
The oldest birotule-bearing freshwater sponges from the Upper Cretaceous–lower Paleocene Deccan volcanic-associated sediments of India
BANDANA SAMANT, ROBERTO PRONZATO, DHANANJAY MAHENDRAKUMAR MOHABEY, TIZIANA CUBEDDU, GIACINTA ANGELA STOCCHINO, KRUTIKA JANGALE, PRANAY THALAL, ANUP DHOBALE, and RENATA MANCONI
Samant, B., Pronzato, R., Mohabey, D.M., Cubeddu, T., Stocchino, G.A., Jangale, K., Thalal, P., Dhobale, A., and Manconi, R. 2023. The oldest birotule-bearing freshwater sponges from the Upper Cretaceous–lower Paleocene Deccan volcanic-associated sediments of India. Acta Palaeontologica Polonica 68 (1): 167–174.
A new fossil occurrence of freshwater sponges (Porifera: Demospongiae) is reported from the Deccan volcanic associated Naskal intertrappean locality, deposited in an interval of <100 kyr across the Cretaceous/Paleogene (K/Pg) boundary. This is the oldest record of siliceous fossil birotule spicules (gemmuloscleres) belonging to asexual resting stages typical of the order Spongillida. The analysis supports the ascription of these fossils to the family Palaeospongillidae. The diagnosis and description of Longibirotula Pronzato and Manconi gen. nov. and its type species Longibirotula antiqua Manconi and Samant sp. nov. from the Naskal intertrappean is based on skeletal and gemmular spicular morphotraits. The findings have provided evidence of the presence of diversified groups of freshwater sponges during the Late Cretaceous on the Indian subcontinent and Gondwanaland. From the biogeographic context, the findings track the evolutionary trends of the oldest continental sponges in the Asian and Australasian/Insular Pacific regions.
Key words: Porifera, Palaeospongillidae, inland water sponges, conservative morphotraits, gemmules, siliceous skeleton, palaeobiodiversity, palaeogeography, Cretaceous, Paleocene, Deccan traps.
Bandana Samant [bandanabhu@gmail.com, ORCID: https://orcid.org/0000-0001-8257-452 ], Dhananjay Mahendrakumar Mohabey [dinomohabey@yahoo.com; ORCID: https://orcid.org.0000-0003-4538-6907 ], Krutika Jangale [krutismile1999@gmail.com; ORCID: https://orcid.org/0000-0002-1452-3545 ],
Pranay Thalal [ppt52525.pt.pt@gmail.com; ORCID: https://orcid.org/0000-0003-3768-1519 ] and Anup Dhobale [anupdhobale@gmail.com; ORCID: https://orcid.org/0000-0001-9496-0069 ], Postgraduate Department of Geology, RTM Nagpur University, Nagpur, Maharashtra 440001, India.
Roberto Pronzato [roberto.pronzato@unige.it; ORCID: https://orcid.org/0000-0003-1355-0936 ], Dipartimento di Scienze della Terra, dell’Ambiente e della Vita (DISTAV), Università di Genova, Genova, Italy.
Renata Manconi [rmanconi@uniss.it; ORCID: https://orcid.org/0000-0002-7619-8493 ], Tiziana Cubeddu [tcubeddu @uniss.it; ORCID: https://orcid.org/0000-0001-7522-8200 ], and Giacinta Angela Stocchino [stocchin@uniss.it; ORCID: https://orcid.org/0000-0002-7005-208X ], Dipartimento di Medicina Veterinaria, Laboratorio Zoologia, Università di Sassari, Sassari, Italy.
Received 30 October 2022, accepted 25 January 2023, available online 8 March 2023.
Copyright © 2023 B. Samant et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The fossil freshwater sponges (Porifera: Demospongiae), known from both Laurasia and Gondwana, belong to the order Spongillida Manconi and Pronzato, 2002, which comprises three families with fossil taxa, i.e., Palaeospongillidae Volkmer-Ribeiro and Reitner, 1991, Potamolepidae Brien, 1967, and Spongillidae Gray, 1867. The oldest Spongillida fossil records (only spicules) dates back to the Permo-Carboniferous (Schindler et al. 2008), and the Late Jurassic (Spongilla purbeckensis Hinde, 1883). Unfortunately, fossil records of inland water sponges are very scarce, due to their fragile skeletal siliceous architecture (Pronzato et al. 2017).
Most fossil Spongillida are hard to identify to the family, genus, or species level because they lack gemmuloscleres, which are one of the most important diagnostic morphotraits for systematics and phylogeny. Till date, the oldest known entire gemmule is of Palaeospongilla chubutensis Volkmer-Ribeiro and Reitner, 1991 (Palaeospongillidae) from the Lower Cretaceous of Patagonia (Volkmer-Ribeiro and Reitner 1991).
The gemmuloscleres remains of other fossil species of the families Spongillidae and Potamolepidae share similar morphotraits with living genera, e.g., Anheteromeyenia Schröder, 1927; Corvospongilla Annandale, 1911; Ephydatia Lamouroux, 1816; Eunapius Gray, 1867; Radiospongilla Penney and Racek, 1968; Spongilla Linnaeus, 1759; Trochospongilla Vejdovsky, 1883; Oncosclera Volkmer-Ribeiro, 1970, and Potamophloios Brien, 1970 (Pisera 2006; Pisera et al. 2013, 2016; Pronzato et al. 2017).
From the Deccan volcanic province, some unidentified spicules of freshwater sponges have been recorded from the Deccan infra (Lameta Formation) and intertrappean sediments of central India and the intertrappean beds of northwestern India (Samant and Mohabey 2009, 2014; Samant et al. 2014), but definite record of fossil freshwater sponges of Spongillida is scarce. Recently, the new genus Palaeocorvospongilla Pronzato and Manconi, 2021, of the family Palaeospongillida was described from the Deccan intertrappean sediments of India (Maastrichtian, Upper Cretaceous) with the new species Palaeocorvospongilla cretacea Manconi and Samant, 2021 (Samant et al. 2021).
The present study records the oldest fossil birotule-bearing freshwater sponge (Porifera: Demospongiae) of a new genus and species of Palaeospongillidae from the Naskal intertrappean deposit in the south-eastern part of the Deccan volcanic province.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:7B9F49CE-E5E9-46CD-BC9A-17C204352BC1.
Institutional abbreviations.—PGDG, Museum Postgraduate Department of Geology, Nagpur University, India; PGNU, Postgraduate Department of Geology, Nagpur University, India.
Other abbreviations.—NSKQ, Naskal Quarry section.
Geological setting
The Upper Cretaceous–lower Paleocene Deccan volcanic-associated sediment covers an area of ~500,000 km2 in the south, western and central parts of India. The Deccan volcanic flows are associated with infratrappean (deposited below the volcanic flows) and intertrappean sediments (deposited in-between the two volcanic flows). Vertebrates, invertebrates, micro and megaflora have been discovered in both infratrappean and intertrappean sediments (Mohabey 1996; Mohabey and Udhoji 2000; Khosla and Sahni 2003). The Naskal section is located at 17°14’21” N, 77°53’16” E, in the Ranga Reddy District in the state of Telangana, India (Fig. 1A). This section has a thickness of <3 m and a lateral exposure of <15 m and an aerial distribution of <5 m2. It occurs between Flow-3 and Flow-4 at an elevation of 624 m. In the Naskal locality there are three geographically separated sections designated as Naskal-A, Naskal-B, and Naskal GSI Quarry (Wilson Mantilla et al. 2022: fig. 3). Based on Ar40/Ar39 plagioclase dating of the flows, it is indicated that the permissible age range of the Naskal intertrappean sediments is between 66.136 and 66.056 Ma at 68% confidence (Wilson Mantilla et al. 2022), thus deposited close to the Cretaceous/Paleogene boundary.
All the three Naskal sections were targeted for the sampling but the sponge spicules were recovered only from the Naskal GSI Quarry section (Fig. 1). The sponge spicule bearing section has a thickness of 120 cm comprises black to grey cherty limestone, hard yellowish shaly mudstone, loose shaly to carbonate mudstone to marlstone, white mudstone, and dark clay with sandy lenses in ascending stratigraphic order (Fig. 1B). Naskal B (Fig. 1C) which is ~7 m west of the Naskal GSI Quarry section yielded only palynomorphs (for details see Wilson Mantilla et al. 2022).
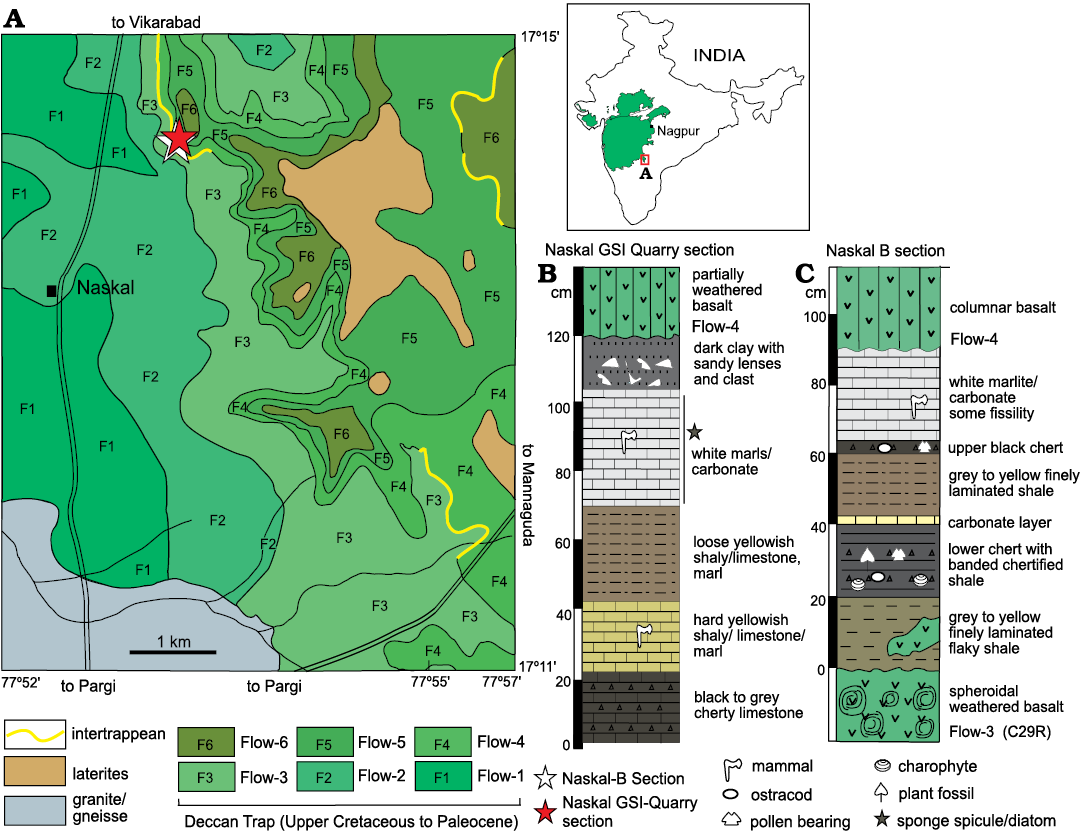
Fig. 1. Map of India showing Deccan volcanic province (green area). A. Location of Naskal intertrappean, Naskal B (white star) and Naskal GSI Quarry sections (red star); map modified after Ahluwalia (1990) and Wilson Mantilla et al. (2022). B. Sponge spicule and diatom bearing horizon in Naskal GSI Quarry section. C. Palynomorph bearing Naskal B section (modified after Wilson Mantilla et al. 2022).
Palaeoecology and age of the Naskal Intertrappean.—The Naskal intertrappean beds have yielded a rich mammal record (summarised in Wilson Mantilla et al. 2022), including the first Indian record of a Cretaceous mammal (Prasad and Sahni 1988). In addition to mammals, it has other vertebrates such as fish, anurans, squamates, sphenodontian, turtles, and crocodilians (Prasad and Sahni 1988; Prasad 2012). Palynoflora of Naskal is represented by marker Maastrichtian taxa, i.e., Crybelosporites intertrappea, Maastrichtian–Paleocene taxa, i.e., Gabonisporis vigourouxii, Mulleripollis bolpurensis, and Paleocene taxa, i.e., Striacolporites striatus and Echistephanocolpites meghalayensis. Overall, palynoassemblage indicates the presence of transitional flora where Maastrichtian palynomorphs were depleted and Paleocene flora was dominating.
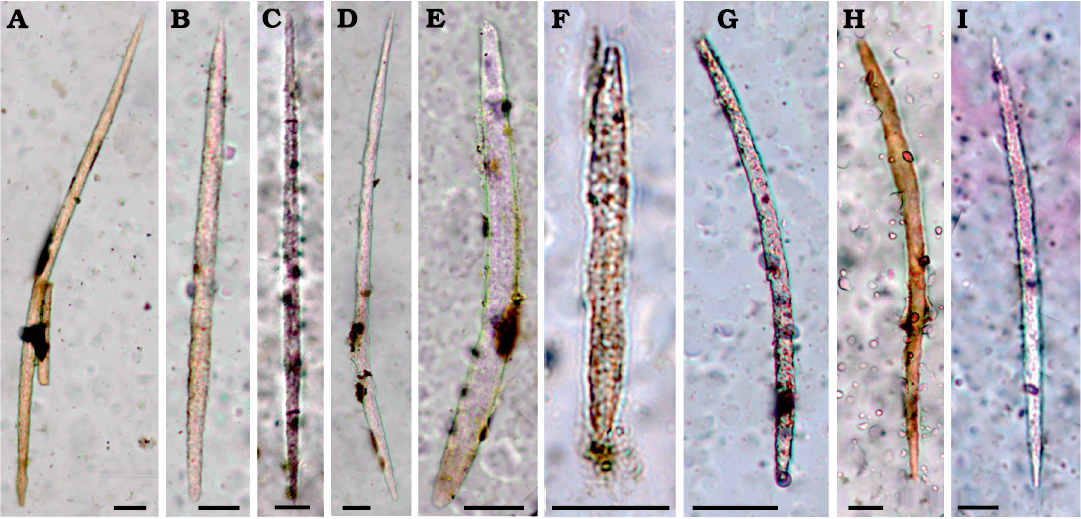
Fig. 2. Megascleres of palaeospongillid sponge Longibirotula antiqua gen. et sp. nov. from Upper Cretaceous–lower Paleocene of Naskal GSI Quarry (India). A–I. Oxeas (slides PGNU/NSKQ/SL-1–13) slim to stout with variably pointed tips. Diagenetic processes affect all spicules to various degree. Scale bars 20 µm. {fig. will be greyscale in printed version}
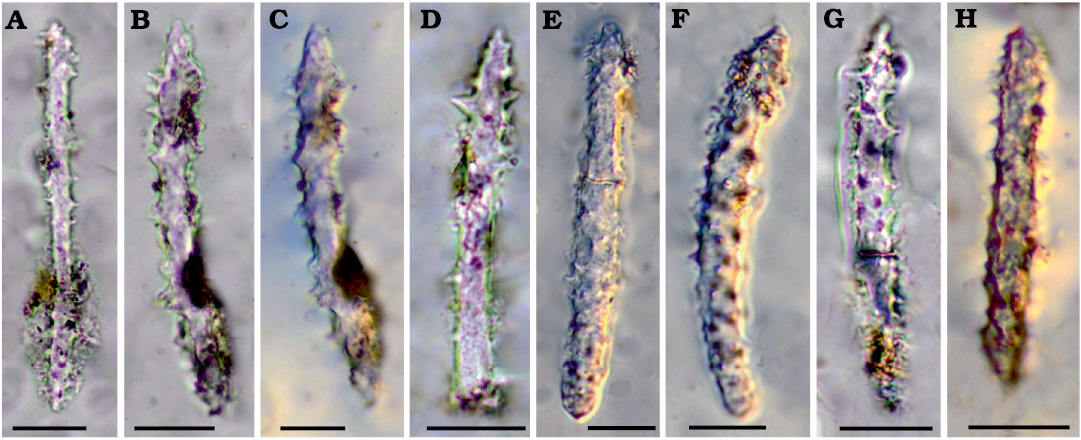
Fig. 3. Megascleres of palaeospongillid sponge Longibirotula antiqua gen. et sp. nov. from Upper Cretaceous–lower of Paleocene of Naskal GSI Quarry (India). A–H. Acanthoxeas (slides PGNU/NSKQ/SL-1–13) with large spines. Diagenetic processes affect all spicules to various degree. Scale bars 20 µm.
Material and methods
Samples of shale, carbonate mudstone, and marlstone (100 g each) were treated with 5% hydrochloric acid (HCl) until the effervescence subsided. This was followed by thorough washing with distilled water and treatment with 5% hydrofluoric acid (HF) for 5 minutes to remove the thin siliceous secondary coating on the fossils. To remove organic matter, samples were sometimes treated with dilute nitric acid (HNO3) and 5% potassium hydroxide (KOH). Every chemical treatment was followed by washing the sample with distilled water. After chemical treatments, the sieved samples were divided into two aliquots, one for the preparation of slides for Light Microscopy (LM) using polyvinyl alcohol and Canada balsam, and the other for the preparation of stubs for Scanning Electron Microscopy (SEM).
All the slides were studied under a transmitted light Olympus BX51 (Japan) microscope, and the photographs were taken with a DP 20 (Olympus, Japan) camera. A total of 25 spicules from each category were studied and measured under the LM microscope. For SEM studies, the sponge spicules bearing sample was spread over a glass slide and the slide was scanned under a microscope using a 10X objective. The spicules were picked up with the help of hair attached to the dissecting needle and placed over the SEM stub. Later, the stub was coated with gold/palladium, and SEM observations were carried out under the Jeol microscope (Japan) at the Jawaharlal Nehru Aluminium Research Development and Design Centre (JNARDDC), Nagpur, India.
The holotype and paratype material, including the rock samples, studied slides and SEM stubs, are housed in the Museum of the Postgraduate Department of Geology (PGDG), RTM Nagpur University, Nagpur, India. The acronym followed by sample number of the (a) holotype rock sample is PGDG/NSKQ/2019; (b) slides (SL) followed by numbers are PGNU/NSKQ/SL-1 to 13; (c) SEM stubs (ST) followed by numbers are PGNU/NSKQ/ST-1, 2.
Terminology of morphotraits follows Manconi and Pronzato (2002). The systematic status of taxa was checked in the World Porifera Database (Voogd et al. 2023).
Systematic palaeontology
Phylum Porifera Grant, 1836
Class Demospongiae Sollas, 1885
Subclass Heteroscleromorpha Cárdenas, Perez, and Boury-Esnault, 2012
Order Spongillida Manconi and Pronzato, 2002
Family Palaeospongillidae Volkmer-Ribeiro and Reitner, 1991
Genus Longibirotula Pronzato and Manconi nov.
Zoobank LSID: urn:lsid:zoobank.org:act:9A389B3D-118E-42AA-AD 9E-1AF484CE7681
Type species: Longibirotula antiqua Manconi and Samant, by monotypy.
Etymology: In reference to the long shaft of birotules gemmuloscleres, gender feminine.
Diagnosis.—Longibirotula is characterised by gemmuloscleres slender birotules with very long spiny shaft, and two types of skeletal megascleres, i.e., long slim oxeas and short acanthoxeas.
Longibirotula antiqua Manconi and Samant sp. nov.
Figs. 1–5.
Zoobank LSID: urn:lsid:zoobank.org:act:A55B4EB4-F32C-41D0-B20 C-C1609F643060
Etymology: From Latin antiqua, old.
Type material: Holotype, rock sample PGDG/NSKQ/2019, slides PGNU/NSKQ/SL-1–13, stubs PGNU/NSK/ST-1, 2.
Type locality: Naskal GSI Quarry, 17°14’21” N, 77°53’16” E, Ranga Reddy District, Telangana State, India.
Type horizon: White marl/carbonate layers of Naskal GSI Quarry, Naskal intertrappean beds (Upper Cretaceous–lower Paleocene).
Diagnosis.—Longibirotula antiqua sp. nov. is characterized by slender gemmuloscleres birotules with long spiny shaft, straight to slightly curved, with large scattered spines variably dense and numerous. Rotules flat with indented margins. Skeletal megascleres of two types, slim oxeas microspiny to smooth and short, stout acanthoxeas.
Description.—Skeletal megascleres monaxial with two morphotypes. Slim, long, microspiny to smooth oxeas (142–425 × 6–13 µm) rarely sinuous. Shorter, stouter acanthoxeas (71–104 × 6–9 µm) straight to slightly curve with dense, large spines. Microscleres absent. Entire gemmules are not found because organic matter associated with gemmular theca is not preserved. Fossil remains are only represented by siliceous spicules. Gemmuloscleres birotules with very long, straight to slightly curved, spiny shaft (47–76 µm in length, 3–6 µm in thickness), with spines large, numerous, variably dense, and rotules flat with indented margins (12–19 µm in diameter).
Remarks.—Spicule deposits suggest that the Naskal intertrappean palaeolake was inhabited by a population of sponges with resting stages at the time of deposition. In addition, centric diatoms (Aulacoseira spp.) and aquatic to semiaquatic flora were also part of the biotic community.
Stratigraphic and geographic range.—Type locality and horizon only.
Discussion
The association of centric diatoms Aulacoseira sp. with sponge spicules in the uppermost part of the intertrappean indicates the development of eutrophic conditions in the lake, likely due to volcanogenic input in the palaeolake. The presence of resting stages in the sponge remains (gemmuloscleres) suggests possible seasonal variations of water level.
Gemmulation processes, synchronised with the local long-term seasonal rhythm, represent the most successful evolutionary strategy in the life cycle of sponges to colonize inland water; gemmules are asexual propagules able to survive in unfavourable climatic/environmental critical phases and to perform dispersal and defensive roles to persist in situ; architecture of gemmules and their morphotraits show a wide adaptive radiation worldwide (Manconi and Pronzato 2002, 2008, 2009, 2015, 2016a, b; Manconi 2008).
Although entire gemmules have not been found in the Naskal fossil remains, Longibirotula antiqua Manconi and Samant sp. nov. is well distinguished from the rest of the known fossil taxa by a unique spicular complement lacking skeletal microscleres, and composed of skeletal monaxons (long slim oxeas, shorter acanthoxeas) and long gemmuloscleres birotules. The combination of these three spicular morphs of Longibirotula antiqua Manconi and Samant sp. nov. partially resembles with that of some fossil and living taxa of the order Spongillida.
Gemmuloscleres morphologies and morphometries of Longibirotula antiqua Manconi and Samant sp. nov. indicate divergence from fossils birotule-bearing species of the worldwide reported genus Ephydatia Lamouroux, 1816 (Pronzato et al. 2017), i.e., (i) Ephydatia fossilis Traxler, 1894 (Miocene in age, western Palaearctic, Romania) with stout, spiny birotules (41–69 μm in length) and rotules with indented margins (Traxler 1894); (ii) Ephydatia kaiseri Rauff, 1926 (pre-middle Eocene in age, south-western Afrotropical, Namibia) with entire gemmules bearing smooth birotules (44–65 μm in length) and non-incised margins of flat rotules (Rauff 1926); (iii) Ephydatia chileana Pisera and Sáez, 2003 (Late Miocene in age, southwestern Neotropical, Atacama Region, Chile) with birotules (39–45 μm in length) bearing spiny shaft and rotules with irregular margins, often deeply incised (Pisera and Saez 2003), and (iv) Ephydatia cf. facunda Weltner, 1895 (middle Eocene in age, Northern Nearctic, Canada) with stout birotules (26–57 μm in length) with spiny shaft and strongly incised margins of both rotules (Pisera et al. 2016).
As for extant birotules-bearing Spongillida, Longibirotula antiqua Manconi and Samant sp. nov. slightly resembles some taxa, but gemmulosclere outline and/or rotules morphs differ from species of e.g., Anheteromeyenia Schröder, 1927; Corvoheteromeyenia Ezcurra de Drago, 1979; Corvomeyenia Weltner, 1913; Dosilia Gray, 1867; Heteromeyenia Potts, 1881; Racekiela Bass and Volkmer-Ribeiro, 1998, and Umborotula Penney and Racek, 1968 (Penney and Racek 1968; Manconi and Pronzato 2002, 2015, 2016b; Pronzato and Manconi 2019a).
The fossil Longibirotula antiqua Manconi and Samant sp. nov. partly shares gemmuloscleres outline and length (47–76 µm) with extant species of Ephydatia (e.g., 45–50 µm, Ephydatia robusta Potts, 1887), and Heteromeyenia (e.g., 75–88 µm, Heteromeyenia stepanowi Dybowsky, 1884). The Longibirotula antiqua Manconi and Samant sp. nov. birotules morphs and length particularly resembles the slender birotules known from the species of the genus Heterorotula Penney and Racek, 1968 e.g., Heterorotula kakahuensis (Traxler, 1896) (38–44 μm from New Zealand), Heterorotula multidentata (Weltner, 1895) (64–84 µm from Australia, Penney and Racek 1968; 32–48 μm from New Caledonia, Rützler 1968) and Heterorotula caledonensis (Rützler, 1968) (30–100 μm from New Caledonia) (Penney and Racek 1968; Rützler 1968; Racek 1969; Pronzato and Manconi 2019b).
As for megascleres, smooth to microspiny long oxeas of Longibirotula antiqua Manconi and Samant sp. nov. (142–425 × 6–13 µm) partly show similarity with the megascleres morphometric values in the species of Heterorotula (Penney and Racek 1968; Racek 1969; Volkmer-Ribeiro and Motta 1995; Rützler 1968; Pronzato and Manconi 2002, 2019b), namely (i) the Australian endemic lineage, e.g., Heterorotula capewelli (Bowerbank, 1863) (195–330 × 13–18 µm, type species), Heterorotula nigra (Lendenfeld, 1887)(224–360 × 7–13 μm), Heterorotula multidentata (284–320 × 10–18 μm), Heterorotula multiformis (Weltner, 1910) (330–420 × 13–20 μm), (ii) the New Zealand–New Caledonia endemic lineage with Heterorotula kakahuensis (170–288 × 8–22 μm) and Heterorotula caledonensis (100–150 × 8–19 μm) and, in addition, with (iii) the Neotropical endemic lineage Heterorotula fistula Volkmer-Ribeiro and Motta, 1995 (196–361 × 16–31 μm).
Furthermore, short acanthoxeas of Longibirotula antiqua (71–104 × 6–9 μm, here interpreted as gemmular cage components) partly show similarity with those of Heterorotula, which were reported as a short skeletal megascleres and/or belonging to the gemmular cage, particularly evident in H. kakahuensis endemic to New Zealand (Pronzato and Manconi 2019b: fig. 7C, D) and H. caledonensis endemic to New Caledonia (Pronzato and Manconi 2019b: fig. 4A, B)
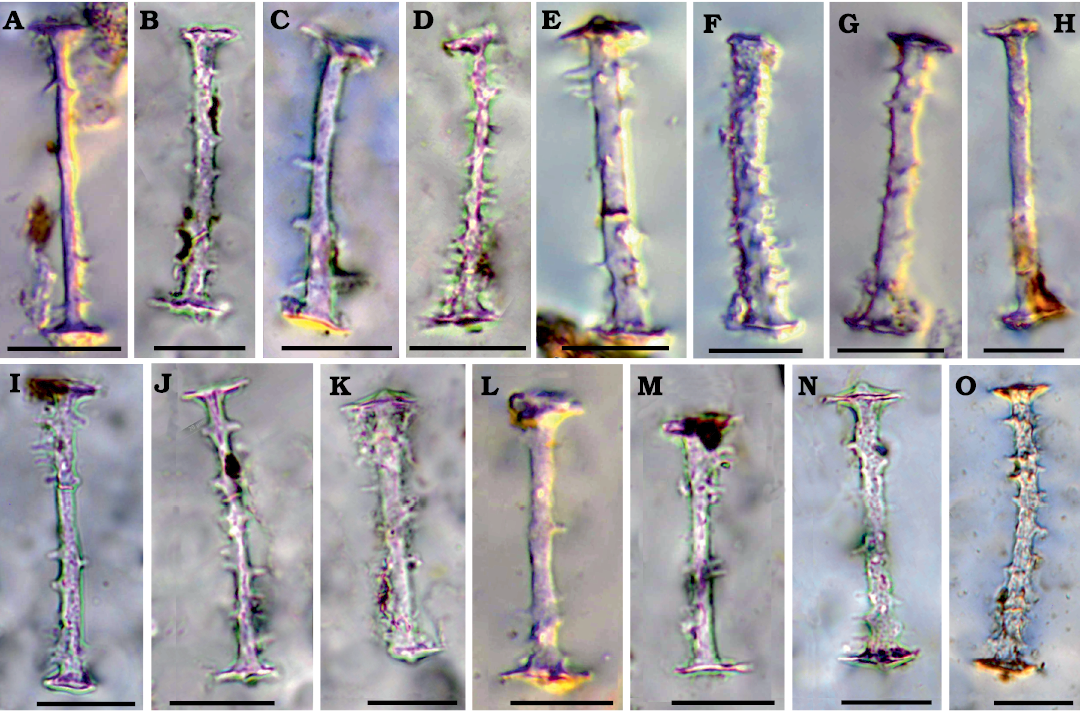
Fig. 4. Gemmuloscleres of palaeospongillid sponge Longibirotula antiqua gen. et sp. non. from Upper Cretaceous–lower Paleocene of Naskal GSI Quarry (India). A–O. Birotules (slides PGNU/NSKQ/SL-1–13) slender, spiny, with long shaft. Diagenetic processes affect all spicules to various degree. Scale bars 20 µm.
In conclusion, the gemmuloscleres and megascleres morphotraits of Longibirotula antiqua gen. et sp. nov. are similar to that of some species of Heterorotula known from living populations in Australia, New Zealand, and New Caledonia and from subequatorial Brazil fossils remains, suggesting a Gondwanan track which is not seen in the Afrotropical Region (Manconi and Pronzato 2009).
The old evolutionary history and radiation of Spongillida seem to be written in the gemmular architecture of coeval fossil remains. The Longibirotula antiqua Manconi and Samant sp. nov. from the Deccan intertrappean sediments close to the Cretaceous/Paleogene boundary, presumably displays a radial arrangement of birotule spicules in the gemmular theca. In contrast, the Deccan Upper Cretaceous (Maastrichtian, Chron 30N) intertrappean sediments of Malwa Group were inhabited by Palaeocorvospongilla cretacea characterized by a totally different spicular complement of spiny stout oxeas/strongyles/strongyloxeas as gemmuloscleres and pseudobirotules as skeletal microscleres (Samant et al. 2021). The gemmules of the Early Cretaceous Palaeospongilla chubutensis Ott and Volkmer, 1972, from Patagonia (Volkmer-Ribeiro and Reitner 1991) has oxeas gemmuloscleres which are almost radially, irregularly arranged in the gemmular theca. On the basis of these record, the family Palaeospongillidae is enlarged and now comprises five monotypic genera: Eospongilla (Eospongilla morrisonensis Dunagan, 1999), Longibirotula Pronzato and Manconi gen. nov., Lutetiospongilla (Lutetiospongilla heili Richter and Wuttke, 1999), Palaeocorvospongilla Pronzato and Manconi, 2021, and Palaeospongilla Ott and Volkmer, 1972.
The discovery of the new species Longibirotula antiqua confirm that the Deccan volcanic province in India is a favourable area to study the natural history of freshwater sponges adaptive processes. The data give us more information about how the anatomy of Spongillida have changed or persisted over time and how they have always been able to drive a morphological diversification by a successful evolution of resistant bodies, i.e., gemmules (Manconi and Pronzato 2002, 2015, 2016a). Importantly, the long lasting structural conservative trend of freshwater sponges (Pisera 2006; Pronzato et al. 2017; Samant et al. 2021) seems to be confirmed by Deccan spicule morphologies that remained almost unchanged through tens of millions of years.
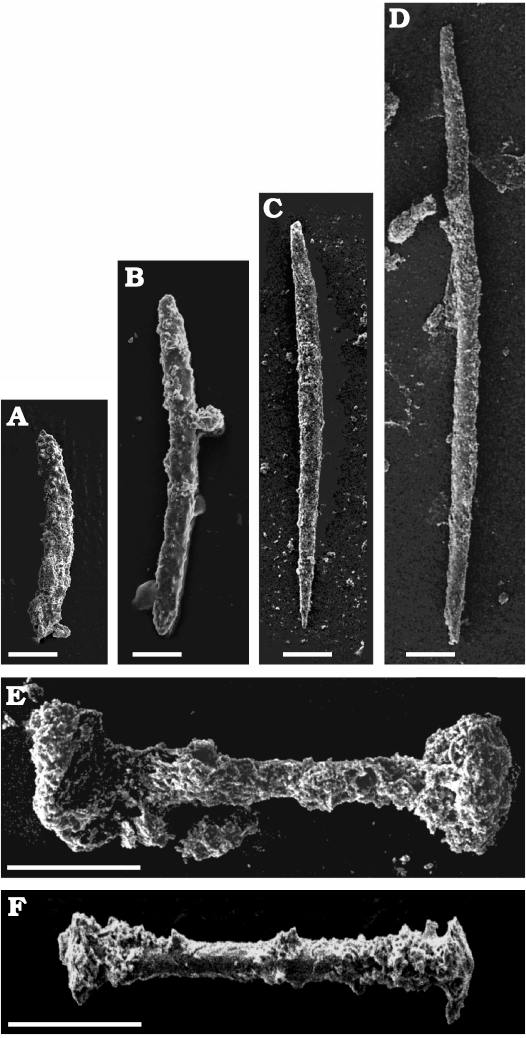
Fig. 5. Spicular complement of skeleton and gemmules of palaeospongillid sponge Longibirotula antiqua gen. et sp. nov. from Upper Cretaceous–lower Paleocene of Naskal GSI Quarry (India) (slides PGNU/NSKQ/ST-1, 2). A, B. Acanthoxeas short with dense spines. C, D. Oxeas fusiform, long and with acute tips. E, F. Birotules with long shaft. Diagenetic processes affect all spicules to various degree. Scale bars 20 µm.
Acknowledgements
BS and DMM are thankful to Kirtikumar R. Randive (Department of Geology, RTM Nagpur University, India), for providing working facilities. We are also thankful to the Jawaharlal Nehru Aluminium Research Development and Design Centre, Nagpur, India for help in SEM study. Special thanks to Laura Negretti (DISTAV, Genova, Italy) for her kind efforts in technical cooperation. We are also thankful to the anonymous reviewer and Joachim Reitner (Georg-August-Universität Göttingen, Germany) for useful comments and suggestions that significantly improved the manuscript. BS and DMM are thankful to the Science and Engineering Research Board, New Delhi under grant (CRG/2020/001339) and National Science Foundation (EAR-1736787) for financial assistance. RM research was co-funded by the Fondazione di Sardegna (grant FdS/RAS-2016/CUP J86C1800082005), Regione Autonoma Sardegna (grant RAS2012-LR7/2007-CRP-60215), Parco Nazionale dell’Asinara (grant PNA2016), and Sassari University (grant 2019-2021).
References
Ahluwalia, A.D. 1990. Detailed geological map and field observations in the Cretaceous mammal bearing locality, Hyderabad District, Andhra Pradesh, India. In: A. Sahni and A. Jolly (eds.), Cretaceous Event Stratigraphy and the Correlation of the Indian Nonmarine Strata. Proceedings Seminar cum Workshop IGCP 216 and 245, 120–122. Chandigarh.
Khosla, A. and Sahni, A. 2003. Biodiversity during the Deccan volcanic eruptive episode. Journal of Asian Earth Sciences 21: 895–908. Crossref
Manconi, R. 2008. The genus Ephydatia (Spongillina: Spongillidae) in Africa: a case of Mediterranean vs. southern Africa disjunct distribution.Biogeographia. The Journal of Integrative Biogeography 29: 19–28. Crossref
Manconi, R. and Pronzato, R. 2002. Spongillina new suborder, Lubomirskiidae, Malawispongiidae n. fam., Metaniidae, Metschnikowiidae, Palaeospongillidae, Potamolepidae, Spongillidae. In: H.J.N Hooper and R.W.M. Van Soest (eds.), Systema Porifera: A Guide to the Classification of Sponges 1, 921–1019. Kluwer Academic/Plenum Publishers, New York. Crossref
Manconi, R. and Pronzato, R. 2008. Global diversity of sponges (Porifera: Spongillina) in freshwater. Hydrobiologia 595: 27–33. Crossref
Manconi, R. and Pronzato, R. 2009. Atlas of African freshwater sponges. Studies in Afrotropical Zoology 295: 1–214.
Manconi, R. and Pronzato, R. 2015. Phylum Porifera. In: J.H. Thorp and D.C. Rogers (eds.), Ecology and General Biology: Thorp and Covich’s Freshwater Invertebrates, vol 1. 4th Edition, 133–157. Academic Press, Elsevier, London. Crossref
Manconi, R. and Pronzato, R. 2016a. How to survive and persist in temporary freshwater? Adaptive traits of sponges (Porifera, Spongillida). A review. Hydrobiologia 782: 11–22. Crossref
Manconi, R. and Pronzato, R. 2016b. Phylum Porifera. In: J.H. Thorp and D.C. Rogers (eds.), Keys to Nearctic Fauna: Thorp and Covich’s Freshwater Invertebrates, vol 2, 4th Edition, 39–83. Academic Press, Elsevier, San Diego. Crossref
Mohabey, D.M. 1996. Depositional environments of Lameta Formation (Late Cretaceous) of Nand-Dongargaon inland basin, Maharashtra: The fossil and lithological evidence. Memoirs Geological Society of India 37: 363–386.
Mohabey, D.M. and Udhoji, S.G. 2000. Vertebrate Fauna of Late Cretaceous dinosaur bearing Lameta Formation of Nand-Dongargaon inland basin, Maharashtra and KT boundary implications. Memoirs Geological Society of India 46: 295–322.
Penney, J.T. and Racek, A.A. 1968. Comprehensive revision of a worldwide collection of freshwater sponges (Porifera, Spongillidae). Bulletin of the United States National Museum 272: 1–184. Crossref
Pisera, A. 2006. Palaeontology of sponges—a review. Canadian Journal of Zoology 84: 242–261. Crossref
Pisera, A. and Saez, A. 2003. Paleoenvironmental significance of a new species of freshwater sponge from the Late Miocene Quillagua Formation (N Chile). Journal of South American Earth Studies 15: 847–852. Crossref
Pisera, A., Manconi, R., Siver, P., and Wolfe, A. 2016. The sponge genus Ephydatia from the Middle Eocene: environmental and evolutionary significance. Paläontologische Zeitschrift 90: 673–680. Crossref
Pisera, A., Siver, P.A., and Wolfe, A.P. 2013. A first account of freshwater potamolepid sponges (Demospongiae, Spongillina, Potamolepidae) from the middle Eocene: biogeographic and paleoclimatic implications. Journal of Paleontology 87: 373–378. Crossref
Prasad, G.V.R. 2012. Vertebrate biodiversity of the Deccan volcanic province of India: a review. Bulletin de la Société géologique de France 183: 597–610. Crossref
Prasad, G.V.R. and Sahni, A. 1988. First Cretaceous mammal from India. Nature 332: 638–640. Crossref
Pronzato, R. and Manconi, R. 2019a. An overview on the freshwater sponge fauna (Demospongiae: Spongillida) of New Zealand and New Caledonia with new insights into Heterorotula from deep thermal vents of the Lake Taupo. Journal of Natural History 53: 2207–2229. Crossref
Pronzato, R. and Manconi, R. 2019b. Phylum Porifera. In: J.H. Thorp and D.C. Rogers (eds.), Keys to Palaearctic Fauna. Thorp and Covich’s Freshwater Invertebrates. 4th Edition, 39–83. Academic Press, London.
Pronzato, R. and Manconi, R. (in press). Phylum Porifera. In: J.H. Thorp and D.C. Rogers (eds.), Keys to Australasian Fauna. Thorp and Covich’s Freshwater Invertebrates. 4th Edition. Academic Press, Elsevier, London.
Pronzato, R., Pisera, A., and Manconi, R. 2017. Fossil freshwater sponges: Taxonomy, geographic distribution, and critical review. Acta Palaeontologica Polonica 62: 467–495. Crossref
Rauff, H. 1926. Überprämitteleozäne fossilfuhrende Süswasser-Hornsteineaus der Namib. In: E. Kaiser (ed.), Die Diamantenwuste Südwest-Afrikas 2: 160–166.
Racek, A.A. 1969. The freshwater sponges of Australia (Porifera: Spongillidae). Marine and Freshwater Research 20: 267–310. Crossref
Rützler, K. 1968. Fresh-water sponges from New Caledonia. Cahiers ORSTOM (Office de la Recherche Scientifique et Technique OutreMer), Series Hydrobiologie 2: 57–66.
Samant, B. and Mohabey, D.M. 2009. Palynoflora from Deccan volcano-sedimentary sequence (Cretaceous–Paleocene transition) of central India: implications for spatio-temporal correlation. Journal of Biosciences 34: 811–823. Crossref
Samant, B. and Mohabey, D.M. 2014. Deccan volcanic eruptions and their impact on flora: palynological evidence. Geological Society of America, Special Papers 505: 171–191. Crossref
Samant, B., Mohabey, D.M., Srivastava, P., and Thakre, D. 2014. Paleocene palynoflora from the intertrappean sediments of Saurashtra, Gujarat: age and paleoenvironments. Journal of Earth System Science 123: 219–232. Crossref
Samant, B., Pronzato, R., Mohabey, D.M., Kumar, D., Dhobale, A., Pizal, P., and Manconi, R. 2021. Insight into the evolutionary history of freshwater sponges: A new genus and new species of Spongillida (Porifera: Demospongiae) from Upper Cretaceous (Maastrichtian) Deccan intertrappean lacustrine deposits of the Malwa Group, Central India. Cretaceous Research 126: 104851. Crossref
Schindler, T., Wuttke, M.P., and Poschmann, M. 2008. Oldest record of freshwater sponges (Porifera: Spongillina) spiculite finds in the Permo-Carboniferous of Europe. Paläontologische Zeitschrift 82: 373–384. Crossref
Traxler, L. 1894. Ephydatia fossilis, eine neu Art der fossilen Spongilliden. Földatni Közlöny 24: 234–237.
Volkmer-Ribeiro, C. and Motta, J.F.M. 1995. Esponjas formador as de espongilitos em lagoas no Triângulo Mineiro e ajacências com indicaçao de preservaçao de habitat. Biociencias 3: 183–205.
Volkmer-Ribeiro, C. and Reitner, J. 1991. Renewed study of the type material of Palaeospongilla chubutensis Ott and Volkheimer (1972). In: J. Reitner and H. Keupp (eds.), Fossil and Recent Sponges, 121–133. Springer, Berlin. Crossref
Voogd, N.J. de, Alvarez, B., Boury-Esnault, N., Carballo, J.L., Cárdenas, P., Díaz, M.-C., Dohrmann, M., Downey, R., Hajdu, E., Hooper, J.N.A., Kelly, M., Klautau, M., Manconi, R., Morrow, C.C., Pisera, A.B., Ríos, P., Rützler, K., Schönberg, C., Vacelet, J., and van Soest, R.W.M. 2023. World Porifera Database [available online, https://www.marinespecies.org/porifera/porifera.php?p=taxdetails&id=845456 on 2023-03-06].
Wilson Mantilla, G.P., Renne, P.R., Samant, B., Mohabey, D.M., Dhobale, A., Tholt, A.J., Tobin, T.S., Widdowson, M., Anantharaman, S., Dassarma, D.C., and Wilson Mantilla, J.A. 2022. New mammals from the Naskal intertrappean site and the age of India’s earliest eutherians. Palaeogeography, Palaeoclimatology, Palaeoecology 591: 110857. Crossref
Acta Palaeontol. Pol. 68 (1): 167–174, 2023
https://doi.org/10.4202/app.01040.2022