. In: G. Arratia, H.-P. Schultze, H. Tischlinger, and G. Viohl (eds.), Solnhofen—Ein Fenster in die Jurazeit, 1, 360–380. Verlag Dr. Friedrich Pfeil, Munich.
Skrzycka, R. 2014. Revision of two relic actinopterygians from the Middle or Upper Jurassic Karabastau Formation, Karatau Range, Kazakhstan. Alcheringa: An Australasian Journal of Palaeontology 38: 364–390. Crossref
Sun, Z.Y., Lombardo, C., Tintori, A., Jiang, D.Y., Hao, W.C., Sun, Y.L., and Lin, H.Q. 2012. Fuyuanperleidus dengi Geng et al. 2012 (Osteichthyes, Actinopterygii) from the Middle Triassic of Yunnan Province, South China. Rivista Italiana di Paleontologia e Stratigrafia 118: 359–373.
Sytchevskaya, E.K. 2006. Iyalepis nom. nov., a new replacement name for Angarichthys Sytchevskaya et Yakovlev V. N., 1985 (Pisces, Palaeonisciformes, Coccolepidae). Paleontological Journal 40: 339. Crossref
Sytchevskaya, E.K. [Sytčevskaâ, E.K.] and Yakovlev, V.N. [Âkovlev, V.N.] 1985. Fishes. Jurassic continental biocenoses of southern Siberia and adjacent areas [in Russian]. Trudy Paleontologičeskogo Instituta RAN SSSR 113: 132–136.
Tischlinger, H. and Schweigert, G. 2020. Neuer Ammonitenfund ermöglicht Alterseinstufung der Fossillagerstätte Ettling. New ammonite record allows dating of the Ettling Fossillagerstätte. Archaeopteryx 36: 54–63.
Tischlinger, H. and Völkl-Costantini, P. 2015. Fundbergung, Präparation, Dokumentation. In: G. Arratia, H.-P. Schultze, H. Tischlinger, and G. Viohl (eds.), Solnhofen. Ein Fenster in die Jurazeit, 101–108. Dr. Friedrich Pfeil, Munich.
Traquair, R.H. 1911. Les poissons wealdiens de Bernissart. Mémoires du Musée royal d’Histoire naturelle de Belgique 21: 1–65.
Vetter, B. 1881. Die Fische aus dem lithographischen Schiefer im Dresdener Museum. Mittheilungen aus dem Koeniglichen Mineralogisch–Geologischen und Praehistorischen Museum in Dresden 4: 1–118.
Viohl, G. 2015. Der geologische Rahmen: die Südliche Frankenalb und ihre Entwicklung. In: G. Arratia, H.-P. Schultze, H. Tischlinger, and G. Viohl (eds.), Solnhofen. Ein Fenster in die Jurazeit, 56–62. Dr. Friedrich Pfeil, Munich.
Wagner, A. 1863. Monographie der fossilen Fische aus den lithographischen Schiefern Bayerns. Zweite Abtheilung. Abhandlungen der Bayerische Akademie von Wissenschaften II 9: 611–748.
Waldman, M. 1971. Fish from the freshwater Lower Cretaceous of Victoria, Australia, with comments on the palaeo-environment. Special Papers in Palaeontology 9: 1–124.
Wang, N.-C. 1977. Jurassic fish and their stratigraphic significance from the Hengyang Region of Lingling Co., Hunan Province. Vertebrata PalAsiatica, 15: 233–243.
Woodward, A.S. 1890. On some new fishes from the English Wealden and Purbeck beds. Proceedings of the Zoological Society of London 1890: 347–353.
Woodward, A.S. 1891. Catalogue of the Fossil Fishes in the British Museum (Natural History). Part 2. 567 pp. Longmans and Co., London. Crossref
Woodward, A.S. 1895. The fossil fishes of the Talbragar Beds (Jurassic?). Memoirs of the Geological Survey of New South Wales, Palaeontology 9: 1–27.
Xu, G.H. and Zhao, L.J. 2016. A Middle Triassic stem-neopterygian fish from China shows remarkable secondary sexual characteristics. Science Bulletin 61: 338–344. Crossref
Acta Palaeontol. Pol. 66 (4): 837–846, 2021
https://doi.org/10.4202/app.00873.2021