
New vertebrate microfossils expand the diversity of the chondrichthyan and actinopterygian fauna of the Maastrichtian–Danian Hornerstown Formation in New Jersey
ZACHARY M. BOLES, PAUL V. ULLMANN, IAN PUTNAM, MARIELE FORD, and JOSEPH T. DECKHUT
Boles, Z.M., Ullmann, P.V., Putnam, I., Ford, M., and Deckhut, J.T. 2024. New vertebrate microfossils expand the diversity of the chondrichthyan and actinopterygian fauna of the Maastrichtian–Danian Hornerstown Formation in New Jersey. Acta Palaeontologica Polonica 69 (2): 173–198.
The abundance of shark and actinopterygian fossils in the Cretaceous and Paleogene strata of the Atlantic Coastal Plain is well documented; but much remains unknown about the survivorship patterns of these major components of shallow marine faunas in the western Atlantic through the K/Pg mass extinction. To shed light on this subject, we describe an assemblage of new actinopterygian, chondrichthyan, and reptilian microfossils recently recovered from the Maastrichtian Navesink and Maastrichtian–Danian Hornerstown formations at the Jean and Ric Edelman Fossil Park at Rowan University in Mantua Township, New Jersey. The new microfossils clarify extinction patterns across the K/Pg, create temporal and geographic range extensions for several taxa, and expand the known fauna of this regionally-rare and important K/Pg-boundary locality. We report 11 new additions to the vertebrate fauna of Edelman Fossil Park, the first Paleocene record of Saurocephalus lanciformis, the first Cretaceous records of Paralbula marylandica and Palaeogaleus vincenti, and the first recovery of gar and dercetid fish remains from the Paleocene in New Jersey (the last indicating that these fish survived the K/Pg extinction in the western Atlantic). Geographic range extensions include: Notidanodon brotzeni into the Western Hemisphere, Saurocephalus into northeastern North America and Phyllodus paulkatoi to the eastern coast of North America. A dentary of a juvenile alligatorid, Bottosaurus harlani, indicate that the mandible exhibited isometric growth through ontogeny. Our findings generally agree with other studies that these groups were significantly impacted by the extinction event, that extinctions were selective, and recovery was slow. This wealth of novel insights garnered from microfossils in this study highlights their critical importance for elaborating past faunas and illuminating the character of ancient ecosystems. We therefore recommend microsieving as a fruitful method for future faunal studies of shallow-marine strata and predict that such efforts will frequently yield similar important insights.
Key words: Actinopterygii, Chondrichthyes, microfossils, K/Pg, Hornerstown Formation, Edelman Fossil Park, New Jersey, USA.
Zachary M. Boles [bolesz@rowan.edu; ORCID: https://orcid.org/0009-0007-4086-6860 ], Department of Geology, Rowan University, Glassboro, New Jersey 08028, USA; Jean and Ric Edelman Fossil Park at Rowan University, Mantua Township, New Jersey 08080, USA.
Paul V. Ullmann [paul.ullmann@und.edu; ORCID: https://orcid.org/0000-0003-1457-6580 ], Harold Hamm School of Geology and Geological Engineering, University of North Dakota, Grand Forks, North Dakota 58202, USA.
Ian Putnam [ianputnamdvc@yahoo.com], Jean and Ric Edelman Fossil Park and Museum of Rowan University, Mantua Township, New Jersey 08080, USA.
Mariele Ford [marieleford3@gmail.com], Biomedical Arts and Visualization, Rowan University, Glassboro, New Jersey 08028, USA.
Joseph T. Deckhut [jdeckhut@ewu.edu; ORCID: https://orcid.org/0000-0001-5034-6788 ], Department of Biology, Eastern Washington University, Cheney, Washington 99004, USA.
Received 1 November 2023, accepted 8 March 2024, published online 12 April 2024.
Copyright © 2024 Z.M. Boles et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The remains of sharks and actinopterygian fish represent the most common vertebrate fossils in Cretaceous and Paleogene shallow-marine deposits across the Gulf and Atlantic Coastal Plain Provinces of North America (Case and Schwimmer 1988). Like today, these groups accounted for the majority of marine vertebrate diversity during these time periods, yet they tend to receive less attention than larger inhabitants of Cretaceous and Paleogene seas (i.e., turtles, crocodilians, extinct marine reptiles, and birds; Gallagher 1993). This unintentional bias continues to result in numerous uncertainties regarding the taxonomic identities (i.e., potential synonymies), phylogenetic relationships, and geographic and temporal ranges of chondrichthyan and actinopterygian taxa that inhabited coastal waters along the eastern margin of Appalachia (Case and Schwimmer 1988; Case 1996; Callahan et al. 2014; Andrews et al. 2023). In many cases, it also remains unclear which taxa survived the K/Pg mass extinction and which did not (Adolfssen and Ward 2014; Schwarzhans and Stringer 2020).
The collective dearth of knowledge of Cretaceous and Paleogene fish arises in-part from their fossil record consisting primarily of shed teeth and disarticulated, isolated specimens, the vast majority of which are “microfossils” <10 mm in size (e.g., Bazzi et al. 2015; Ebersole et al. 2019). This problem is compounded for sharks due to the cartilaginous composition of the majority of their skeleton (Heinicke et al. 2009), leaving only their teeth and vertebrae as mineralized remains (e.g., Adolfssen and Ward 2014, 2015); as well as by their routine shedding and replacement of teeth (e.g., Cappetta 1987). Teeth of select, larger sharks and fish are sizable enough that they may be found by surface collecting at several prolific fossil sites across the Gulf and Atlantic Coastal Plain Provinces (e.g., Jean and Ric Edelman Fossil Park at Rowan University, Gallagher 1993; Holmdel Park, New Jersey, Callahan et al. 2014; Dinosaur Park, Maryland, Frederickson et al. 2018; Ten Bits Microsite, Texas, Schubert et al. 2017); but the teeth and bones of many selachians and fish are so small that they can only be found by sieving/screen-washing (Peterson et al. 2011). For this reason, screen-washing has become a standard method to explore the diversity of microvertebrates in unconsolidated sediments around the world (e.g., Case and Schwimmer 1988; Purdy et al. 2001; Kirkland et al. 2013; Adolfssen and Ward 2014, 2015; Callahan et al. 2014; Schubert et al. 2017; Frederickson et al. 2018).
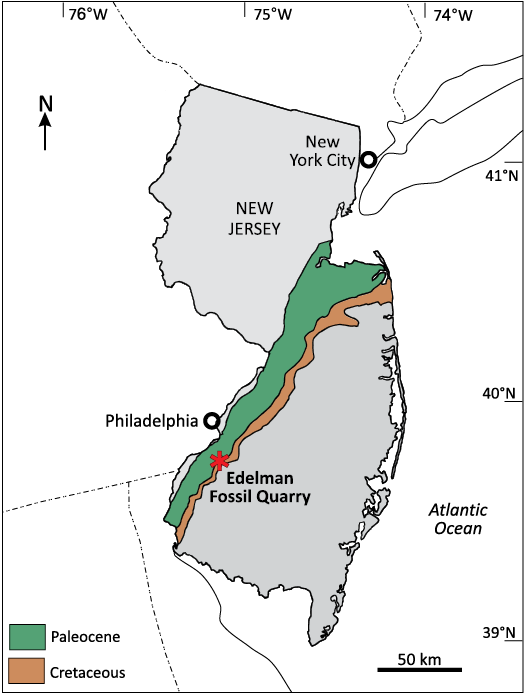
Fig. 1. Map showing the location of the outcrops of Cretaceous and Paleocene strata and Jean and Ric Edelman Fossil Park Quarry in Mantua Township, New Jersey, USA. Reproduced, with permission, from Ullmann et al. (2018).
In this study, we describe a microvertebrate assemblage recovered from the Jean and Ric Edelman Fossil Park Quarry (EFPQ) at Rowan University (formerly an Inversand Company marl quarry) in Mantua Township, New Jersey (Fig. 1). The EFPQ preserves one of the best exposures of Cretaceous–Paleogene strata along the eastern coast of the United States (Gallagher 2003), and fossils from this famous locality provide a nearly-unrivaled snapshot into the diversity of west-Atlantic, shallow-marine faunas across the K/Pg boundary (Gallagher 1993). Strata in the EFPQ not only host evidence of the K/Pg impact event, but also of its subsequent effects on shallow-marine invertebrate and vertebrate faunas and ecosystems (e.g., Koch and Olsson 1977; Obasi et al. 2011; Esmeray-Senlet et al. 2017; Wiest et al. 2016). Our study of vertebrate microfossils recovered from the EFPQ expands the known fauna from this K/Pg locality, reveals patterns of survivorship and extinction across this mass extinction, and yields temporal and geographic range extensions for several chondrichthyan and actinopterygian taxa.
Institutional abbreviations.—ANSP, Academy of Natural Science of Drexel University, Philadelphia, USA; RU-EFP, Jean and Ric Edelman Fossil Park, Rowan University, Mantua Township, USA; NJSM, New Jersey State Museum, Trenton, USA.
Other abbreviations.—EFPQ, Jean and Ric Edelman Fossil Park Quarry; MFL, main fossiliferous layer.
Geological setting
The majority of the fossils described herein were recovered from the Main Fossiliferous Layer (MFL) of the Maastrichtian–Danian Hornerstown Formation. Within the EFPQ, the Hornerstown Formation is underlain by the Maastrichtian Navesink Formation and overlain by the Thanetian Vincentown Formation (see Gallagher 1993 for a graphic representation of the full stratigraphic sequence exposed at the EFPQ). All three of these formations consist of glauconitic greensands deposited in organic-rich, siliciclastic, shallow-marine environments (Gallagher 1993). More specifically, in the EFPQ only the uppermost ~1.5 m of the underlying Navesink Formation is exposed in the bottom of the quarry; it is composed of olive-gray to dusky brown, clayey-glauconitic sand that was deposited on the inner shelf (Wiest et al. 2016). The overlying Hornerstown Formation is 6.2 m thick and consists of fine- to medium-grained, grayish-olive-green glauconitic sand that was deposited on the mid-to-outer shelf (Wiest et al. 2016). The overlying Vincentown Formation, which is above the interval examined herein, represents the end of the conformable sequence of strata in the lower portion of the EFPQ; it is 1.6 m thick and composed of very fine- to medium-grained, grayish-olive-green to grayish-yellow, clayey-glauconitic sand that was deposited on the inner shelf (Gallagher 1993).
Within the lower Hornerstown Formation, the MFL is a prolific, ~10 cm-thick, regionally-vast bonebed located approximately ~20–30 cm above the Navesink-Hornerstown formational contact (Voegele et al. 2021). Remains of numerous shallow-marine invertebrates and paralic turtles, birds, crocodilians, fish, sharks, and rays are preserved within the MFL (Gallagher 1993, 2003); as well as occasional remains of mosasaurs and “bloat-and-float” dinosaur carcasses (e.g., Schein and Poole et al. 2011).
The taphonomic origin of the MFL has been debated for decades, with explanations ranging from attritional accumulation (i.e., a condensed assemblage; Gallagher 2003, 2012; Schein et al. 2008), reworking of Cretaceous remains into an early Danian storm/tsunami lag deposit (Minard et al. 1969; Gallagher 2002; Olsson et al. 2002; Horner et al. 2016), or relatively-rapid accumulation of remains via mass-death following the Chicxulub impact (Gallagher 1993; Obasi et al. 2011; Boles 2016; Wiest et al. 2016). We continue to conduct excavations in the EFPQ to resolve this question; but recent discoveries of shocked quartz grains within a burrow immediately below the MFL (Obasi et al. 2011) and a spike in iridium concentrations within the MFL in the EFPQ (Esmeray-Senlet et al. 2017) indicate that the MFL may represent a thanatocoenosis recording ecosystem collapse during the K/Pg mass extinction. Based on the inclusion of impact indicators and the definition of the K/Pg boundary as the moment of impact (Molina et al. 2006), we regard the MFL as Danian in age.
Material and methods
The majority of the fossils described herein were recovered by traditional screen-washing of sediments from the uppermost Navesink Formation and the lower Hornerstown Formation. A few (primarily-larger) fossil specimens were recovered either by surface collecting from outcrops of the Hornerstown Formation within the current quarry or during field excavations through the MFL in the EFPQ. Many specimens we describe were recovered during a recent, detailed microstratigraphic study of the MFL (Voegele et al. 2021). A 1.4 mm mesh sieve was used, at times in the field and at other times with collected sediment samples in a lab setting, to produce concentrates which were dried and hand-picked under a dissecting microscope, following traditional methods (e.g., Cifelli et al. 1996). Screen-washing of the 61 sediment samples collected as part of that project yielded abundant microvertebrate fossils (e.g., teeth, vertebrae, scales) whose positions relative to the Navesink-Hornerstown formational contact are constrained with centimeter resolution. A number of additional specimens were recovered by earlier screen-washing of excavated sediments sourced from our ongoing field excavations through this same, roughly half-meter, stratigraphic interval spanning the MFL; the precise origin of these sediments, and thus the microfossils within them, were only constrained to “above MFL”, “within MFL”, or “(within the basal Hornerstown Fm.) below the MFL” (this last category equates with the lowest ~20 cm of the Hornerstown Formation; cf. Voegele et al. 2021). All but the largest few specimens were imaged using a Zeiss Stemi 508 dissecting microscope with an integrated AxioCam 506 camera, occasionally using the Manual Extended Depth of Focus (Z-stack) module (where needed for clarity).
Systematic palaeontology
Chondrichthyes Huxley, 1880
Carcharhiniformes Compagno, 1973
Triakidae Gray, 1851
Genus Palaeogaleus Gurr, 1962
Type species: Galeocerdo vincenti Daimeries, 1888; Thanetian, Orple-Grand, Belgium.
Palaeogaleus vincenti (Daimeries, 1888)
Fig. 2A.
Material.—Seventeen teeth: one lateral tooth (RU-EFP- 00284-1) from the upper Navesink Formation; one anterior tooth (RU-EFP-00268-1) from the lower Hornerstown Formation below the MFL; two anterior teeth (RU-EFP- 00248-1 and 00254-1) from the MFL; one anterior tooth (RU-EFP-04143) and four lateral teeth (RU-EFP-00158-1, 00225-1, 00233-1, and 00238-1) from the lower Hornerstown Formation above the MFL. The remainder (RU-EFP-04144, 04145, and 00157-1–00157-6) were collected as float. All from the Maastrichtian Navesink and Maastrichtian–Danian Hornerstown formations, Jean and Ric Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The principal cusp is broad, strongly convex lingually, labiolingually thick, and angled slightly apicodistally in anterior teeth; in lateral teeth, it is more strongly apicodistally directed. The mesial and distal cutting edges are smooth. Distal cusplets vary in distinction and range from one to four in number. The first distal cusplet is generally around one-third the height of the principal cusp, and succeeding distal cusplets (when present) are shorter. Mesial cusplets range from one to four in number on anterior teeth, and are generally absent in lateral teeth (cf. Cappetta 1987: fig. 100D). Only one of the lateral teeth possesses a subtle, undulating peak along the mesial heel. In lateral teeth, the distal heel is shorter than the mesial heel. The labial base of the crown strongly overhangs the root in all specimens preserving roots, and appears to curl lingually in the same manner in specimens lacking the root. Coarse vertical costules cover the entire labial crown base on most specimens, as well as basal portions of each heel on the lingual face; however, a single lateral and anterior tooth each exhibit nearly smooth labial heels. The basal border of the crown ranges from flat to slightly concave. The root is labiolingually thick and holaulacorhizid in structure, with a deep central groove vertically traversing the lingual protuberance. A small, circular pit occupies the center of this groove on the lingual face in six of the specimens. Where preserved, lateral branches of the root are broad and have flat, wide basal faces.
Remarks.—These teeth can be assigned to Palaeogaleus owing to strong overhang of the labial crown base over the root, the strongly convex lingual face of the principal cusp, relative shortness of the principal cusp, and having a thick, holaulacorhizid root (Cappetta 1987; Hovestadt and Hovestadt-Euler 2002). Among the eight species of Palaeogaleus currently recognized, the combination of coarse, vertical costules on the labial crown base and overall weak development of mesial and distal cusplets in the Edelman Fossil Park teeth compares best with Palaeogaleus vincenti. Cusplets are generally more well developed in Palaeogaleus havreensis Herman, 1977, Palaeogaleus dahmanii Noubhani & Cappetta, 1999, and Palaeogaleus larachei Noubhani & Cappetta, 1999, and labial crown base ornamentation is absent or poorly developed in Palaeogaleus brivesi Arambourg, 1952, Palaeogaleus faujasi Geyn, 1937, and Palaeogaleus prior Arambourg, 1952 (Hovestadt and Hovestadt-Euler 2002). Also, in contrast to Palaeogaleus prior, Palaeogaleus sublaevis Noubhani & Cappetta, 1999, and Palaeogaleus larachei (Hovestadt and Hovestadt-Euler 2002), the principal cusp is not serrated in any of the RU-EFP specimens. A pit within the midline groove was also described in Palaeogaleus vincenti teeth by Cvancara and Hoganson (1993).
Palaeogaleus vincenti represents a new addition to the fauna of the EFPQ. Case (1996) briefly described 12 specimens of Palaeogaleus vincenti from the lower Hornerstown Formation of Monmouth County, New Jersey, which, at the time, represented the first occurrence of the species in eastern North America. However, the stratigraphic occurrences of Palaeogaleus vincenti teeth from the EFPQ are more precisely constrained and demonstrate that it is present throughout all portions of the lower Hornerstown Formation (specifically above, within, and below the MFL) as well as in the upper Navesink Formation. All previous occurrences, including that described by Case (1996), were from the Paleogene (Cappetta 2012; Maisch et al. 2020); thus, the two specimens of Palaeogaleus vincenti recovered the Navesink Formation and beneath the MFL in the Hornerstown Formation represent the first Cretaceous occurrences of this species, indicating that Palaeogaleus vincenti evolved before the K/Pg extinction.
Stratigraphic and geographic range.—Upper Cretaceous, Maastrichtian to Paleocene of New Jersey in the USA, Belgium, England, France, and Morocco
Chondrichthyes Huxley, 1880
Chimaeriformes Obruchev, 1953
Callorhynchidae Garman, 1901
Genus Ischyodus Egerton, 1843
Type species: Ischyodus townsendi Buckland, 1835; Jurassic, Stonesfield, England.
Ischyodus bifurcatus (Case, 1978)
Fig. 2B.
Material.—Seventeen tooth plates: four left mandibulars, six right mandibulars, five left palatines, and two right palatines from the MFL of the Maastrichtian–Danian Hornerstown Formation, Jean and Ric Edelman Fossil Park, Mantua Township, New Jersey, USA. (SOM 1: table 1, Supplementary Online Material available at http://app.pan.pl/SOM/app69-Boles_etal_SOM.pdf).
Description.—Mandibular tooth plates are robust, labiolingually narrow, and possess a short mesial ramus and “stepped” labial margin (cf. Cicimurri and Ebersole 2014). They possess four tritors in occlusal view: anterior inner tritor, “median” tritor (fused middle and posterior inner tritors; Case 1978), anterior external tritor, and posterior external tritor. The “median” tritor is expansive with an embayment into its mesial margin that creates bifurcations that are unequal in length: the outer bifurcation extends further mesially than the inner bifurcation. The mandibular symphysis is thin and curves longitudinally. The anterior external tritor is positioned slightly ventrally compared to the surface of the “median” tritor and is commonly angled slightly anterolaterally relative to the sagittal axis. Axes of the other tritors are nearly parallel and parasagittal. Where preserved, the labial face is smooth and slightly concave.
Palatine tooth plates are roughly triangular in occlusal view and narrower than the mandibular tooth plates. Four tritors are present: anterior inner tritor, posterior inner tritor, median tritor, and outer tritor. Each of these tritors are long and narrow labiolingually. The anterior inner tritor is positioned on an elevated, roughly flat platform compared to the rest of the lingual face and is commonly angled slightly anterolaterally relative to the sagittal axis. The posterior inner and median tritors are approximately equal in length, though the median tritor occasionally extends slightly further mesially in some specimens. The medial face is flat and straight. The aboral surface is roughly flat in most specimens, except those in which surficial bone and internal tissue of the anterior inner tritor have been lost; in these specimens, erosion has created a deep, rounded longitudinal groove representing the lingual border of the anterior inner tritor tissue. The labial face is flat with a sigmoidal lateral margin.

Fig. 2. Chondrichthyan ichthyoliths from the Cretaceous–Paleogene lower Hornerstown Formation at the Jean and Ric Edelman Fossil Park in Mantua Township, New Jersey, USA. Anterolateral (A, I, K) and lateral (C–H) teeth, mandibular toothplate (B), and spine (J). A. Triakid carchariniform Palaeogaleus vincenti (Daimeries, 1888) (RU-EFP-04145) in labial (A1) and lingual (A2) views. B. Callorhynchid chimaeriform Ischyodus bifurcatus (Case, 1978) (RU-EFP-03717) in oral (B1) and labial (B2) views. C. Hexanchid Heptranchias howellii (Reed, 1946) (RU-EFP-04139) in labial (C1) and lingual (C2) views. D. Hexanchid Hexanchus sp. (RU-EFP-02633) in labial (D1) and lingual (D2) views. E, F. Hexanchid Notidanodon brotzeni (Siverson, 1995), RU-EFP-04141 (E) and RU-EFP-03586 (F) in labial (E1, F1) and lingual (E2, F2) views. G. Hexanchid Weltonia ancistrodon (Arambourg, 1952) (RU-EFP-04142) in labial (G1) and lingual (G2) views. H. Pseudocoracid lamniform Pseudocorax affinis (Münster in Agassiz, 1843) (RU-EFP-02832) in labial (H1) and lingual (H2) views. I, J. Squalid Squalus sp. I. RU-EFP-02582 in lateral view. J. RU-EFP-00157-7 in labial (J1) and lingual (J2) views. K. Orthacodontid synechodontiform Sphenodus lundgreni (Davis, 1890) (RU-EFP-02913) in labial (K1), lingual (K2), mesial or distal (K3), occlusal (K4), and basal (K5) views.
Remarks.—Gallagher (1993, 2003) listed five chimaeroid fish among the fauna recovered from the EFPQ, including Ischyodus bifurcatus, Ischyodus thurmanni Pictet & Campiche, 1858, and three species of Edaphodon Buckland, 1838. The specimens described here most notably differ from Edaphodon in their possession of four tritors (rather than three) in both mandibular and palatine tooth plates (Cicimurri and Ebersole 2014; Ward and Grande 1991; Duffin and Reynders 1995; Stahl 1999). Additionally, the mandibular symphysis is thin and curved in mandibular tooth plates described herein, whereas in Edaphodon it is thick and nearly straight/flat (Cicimurri and Ebersole 2014; Ward and Grande 1991). Finally, the aboral surface of the palatine tooth plates are generally flat, as in Ischyodus, whereas those of Edaphodon usually exhibit a deep longitudinal sulcus (Cicimurri 2010, and references therein). Among the two species of Ischyodus listed by Gallagher (1993, 2003), the specimens described here can be assigned to I. bifurcatus based on attributes of the tritors: mandibulars exhibit an embayment into the anterior margin of the “median” tritor characteristic of this species, and the median and posterior inner tritors of the palatines extend approximately the same length anteriorly (whereas in I. thurmanni the posterior inner tritor commonly extends further anteriorly; Case 1978). Also as in I. bifurcatus (Case 1978), the outer tritor of the palatine is longer and narrower than in I. thurmanni. Mandibular tooth plates of I. rayhaasi Hoganson & Erickson, 2005, also exhibit an embayment into the “median” tritor, but it is the inner bifurcation that extends further anteriorly as opposed to the outer as in our specimens and I. bifurcatus (Hoganson and Erickson 2005; Hoganson et al. 2015).
Gallagher (1993, 2003) listed I. thurmanni as the only species of the genus present in the MFL, with I. bifurcatus being recovered from the underlying Navesink Formation. Elsewhere, I. bifurcatus has been described from Santonian to Maastrichtian deposits in Belgium and across the United States (Case and Schwimmer 1992). Our identification of numerous specimens of I. bifurcatus in the MFL extends its range across the K/Pg boundary into (presumably) the initial few years of the Danian (cf. Esmeray-Senlet et al. 2017).
Stratigraphic and geographic range.—Upper Cretaceous (Santonian) to lower Paleocene (Danian) of California, Delaware, Montana, New Jersey, and North Carolina in the USA, Belgium, and Russia.
Hexanchiformes Buen, 1926
Hexanchidae Gray, 1851
Genus Heptranchias Rafinesque, 1810
Type species: Squalus perlo Bonnaterre, 1788; Recent, in temperate waters worldwide.
Heptranchias howellii (Reed, 1946)
Fig. 2C.
Material.—Four teeth: three (RU-EFP-03582, 4137, and 4138) from the MFL and one (RU-EFP-04139) collected as float. All from the Maastrichtian–Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The principal cusp is fairly slender, apicodistally directed, and approximately twice as tall as the first few distal cusplets. The labial face of the principal cusp is nearly flat, whereas the lingual face is distinctly convex. The mesial and distal cutting edges of the principal cusp are nearly straight. Six mesial cusplets are present on each tooth. In the two nearly complete teeth, the first mesial cusplet is small and appears almost confluent with the mesial edge of the principal cusp, and it is followed by a larger, more distinct, second mesial cusplet. In the incomplete MFL specimen, the first mesial cusplet is large and distinct. Additional mesial cusplets steadily decrease in size. The mesial-most cusplet is directed mesially whereas succeeding cusplets turn to point apically then apicodistally as one follows their sequence up to the base of the principal cusp. The first three to four distal cusplets are roughly equal in breadth and height, while the final, distal-most cusplet is considerably smaller. Distal cusplets are directed apicodistally and exhibit roughly straight mesial and distal edges. The crown of RU-EFP-04138 has five distal cusplets, and RU-EFP-04139 has six. The basal border of the crown in these two specimens is slightly concave. The root is roughly rectangular in shape, with a pronounced mesial depression. In each tooth, the root is nearly as tall as the principal cusp, with a flat labial face and a rounded, mesio-distally directed ridge near the top of the lingual face. The basal border of RU-EFP-04139 (the only specimen with this edge complete) is flat.
Remarks.—The irregular decrease in size of the distal cusplets identify these teeth as belonging to Heptranchias (Cappetta 1987). Among described species, only Heptranchias howellii is known to commonly have more than three mesial cusplets (Welton 1974; Cappetta 1981; Adolfssen and Ward 2015); we therefore assign these teeth to that species. The Oligocene taxon Heptranchias ezoensis Applegate & Uyeno, 1968, is known to possess three or fewer mesial cusplets (Cappetta 1981; Carlsen and Cuny 2014), and Heptranchias tenuidens Leriche, 1938, is now considered a likely synonym of H. howellii (Cappetta 1987; Carrillo-Briceño et al. 2016).
Case (1973, 1980) mentioned the potential discovery of Heptranchias howellii from the Hornerstown Formation in Cream Ridge, New Jersey, which was viewed as suspect by Siverson (1995) based on similarity to teeth he described as Heptranchias sp. from the Danian of Sweden. We do not agree that the three features listed by Siverson (1995), namely slenderness of the principal cusp and sizes of the first distal and first mesial cusplets, readily distinguish any of these specimens from H. howellii. Rather, we find that the very subtle variations noted by Siverson (1995) to fit within the spectrum of morphologies in teeth attributed to H. howellii (e.g., Welton 1974; Cappetta 1987; Adolfssen and Ward 2015; Carrillo-Briceño et al. 2016). Thus, the specimens described here constitute a second recovery of H. howellii from the lower Hornerstown Formation of New Jersey and a new addition to the fauna of the EFPQ. Occurrence of two specimens in the MFL also indicates that H. howellii already existed in the earliest Danian, slightly older than other reports of Danian specimens from New Jersey (Case 1980) and elsewhere (Mannering and Hiller 2008).
Stratigraphic and geographic range.—Lower Paleocene (Danian) to lower Oligocene of Oregon and New Jersey in USA, Poland, and New Zealand.
Genus Hexanchus Rafinesque, 1810
Type species: Squalus griseus Bonnaterre, 1788; Recent, worldwide.
Hexanchus sp.
Fig. 2D.
Material.—Four lower lateral teeth: one (RU-EFP-02633) from the lower Hornerstown Formation above the MFL, three (RU-EFP-04140-1–04140-3) collected as float. All from the Maastrichtian-Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The teeth are labiolingually narrow and, in lingual view, are low and mesiodistally elongate with up to nine distal cusplets (in the sole complete specimen). In all specimens where it is preserved, the principal cusp is modestly larger than the first distal cusplet, termed “grisiform” by Ward (1979), with a nearly straight and unserrated distal cutting edge. Fine serrations are present along the lower half of the mesial edge of the principal cusp. The tip of the principal cusp is broadly triangular and apicodistally directed. Distal cusplets decrease slowly and regularly in size and their tips are all directed apicodistally; their mesial edges are strongly convex, and their distal edges are weakly convex. The basal border of the crown is nearly straight, with only a very slight convexity developed beneath the principal cusp. The root is nearly twice as tall as the principal cusp and exhibits a distinct mesial depression. A longitudinal ridge traverses mesiodistally across the lingual face near the top of the root. The labial face of the root is flat, and its basal border is nearly straight and slightly angled to the basal border of the crown such that the root height decreases distally.
Remarks.—These teeth are assigned to Hexanchus based on their low, mesiodistally elongate form, fine serrations on the mesial cutting edge of the principal cusp, and distally-reducing distal cusplets (Ward 1979). Serrations along the mesial cutting edge are larger in Notorynchus Ayres, 1855 (Ward 1979) than in the specimens described here. Among the many species names ascribed to fossil Hexanchus teeth over the last 150 years, only two are generally still considered valid: H. microdon Agassiz, 1843, and H. agassizi Cappetta, 1976 (Adolfssen and Ward 2014, 2015). Adolfssen and Ward (2014) note that there are no appreciable anatomical differences among teeth of these two species and that they are normally used for Hexanchus material of different ages: H. microdon occurs in the Maastrichtian through the Danian whereas H. agassizi is restricted to the Eocene. Although the Danian age of the four specimens described here is hence more consistent with H. microdon, we refrain from assigning them to a particular species at this time because multiple authors note that tooth morphology in modern Hexanchus is known to vary ontogenetically, by gender, among individuals, and within the jaw (= monognathic heterodonty), and that, because of this, fossil species within the genus are in need of revision (Adnet 2006; Adolfssen and Ward 2014, 2015).
Hexanchus sp. has previously been reported from the Paleocene–Eocene Vincentown, Manasquan, and Shark River formations in New Jersey (Gallagher 1993) and from the MFL at the EFPQ by Gallagher (Gallagher et al. 1986; Gallagher 1993); though his more recent faunal lists for the MFL have not included this taxon (perhaps by accidental omission in Gallagher 2002 and 2003?). Case (1996) also described several teeth he assigned to Hexanchus sp. and H. microdon from the lower Hornerstown Formation in “Deep Run” in Monmouth County, NJ. Recovery of at least one specimen from above the MFL extends Gallagher’s (1993) stratigraphic range for Hexanchus sp. in the EFPQ beyond the MFL into the upper Hornerstown Formation.
Genus Notidanodon Cappetta, 1975
Type species: Notidanus pectinatus Agassiz, 1843; Late Cretaceous, Sussex, England.
Notidanodon brotzeni (Siverson, 1995)
Fig. 2E, F.
Material.—Three lower lateral teeth: two (RU-EFP-00244-1, 03586) from the MFL and one (RU-EFP-04141) collected as float. All from the Maastrichtian–Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The principal cusp and distal cusplets are weakly convex both labially and lingually. In RU-EFP-04141 and 00244-1, the preserved principal cusp is broad, only slightly taller than the first distal cusplet, and apicodistally directed. It has a convex mesial edge and slightly concave distal edge. Mesial cusplets vary in number, with three present in the RU-EFP-03586, seven in RU-EFP-00244-1, and eight in the RU-EFP-04141. They are usually apicodistally directed, though the mesial-most mesial cusplets of the RU-EFP-00244-1 and 04141 are directed apically. The distal edges of both the mesial and distal cusplets are concave. Distal cusplets decrease slowly and regularly in size, and the basal border of the crown is distinctly convex beneath the principal cusp. The nearly complete root of RU-EFP-03586 is mesiodistally wide, decreases in height distally, and would have been roughly two to three times as tall as the principal cusp if it were complete. A pronounced horizontal ridge traverses the lingual face of the root of both RU-EFP-03586 and 04141, roughly one-third of the way down from the basal border of the crown. Apicobasally oriented ridges are present on the labial face of the root and are roughly positioned such that they descend beneath the notches between each cusp of the crown. A mesial depression is present in the root of both as well.
Remarks.—The combination of distal cusplets that regularly decrease in size from a modestly large acrocone that exhibits multiple, distinct, large cusplets along its mesial edge identify these specimens as belonging to the genus Notidanodon (Cappetta 1987; Siverson 1995). As in all known species except N. loozi Vincent, 1876, the mesial cusplets are apicodistally directed (Ward 1979; Ward and Thies 1987; Siverson 1995). The distal cutting edges of the principal cusp and mesial cusplets are slightly concave, similar to those of N. brotzeni, N. pectinatus Agassiz, 1843, and N. dentatus Woodward, 1886, but in contrast to those of N. loozi and N. lanceolatus Woodward, 1886, which are convex (Ward 1979; Cione and Medina 1987; Siverson 1995). Also as in N. brotzeni, the basal border of the crown in these specimens is distinctly convex beneath the principal cusp; this differs from N. dentatus and N. pectinatus which exhibit a flatter border (Ward and Thies 1987; Siverson 1995). N. dentatus and N. loozi also generally tend to exhibit fewer and larger distal cusplets (Ward 1979; Cione and Medina 1987) than in the specimens described here. Similar to N. brotzeni and N. loozi, the mesial margins of the principal cusp and distal cusplets are convex, whereas in N. lanceolatus they are nearly straight (Ward and Thies 1987; Siverson 1995). The proportionately tall root seen in our specimens compares best with the tall roots of N. brotzeni and N. loozi (Siverson 1995; Cappetta 1987; Ward and Thies 1987).
To summarize, among known species of Notidanodon, the EFPQ specimens share the most similarities with N. brotzeni. These features include apicodistally directed mesial cusplets, slightly concave distal edges of the principal cusp and mesial cusplets, a convex basal borderline of the crown, and convex mesial margins of the principal cusp and distal cusplets (Siverson 1995). We therefore assign the specimens to N. brotzeni, and further note that N. brotzeni is also the only one of the five known species reported from the Danian, the same age as the majority of the Hornerstown Formation at the EFPQ.
Notidanodon brotzeni was previously only known from the middle Danian of Sweden and Denmark (Siverson 1995), with a second possible occurrence later in the Paleocene of New Zealand (Mannering and Hiller 2008). The three specimens described here represent the first occurrence of N. brotzeni in the Western Hemisphere. Further, occurrence of two of the new specimens in the MFL demonstrates that N. brotzeni existed in the very beginning of the Danian and most likely evolved in the Maastrichtian, if not earlier.
Stratigraphic and geographic range.—Lower Paleocene (Danian) of New Jersey in USA, Denmark, Sweden, and Morocco.
Genus Weltonia Ward, 1979
Type species: Notidanus (Heptranchias) ancistrodon Arambourg, 1952, Thanetian, Ouled Abdoun basin, Morocco.
Weltonia ancistrodon (Arambourg, 1952)
Fig. 2G.
Material.—One lower lateral tooth (RU-EFP-04142) from either the upper Danian portion of the Hornerstown Formation or the Thanetian Vincentown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The principal cusp is elongate, slender, distally directed, and sigmoidal. It is over four times taller than the first distal cusplet. The lingual face of the principal cusp is strongly convex whereas its labial face is slightly convex. A single, subtle protuberance (“serration”) occurs near the base of the mesial edge of the principal cusp. Four distal cusplets are present, which are distally directed, slightly labially inclined, and decrease regularly in size. The root is trapezoidal, lessening in height distally, with a distinct mesial depression. Its labial face is flat whereas a distinct mesiodistal ridge runs along the upper portion of the lingual face, creating a slightly concave region below it.
Remarks.—Possession of an extremely tall principal cusp (≥50% taller than the first distal cusplet) that is slender and sigmoidally curved identifies this specimen as Weltonia (Ward 1979; Cappetta 1987). The extremely elongate nature of the principal cusp (over four times as tall as the first distal cusplet) is consistent with the condition in W. ancistrodon, whereas in the only other known species, W. burnhamensis Ward, 1979, the principal cusp is only about 50% taller than the first distal cusplet (Ward 1979).
Case (1996) reported eight specimens of Weltonia ancistrodon recovered from an exposure of the lower Hornerstown Formation in “Deep Run”, a tributary of Crosswicks Creek in Monmouth County, New Jersey. This was the first report of W. ancistrodon from North America. The new specimen described here also likely derives from the Danian portion of the upper Hornerstown Formation and represents a new addition to the fauna of the EFPQ.
Stratigraphic and geographic range.—Lower Paleocene (Danian) to lower Eocene (Ypresian) of New Jersey in USA, England, and Morocco.
Lamniformes Berg, 1958
Pseudocoracidae Cappetta, 2012
Genus Pseudocorax Priem, 1897
Type species: Corax affinis Münster in Agassiz, 1843, Maastrichtian, Netherlands.
Pseudocorax affinis (Münster in Agassiz, 1843)
Fig. 2H.
Material.—Six isolated teeth (SOM: table 1): five (RU-EFP- 02463, 2832, 4158, 4166, 4167) from the MFL and one (RU-EFP-04168) collected as float. All from the Maastrichtian–Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Table 1. Comprehensive list of the invertebrate and vertebrate fauna recovered from Jean and Ric Edelman Fossil Park, sorted by geologic Formation and fossil-rich horizons. Revised from prior lists by Gallagher (2002, 2003) to include more recent additions from previous studies (e.g., Schein et al. 2011; Brochu et al. 2012; Callahan et al. 2012), taxonomic revisions and/or recombinations, and synonymizations over the last 20 years. New additions to the vertebrate fauna identified herein are denoted by an asterisk (*) after their name. Based on recent discoveries, a few additions to the invertebrate and ichnotaxa lists are also included (denoted by a superscript diamond after their name, ◊); these are being formally described in a separate manuscript currently in preparation. Abbreviation: MFL, Main Fossiliferous Layer.
|
Taxon |
Navesink Formation |
Main Fossiliferous Layer (MFL) |
Hornerstown Formation |
Vincentown Formation |
|
Porifera |
Cliona cretacea |
Cliona cretacea |
Peronidella dichotoma |
|
|
Cnidaria |
|
Flabellum mortoni |
Flabellum mortoni |
|
|
Brachiopoda |
Choristothyris plicata |
Terebratulina atlantica |
Oleneothyris harlani Oleneothyris cf. manasquani Terebratulina atlantica◊ |
Oleneothyris harlani |
|
Bryozoa |
Indet. encrusting bryozoans |
|
|
Indet. encrusting bryozoans |
|
Bivalvia |
Agerostrea nasuta Crassatellites vadosus Cucullaea antrosa Cucullaea neglecta Cucullaea vulgaris Exogyra costata Glycymeris mortoni Gryphaeostrea vomer Liopistha protexta Lithophaga ripleyana Pachycardium spillmani Pycnodonte convexa Solyma cf. lineolatus Spondylus echinata Trigonia mortoni |
Cardium tenuistriatum Crassatellites vadosus Cucullaea vulgaris Cuspidaria sp.◊ Etea delawarensis Gervilliopsis ensiformis Glycymeris mortoni Gryphaeostrea vomer Linearia sp.◊ Lithophaga ripleyana Nuculana stephensoni Panopea decisa Pycnodonte dissimilaris Veniella conradi |
Caryatis veta Clavipholas sp.◊ Crassatellites cf. littoralis Cucullaea macrodonta Ostrea glandiformis Veniella conradi |
Cyprimeria densata Polorthus tibialis Pycnodonte sp. Venericardia antiqua |
|
Gastropoda |
Anchura cf. abrupt Anchura pennata Euspira halli Gyrodes petrosus Pleurotomaria sp.◊ Pyrifusus macfarlandi Turbinopsis curta Turritella cf. vertebroides Volutomorpha ponderosa |
Acteon cretacea Anchura abrupt Avellana sp.◊ Cylichna sp.◊ Euspira halli Gyrodes abyssinus Pleurotomaria sp.◊ Pyropsis trochiformis Turbinella parva Turbinella subconical Turritella vertebroides Volutoderma ovata |
cf. Volutocorbis sp. |
|
|
Nautiloidea |
Eutrephoceras dekayi |
Eutrephoceras dekayi Hercoglossa sp.◊ |
cf. Aturia sp. |
|
|
Ammonoidea |
Baculites ovatus Discoscaphites conradi |
Baculites sp. Discoscaphites iris Pachydiscus mokotibensis Sphenodiscus lobatus |
|
|
|
Echinodermata |
Hemiaster sp. |
|
|
|
|
Crustacea |
|
Hoploparia sp. |
|
|
|
Chondrichthyes |
Heptranchias howellii* Odontaspis sp. Palaeogaleus vincenti* Squalicorax pristodontus |
Cretolamna appendiculata Edaphodon mirificus Edaphodon stenobyrus Heptranchias howellii* Hexanchus sp. Ischyodus bifurcatus* Ischyodus thurmanni Myliobatis cf. leidyi Notidanodon brotzeni* Odontaspis cuspidata Palaeogaleus vincenti* Pseudocorax affinis* |
Edaphodon agassizi Hexanchus sp.* Odontaspis sp. Otodus obliquus Palaeocarcharodon sp. Palaeogaleus vincenti* Squalus sp.* Weltonia ancistrodon* |
Lamnidae gen. et sp. indet. |
|
Taxon |
Navesink Formation |
Main Fossiliferous Layer (MFL) |
Hornerstown Formation |
Vincentown Formation |
|
Chondrichthyes |
|
Rhinoptera sp. Rhombodus laevis Sphenodus lundgreni Squalicorax pristodontus Squalus sp.* Squatina sp. Batomorpha gen. et sp. indet. |
|
|
|
Osteichthyes |
Paralbula marylandica* Anomoeodus phaseolus Enchodus ferox Dercetidae gen. et sp. indet.* Osteichthyes gen. et sp. indet. |
Acipenser cf. albertensis Anomoeodus phaseolus* Atractosteus sp.* cf. Bananogmius sp. Enchodus ferox Enchodus gladiolus* Paralbula marylandica Phyllodus paulkatoi* Dercetidae gen. et sp. indet.* Teleostei gen. et sp. indet. |
Anomoeodus phaseolus* Atractosteus sp.* Paralbula marylandica* Saurocephalus lanciformis* Dercetidae gen. et sp. indet.* |
|
|
Testudines |
Cheloniidae gen. et sp. indet. Peritresius ornatus |
Adocus beatus Agomphus turgidus Bothremys sp. Euclastes wielandi Peritresius ornatus Taphrosphys sulcatus |
Euclastes wielandi Testudines gen. et sp. indet. |
|
|
Crocodylia |
|
Borealosuchus threeensis Bottosaurus harlani cf. Procaimanoidea sp. Hyposaurus rogersii Thoracosaurus neocesariensis |
Hyposaurus rogersii Thoracosaurus neocesariensis |
|
|
Mosasauridae |
Halisaurus platyspondylus Mosasaurus hoffmanni Prognathodon rapax |
Mosasaurus hoffmanni* Plioplatecarpus sp. |
|
|
|
Dinosauria |
Hadrosaurus minor Hadrosauridae gen. et sp. indet. |
Hadrosauridae gen. et sp. indet. |
|
|
|
Aves |
|
Graculavis velox Novacaesareala hungerfordi Palaeotringa littoralis Telmatornis priscus Tithostonyx glauconiticus |
|
|
|
Plantae |
|
Plantae gen. et sp. indet. |
|
|
|
Ichnotaxa |
Thalassinoides isp. |
Gnathichnus◊ Radulichnus◊ Rhizocorallium isp.◊ Thalassinoides isp. Xylophagella irregularis |
Thalassinoides isp. |
|
Description.—All of the teeth are small, measuring less than 13 mm in mesiodistal length at the base of the crown. The crown is labiolingually thin and moderately inclined posteriorly with a flat labial face and weakly-convex lingual face. The base of the crown extends into distinct, obliquely-oriented mesial and distal heels, with the rounded distal heel being nearly detached from the central cusp whereas the mesial heel forms a comparatively-weaker convex berm. The mesial heel possesses a weakly-pointed apex in specimens RU-EFP-04167 and 4168. Fine serrations are present along the entire mesial and distal margins of the central cusp and both heels. The root is tall, generally of approximately the same height as the central cusp, with large mesial and distal lobes that are divided by a broadly-triangular basal notch. A shallow nutrient groove bisects a weak, yet distinct, lingual protuberance. Subtle, rounded ridges extend just beneath the crown-root boundary from this protuberance to the mesial and distal margins of the root. These margins each exhibit a concavity just below the crown-root boundary.
Remarks.—The specimens can be differentiated from Squalicorax Whitley, 1939, by their gracile morphology and presence of a nutrient groove (Hamm and Cicimurri 2011). The specimens can be assigned to the genus Pseudocorax based on their possession of a weakly-inclined and triangular central cusp that is flanked mesially and distally by oblique heels, and a large root with weakly-separated lobes that have slightly-concave mesial and distal edges (Jambura et al. 2021). Among the six currently-recognized species of Pseudocorax, only Pseudocorax affinis possesses serrations (Hamm and Shimada 2007). The presence of a nutrient groove on the lingual side of the root is characteristic of Pseudocorax affinis (Case et al. 2017).
The specimens described here represent the first recovery of Pseudocorax from the MFL. Case and Cappetta (2004) reported two teeth of Pseudocorax affinis from the lower Maastrichtian Navesink Formation at Big Brook in Marlboro, NJ. “Pseudocorax affinis” has also been reported from the Campanian of Georgia (Case and Schwimmer 1988) and Alabama (Applegate 1970), though due to their lack of serrations, those specimens should be assigned to Pseudocorax laevis Leriche, 1906 (Shimada 2009). Teeth of the species Pseudocorax laevis (possibly including Pseudocorax granti Cappetta & Case, 1975, Hamm and Shimada 2007) have also been described from the Campanian Marshalltown and Mount Laurel formations near the Chesapeake and Delaware Canal area in Delaware (Lauginger and Hartstein 1983). Pseudocorax teeth have been reported from the Paleocene only once before: Eaton et al. (1989) described several specimens from the Shotgun Member of the Fort Union Formation of Wyoming, although they were interpreted as being reworked from Cretaceous deposits.
Stratigraphic and geographic range.—Upper Cretaceous (Maastrichtian) to lower Paleocene (Danian) of New Jersey in USA, Denmark, Netherlands, and Africa.
Squaliformes Goodrich, 1909
Squalidae Bonaparte, 1834
Genus Squalus Linnaeus, 1758
Type species: Squalus acanthias Linnaeus, 1758, Recent, temperate waters, worldwide.
Squalus sp.
Fig. 2I, J.
Material.—One lateral tooth (RU-EFP-00157-7) from the MFL; eight partial squalid fin spines including one (RU-EFP-00267-1) from the lower Hornerstown Formation below the MFL, two (RU-EFP-02582 and 3942) from the MFL, one (RU-EFP-03567) from the lower Hornerstown Formation above the MFL, and four (RU-EFP-00157-8, 0157-9, 0267, and 4146) collected as float. All from the Maastrichtian–Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA. Because the tooth can be assigned to Squalus, we tentatively assign the fin spines to this genus as well.
Description.—The single cusp is broad, low, labiolingually narrow, and strongly turned distally. Weak serrations are present along its mesial cutting edge. The distal heel is proportionately short, with a tall, rounded profile, and is separated from the cusp by an acute notch. A distinct, tongue-shaped apron of the crown extends below the basal border of the root from the center of the labial face. The sides of the apron are parallel, and its distal tip is circular. The mediolingual uvula of the crown is large and forms a rounded, linguodistally directed apex. The root is short and approximately the same breadth as the crown, with a well-developed basal lobe descending from its midpoint nearly to the apex of the apron. A pronounced horizontal shelf traverses the lingual face of the root. It is perforated beneath the uvula by a subcircular, moderately-broad infundibulum. The basal face of the root is shallowly concave and only slightly turned lingually.
Remarks.—Among Squalinae, the tooth can best be assigned to Squalus based on the basal face being only slightly lingually turned, presence of a modestly broad infundibulum, and a long mediolabial apron on the crown with parallel sides (Cappetta 1987; Adolfssen and Ward 2014). Although teeth of Centrophoroides Davis, 1887, and Centrosqualus Signeux, 1950, are very similar in form (Adolfssen and Ward 2014), the basal face is more strongly turned lingually in Centrophoroides and the apron in Centrosqualus is more bulbous in form owing to narrowing of its base (Müller and Schöllmann 1989). The apron is comparatively broader and more triangular in Protosqualus Cappetta, 1977, and Megasqualus Herman, 1982 (Cappetta 1987). Teeth of Deania Jordan & Snyder, 1902, and Centrophorus Müller & Henle, 1837, lack the projection of the apron beneath the basal plane of the root (Cappetta 1987). While absence of well-developed serrations suggests the specimen is not S. serriculus Jordan & Hannibal, 1923, or S. crenatidens Arambourg, 1952 (Cappetta 1987), other features used to diagnose species within the genus are known to vary in form ontogenetically (Adolfssen and Ward 2014), individually (Cappetta et al. 2016), and by gender (Siverson 1993). Thus, we refrain from assigning the specimen to any particular species at this time.
To our knowledge, among Squalus species only S. minor Daimeries, 1888, has been reported in Paleocene deposits in northeastern North America. This species is reported from the Danian Brightseat Formation in Maryland (Ward and Wiest 1990) and from the lower (Danian) portion of the Hornerstown Formation in Monmouth County, New Jersey (Case 1996). The teeth described by Case (1996) thus derive from the same stratigraphic range as our tooth. However, Case (1996) did not provide a differential diagnosis among Squalus species; therefore, we view his species assignment as inconclusive and do not follow it; instead assigning RU-EFP-00157-7 to Squalus sp. This specimen derives from the MFL, placing it in the very earliest Danian, and the fin spines tentatively inferred to also pertain to Squalus include a specimen from beneath the MFL in the Hornerstown Formation, indicating squalid sharks crossed the K/Pg at this site.
Synechodontiformes Duffin & Ward, 1993
Orthacodontidae Glikman, 1957
Genus Sphenodus Agassiz, 1843
Type species: Lamna longidens Agassiz, 1843, Late Jurassic, Switzerland.
Sphenodus lundgreni (Davis, 1890)
Fig. 2K.
Material.—Two teeth: one (RU-EFP-02913) from the MFL and another (RU-EFP-04159) collected as float. All from Fossil Park, Mantua Township, New Jersey, USA.
Description.—RU-EFP-02913 is essentially complete, measuring 30.2 mm in maximum height, with a root that is 25.5 mm in width and 12.3 mm in length. The crown is nearly vertically oriented and weakly sigmoidal in mesial and distal views. The crown is smooth, with the lingual surface being more convex than the labial surface. Two sharp carinae extend from the crown to form lateral heels on the root. The carinae thin towards the tooth tip and at the base of the crown. Numerous folds are present along the crown-root junction, with enamel ridges extending onto the root. Folds are more tightly spaced and shorter on the lingual side of the tooth. The root is short with a nearly flat basal surface. Abundant nutrient foramina are present, especially on the lingual surface, as well as distinct labiolingually-oriented grooves on its basal surface.
Remarks.—The teeth can be confidently assigned to Sphenodus due to their possession of a straight, sharp central cusp, lateral heels of each carina extending onto the lingual surface of the root, a more lingually convex crown (in cross section), and shallow, basally-flat root with labiolingually-oriented vascularization (Cusumano et al. 2021). Among the currently-recognized species of Sphenodus, S. lundgreni is the only species known from the Maastrichtian (Adolfssen and Ward 2014) and Danian (Cappetta 2012). The presence of numerous folds across the crown-root junction further supports assignment to this species (Kanno et al. 2017).
Callahan et al. (2012) reported the first known occurrence of the synechodontiform shark S. lundgreni, a single lateral tooth (NJSM 23223), from the EFPQ, which was also the first report of this taxon from North America. However, NJSM 23223 was not found in situ, thus its exact stratigraphic provenance within the Hornerstown Formation remains uncertain. Recovery of RU-EFP-02913 constrains the occurrence of this taxon at least to the MFL. We also report the second and third known occurrences of this taxon from the EFPQ, and thus from North America.
Stratigraphic and geographic range.—Upper Cretaceous (Maastrichtian) to Paleocene (Thanetian) of New Jersey in USA, Denmark, Sweden, UK, and Kazakhstan.
Osteichthyes Huxley, 1880
Actinopterygii Klein, 1885
Teleostei Müller, 1845
Aulopiformes Rosen, 1973
Dercetidae Pictet, 1850
Dercetidae gen. et sp. indet.
Fig. 3A, B.
Material.—Twenty-eight dercetid flank scutes (SOM 1: table 1): one (RU-EFP-00282-1) from the upper Navesink Formation, two (RU-EFP-00269-1 and 0277-1) from the Hornerstown Formation below the MFL, ten (including RU-EFP-00228-1, 02490, and 03667) from the MFL, and seven from the Hornerstown Formation above the MFL; the precise stratigraphic origin of the remaining eight within the Hornerstown Formation is uncertain. All from the Maastrichtian Navesink and Maastrichtian–Danian Hornerstown formations, Edelman Fossil Park, Mantua Township, New Jersey, USA. Please see SOM 1: table 1 for the remaining specimen numbers.
Description.—Scutes vary from tripartite to cordiform in shape with a pronounced, elevated median crest along the central axis. Where preserved, the anterior extension is always longer than the two posterior extensions. The largest specimen (RU-EFP-02490) collected from the MFL, is cordiform in shape with large, distinct, randomly arranged tubercles ornamenting its external surface. The second largest specimen (RU-EFP-03667), also from the MFL, exhibits similar ornamentation but is tripartite in shape with a less pronounced median crest. Surface ornamentation is variably developed among the smaller specimens, with some scutes exhibiting well-developed, minute tubercles (occasionally oriented in radial rows) whereas others are essentially smooth. The median crest terminates in a posterolaterally-directed spine in many of the scutes, and its keel ranges from sharp to rounded in profile; this variation does not appear to correlate with scute size. The posterior extensions diverge from near the posterior end of the median crest and each possess a distinct ridge along the posterior margin. In lateral view, these two ridges range from straight to posteriorly concave in orientation. These ridges rotate to become parallel behind the median keel in one scute, but in most specimens the posterior extensions diverge to form an acute or right angle. The inner surface is smooth and occasionally exhibits a shallow, triangular hollow beneath the posterior half of the median crest.
Remarks.—The tripartite and cordiform shapes of these scutes with a distinct median crest identify them as belonging to a dercetid fish (Chalifa 1989; Figueiredo and Gallo 2006; Friedman 2012; Taverne and Goolaerts 2015). Ornamentation on many of the scutes with large or minute tubercles occasionally aligned in radial rows is similar to some dercetids (e.g., Brazilodercetis longirostris Figueiredo & Gallo, 2006) but differs from others in the family which are smooth (e.g., Apuliadercetis indeherbergei Taverne & Goolaerts, 2015; Rhynchodercetis gracilis Chalifa, 1989; Dercetoides venator Chalifa, 1989).
Members of the family Dercetidae are known from the Late Cretaceous and early Paleocene around the world (Silva and Gallo 2011), though it appears only a few species survived the K/Pg mass extinction (Adolfssen et al. 2017). The only previous report of dercetid material from the northeastern United States was by Oman et al. (2016) who described similarly-sized, small, tripartite and cordiform scutes of an indeterminate dercetid from the Campanian Woodbury Formation in Cherry Hill, New Jersey. Recovery of dercetid scutes from the Navesink and Hornerstown formations in the EFPQ extends the known temporal range of these fish in New Jersey through the end of the Cretaceous and into the early Danian. The indeterminate dercetid represented by these fossils also represents a new addition to the fauna.
Enchodontidae Lydekker, 1889
Genus Enchodus Agassiz, 1835
Type species: Esox lewesiensis Mantell, 1822, Turonian, Sussex, England.
Enchodus gladiolus (Cope, 1872)
Fig. 3F, G.
Material.—Eight specimens: two (RU-EFP-00277-2 and 4156) from the lower Hornerstown Formation below the MFL, four (RU-EFP-01916, 2188, 2956, 3698) from the MFL, and two (RU-EFP-04157-1, 4157-2) collected as float. All from the Maastrichtian-Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The teeth are sigmoidal and range in apicobasal height 6–14 mm. Longitudinal striations are present on the rounded posterior face of the crown which extend from the base of the crown to just beneath a pronounced post-apical barb. A longitudinal carina is present along the mesial margin and extends the entire height of the crown. There is subtle variation in overall morphology among the specimens, with some being narrow and elongate whereas others are comparatively shorter and mesiodistally wider. Teeth of the latter morphology have an ovoid-shaped basal cross section, whereas those of the former possess a shallow depression on one side of the basal portion of the crown, making their basal cross section crescentic. This variation may reflect differing tooth positions (i.e., palatine versus mandibular crowns).

Fig. 3. Exemplar actinopterygian scutes (A, B) and teeth (C–I) recovered from the Cretaceous–Paleogene lower Hornerstown Formation at the Jean and Ric Edelman Fossil Park in Mantua Township, New Jersey, USA. A, B. Aulopiform teleost Dercetidae gen. et sp. indet., RU-EFP-02490 (A) and RU-EFP-00228-1 (B), in dorsal (A1, B1) and lateral (A2, B2) views. C. Phyllodontid elopiform Phyllodus paulkatoi Estes & Hiatt, 1978 (RU-EFP-04165-1) in occlusal (C1) and lateral (C2) views. D. Phyllodontid elopiform Paralbula marylandica Blake, 1940 (RU-EFP-00228-2) in occlusal (D1) and lateral (D2) views. E. Saurodontid ichthyodectiform Saurocephalus lanciformis Harlan, 1824 (RU-EFP-04151) in labial (E1) and lingual (E2) views. F, G. Enchodontid aulopiform Enchodus gladiolus (Cope, 1872). F. RU-EFP-04157-1 in lateral view. G. RU-EFP-02188 in lateral (G1) and basal (G1) views. H. Lepisosteid lepisosteiform Atractosteus sp. (RU-EFP-02939) in labial (H1) and mesial or distal (H2) views. I. Pycnodontid pycnodontiform Anomoeodus phaseolus (Hay, 1899) (RU-EFP-02858) in lateral view.
Remarks.—The sigmoidal, fang-like character of the teeth is characteristic of aulopiform actinopterygians, especially of the genus Enchodus (Kriwet 2003; Kriwet et al. 2006). Enchodus is a diverse genus of fish known from the early Cretaceous through the Paleocene. Approximately 30 taxa have been recognized, with five currently-valid species known from North America (Goody 1976; Fielitz 1997; Holloway et al. 2017). The specimens described here are best assignable to the species E. gladiolus due to their possession of a post-apical barb, full-length mesial carina, and striations on the basal portion of the distal face (Goody 1976; Parris et al. 2007; Becker et al. 2010; Alvarado-Ortega et al. 2020; Gouric-Cavalli et al. 2021).
The only species of Enchodus previously reported from the MFL was E. ferox Leidy, 1855 (Miller 1955; Gallagher 1993, 2003), although elsewhere in New Jersey E. gladiolus has been reported from the basal lag of the Navesink Formation at Holmdel Park and in cut bank exposures along Ramanessin Brook (Callahan et al. 2014). Here, we report the first occurrence of E. gladiolus from the MFL, indicating it survived in the western Atlantic until the K/Pg mass extinction.
Stratigraphic and geographic range.—Upper Cretaceous (Coniacian to Maastrichtian) of Arkansas, Colorado, Iowa, Kansas, Nebraska, New Jersey, New Mexico, and South Dakota in USA, Argentina, Canada, Mexico, and Russia.
Elopiformes Sauvage, 1875
Phyllodontidae Sauvage, 1875
Paralbulinae Estes, 1969
Genus Paralbula Blake, 1940
Type species: Paralbula marylandica Blake, 1940, Eocene, Maryland, USA.
Paralbula marylandica Blake, 1940
Fig. 3D.
Material.—Twenty-eight teeth (for repository numbers see SOM 1: table 1): five from the lower Hornerstown Formation below the MFL, seven from the MFL, six from the lower Hornerstown Formation above the MFL, and ten collected as float. All from the Maastrichtian-Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The teeth are roughly circular in occlusal view and hemispherical in form. The enamel of each tooth is smooth, and many exhibit fairly thin enamel structure when viewed in profile. A basilar foramen is present in several complete specimens. A few specimens consist of only the enamel cap, with the underlying dentine broken away/not preserved. One specimen consists solely of a dentine core. All specimens range ~0.5–2 mm in diameter.
Remarks.—The hemispherical shape of the teeth, in combination with possession of a basilar foramen (in some specimens), identify them as pertaining to a paralbuline fish (Estes 1969). Their round shape in occlusal view is more consistent with Paralbula than Pseudoegertonia, the only other genus within Paralbulinae Estes, 1969. Further, the smooth, unornamented enamel of these teeth is consistent with Paralbula marylandica Blake, 1940, but differs from the condition in the only other known Paralbula species from North America, Paralbula casei Estes, 1969, which exhibits intricate and often radiating sculpturing of the enamel surface (Estes 1969).
As all other previously-described specimens of this taxon derive from the Paleocene and Eocene (Blake 1940; Estes 1969; Ebersole et al. 2019; Schein et al. 2011), our recovery of teeth in Hornerstown sediments beneath the MFL constitutes a range extension for Paralbula marylandica back into the latest Maastrichtian, indicating it survived the K/Pg mass extinction.
Stratigraphic and geographic range.—Upper Cretaceous (Maastrichtian) to lower Eocene (Lutetian) of Alabama, Arkansas, Maryland, New Jersey, Texas, and Virginia in USA.
Phyllodontinae Darteville & Casier, 1943
Genus Phyllodus Agassiz, 1843
Type species: Phyllodus toliapicus Agassiz, 1844, Ypresian, England.
Phyllodus paulkatoi Estes & Hiatt, 1978
Fig. 3C.
Material.—One incomplete tooth plate (RU-EFP-04165-1), partially encased in concretion, found as float, and an isolated tooth (RU-EFP-04165-2) from the Hornerstown Formation. The presence of mature, bright green glauconite within the concretion matrix suggests that the tooth plate almost certainly derives from the Hornerstown Formation. All from the Maastrichtian–Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—All teeth within the partial tooth plate are circular in occlusal view, tightly packed, and phyllodont. The teeth are superimposed in directly successional stacks, being up to three deep (as preserved). Each tooth is ~3 mm in diameter, very flat, and thin when viewed in profile. Two of the “highest” exposed crowns exhibit a finely punctate occlusal surface texture, whereas four of the other crowns exposed slightly deeper within the tooth plate exhibit intricate, radially-oriented ridges which anastomose with one another, indicating they were each covered by another tooth which is not preserved. RU-EFP-04165-2 exhibits the same round outline in occlusal view and flat shape in profile as those in the partial tooth plate; its occlusal surface is covered by similarly faint, radially-oriented, anastomosing ridges, indicating that it is an unerupted replacement tooth.
Remarks.—Stacking of teeth in direct succession identifies RU-EFP-04165-1 as pertaining to a phyllodontine fish, differentiating it from paralbulines (Estes 1969). The very flat, thin shape of the teeth in profile is more consistent with the genus Phyllodus than Egertonia (the only two genera within Phylodontinae; Halliday et al. 2016), the latter of which tends to exhibit more domed teeth with thicker enamel (Estes 1969). Among the two currently-recognized species of Phyllodus, the consistently-round shape of all tooth stacks and degree of punctate sculpturing on the occlusal surface in the tooth plate are both consistent with Phyllodus paulkatoi. In contrast, the teeth of Phyllodus toliapicus Agassiz, 1839, are slightly smoother and both the basibranchial and parasphenoid tooth plates possess transversely-elongate teeth in their central rows (Estes and Hiatt 1978).
Phyllodus paulkatoi was previously only known from the Maastrichtian and Danian of Montana (Estes and Hiatt 1978) and the Maastrichtian of Wyoming (Brinkman et al. 2021). Recovery of two specimens from the Hornerstown Formation at Edelman Fossil Park extends its geographic range to the eastern coast of North America at this time (Maastrichtian–Danian).
Stratigraphic and geographic range.—Upper Cretaceous (Maastrichtian) to Paleocene (Danian) of Montana, New Jersey, and Wyoming in USA.
Ichthyodectiformes Bardack & Sprinkle, 1969
Saurodontidae Cope, 1871
Genus Saurocephalus Harlan, 1824
Type species: Saurocephalus lanciformis Harlan, 1824, Campanian, Kansas, USA.
Saurocephalus lanciformis Harlan, 1824
Fig. 3E.
Material.—One tooth (RU-EFP-04151) from the lower Hornerstown Formation above the MFL, Danian, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The crown is triangular and strongly laterally compressed, giving it a flat, spatulate form. Its tip is directed apically and forms a roughly 55° angle in labial view. A sharp, unserrated carina is present on both cutting edges of the crown. The base of the crown, which is slightly wider than the root, curves smoothly and evenly to join it. The root is straight, hollow, ~2.5 times as tall as wide, gradually tapers toward its base, and exhibits a faint, longitudinal groove on both flat sides; these grooves give the root an hourglass cross section when viewed basally.
Remarks.—The broad, laterally-compressed shape of the crown and length of the root identify this tooth as belonging to a saurodontid fish (Bardack and Sprinkle 1969; Stewart 1999). Among the three genera of saurodontids, the flat, broad character of the crown is most similar to that of Saurocephalus (Hays 1830). In Saurodon and Prosaurodon, the teeth are more acutely lanciform (Hays 1830; Stewart 1999). Additionally, the teeth of Prosaurodon are inclined anteriorly and exhibit a more ovoid shape in cross section (Stewart 1999; Everhart 2005), and the crown in Saurodon leanus Hays, 1830, appears more cuspate/curved (cf. Hays 1830: pl. 16: 7, 8). Of the two known species of Saurocephalus, only S. lanciformis has been described from North America (Friedman 2012). The only other species, S. woodwardii Davies, 1878, known exclusively from Europe (Netherlands and Belgium), is based on disparate geographic occurrences compared to known specimens of S. lanciformis rather than an anatomical diagnosis. For this reason, Friedman (2012) noted that S. woodwardii may be a synonym of S. lanciformis and that detailed taxonomic review of specimens assigned to the genus is needed.
Fossils of Saurocephalus lanciformis have been recovered from Campanian strata in the Western Interior and Gulf Coast of the United States (Friedman 2012; Irwin and Fielitz 2013). The only previous report of a saurodontid from New Jersey was the description of a partial skull of Saurodon leanus collected from the Navesink Formation in Pensauken Creek near Moorestown (Hays 1830; Fowler 1911). The tooth described here constitutes the first record of Saurocephalus from northeastern North America and the first record of the genus in the Danian, thus extending the geographic range of S. lanciformis (from the Campanian) and demonstrating that it survived briefly into the early Paleocene.
Stratigraphic and geographic range.—Upper Cretaceous (Campanian) to lower Paleocene (Danian) of Arkansas, Kansas, and New Jersey in USA, and Russia.
Lepisosteiformes Hay, 1926
Lepisosteidae Cuvier, 1825
Genus Atractosteus Rafinesque, 1820
Type species: Atractosteus spatula Lacépède, 1803, Recent, southern USA.
Atractosteus sp.
Fig. 3H.
Material.—Eleven teeth (SOM 1: table 1): two (RU-EFP- 00265-2, 268-2) from the lower Hornerstown Formation below the MFL; five (RU-EFP-00159-3, 2761, 2939, 2970, 4068) from the MFL; one (RU-EFP-00234-1) from the lower Hornerstown Formation above the MFL; and three (RU-EFP-00157-23, 3374, 4152) collected as float. All from the Maastrichtian–Danian Hornerstown Formation, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The teeth are apicobasally tall and exhibit the plicidentine structure unique to lepisosteids (Grande 2010): a circular cross section with longitudinal striae/grooves present along the lower half of the dentine tooth base. The enameled crown is restricted to the tip of the tooth and comprises 10% or less of the apicobasal height; its ventral border is broadly concave in labial and lingual views. The crown is lanceolate in form, being labiolingually flattened with distinct, unserrated carinae along the mesial and distal edges that gently curve inward to meet the dentine at their bases. This morphology gives the impression of a slight constriction beneath the crown, but it is actually due to broadening of the basal crown rather than narrowing of the subjacent dentine. The tip of the crown forms a rounded, obtuse angle in most specimens, but one exhibits a semicircular profile without an angled apex.
Remarks.—Plicidentine tooth structure and enamel restricted to the tip of the crown identify these teeth as belonging to a lepisosteid gar (Grande 2010). Among lepisosteids, only Lepisosteus Lacépède, 1803, and Atractosteus possess tall, fang-like teeth (Grande 2010). Though teeth of these genera are morphologically similar in most respects, Atractosteus possesses more lanceolate, blade-like crowns with a slight constriction beneath the enamel tip (Sigé et al. 1997; Szabó et al. 2016); whereas the enamel tip in Lepisosteus is rounder and lacks such constriction (Grande 2010). We therefore refer the teeth described here to Atractosteus. At present, no features of the teeth are known to be diagnostic at the species level, so we refer the specimens to Atractosteus sp.
Fossils of gars have previously been described from the Cretaceous of New Jersey, including remains of Lepisosteus collected from the Campanian Marshalltown, Wenonah, and Mount Laurel formations (Gallagher 1993) and Atractosteus material from the Marshalltown Formation at Ellisdale (Denton et al. 1996). Scales of an indeterminate lepisosteid have also been recovered from the basal portion of the Maastrichtian Navesink Formation at Holmdel Park in Monmouth County (Callahan et al. 2014). The six teeth described here thus constitute the first gar fossils from the Hornerstown Formation and include the first records of Atractosteus in the Paleocene (Danian) in New Jersey and a new addition to the fauna of Edelman Fossil Park.
Pycnodontiformes Berg, 1940
Pycnodontidae Agassiz, 1833
Genus Anomoeodus Forir, 1887
Type species: Anomoeodus subclavatus Agassiz, 1834, Maastrichtian, Netherlands.
Anomoeodus phaseolus (Hay, 1899)
Fig. 3I.
Material.—Eight teeth: four branchial teeth (RU-EFP- 00159-4, 2474, 2858, 4153), one prearticular/vomerine tooth (RU-EFP-03620) from the MFL, one branchial tooth (RU-EFP-04154) from the lower Hornerstown Formation above the MFL. One branchial tooth (RU-EFP-04155-2) and one prearticular/vomerine tooth (RU-EFP-04155-1) were collected as float. All from Danian, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The prearticular/vomerine teeth are low crowned with a smooth occlusal surface. They are reniform in shape and have a concave base. The larger prearticular/vomerine tooth (RU-EFP-03620) is considerably more elongate that the smaller specimen and measures 7.3 mm in length by 3.3 mm in maximum width. In occlusal view, one end of this tooth is more acute whereas the other is more rounded.
The six branchial teeth are mediolaterally compressed with an apically or apicodistally-directed dorsal hook on the crown. These teeth vary considerably in size, development of a ventral crown prominence (sensu Kriwet 1999), and degree of curvature of the dorsal hook. The largest specimen, recovered from the MFL, has the largest ventral prominence and its dorsal hook forms the most acute angle with the base of the crown (approximately 40°). In contrast, the smallest teeth have a poorly developed ventral prominence and a short, nearly apically-directed dorsal hook. It therefore appears that the ventral prominence may grow in size and the dorsal hook may lengthen and rotate to curve more acutely through ontogeny. In the two specimens retaining their root, the root gradually tapers toward the jaw in the large specimen but exhibits nearly parallel mesial and distal margins in the smaller specimen. The crown-root border is rounded; in lateral view, it appears as a quarter-circle that is horizontally oriented near the distal margin and curves dorsally toward the mesial margin.
Remarks.—Identification of isolated, laterally compressed, hooked teeth has been a subject of debate and confusion for over a century, with teeth of this form previously being assigned to Pycnodontidae, Sclerodontidae, Semionotidae, Amiidae, and Trigonodontidae (Case and Schwimmer 1988; Kriwet 1999, and references therein). Traditionally, many such specimens were assigned to a tetraodontiform, Stephanodus (for a historical review see Kriwet 1999). However, studies of fish skulls in the last few decades have resolved the identity of such teeth as branchial (or “pharyngeal”) teeth of pycnodont fish (Thurmond and Jones 1981; Kriwet 1999). Given concurrent reports of “Stephanodus” (pycnodont branchial teeth) and Anomoeodus (palatine/vomerine teeth) from multiple formations in New Jersey (see below) and Gallagher’s (2003) listing of only Anomoeodus phaseolus at Edelman Fossil Park, we provisionally assign the branchial teeth as belonging to this species.
The smooth, reniform shape of the palatine/vomerine teeth is consistent with the genus Anomoeodus (cf. Case and Schwimmer 1988; Bazzi et al. 2015). Possession of a concave base identifies these teeth as pertaining to the species Anomoeodus phaseolus (Case and Schwimmer 1988), and this assignment is circumstantially supported by their occurrence in North America where this species was known to have been widespread (see below).
Gallagher (2003) previously reported Anomoeodus phaseolus from the Navesink Formation at EFPQ, and Gallagher (1993) reported recovery of branchial teeth of this taxon from multiple other Cretaceous formations across New Jersey, including the Marshalltown, Wenonah, and Mount Laurel formations (as Stephanodus sp.). Fowler (1911) also described a partial jaw from an indeterminate Cretaceous formation from Crosswicks Creek in Burlington County (as Pycnodus phaseolus), and Baird and Case (1966) reported recovery of branchial and prearticular/vomerine teeth of Anomoeodus from the Navesink Formation in Burlington County (with the branchial teeth again listed as Stephanodus). Thus, the new specimens described here include the first report of Anomoeodus phaseolus from the Hornerstown Formation and the Danian; all previous reports of this taxon across North America derive from Campanian and Maastrichtian deposits (references above, and Bazzi et al. 2015).
As only a single, small, worn specimen has been found above the MFL, it may be that this specimen was reworked up section by bioturbation and that the more abundant occurrences of Anomoeodus teeth in the MFL represent the actual latest occurrence of the species (i.e., A. phaseolus only briefly survived into the very earliest Danian, as recorded by the MFL death assemblage).
Stratigraphic and geographic range.—Upper Cretaceous (Coniacian) to lower Paleocene (Danian) of Delaware, Georgia, Maryland, Mississippi, New Jersey, North Carolina, and South Carolina in USA.
Sarcopterygii Romer, 1955
Reptilia Laurenti, 1768
Crocodilia Owen, 1842
Alligatoridae Cuvier, 1807
Genus Bottosaurus Agassiz, 1849
Type species: Crocodilus harlani von Meyer, 1832, Maastrichtian and Danian, Fieldsboro, New Jersey, USA.
Bottosaurus harlani (von Meyer, 1832)
Fig. 4A–C.
Material.—A relatively large, isolated posterior tooth (RU-EFP-02450), a left juvenile dentary (RU-EFP-03820), and two small partial crowns (RU-EFP-03876, 4169) from the MFL. All from Danian, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—RU-EFP-02450 is large, measuring 56.8 mm in height and 24.0 mm in mesiodistal width. The crown itself is short and blunt, measuring only 12.7 mm in height. The tooth is strongly labiolingually compressed, making the crown subrectangular in cross section. The enamel is strongly folded and exhibits abundant apical striations, giving the crown a wrinkled appearance. A modest, smooth carina is present on the mesial and distal edges of the crown. This keel extends to the base of the crown and would likely have extended over the tip of the tooth, which is worn. Enamel extends slightly further down the root on the lingual side than the labial side.
The left dentary (RU-EFP-03820) is significantly smaller than any previously described specimen of the genus, measuring approximately 58.2 mm (Fig. 4C). Its mandibular symphysis is intact, but its rostral and posterior regions are damaged, making the exact number of alveoli difficult to determine. The specimen possesses either 11 or 12 alveoli with the largest being the most-posteriorly positioned. Based on the morphology of the articulation facet for the splenial, the splenial possessed an anterior tip that passed dorsal to the Meckelian groove, but did not contribute to the mandibular symphysis. The mandibular symphysis extends posteriorly to about the fourth or fifth alveolus, and the tooth row forms a gentle, medially-concave curve between the ?fourth and ?tenth alveolus.
Two small isolated teeth (RU-EFP-03876, 4169) were also recovered within relatively close proximity (~4 m) to the dentary. Each is incomplete, consisting of most of the crown and a small portion of the root (e.g., Fig. 4B). They are similar to RU-EFP-02450 in being labiolingually compressed, possessing wrinkled enamel with apicobasally-aligned striations, and development of distinct mesial and distal carinae. Their morphology suggests that they are posterior teeth and are of the appropriate size to have derived from the dentary.

Fig. 4. Remains of the alligatorid crocodilian Bottosaurus harlani (von Meyer, 1832) (A–C) and mosasaurid cf. Mosasaurus hoffmannii Mantell, 1829 (D) recovered from the Cretaceous–Paleogene lower Hornerstown Formation at the Jean and Ric Edelman Fossil Park in Mantua Township, New Jersey, USA. A. Posterior tooth (RU-EFP-02450) in labial (A1), lingual (A2), and mesial or distal (A3) views. B. ?Posterior tooth (RU-EFP-03876) in labial (B1), lingual (B2), and occlusal (B3) views. C. Left dentary (RU-EFP-03820) in dorsal (C1), medial (C2), lateral (C3), and ventral (C4) views. D. Right pterygoid (RU-EFP-03592) in medial (D1), ventral (D2), and lateral (D3) views. Abbreviations: av, alveolus; t, tooth.
Remarks.—RU-EFP-02450 differs from durophagous mosasaurs in that the base of the crown is not inflated and by its lack of multiple cusps, as seen in the mosasaur Carinodens (e.g., Schulp et al. 2004). Of the five crocodilians known from the Hornerstown Formation, namely Hyposaurus Owen, 1849, cf. Procaimanoidea Gilmore, 1946, Bottosaurus, Thoracosaurus Leidy, 1852, and Borealosuchus Brochu, 1997 (Gallagher 2003; Brochu et al. 2012), only Bottosaurus possesses similar blunt, crushing-style teeth. In particular, RU-EFP-02450 is assigned to Bottosaurus harlani based on its large size, short and blunt crown, and wrinkled enamel, each of which are characteristic of this taxon (Cossette and Brochu 2018). This specimen is nearly identical to the paralectotype (ANSP 9174) and to the teeth described from a new, partially-articulated specimen of Bottosaurus harlani from the MFL in the EFPQ (Cossette and Brochu 2018).
Although the dentary (RU-EFP-03820) is incomplete, its morphology is inconsistent with that of Borealosuchus because the mandibular symphysis extends posterior to the fourth alveolus and the ninth alveolus is distinctly larger than the eighth. In Borealosuchus, alveoli diameter doesn’t increase until the tenth alveolus. The majority of the complete alveoli in our specimen are slightly labiolingually compressed (~20%), implying that the teeth within them would also have been compressed, consistent with the genus Bottosaurus (Cossette 2021).
Bottosaurus harlani is a rare and enigmatic caimanine known only from the Maastrichtian to early Danian of the Atlantic Coastal Plain (Erickson 1998; Schwimmer et al. 2015; Cossette and Brochu 2018). Bottosaurus harlani has previously been described from the MFL by Gallagher (1993) and Cossette and Brochu (2018), yet the small dentary described here represents the first juvenile remains for this taxon. The morphology of RU-EFP-03820 is strikingly consistent with that of adult specimen NJSM 11265 (Cossette and Brochu 2018: fig. 5), indicating that this alligatorid acquired its mandibular morphology early in life.
Stratigraphic and geographic range.—Upper Cretaceous (Maastrichtian) to lower Paleocene (Danian) of New Jersey, USA.
Squamata Oppel, 1811
Mosasauridae Gervais, 1853
Genus Mosasaurus Coneybeare, 1822
Type species: Mosasaurus hoffmannii Mantell, 1829, Maastrichtian, Maastricht, Netherlands.
cf. Mosasaurus hoffmannii Mantell, 1829
Fig. 4D.
Material.—A nearly-complete right pterygoid (RU-EFP- 03592) from the MFL, nine isolated teeth (RU-EFP-00109, 01826, 01901, 02703, 02916, and 00120-1–00120-4) and seven isolated vertebral centra (RU-EFP-00033, 02645, 03640, and 00043-1–00043-4) of mosasaurids have also been recovered from the MFL; many, if not all, of these specimens likely pertain to this same taxon, but they are too incomplete or lack enough diagnostic features for confident referral. All from Danian, Edelman Fossil Park, Mantua Township, New Jersey, USA.
Description.—The pterygoid is well preserved and represents one of the rarer skeletal elements of a mosasaurid recovered from the EFPQ in the last half century. Its preserved anteroposterior length is ~33 cm. Five erupted and three replacement teeth are present, although the specimen possesses a total of nine alveoli. The enamel of the preserved teeth is generally smooth or faintly faceted. Where these facets are present, they are much less well pronounced than in the marginal teeth of Mosasaurus hoffmannii. Faintly-serrated carinae are present on both the anterior and posterior margins of each tooth. As in other Mosasaurus species, the middle teeth are larger (crown height = 28 mm) than the posteriormost ones (~18 mm). As described for the holotype of M. hoffmannii by Street and Caldwell (2017), the tooth row is spindle shaped in ventral view with a rather flat parapet on the medial side and a bowed outwards parapet laterally. The basisphenoid and quadratic rami are each thin, with the quadratic ramus also being dorsomedially concave.
Remarks.—Several mosasaurs are known from Cretaceous strata in New Jersey, including the large-bodied taxa Mosasaurus conodon Cope, 1881, M. hoffmannii, and Prognathodon rapax Hay, 1902 (see Gallagher 2005, 2015; Gallagher et al. 2012, for full taxa lists). Of these, M. cf. hoffmannii is known from the MFL whereas M. conodon and Prognathodon rapax are known from the underlying Navesink and Mount Laurel formations (Gallagher 2005; Gallagher et al. 2012). RU-EFP-03592 differs from Prognathodon in having teeth with distinct facets (Dortangs et al. 2002) and both anterior and posterior carinae (Konishi et al. 2011). It also differs from M. conodon in having serrated carinae (Ikejiri and Lucas 2015). The overall size and morphology of the pterygoid is consistent with that of M. hoffmannii; although the only diagnostic feature reported for this species is the presence of eight teeth in the pterygoid (Street and Caldwell 2017). However, other mosasaur genera have eight pterygoid teeth as well (e.g., M. conodon), and tooth count can vary within mosasaurid genera or species (Ikejiri and Lucas 2015). Given this uncertainty, we conservatively identify RU-EFP-03592 as belonging to cf. M. hoffmannii.
Mosasaur material is relatively rare in the MFL, being more common and more complete in the underlying formations (Gallagher 2005, 2015; Gallagher et al. 2012). Within the MFL, typical finds include isolated teeth and worn centra. Thus, the recovery of a relatively complete and well-preserved pterygoid with teeth represents an exceptional discovery. These better-preserved specimens (RU-EFP-03592 and 00033, a large anterior ?caudal retaining portions of the transverse processes and neural spine) also suggest that mosasaur material in the MFL is not reworked (contra Gallagher 1993, 2005).
Stratigraphic and geographic range.—Upper Cretaceous (Campanian to Maastricthtian) of Maryland, Missouri, New Jersey, South Dakota, Tennessee, and Texas in USA, Angola, Belgium, Brazil, Bulgaria, Canada, Denmark, Germany, Italy, Japan, Jordan, Morocco, Netherlands, New Zealand, Niger, Poland, Russia, South Africa, Spain, Syria, and Turkey.
Discussion
Faunal insights.—Ichthyofaunal assemblages spanning the K/Pg boundary are rare (Cavin 2001; Becker and Chamberlain 2012; Argyriou and Davesne 2021), as are surface outcrops of the K/Pg boundary along the Atlantic and Gulf Coastal Plains. This is especially true along the northeastern seaboard where the boundary has often become deeply buried by sediments derived from ongoing erosion of the Appalachian Mountains since the Mesozoic (e.g., Olsson et al. 1997; Miller et al. 1999). Former excavations by the Inversand Company in Mantua Township, New Jersey, allowed geologists and paleontologists to circumvent this challenge for nearly a century. This property, now the Jean and Ric Edelman Fossil Park and Museum, continues to afford a regionally-unrivaled window into the Maastrichtian–Thanetian strata and fossil faunas of the western Atlantic, making it one of the most informative K/Pg boundary sites along the Atlantic Coastal Plain (Gallagher 1993).
Prior studies of the fossilized faunas preserved in the EFPQ (e.g., Gallagher 1993, 2003) identified a diverse suite of 67 taxa within the MFL of the lower Hornerstown Formation, as well as 42 taxa within the underlying Navesink Formation and 19 taxa within overlying sediments of the Hornerstown Formation above the MFL (Table 1). Our study of vertebrate microfossils recovered from the EFPQ adds 11 taxa to the known faunal list for this K/Pg locality (Table 1): Palaeogaleus vincenti, Heptranchias howellii, Notidanodon brotzeni, Weltonia ancistrodon, Pseudocorax affinis, Squalus sp., Dercetidae indet., Enchodus gladiolus, Phyllodus paulkatoi, Saurocephalus lanciformis, and Atractosteus sp. The remains of the majority of these new taxa were recovered from the MFL but as shown in Table 1, several new species occurrences were also identified in the Navesink Formation and Hornerstown Formation above the MFL. Each of the newly-identified taxa are generally compatible with prior paleoecological interpretations by Gallagher (1993), Wiest et al. (2016), and Vellekoop et al. (2016) that the Hornerstown Formation records deposition on the mid-to-outer shelf under warm/subtropical conditions.
Geographic and temporal range extensions.—Taken as a whole, identification of these new microfossils reveals important new insights into patterns of vertebrate survivorship and extinction across the K/Pg boundary, as well as temporal and geographic range extensions for several chondrichthyan and actinopterygian taxa (Fig. 5). Of primary significance, we have identified the first Paleocene (Danian) record of Saurocephalus and first Cretaceous (Maastrichtian) records of Paralbula marylandica and Palaeogaleus vincenti, demonstrating for the first time that these taxa survived the K/Pg mass extinction. Our recovery of dercetid scutes from both the Navesink and Hornerstown formations (including above the MFL) reveals that these predatory needlefish also survived the mass extinction in the western Atlantic (contra Cavin 2001). Further, we have identified the first record of gars in the Hornerstown Formation, including the first Paleocene (Danian) occurrences of Atractosteus in New Jersey, as well as the first records of Enchodus gladiolus, Ischyodus bifurcatus, Anomoeodus phaseolus, and Pseudocorax affinis from the MFL (which represent the youngest occurrences of these taxa in the region). These latter discoveries indicate that those four taxa survived all the way through the Maastrichtian in the western Atlantic, ultimately perishing during the collapse of the local ecosystem in the earliest Danian following the Chicxulub impact (cf. Obasi et al. 2011; Esmeray-Senlet et al. 2017). Three geographic range extensions were also identified in this study, namely the first records of Notidanodon brotzeni from the Western Hemisphere, Saurocephalus from northeastern North America, and Phyllodus paulkatoi along the eastern coast of North America. Finally, the dentary of Bottosaurus we describe from the MFL indicates that the mandible of this poorly-known alligatorid likely grew isometrically through ontogeny (cf. Cossette and Brochu 2018).
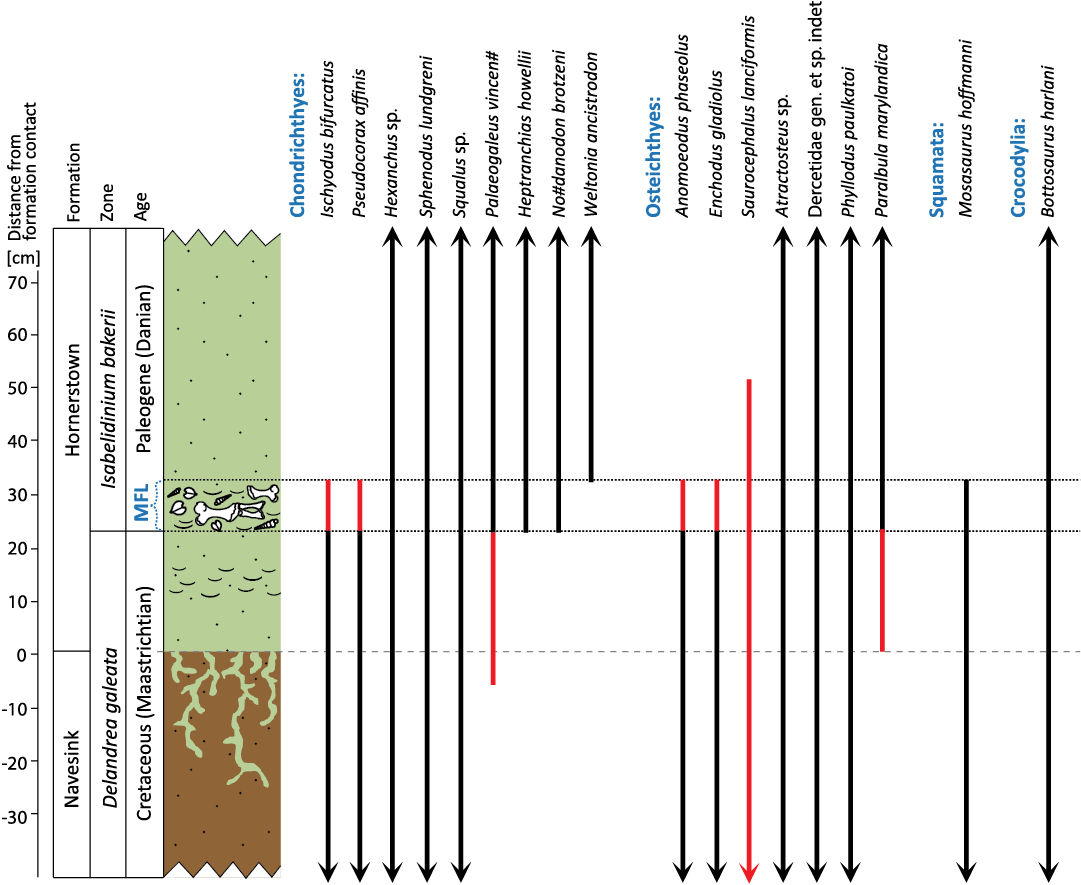
Fig. 5. Stratigraphic occurrences of each taxon discussed in this study, with new range extensions noted in red. The range extensions shown are based on the global record of each taxon, and thus represent global temporal range extensions (i.e., they are not merely extensions in the record solely at Edelman Fossil Park). Placement and thickness of the Main Fossiliferous Layer (MFL) and “oyster layer” beneath it within the lower Hornerstown Formation are based on the findings of Voegele et al. (2021), and assignment of the dinoflagellate zones is based on Koch and Olsson (1977) and Aurisano (1989).
These novel insights highlight the importance of microfossils for expounding knowledge of past faunas and illuminating the character of ancient ecosystems (Sankey and Baszio 2008). Moreover, given the wealth of microfossil specimens recovered in our recent projects in the EFPQ (e.g., Boles 2016; Voegele et al. 2021), it is almost certain that further microsieving of sediments of each of the formations examined herein, as well as from other unconsolidated Cretaceous and Paleocene formations across the Atlantic and Gulf Coastal Plains, would yield a wealth of microfossil specimens stemming from as-yet unrecognized taxa (and potentially taxa new to science). For instance, despite over a century of study of the Cretaceous–Paleocene faunas of New Jersey (e.g., Cope 1869), to this day new vertebrate taxa continue to be named based on newly-discovered macrofossils from these deposits (e.g., Borealosuchus threeensis Brochu et al., 2012, Iridopristis parrisi Andrews et al., 2023). We therefore infer that this study likely only elucidates a portion of the “hidden” microfauna of the EFPQ and predict that further microsieving studies here and elsewhere across the region will continue to yield similar insights.
Comparisons.—The late Maastrichtian–early Danian ichthyofauna of the EFPQ is broadly similar to those of many other K/Pg boundary localities across the Atlantic and Gulf Coastal Plains, and from deposits of the Western Interior Seaway (e.g., Becker et al. 2010, and references therein). Further, as at Stevns Klint in Denmark, hexanchids were common at EFPQ and chondrichthyan macrofossils are more abundant than those of osteichthyans (Adolfssen and Ward 2015; Adolfssen et al. 2017). Aulopiforms (e.g., Enchodus) were especially abundant during the Cretaceous at EFPQ, as in the Pindos Unit in Greece (Argyriou and Davesne 2021); and as in the vertebrate record of the López de Bertodano Formation of Seymour Island, Antarctica (Cione et al. 2018), there is no apparent decline in fish diversity leading up to the K/Pg at EFPQ.
Our findings also generally agree with the growing consensus (Noubhani and Cappetta 1997; Becker et al. 2011; Adolfssen and Ward 2014; Poyato-Ariza and Martín-Abad 2016; Guinot and Condamine 2023) that both osteichthyans and chondrichthyans were significantly impacted by the K/Pg extinction. For instance, at the EFPQ 50% of actinopterygian (4 of 8) and 71% of chondrichthyan genera (10 of 14) disappear at the top of the MFL (Table 1). These extinctions were selective, with our results from the EFPQ showing strong correspondence with those of other studies that crocodilians (Jouve and Jalil 2020), teleosts (Poyato-Ariza and Martín-Abad 2016), durophagous actinopterygians (e.g., Paralbula, Anomoeodus), gars (e.g., Atractosteus), hexanchids (e.g., Hexanchus), and squalids (e.g., Squalus) weathered the extinction well while durophagous chondrichthyans (e.g., edaphodontids), rhombodontids (e.g., Rhombodus), and large, fast, open-water predators (e.g., enchodontids and anacoracids, such as Enchodus and Squalicorax) went extinct (Cavin 2001; Kriwet and Benton 2004; Friedman 2009; Friedman and Sallan 2012; Adolfssen et al. 2017; Schwarzhans and Stringer 2020; Guinot and Condamine 2023). Perhaps due to the decimation of invertebrates, rays also seem to disappear above the MFL at the EFPQ; this finding matches global trends of severe declines for myliobatiforms in the immediate aftermath of the Chicxulub impact (Kriwet and Benton 2004; Guinot and Condamine 2023).
The comparative rarity of fish fossils in the Hornerstown Formation above the MFL implies that recovery of elasmobranchs and fish after the extinction was slow. The recovery of chondrichthyans, especially sharks, in New Jersey appears to have occurred more slowly than in Denmark (cf. Adolfssen and Ward 2014) but faster than along the Gulf Coastal Plain in Arkansas (cf. Maisch et al. 2020), perhaps reflecting its intermediate distance from the Chicxulub impact site compared to these other localities. Collectively, these trends align well with slow recovery patterns inferred from ichthyoliths in the southern Atlantic Ocean (Sibert et al. 2014) and based on studies of the global fossil record of Cretaceous–Paleogene fish (Friedman 2009; Guinot and Condamine 2023).
Macro- and microfossils collected from above the MFL at the EFPQ in this study, and by Gallagher (1993), indicate that the local ichthyofauna during the initial phase of recovery after the Chicxulub impact was dominated by durophagous fish and hexanchid and lamniform sharks. This is evident by sediments of the Hornerstown Formation above the MFL yielding a relative abundance of teeth of Odontaspis, Otodus (personal observations by ZMB and PVU), and Paralbula (this study), as well as by hexanchids and lamniforms representing five of the eight (63%) known chondrichthyans in the upper Hornerstown Formation (Table 1). Such abundance of odontaspid and otodontid teeth differs from the early Danian ichthyofaunas of Antarctica, where these sharks appear to have been rare (Cione et al. 2018). However, similar recovery patterns have been noted in the Paleocene Clayton Limestone of Arkansas, where scarce food may have favored the survival of durophagous taxa (Becker and Chamberlain 2012), and in Danian strata across Denmark, where cooler waters stemming from the impact winter may have provided suitable conditions for sharks adapted to cool, deep settings (Adolfssen and Ward 2015; Adolfssen et al. 2017).
Finally, in agreement with global trends inferred by Bazzi et al. (2018) and Guinot and Condamine (2023), there is no significant shift in shark tooth shape disparity across the K/Pg at the EFPQ. Wide-ranging Cretaceous chondrichthyan genera (4 of the 11, or 36%) present at EFPQ survived the K/Pg slightly better than endemic taxa (1 of the 5, or 20%), and lamniforms diversified in its aftermath (i.e., two new genera, Otodus and Palaeocarcharodon, are first observed in the Hornerstown Formation above the MFL; Table 1). However, unlike these studies, at the EFPQ there are no signs of carcharhiniform sharks rising to dominance following the K/Pg (cf. Bazzi et al. 2018) nor of differential survivorship by short-lived (37%) versus long-lived (33%) shark genera (Table 1; cf. Guinot and Condamine 2023). These contrasts highlight the value of detailed studies of individual sites, like this one, toward characterizing how regional geography, climate, and ecological relationships interact to control survivorship and recovery patterns through mass extinction events.
Acknowledgements
Many of the specimens described herein could not have been collected without the support of staff of the Inversand Mining Company, who graciously allowed site access and assisted with overburden removal while they owned the site. We also thank Jean and Ric Edelman for continued support of the Fossil Park which bears their name, as well as the Economic Development Committee of Mantua Township for their efforts to preserve the site in perpetuity prior to its purchase by Rowan University in 2015. We further extend gratitude to Ned Gilmore (Academy of Natural Sciences of Drexel University, Philadelphia, Pennsylvania, USA) for opening their collections for comparative studies. We would like to sincerely thank our dedicated volunteers and especially Arya Valcarcel (Gwynedd Valley, Pennsylvania, USA) for discovering and donating a Sphenodus lundgreni tooth. Finally, we would like to thank Kerin Claeson (Philadelphia College of Osteopathic Medicine, Philadelphia, Pennsylvania, USA) and an anonymous reviewer for their constructive feedback. This research was supported by Rowan University.
References
Adnet, S. 2006. Biometric analysis of the teeth of fossil and Recent hexanchid sharks and its taxonomic implications. Acta Palaeontologia Polonica 51: 477–488.
Adolfssen, J.S. and Ward, D.J. 2014. Crossing the boundary: an elasmobranch fauna from Stevns Klint, Denmark. Palaeontology 57: 591–629. Crossref
Adolfssen, J.S. and Ward, D.J. 2015. Neoselachians from the Danian (early Paleocene) of Denmark. Acta Palaeontologia Polonica 60: 313–338.
Adolfssen, J.S., Milàn, J., and Friedman, M. 2017. Review of the Danian vertebrate fauna of southern Scandinavia. Bulletin of the Geological Society of Denmark 65: 1–23. Crossref
Agassiz, L. 1839. Recherches sur les poissons fossiles, Vol. 2. 336 pp. Imprimerie de Patitpierre, Neuchatel.
Agassiz, L. 1843. Recherches sur les poissons fossils, Vol. 3. 422 pp. Imprimerie de Patitpierre, Neuchatel.
Alvarado-Ortega, J., Cantalice, K.M., Díaz-Cruz, J.A., Castañeda-Posadas, C., and Zavaleta-Villareal, V. 2020. Vertebrate fossils from the San José de Gracia quarry, a new Late Cretaceous marine fossil site in Puebla, Mexico. Boletín de la Sociedad Geológica Mexicana 72: e160819. Crossref
Andrews, J.V., Schein, J.P., and Friedman, M. 2023. An earliest Paleocene squirrelfish (Teleostei: Beryciformes: Holocentroidea) and its bearing on the timescale of holocentroid evolution. Journal of Systematic Palaeontology 21: 2168571. Crossref
Applegate, S.P. 1970. The vertebrate fauna of the Selma Formation in Alabama. Part VIII: The fishes. Fieldiana, Geology Memoirs 3: 385–433. Crossref
Applegate, S.P. and Uyeno, T. 1968. The first discovery of a a fossil tooth belonging to the shark genus Heptranchias, with a new Pristiophorus spine, both from the Oligocene of Japan. Bulletin of the National Science Museum, Tokyo 11 (2): 195–200.
Arambourg, C. 1952. Les vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). Notes et Mémoires du Service Géologique du Maroc 92: 1–372.
Argyriou, T. and Davesne, D. 2021. Offshore marine actinopterygian assemblages from the Maastrichtian–Paleogene of the Pindos Unit in Eurytania, Greece. PeerJ 9: e10676. Crossref
Aurisano, R.W. 1989. Upper Cretaceous dinoflagellate biostratigraphy of the subsurface Atlantic Coastal Plain of New Jersey and Delaware, U.S.A. Palynology 13: 143–179. Crossref
Ayres, W.O. 1855. A shark of a new generic type: Notorynchus maculatus. Proceedings of the California Academy of Natural Sciences 1: 72–73.
Baird, D. and Case, G.R. 1966. Rare marine reptiles from the Cretaceous of New Jersey. Journal of Paleontology 40: 1211–1215.
Bardack, D. and Sprinkle, G. 1969. Morphology and relationships of saurocephalid fishes. Fieldiana Geology 16: 297–340. Crossref
Bazzi, M., Einarsson, E., and Kear, B.P. 2015. Late Cretaceous (Campanian) actinopterygian fishes from the Kristianstad Basin of southern Sweden. In: B.P. Kear, J. Lindgren, J. Hurum, J.H. Milàn, and V. Vajda (eds.), Mesozoic Biotas of Scandinavia and its Arctic Territories, Geological Society of London Special Publications 434: 277–292. Crossref
Bazzi, M., Kear, B.P., Blom, H., Ahlberg, P.E., and Campione, N.E. 2018. Static dental disparity and morphological turnover in sharks across the end-Cretaceous mass extinction. Current Biology 28: 2607–2615. Crossref
Becker, M.A. and Chamberlain, J.A. 2012. Osteichthyans from the Paleocene Clayton Limestone of the Midway Group, Hot Spring County, Arkansas, USA: Bony fish evolution across the Cretaceous–Paleogene Boundary. Paludicola 8 (4): 194-207.
Becker, M.A., Mallery, Jr. , C.S., and Chamberlain, Jr., J.A. 2010. Osteichthyans from an Arkadelphia Formation-Midway Group lag deposit (Late Maastrichtian–Paleocene), Hot Spring County, Arkansas, U.S.A. Journal of Vertebrate Paleontology 30: 1019–1036. Crossref
Blake, S. 1940. Paralbula, a new fossil fish based on dental plates from the Eocene and Miocene of Maryland. Journal of the Washington Academy of Science 30: 205–209.
Boles, Z.M. 2016. Vertebrate Taphonomy and Paleoecology of a Cretaceous–Paleogene Marine Bonebed. 287 pp. Ph.D. Thesis, Drexel University, Philadelphia.
Brinkman, D.B., Divay, J.D., DeMar, Jr., D.G., and Wilson Mantilla, G.P. 2021. A systematic reappraisal and quantitative study of the nonmarine teleost fishes from the late Maastrichtian of the Western Interior of North America: evidence from vertebrate microfossil localities. Canadian Journal of Earth Sciences 58: 936–967. Crossref
Brochu, C.A. 1997. A review of “Leidyosuchus” (Crocodyliformes, Eusuchia) from the Cretaceous through Eocene of North America. Journal of Vertebrate Paleontology 17: 679–697. Crossref
Brochu, C.A., Parris, D.C., Grandstaff, B.S., Denton, Jr., R.K., and Gallagher, W.B. 2012. A new species of Borealosuchus (Crocodyliformes, Eusuchia) from the Late Cretaceous–Early Paleogene of New Jersey. Journal of Vertebrate Paleontology 32: 105–116. Crossref
Buckland, W. 1838. On the discovery of fossil fishes in the Bagshot Sands at Goldworth Hill, 4 miles north of Guilford. Proceedings of the Geological Society of London 2: 687–688.
Callahan, W.R., Mehling, C.M., Denton, R.K., and Parris, D.C. 2014. Vertebrate paleontology and stratigraphy of the Late Cretaceous Holmdel Park Site, Monmouth County, New Jersey. Dakoterra 6: 163–169.
Callahan, W.R., Schein, J.P., Schroeter, E.R., Parris, D.C., and Lacovara, K.J. 2012. First record of the synechodontiform shark Sphenodus (Neoselachii, Orthacodontidae) from the Danian of North America. Program and Abstracts, Journal of Vertebrate Paleontology 32 (Supplement 2): 1–207. Crossref
Cappetta, H. 1976. Sélaciens nouveaux du London Clay de l’Essex (Yprésien du Bassin de Londres). Geobios 9: 551–575. Crossref
Cappetta, H. 1977. Sélaciens nouveaux de l’Albien supérieur de Wissant (Pas-de-Calais). Géobios 10 (6): 967–973. Crossref
Cappetta, H. 1981. Additions à la faune de sélaciens fossils du Maroc. 1: Sur la presence des genres Heptranchias, Alopias et Odontorhytis dans l’Yprésien des Ouled Abdoun. Geobios 14: 563–575. Crossref
Cappetta, H. 1987. Chondrichthyes II: Mesozoic and Cenozoic Elasmobranchii. 193 pp. Verlag, New York.
Cappetta, H. 2012. Chondrichthyes: Mesozoic and Cenozoic Elasmobranchii. Handbook of Palaeoichthyology. Vol. 3E. 512 pp. Verlag, Munich.
Cappetta, H. and Case, G.R. 1975. Sélaciens nouveaux du Crétacé du Texas. Geobios 8: 303–307. Crossref
Cappetta, H., Gregorová, R., and Adnet, S. 2016. New selachian assemblages from the Oligocene of Moravia (Czech Republic). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 280: 259–284. Crossref
Carlsen, A.W. and Cuny, G. 2014. A study of the sharks and rays from the Lillebælt Clay (Early–Middle Eocene) of Denmark, and their palaeoecology. Bulletin of the Geological Society of Denmark 62: 39–88. Crossref
Carrillo-Briceño, J.D., Argyriou, T., Zapata, V., Kindlimann, R., and Jaramillo, C. 2016. A new Early Miocene (Aquitanian) Elasmobranchii assemblage from the La Guajira Peninsula, Colombia. Ameghiniana 53: 77–99. Crossref
Case, G.R. 1973. Fossil Sharks: A Pictorial Review. 64 pp. Pioneer Litho Co. Inc., New York.
Case, G.R. 1978. Ischyodus bifurcatus, a new species of chimaeroid fish from the Upper Cretaceous of New Jersey. Geobios 11: 21–29. Crossref
Case, G.R. 1980. A selachian fauna from the Trent Formation, Lower Miocene (Aquitanian) of eastern North Carolina. Palaeontographica Abteilung A 171: 75–103.
Case, G.R. 1996. A new selachian fauna from the lower Hornerstown Formation (Early Paleocene/Montian) of Monmouth County, New Jersey. Palaeontographica Beiträge zur Naturgeschichte der Vorzeit Abteilung A 242: 1–14. Crossref
Case, G.R. and Cappetta, H. 2004. Additions to the elasmobranch fauna from the late Cretaceous of New Jersey (lower Navesink Formation, early Maastrichtian). Palaeovertebrata 33: 1–16.
Case, G.R. and Schwimmer, D.R. 1988. Late Cretaceous fish from the Blufftown Formation (Campanian) in western Georgia. Journal of Paleontology 62: 290–301. Crossref
Case, G.R. and Schwimmer, D.R. 1992. Occurrence of the chimaeroid Ischyodus bifurcatus Case in the Cusseta Formation (Upper Cretaceous, Campanian) of western Georgia and its distribution. Journal of Paleontology 66: 347–350. Crossref
Case, G.R., Cook, T.D., Sadorf, E.M., and Shannon, K.R. 2017. A late Maastrichtian selachian assemblage from the Peedee Formation of North Carolina, USA. Vertebrate Anatomy Morphology Palaeontology 3: 63–80. Crossref
Cavin, L. 2001. Effects of the Cretaceous–Tertiary boundary event on bony fishes. In: E. Buffetaut and C. Koeberl (eds.), Geological and Biological Effects of Impact Events, 141–158. Springer, Berlin. Crossref
Chalifa, Y. 1989. Two new species of longirostrine fishes from the Early Cenomanian (Late Cretaceous) of Ein-Yabrud, Israel, with comments on the phylogeny of the Dercetidae. Journal of Vertebrate Paleontology 9: 314–328. Crossref
Cicimurri, D.J. 2010. Fossil chimaeroid remains (Chondrichthyes: Holocephali) from Williamsburg County, South Carolina, USA. Paludicola 8: 37–48.
Cicimurri, D.J. and Ebersole, J.A. 2014. Late Cretaceous chimaeroids (Chondrichthyes: Holocephali) from Alabama, USA. PaleoBios 31: 1–14. Crossref
Cifelli, R.L., Madsen, S.K., and Larson, E.M. 1996. Screenwashing and associated techniques for the recovery of microvertebrate fossils. In: R.L. Cifelli (ed.), Techniques for Recovery and Preparation of Microvertebrate Fossils. Oklahoma Geological Survey Special Publication 96-4: 1–24.
Cione, A.L. and Medina, F.A. 1987. A record of Notidanodon pectinatus (Chondrichthyes, Hexanchiformes) in the Upper Cretaceous of the Antarctic Peninsula. Mesozoic Research 1: 79–88.
Cione, A.L., Santillana, S., Gouiric-Cavalli, S., Hospitaleche, C.A., Gelfo, J.N., López, G.M., and Reguero, M. 2018. Before and after the K/Pg extinction in West Antarctica: New marine fish records from Marambio (Seymour) Island. Cretaceous Research 85: 250–265. Crossref
Cope, E.D. 1869. Synopsis of the extinct Reptilia found in the Mesozoic and Tertiary strata of New Jersey. In: G.H. Cook (ed.), Geology of New Jersey, 733–738. New Jersey Geological Survey, Trenton.
Cope, E.D. 1881. A new Clidastes from New Jersey. The American Naturalist 15: 587–588. Crossref
Cossette, A.P. 2021. A new species of Bottosaurus (Alligatoroidea: Caimaninae) from the Black Peaks Formation (Palaeocene) of Texas indicates an early radiation of North American caimanines. Zoological Journal of the Linnean Society 191: 276–301. Crossref
Cossette, A.P. and Brochu, C.A. 2018. A new specimen of the alligatoroid Bottosaurus harlani and the early history of character evolution in alligatorids. Journal of Vertebrate Paleontology 38: 1–22. Crossref
Cusumano, A., D’Arpa, C., and Duffin, C.J. 2021. Taxonomic revision of the shark Sphenodus in the Jurassic collections of the “Museo Geologico G.G. Gemmellaro” (Palermo, Sicily). Bollettino della Società Paleontologica Italiana 60: 169–181.
Cvancara, A.M. and Hoganson, J.W. 1993. Vertebrates of the Cannonball Formation (Paleocene) in North and South Dakota. Journal of Vertebrate Paleontology 13: 1–23. Crossref
Daimeries, A. 1888. Notes ichthyologiques I. Annales de la Société royale malacologique de Belgique, Bulletin des des Séances 23: 42–43.
Davis, J.D. 1887. The fossil fishes of the chalk of Mount Lebanon, in Syria. The Scientific Transactions of the Royal Dublin Society, Series II 3: 457–634.
Denton, R.K., Jr., O’Neill, R.C., Grandstaff, B., and Parris, D.C. 1996. The Ellisdale dinosaur site: An overview of its geology and paleontology. In: W.B. Gallagher and D.C. Parris (eds.), Cenozoic and Mesozoic vertebrate paleontology of the New Jersey coastal plain, 19–34. New Jersey State Museum Investigation 5: 1–58.
Dortangs, R.W., Schulp, A.S., Mulder, E.W.A., Jagt, J.W.M., Peeters, H.H.G., and de Graaf, D.T. 2002. A large new mosasaur from the Upper Cretaceous of the Netherlands. Netherlands Journal of Geosciences 81: 1–8. Crossref
Duffin, C.J. and Reynders, J.P.H. 1995. A fossil chimaeroid from the Gronsveld Member (Late Maastrichtian, Late Cretaceous) of northeast Belgium. Belgian Geological Survey Professional Paper 278: 111–156.
Eaton, J.G., Kirkland, J.I., and Doi, K. 1989. Evidence of reworked Cretaceous fossils and their bearing on the existence of Tertiary dinosaurs. Palaios 4: 281–286. Crossref
Ebersole, J.A., Cicimurri, D.J., and Stringer, G.L. 2019. Taxonomy and biostratigraphy of the elasmobranchs and bony fishes (Chondrichthyes and Osteichthyes) of the lower-to-middle Eocene (Ypresian to Bartonian) Claiborne Group in Alabama, USA, including an analysis of otoliths. European Journal of Taxonomy 585: 1–274. Crossref
Erickson, B.R. 1998. Crocodilians of the Black Mingo Group (Paleocene) of the South Carolina coastal plain. Transactions of the American Philosophical Society 88: 196–214. Crossref
Esmeray-Senlet, S., Miller, K.G., Sherrell, R.M., Senlet, T., Vellekoop, J., and Brinkhuis, H. 2017. Iridium profiles and delivery across the Cretaceous/Paleogene boundary. Earth and Planetary Science Letters 457: 117–126. Crossref
Estes, R. 1969. Studies on fossil phyllodont fishes: interrelationships and evolution in the Phyllodontidae (Albuloidei). Copeia 1969: 317–331. Crossref
Estes, R. and Hiatt, R. 1978. Studies on fossil phyllodont fishes: a new species of Phyllodus (Elopiformes, Albuloidea) from the Late Cretaceous of Montana. PaleoBios 28: 1–12.
Everhart, M.J. 2005. Oceans of Kansas: A Natural History of the Western Interior Sea. 322 pp. Indiana University Press, Bloomington.
Fielitz, C. 1997. A preliminary phylogenetic analysis of the fish genus Enchodus (Teleostei: Aulopiformes). Journal of Vertebrate Paleontology 17 (Supplement 3): 45A–46A.
Figueiredo, F.J. and Gallo, V. 2006. A new dercetid fish (Neoteleostei: Aulopiformes) from the Turonian of the Pelotas Basin, southern Brazil. Palaeontology 49: 445–456. Crossref
Fowler, H.W. 1911. A description of the fossil fish remains of the Cretaceous, Eocene and Miocene formations of New Jersey. Geological Survey of New Jersey Bulletin 4: 1–192. Crossref
Frederickson, J.A., Lipka, T.R., and Cifelli, R.L. 2018. Faunal composition and paleoenvironment of the Arundel Clay (Potomac Formation; Early Cretaceous), Maryland, USA. Palaeontologia Electronica 21.2.31A: 1–24. Crossref
Friedman, M. 2009. Ecomorphological selectivity among marine teleost fishes during the end-Cretaceous extinction. Proceedings of the National Academy of Sciences 106: 5218–5223. Crossref
Friedman, M. 2012. Ray-finned fishes (Osteichthyes, Actinopterygii) from the type Maastrichtian, the Netherlands and Belgium. In: J.W.M. Jagt, S.K. Donovan, and E.A. Jagt-Yazykova (eds.), Fossils of the Type Maastrichtian (Part 1). Scripta Geologica Special Issue 8: 113–142.
Friedman, M. and Sallan, L.C. 2012. Five hundred million years of extinction and recovery: a Phanerozoic survey of large-scale diversity patterns in fishes. Palaeontology 55: 707–742. Crossref
Gallagher, W.B. 1993. The Cretaceous/Tertiary mass extinction in the northern Atlantic coastal plain. The Mosasaur 5: 75–154.
Gallagher, W.B. 2002. Faunal changes across the Cretaceous–Tertiary (K-T) boundary in the Atlantic coastal plain of New Jersey: restructuring the marine community after the K-T mass-extinction event. Geological Society of America Special Paper 356: 291–301. Crossref
Gallagher, W.B. 2003. Oligotrophic oceans and minimalist organisms: collapse of the Maastrichtian marine ecosystem and Paleocene recovery in the Cretaceous–Tertiary sequence of New Jersey. Netherlands Journal of Geosciences 82: 225–231. Crossref
Gallagher, W.B. 2005. Recent mosasaur discoveries from New Jersey and Delaware, USA: stratigraphy, taphonomy and implications for mosasaur extinction. Netherlands Journal of Geosciences 84: 241–245. Crossref
Gallagher, W.B. 2012. Comparative taphonomy of Late Cretaceous vertebrate fossil occurrences in the Atlantic Coastal Plain deposits of Appalachia: testing the hypothesis of mass mortality at the K/Pg boundary. Society of Vertebrate Paleontology Program and Abstracts: 98.
Gallagher, W.B. 2015. Greensand mosasaurs of New Jersey and the Cretaceous–Paleogene transition of marine vertebrates. Netherlands Journal of Geosciences 94: 87–91. Crossref
Gallagher, W.B., Miller, K.G., Sherrell, R.M., Browning, J.V., Field, M.P., Olsson, R.K., Sugarman, P.J., Tuorto, S., and Wahyudi, H. 2012. On the last mosasaurs: Late Maastrichtian mosasaurs and the Cretaceous–Paleogene boundary in New Jersey. Bulletin de la Société Géologique de France 183: 145–150. Crossref
Gallagher, W.B., Parris, D.C., and Spamer, E.E. 1986. Paleontology, biostratigraphy, and depositional environments of the Cretaceous–Tertiary transition in the New Jersey coastal plain. The Mosasaur 3: 1–35.
Geyn. W. van de 1937. Les élasmobranches du Crétacé marin du Limbourg Hollandais. Natuurhistorich Maandblad 26: 16–69.
Gilmore, C.W. 1946. A new crocodilian from the Eocene of Utah. Journal of Paleontology 20: 62–67.
Goody, P.C. 1976. Enchodus (Teleostei: Enchodontidae) from the Upper Cretaceous Pierre Shale of Wyoming and South Dakota with an evaluation of the North American enchodontid species. Palaeontographica, Abteilung A 152: 91–112.
Gouric-Cavalli, S., Cione, A.L., Tineo, D.E., Pérez, L.M., Iribarne, M., Allcca Torres, M.A., and Poiré, D.G. 2021. The first Peruvian record of Enchodus (Actinopterygii, Aulopiformes, Enchodontidae) in the Upper Cretaceous Vivan Formation. Andean Geology 48: 303–315. Crossref
Grande, L. 2010. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy: the resurrection of Holostei. Copeia, Supplement 10 (2A): 1–871.
Guinot, G. and Condamine, F.L. 2023. Global impact and selectivity of the Cretaceous–Paleogene mass extinction among sharks, skates, and rays. Science 379: 802–806. Crossref
Halliday, T.J.D., Cuff, A.R., Prasad, G.V.R., Thanglemmoi, M.S., and Goswami, A. 2016. New record of Egertonia (Elopiformes, Phyllodontidae) from the Late Cretaceous of south India. Papers in Palaeontology 2: 287–294. Crossref
Hamm, S.A. and Cicimurri, D.J. 2011. Early Coniacian (Late Cretaceous) selachian fauna from the basal Atco Formation, lower Austin Group, north central Texas. Paludicola 8: 107–127.
Hamm, S.A. and Shimada, K. 2007. The Late Cretaceous lamniform shark, Pseudocorax laevis, from the Niobrara Chalk in western Kansas. Transactions of the Kansas Academy of Science 110: 44–52.
Hay, O.P. 1902. Bibliography and catalogue of the fossil vertebrata of North America. Bulletin of the United States Geological Survey 179:1–868. Crossref
Hays, I. 1830. Description of a fragment of the head of a new fossil animal, discovered in a marl pit near Moorestown, New Jersey. Transactions of the American Philosophical Society 3: 471–477. Crossref
Heinicke, M.P., Naylor, G.J.P., and Hedges, S.B. 2009. Cartilaginous fishes (Chondrichthyes). In: S.B. Hedges and S. Kumar (eds.), The Timetree of Life, 320–327. Oxford University Press, Oxford. Crossref
Herman, J. 1977. Les sélaciens des terrains néocrétacés et paléocènes de Belgique et des contrées limitrophes. Eléments d’une biostratigraphie intercontinentale. Mémoires pour servir à l’explication des Cartes géologiques et minières de la Belgique. Service Geologique de Belgique 15: 1–401.
Herman, J. 1982. Additions to the fauna of Belgium 6. The Belgian Eocene Squalidae. Tertiary Research 4 (1): 1-6.
Hoganson, J.W. and Erickson, J.M. 2005. A new species of Ischyodus (Chondrichthyes: Holocephali: Callorhynchidae) from Upper Maastrichtian shallow marine facies of the Fox Hills and Hell Creek formations, Williston Basin, North Dakota, USA. Palaeontology 48: 709–721. Crossref
Hoganson, J.W., Erickson, J.M., and Everhart, M.J. 2015. Ischyodus rayhaasi (Chimaeroidei; Callorhynchidae) from the Campanian–Maastrichtian Fox Hills Formation of northeastern Colorado, USA. Transactions of the Kansas Academy of Science 118: 27–40. Crossref
Holloway, W.L., Claeson, K.M., Sallam, H.M., El-Sayed, S., Kora, M., Sertich, J.J.W., and O’Connor, P.M. 2017. A new species of the neopterygian fish Enchodus from the Duwi Formation, Campanian, Late Cretaceous, Western Desert, central Egypt. Acta Palaeontologica Polonica 62: 603–611. Crossref
Horner, R.J., Wiest, L.A., Buynevich, I.V., Terry, Jr. , D.O., and Grandstaff, D.E. 2016. Chemical composition of Thalassinoides boxwork across the marine K-Pg boundary of central New Jersey, U.S.A. Journal of Sedimentary Research 86: 1444–1455. Crossref
Hovestadt, D.C. and Hovestadt-Euler, M. 2002. The remains of a carcharhinid shark with a new triakid species in its digestive tract from the Oligocene of Germany. Tertiary Research 21: 171–182.
Ikejiri, T. and Lucas, S.G. 2015. Osteology and taxonomy of Mosasaurus conodon Cope 1881 from the Late Cretaceous of North America. Netherlands Journal of Geosciences 94: 39–54. Crossref
Irwin, K.J. and Fielitz, C. 2013. Ichthyodectiform fishes from the Late Cretaceous (Campanian) of Arkansas, USA. In: G. Arratia and H.-P. Schultze (eds.), Mesozoic Fishes 5—Global Diversity and Evolution, 247–266. Verlag, Munich.
Jambura, P.L., Stumpf, S., and Kriwet, J. 2021. Skeletal remains of the oldest pseudocoracid shark Pseudocorax kindlimanni sp. nov. (Chondrichthyes, Lamniformes) from the Late Cretaceous of Lebanon. Cretaceous Research 125: 104842. Crossref
Jordan, D.S. and Hannibal, H. 1923. Fossil sharks and rays of the Pacific slope of North America. Bulletin of the Southern California Academy of Sciences 22: 27–63.
Jordan, D.S. and Snyder, J.O. 1902. Descriptions of two new species of squaloid sharks from Japan. Proceedings of the United States National Museum 25: 79–81. Crossref
Jouve, S. and Jalil, N.-E. 2020. Paleocene resurrection of a crocodylomorph taxon: Biotic crises, climatic and sea level fluctuations. Gondwana Research 85: 1–18. Crossref
Kanno, S., Nakajima, Y., Hikida, Y., and Sato, T. 2017. Sphenodus (Chondrichthyes, Neoselachii) from the Upper Cretaceous in Nakagawa Town, Hokkaido, Japan. Paleontological Research 21: 122–130. Crossref
Kirkland, J.I., Eaton, J.G., and Brinkman, D.B. 2013. Elasmobranchs from Upper Cretaceous freshwater facies in southern Utah. In: A.L. Titus and M.A. Loewen (eds.), At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah, 153–194. Indiana University Press, Bloomington.
Koch, R.C. and Olsson, R.K. 1977. Dinoflagellate and planktonic foraminiferal biostratigraphy of the uppermost Cretaceous of New Jersey. Journal of Paleontology 51: 480–491.
Konishi, T., Brinkman, D., Massare, J.A., and Caldwell, M.W. 2011. New exceptional specimens of Prognathodon overtoni (Squamata, Mosasauridae) from the upper Campanian of Alberta, Canada, and the systematics and ecology of the genus. Journal of Vertebrate Paleontology 31 (5): 1026–1046. Crossref
Kriwet, J. 1999. Pycnodont fishes (Neopterygii, Pycnodontiformes) from the Lower Cretaceous of Uña (E-Spain) with comments on branchial teeth in pycnodontid fishes. In: G. Arratia and H.-P. Schultze (eds.), Mesozoic Fishes 2—Systematics and Fossil Record, 215–238. Verlag, Munich.
Kriwet, J. 2003. Lancetfish teeth (Neoteleostei, Alepisauroidei) from the Early Cretaceous of Alcaine, NE Spain. Lethaia 36: 323–332. Crossref
Kriwet, J. and Benton, M.J. 2004. Neoselachian (Chondrichthyes, Elasmobranchii) diversity across the Cretaceous–Tertiary boundary. Palaeogeography, Palaeoclimatology, Palaeoecology 214: 181–194. Crossref
Kriwet, J., Lirio, J.M., Nuñez, H.J., Puceat, E., and Lécuyer, C. 2006. Late Cretaceous Antarctic fish diversity. In: J.E. Francis, D. Pirrie, and J.A. Crame (eds.), Cretaceous–Tertiary High-Latitude Palaeoenvironments, James Ross Basin, Antarctica. Geological Society of London Special Publication 258: 83–100. Crossref
Lauginger, E.M. and Hartstein, E.F. 1983. A guide to fossil sharks, skates, and rays from the Chesapeake and Delaware Canal area, Delaware. Delaware Geological Survey Open File Report 21: 1–64.
Leidy, J. 1852. Descriptions of Delphinus conradi and Thoracosaurus grandis. Proceedings of the Academy of Natural Sciences of Philadelphia 6: 35.
Leidy, J. 1855. Indications of twelve species of fossil fishes. Proceedings of the Academy of Natural Sciences of Philadelphia 7 (10): 395–397.
Leriche, M. 1906. Contribution à lètude des poisons fossils du Nord de la France et des regions voisines. Mémoires de la Société géologique du Nord 5: 1–430.
Leriche, M. 1938. Contribution à l’étude des poissons fossiles des pays riverains de la Méditerranée américaine (Venezuela, Trinité, Antilles, Mexique). Mémoires de la Société Paléontologique Suisse 61 (1): 1–42.
Maisch, H.M., Becker, M.A., and Griffiths, M.L. 2020. Chondrichthyans from the Lower Clayton Limestone Unit of the Midway Group (Paleocene) near Malvern, Arkansas, USA, with comments on the K/Pg boundary. PalZ 94: 561–593. Crossref
Mannering, A.A. and Hiller, N. 2008. An early Cenozoic neoselachian shark fauna from the southwest Pacific. Palaeontology 51: 1341–1365. Crossref
Miller, Jr. , H.W. 1955. A check-list of the Cretaceous and Tertiary vertebrates of New Jersey. Journal of Paleontology 29: 903–914.
Miller, K.G., Sugarman, P.J., Browning, J.V., Cramer, B.S., Olsson, R.K., de Romero, L., Aubry, M.-P., Pekar, S.F., Georgescu, M.D., Metzger, K.T., Monteverde, D.H., Skinner, E.S., Uptegrove, J., Mullikin, L.G., Muller, F.L., Feigenson, M.D., Reilly, T.J., Brenner, G.J., and Queen, D. 1999. Ancora Site. In: K.G. Miller, P.J. Sugarman, J.V Browning, et al. (eds.), Proceedings of the Ocean Drilling Project, Initial Reports, Volume 174AX (Supplement), 1–65. Ocean Drilling Program, College Station.
Minard, J.P., Owens, J.P., Sohl, N.F., Gill, H.E., and Mello, J.F. 1969. Cretaceous–Tertiary boundary in New Jersey, Delaware, and eastern Maryland. Geological Survey Bulletin 1274: H1–H33.
Molina, E., Alegret, L., Arenillas, I., Arz, J.A., Gallala, N., Hardenbol, J., von Salis, K., Sterurbaut, E., Vandenberghe, N., and Zaghbib-Turki, D. 2006. The Global Boundary Stratotype Section and Point for the base of the Danian Stage (Paleocene, Paleogene, “Tertiary”, Cenozoic) at El Kef, Tunisia: original definition and revision. Episodes 29: 263–273. Crossref
Müller, J. and Henle, F.G.J. 1837. Gattungen der Haifische und Rochen, nach ihrer Arbeit: Über die Naturgeschichte der Knorpelfische. Bericht der Königlisch Preussischen Akademie der Wissenschaften zu Berlin 1836: 111–118.
Müller, A. and Schöllmann, L. 1989. Neue selachier (Neoselachii, Squalomorphii) aus dem Campanium Westfalens (NW-Deutschland). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 178: 1–35.
Noubhani, A. and Cappetta, H. 1997. Les Orectolobiformes, Carchariniformes et Myliobatiformes (Elasmobranchii, Neoselachii) des Bassins à phosphate du Maroc (Maastrichtien–Lutâetien basal): systématique, biostratigraphie, évolution et dynamique des faunes. Paleo Ichthyologica 8: 1–327.
Obasi, C.C., Terry, D.O., Myer, G.H., and Grandstaff, D.E. 2011. Glauconite composition and morphology, shocked quartz, and the origin of the Cretaceous(?) main fossiliferous layer (MFL) in southern New Jersey, U.S.A. Journal of Sedimentary Research 81: 479–494. Crossref
Olsson, R.K., Miller, K.G., Browning, J.V, Habib, D., and Sugarman, P.J. 1997. Ejecta layer at the Cretaceous–Tertiary boundary, Bass River, New Jersey (Ocean Drilling Program Leg 174AX). Geology 25: 759–762. Crossref
Olsson, R.K., Miller, K.G., Browning, J.V., Wright, J.D., and Cramer, B.S. 2002. Sequence stratigraphy and sea level change across the Cretaceous–Tertiary boundary on the New Jersey passive margin. Geological Society of America Special Paper 356: 97–108. Crossref
Oman, L.D., Manning, E.M., Badger, R.F., and Whitley, J.B. 2016. A close examination of an aragonitic upper Woodbury Formation fauna. The Mosasaur 9: 83–126.
Owen, R. 1849. Notes on the remains of a fossil reptile discovered by professor Henry Rogers of Pennsylvania, United States, in greensand formations of New Jersey. Proceedings of the Geological Society of London 5: 380–383. Crossref
Parris, D.C., Grandstaff, B.S., and Gallagher, W.B. 2007. Fossil fish from the Pierre Shale Group (Late Cretaceous): Clarifying the biostratigraphic record. In: J.E. Martin and D.C. Parris (eds.), The Geology and Paleontology of the Late Cretaceous marine deposits of the Dakotas. Geological Society of America Special Paper 427: 99–109. Crossref
Peterson, J.E., Scherer, R.P., and Huffman, K.M. 2011. Methods of microvertebrate sampling and their influences on taphonomic interpretations. Palaios 26: 81–88. Crossref
Pictet, F.J. and Campiche, G. 1858. Description des fossiles du terrain crétacé des environs de Sainte-Croix, 1ère partie. Matériaux pour la Paléontologie Suisse 2: 1–380.
Poyato-Ariza, F.J. and Martín-Abad, H. 2016. The Cretaceous in the evolutionary history of the Actinopterygii. In: A. Khosla and S.G. Lucas (eds.), Cretaceous Period: Biotic Diversity and Biogeography. New Mexico Museum of Natural History and Science Bulletin 71: 275–286.
Purdy, R.W., Schneider, V.P., Applegate, S.P., McLellan, J.H., Meyer, R.L., and Slaughter, B.H. 2001. The Neogene sharks, rays, and bony fishes from Lee Creek Mine, Aurora, North Carolina. In: C.E. Ray and D.J. Bohaska (eds.), Geology and Paleontology of the Lee Creek Mine, North Carolina, III, 71–202. Smithsonian Institution Press, Washington. Crossref
Sankey, J.T. and Baszio, S. 2008. Vertebrate Microfossil Assemblages: Their Role in Paleoecology and Paleobiogeography. 296 pp. Indiana University Press, Bloomington.
Schein, J. and Poole, J. 2014. A shark-bitten dinosaur (Hadrosauridae) femur from the latest Maastrichtian basal Hornerstown Formation, New Jersey, U.S.A. The Mosasaur 8: 15–22.
Schein, J., Grandstaff, B.S., Gallagher, W.B., Poole, J.C., and Lacovara, K.J. 2011. Paralbula in North America: revisiting an enigmatic Campanian–Late Paleocene teleost with hope for new insights. Geological Society of America Abstracts with Programs 43: 87.
Schein, J., Lacovara, K.J., Gallagher, W., and Poole, J. 2008. Transgressive lag or condensed section?: taphonomic evidence from the latest Cretaceous–earliest Tertiary basal Hornerstown Formation (New Jersey, U.S.A.). Journal of Vertebrate Paleontology 28 (3 – Supplement): 136A.
Schubert, J.A., Wick, S.L., and Lehman, T.M. 2017. An Upper Cretaceous (middle Campanian) marine chondrichthyan and osteichthyan fauna from the Rattlesnake Mountain sandstone member of the Aguja Formation in West Texas. Cretaceous Research 69: 6–33. Crossref
Schulp, A.S., Jagt, J.W.M., and Fonken, F. 2004. New material of the mosasaur Carinodens belgicus from the Upper Cretaceous of the Netherlands. Journal of Vertebrate Paleontology 24: 744–747. Crossref
Schwarzhans, W. and Stringer, G.L. 2020. Fish otoliths from the late Maastrichtian Kemp Clay (Texas, USA) and the early Danian Clayton Formation (Arkansas, USA) and an assessment of extinction and survival of teleost lineages across the K-Pg boundary based on otoliths. Rivista Italiana di Paleontologia e Stratigrafia 126: 395–446.
Schwimmer, D.R., Sanders, A.E., Erickson, B.R., and Weems, R.E. 2015. A Late Cretaceous dinosaur and reptile assemblage from South Carolina, USA. Transactions of the American Philosophical Society 105: 1–152.
Shimada, K. 2009. The first associated teeth of the Late Cretaceous anacoracid shark, Pseudocorax laevis (Leriche), from the Mooreville Chalk of Alabama. Transactions of the Kansas Academy of Science 112: 164–168. Crossref
Sibert, E.C., Hull, P.M., and Norris, R.D. 2014. Resilience of Pacific pelagic fish across the Cretaceous/Palaeogene mass extinction. Nature Geoscience 7: 667–670. Crossref
Sigé, B., Buscalioni, A.D., Duffaud, S., Gayet, M., Orth, B., Rage, J.-C., and Sanz, J.L. 1997. Etat des données sur le gisement Crétacé supérieur continental de Champ-Garimond (Gard, Sud de la France). Münchner Geowissenschaftlicher Abhandlungen 34: 11–130.
Signeux, J. 1950. Notes Paléoichthyologiques. III: Squalidae fossiles du Sénonien de Sahel-Alma. Bulletin du Muséum National d’Histoire Naturelle (Série 2) 22 (2): 315–319.
Silva, H.M.A and Gallo, V. 2011. Taxonomic review and phylogenetic analysis of Enchodontoidei (Teleostei: Aulopiformes). Anais da Academia Brasileira de Ciências 83: 483–511. Crossref
Siverson, M. 1993. Maastrichtian squaloid sharks from southern Sweden. Palaeontology 36: 1–19.
Siverson, M. 1995. Revision of the Danian cow sharks, sand tiger sharks, and goblin sharks (Hexanchidae, Odontaspididae, and Mitsukurinidae) from southern Sweden. Journal of Vertebrate Paleontology 15: 1–12. Crossref
Stahl, B.J. 1999. Chondrichthyes III: Holocephali. Handbook of Paleoichthyology, Vol. 4. 164 pp. Verlag Dr. Friedrich Pfeil, München.
Stewart, J.D. 1999. A new genus of Saurodontidae (Teleostei: Ichthyodectiformes) from Upper Cretaceous rocks of the Western Interior of North America. In: G. Arratia and H.-P. Schultze (eds.), Mesozoic Fishes 2—Systematics and Fossil Record, 335–360. Verlag, Munich.
Street, H.P. and Caldwell, M.W. 2017. Rediagnosis and redescription of Mosasaurus hoffmannii (Squamata: Mosasauridae) and an assessment of species assigned to the genus Mosasaurus. Geological Magazine 154: 521–557. Crossref
Szabó, M., Gulyás, P., and Ösi, A. 2016. Late Cretaceous (Santonian) Atractosteus (Actinopterygii, Lepidosteidae) remains from Hungary (Iharkút, Bakony Mountains). Cretaceous Research 60: 239–252. Crossref
Taverne, L. and Goolaerts, S. 2015. The dercetid fishes (Teleostei, Aulopiformes) from the Maastrichtian (Late Cretaceous) of Belgium and the Netherlands. Geologica Belgica 18: 21–30.
Thurmond, J.T. and Jones, D.E. 1981. Fossil Vertebrates of Alabama. 244 pp. University of Alabama Press, Tuscaloosa.
Vellekoop, J., Esmeray-Senlet, S., Miller, K.G., Browning, J.V., Sluijs, A., van de Schootbrugge, B., Sinninghe-Damsté, J.S., and Brinkhuis, H. 2016. Evidence for Cretaceous–Paleogene Boundary Bolide “Impact Winter” Condition From New Jersey, USA. Geology 44: 619–622. Crossref
Vicent, G. 1876. Description de la faune de l’étage Landénien inférieur de Belgique. Annales de la Société royale malacologique de Belgique 11: 111–160.
Voegele, K.K., Ullmann, P.V., Lonsdorf, T., Christman, Z., Heierbacher, M., Kibelstis, B.J., Putnam, I., Boles, Z.M., Walsh, S., and Lacovara, K.J. 2021. Microstratigraphic analysis of fossil distribution in the lower Hornerstown and upper Navesink formations at the Edelman Fossil Park, NJ. Frontiers in Earth Science 9: 756655. Crossref
Ward, D.J. 1979. Additions to the fish fauna of the English Palaeogene. 3: A review of the hexanchid sharks with a description of four new species. Tertiary Research 2: 111–129.
Ward, D.J. and Grande, L. 1991. Chimaeroid fish remains from Seymour Island, Antarctic Peninsula. Antarctic Science 3: 323–330. Crossref
Ward, D.J. and Thies, D. 1987. Hexanchid shark teeth (Neoselachii, Vertebrata) from the Lower Cretaceous of Germany and England. Mesozoic Research 1: 89–106.
Ward, D.J. and Wiest, R.L. 1990. A checklist of Palaeocene and Eocene sharks and rays (Chondrichthyes) from the Pamunkey Group, Maryland and Virginia, USA. Tertiary Research 12: 81–88.
Welton, B.J. 1974. Heptranchias howellii (Reed, 1946) (Selachii: Hexanchidae) in the Eocene of the United States and British Columbia. PaleoBios 17: 1–15.
Whitley, G.P. 1939. Taxonomic notes on sharks and rays. Australian Zoologis 9 (3): 227–262.
Wiest, L.A., Buynevich, I.V., Grandstaff, D.E., Terry, D.O., Maza, Z.A., and Lacovara, K.J. 2016. Ichnological evidence for endobenthic response to the K-Pg event, New Jersey, U.S.A. Palaios 31: 231–241. Crossref
Woodward, A.S. 1886. On the palaeontology of the Selachian genus Notidanus Cuvier. Geological Magazine 3 (5): 205–259. Crossref
Acta Palaeontol. Pol. 69 (2): 173–198, 2024
https://doi.org/10.4202/app.01117.2023