
First palaeoscolecid from the Cambrian (Drumian, Miaolingian) Marjum Formation of western Utah, USA
WADE W. LEIBACH, RUDY LEROSEY-AUBRIL, ANNA F. WHITAKER, JAMES D. SCHIFFBAUER, and JULIEN KIMMIG
Leibach, W.W., Lerosey-Aubril, R., Whitaker, A.F., Schiffbauer, J.D., and Kimmig, J. 2021. First palaeoscolecid from the Cambrian (Drumian, Miaolingian) Marjum Formation of western Utah, USA. Acta Palaeontologica Polonica 66 (3): 663–678.
The middle Marjum Formation is one of five Miaolingian Burgess Shale-type deposits in Utah, USA. It preserves a diverse non-biomineralized fossil assemblage, which is dominated by panarthropods and sponges. Infaunal components are particularly rare, and are best exemplified by the poorly diverse scalidophoran fauna and the uncertain presence of palaeoscolecids amongst it. To date, only a single Marjum Formation fossil has been tentatively assigned to the palaeoscolecid taxon Scathascolex minor. This specimen and two recently collected worm fragments were analysed in this study using scanning electron microscopy and energy dispersive X-ray spectrometry. The previous occurrence of a Marjum Formation palaeoscolecid is refuted based on the absence of sclerites in the specimen, which we tentatively assign to an unidentified species of Ottoia. The two new fossils, however, are identified as a new palaeoscolecid taxon, Arrakiscolex aasei gen. et sp. nov., characterized by the presence of hundreds of size-constrained (20–30 µm), smooth-rimmed, discoid plates on each annulus. This is the first indisputable evidence for the presence of palaeoscolecids in the Marjum biota, and a rare occurrence of the group in the Cambrian of Laurentia. Palaeoscolecids are now known from nine Cambrian Stage 3–Guzhangian localities in Laurentia, but they typically represent rare components of the biotas.
Key words: Scalidophora, Burgess Shale-type preservation, Great Basin, House Range, Laurentia.
Wade W. Leibach [wade.leibach@gmail.com], Department of Geological Sciences, University of Missouri, Columbia, MO 65211, USA; X-ray Microanalysis Core, University of Missouri, Columbia, MO 65211, USA; Biodiversity Institute, University of Kansas, Lawrence, KS 66045, USA and Department of Geology, University of Kansas, Lawrence, KS 66045, USA.
Rudy Lerosey-Aubril [rudy_lerosey@fas.harvard.edu], Department of Organismic and Evolutionary Biology and Museum of Comparative Zoology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, USA.
Anna F. Whitaker [a.whitaker@mail.utoronto.ca], University of Toronto Mississauga, Department of Chemical and Physical Sciences, 3359 Mississauga Road, Mississauga, Ontario L5L 1C6, Canada.
James D. Schiffbauer [schiffbauerj@missouri.edu], Department of Geological Sciences, University of Missouri, Columbia, MO 65211, USA and X-ray Microanalysis Core, University of Missouri, Columbia, MO 65211, USA.
Julien Kimmig [jkimmig@psu.edu], Earth and Mineral Sciences Museum and Art Gallery, Pennsylvania State University, University Park, PA 16802, USA.
Received 14 January 2021, accepted 18 March 2021, available online 6 September 2021.
Copyright © 2021 W.W. Leibach et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Marjum Formation (Drumian Stage, Miaolingian; ca. 504.5–500.5 Ma) of western Utah preserves one of the most diverse assemblages of Cambrian soft-bodied fossils in Laurentia (Robison et al. 2015; Pates et al. 2021). It is one of five Miaolingian Burgess Shale-type Lagerstätten in Utah, and one of three located in the House Range (Robison et al. 2015; Foster and Gaines 2016; Lerosey-Aubril et al. 2018, 2020; Kimmig 2021). Its fauna comprises close to a hundred species, among which roughly 45% are non-biomineralizing taxa. These “soft-bodied” components belong to a variety of groups, including annelids, cnidarians, cyanobacteria, deuterostomes, scalidophorans, and sponges (Robison et al. 2015). Worms are particularly rare and essentially represented by species of the scalidophoran genera Ottoia and Selkirkia (Conway Morris and Robison 1986). Another taxon, the palaeoscolecid Scathascolex minor, was tentatively regarded as present in the Marjum Formation (Smith 2015) based on the reinterpretation of a fossil originally assigned to Ottoia prolifica (Conway Morris and Robison 1986). However, this assumption has yet to be confirmed by detailed examination of the specimen. To date, this remains the only claim for the presence of palaeoscolecids in the Marjum Formation, while these organisms are known from many other Cambrian Burgess Shale-type faunas.
With more than 60 species and 45 genera described, the class Palaeoscolecida is the most diverse group of fossil scalidophorans. Traditionally regarded as part of total-group Priapulida, the class may not represent a clade, and no consensus has been reached as to how it should be defined (Harvey et al. 2010; Smith 2015). Morphologically, palaeoscolecids are scalidophorans with an elongate, parallel-sided, annulated body bearing phosphatic plates and a straight alimentary canal (Smith 2015). Taphonomically, palaeoscolecids are preserved as macroscopic carbonaceous compressions, though their presence has been additionally surmised from recovery of isolated plates and scleritome fragments. In most forms, considered palaeoscolecids sensu stricto by Harvey et al. (2010), numerous, micrometric plates form a scleritome that essentially covers the whole body, except the introvert. These sclerites often vary in size, morphology, and arrangement, which allows the discrimination of two or more types (“plates”, ”platelets”, and ”microplates”, see Topper et al. 2010; García-Bellido et al. 2013; and Whitaker et al. 2020, for different definitions of those terms). Conversely, a handful of Cambrian taxa referred to as “palaeoscolecids” share with palaeoscolecids sensu stricto an elongate body bearing plates, but differ by characters pertaining to those plates: their limited number, their unusually large sizes, and, in some taxa, their non-phosphatic composition (Harvey et al. 2010). This is well-exemplified by Cricocosmia, in which each annulus bears a single pair of large, non-biomineralized plates (Han et al. 2007a; Steiner et al. 2012; Vannier and Martin 2017).
Spanning from Cambrian Stage 3 to Silurian strata, palaeoscolecids were globally distributed, with occurrences of compressed representatives reported from: (i) Avalonia (Whittard 1953; Conway Morris 1997; Botting et al. 2012); (ii) eastern (Glaessner 1979; García-Bellido et al. 2013) and western Gondwana (Conway Morris and Robison 1986; Kraft and Mergl 1989; García-Bellido et al. 2013; Gutiérrez-Marco and García-Bellido 2015; Martin et al. 2016; Kouraiss et al. 2018); (iii) Laurentia (Robison 1969; Conway Morris and Robison 1986; Butterfield 1994; LoDuca et al. 1997; Balthasar 2004; Johnston et al. 2009; Conway Morris and Peel 2010; Smith 2015; Lerosey-Aubril et al. 2018; Wendruff et al. 2020; Whitaker et al. 2020); (iv) Baltica (Streng et al. 2017); (v) South China (e.g., Sun and Hou 1987; Hou and Sun 1988; Lin 1995; Luo et al. 1999; Han et al. 2003a, b, 2007a, b; Hu et al. 2008, 2012; Wang et al. 2014; Muir et al. 2014; Liu et al. 2016, 2018; Peng et al. 2016, 2020; Yang and Zhang 2016a, b; Zhu et al. 2016; Yang et al. 2018); and (vi) Siberia (Barskov and Zhuravlev 1988; Ivantsov and Wrona 2004; Ivantsov and Zhuralev 2005). From these occurrences, about 30 genera have thus far been described. Microscopic fragments of secondarily phosphatized cuticle and associated scleritome are known from Cambrian Stage 3 to Paibian Orsten-type deposits of eastern Gondwana (Brock and Cooper 1993; Müller and Hinz-Schallreuter 1993; Harvey et al. 2010; Topper et al. 2010), Siberia (Ivantsov and Wrona 2004), and South China (Zhang and Pratt 1996; Harvey et al. 2010; Duan et al. 2012; Duan and Dong 2013; Liu et al. 2018). Though inherently more limited, these microfossils have allowed for the description of 15 additional palaeoscolecid genera. Importantly, none of the genera described from compression macrofossils are known from Orsten-type scleritome fragments, and vice versa. Palaeoscolecid isolated plates are also common components of Cambrian Stage 2 to Silurian small shelly faunas worldwide (Topper et al. 2010), where they have been assigned to several distinct genera including: Hadimopanella, Kaimenella, Milaculum, and Utahphospha (e.g., Müller 1973; Müller and Miller 1976; Gedik 1977; Müller and Hinz-Schallreuter 1993). These generic names have also been assigned to gross plate morphotypes in both compression and Orsten-type fossils (e.g., Hadimopanella-type plate). Plates have also been preserved as carbonaceous microfossils (Butterfield and Harvey 2012; Slater et al. 2017).
In this contribution, we describe Arrakiscolex aasei gen. et sp. nov., a new palaeoscolecid worm from the Marjum Formation, and re-assess Smith’s (2015) suggestion of the presence of palaeoscolecid Scathascolex minor in these deposits. Further, we also discuss the possible reasons and consequences of the variability in palaeoscolecid biodiversity and abundance between Cambrian Lagerstätten in Laurentia.
Institutional abbreviations.—KUMIP, Division of Invertebrate Paleontology, Biodiversity Institute, University of Kansas, Lawrence, Kansas, USA.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:AC0E9094-6BDC-4BC1-B45A-3EF8A88AAA35.
Material and methods
The Miaolingian (Drumian–Guzhangian) Marjum Formation outcrops in the House Range of western Utah, within the Great Basin of the western United States (Fig. 1A, B). It represents a 430-meter-thick succession of thin-bedded limestone inter-bedded with shale/mudstone (Foster and Gaines 2016; Fig. 1B), occupying the upper Ptychagnostus atavus Zone through the lower Lejopyge laevigata Zone (Robison and Babcock 2011). Exceptional preservation in the Marjum Formation is restricted to an interval extending from 60 to 200 m from its base, possibly up to 300 m, that corresponds to the lower P. punctuosus Zone (Pates et al. 2021). The Marjum Formation is conformably underlain by the Wuliuan–Drumian Wheeler Formation and overlain by the Guzhangian Weeks Formation. The three units form a continuous depositional sequence that filled a local asymmetric basin, the House Range Embayment (Rees 1986). The Wheeler Formation is regionally extensive, found throughout the House Range and into the neighbouring Drum Mountains. Comparatively, the Marjum Formation is restricted to the House Range, thinning northward, and the Weeks Formation outcrops only in the southern part of the House Range (Foster and Gaines 2016). The Marjum Formation correlates to the Pierson Cove Formation in the Drum Mountains, Wah Wah Mountains, Fish Springs Range, and Dugway Range; the upper Limestone of the Cricket Mountains; and the upper Abercrombie Formation and Young Peak Dolomite in the Deep Creek Range (Foster and Gaines 2016). The Marjum Formation was deposited on a gently sloping ramp connecting the high-energy, shallow-water carbonate platform to the quiet, deep-water House Range Embayment (Foster and Gaines 2016), and accordingly records important spatial and temporal variations in depositional settings.
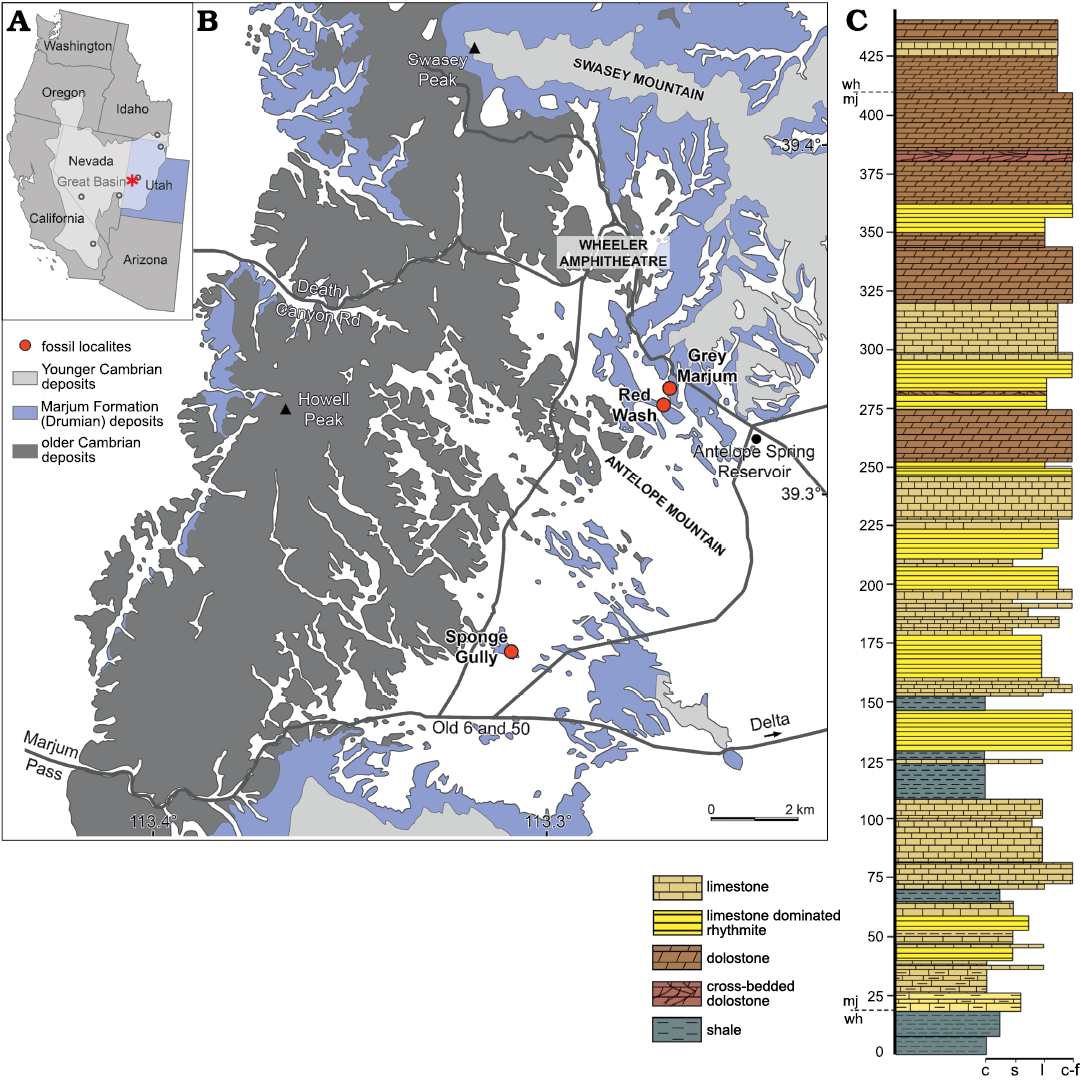
Fig. 1. Geographic and geologic setting of the Marjum Formation. A. Map of western United States showing the locations of the main Cambrian Konservat-Lagerstätten (circles) of the Great Basin (light grey area), including the Marjum Formation in the House Range (asterisk) of western Utah. B. Simplified geological map of the central House Range, showing the geographic distribution of the Marjum Formation and the localities mentioned in the text. C. Stratigraphic column of the Marjum Formation (mj) at the Wheeler Amphitheater (wh) (adapted from Smith 2007). Data relating to the spatial distributions of Cambrian strata are derived from Hintze (1980). Abbreviations: c, cover; s, slope; l, ledge; c-f, cliff-forming.
The material considered in this study comprises three specimens, reposited at the University of Kansas, Biodiversity Institute, Division of Invertebrate Paleontology collections (KUMIP 204770, 314100, 314101). KUMIP 204770 was collected by Thomas T. Johnson at the Sponge Gully locality (“Sponge Gulch”, locality 347 of Robison and Babcock 2011; Fig. 1A2), about 5 km NE of Marjum Pass. This site has yielded agnostoids and other trilobites (Robison and Babcock 2011), along with a diverse soft-bodied panarthropod assemblage (Richard Robison, personal communication 2019) that includes Branchiocaris pretiosa, Leanchoilia? cf. L. protagonia, Meristosoma sp., Naraoia compacta, Nettapezoura basilikos, Perspicaris? ellipsopelta, Tuzoia guntheri, and possibly Dicranocaris guntherorum (Robison 1984; Briggs and Robison 1984; Briggs et al. 2008; Pates et al. 2018, 2021). These taxa co-occur at this locality with a rich poriferan fauna composed of Choia carteri, Choia utahensis, Diagoniella hindei, Diagnoniella sp., Leptomitella metta, Protospongia? elongata, Testiispongia venula, and Valospongia gigantis (Rigby 1983; Richard Robison, personal communication 2019). The holotype and sole Marjum Formation specimen of Selkirkia willoughbyi was also recovered from the Sponge Gully locality (Conway Morris and Robison 1986). The main excavation pits that used to form the Sponge Gully locality were filled by the Bureau of Land Management.
The two other KUMIP specimens, KUMIP 314100 and 314101, were collected by Arvid Aase at a locality hereby called the Grey Marjum, a couple hundred meters north of the Red Wash locality near the entrance of the Wheeler Amphitheatre (Fig. 1A2). This relatively recent excavation site is less well-known. It seems to expose the same strata as those of the Red Wash, with the differences in rock colouration between the two sites likely resulting from distinct weathering histories (Robert Gaines and Jacob Skabelund, personal communication 2019). The soft-bodied fauna recovered here includes panarthropods (e.g., Naraoia compacta, a possible leanchoiliid), sponges (Crumillospongia? sp.; Diagoniella? sp.), a hemichordate (Tarnagraptus sp.), and the possible vetulicolan Skeemella clavula (Kimmig et al. 2020).
The specimens were photographed under non-polarized light when immersed in dilute ethanol using a GIGAmacro Magnify2 robotic imaging system. High-resolution composite images of the whole specimens were created by stitching together z-stacked photographs of their parts using GIGAmacro Flow. Affinity Photo was used for basic photomanipulation (colour, contrast, and brightness) of those composite images.
The specimens were examined using scanning electron microscopy (SEM) with integrated energy dispersive X-ray spectrometry (EDS). SEM-EDS analyses were conducted using a Zeiss Sigma 500 VP variable pressure SEM with dual, co-planar Bruker XFlash EDS detectors (30 mm2), located at the X-ray Microanalysis Core Facility at the University of Missouri. Using the Fibics Incorporated ATLAS interface, large-area secondary and backscattered electron mosaics were conducted for all three specimens, two of which (KUMIP 204770 and 314100) included both part and counterpart imaging. Secondary electron images (topography) were collected using a low-vacuum-specific cascade current detector (C2D), and backscattered images (composition) using a high-definition 5-segment backscatter detector (HDBSE). All imaging analyses were conducted with the following operating conditions: 14 mm ± 1 mm working distance (minor differences to allow for variation in sample thickness or topography) for basic imaging and EDS, 16.5 mm minimum working distance for ATLAS mosaic imaging, 20 keV beam accelerating voltage, 40 nA beam current, 20 Pa chamber pressure (low vacuum), 60 µm aperture for imaging, and 120 µm aperture for EDS analysis. EDS analyses were conducted with both detectors active (in tandem), aiding in the removal of topographic artefacts and improving X-ray count rate (> 150 kilocounts per second, combined, on average).
The terminology used in this contribution follows Whitaker et al. (2020), including the use of the term “plates”, instead of “sclerites” (see also Smith 2015).
Systematic palaeontology
Unranked Scalidophora Lemburg, 1995
Phylum uncertain
Class uncertain
Family Ottoiidae Walcott, 1911
Genus Ottoia Walcott, 1911
Type species: Ottoia prolifica Walcott, 1911; Burgess Shale Formation, Wulian, Cambrian, British Columbia, Canada.
Ottoia? sp.
Figs. 2, 3.
1986 Ottoia prolifica (Walcott, 1911); Conway Morris and Robison 1986: figs. 1, 2.
1991 Ottoia prolifica (Walcott, 1911); Robison 1991: fig. 6.2.
2015 Scathascolex minor gen. et sp. nov.; Smith 2015: 974.
2016 Ottoia prolifica; Foster and Gaines 2016: 311, fig. 20A.
2017 Ottoia prolifica; Broce and Schiffbauer 2017: 602, fig. 1A–C.
Material.—KUMIP 204770 (part and counterpart) from the Sponge Gully locality, Utah, USA (Fig. 1B2). Middle part (Ptychagnostus punctuosus Agnostoid Zone) of the Drumian Marjum Formation.
Description.—KUMIP 204770 measures 4.92 mm in maximum width and 48 mm in length, when uncoiled and represents a complete individual (Fig. 2). It displays anteriorly a partially everted proboscis (eversion stage 3 of Conway Morris 1977; Fig. 2A1, A2, A6), which is approx. 1.87 mm long and widest distally (approx. 1.28 mm); there is no clear evidence for the preservation of teeth on this organ, however, there are several observable introvert hooks present (Fig. 2A2, A6). The trunk is almost parallel-sided, except in its posterior fifth where it progressively tapers posteriorly. About 110 well-delimitated, densely distributed (approx. 5 annuli per mm), thin trunk annuli are visible (Fig. 2A4). The presence of caudal hooks cannot be confidently established, but two rather large structures resembling the “conical projections” of Harvey et al. (2010: fig. 2E–G) are interpreted as the basal parts of a single pair of strong hooks (Fig. 2A3). A non-everted part of the proboscis, about as long as the everted part, is visible in the anterior trunk region (Fig. 2A2). It is continued posteriorly by a short portion of a slightly wider and not as well-delimitated intestine, represented by a darker coloured, carbon-rich region. No plates, phosphatic or otherwise, were observed, despite careful examination of the surface of the trunk using SEM/EDS (Fig. 3). EDS analysis reveals that the fossil is predominantly composed of C, O, Fe, and Mg.

Fig. 2. Ottoid scalidophoran Ottoia? sp. (KUMIP 204770) from the Drumian, Cambrian Marjum Formation in the House Range of Utah, USA (Sponge Gully locality). A1, general view showing body orientation and location of close-ups; A2, detailed view of anterior region, showing the partially everted introvert and digestive tract; A3, detailed view of posterior region, showing location of possible bases of caudal hooks; A4, SEM micrograph showing trunk annulation, arrowheads point to fossil margins; A5, SEM micrograph of anterior region, showing introvert and a few introvert hooks, arrowheads point to fossil margins; A6, SEM micrograph of introvert hooks.

Fig. 3. SEM micrograph and SEM-EDS elemental maps of the posterior region of KUMIP 204770 (Ottoia? sp.), from the Drumian, Cambrian Marjum Formation in the House Range of Utah, USA (Sponge Gully locality). A1, SEM micrograph of the analyzed area (posterior trunk region); A2–A6, SEM-EDS elemental maps of P, Ca, C, Fe, and Mg, respectively.
Remarks.—This specimen was originally assigned to the scalidophoran Ottoia prolifica by Conway Morris and Robison (1986), which purportedly extended the geographic and biostratigraphic distributions of this Wuliuan Burgess Shale species to the Drumian strata of Utah. More recently, Smith et al. (2015) questioned this Marjum Formation occurrence of Ottoia prolifica on the grounds that it lacked a detailed description. Later that year, Smith (2015: 974) tentatively reassigned KUMIP 204770 to the new palaeoscolecid taxon, Scathascolex minor Smith, 2015, but provided no support for this claim.
Palaeoscolecid affinities for KUMIP 204770 might be argued based on the presence of a subparallel-sided trunk, a feature exhibited by all members of the group, but not exclusive to it. The possible remains of a single pair of large caudal hooks would also be more compatible with an assignment to Palaeoscolecida than Ottoia (Harvey et al. 2010), the latter taxon possessing eight smaller caudal hooks (Conway Morris 1977). However, our SEM-EDS investigations demonstrate the complete absence of plates covering the trunk in this specimen, which strongly argues against a close relationship with S. minor or any member of the Palaeoscolecida. Moreover, S. minor has four, not two, caudal hooks and a significantly more elongate body (approx. 16 times longer than wide) than KUMIP 204770 (approx. 10 times longer than wide), despite being a rather short form for a palaeoscolecid. On the other hand, the present description of the Marjum specimen does not provide support for its assignment to Ottoia prolifica either. Emended by Smith et al. (2015), the diagnosis of this species now solely concerns characters pertaining to the teeth borne by the proboscis, whereas KUMIP 204770 preserves no such structures. A tentative assignment to the genus Ottoia may be proposed based on the observations of many of the characters forming the unique combination differentiating this genus: a cylindrical and extensible body with a pronounced external symmetry anteriorly, which is divided into an introvert with an extensible anterior end, and a trunk flexible in a single direction (U-shaped body) and comprising about a hundred annuli (Conway Morris 1977; Smith et al. 2015; Yang et al. 2016). Most of the diagnostic characters of Ottoia that cannot be observed, such as those pertaining to the proboscis teeth or the internal anatomy (gut, musculature), are likely absent due to the insufficient quality of preservation of KUMIP 204770. Additionally, whether a bursa is present along with the exact number of caudal hooks remain unresolved for KUMIP 204770. The investigated specimen only departs from the diagnosis of Ottoia in the absence of an expanded and bulbous posterior trunk region (Conway Morris 1977; Smith et al. 2015; Yang et al. 2016). However, if this morphological trait is generally well-expressed in Ottoia prolifica (for an exception, see Conway Morris 1977: pl. 2: 3) and Ottoia tricuspida (Smith et al. 2015), it is absent in Ottoia guizhouensis (Yang et al. 2016) and may therefore be inadequate for the definition of the whole genus Ottoia.
In summary, our investigations do not support a palaeoscolecid affinity for KUMIP 204770, but we concur with Smith et al. (2015) that this specimen cannot be confidently assigned to Ottoia prolifica either. Awaiting the study of three other specimens of this scalidophoran (Robison et al. 2015: fig. 52; Broce and Schiffbauer 2017: fig. 1F–H; RLA personal observation), we propose to tentatively assign this fossil to an undetermined species of Ottoia.
Class Palaeoscolecida Conway Morris and Robison, 1986 (sensu Smith 2015)
Family uncertain
Genus Arrakiscolex nov.
Zoobank LCID: urn:lsid:zoobank.org:act:04D3C924-950D-4E27-97 D6-C2F0E3B1D79F
Type species: Arrakiscolex aasei gen. et sp. nov. (by monotypy); see below.
Etymology: The name refers to the fictional planet of Arrakis in the novel “Dune” by Frank Herbert, which is inhabited by a species of armoured worm.
Diagnosis.—As for the monotypic type species.
Remarks.—The assignment of the new genus to the class Palaeoscolecida (sensu Smith 2015) is supported by the presence of a markedly annulated cuticle (Fig. 4) bearing numerous phosphatic plates. Arrakiscolex gen. nov. possesses small plates distributed over the surfaces of the preserved cuticle fragments, and as such can be regarded as a representative of the Palaeoscolecida sensu stricto of Harvey et al. (2010). The plate size, abundance, and distribution easily differentiate members of this new genus from taxa with few, large plates per annulus, such as Cricocosmia Hou and Sun, 1988 and Tabelliscolex Han, Zhang, Zhang, and Shu, 2003a. The small plates of the new genus are discoid, which excludes close relationships with a series of derived, mostly Ordovician genera characterized by elongate plates, whether these plates are oblong (e.g., Milaculum Müller, 1973, Palaeoscolex Whittard, 1953, Shergoldiscolex Müller and Hinz-Schallreuter, 1993) or rectangular (e.g., Loriciscolex Botting, Muir, Van Roy, Bates, and Upton, 2012, “Palaeoscolex” tenensis Kraft and Mergl, 1989, Plasmuscolex Kraft and Mergl, 1989, Pluoscolex Botting, Muir, Van Roy, Bates, and Upton, 2012). Overall, the plates of Arrakiscolex gen. nov. are somewhat reminiscent to Hadimopanella-type plates, except for the presence of a well-differentiated marginal rim and, more importantly, their entirely smooth surface. The latter character alone allows the discrimination of the new genus from most of the aforementioned genera and the remaining ones composing the class. Indeed, the largest plates of the scleritomes of the overwhelming majority of palaeoscolecids bear at least one, and usually more, nodes, whether they are granules (e.g., Bullascolex Botting, Muir, Van Roy, Bates, and Upton, 2012, Wudingscolex Hu, Steiner, Zhu, Luo, Forchielli, Keupp, Zhao, and Liu, 2012), well-developed tubercles (e.g., Palaeoscolex, some Wronascolex Ivantsov and Zhuralev, 2005; García-Bellido et al. 2013; Conway Morris 1997), or even fully formed spines (e.g., some Wronascolex; Ivantsov and Wrona 2004). Completely smooth discoid plates have been previously reported in only two formally described palaeoscolecid genera, the Wuliuan Scathascolex Smith, 2015 and Utahscolex Whitaker, Jamison, Schiffbauer, and Kimmig, 2020.
The scleritomes of Arrakiscolex gen. nov. and the Burgess Shale Formation Scathascolex (Smith, 2015) are both solely composed of homogeneously sized discoid plates (i.e., no platelets or microplates). The diameters of those plates are comparable in the two taxa (23–28 μm and 24–25 μm in the Marjum Formation taxon, 15–25 μm in the Burgess Shale Formation taxon), as are their gross morphologies, a moderately swollen, essentially smooth central region surrounded by a marginal rim. Yet, the marginal rim is more pronounced in the plates of Arrakiscolex gen. nov., which also show no indication of the five nodes/depressions sometimes observed on those of Scathascolex; the absence of these faint ornamental features is not taphonomic in origin, as the plates are excellently preserved in some parts of the Marjum Formation specimens (Fig. 5). More importantly, the two taxa can be easily distinguished by the radically different distribution patterns of their plates. In Scathascolex, each annulus bears two adjacent rows of plates near each of its boundaries with neighbouring annuli, and these pairs of rows are separated by a wide plate-free central region (Smith 2015). This strongly contrasts with the condition observed in Arrakiscolex gen. nov., where the plates are homogeneously distributed over most of the annulus, only loosely forming “rows” (18–25 per annulus) and with only rare and small plate-free areas associated with annulus boundaries.
The Spence Shale Utahscolex resembles Scathascolex in possessing homogeneously single-sized plates, which form two sets of two rows separated by a plate-free central area on each annulus. However, those rows of plates may occasionally bifurcate when approaching one body margin (Whitaker et al. 2020), a feature unreported in Scathascolex. As mentioned above, this organization remains fundamentally different from that of Arrakiscolex gen. nov. Additionally, the new Marjum Formation taxon possesses smaller and morphologically more complex plates compared to Utahscolex, in which the plates are approx. 70–80 μm in diameter and devoid of marginal rim or any morphological feature. In summary, the two Cambrian palaeoscolecids from Utah are not only found in stratigraphically different (Wuliuan vs. Drumian) deposits roughly 275 km apart, they also possess highly distinguishable plates and plate organisation.
Arrakiscolex aasei sp. nov.
Figs. 4–6.
Zoobank LSID: urn:lsid:zoobank.org:act:6B251ADA-C57D-4E34-BA E3-F8325F504C91
?2010 Priapulida indet.; Bonino and Kier 2010: pl. 74d.
?2015 Wronascolex sp.; Robison et al. 2015: 88, fig. 54 (right).
?2018 Palaeoscolecida gen. et sp. nov. A; Lerosey-Aubril et al. 2018: 708, 711, figs. 3d–g, 5e, f, supplemental tables 1, 3.
Etymology: In honour of Arvid Aase, the collector of the type material.
Type material: Holotype, KUMIP 314100, a fragment of cuticle. Paratype, KUMIP 314101, a fragment of cuticle, from the type locality and horizon.
Type locality: The Grey Marjum locality, House Range, Utah, USA.
Type horizon: Middle part (Ptychagnostus punctuosus Zone) of the Drumian Marjum Formation.
Material.—The type material and possibly up to fifty, mostly incomplete specimens from the Guzhangian (Proagnostus bulbus Agnostoid Zone, Cedaria Trilobite Zone) upper Weeks Formation of the House Range of Utah, USA (see Remarks below).
Diagnosis.—Each annulus homogeneously covered by hundreds of plates (>500 on average). Plates equally sized, approx. 25 μm in diameter, discoid, and smooth, each composed of a slightly depressed to slightly swollen inner region surrounded by a pronounced marginal rim of varying width. Platelets and microplates are absent. Interannular boundary is represented by plate-free areas that alternate with plate-bearing areas (“columns”) connecting one annulus to the other.
Description.—The fossils are fragments preserved as two-dimensional compressions, either straight or slightly curved (Fig. 4). The holotype (KUMIP 314100) has a width of 3.95 mm at its widest and a length (uncoiled) of 38.14 mm (Fig. 4A). The paratype (KUMIP 314101) has a width of 1.98 mm at its widest and a length (uncoiled) of 25.12 mm (Fig. 4B). In both KUMIP 314100 and KUMIP 314101, there are approx. 3 annuli per mm (Fig. 4A2, B). The plates are similar in discoid morphology and size, ranging from 23–28 μm in diameter in KUMIP 314100 and 24–25 μm in diameter in KUMIP 314101 (Fig. 5A, B). Their external surface exhibits a smooth inner region, which varies from slightly depressed to slightly swollen compared to the thin raised rim surrounding it (Fig. 5B2). The width of this marginal rim is not constant both between and within individual plates, varying from 1.5–3 μm between different plates in KUMIP 314100 and 3–6 μm in KUMIP 314101, and 1–3 μm in thickness within plates in both specimens. With regard to their arrangement, the plates are evenly distributed, with one plate being typically surrounded by six others located, on average, at a spacing distance of approx. 13 μm (Fig. 5B1). Alignments of plates in any given direction have proven particularly challenging to follow, and therefore we use the term “rows” in a loose sense to depict gross alignments of plates along the transverse axis of the body. In KUMIP 314100, each annulus comprises 8–14 of such “rows”, which are composed of 60–90 plates as measured in the widest preserved parts of the scleritome (Fig. 5A1). Counting of all plates present on at least three annuli per specimen results in a varying plate number per annulus, from 300–860 in KUMIP 314100 and 370–630 in KUMIP 314101. Annuli boundaries are marked by areas lacking plates. These areas never form continuous transverse bands but are interrupted by plate-covered zones or “columns” connecting two consecutives annuli (Fig. 5A2). It is unclear whether the presence of bands of discontinuous plate-free areas is taphonomic. Both the holotype and paratype of Arrakiscolex aasei gen. et sp. nov. are relatively poorly preserved, missing both their anterior and posterior ends as well as gut tract. Accordingly, the morphology of the proboscis and the presence of caudal spines are unknown in the new taxon. It is unlikely, however, that these organisms were transported or disturbed much either prior to or after burial, given their relatively well-ordered delicate plates.
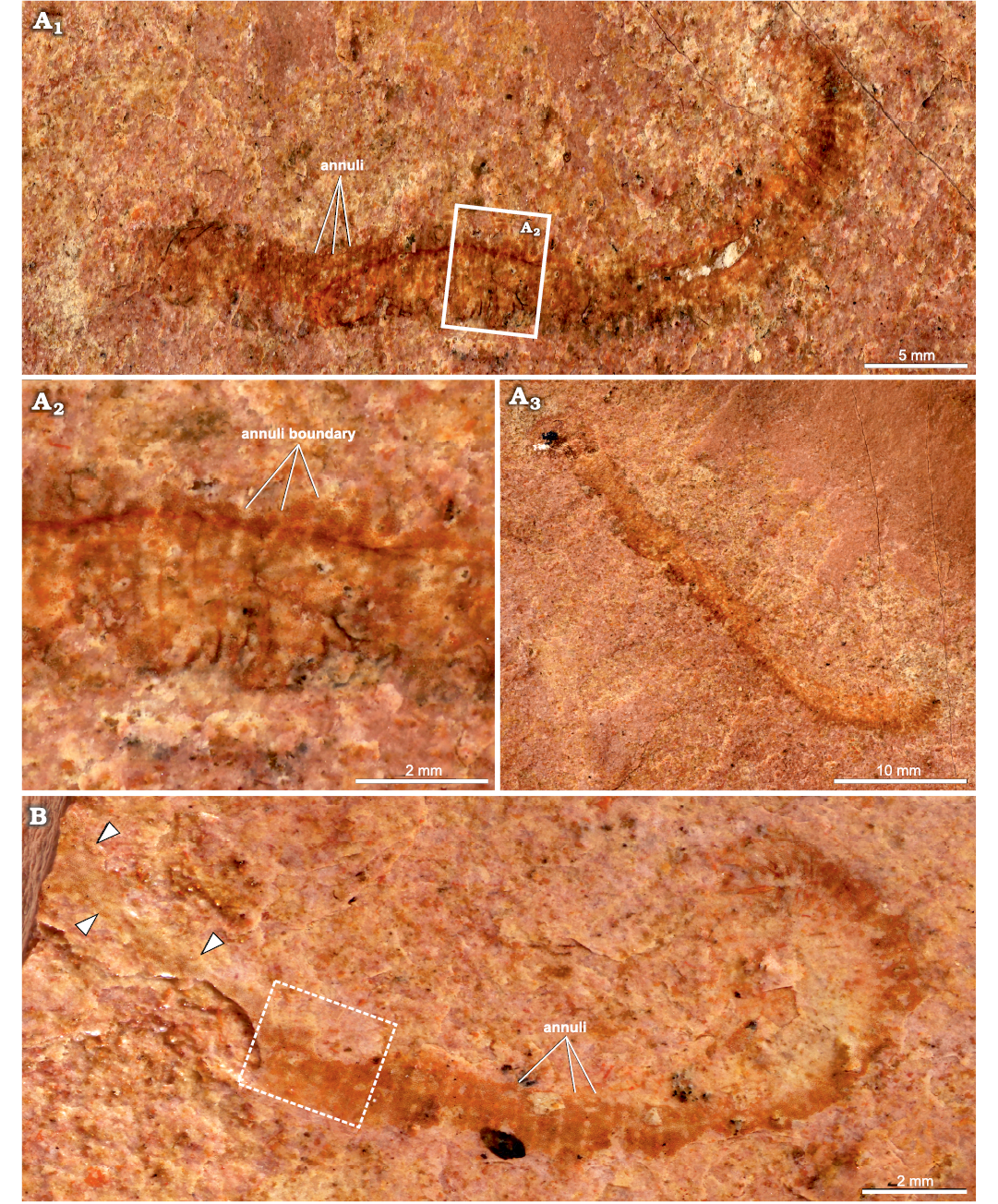
Fig. 4. Palaeoscolecid Arrakiscolex aasei gen et sp. nov. from the Drumian, Cambrian Marjum Formation in the House Range of Utah, USA (Grey Marjum locality). A. Holotype (KUMIP 314100). A1, general view of counterpart with location of close-up; A2, detailed view of trunk annulation; A3, general view of part. B. Paratype (KUMIP 314101), general view, showing trunk annulation and location of SEM micrograph in Fig. 5A1 (dashed line rectangle, arrowheads point to the limits of poorly preserved part of the fossil).
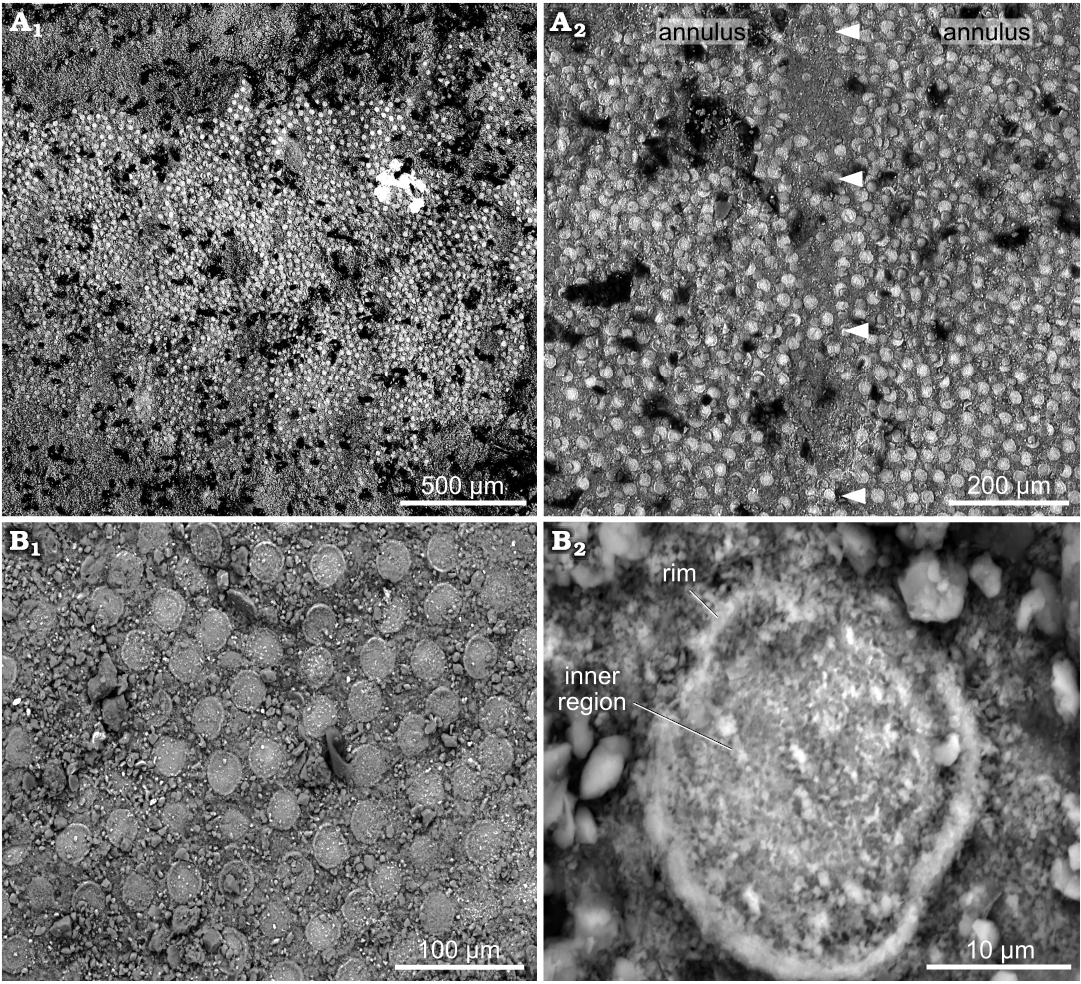
Fig. 5. Scleritome of the palaeoscolecid Arrakiscolex aasei gen et sp. nov. from the Drumian, Cambrian Marjum Formation in the House Range of Utah, USA (Grey Marjum locality). A. SEM micrographs of paratype (KUMIP 314101). A1, dense and homogeneous distribution of the plates on the surfaces of annuli; A2, interannular region showing alternation between plate-free areas and plate-covered areas (“columns”; arrowheads). B. SEM micrographs of holotype (KUMIP 314100). B1, homogeneous distribution of the plates; B2, single plate, showing the swollen inner region surrounded by a raised marginal rim.
Plates of the holotype (KUMIP 314100) are preserved as Fe, Mg, and P (Fig. 6A), and plates on the paratype (KUMIP 314101) are preserved as Fe, Mg, P, and Ca (Fig. 6B), which is not atypical of palaeoscolecid plates (e.g., Harvey et al. 2010; Botting et al. 2012; Martin et al. 2016; Whitaker et al. 2020). While both holotype and paratype have similar Fe, Mg, and P elemental maps, KUMIP 314100 has a visibly weaker Ca signature (Fig. 6A2) likely owing to the presence of two mineral bands with comparatively high Ca signal overwhelming the signature of the plates in relative abundance EDS elemental maps. Despite the presence of well-preserved plates, little soft tissue is preserved within either KUMIP 314100 or 314101, and what remains is primarily replaced by calcium carbonate, later altered to dolomite or high-Mg calcite (Fig. 6A2, B2).
Remarks.—We tentatively assigned to Arrakiscolex aasei gen. et sp. nov. about fifty specimens recovered from the overlying Guzhangian Weeks Formation, and reposited in the Department of Geology of the University of Utah and the Natural History Museum of Utah (Salt Lake City, USA), and the Back to the Past Museum (Cancun, Mexico). When studied macroscopically, these specimens all exhibit similar scleritomes (e.g., Lerosey-Aubril et al. 2018: figs. 3d–g, 5e, f), which are characterized by the presence of numerous, densely distributed plates over the whole surface of each annulus. Preliminary microscopic investigations on two specimens also revealed plates comparable in size and morphology to those described in the type material of the new Marjum Formation taxon (Lerosey-Aubril et al. 2018: fig. 5f). However, these microscopic observations are too scarce and spatially limited to allow their generalization to the whole scleritomes of those two individuals, and a fortiori to all the palaeoscolecid fossils found in the Weeks Formation, hence their tentative assignment to Arrakiscolex aasei gen. et sp. nov. Confirming this assignment will be a major objective of our investigations on this abundant, stratigraphically younger palaeoscolecid material.
Stratigraphic and geographic range.—Type locality and horizon only.
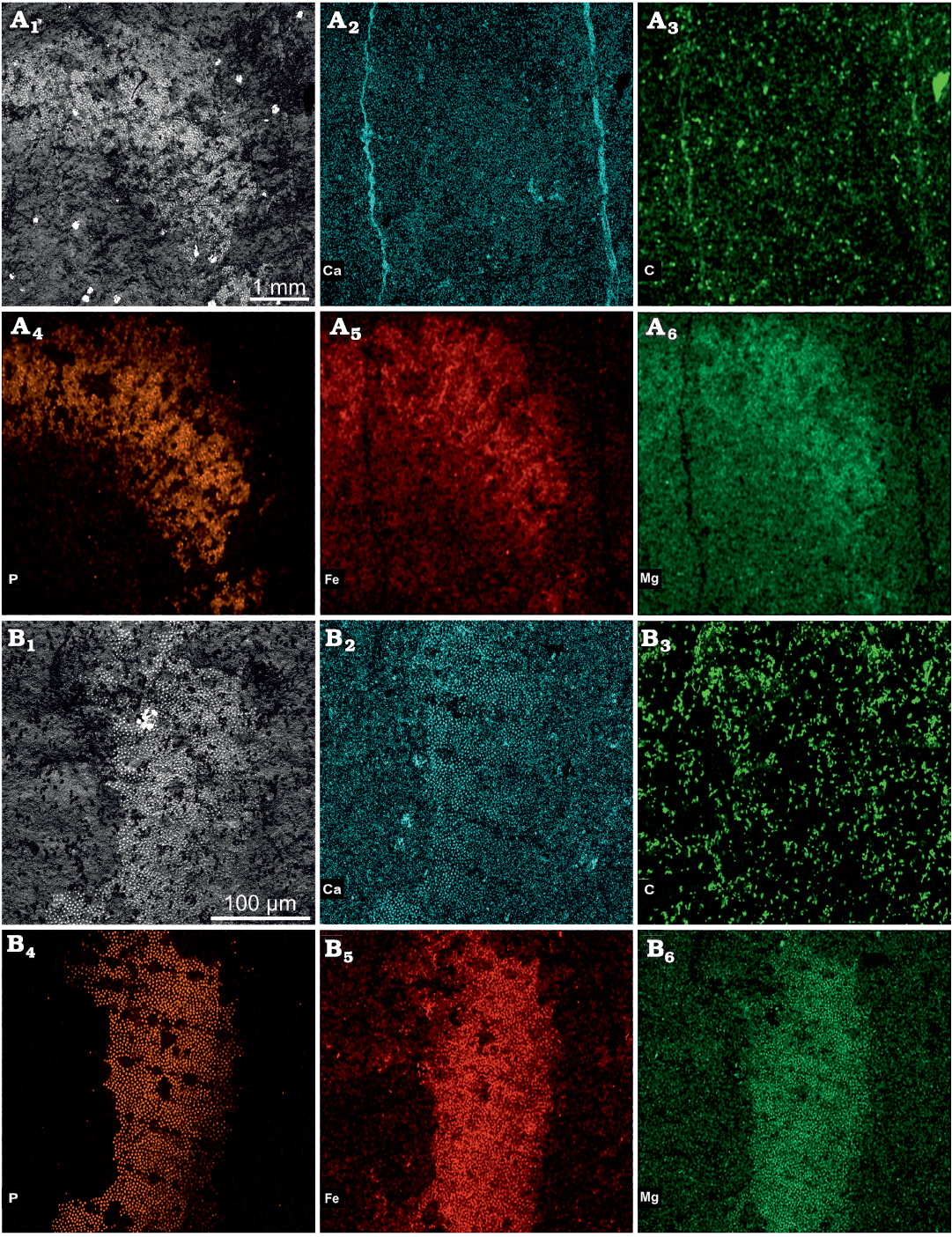
Fig. 6. SEM micrograph and SEM-EDS elemental maps of the type specimens of the palaeoscolecid Arrakiscolex aasei gen et sp. nov. from the Drumian, Cambrian Marjum Formation in the House Range of Utah (Grey Marjum locality). A. Holotype (KUMIP 314100). A1, detailed view of the analyzed area (curved extremity); A2–A6, SEM-EDS elemental maps of Ca, C, P, Fe, and Mg, respectively. B. Paratype (KUMIP 314101). B1, detailed view of the analyzed area (straight extremity); B2–B6, SEM-EDS elemental maps of Ca, C, P, Fe, and Mg, respectively.
Discussion
The Marjum palaeoscolecids and controls on the preservation of Cambrian palaeoscolecids in Laurentia.—Our reinvestigation of KUMIP 204770 shows that this fossil is demonstrably not a palaeoscolecid and possesses no features attributable to Scathascolex minor (contra Smith 2015); it is instead better tentatively interpreted as an undetermined species of Ottoia. Nonetheless, we provide clear evidence for the presence of the class Palaeoscolecida in the Marjum Formation, and more specifically a new representative, Arrakiscolex aasei gen. et sp. nov. Fossils from the Marjum Formation have been intensively collected by academic and non-academic palaeontologists for more than fifty years, resulting in the reports of about a hundred species from the interval associated with exceptional preservation (Pates et al. 2021). The discovery of Arrakiscolex aasei gen. et sp. nov. after such extensive collection and study, and the fact that it is represented by scant material, together suggest that this palaeoscolecid was a rare, or rarely preserved, component of the Marjum biota. Alternatively, it might be that palaeoscolecid fragments have been left in the field, as they are relatively small and not particularly appealing to collectors (Whitaker and Kimmig 2020).
Speaking in favour of scarcity of palaeoscolecids in the Marjum biota is the somewhat similar situation of the Burgess Shale Scathascolex minor, which is known from only seven specimens collected at the Walcott Quarry (Smith 2015), despite a preservation potential greater than that of the thousands of entirely non-biomineralized fossils recovered from this locality (Caron 2009). In general, palaeoscolecids have proven particularly elusive in the Cambrian of Laurentia, despite a fossil record that includes 11 Cambrian Stage 3–Guzhangian occurrences (Fig. 7). This is well-illustrated by the particularly limited number of specimens recovered in most Laurentian Lagerstätten: one each from the Mural Formation (Sperling et al. 2018; Uwe Balthasar personal communication 2021), the Colville Hills Lagerstätte of the Mount Cap Formation (Harvey and Butterfield 2011), the Latham Shale (Conway Morris and Peel 2010), and the Duchesnay Unit of the Chancellor Group (Johnston et al. 2009); two each from the Spence Shale (Whitaker et al. 2020) and the Marjum Formation (this study); three from the Kinzers Formation (Conway Morris and Peel 2010); and five from the Rosella Formation (two species; RLA unpublished data). Abundant palaeoscolecid materials (>20 specimens) have only been recovered from the Buen Formation (Sirius Passet Lagerstätte) in Greenland (Conway Morris and Peel 2010) and the Weeks Formation in Utah (Lerosey-Aubril et al. 2018). The relative inconspicuousness of this group in Laurentia is also evidenced by the presence of a single palaeoscolecid species in each of these remarkable Cambrian assemblages, excepting those from the Buen and Rosella formations which each contain two species (Fig. 7; Conway Morris and Peel 2010; RLA unpublished data).
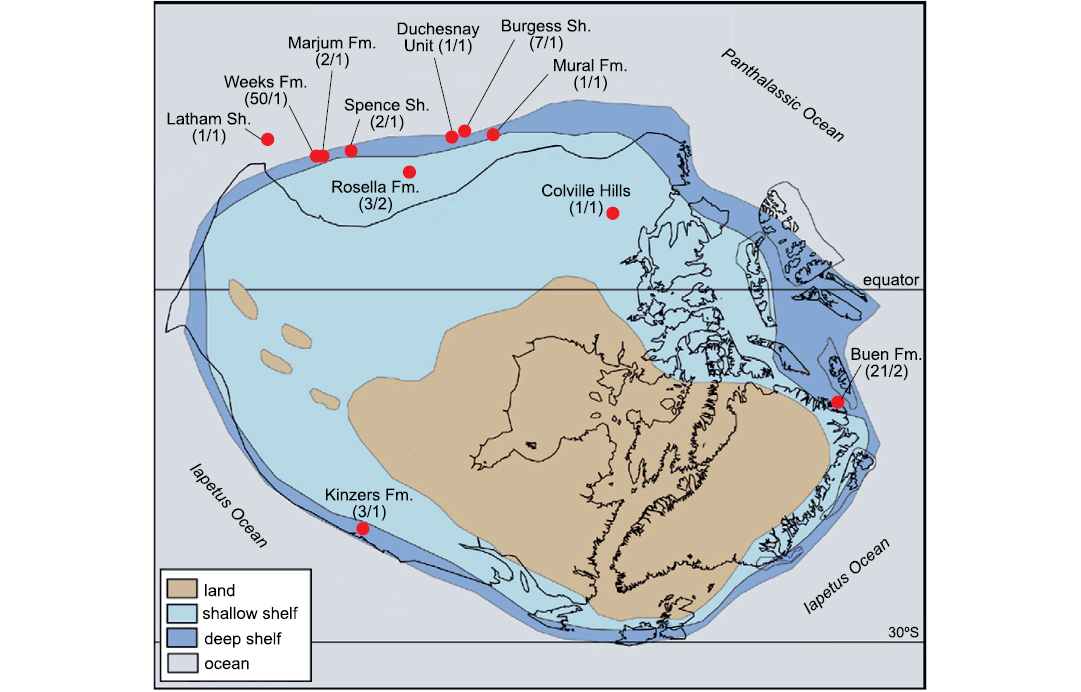
Fig. 7. Palaeogeographic distribution of Cambrian palaeoscolecids in Laurentia. For each occurrence, the numbers in parentheses refer to the number of palaeoscolecid specimens known (left) and the number of species they represent (right). The Rosella Formation Lagerstätte is positioned between the Lagerstätten of the American Great Basin and those of the Canadian Rockies, because the Cassiar terrane was located near present-day Idaho in the Cambrian, and subsequently migrated more than 2000 km to the North (Pope and Sears 1997). Abbreviations: Fm., Formation; Sh., Shale. Background map adapted from Cocks and Torsvik (2011).
The causes underlying the observed rarity of palaeoscolecids from the Cambrian of Laurentia is unclear. Smith (2015) convincingly argued that the scarcity of palaeoscolecid remains in the Burgess Shale Formation relates to the abundance of ecological competitors (e.g., annelids, priapulids), as opposed to any environmental or preservational specificities. To what extent this ecological explanation could be generalized to other Laurentian Lagerstätten is uncertain. Sharing similar depositional settings (Lerosey-Aubril et al. 2018), the Wheeler, Marjum, and Weeks formations provide strong evidence against a tight relationship between the abundance of palaeoscolecids at a given locality and parameters of the physical environment. These units represent different time intervals when conditions conducive to exceptional preservation developed within the same basin. However, around fifty palaeoscolecid fossils have been recovered from the Weeks Formation, versus only two in the Marjum Formation and none yet from the Wheeler Formation. Both the Weeks and Buen formations are notable outliers amongst other Cambrian Laurentian Lagerstätten, showing taphonomic features less common in most Burgess Shale-type deposits (Gaines 2014; Lerosey-Aubril et al. 2018; Harper et al. 2019). While fossils from some Burgess Shale-type localities contain phosphatized structures, such as secondarily phosphatized guts (Butterfield 2002, 2009; Vannier et al. 2014; Zacaï et al. 2016, Legg and Pates 2017; Lerosey-Aubril et al. 2017, 2020), the incidence of such structures is significantly higher in the Buen (Budd 1998, 1999; Stein et al. 2013; Vannier et al. 2014; Peel 2017; Strang et al. 2016; Young and Vinther 2017; Nielsen et al. 2019) and Weeks (Lerosey-Aubril 2015; Lerosey-Aubril et al. 2012, 2018) formations. Accordingly, some chemical distinction in the early diagenetic settings of these two sites may help to explain their greater representation of palaeoscolecids.
Assessing the role of ecological competition on the abundance of palaeoscolecids in a given Lagerstätte remains a challenge, as data concerning diversity of the fauna, the relative abundance of its components, and their ecologies are only available for the Burgess Shale Formation. The infauna of the Buen Formation comprises two palaeoscolecid species, a priapulid, and two annelids (Harper et al. 2019), the most common of which is Phragmochaeta (which is represented by specimens numbering roughly twice the whole palaeoscolecid material; Conway Morris and Peel 2008). If this annelid truly was an ecological competitor of the palaeoscolecids, it did not prevent the development of abundant palaeoscolecids in the Buen biota. As for the Weeks biota, it includes a variety of worms that are often too poorly preserved to allow confident assignment. However, the palaeoscolecid and an undescribed annelid (Lerosey-Aubril et al. 2018: fig. 5g, h) are by far the most common of those worms (ca. 50 specimens each). Whether these organisms originally occupied similar ecological niches in the same environment is not known, but if they did, it did not impact their development in the Weeks biota.
In summary, we concur with Smith (2015) that physical characteristics of the depositional environment (e.g., bathymetry, distance from the shore, submarine relief) do not explain the variation of palaeoscolecid abundance observed in the Cambrian Lagerstätten of Laurentia. We instead argue that the preservation of more abundant palaeoscolecid materials in the Buen and Weeks formations might have been facilitated by early diagenetic chemical settings favorable to the preservation and possibly the precipitation of apatite. Ecological competition remains a valid explanation for the limited representation of the group in Laurentian Lagerstätten, but we could not find supporting evidence for this outside the Burgess Shale Formation. Finally, it is likely that collections and research bias (sensu Whitaker and Kimmig 2020) also plays a role in the scarcity of palaeoscolecids from certain Laurentian deposits.
Taphonomy.—Marjum Formation soft-bodied fossils are traditionally regarded as carbonaceous compressions similar to those of the Burgess Shale Formation (Gaines et al. 2005; Foster and Gaines 2016). However, neither KUMIP 314100 nor 314101 provide a distinctive carbon signature in SEM-based analyses (Fig. 6A6, B6). Using EDS, Broce and Schiffbauer (2017) detected an elevated carbon signal associated with a Marjum Formation worm fossil, but observed no discernible carbonaceous film via backscattered or secondary electron imaging. They inferred that the higher carbon concentration observed locally stemmed from accumulation of interstitial carbonaceous material. The absence of detectable carbonaceous films or local enrichments of carbon in the palaeoscolecid fossils might indicate a few possible taphonomic scenarios, involving the complete removal of organic carbon. For example, given their infaunal nature, the organisms plausibly died within the uppermost oxic- to sub-oxic layers of sediment, allowing for efficient decay by oxygenic microbes. If burial occurred, this very likely occurred rapidly after death, for the two specimens preserve distinct annuli boundaries and organized plates. Scavenging was prevented, but not the decay of labile structures (e.g., proboscis, gut). Organic carbon removal/dispersion could have continued through diagenesis as well. While such information may provide insights onto the patterns of palaeoscolecid abundance and diversity in Laurentian Lagerstätten, additional investigation of the diagenetic histories between House Range and other Laurentian fossil localities is required before testing any potential hypotheses.
Conclusions
The description of KUMIP 314100 and 314101 represents the first confident report of the class Palaeoscolecida in the Marjum Formation. Both specimens are fragments of articulated scleritomes, the preservation of which required exceptional depositional circumstances. However, the absence of internal organs (e.g., the gut), the plate-barren body extremities, and the structures borne by these latter (proboscis, caudal hooks) indicates that the most fragile parts of the body were lost during diagenesis, which considerably limits inferences on the palaeoecology of the new palaeoscolecid taxon. The scarcity of palaeoscolecid remains in the Marjum Formation mirrors the biotic compositions observed from most Cambrian Lagerstätten of Laurentia, except for the Buen and Weeks formations. The reasons underlying this palaeogeographically uneven representation of palaeoscolecids are uncertain, but it might suggest that more than a single taphonomic pathway permitted the preservation of macroscopic soft-bodied fossil assemblages in the Cambrian, such that some taxa may have been more or less favourably preserved within specific taphonomic settings.
Authors’ contributions
WL and JK designed the project. WL and AFW photographed material. WL and JDS obtained the SEM and EDS data. WL and RL-A prepared figures with input from all authors. WL, RL-A, JK, and JDS interpreted the data. WL, RL-A, and JK wrote the manuscript with input from all co-authors. All authors performed research, discussed, and approved the final manuscript.
Acknowledgements
We thank Arvid Aase (Kemmerer, Wyoming, USA) and Thomas T. Johnson (Peebles, Ohio, USA) for donating the specimens, Uwe Balthasar (University of Plymouth, UK) for information on the palaeoscolecid occurrence in the Mural Formation, and J. Moore and an anonymous colleague for their thorough and constructive reviews. WWL was supported by a University of Kansas Undergraduate Research Grant, and a student research grant provided by the South-Central Section of the Geological Society of America. JDS is supported by NSF CAREER 1652351, and the University of Missouri X-ray Microanalysis Core by NSF IF 1636643. RL-A is supported by the J. Ortega-Hernández Laboratory for Invertebrate Palaeobiology, Harvard University.
References
Balthasar, U. 2004. Shell structure, ontogeny and affinities of the Lower Cambrian bivalved problematic fossil Mickwitzia muralensis Walcott, 1913. Lethaia 37: 381–400. Crossref
Barskov, I.S. and Zhuravlev, A.Y. [Žuravlev, A.Y.] 1988. Soft-bodied organisms from the Cambrian of the Siberian Platform [in Russian]. Paleontologičeskij žurnal 1988 (1): 3–9.
Bonino, E. and Kier, C. 2010. The Back to the Past Museum Guide to Trilobites. 495 pp. Casa Editrice Marna, Lecco.
Botting, J.P., Muir, L.A., Van Roy, P., Bates, D., and Upton, C., 2012. Diverse Middle Ordovician palaeoscolecidan worms from the Builth-Llandrindod Inlier of central Wales. Palaeontology 55: 501–528. Crossref
Briggs, D.E.G. and Robison, R.A. 1984. Exceptionally preserved nontrilobite arthropods and Anomalocaris from the middle Cambrian of Utah. The University of Kansas Paleontological Contributions 111: 1–24.
Briggs, D.E.G., Lieberman, B.S., Hendricks, J.R., Halgedahl, S.L., and Jarrard, R.D. 2008. Middle Cambrian arthropods from Utah. Journal of Paleontology 82: 238–254. Crossref
Broce, J.S. and Schiffbauer, J.D. 2017. Taphonomic analysis of Cambrian vermiform fossils of Utah and Nevada, and implications for the chemistry of Burgess Shale-type preservation of vermiform fossils. Palaios 32: 600–619. Crossref
Brock, G.A. and Cooper, B.J. 1993. Shelly fossils from the early Cambrian (Toyonian) Wirrealpa, Aroona Creek, and Ramsay Limestones of South Australia. Journal of Paleontology 67: 758–787. Crossref
Budd, G.E. 1998. Stem group arthropods from the lower Cambrian Sirius Passet fauna of North Greenland. In: R.A. Fortey and R.H. Thomas (eds.), Arthropod Relationships. The Systematics Association Special Volume Series, vol. 55, 125–138. Springer, Dordrecht. Crossref
Budd, G.E. 1999. The morphology and phylogenetic significance of Kerygmachela kierkegaardi Budd (Buen Formation, lower Cambrian, N Greenland). Earth and Environmental Science Transactions of The Royal Society of Edinburgh 89: 249–290. Crossref
Butterfield, N.J. 1994. Burgess Shale-type fossils from a lower Cambrian shallow-shelf sequence in northwestern Canada. Nature 369: 477–479. Crossref
Butterfield, N.J. 2002. Leanchoilia guts and the interpretation of three-dimensional structures in Burgess Shale-type fossils. Paleobiology 28: 155–171. Crossref
Butterfield, N.J. 2009. Fossil preservation in the Burgess Shale. In: J.-B. Caron and D.M. Rudkin (eds.), A Burgess Shale Primer: History, Geology, and Research Highlights, Field Trip Companion Volume, ICCE 2009, 63–69. Burgess Shale Consortium, Toronto.
Butterfield, N.J. and Harvey, T.H.P. 2012. Small carbonaceous fossils (SCFs): a new measure of early Paleozoic paleobiology. Geology 40: 71–74. Crossref
Caron, J.-B. 2009. The Greater Phyllopod Bed community, historical variations and quantitative approaches. In: J.-B. Caron and D.M. Rudkin (eds.), A Burgess Shale Primer: History, Geology, and Research Highlights, Field Trip Companion Volume, ICCE 2009, 71–89. Burgess Shale Consortium, Toronto.
Cocks, L.R.M. and Torsvik, T.H. 2011. The Paleozoic geography of Laurentia and western Laurussia: a stable craton with mobile margins. Earth-Science Reviews 106: 1–51. Crossref
Conway Morris, S. 1977. Fossil priapulid worms. Special Papers in Palaeontology 20: 1–95.
Conway Morris, S. 1997. The cuticular structure of the 495-Myr-old type species of the fossil worm Palaeoscolex, P. piscatorum (?Priapulida). Zoological Journal of the Linnean Society 119: 69–82. Crossref
Conway Morris, S. and Peel, J.S. 2008. The earliest annelids: lower Cambrian polychaetes from the Sirius Passet Lagerstätte, Peary Land, North Greenland. Acta Palaeontologica Polonica 53: 137–148. Crossref
Conway Morris, S. and Peel, J.S. 2010. New palaeoscolecidan worms from the lower Cambrian: Sirius Passet, Latham Shale and Kinzers Shale. Acta Palaeontologica Polonica 55: 141–156. Crossref
Conway Morris, S. and Robison, R.A. 1986. Middle Cambrian priapulids and other soft-bodied fossils from Utah and Spain. The University of Kansas Paleontological Contributions 117: 1–22.
Duan, B. and Dong, X. 2013. Furongian (late Cambrian) palaeoscolecid cuticles from Hunan Province, South China: the growth impact on the worm cuticle. Acta Scientiarum Universitatis Pekinensis 49: 591–602.
Duan, B., Dong, X.-P., and Donoghue, P.C.J. 2012. New palaeoscolecid worms from the Furongian (upper Cambrian) of Hunan, South China: is Markuelia an embryonic palaeoscolecid? Palaeontology 55: 613–622. Crossref
Foster, J.R. and Gaines, R.R. 2016. Taphonomy and paleoecology of the “middle” Cambrian (Series 3) formations in Utah’s West Desert: recent finds and new data. Utah Geological Association Publication 45: 291–336.
Gaines, R.R. 2014. Burgess Shale-type preservation and its distribution in space and time. The Paleontological Society Papers 20: 123–146. Crossref
Gaines, R.R., Kennedy, M.J., and Droser, M.L. 2005. A new hypothesis for organic preservation of Burgess Shale taxa in the middle Cambrian Wheeler Formation, House Range, Utah. Palaeogeography, Palaeoclimatology, Palaeoecology 220: 193–205. Crossref
García-Bellido, D.C., Paterson, J.R., and Edgecombe, G.D. 2013. Cambrian palaeoscolecids (Cycloneuralia) from Gondwana and reappraisal of species assigned to Palaeoscolex. Gondwana Research 24: 780–795. Crossref
Gedik, I. 1977. Orta Toroslar’da konodont biyostratigrafisi. Türkiye Jeoloji Kurumu Bülteni 20: 35–48.
Glaessner, M.F. 1979. Lower Cambrian Crustacea and annelid worms from Kangaroo Island, South Australia. Alcheringa 3: 21–31. Crossref
Gutiérrez-Marco, J.C. and García-Bellido, D.C. 2015. Micrometric detail in palaeoscolecid worms from Late Ordovician sandstones of the Tafilalt Konservat-Lagerstätte, Morocco. Gondwana Research 28: 875–881. Crossref
Han, J., Liu, J., Zhang, Z., Zhang, X., and Shu, D. 2007a. Trunk ornament on the palaeoscolecid worms Cricocosmia and Tabelliscolex from the early Cambrian Chengjiang deposits of China. Acta Palaeontologica Polonica 52: 423–431.
Han, J., Yao, Y., Zhang, Z., Liu, J., and Shu, D. 2007b. New observations on the palaeoscolecid worm Tylotites petiolaris from the Cambrian Chengjiang Lagerstätte, south China. Paleontological Research 11: 59–69. Crossref
Han, J., Zhang, X.-L., Zhang, Z.-F., and Shu, D.-G. 2003a. A new platy-armored worm from the early Cambrian Chengjiang Lagerstätte, South China. Acta Geologica Sinica (English Edition) 77: 1–6.
Han, J., Zhang, X.-L., Zhang, Z.-F., and Shu, D.-G. 2003b. Discovery of the proboscis on Tylotites petiolaris. Journal of Northwestern Geology (Natural Science Edition) 36: 87–91.
Harper, D.A.T., Hammarlund, E.U., Topper, T.P., Nielsen, A.T., Rasmussen, J.A., Park, T.Y.S., and Smith, M.P. 2019. The Sirius Passet Lagerstätte of North Greenland: a remote window on the Cambrian explosion. Journal of the Geological Society 176: 1023–1037. Crossref
Harvey, T.H.P. and Butterfield, N.J. 2011. Great Canadian Lagerstätten 2. Macro- and microfossils of the Mount Cap Formation (early and middle Cambrian, Northwest Territories). Geoscience Canada 38: 165–173.
Harvey, T.H.P., Dong, X., and Donoghue, P.C.J. 2010. Are palaeoscolecids ancestral ecdysozoans? Evolution and Development 12: 177–200. Crossref
Hintze, L.F. 1980. Geologic Map of Utah, Scale 1:500,000. Utah Geological and Mineral Survey, Salt Lake City.
Hou, X.-G. and Sun, W.-G. 1988. Discovery of Chengjiang fauna at Meishucun, Jinning, Yunnan [in Chinese, with English abstract]. Acta Palaeontologica Sinica 27: 1–12.
Hu, S.X., Li, Y., Luo, H.L., Fu, X.P., You, T., Pang, J.Y., Liu, Q., and Steiner, M. 2008. New record of palaeoscolecids from the early Cambrian of Yunnan, China. Acta Geologica Sinica (English Edition) 82: 244–248. Crossref
Hu, S.X., Steiner, M., Zhu, M., Luo, H., Forchielli, A., Keupp, H., Zhao, F., and Liu, Q. 2012. A new priapulid assemblage from the early Cambrian Guanshan fossil Lagerstätte of SW China. Bulletin of Geosciences 87: 93–106. Crossref
Ivantsov, A.Y. and Wrona, R. 2004. Articulated palaeoscolecid sclerite arrays from the lower Cambrian of eastern Siberia. Acta Geologica Polonica 54: 1–22.
Ivantsov, A.Y. [Ivancov, A.Y.] and Zhuravlev, A.Y. [Žuravlev, A.Y.] 2005. Cephalorhynchs [in Russian]. In: A.G. Pomarenko (ed.), Unikal’nye sinskie mestonahoždeniâ rannekembrijskih organizmov. Trudy Paleontologičeskogo Instituta 2005: 61–72.
Johnston, K.J., Johnston, P.A., and Powell, W.G. 2009. A new, middle Cambrian, Burgess Shale-type biota, Bolaspidella Zone, Chancellor Basin, southeastern British Columbia. Palaeogeography, Palaeoclimatology, Palaeoecology 277: 106–126. Crossref
Kimmig, J. 2021. Burgess Shale fauna. In: S. Elias and D. Alderton (eds.), Encyclopedia of Geology, 2nd ed., 576–582. Elsevier, Oxford, Crossref
Kimmig, J., Leibach, W.W., and Lieberman, B.S. 2020. First occurrence of the problematic vetulicolian Skeemella clavula in the Cambrian Marjum Formation of Utah, USA. Carnets de Géologie 20: 215–221. Crossref
Kouraiss, K., El Hariri, K., El Albani, A., Azizi, A., Mazurier, A., and Vannier, J. 2018. X-ray microtomography applied to fossils preserved in compression: palaeoscolescid worms from the Lower Ordovician Fezouata Shale. Palaeogeography, Palaeoclimatology, Palaeoecology 508: 48–58. Crossref
Kraft, P. and Mergl, M. 1989. Worm-like fossils (Palaeoscolecida; ?Chaetognatha) from the Lower Ordovician of Bohemia. Sborník geologických vĕd Paleontologie 30, 9–36.
Legg, D.A. and Pates, S. 2017. A restudy of Utahcaris orion (Euarthropoda) from the Spence Shale (middle Cambrian, Utah, USA). Geological Magazine 154: 181–186. Crossref
Lemburg, C. 1995. Ultrastructure of sense organs and receptor cells of the neck and lorica of the Halicryptus spinulosus larva (Priapulida). Microfauna Marina 10: 7–30.
Lerosey-Aubril, R. 2015. Notchia weugi gen. et sp. nov.: a new short-headed arthropod from the Weeks Formation Konservat-Lagerstätte (Cambrian; Utah). Geological Magazine 152: 351–357. Crossref
Lerosey-Aubril, R., Gaines, R.R., Hegna, T.A., Ortega-Hernández, J., Van Roy, P., Kier, C., and Bonino, E. 2018. The Weeks Formation Konservat-Lagerstätte and the evolutionary transition of Cambrian marine life. Journal of the Geological Society 175: 705–715. Crossref
Lerosey-Aubril, R., Hegna, T.A., Kier, C., Bonino, E., Habersetzer, J., and Carré, M. 2012. Controls on gut phosphatisation: the trilobites from the Weeks Formation Lagerstätte (Cambrian; Utah). PLoS One 7: e32934. Crossref
Lerosey-Aubril, R., Kimmig, J., Pates, S., Skabelund, J., Weug, A., and Ortega-Hernández, J. 2020. New exceptionally preserved panarthropods from the Drumian Wheeler Konservat-Lagerstätte of the House Range Utah. Papers in Palaeontology 6: 501–531. Crossref
Lerosey-Aubril, R., Paterson, J.R., Gibb, S., and Chatterton, B.D. 2017. Exceptionally-preserved late Cambrian fossils from the McKay Group (British Columbia, Canada) and the evolution of tagmosis in aglaspidid arthropods. Gondwana Research 42: 264–279. Crossref
Lin, T. 1995. Discovery of late early Cambrian worm from Huainan, Anhui. Acta Palaeontologica Sinica 34: 504–508.
Liu, J., Han, J., Li, J., Wu, Y., Peng, J., Qi, N., Yang, Y., and Li, J. 2016. New localities and palaeoscolecid worms from the Cambrian (Stage 4, Series 2) Guanshan Biota in Kunming, Yunnan, South China. Acta Geologica Sinica (English Edition) 90: 1939–1945. Crossref
Liu, Y., Wang, Q., Shao, T., Zhang, H., Qin, J., Chen, L., Liang, Y., Chen, C., Xue, J., and Liu, X. 2018. New material of three-dimensionally phosphatized and microscopic cycloneuralians from the Cambrian Paibian stage of South China. Journal of Paleontology 92: 87–98. Crossref
LoDuca, S.T., Brett, C.E., and Baird, G.C. 1997. The Medusaegraptus epibole and lower Ludlovian Konservat-Lagerstätten of eastern North America. In: C.E. Brett (ed.), Paleontological Events: Stratigraphic, Ecological, and Evolutionary Implications, 369–405. Columbia University Press, New York.
Luo, H., Hu, S., Chen, L., Zhang, S., and Tao, Y. 1999. Early Cambrian Chengjiang Fauna >From Kunming Region, China. 162 pp. Yunnan Science and Technology Press, Kunming.
Martin, E.L.O., Lerosey-Aubril, R., and Vannier, J. 2016. Palaeoscolecid worms from the Lower Ordovician Fezouata Lagerstätte, Morocco: Palaeoecological and palaeogeographical implications. Palaeogeography, Palaeoclimatology, Palaeoecology 460: 130–141. Crossref
Muir, L.A., Ng, T.-W., Li, X.-F., Zhang, Y.-D., and Lin, J.-P. 2014. Palaeoscolecidan worms and a possible nematode from the Early Ordovician of South China. Palaeoworld 23: 15–24. Crossref
Müller, K.J. 1973. Milaculum n.g., ein phosphatisches Mikrofossil aus dem Altpalaeozoikum. Paläontologische Zeitschrift 47: 217–228. Crossref
Müller, K.J. and Hinz-Schallreuter, I. 1993. Palaeoscolecid worms from the middle Cambrian of Australia. Palaeontology 36: 549–592.
Müller, K.J. and Miller, J.F. 1976. The problematic microfossil Utahphospha from the upper Cambrian of the western United States. Lethaia 9: 391–395. Crossref
Nielsen, M., Lee, M., Wilby, P., Park, T.-Y., Ng, H.C., Hendry, K., Nielsen, A., and Vinther, J. 2019. Selective transformation and ontogenetic biases of phosphatised soft tissues in the Sirius Passet Lagerstätte, North Greenland. PaleoBios 36 (Supplement 1): 265–266.
Pates, S., Daley, A.C., and Lieberman, B.S. 2018. Hurdiid radiodontans from the middle Cambrian (Series 3) of Utah. Journal of Paleontology 92: 99–113. Crossref
Pates, S., Lerosey-Aubril, R., Daley, A.C., Kier, C., Bonino, E., and Ortega-Hernández, J. 2021. Ecology and affinities of the diverse radiodont fauna from the Marjum Formation of Utah, USA (Cambrian, Drumian). PeerJ 9: e10509. Crossref
Peel, J.S. 2017. Mineralized gutfills from the Sirius Passet Lagerstätte (Cambrian Series 2) of North Greenland. GFF 139: 83–91. Crossref
Peng, J., Huang, D., Zhao, Y., and Sun, H. 2016. Palaeoscolecids from the Balang Fauna of the Qiandongian (Cambrian Series 2), Guizhou, China. Geological Magazine 153: 438–448. Crossref
Peng, S.C., Yang, X.F., Liu, Y., Zhu, X.J., Sun, H.J., Zamora, S., Mao, Y.C., and Zhang, Y.C. 2020. Fulu biota, a new exceptionally-preserved Cambrian fossil assemblage from the Longha Formation in southeastern Yunnan. Palaeoworld 29: 453–461. Crossref
Pope, M.C. and Sears, J.W. 1997. Cassiar platform, north-central British Columbia: a miogeoclinal fragment from Idaho. Geology 25: 515–518. Crossref
Rees, M.N. 1986. A fault-controlled trough through a carbonate platform: the middle Cambrian House Range Embayment. Geological Society of America Bulletin 97: 1054–1069. Crossref
Rigby, J.K. 1983. Sponges of the middle Cambrian Marjum Limestone from the House Range and Drum Mountains of western Millard County, Utah. Journal of Paleontology 57: 240–270.
Robison, R.A. 1969. Annelids from the middle Cambrian Spence Shale of Utah. Journal of Paleontology 43: 1169–1173.
Robison, R.A. 1984. New occurrences of the unusual trilobite Naraoia from the Cambrian of Idaho and Utah. University of Kansas Paleontological Contributions 112: 1–8.
Robison, R.A. 1991. Middle Cambrian biotic diversity: examples from four Utah Lagerstätten. In: A. Simonetta and S. Conway Morris (eds.), The Early Evolution of Metazoa and the Significance of Problematic Taxa, 77–98. Cambridge University Press, Cambridge.
Robison, R.A. and Babcock, L.E. 2011. Systematics, paleobiology, and taphonomy of some exceptionally preserved trilobites from Cambrian Lagerstätten of Utah. Kansas University Paleontological Contributions 5: 1–47.
Robison, R.A., Babcock, L.E., and Gunther, V.G. 2015. Exceptional Cambrian fossils from Utah: A window into the age of trilobites. Utah Geological Survey, Miscellaneous Publications 15: 1–97.
Slater, B.J., Harvey, T.H., Guilbaud, R., and Butterfield, N.J. 2017. A cryptic record of Burgess Shale-type diversity from the early Cambrian of Baltica. Palaeontology 60: 117–140. Crossref
Smith, D.D. 2007. Sequence Stratigraphy of the Middle Cambrian Marjum Formation: Response of Sedimentary Facies and Biota to Sea-level Changes. 156 pp. Unpublished M.Sc. Thesis, Utah State University, Logan.
Smith, M.R. 2015. A palaeoscolecid worm from the Burgess Shale. Palaeontology 58: 973–979. Crossref
Smith, M.R., Harvey, T.H.P., and Butterfield, N.J. 2015. The macro- and microfossil record of the priapulid Ottoia. Palaeontology 58: 705–721. Crossref
Sperling, E.A., Balthasar, U., and Skovsted, C.B. 2018. On the edge of exceptional preservation: insights into the role of redox state in Burgess Shale-type taphonomic windows from the Mural Formation, Alberta, Canada. Emerging Topics in Life Sciences 2: 311–323. Crossref
Stein, M., Budd, G.E., Peel, J.S., and Harper, D.A.T. 2013. Arthroaspis n. gen., a common element of the Sirius Passet Lagerstätte (Cambrian, North Greenland), sheds light on trilobite ancestry. BMC Evolutionary Biology 13: 99. Crossref
Steiner, M., Hu, S. X., Liu, J., and Keupp, H. 2012. A new species of Hallucigenia from the Cambrian Stage 4 Wulongqing Formation of Yunnan (South China) and the structure of sclerites in lobopodians. Bulletin of Geosciences 87: 107–124. Crossref
Strang, K.M., Armstrong, H.A., and Harper, D.A.T. 2016. Minerals in the gut: scoping a Cambrian digestive system. Royal Society Open Science 3: 160420. Crossref
Streng, M., Ebbestad, J.O.R., and Berg‐Madsen, V. 2017. Cambrian palaeoscolecids (Cycloneuralia) of southern Scandinavia. Papers in Palaeontology 3: 21–48. Crossref
Sun, W.-G. and Hou, X.-G. 1987. Early Cambrian worm from Chengjiang, Yunnan, China [in Chinese, with English abstract]. Acta Palaeontologica Sinica 26: 257–271.
Topper, T.P., Brock, G.A., Skovsted, C.B., and Paterson, J.R. 2010. Palaeoscolecid scleritome fragments with Hadimopanella sclerites from the early Cambrian of South Australia. Geological Magazine 147: 86–97. Crossref
Vannier, J. and Martin, E.L. 2017. Worm-lobopodian assemblages from the early Cambrian Chengjiang biota: insight into the “pre-arthropodan ecology”? Palaeogeography, Palaeoclimatology, Palaeoecology 468: 373–387. Crossref
Vannier, J., Liu, J., Lerosey-Aubril, R., Vinther, J., and Daley, A.C. 2014. Sophisticated digestive systems in early arthropods. Nature Communications 5: 1–9. Crossref
Walcott, C.D. 1911. Cambrian geology and paleontology II. Middle Cambrian annelids. Smithsonian Miscellaneous Collections 57: 109–144.
Wang, W., Muir, L. A., Botting, J. P., Feng, H., Servais, T., and Li, L. 2014. A Tremadocian (Early Ordovician) palaeoscolecidan worm from graptolitic shales in Hunan Province, South China. Palaeontology 57: 657–671. Crossref
Wendruff, A.J., Babcock, L.E., Kluessendorf, J., and Mikulic, D.G. 2020. Paleobiology and taphonomy of exceptionally preserved organisms from the Waukesha Biota (Silurian), Wisconsin, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 546: 109631. Crossref
Whitaker, A.F. and Kimmig, J. 2020. Anthropologically introduced biases in natural history collections, with a case study on the invertebrate paleontology collections from the middle Cambrian Spence Shale Lagerstätte. Palaeontologia Electronica 23: a58. Crossref
Whitaker, A.F., Jamison, P.G., Schiffbauer, J.D.,
and Kimmig, J. 2020. Re-description of the Spence Shale palaeoscolecids
in light of new morphological features with comments on palaeoscolecid
taxonomy and taphonomy. Paläontologische
Zeitschrift 94: 661–674. Crossref
Whittard, W.F. 1953. Palaeoscolex piscatorum gen. et sp. nov., a worm from the Tremadocian of Shropshire. Quarterly Journal of the Geological Society 109: 125–135. Crossref
Yang, Y. and Zhang, X. 2016a. Distinctive scleritome with marginal tubercles of a new palaeoscolecid worm from the Shipai fauna (Cambrian Epoch 2) at Three Gorges, South China. Acta Geologica Sinica (English Edition) 90: 807–817. Crossref
Yang, Y. and Zhang, X. 2016b. The Cambrian palaeoscolecid Wronascolex from the Shipai fauna (Cambrian Series 2, Stage 4) of the three gorges area, South China. Papers in Palaeontology 2: 555–568. Crossref
Yang, Y., Zhang, X., Zhao, Y., Qi, Y., and Cui, L. 2018. New paleoscolecid worms from the early Cambrian north margin of the Yangtze Platform, South China. Journal of Paleontology 92: 49–58. Crossref
Yang, Y., Zhao, Y., and Zhang, X. 2016. Fossil priapulid Ottoia from the Kaili biota (Cambrian Series 3) of South China. Journal of Systematic Palaeontology 14: 527–543. Crossref
Young, F.J. and Vinther, J. 2017. Onychophoran‐like myoanatomy of the Cambrian gilled lobopodian Pambdelurion whittingtoni. Palaeontology 60: 27–54. Crossref
Zacaï, A., Vannier, J., and Lerosey-Aubril, R. 2016. Reconstructing the diet of a 505-million-year-old arthropod: Sidneyia inexpectans from the Burgess Shale fauna. Arthropod Structure and Development 45: 200–220. Crossref
Zhang, X.-G. and Pratt, B.R. 1996. Early Cambrian palaeoscolecid cuticles from Shaanxi, China. Journal of Paleontology 70: 275–279. Crossref
Zhu, X., Peng, S., Zamora, S., Lefebvre, B., and Chen, G. 2016. Furongian (upper Cambrian) Guole Konservat-Lagerstätte from South China. Acta Geologica Sinica 90: 30–37. Crossref
Acta Palaeontol. Pol. 66 (3): 663–678, 2021
https://doi.org/10.4202/app.00875.2021