
Bathyal molluscs from Upper Pleistocene methane seeps in Krishna-Godavari Basin, offshore eastern India
CRISPIN T.S. LITTLE, RAJENDAR KUMAR, JOEL E. JOHNSON, and LEON HOFFMAN
Little, C.T.S. Kumar, R., Johnson, J.E., and Hoffman, L. 2025. Bathyal molluscs from Upper Pleistocene methane seeps in Krishna-Godavari Basin, offshore eastern India. Acta Palaeontologica Polonica 70 (3): 443–477.
Compared to other ocean basins there are few reported Recent methane seep communities from the Indian Ocean, with records from offshore Indonesia and Pakistan, and, more recently the east coast of India, in the Krishna-Godavari and Mannar Basins in bathyal water depths. Also from the former area, Upper Pleistocene aged fossil methane seep assemblages have been recovered from sediment cores. Here we describe systematically bivalves, gastropods and scaphopods from a methane seep assemblage penetrated by two sediment cores, drilled in 1045 m and 1050 m water depth, at horizons dated to between 40 and 52 kyrBP. The fossil molluscs comprise 29 taxa: 15 gastropods, 12 bivalves and two scaphopods. Of these, nine are new species: six gastropods (Paralepetopsis bathyalus Hoffman & Little sp. nov., Mesopelex godavariensis Hoffman & Little sp. nov., Anatoma sahlingi Hoffman & Little sp. nov., Cirsonella aperta Hoffman & Little sp. nov., Dikoleps? magnarota Hoffman & Little sp. nov., and Alvania axistriata Hoffman & Little sp. nov.) and three bivalves (Ledella favus Hoffman & Little sp. nov., Yoldiella umbostriata Hoffman & Little sp. nov., and Vesicomya prashadi Hoffman & Little sp. nov.). Six of the molluscan taxa likely had chemosymbionts: (Acharax sp., Gigantidas cf. platifrons, Conchocele sp., Pliocardia cf. solidissima, Callogonia cf. leeana, and Archivesica cf. kawamurai), representing 21% of the diversity in the seep assemblage. Apart from Acharax sp., all these putative chemosymbiotic taxa were likely obligate to seeps, as was probably the case for Paralepetopsis bathyalus Hoffman & Little sp. nov. and Anatoma sahlingi Hoffman & Little sp. nov. The other bivalve, gastropod and scaphopod species in the assemblage have living relatives common in bathyal habitats and can thus be considered as facultative or ‘background’ fauna. The fossil seep assemblage shares some taxa with recent seep communities in the east coast of India and elsewhere in the Indian Ocean, although additional systematic work is needed on the living taxa for a full comparison to be made.
Key words: Mollusca, Gastropoda, Bivalvia, Scaphopoda, taphonomy, hydrocabon seeps, Bay of Bengal, Indian Ocean.
Crispin T.S. Little [earctsl@leeds.ac.uk, ORCID 0000-0002-1917-4460 ], School of Earth and Environment, University of Leeds, Woodhouse Lane, Leeds LS2 9JT, UK; Life Sciences Department, Natural History Museum, London, Cromwell Road, London SW7 5BD, UK.
R. Rajendar Kumar [rrkumar@zsi.gov.in, ORCID 0000-0003-3711-4178 ], Zoological Survey of India, Marine Biology Regional Centre, 130 Santhome High Road, Chennai, Tamil Nadu, 600 028, India.
Joel E. Johnson [joel.johnson@unh.edu, ORCID 0000-0002-5671-7209 ], Department of Earth Sciences, University of New Hampshire, Durham, NH 03824, USA.
Leon Hoffman [Leon.Hoffman@senckenberg.de, ORCID 0000-0002-0205-186X ], Marine Research Department, Senckenberg am Meer, Südstrand 40, 26382 Wilhelmshaven, Germany.
Received 10 June 2024, accepted 13 June 2025, published online 25 August 2025.
Copyright © 2025 C.T.S. Little et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Marine hydrocarbon seeps are discrete sites where fluids rich in hydrocarbons, principally methane, flow onto the seafloor (e.g., Sibuet and Olu 1998; Levin 2005; Cordes et al. 2010; Joye 2020). They have subsequently proved to be common features of active and passive continental margins at bathymetric depths ranging from a few metres to more than seven kilometres (e.g., Fujikura et al. 1999; Leifer 2019). The abundance of reduced compounds at seeps (primarily methane and hydrogen sulphide) serve as the principal energy source for distinctive animal communities (e.g., Sibuet and Olu 1998; Levin 2005; Cordes et al. 2010). In water depths greater than about 400 m these communities are dominated in terms of biomass by larger animals that have symbiotic relationships with methanotrophic and/or sulphide-oxidizing bacteria, such as siboglinid tubeworms, and solemyid, bathymodiolin, vesicomyid, lucinid, and some thyasirid bivalves (e.g., Sibuet and Olu 1998; Sahling et al. 2003; Levin 2005; Dubilier et al. 2008; Cordes et al. 2010). However, there is more diversity amongst the smaller animals, such as provannid gastropods, some of which may be grazers of microbial mats (e.g., Levin 2005). In addition to these obligate animals, hydrocarbon seep communities comprise taxa that can also be found in surrounding deep-sea areas and are thus considered as “background” fauna. Examples amongst the bivalves are the solemyids, nuculanoids, pectinoids and small vesicomyids and thyasirids, and for the gastropods, the buccinoids (e.g., Levin 2005; Cordes et al. 2010). Because of the presence of methane derived authigenic carbonate (MDAC) and mollusc shells at seeps (exhumed from the subsurface in most cases) hard substrates are available for encrusting fauna, such as cirripede barnacles, serpulid polychaetes and corals (e.g., Pereira et al. 2021).
In contrast to the Pacific and Atlantic, relatively few hydrocarbon seeps have been discovered in the Indian Ocean. The Semeulue Seep (2°33.8’ N, 96°45.4’ E) is at 1134 m water depth in the Semeulue Basin, Sumatra Forearc, off northern Sumatra. The site has exposed MDAC and patches of microbial mats with a macrofauna comprising serpulid and probable siboglinid tubeworms, mytilid bivalves, and galatheoid squat lobsters (Wiedicke-Hombach et al. 2006; Siegert et al. 2011). Further South along the Sunda Arc is the Snail Hill Seep (7°57.5’ S, 106°17.7’ E) in 2900 m water depth in the Sunda Forearc Basin, off eastern Java (Wiedicke et al. 2002). Here is exposed MDAC, two genera of siboglinid tubeworms, probable vesicomyids, thyasirids, Acharax (confirmed genetically in Neulinger et al. 2006) and buccinoid gastropods (Wiedicke et al. 2002). Additional large vesicomyid specimens have been recovered from the seafloor in 2100 m water depth off eastern Java (7°24.6’ S, 105°46.9’ E) at what is likely to be one or more seep sites. A few of these specimens were studied genetically by Kojima et al. (2004) and others were described as Calyptogena (Archivesica) garuda Okutani & Soh, 2005. It is probable that these specimens are conspecific. Another seep site in the Java Trench is indicated by the presence of specimens of the siboglinid Paraescarpia echinospica Southward et al., 2002, dredged from 6°25.01’ S, 104°49.53’ E at a depth of approximately 1100–1550 m (Southward et al. 2002). From the western side of the Indian Ocean a large number of seeps have been found along the Makran accretionary margin, Pakistan, between 575 and 2870 m water depth (Bohrmann 2008; Römer et al. 2012; Wei et al. 2021). Below the Oxygen Minimum Zone at around 1000 m the seeps have exposed MDAC, microbial mats and are populated by siboglinid tubeworms, galatheoids, neolepadid barnacles, bathymodiolins, solemyids, and large vesicomyids (Bohrmann 2008; Wei et al. 2021). From this fauna Calyptogena makranensis Krylova & Sahling, 2006, has been described by Krylova and Sahling (2006), with a depth range of 2215–2336 m, and Archarax was genetically confirmed by Neulinger et al. (2006), from one of the seeps at 24°33.0’ N, 64°15.6’ E, in 2220 m water depth.
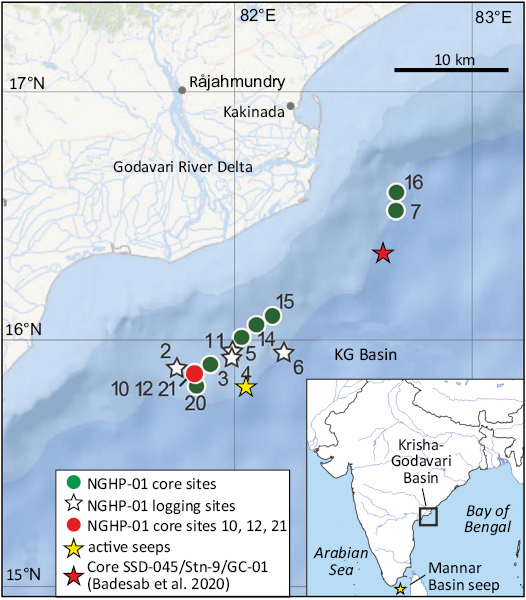
Fig. 1. Map showing NGHP-01 core sites (circles) and logging sites (stars) in the offshore Krishna-Godavari Basin (modified from Lorenson and Collett 2018). Site 12 is co-located with sites 10 and 21 (red circle). Yellow star indicates the position of the active seeps. Red star indicates the position of sediment core SSD-045/Stn-9/GC-01 of Badesab et al. (2020). Inset shows the study area in Krishna-Godavari Basin located on the eastern margin of peninsular India in the Bay of Bengal. Yellow star indicates the position of the active seep in the Mannar Basin, between Sri Lanka and Southern India.
More recently active seeps and associated animal communities have been found on the continental margin off eastern India in the Krishna-Godavari Basin and the Mannar Basin (Mazumdar et al. 2019, 2021; Dewangan et al. 2021; Sangodkar et al. 2022; Peketi et al. 2022) (Fig. 1). The Krishna-Godavari Basin seeps are located in a small area between 15°41.0’ N, 82°2.0’ ‘E and 15°43.7’ N, 82°4’48.7’ E in 1752–1756 m water depth (Mazumdar et al. 2019). The living macrofauna comprises siboglinids and a diversity of other polychaetes, galatheoids (both galatheids and munidopsids), carid shrimps, neolepadids, ophiuroids, echinoids, and gastropod and bivalve molluscs (Mazumdar et al. 2019; Dewangan et al. 2021; Gonsalves et al. 2022; Sangodkar et al. 2022). The bivalves are bathymodiolins, large vesicomyids, thyasirids, soleymids, and pectinids; the gastropods are limpets, provannids, and nertitids. Apart from Shinkaia crosnieri Baba & Williams, 1998 (Gonsalves et al. 2022) and Gigantidas niobengalensis Oliver et al., 2024, none of the animals in these seep communities have been formally described. However, their trophic positions have been investigated using stable isotopes (Peketi et al. 2022). The Mannar Basin seep is located in a pockmark at 7°51.4’ N, 78°36.46’ E in a water depth of 1644 m (Mazumdar et al. 2021) (Fig. 1). The fauna here comprises living specimens of siboglinids, neolepadids, munidopsids, and buccinoid, and relict shells of large vesicomyids. The thyasirid Ascetoaxinus ravichandrani Ravinesh et al., 2024, is the only species to have been described from the Mannar Basin although a variety of lucinids is currently under study (Graham Oliver, personal communication 2024).
Prior to the discovery of active seeps in the Krishna-Godavari Basin, Upper Pleistocene aged bathyal fossil seepage in the same area was inferred by the presence of MDAC and associated bivalve shells from several sediment cores (Mazumdar et al. 2009; Teichert et al. 2014; Collett et al. 2014). Subsequently, a diverse fossil shell assemblage was recovered from two of these cores by CTSL, from which Gale et al. (2020) described the neolepadid cirripede Ashinkailepas indica Gale in Gale et al., 2020. In this paper we systematically describe the molluscan fauna (bivalves, gastropods, and scaphopods) from this material, and discuss it in terms of palaeoecology, and its relationship to the fossil and modern and seep communities off the eastern coast of India, and in other areas of the Indian Ocean.
Institutional abbreviations.—NHMUK, Natural History Museum, London, UK; SMF, Senckenberg Museum, Frankfurt am Main, Germany.
Other abbreviation.—MDAC, methane derived authigenic carbonate.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: LSIDurn:lsid:zoobank.org:pub:1A8C3C31-0761-41D1-925B-E7063975567.
Material and methods
The studied specimens come from two sediment cores drilled in 2006 by the D/V JOIDES Resolution during the Indian National Gas Hydrate Program Expedition 01 (NGHP-01) in the Krishna-Godavari Basin, Bay of Bengal, offshore eastern India (Collett et al. 2014, 2015). Core NGHP-01-12A was drilled in 1045.8 m of water at 15°51.6335’ N, 81°50.2274’ E; core NGHP-01-10D was drilled approximately 500 m to the northeast in 1050.4 m water depth at 15°51.8647’ N, 81°50.0709’ E (Fig. 1). The sediment in the cores and throughout the offshore Krishna-Godavari Basin consists of variably coloured hemipelagic clay containing greater or lesser quantities of organic carbon (Collett et al. 2015; Phillips et al. 2014). Slumping, debris flows, and turbidites are also common (Johnson et al. 2014, 2021; Flores et al. 2014). The sediment subsamples used in this study were taken from the two cores at depths below seafloor of 16–17 mbsf in core NGHP-01-12A, 15–17 mbsf, 41–42 mbsf, and at 61 mbsf in core NGHP-01-10D. Carbon-14 (14C) ages obtained from foraminiferan and mollusc shells at 16.35 mbsf from core NGHP-01-12A have an age of 40 100 ± 360 and 50 300 ± 790 a (BP 1950), respectively (Teichert et al. 2014). Mollusc shells at 16.99 mbsf have a 14C age of 51 600 ± 910 a (BP 1950). These dates correlate to the Upper Pleistocene. No radiocarbon dates have been obtained from concretions or shells from deeper in either of the cores as they are likely to fall outside the range of applicability for this dating method (Teichert et al. 2014).
Variable sediment thickness of sediment (2–10 cm; Table 1) was sampled from the working halves of the cores and this was later sieved (1.7 mm, 300 µm and 125 µm mesh sizes) at the University of Leeds to remove the fine grains. The residual material comprised variably preserved macrofossils and microfossils, mineral grains, and variably cemented mudstone concretions of varying sizes, most of which had a shelly component (Fig. 2). The planktonic elements present in the sieved residues included fish otoliths and the shells of foraminifera, as well as pteropod and heteropod gastropods. The benthic elements comprised foraminifera, serpulid tubes, scleractinian corals, ostracod valves, isolated neolepadid plates, isolated echinoid plates and spines, and mollusc shells (chitons, bivalves, gastropods, and scaphopods). The bivalve, gastropod, and scaphopod shells were removed from the residues for identification under a low magnification binocular microscope. Subsequently, some of the larger specimens were then coated with ammonium chloride for photographic documentation using a digital camera at the University of Leeds. Some of smaller specimens were imaged using Scanning Electron Microscopes (SEM) at the University of Leeds, UK and Senckenberg am Meer, Wilhelmshaven, Germany, with gold coating to enhance image quality. The electron incident energy was 20 keV at the University of Leeds and 10 keV at Senckenberg am Meer. Imaging was performed using direct and back-scatter electrons. The SEM provided calibrated measurements for shell sizes for the smaller specimens. Counts of specimens were limited to those that were complete enough for reliable identification, both in the sieved material and on the surfaces of the concretions. Single gastropod and scaphopod shells were counted as one individual. Articulated bivalve shells were counted as one individual, as were left and right valves from the same sediment sample that obviously belonged to the same specimen; single left and right valves were counted as one individual. Many of the bivalve shell fragments were identifiable only to higher taxonomic level; these were recorded as “present” in the respective samples, but not counted. This was also done for other benthic taxa (chitons, serpulid tubes, corals, cirripedes, echinoids, and ichnofossils) for palaeoecological analysis (Table 1). Species identifications were done to the lowest taxonomic level possible using existing literature. The holotypes and subsets of paratypes (including all those figured) for the new species were deposited in the collection of the Senckenburg Museum, Frankfurt am Main, Germany (SMF). A representative suite of non-type specimens has been accessioned in the collections of the Palaeozoology Division, Zoological Survey of India, Kolkata (for details, see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app70-Little_etal_SOM.pdf).

Fig. 2. Upper Pleistocene cemented mudstone concretion from core NGHP-01-12A, sample SMF373061 from Krishna-Godavari Basin, offshore eastern India, with shells of Conchocele sp. (Co), Parvamussium? aff. scitulum (Smith, 1885) (Pa), Cirsonella aperta Hoffman & Little sp. nov. (Ci), Ashinkailepas indica Gale in Gale et al., 2020 (Sh), and serpulid tubes (Se).
Systematic palaeontology
Class Gastropoda Cuvier, 1795
Subclass Patellogastropoda Lindberg, 1986
Family Neolepetopsidae McLean, 1990
Genus Paralepetopsis McLean, 1990
Type species: Paralepetopsis floridensis McLean, 1990, by original designation; Recent, USA, off Florida, in cold seeps.
Paralepetopsis bathyalus Hoffman & Little sp. nov.
Fig. 3.
Zoobank LCID: urn:lsid:zoobank.org:act:411077AB-6C98-422E-A455- 17BFE8841F10.
Etymology: After the habitat in a bathyal (palaeo-) depth range.
Type material: Holotype, SMF373134. Paratypes, 11 shells, SMF373131; seven shells, SMF373132; one shell, SMF373135; five shells, SMF 373136; four shells, SMF373137; one shell, SMF373133. All from the type locality and horizon.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.18–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.—Type material and several specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Small, thin, solid, limpet shell, oval apertural outline, apex anterior of midline. Protoconch simple exposed whorl, smooth sculpture, often eroded in adult specimens.
Dimensions.—Holotype height 2.5 mm, length 3.8 mm, width 1.2 mm. Shells in type set maximum length 5 mm, height 3 mm, width 1.6 mm.
Description.—Protoconch single stage only, only present on juvenile shells, smooth, planispiral aligned along major height and length axes, 30% from anterior margin, shell leaning posteriorly, fused with teleoconch. Transition to teleoconch clear by change in sculpture, margin sharp. Length 200 μm. Teleoconch convex outline on all margins in dorsal view, anterior margin highest convexity, shell broader posteriorly. Apical angle in side view 120°; anterior outline short, straightened; several convex stages posteriorly. Apex in adult shells with oblong scar and imprint of lost protoconch posteriorly; often worn apex. Sculpture with fine irregular growth lines, fine radial ridges, frequently with nodules at interfaces with major growth stages. Margin smooth, sharp. Internally smooth, glossy, with inconspicuous imprint of muscle. Deep smooth pit at apex.
Remarks.—Eight species in the genus Paralepetopsis have been described from hydrothermal vent and seep habitats in the Atlantic and the Pacific Ocean (McLean 1990, 2008; Beck 1996; Warén and Bouchet 2001, 2009; Chen et al. 2023). Paralepetopsis floridensis McLean, 1990, has a near identical protoconch compared to Paralepetopsis bathyalus Hoffman & Little sp. nov., but the outline is more oval and it has a stronger radial sculpture (McLean 1990). Beck (1996: 89–90, pl. 1) described Paralepetopsis rosemariae Beck, 1996, based on a single specimen from a seamount in the western Pacific; the decapitated shell has a raised outline with little sculpture apart from a few major growth stages. The eastern Pacific Paralepetopsis tunnicliffae McLean, 2008, has a similar morphology compared to Paralepetopsis bathyalus Hoffman & Little sp. nov., but it has a strong radial sculpture (McLean 2008: 18–19, fig. 2A). The Mid-Atlantic Ridge hydrothermal vent species Paralepetopsis ferrugivora Warén & Bouchet, 2001, has a strong radial as well as concentric sculpture (Warén and Bouchet 2001: 123–124, fig. 2e–f). The northwestern Pacific seep species Paralepetopsis lepichoni Warén & Bouchet, 2001, is most similar to Paralepetopsis bathyalus Hoffman & Little sp. nov., but has a coarse as well as fine radial sculpture (Warén and Bouchet 2001: 125, figs. 2c, 5a–f). The eastern Atlantic seep species Paralepetopsis sasakii Warén & Bouchet, 2009, has a coarse radial sculpture and a similar protoconch (Warén and Bouchet 2009: 2331–2332, fig. 3). Paralepetopsis polita Chen et al., 2023, from the South China Sea has a polished shell and all known specimens lack a protoconch. Paralepetopsis bathyalus Hoffman & Little sp. nov. has co-marginal ridges with a few growth stages and juveniles show a protoconch. Chen et al. (2023) found a mitochondrial molecular sequence (COI) of Paralepetopsis polita similar to that of Lepetidae, but radular characteristics similar to those in Neolepetopsidae.
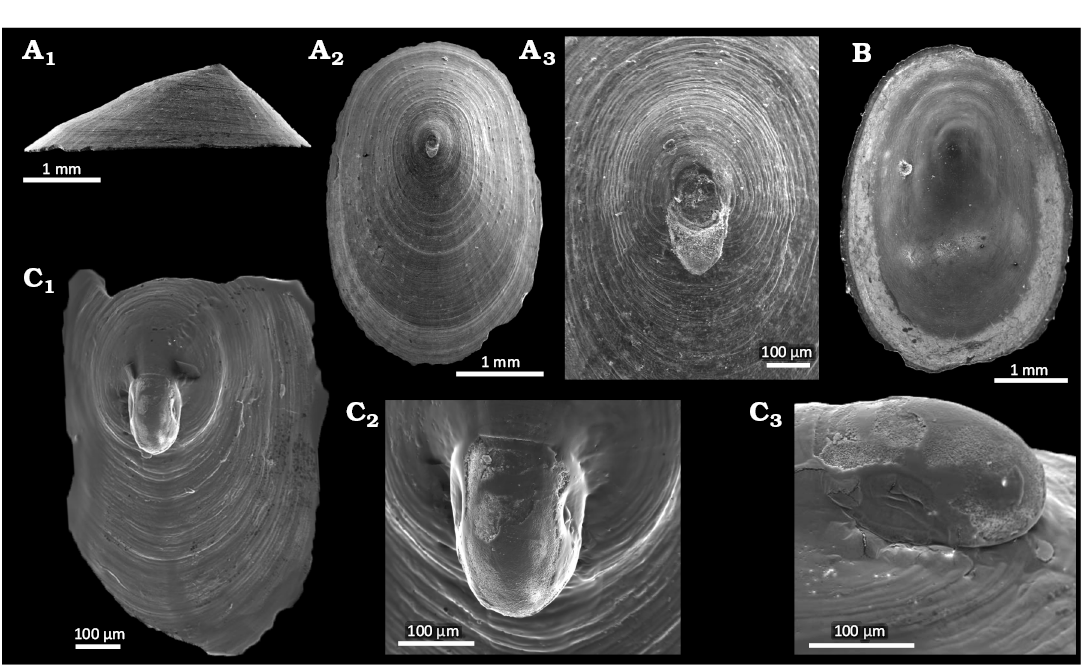
Fig. 3. The neolepetopsid gastropod Paralepetopsis bathyalus Hoffman & Little sp. nov. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373134, holotype, lateral (A1) and external (A2) views, apical detail with protoconch scar (A3). B. SMF373135, paratype, internal view. C. SMF373133, paratype, partial specimen with protoconch, external view (C1), protoconch details (C2, C3).
Stratigraphic and geographic range.—Type locality and horizon only.
Subclass Vetigastropoda Salvini-Plawen, 1989
Family Pseudococculinidae Hickman, 1983
Genus Mesopelex Marshall, 1986
Type species: Mesopelex zelandica Marshall, 1986; Recent, New Zealand.
Mesopelex godavariensis Hoffman & Little sp. nov.
Fig. 4.
Zoobank LCID: urn:lsid:zoobank.org:act:13902726-1A3C-49F9-864E- 98E4D1E3C46B.
Etymology: After the type locality in the Krishna-Godavari Basin.
Type material: Holotype, SMF373138, relatively well preserved limpet shell with protoconch. Paratype, SMF373139, relatively well preserved limpet shell with protoconch.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.28–16.48 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.—Type material and several additional specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Small, thin, fragile, patelliform shell, oval apertural outline, apex posterior of midline. Protoconch simple exposed whorl with pitted sculpture.
Dimensions.—Holotype height 2.0 mm, length 2.4 mm, width 0.8 mm.
Description.—Protoconch single stage only, network of spirally and axial line segments, spirally aligned pitted sculpture, planispiral aligned along major height and length axes, 40% from posterior margin, shell leaning posteriorly, fused with teleoconch. Transition to teleoconch clear by change in sculpture, margin indistinct. Length 150 μm. Teleoconch convex sub-elliptical in dorsal view; anterior margin highest convexity. In side view, weakly convex anterior and posterior margins; apical angle high (approximately 160°). Sculpture smooth with fine irregular growth lines. Margin smooth, sharp. Internally smooth, glossy.
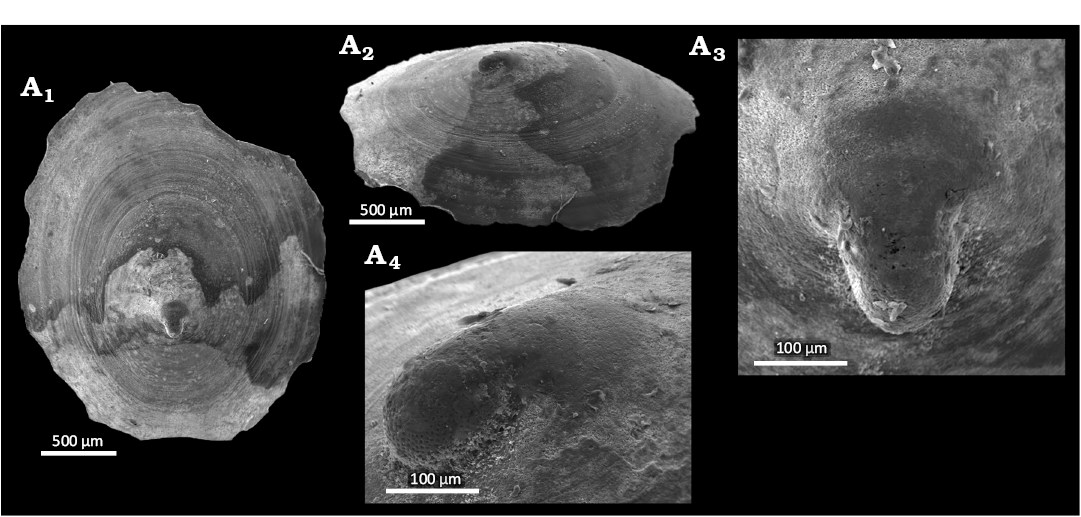
Fig. 4. The pseudococculinid gastropod Mesopelex godavariensis Hoffman & Little sp. nov. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373138, holotype, external (A1) and oblique lateral (A2) views, protoconch details (A3, A4).
Remarks.—A single species has been described in Mesopelex Marshall, 1986 (MolluscaBase 2024). The type species Mesopelex zelandica Marshall, 1986, was found off New Zealand on muddy sediment (Marshall 1986: 535–536, fig. 10a–d). Its apex is more raised and its protoconch is less elongated than in Mesopelex godavariensis Hoffman & Little sp. nov. The dorsal outline, protoconch shape and sculpture are very similar to Mesopelex godavariensis Hoffman & Little sp. nov. Pseudococculinid species in other genera have a strong sculpture (for example in the genera Copulabyssia Haszprunar, 1988, Kaikarapelta Marshall, 1986, Notocrater Finley, 1926), a raised shell (for example in the genera Pseudococculina Schepman, 1908, Pterodacna Herbert, 2024, Tentaoculus Marshall, 1986) or a pointed conical outline (Punctabyssia McLean, 1991).
Stratigraphic and geographic range.—Type locality and horizon only.
Family Fissurellidae Fleming, 1822
Fissurellidae gen. et sp. indet.
Fig. 5.
Material.—One shell (SMF373112) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.38 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Small cap-shaped shell, planispiral coiling. Protoconch paucispiral, three-quarters whorl, smooth, large nucleus, diameter 200 µm. Teleoconch in two stages. First stage half whorl, without selenizone, sculpture with fine radial bifurcating riblets, diameter about 600 µm. Second stage with about 15 radial ribs and commarginal somewhat regular growth margins, weak blunt nodules are formed at the crossings. Shell fragment length about 1 mm, width about 1 mm, thickness about 600 µm.

Fig. 5. Fissurellid gastropod Fissurellidae gen. et sp. indet. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373112, external (A1) and lateral (A2) views, protoconch detail (A3).
Remarks.—The shell is too fragmentary to identify below family level.
Family Anatomidae McLean, 1989
Genus Anatoma Woodward, 1859
Type species: Scissurella crispata J. Fleming, 1828, by monotypy; Recent, NE Atlantic and Mediterranean.
Anatoma sahlingi Hoffman & Little sp. nov.
Fig. 6.
Zoobank LCID: urn:lsid:zoobank.org:act:C51A4420-6F20-4CF1-9748- 8C5B3489F090.
Etymology: After Heiko Sahling (1969–2018), to celebrate his pioneering work on hydrocarbon seep community ecology.
Type material: Holotype, SMF373130. Paratypes, five shells, SMF 373124; one shell, SMF373125; one shell, SMF373126; one shell, SMF373127; 12 shells, SMF373128; four shells, SMF373129.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.18–16.68 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.—Type material and several additional specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Small, thin, fragile, globose elevated shell, apertural wide anal slit, strong sculpture with spirals and ribs on shoulder, weak spiral sculpture below selenizone, selenizone sculpture weak.
Dimensions.—Maximum length 2.5 mm, height 2.3 mm.
Description.—Protoconch single stage only, three quarters flattened whorl. Sculpture rough, grooved network of dominating spirals and short axial line segments. Transition to protoconch clear by terminating smooth rim and change in sculpture. Length 200 μm. Teleoconch with two stages. First stage without selenizone three quarters whorl, convex, sculpture worn; suture to protoconch deep. Second stage one and three quarters globose whorls, spirals and dominating raised prosocline ribs above selenizone; 50 ribs, 15 spirals on body whorl; ribs extending on margin above selenizone, rib frequency increased near lip. Outline raised with early development of area below selenizone and suture. Very broad selenizone at periphery; sculpture reverse-arched ribs, frequency twice of axial ribs above; extended margins above and below eroded. Lower half whorl evenly convex; sculpture fine spiral lines and fine growth lines, flexuous near base. Deep open umbilicus with spiral lines, near vertical umbilical keel ending on columellar callus. Aperture circular, smooth internally with sharp lip; peripheral slit one fifth whorl deep. Union of parietal lip and shoulder area at 90°. Parietal and columellar lip united, concave, ending in pointed extended lip area.

Fig. 6. The anatomid gastropod Anatoma sahlingi Hoffman & Little sp. nov. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373130, holotype, lateral (A1) and adapical (A2) views, detail of umbilicus (A3), detail of selenizone (A4). B. SMF373126, paratype, apical view (B1), detail of protoconch (B2). C. SMF373125, paratype, lateral (C1) and apical (C2) views. D. SMF373127, paratype, lateral view (D1), oblique detail of protoconch (D2), detail of protoconch microsculpture (D3), detail of selenizone (D4).
Remarks.—Anatomids in the Indian Ocean were reviewed by Geiger (2012) and Hoffman et al. (2022). Hoffman et al. (2022) identified 17 species in the Indian Ocean of which six species were found alive in the lower (>2000 m) bathyal depth range; four species were associated with hydrothermal vents on the Central Indian Ridge and the SE Indian Ridge. The combination of strong ribs and spirals on the shoulder, weak spirals on the lower whorl and very wide selenizone in the specimens here is different from all known species in the Indian Ocean. The strong sculpture on the shoulder is common in anatomids, but this is commonly combined with ribs and occasionally spirals at the base. The lack of basal ribs is unique. For example, Anatoma paucisculpta Hoffman et al., 2022, has a weak basal rib and spiral sculpture but lacks the rib sculpture on the shoulder (Hoffman et al. 2022: 155–156, fig. 11). Anatoma sagamiana (Okutani, 1964) from the Western Pacific has a similar outline and wide selenizone with thin margins like Anatoma sahlingi Hoffman & Little sp. nov., but it has a weak reticulated sculpture above and below the selenizone and its umbilicus is closed (Geiger 2012: 1075–1081, figs. 875–880); Anatoma sahlingi Hoffman & Little sp. nov. has a stronger sculpture on the shoulder and a weak spiral sculpture below, and its umbilicus is open. The similar Anatoma soyoae (Habe, 1951) is found off NW Honshu, Japan. It has a similar wide selenizone with thin margins and its umbilicus is open, but its upper sculpture is finer and the outline is more compressed (Geiger 2012: 1095–1097, figs. 895–896).
Stratigraphic and geographic range.—Type locality and horizon only.
Superfamily Seguenzioidea Verrill, 1884
Seguenzioidea gen. et sp. indet.
Fig. 7.
Material.—Two shells, SMF373111 and one lost in processing. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.18–16.38 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Small shell with broad spire, umbilicate. Protoconch paucispiral, three quarters whorl, small nucleus, sculpture with weak tiny dimples, terminal smooth straight rim, diameter about 300 µm. Teleoconch with two convex whorls with slightly cyrtoconoid outline, sculpture with numerous flexuous growth lines, suture deep, umbilicus filled but probably wide and deep. Aperture broad piriform, outer lip thin, prosocline, columellar lip sharp.

Fig. 7. Seguenzioid gastropod Seguenzioidea gen. et sp. indet. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373111, lateral view. B. Shell lost in processing, lateral view (B1), detail of protoconch (B2).
Remarks.—Specific characters are the flexuous growth lines and prosocline lip. The general outline and protoconch are clearly skeneimorph, but the small number of collected specimens precludes us identifying them at a lower taxonomic level. The shells are very similar to Akritogyra curvilineata Warén, 1992, from the NE Atlantic. This genus has not yet been assigned to a family in the Seguenzioidea.
Family Skeneidae Clark, 1851
Genus Cirsonella Angas, 1877
Type species: Cirsonella australis Angas, 1877, junior synonym of Cirsonella weldii (Tenison Woods, 1877), by monotypy; Recent, New South Wales, Australia.
Cirsonella aperta Hoffman & Little sp. nov.
Fig. 8.
Zoobank LCID: LSIDurn:lsid:zoobank.org:act:60068CBB-196B-4C4E- 82E5-8F4CD3D84E52.
Etymology: With reference to the partly open umbilicus.
Type material: Holotype, shell, SMF373123. Paratypes, one shell, SMF 373118; one shell, SMF373119; one shell, SMF373120; one shell, SMF373121; two shells, SMF373122, from the type locality and horizon.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.18–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.— Type material and several specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Small shell with moderately raised spire, solid, smooth, narrow open umbilicus, umbilical chink with two broad spiral cords, suture shallow impressed.
Dimensions.—Holotype height 1.7 mm, length 2.0 mm. Shells in type set maximum height 2.1 mm, length 2.3 mm.
Description.—Protoconch single stage only, half whorl, flattened, smooth, curved sharp lip. Transition to dissoconch clear by labial margin. Length 330 µm. Teleoconch raised with flat apex, two convex whorls. Smooth, slightly flexuous growth lines. Umbilical area demarcated with rim, few axial, curved ridges with variable strength fill area to columella, narrow open umbilical slit. Aperture subcircular, internally smooth with (operculum?) edge near external margin, external lip blunt.

Fig. 8. The skeneid gastropod Cirsonella aperta Hoffman & Little sp. nov. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373123, holotype, lateral (A1) and apical (A2) views, detail of protoconch (A3). B. SMF373119, paratype, lateral (B1) and apical (B2) views, detail of protoconch (B3). C. SMF373120, paratype, lateral (C1) and apical (C2) views, detail of protoconch (C3). D. SMF373121, paratype, lateral (D1) and apical (D2) views.
Remarks.—We place the species in Cirsonella (Skeneidae) on the basis of the morphology of the protoconch, teleoconch and the partly closed umbilicus. Alternatively, it could be placed in the genus Skenea Fleming, 1825. Species in Cirsonella are widely distributed, e.g., the Northern Atlantic, the SW Pacific (off Australia and New Zealand), South Africa and the Kerguelen Islands (MolluscaBase 2024). This is the first species from the Northern Indian Ocean. An open umbilicus is typical in young specimens and a narrow umbilical slit is common in for example the NE Atlantic Cirsonella ateles (Dautzenberg & Fischer, 1896) and Cirsonella gaudryi (Dautzenberg & Fischer, 1896) (Hoffman et al. 2020: 55–59, figs. 48–49). Known species show a well-defined chink in the umbilical area; the open umbilicus is often covered by smooth columellar callus in adult specimens. The umbilical grooves and ridges distinguish Cirsonella aperta Hoffman & Little sp. nov. from other described species.
Stratigraphic and geographic range.—Type locality and horizon only.
Genus Dikoleps Høisaeter, 1968
Type species: Margarita pusilla Jeffreys, 1847, junior synonym of Dikoleps nitens (Philippi, 1844), by original designation; Recent, NE Atlantic.
Dikoleps? magnarota Hoffman & Little sp. nov.
Fig. 9.
Zoobank LCID: urn:lsid:zoobank.org:act:2B99F77B-E09E-4802-8D26- DDE333A81C5F.
Etymology: From the resemblance of the species to a big wheel, magna rota in Latin.
Type material: Holotype, SMF376542, well preserved shell with protoconch. Paratypes, two shells, SMF 376541; one shell, SMF376543; one shell, SMF376544, from the type locality and horizon.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.18–16.48 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.— Type material and several specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Very small shell with flattened spire, fragile, sculpture with broadly undulating ribs on shoulder, suture narrow and deep.
Dimensions.—Holotype height 0.8 mm, length 1.3 mm. Maximum dimensions of shells in type set as in holotype.
Description.—Protoconch single stage only, three quarters flattened smooth whorl, curved broken lip. Transition to protoconch clear by change in sculpture. Length 220 μm Teleoconch slightly raised, two and a half rounded whorls. Sculpture smooth with fine growth lines and flexuous undulations on shoulder, undulations disappearing near periphery; base smooth with fine flexuous growth lines. Umbilicus open, deep. Aperture well rounded convex, flexuous, sharp external and columellar lip; parietal side concave; callus very thin.

Fig. 9. The skeneid gastropod Dikoleps? magnarota Hoffman & Little sp. nov. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF376542, holotype, apical (A1), adapical (A2), and lateral (A3, A4) views, detail of protoconch (A5). B. SMF376543, paratype, apical (B1) and lateral (B2) views, detail of protoconch (B3).
Remarks.—We are somewhat uncertain about the generic assignment of the species, but place it in the genus Dikoleps (Skeneidae) on the basis of the morphology of the protoconch, teleoconch and the flexuous base of the lip and the growth lines. Currently, nine Dikoleps species are known; eight species in the NE Atlantic and Mediterranean Sea, mostly living subtidally down to an upper bathyal depth range, and one species is known from South Africa (MolluscaBase 2024). This is the first species from the northern Indian Ocean.
Stratigraphic and geographic range.—Type locality and horizon only.
Subclass Caenogastropoda Cox, 1960
Family Rissoidae Gray, 1847
Genus Alvania Risso, 1826
Type species: Alvania europea Risso, 1826, junior synonym of Alvania cimex (Linnaeus, 1758), by subsequent designation; Recent, NE Atlantic.
Alvania axistriata Hoffman & Little sp. nov.
Fig. 10.
Zoobank LCID: urn:lsid:zoobank.org:act:63375D11-3BF0-4D60- B958- B7D3A0901AC5.
Etymology: After the axial ribs that are aligned with the spire axis.
Type material: Holotype, SMF373067, well preserved shell with protoconch.. Paratypes, three shells, SMF 373063; five shells, SMF373065; two shells, SMF373068; one shell, SMF373064; six shells, SMF373066, from the type locality and horizon.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.18–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.—Type material and several specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Small shell with raised rounded outline with pointed apex, fragile, broad ribs and weaker broad spiral cords on shell face, suture deeply impressed.
Dimensions.—Holotype height 2.2 mm, length 1.4 mm. Maximum dimensions of shells in type set as in holotype.
Description.—Protoconch single stage only; raised, slightly over one whorl; embryonic shell smooth; last half whorl with shallow vertical line segments, somewhat spirally aligned, length 10–20 µm, width 2 µm; thin sharp margin. Transition to protoconch clear by change in sculpture. Maximum diameter 470 μm. Teleoconch conical, apical angle 50–60°, two and half convex whorls. Sculpture wide axial ribs, interspaces of comparable width, about 30 ribs on each whorl; fine spiral threads overrunning the ribs; one broad spiral channel high on the shoulder. Base whorl with 6–7 strong spiral cords, fading near periphery. Thickened rim near lip. Aperture orthocline, pyriform, bevelled external lip, thick callus, peristome complete.

Fig. 10. The rissoid gastropod Alvania axistriata Hoffman & Little sp. nov. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373064, paratype, lateral (A1, A2) and apical (A3) views, detail of protoconch, lateral view (A4), detail of protoconch microsculpture (A5), detail of protoconch, apical view (A6). B. SMF373067, holotype, lateral view (B1, B2), detail of protoconch, apical view (B3).
Remarks.—We place the specimens in the genus Alvania on the basis of the combination of morphological features, for example size, outline, aperture, sculpture and protoconch. The genus is cosmopolitan, but most diverse in the NE Atlantic and Mediterranean Sea. Alvania stigmata Frauenfeld, 1867, is a similar species described from the Nicobar Islands. However, that species has 16–18 vertical ribs and 3–4 basal cords and a few subsutural lines; our species has 30 ribs, 6–7 basal cords and one subsutural channel (Frauenfeld 1867: 12). The Indian Ocean Alvania versoverana (Melvill, 1893) has a similar outline and size, but its axial ribs are curved and the spiral cords cover the full shell face (Mellvill 1893: 61, pl. 1: 15). Alvania mahimensis (Melvill, 1893) has a similar outline and size, but its suprasutural sculpture has 3–4 rows of broad nodules (Mellvill 1893: 61, pl. 1: 17). The NE Atlantic Alvania electa (Monterosato, 1874) is very similar to our species, but has a strong spiral sculpture on the shell face and it lacks the spiral channel on the shoulder (Bouchet and Warén 1993: 633–634, figs. 1410–1420). Alvania porcupinae Gofas & Warén, 1982, is also very similar but has a larger protoconch with two whorls (Bouchet and Warén 1993: 636–638, figs. 1379, 1421–1429).
Stratigraphic and geographic range.—Type locality and horizon only.
Genus Benthonellania Lozouet, 1990
Type species: Benthonellania gofasi Lozouet, 1990, by original designation; Recent, NE Atlantic.
Benthonellania sp.
Fig. 11A.
Material.—One shell, SMF373110. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.68 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Elevated, conical, strong rissoid shell. Protoconch paucispiral with large nucleus, one and a quarter whorls, smooth, diameter about 350 µm. Teleoconch with three flattened whorls, sculpture with about 14 broadly undulating axial ribs with protruding pointed nodules on the shoulder, very fine growth lines, narrow undulating shoulder area, deeply impressed suture. Abapically pointed. Ultimate whorl shows fading axial ribs and few weak chords below periphery. Lip broken. Estimated height 2.5 mm.

Fig. 11. Gastropods from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. Rissoid Benthonellania sp., SMF373110, lateral (A1, A3) and apical (A2) views, detail of protoconch (A4). B. Conoidea gen. et sp. indet., SMF373107, lateral view (B1, B2). C. Columbellidae gen. et sp. indet., SMF373108, lateral view (C1), detail of protoconch (C2).
Remarks.—The single shell is well preserved, apart from a broken lip. The shell is morphologically very similar to the East Atlantic type species. As several congeneric species are similar we refrain from a species determination based on only one specimen.
Order Neogastropoda Wenz, 1938
Superfamily Buccinoidea Rafinesque, 1815
Family Columbellidae Swainson, 1840
Columbellidae gen. et sp. indet.
Fig. 11C.
Material.—Two shells, SMF373108 and SMF373109, Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.18–16.68 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1.
Table 1. Occurrence of bivalves, gastropods, and scaphopods, and selected other benthic taxa from cores NGHP-01-12A and NGHP-01-10D. p, present. Counts include both type and non-type material. CSV version of this table is available in SOM 2.
|
Sample |
Average (mbsf) |
Paralepetopsis bathyalus sp. nov. |
Mesopelex godavariensis sp. nov. |
Fissurellidae sp. indet. |
Anatoma sahlingi sp. nov. |
Seguenzioidea sp. indet. |
Cirsonella aperta sp. nov. |
Dikoleps? magnarota sp. nov. |
Alvania axistriatus sp. nov. |
Benthonellania sp. |
Columbellidae sp. indet. |
Conoidea sp. indet. |
Eulimella sp. 1 |
Eulimella sp. 2 |
Odostomia sp. |
Cylichnidae? sp. indet. |
Acharax sp. |
Ledella favus sp. nov. |
Yoldiella umbostriatus sp. nov. |
Gigantidas cf. platifrons |
Parvamussium? aff. scitulum |
Propeamussiidae? sp. indet. |
Conchocele sp. |
Vesicomya prashadi sp. nov. |
Pliocardia cf. solidissima |
Callogonia cf. leeana |
Archivesica cf. kawamurai |
Cuspidariidae? sp. indet. |
Dentaliidae? sp. indet. |
Cadulus chuni |
Bathymodiolinae shell fragments |
Propeamussiidae shell fragments |
Vesicomyidae shell fragments |
Chitons |
Caryophyllia sp. |
Serpulids |
Ashinkailepas indica |
Echinoid test plates |
Radulichnus sp. |
|
Core NGHP-01-10D |
|||||||||||||||||||||||||||||||||||||||
|
CLM 3H2 20-22 cm |
15.3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
|
|
CLM 3H2 42-46 cm |
15.52 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
|
|
CLM 3H2 44-46 cm |
15.54 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
CLM 3H2 60-62 cm |
15.7 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
p |
|
CLM 3H2 62-64 cm |
15.72 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
|
|
CLM 3H2 63-65 cm |
15.73 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
CLM 7x1 44-46 cm |
41.54 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
p |
|
CLM 7x1 82-83 cm |
41.92 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
p |
|
CLM 7x1 91-93 cm |
42.01 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
|
|
|
CLM 7x1 117-119 cm |
42.27 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
p |
|
CLM 10x2 1-1 cm |
61.03 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
|
|
Abundance |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
1 |
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Core NGHP-01-12A |
|||||||||||||||||||||||||||||||||||||||
|
CLM 3H2 30-40 cm |
16.18 |
12 |
|
|
6 |
1 |
8 |
2 |
4 |
|
1 |
|
|
|
|
|
|
|
13 |
13 |
8 |
3 |
1 |
|
|
|
2 |
|
|
|
p |
p |
p |
p |
|
p |
|
p |
p |
|
CLM 3H2 40-50 cm |
16.28 |
16 |
2 |
|
19 |
|
2 |
3 |
5 |
|
|
1 |
|
|
|
|
|
1 |
1 |
13 |
6 |
5 |
|
|
|
|
3 |
|
|
|
p |
p |
p |
p |
|
p |
p |
|
p |
|
CSD 3H2 52-54.5 cm |
16.36 |
|
|
|
7 |
|
2 |
2 |
8 |
|
|
|
|
|
|
|
|
|
2 |
2 |
1 |
|
|
|
|
|
|
|
|
|
p |
p |
p |
|
|
p |
p |
|
p |
|
FOS 3H2 52-54 cm |
16.36 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
CLM 3H2 50-60 cm |
16.38 |
10 |
1 |
|
23 |
1 |
4 |
1 |
11 |
|
|
|
|
|
|
|
|
|
7 |
4 |
|
4 |
|
1 |
|
|
3 |
|
|
|
p |
p |
p |
p |
|
|
p |
|
p |
|
CSD 3H2 52-57 cm |
16.38 |
2 |
1 |
1 |
4 |
2 |
|
4 |
7 |
|
|
|
|
1 |
1 |
|
|
|
2 |
2 |
1 |
3 |
|
|
|
|
|
|
|
|
p |
p |
p |
p |
|
p |
p |
|
|
|
CLM 3H2 60-70 cm |
16.48 |
7 |
1 |
|
6 |
1 |
2 |
1 |
10 |
|
1 |
|
|
|
|
|
|
1 |
4 |
1 |
|
1 |
|
2 |
|
|
2 |
|
|
|
|
|
|
p |
|
p |
p |
|
|
|
CLM 3H2 80-90 cm |
16.68 |
8 |
|
|
3 |
1 |
2 |
|
11 |
1 |
1 |
|
1 |
|
|
|
|
2 |
1 |
1 |
3 |
2 |
2 |
5 |
1 |
|
2 |
|
|
|
|
p |
p |
|
|
p |
p |
p |
p |
|
CSD 3H2 84-86 cm |
16.68 |
|
|
|
|
|
|
|
7 |
|
|
|
|
|
|
|
|
|
|
|
1 |
1 |
|
|
|
|
|
|
|
|
|
p |
p |
p |
|
|
p |
|
p |
|
CLM 3H2 90-100 cm |
16.78 |
5 |
|
|
|
|
|
|
18 |
|
|
|
|
|
|
|
|
6 |
1 |
2 |
1 |
3 |
|
|
|
|
3 |
|
6 |
4 |
|
p |
p |
|
|
p |
p |
|
|
|
CLM 3H2 100-110 cm |
16.88 |
3 |
|
|
|
|
2 |
|
10 |
|
|
|
|
|
|
1 |
1 |
4 |
1 |
1 |
|
3 |
1 |
1 |
|
|
4 |
1 |
|
1 |
|
p |
p |
p |
|
p |
p |
|
p |
|
CSD 3H2 116-117 cm |
17 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
|
|
|
|
CSD 3H2 116-118 cm |
17 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p |
|
|
|
Abundance |
|
63 |
5 |
1 |
68 |
6 |
22 |
13 |
91 |
1 |
3 |
1 |
1 |
1 |
1 |
1 |
1 |
14 |
32 |
39 |
21 |
25 |
4 |
9 |
1 |
|
19 |
1 |
6 |
5 |
|
|
|
|
|
|
|
|
|
Description.—Elevated, small, smooth collumbellid shell with blunt apex. Protoconch paucispiral, one whorl with large nucleus, smooth, diameter about 600 µm. Teleoconch juvenile only, two whorls, smooth with very fine growth lines, impressed suture, very narrow shoulder. Outline straightened suprasutural whorl face, widely convex periphery, concave below, columellar area straightened. Aperture piriform with open straight siphonal channel, outer lip orthocline, thin.
Remarks.—All three shells are juveniles, so identification below family level is not possible.
Superfamily Conoidea Fleming, 1822
Conoidea gen. et sp. indet.
Fig. 11B.
Material.—One shell (SMF373107) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.28 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Elevated conical shell, raphitomiform, strongly sculptured with blunt apex. Protoconch paucispiral, about one whorl with large nucleus, smooth, diameter about 350 µm. Teleoconch juvenile only, three whorls with flexuous growth lines. Sculpture with three spiral cords, one shoulder cord, one at the suture and one below the suture, about 16 flexuous axial ribs with pointed nodules at crossings with cords. Aperture piriform.
Remarks.—The apex and part of the teleoconch is obscured, so the material is inadequate for a determination or a complete description. However, it is possible the shells belongs to the genus Taranis Jeffreys, 1870 (Raphitomidae Bellardi, 1875).
Subclass Heterobranchia Burmeister, 1837
Superfamily Pyramidelloidea Gray, 1840
Family Pyramidellidae Gray, 1840
Genus Eulimella Forbes & M’Andrew, 1846
Type species: Eulima macandrei Forbes, 1844; original designation, junior synonym of Eulimella scillae (Scacchi, 1835); Recent, NE Atlantic.
Eulimella sp. 1
Fig. 12A.
Material.—One shell (SMF373103) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.68 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Highly elevated shell, eullimelliform, smooth with fine apex. Protoconch sinistral, axis rotated about 120° from spire axis, one whorl, smooth, diameter about 300 µm. Teleoconch dextral, smooth, with fine orthocline growth lines, seven whorls, slightly convex above deeply impressed suture, ultimate whorl below suture with straightened columella. Aperture trapezoidal.

Fig. 12. Heterobranch gastropods from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373103, Eulimella sp. 1, lateral view (A1, A2). B. SMF373104, Eulimella sp. 2, lateral view. C. SMF373105, Odostomia sp., lateral view (C1, C2). D. SMF373106 Cylichnidae? gen. et sp. indet, lateral view (D1, D2). H, height.
Remarks.—Placement in Eulimella is based on the smooth elevated shell and the rotated heterotrophic protoconch. However, the single shell is considered as inadequate for a description.
Eulimella sp. 2
Fig. 12B.
Material.—One shell (SMF373104) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.38 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Highly elevated shell, eullimelliform, smooth, with large apex. Protoconch sinistral, axis rotated about 100° from spire axis, one whorl, smooth, diameter about 300 µm. Teleoconch smooth with fine orthocline growth lines, four and half whorls, maximum convexity at periphery, deeply impressed suture, ultimate whorl below suture with straightened columella. Aperture trapezoidal.
Remarks.—Placement in Eulimella is based on the smooth elevated shell and the rotated heterotrophic protoconch. However, the single shell is considered as inadequate for a description.
Genus Odostomia Fleming, 1813
Type species: Turbo plicatus Montagu, 1803, accepted as Odostomia plicata (Montagu, 1803), type by subsequent designation.
Odostomia sp.
Fig. 12C.
Material.—One shell (SMF373105) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.38 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Highly elevated shell, eullimellid shape, smooth with large apex. Protoconch sinistral, axis rotated about 100 degrees from spire axis, one whorl, smooth, diameter about 300 µm. Teleoconch smooth with fine orthocline growth lines, four and half whorls, maximum convexity at periphery, deeply impressed suture, ultimate whorl below suture straightened columella. Aperture trapezoidal.
Remarks.—Placement in Odostomia is based on the protoconch and on the smooth conical shell with flattened whorls. The single shell is too poorly preserved for a more detailed description.
Order Cephalaspidea Fischer, 1883
Superfamily Cylichnoidea Adams & Adams, 1854
Family Cylichnidae Adams & Adams, 1854
Cylichnidae? gen. et sp. indet.
Fig. 12D.
Material.—One shell (SMF373106) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Small elongated, convolved shell, cylichnid shape. Protoconch not seen, intorted. Teleoconch, ultimate whorl only, smooth with fine curved growth lines. Outline ovate, slightly angular, maximum convexity apically, periphery weakly convex. Aperture elongated ovate, widening abapically, narrow adapically.
Remarks.—The single shell is poorly preserved for a detailed description and consequently the family-level placement is uncertain.
Class Bivalvia Linnaeus, 1758
Subclass Protobranchia Pelseneer, 1889
Order Solemyida Dall, 1889b
Family Solemyidae Gray, 1840
Genus Acharax Dall, 1908a
Type species: Solemya johnsoni Dall, 1891, by original designation; Recent, Baja California.
Acharax sp.
Fig. 13A.
Material.—One shell (SMF373102) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India; 15°51.8647’ N, 81°50.0709’ E, 1050 m water depth, 15.54 mbsf level, core NGHP-01-10D; Upper Pleistocene. For additional material see Table 1.
Description.—Large elongated, oblong, thick solemyid shell. Prodissoconch inconspicuous. Dissoconch flattened, oblong with coarse irregular growth lines, irregular radial lines from umbo towards antero-ventral margin. Outline blunt marginally protruding umbo, straight antero-dorsal margin, postero-dorsal marging slightly concave. Anterior and posterior ends most convex, ventral margin weakly convex. External ligament groove present. Maximum length of fragmented shell 34 mm.
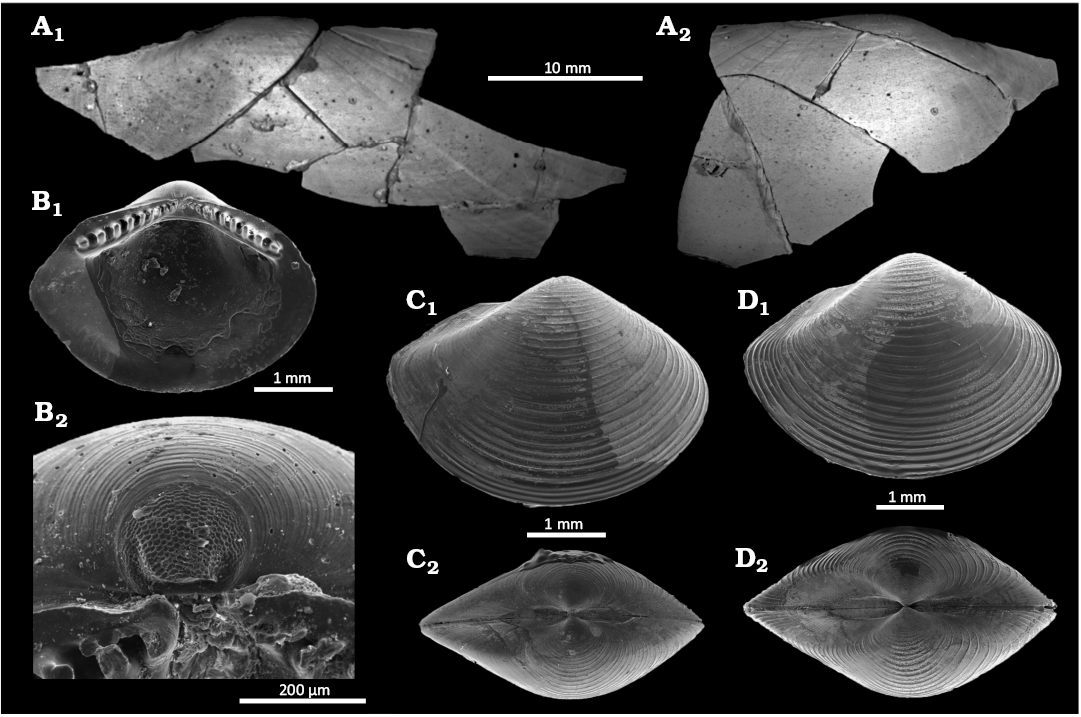
Fig. 13. Solemyid and nuculanid bivalves from Upper Pleistocene seep deposits from cores NGHP-01-12A and NGHP-01-10D, Krishna-Godavari Basin, offshore eastern India. A. Acharax sp. SMF373102, external views of partial right (A1) and left (A2) valves. B–D. Ledella favus Hoffman & Little sp. nov. B. SMF373071, paratype 2, internal view of left valve (B1), and detail of prodissoconch (B2). C. SMF373070, paratype 1, external view of right valve (C1), and dorsal view (C2). D. SMF373069, holotype, external view of right valve (D1), and dorsal view (D2).
Remarks.—Our material is too poorly preserved to conclusively compare with a described species or describe it as new. It is placed in Acharax because of presence of an external ligament pit, but no sign of any internal ligament scars.
Order Nuculanida Carter et al., 2000
Superfamily Nuculanoidea Adams & Adams, 1858 (1854)
Family Nuculanidae Adams & Adams, 1858 (1854)
Genus Ledella Verrill & Bush, 1897
Type species: Ledella bushae Warén, 1978, junior synonym of Ledella ultima (Smith, 1885), by subsequent designation; Recent, Atlantic.
Ledella favus Hoffman & Little sp. nov.
Fig. 13B–D.
Zoobank LCID: urn:lsid:zoobank.org:act:C0663527-E813-4D74-B94B -B85AAE34D9F6.
Etymology: After the honeycomb (favus in Latin) sculpture on the prodissoconch.
Type material: Holotype, one shell, SMF373069. Paratypes, one valve, SMF373071; one shell, SMF373070, from the type locality and horizon.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.28–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.—Type material and several specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Solid shell, oval outline, posteriorly truncated obliquely: strong co-marginal growth margins; length up to 6 mm. Internally smooth, broad hinge plates and heavy dentation.
Dimensions.—Holotype height 4.2 mm, length 5.8 mm, tumidity 3.2 mm. Maximum dimensions of shells in type set as in holotype.
Description.—Prodissoconch single stage evident only, oblong, truncated dorsally, flattened, raised ridge on dorsal side with pointed end; sculpture with central hexagonal network (honeycomb-like), smooth at margins. Transition to dissoconch clear by change in sculpture. Length 200 μm. Dissoconch with straightened dorsal outline posteriorly; ventral and anterior margins convex; posterior with obliquely truncated cusp at end of ridge; shell moderately inflated. Lunule and escutcheon long and lanceolate, escutcheon somewhat inconspicuous, both with fine growth lines. Margins sharp and smooth. Sculpture with regular co-marginal growth stages, width growth lines equal to inter-spaces. Internal surface and margin smooth; muscle scars and pallial line inconspicuous. Pallial line without sinus. Hinge only known from right valve; wide hinge plates at 120°; nine sharp teeth on either side (in right valve with length 4 mm), rectangular in cross-section, central teeth in either row most pronounced. Triangular resilifer placed centrally below umbo.
Remarks.—Placement in Ledella is based on the truncated-oval outline with a cusp, the solid shell, the wide hinge plates each with few teeth and the clear resilifer. The bathyal Ledella procumbens (Prashad, 1932) is known from the Bay of Bengal in 450–1820 m (Huber 2010). This species has a long pointed rostrum, unlike our specimens (Prashad 1932: 20, pl. 1: 21–22). The cosmopolitan type species Ledella ultima (Smith, 1885) has a more globose shell, convex dorsal outline posteriorly, and has thickened (box-shaped) margins when adult (Knudsen 1970). Tan (2021) reported a similarly shaped species, Nuculana cf. corbuloides (Smith, 1885), from deep water off Java but this species has a more pointed rostrum and coarser co-marginal ribs. The prodissoconch sculpture on Ledella favus Hoffman & Little sp. nov. resembles that of other modern and fossil nuculanids, for example Nuculana grasslei Allen, 1993 (Allen 1993: figs. 2, 3; Kiel 2006).
Stratigraphic and geographic range.—Type locality and horizon only.
Family Yoldiidae Dall, 1908b
Genus Yoldiella Verrill & Bush, 1897
Type species: Yoldia lucida Lovén, 1846, by original designation; Recent, NE Atlantic.
Yoldiella umbostriata Hoffman & Little sp. nov.
Fig. 14.
Zoobank LCID: urn:lsid:zoobank.org:act:40B9B90E-1FB5-42F7-811D- 4EFEAB577ED6.
Etymology: After the striated sculpture on the prodissoconch.
Type material: Holotype, one shell, SMF373114. Paratypes, six shells, SMF373113; one shell, SMF373116; one valve, SMF373117; two shells, SMF373115, from the type locality and horizon.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.18–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.—Type material and several specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Small protobranch, solid, equivalve, inequilateral shell oblong outline, beaks anteriorly, smooth. Sculpture prodissoconch with parallel lines.
Dimensions.—Holotype height 1.6 mm, length 2.5 mm, tumidity 1.0 mm. Maximum dimensions of shells in type set height 2.2 mm, length 3.3 mm, tumidity 1.3 mm.
Description.—Prodissoconch single stage evident only, oblong, truncated dorsally, flattened; sculpture with central irregular fine ridges, aligned parallel, occasionally connected by thin ridge segments, smooth at margins. Transition to dissoconch clear by change in sculpture. Length 200 μm. Dissoconch with straightened dorsal outline posteriorly; ventral, posterior and anterior margins convex; anterior most convex; shell rather flattened. Lunule and escutcheon absent. Sculpture irregularly spaced weak growth margins; some specimens nearly smooth. Margins sharp and smooth. Internal surface and margin smooth; muscle scars and pallial line inconspicuous. Hinge only known from right valve; wide hinge plates at 150°; posterior plate longer; nine anterior teeth, 13 posterior teeth (in right valve with length 3.3 mm), chevron-shaped in cross-section, central teeth in either row most pronounced. Triangular, oblique resilifer behind umbo.
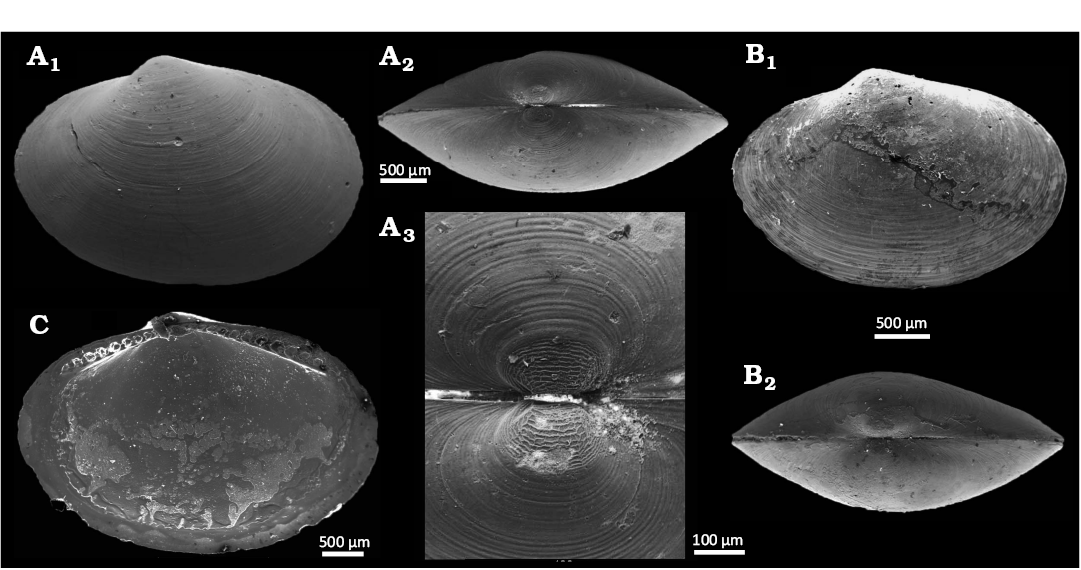
Fig. 14. The yoldiid bivalve Yoldiella umbostriata Hoffman & Little sp. nov. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373114, holotype, external view of right valve (A1), dorsal view (A2), detail of prodissoconch (A3). B. SMF373116, paratype 1, external view of right valve (B1), dorsal view (B2). C. SMF373117, paratype 2, internal view of left valve.
Remarks.—Yoldiella retusa (Hinds, 1843) is known from the Northern Indian Ocean (Huber 2010) and is similar to Yoldiella umbostriata Hoffman & Little sp. nov. Hinds (1843: 99) reported the outline of this species as being nearly equilateral with a pointed anterior margin, and a length of 4.6 mm. Our shells are inequilateral with a more convex anterior and a maximum length of 3.3 mm.
Stratigraphic and geographic range.—Type locality and horizon only.
Infraclass Pteriomorphia Beurlen, 1944
Order Mytilida Férussac, 1822
Family Modiolidae Termier & Termier, 1950
Genus Gigantidas Cosel & Marshall, 2003
Type species: Gigantidas gladius Cosel & Marshall, 2003, by original designation; Recent, SW Pacific, off North Island, New Zealand, cold seeps and active hydrothermal vent sites.
Gigantidas cf. platifrons (Hashimoto & Okutani, 1994)
Fig. 15.
Material.—One adult valve, SMF373100; one adult valve, SMF373094; one juvenile shell, SMF373099; one juvenile valve, SMF373096; two juvenile valves, SMF373098; one juvenile valve, SMF373101; one shell and three valves (all juveniles), SMF373097. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.18–16.78 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1. A single, small poorly preserved valve SMF373095 from 42.01 mbsf level, core NGHP-01-10D may belong to this species.
Description.—Large bathymodiolid shell, piriform. Prodissconch, ovate with straightened dorsal margin and inflated umbo, length 470 μm. Dissoconch piriform with slightly raised umbo near anterior end on dorsal margin. Posterodorsal margin concave near umbo, convex posteriorly, straight ventral margin, maximum convexity at anterior end. Sculpture fine irregular growth lines. Maximum length 52 mm.
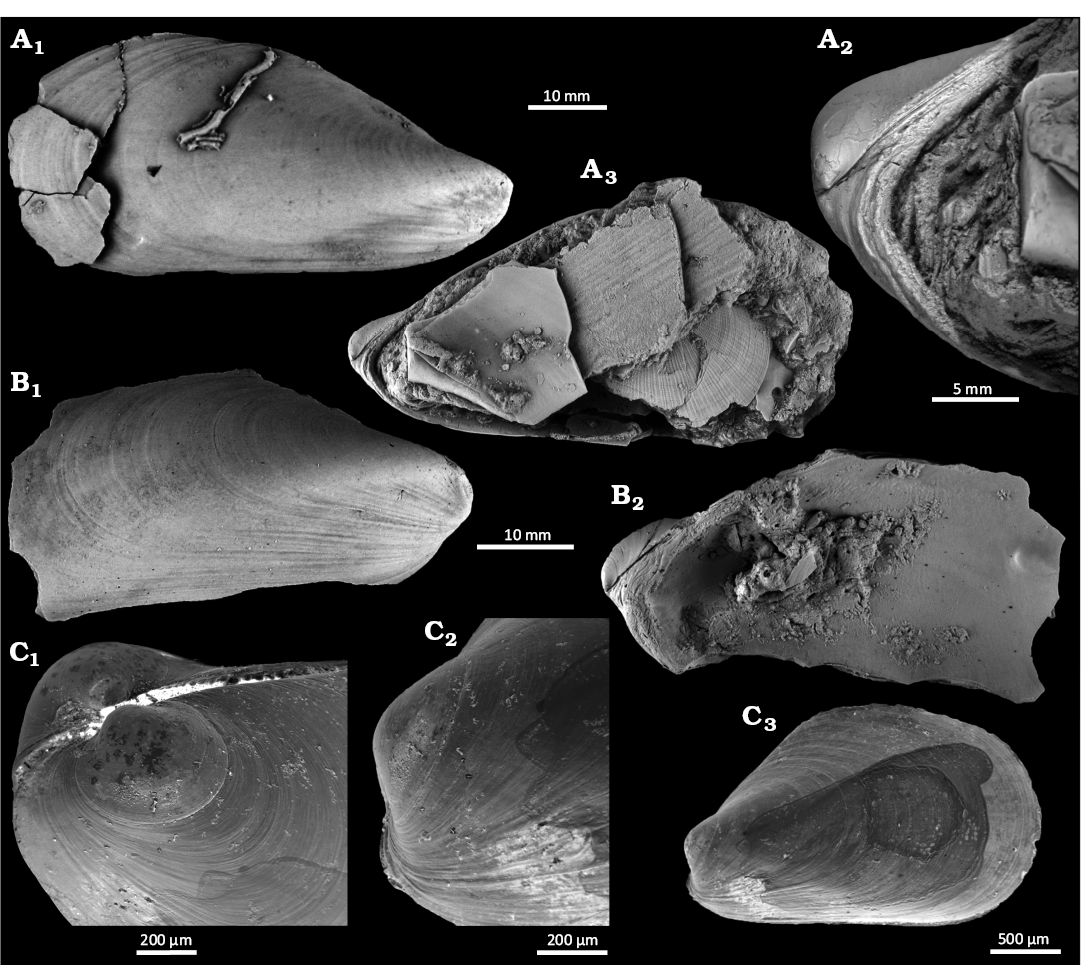
Fig. 15. The bathymodiolin bivalve Gigantidas cf. platifrons (Hashimoto & Okutani, 1994) from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373094, right valve external view (A1), internal umbonal detail (A2), and internal view (A3). B. SMF373100, right valve external (B1) and external (B2) views. C. SMF373099, articulated juvenile specimen, oblique dorsal view showing prodissoconchs (C1), left valve detail of umbo (C2), and left valve (C3).
Remarks.—Our species is close in shape to Gigantidas platifrons Hashimoto & Okutani, 1994, which was described from chemosynthetic environments off Japan. However, the umbones in the Japanese specimens are at the anterior end (Hashimoto and Okutani 1994), whereas our shells have umbones protruding above the anterior-dorsal end. Gigantidas niobengalensis Oliver et al., 2024, has been described from an active cold seep in the Krishna-Godavari Basin, depth 1750 m. This species has its umbo located anteriorly on the dorsal margin and has a convex anterior margin and a straightened posterior-dorsal margin (Oliver et al. 2024: holotype figs. 3A–C). Our species has a terminal umbo at the pointed anterior end and its posterior-dorsal margin is convex. For these reasons we think our fossil specimens do not belong to Gigantidas niobengalensis, although we note there is quite a lot of morphological variation in bathymodiolin species. The type species of Gigantidas has large narrowly elongated shells with protruding umbones on the dorsal margin (Cosel and Marshall 2003). Currently, the genus contains 11 species (MolluscaBase 2024) with a wide morphological variation; ranging from typical modioliform shape towards the elongated shape of the type species. The juvenile shell (Fig 13C1) of the fossil Gigantidas cf. platifrons specimens has a typical modiolid outline and the large prodissoconch (length 470 μm) confiming a planktonic stage.
Order Pectinida Gray, 1854
Superfamily Pectinoidea Rafinesque, 1815
Family Propeamussiidae Abbott, 1954
Genus Parvamussium Sacco, 1897
Type species: Pecten duodecimlamellatus Bronn, 1831, by original designation; Cenozoic, Italy.
Parvamussium? aff. scitulum (Smith, 1885)
Fig. 16A, B.
Material.—One valve, SMF373093; one valve, SMF376540; one valve, SMF373094. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.18–16.28 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1.
Description.—Small fragile, thin, flat, pectiniform shell. Right valve flattened with regular commarginal growth lines and microscopic radial etchings; large anterior auricle with irregular growth lines and deep byssal notch; smaller posterior auricle. Left valve flattened smooth with irregular growth lines; large anterior auricle without byssal notch; smaller posterior auricle. Largest incomplete valve diameter 14 mm.

Fig. 16. Propeamussiid bivalves from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A, B. Parvamussium? aff. scitulum (Smith, 1885). A. SMF373093, left valve external view. B. SMF376540, left valve external view. C–F. Propeamussiidae? gen. et sp. indet. C. SMF373091, left valve external view. D. SMF373092, partial right valve external view. E. SMF373089, partial right valve, external view. F. SMF373090, partial right valve, internal view.
Remarks.—Parvamussium scitulum was reported from the South-West Indian Ocean (Dijkstra and Maestrati 2015) and the Red Sea (Dijkstra and Janssen 2013). Our species has a similar sculpture and outline but it is larger and margins of the auricles are aligned with anterior and posterior margins; the auricles of Parvamussium scitulum end at an angle to the dorsal margins. We refrain from describing a new species on the basis of our fragmentary material.
Propeamussiidae? gen. et sp. indet.
Fig. 16C–F.
Material.—One valve, SMF373091; one valve, SMF373092; one valve, SMF373089; one valve, SMF373090. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.18–16.48 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1.
Description.—Small fragile, thin, flat, pectiniform shell. Right valve flattened with regular commarginal growth lines and microscopic radial etchings; large anterior auricle with irregular growth lines and deep byssal notch; smaller posterior auricle. Left valve flattened smooth with irregular growth lines; large anterior auricle without byssal notch; smaller posterior auricle. Prodissoconch subcircular, convex, length 100 μm. Largest incomplete valve has a diameter of 3 mm.
Remarks.—Only fragments of smooth left and right valves were found. The placement in Propeamussiidae is speculative; an alternative placement in Pectinidae is feasible.
Infraclass Heteroconchia Gray, 1854
Order Lucinida Gray, 1854
Superfamily Thyasiroidea Dall, 1900 (1895)
Family Thyasiridae Dall, 1900 (1895)
Genus Conchocele Gabb, 1866
Type species: Conchocele disjuncta Gabb, 1866, junior synonym of Conchocele bisecta (Conrad, 1849), by monotypy; Miocene, Oregon, USA.
Conchocele sp.
Fig. 17A.
Material.—One valve, SMF373083. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1.
Description.—Medium-sized, solid, subcircular, thyasirid shell, inequivalve, equilateral. Large protruding umbo directed anteriorly, posteriorly two keeled radial ridges with sharp intermediate channel. Sculpture of coarse irregular growth lines and margins, a small semi-circular lunule and a long deep escutcheon demarcated by a sharp ridge. Hinge with elongated grooves both anteriorly and posteriorly. Length of largest incomplete valve 29 mm
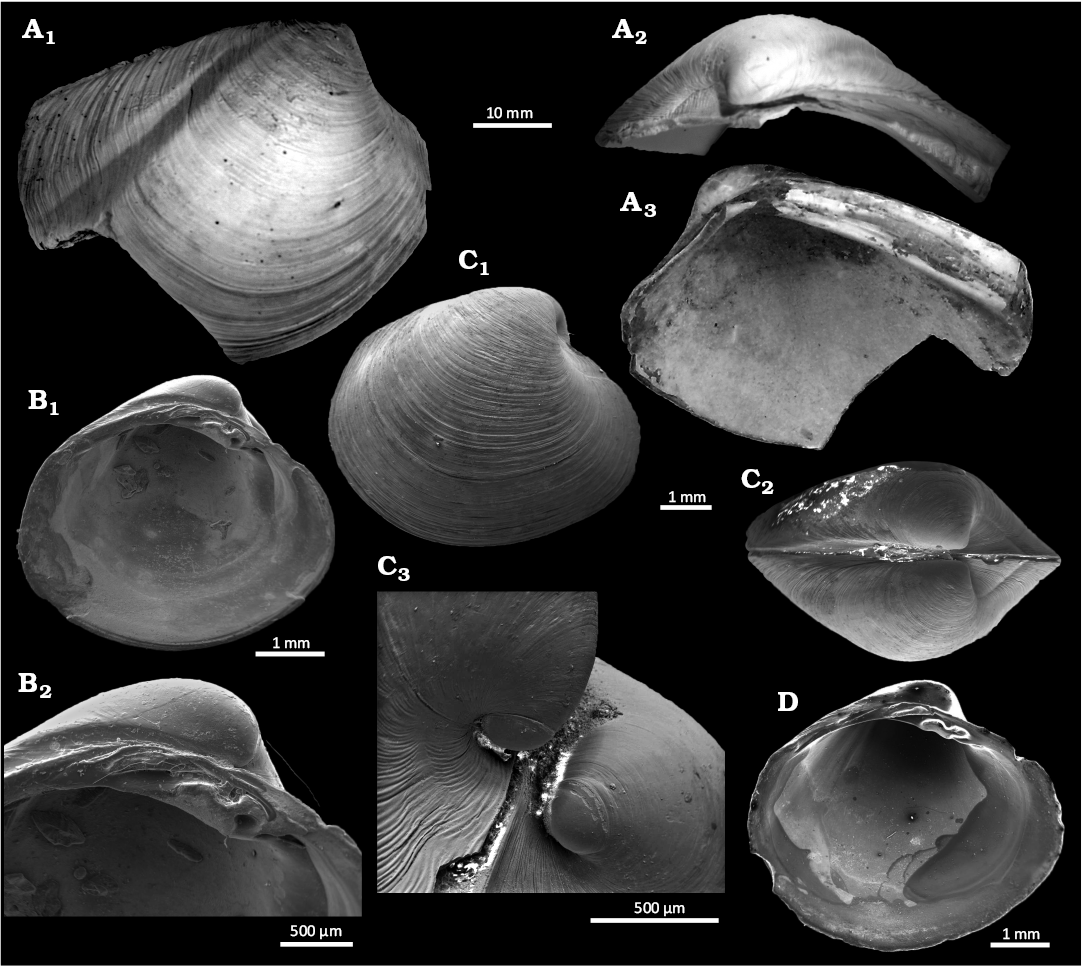
Fig. 17. Bivalves from Upper Pleistocene seep deposits from cores NGHP-01-12A and NGHP-01-10D, Krishna-Godavari Basin, offshore eastern India. A. Thyasirid Conchocele sp., SMF373083, right valve external (A1), dorsal (A2), and internal (A3) views. B–D. Vesicomyid Vesicomya prashadi Hoffman & Little sp. nov. B. SMF373072, paratype, left valve internal view (B1), internal view hinge detail (B2). C. SMF373073, holotype, articulated specimen right valve (C1), dorsal view (C2), detail of umbonal area from anterior (C3), prodissoconch. D. SMF373074, paratype, left valve internal view.
Remarks.—We refrain from formally naming this species based on the small number of fragmentary specimens. Our species is similar to the Miocene Conchocele majimai Kiel et al., 2020, and Conchocele visayaensis Kiel et al., 2020, from the Philippines (Kiel et al. 2020); both these species have a large keeled area surrounding the lunule anteriorly; the lunule in our species is small without a surrounding demarcated area. Another similar thyasirid species is Ascetoaxinus ravichandrani Ravinesh et al., 2024, that was described off Tamil Nadu, southern India, depth 1007–1038 m. This species has narrow sulci placed more posteriorly and its lunule is demarcated by a coarsely sculptured margin (Ravinesh et al. 2024: figs. 3–5). Our species has broader sulci starting immediately behind the umbo and the demarcation of the lunule is smooth. Since we have only few fragmentary shells, we cannot rule out that the species belongs in the genus Ascetoaxinus Oliver & Frey, 2014.
Order Venerida Gray, 1854
Superfamily Glossoidea Gray, 1847 (1840)
Family Vesicomyidae Dall & Simpson, 1901
Subfamily Vesicomyinae Dall & Simpson, 1901
Remarks.—Currently, the subfamily only contains the genus Vesicomya Dall, 1886. Species in Vesicomyinae are not known to harbour chemosymbiotic bacteria (e.g., Krylova et al. 2018).
Genus Vesicomya Dall, 1886
Type species: Callocardia atlantica Smith, 1885, by original designation; Recent, Atlantic.
Vesicomya prashadi Hoffman & Little sp. nov.
Fig. 17B–D.
Zoobank LCID: urn:lsid:zoobank.org:act:8D92F14F-BB4A-41D9-92 BF-F326FA809A7F.
Etymology: After Baini Prashad (1894–1969), Indian malacologist, particularly known for his study of the deep-water bivalves from the Siboga expedition (Prashad 1932).
Type material: Holotype, one shell, SMF373074. Paratypes, one valve, SMF373072; one valve, SMF373073, from the type locality and horizon.
Type locality: Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth.
Type horizon: 16.38–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Material.—Type material and several specimens from core NGHP-01-12A (for details see Table 1).
Diagnosis.—Small, solid, smooth, equivalve, inequilateral shell with oval outline, convex on all sides, beaks anteriorly, demarcated lunule and escutcheon, smooth prodissoconch.
Dimensions.—Holotype height 5.0 mm, length 6.0 mm, tumidity 4.2 mm. Maximum dimensions of shells in type set height as in holotype.
Description.—Prodissoconch single stage evident only, subcircular, truncated dorsally, flattened, smooth. Transition to dissoconch clear by change in sculpture. Length 300 μm. Dissoconch widely convex with prosogyrous umbos at 30% from anterior end; moderately inflated. Posterio-dorsal margin straightened, slightly convex, obliquely truncated posteriorly; ventral margin widely convex; anterior most convex; anterio-dorsal margin with indent at lunule; external sculpture with numerous fine growth margins of variable strength. Long lanceolate escutcheon, with fine growth lines, large lanceolate area for external ligament. Large triangular lunule with indented margins, with somewhat regular coarse growth lines. Ventral margin sharp, dorsal margin blunt. Internally surface and internal margin smooth; muscle scars and pallial line inconspicuous. Adductor scars of equal size; pallial line with minor sinus. Hinge only known from left valve; one curved elongated tooth posterior of umbo, elongated cavity for tooth right valve below; a second inverted w-shaped tooth below umbo, a short cavity for tooth right valve below posterior end w-tooth; one weak lateral tooth below posterio-dorsal margin; no anterior lateral tooth.
Remarks.—The genus Vesicomya has been based on the morphology of dentition in the left valve. The proximal curved tooth and the, long, more distal flexuous tooth Vesicomya prashadi Hoffman & Little sp. nov. is most similar to the dentition in the type species Vesicomya atlantica (Krylova et al. 2018: fig. 2C: teeth 2a, b, 4b) or that of the congener Vesicomya alleni (Krylova et al. 2018: fig. 3D). The type species of Vesicomya has a thin shell and narrow hinge plates whereas Vesicomya prashadi Hoffman & Little sp. nov. has a thick shell and heavy hinge plates. Vesicomya alleni has nearly identical teeth and hinge plate in the left valve but its shell is more elongated and the umbo is located further to the anterior (Krylova et al. 2018). Vesicomya prashadi Hoffman & Little sp. nov. has a more elevated outline and its umbos are in a nearly central position. Vesicomya indica (Knudsen, 1970) is known from abyssal depths in the Bay of Bengal (Huber 2010). That species has similar teeth compared to Vesicomya prashadi sp. nov but its hinge plate is narrower, its triangular outline is more elevated (height/length ±1) and the lunule is rounded. The escutcheon is also less pronounced; the co-marginal sculpture is fine and regular. Vesicomya prashadi Hoffman & Little sp. nov. is more flattened with a more triangular lunule, a pronounced escutcheon, an elongate-oval outline and a more irregular growth sculpture.
We considered placement of Vesicomya prashadi Hoffman & Little sp. nov. in other genera of small vesicomyids, for example Waisiuconcha Beets, 1942, and Isorropodon Sturany, 1896. Waisiuconcha was rejected as the dentition of the type species Waisiuconcha alberdinae Beets, 1942, are clearly different (Beets 1942: figs. 147–148: thick strong teeth). The same character is observed in, for example, the Atlantic Waisiuconcha haeckeli Cosel & Salas, 2001 (Zettler and Hoffman 2021: figs. 4, 12). We also rejected placement in Isorropodon because species in this genus have elongated, dorsally aligned teeth, as in evident in the original illustrations of the type species Isorropodon perplexum Sturany, 1896 (Sturany 1896: pl. 1: 24–27).
Stratigrapic and geographic range.—Type locality and horizon only.
Subfamily Pliocardiinae Woodring, 1925
Remarks.—The subfamily currently contains 19 genera. Species from all genera in Pliocardiinae are known to harbour chemosymbiotic bacteria (e.g., Krylova et al. 2018). In the Krishna-Godavari Basin fossil material species in the genera Pliocardia Woodring, 1925, Callogonia Dall, 1889a, and Archivesica Dall, 1908a, are present.
Genus Pliocardia Woodring, 1925
Type species: Anomalocardia bowdeniana Dall, 1903, by original designation; Pliocene, Jamaica.
Pliocardia cf. solidissima (Prashad, 1932)
Fig. 18A.
Material.—One valve (SMF373088) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.68 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Medium-sized, solid vesicomyid shell, equivalve, inequilateral. Outline oblong-ovate, strongly protruding umbo directed anteriorly, antero-dorsal margin concave, postero-dorsal and ventral margins weakly convex, anterior margin most convex, posterior widely convex. Sculpture with numerous irregular coarse growth lines. Elongated yet widely oval lunule, narrow elongated escutcheon. Internally, large teeth and hinge plate, pallial sinus absent. Length 36 mm.
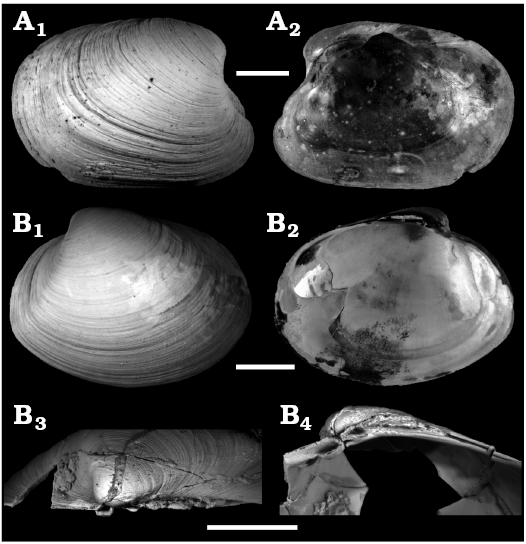
Fig. 18. Vesicomyid bivalves from core NGHP-01-12A and NGHP-01-10D, Krishna-Godavari Basin, offshore eastern India. A. SMF373088, Pliocardia cf. solidissima (Prashad, 1932), right valve external (A1) and internal (A2) views. B. SMF373082, Callogonia cf. leeana (Dall, 1889a), left valve external (B1) and internal (B2) views, partial right valve dorsal (B3), and internal (B4) views. Scale bars 10 mm.
Remarks.—Prashad’s (1932) species is determined by its medium size, thick globose shell, elongated trapezoidal outline, demarcated lunule, hinge and pallial line without sinus. The type locality is from off Bali, Indonesia (SIBOGA sta. 18, 7°28.2’ S, 115°24.6’ E, 1018 m). A similar species is Pliocardia kuroshimana (Okutani et al., 2000), which is known in the Western Pacific off Japan (Okutani et al. 2000) and in the Miocene on Leyte, Philippines (Kiel et al. 2020). Another similar species is Pliocardia atalantae (Cosel & Olu, 2009) from off western Africa. Pliocardia indica (Smith, 1904) also lives in the Bay of Bengal (Smith 1904: 9; Huber 2010) in 767–1985 m. Pliocardia indica has a convex ventral margin, whereas our specimen has a flattened ventral margin and a trapezoidal outline.
Genus Callogonia Dall, 1889a
Type species: Callocardia (Callogonia) leeana Dall, 1889a; Recent, North-West Atlantic.
Callogonia cf. leeana (Dall, 1889a)
Fig. 18B.
Material.—One shell, SMF373082. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.8647’ N, 81°50.0709’ E, 1050 m water depth, 15.73 mbsf level, core NGHP-01-10D; Upper Pleistocene.
Description.—Medium-sized, fragile vesicomyid shell, equivalve, subequilateral. Outline ovate, strongly protruding umbo directed anteriorly, all margins convex, anterior and posterior weakly truncated obliquely. Sculpture with numerous irregular coarse growth lines. Lunule not conspicuous, narrow elongated escutcheon. Internally, small elongated teeth, thin hinge plate, pallial sinus. Length 38 mm
Remarks.—Outline, sculpture, hinge and pallial sinus are similar to Callogonia leeana (Dall, 1889a) from the NW Atlantic; even though there is a morphological similarity, it may be a different species in view of the large oceanic distance to the type locality. The left valve of the present species shows a clear sinus in the pallial line, which is characteristic for the genus.
Genus Archivesica Dall, 1908b
Type species: Callocardia gigas Dall, 1896, by original designation; Recent, E Pacific.
Archivesica cf. kawamurai (Kuroda, 1943)
Fig. 19.
Material.—One juvenile shell, SMF373080; one adult valve, SMF373081; one adult valve, SMF373078; one adult valve, SMF373079; one juvenile valve, SMF373077. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.18–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1.
Description.—Large, solid vesicomyid shell, equivalve, inequilateral. Outline oblong-ovate, weakly protruding umbo at 25% from anterior margin, all margins convex, anterior and posterior most convex. Sculpture with numerous irregular coarse growth lines. Lunule not conspicuous, long lanceolate escutcheon. Length largest valve 43 mm
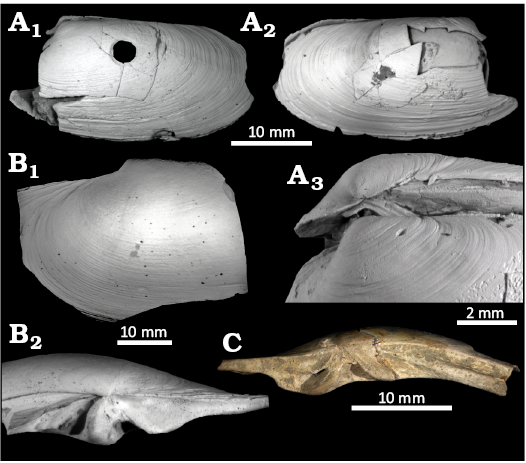
Fig. 19. The vesicomyid bivalve Archivesica cf. kawamurai (Kuroda, 1943) from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373080, articulated specimen, left valve with Oichnus paraboloides Bromley, 1981 boring (A1), right valve (A2), and oblique dorsal view showing right valve hinge (A3). B. SMF373081, left valve external (B1) and hinge (B2) views. C. SMF373078, right valve hinge view. A, B whitened and C not whitened.
Remarks.—The large size, outline, sculpture and hinge of our specimens suggest that they are probably conspecific with Archivesica kawamurai, a species described off Japan, Sagami Bay (Kuroda 1943); the species also known from the Pliocene and Recent from various locations around Japan (Kuroda 1943; Amano and Kiel 2010: 158) and around Taiwan (Kiel et al. 2024: 10, figs. 8P–R). Archivesica kawamurai can be identified by the hinge and the widely convex ventral margin, a feature found in the holotype (Amano and Kiel 2010: 156–158, fig. 18). Many species in Archivesica have a straightened or slightly concave ventral margin. A morphologically similar species to our specimens is Archivesica arctica Hansen et al., 2017, from off NW Svalbard; this species has a similar outline and hinge, and its anterior margin seems less convex (Hansen et al. 2017). Calyptogena marissinica Chen et al., 2018, has a similar outline to our specimens, but this species has a different hinge with symmetrical cardinal teeth 2a, b of equal strength; our species has a strong 2b tooth and a weak 2a tooth, which are placed obliquely (Chen et al. 2018: figs. 2, 3).
Superorder Analodesmata Dall, 1889b
Family Cuspidariidae Dall, 1886
Cuspidariidae? gen. et sp. indet.
Fig. 20.
Material.—One fragmentary valve (SMF373084) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene.
Description.—Small fragile, flattened shell, inequilateral. Outline uncertain. Sculpture with fine radial and commarginal lines. Internal hinge with deep notch in external margin below the umbo, no teeth seen. Length about 3 mm.
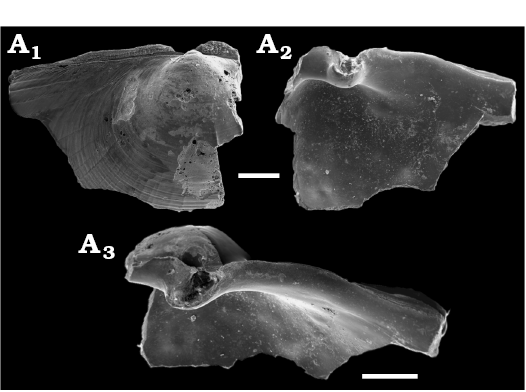
Fig. 20. The bivalve Cuspidariidae? gen. et sp. indet. from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A. SMF373084, partial right valve external (A1), internal (A2), and oblique dorsal (A3) views. Scale bars 500 µm.
Remarks.—Placement in Cuspidariidae is feasible from the external morphology and hinge. The radial sculpture suggests a species in the genus Cardiomya Adams, 1864, albeit that our species has relatively flat valves and known species in the Indo-Pacific are rather globose. Alternatively, it could be placed in another genus in Cuspidariidae or Protocuspidariidae. The material was deemed inadequate for a determination, or description as a new species.
Class Scaphopoda Bronn, 1862
Order Dentalida Starobogatov, 1974
Family Dentaliidae Children, 1834
Dentaliidae? gen. et sp. indet.
Fig. 21A–C.
Material.—Three shells (SMF373085–87) from Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.78 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1.
Description.—small, weakly curved, solid scaphopod tooth-shaped shell with strong axial rib sculpture. small, globose, solid inflated gadilid-shaped shell. Apex small, pointed with circular hole, rectangular outline. Intial strong increase in diameter, less thickening in adult stage. Sculpture with large rounded axial ribs, apically 4 ribs increasing to 9 ribs abapically with incremental ribs progressively formed in between. Maximum length 7 mm.
Remarks.—Placement in Dentaliidae is plausible from the external morphology and outline in the few fragmentary shells. They likely belong to a single species in the genus Antalis Adams & Adams, 1854. Alternatively, they could be placed in another genus in Dentaliidae or in the genus Entalina Monterosato, 1872, in family Entalinidae Chistikov, 1979. The material was deemed inadequate for a determination, or description as a new species.
Order Gadilida Starobogatov, 1974
Family Gadilidae Stoliczka, 1868
Genus Cadulus Philippi, 1844
Type species: Cadulus ovulum (Philippi, 1844)†; Miocene, Sicily, Italy.
Cadulus chuni Jaeckel, 1932
Fig. 21D, E.
1932 Cadulus chuni Jaeckel, 1932: 309, text-fig. 6.
1995 Cadulus chuni Scarabino, 1995: 348–349, figs. 153, 160e.
Material.—One shell, SMF373076; one shell, SMF373075. Northern Indian Ocean, Krishna-Godavari Basin, Bay of Bengal, offshore eastern India, 15°51.6335’ N, 81°50.2274’ E, 1046 m water depth, 16.78–16.88 mbsf level, core NGHP-01-12A; Upper Pleistocene. For additional material see Table 1.
Description.—Small, smooth, swollen, solid gadilid-shaped shell. Apical aperture circular with thick rim. externally 0.6 mm, internally 0.4 mm; abapical aperture circular with thin rim, external diameter 0.9 mm. Lateral outline convexly swollen centre, slightly concave on apical and abapical shell faces. Maximum length 3 mm.
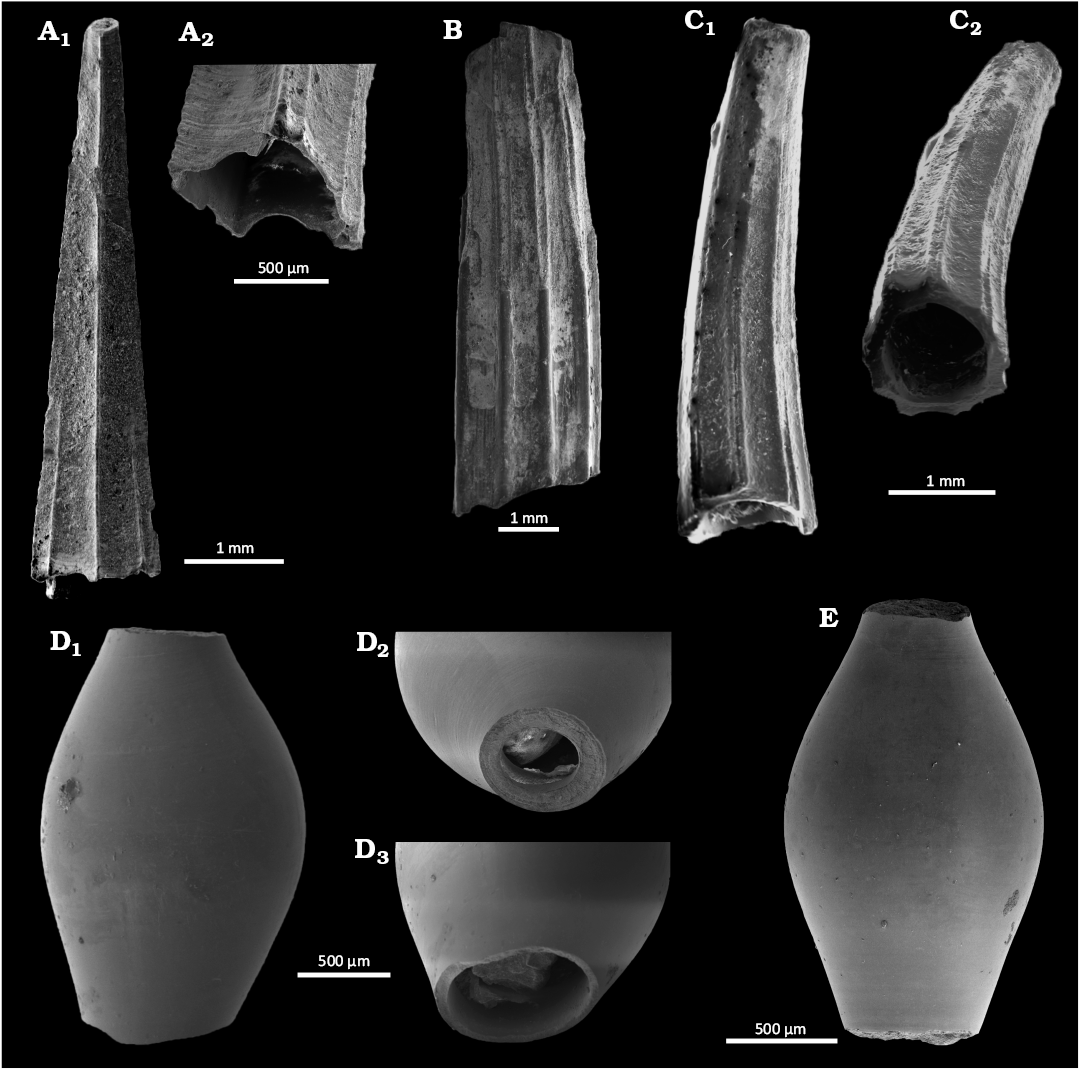
Fig. 21. Scaphopods from Upper Pleistocene seep deposits from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A–C. Dentalidae? gen. et sp. indet. A. SMF373085, side view (A1) and oblique view of aperture (A2). B. SMF373086, side view. C. SMF373087, side view (C1), oblique view of aperture (C2). D, E. Gadilid Cadulus chuni Jaeckel, 1932. D. SMF373076, side view (D1), abapical (D2), and apical (D3) openings. E. SMF373075, dorsal view.
Remarks.—The species was described from off Somalia, eastern Africa, “Valdivia”, station 256, 01°49’ N, 45°30’ E, 1134 m. It is known as a recent species from the type locality and two locations in the Western central Pacific from Indonesia and the Philippines (Scarabino 1995: 349, fig. 153); the bathymetric range is 205–1134 m.
Stratigraphic and geographic range.—Recent, off Somalia, Philippines and Indonesia. Upper Pleistocene, Krishna-Godavari Basin, off eastern India.
Discussion
Palaeocology of the Upper Pleistocene Krishna-Godavari Basin seep assemblage.—The majority of the studied specimens come from the shell-rich horizon in core NGHP-01-12A situated between 16.18 mbsf and 17 mbsf (Table 1). Based on 14C dating the age range of this material spans at least 10 000 years, and thus should be considered an assemblage of fossils representing a series of living seep communities. Nonetheless, the species composition seems to have been similar, so there is no obvious signature of changing community structure over time. Although the material from core NGHP-01-10D is undated, Mazumdar et al. (2019) report from a nearby sediment core U-Th based dates of carbonate tubules of ~46.2 ± 3.7 to 53 ± 1.6 ka at 16 mbsf. Based on these various dates Teichert et al. (2014; fig. 10) correlated the MDAC and shell-rich horizons between NGHP-01-12A and NGHP-01-10D. Consequently, we here consider the material present between 16.18 mbsf and 17 mbsf in the former and 15.3 mbsf and 15.73 mbsf in the latter to be roughly contemporary, and, therefore, below consider them as a single fossil seep assemblage.
The seep gastropod, bivalve and scaphopod assemblage comprises 29 species: 15 gastropods, 12 bivalves, and two scaphopods. Of these species nine are identified as new: six gastropods and three bivalves (Table 1). A comparison with living conspecifics and congenerics (e.g., Dubilier et al. 2008) suggests that six of these species are likely to have had chemosymbionts (Acharax sp., Gigantidas cf. platifrons, Conchocele sp., Pliocardia cf. solidissima, Callogonia cf. leeana, and Archivesica cf. kawamurai). These represent 21% of the diversity, but only 14% of the specimens complete enough for identification (Tables 1, 2). However, shell fragments of bathymodiolins and vesicomyids are very common in most of sediment sub-samples, suggesting that originally these taxa represented a larger proportion of the molluscan faunal abundance in the seep communities. With the exception of Acharax sp., all these putative chemosymbiotic taxa were likely to have been obligate to seeps, based on the occurrence of living conspecifics and congenerics (e.g., Imhoff et al. 2003; Sasaki et al. 2005; Krylova and Sahling 2010; Lorion et al. 2013; Zanzerl et al. 2019). Most occurrences of Acharax species are from seeps (e.g., Neulinger et al. 2006; Mazumdar et al. 2019; Yang et al. 2024), but they are also known from organic-rich sediments associated with oxygen minimum zones, for example off Baja California (Suárez-Mozo et al. 2019), so this taxon is not always obligate to the seep environment. Obligacy is likely to have been the case for Paralepetopsis bathyalus Hoffman & Little sp. nov., as most living Paralepetopsis species are restricted to either seep or hydrothermal vent habitats (e.g., McLean 1990; Warén and Bouchet 2001, 2009). This could possibly also have been the case for Anatoma sahlingi Hoffman & Little sp. nov., as a small number of living Anatoma species have been reported from hydrothermal vent sites in the central Indian Ocean and not elsewhere (Hoffman et al. 2022). However, anatomids are found in a large variety of habitats from intertidal to deep bathyal depths in all oceans of the world (Geiger 2012). Including this taxon gives a value for obligacy in the Krishna-Godavari Basin seep assemblage diversity of 24% (Table 2). The remaining bivalve, gastropod and scaphopod species described here also have living relatives common in bathyal habitats and can probably be considered as facultative or “background” fauna in the assemblages (e.g., Scarabino 1995; Dijkstra and Janssen 2013; Dijkstra and Maestrati 2015; Krylova et al. 2018). They represent a wide variety of infaunal and epifaunal palaeoecologies, ranging from less specialized grazers to specialized microcarnivores (Table 2). Additional palaeoecological information comes from the small number of penetrative borings present in vesicomyid shells (Fig. 19A1) and neolepadid plates (Fig. 22D). These are consistent with the ichnotaxon Oichnus paraboloides Bromley, 1981, which is commonly ascribed to the predatory drilling activities of naticid and muricid gastropods (e.g., Breton et al. 2017). Intriguingly, neither taxon is present as shells in the Krishna-Godavari Basin seep assemblage (Table 1).

Fig. 22. Trace fossils from Upper Pleistocene seep deposit from core NGHP-01-12A, Krishna-Godavari Basin, offshore eastern India. A, B. Fungal microborings. A. SMF373120, external surface of Cirsonella aperta Hoffman & Little sp. nov. (see Fig. 8C). B. SMF373062, bivalve shell fragment. C. Grazing trace Radulichnus inopinatus Voigt, 1977, on vesicomyid bivalve shell fragment. D. NHMUK IC 1416, drillhole Oichnus paraboloides Bromley, 1981, boring in upper latus plate of Ashinkailepas indica Gale in Gale et al., 2020.
Undoubtedly, both the species and palaeoecological diversity in the fossil seep assemblage was enhanced by the presence of abundant hard substrate, as shells and MDAC, in what was otherwise a fine-grained sedimentary environment. These hard substrates served as attachment sites for a diversity of epifauna, including the byssate bivalves (i.e., Gigantidas cf. platifrons, and probably the pectinids), corals (Caryophyllia sp.), serpulids and neolepadids (Fig. 2), sponges (suggested by the presence of spongivorous fissurellids), as well as, presumably, for the larvae of these and many other taxa. The bivalve and gastropod shells also acted as substrates for fungal microborings (Fig. 22A, B) and were grazed extensively, as inferred from the presence of Radulichnus inopinatus Voigt, 1977, trace fossils on many of the larger bivalve shell fragments (Fig. 22C), which probably resulted from gastropod and/or chiton grazing (e.g., Voigt 1977; De Gibert et al. 2007).
Material from deeper in core NGHP-01-10D (between 41.54 mbsf and 42.27 mbsf, and at 61.03 mbsf) represents older fossil assemblages. The presence of MDAC at these levels (Teichert et al. 2014), fragments of large vesicomyid shells and a single, probable Gigantidas cf. platifrons shell at 42.01 mbsf (Table 1) strongly suggests that hydrocarbon seepage and associated faunas were present at this site before ~52 ka. A using the 14C ages from Teichert et al. (2014) for core NGHP-01-12A to calculate a sedimentation rate (1 m per 3.25 ka) would give an age estimate of ~136 ka for the shell at 42.01 mbsf in core NGHP-01-10D.
Comparison with other putative fossil seep assemblages from offshore eastern India.—In addition to the material reported here there are several other likely occurrences of fossil seep assemblages from the Krishna-Godavari Basin. Mazumdar et al. (2009: fig. 4c) figure articulated and fragmented bivalves shells identified as Calyptogena sp. from 16 mbsf to 18 mbsf in a sediment core (MD161-8) recovered during cruise MD-161 of the R/V Marian Dufresne (May 2007). This site is close to NGHP-01-10D at 15°51.8624’ N, 81°50.0692’ E in a water depth of 1033 m (Fig. 1). The figured shells do not show the internal features that are needed for generic determination of vesicomyid genera, but the overall elongation and rounded posterior margin of the specimens are similar to the Archivesica cf. kawamurai specimens described here, so may be conspecific. Additional bivalve shell fragments from the same cruise were figured by Joshi et al. (2014) at 27–31 mbsf in sediment core MD161-15, recovered from 983 m water depth. These comprise one anterior portion of a shell identified as Calyptogena sp. (Joshi et al. 2014: fig. 9a, b) and the ventral portion of a shell identified as Lucinoma sp. (Joshi et al. 2014: fig. 9c). None of this material is complete enough for generic determination, but the shape of the vesicomyid shell is similar to the Archivesica cf. kawamurai specimens herein, and the “Lucinoma sp.” shell bears some resemblance to our Conchocele sp. specimens. Joshi et al. (2014) state that scaphopods are incorporated into MDAC from this core. One specimen (Joshi et al. 2014: fig. 8v, vi) shows closely spaced longitudinal ribbing, so is likely to be a dentaliid, but being considerably larger than any of the specimens herein, cannot be compared directly with them. According to Joshi et al. (2014) age of the shells from MD 161-15 is undetermined.
Badesab et al. (2020) figure fragmentary bivalve shells from a short sediment core (SSD-045/Stn-9/GC-01) recovered from the Krishna-Godavari Basin during cruise SSD-045 of the R/V Sindhu Sadhana at 16°18.6027’ N, 82°37.9809’ E in 1 673 m water depth (Fig. 1). Associated with MDAC Badesab et al. (2020) identified Calyptogena sp. from 2.7 mbsf and 5.1 mbsf (Badesab et al. 2020: fig. 8j, k). Again, these shells are too incomplete for generic determination, although they are most similar in size and shape to our Archivesica cf. kawamurai specimens. The shells recorded in Badesab et al. (2020) are likely to be considerably younger than our Krishna-Godavari Basin seep specimens, as the former come from much shallower core depth horizons.
Comparison of the Krishna-Godavari Basin Upper Pleistocene seep assemblage with modern seep communities in the Indian Ocean.—Until all the taxa present in the modern seep communities from Krishna-Godavari Basin and Mannar Basin have been formally described it will be difficult to make definitive species comparisons between these and those present in the Upper Pleistocene Krishna-Godavari Basin seep assemblage. However, using the figures in Mazumdar et al. (2019), Dewangan et al. (2021) and Sangodkar et al. (2022), and the identifications of He et al. (2023) (which were based on the images in the above publications) for the Krishna-Godavari Basin and Mazumdar et al. (2021) for the Mannar Basin seeps allows us to make some initial comparisons.
Based on the seafloor photographs in Mazumdar et al. (2019: fig. 3a) and Dewangan et al. (2021: fig. 5a) the Krishna-Godavari Basin seep communities appear to be dominated by bathymodiolins and vesicomyids. The bathymodiolin specimen figured in Sangodkar et al. (2022: fig. 3a) looks similar to our Gigantidas cf. platifrons, so could be conspecific, but is also very similar to Gigantidas niobengalensis Oliver et al., 2024. The bathymodiolin images in Mazumdar et al. (2019: fig. 3a,q) and Dewangan et al. (2021: fig. 5a) are probably also the same species, but the images are not detailed enough to be sure. It is worth noting here that several superficially similar bathymodiolin species can be present in modern seeps, for example, the Haima Seep in the South China Sea has four co-occurring species (Lin et al. 2022). The large vesicomyid species figured in Mazumdar et al. (2019: fig. 3b, c) differs from the Archivesica cf. kawamurai specimens herein in being shorter and having a slightly concave ventral margin, so probably represents a different species. This is also the case for the vesicomyids figured in Sangodkar et al. (2022: fig. 3b) and Dewangan et al. (2021: fig. 5d). He et al. (2023) identify this taxon as Calyptogena makranensis.
Solemyids, thyasirids and pectinoids are present in the modern Krishna-Godavari Basin seep communities. The Archarax sp. figured in Mazumdar et al. (2019: fig. 3e) and Sangodkar et al. (2022: fig. 3c) look to be the same specimen. The Archarax sp. figured in Dewangan et al. (2021: fig. 5c) is probably the same species. It is likely that our Acharax sp. is conspecific with these modern specimens, but the fossil material is too fragmentary to be sure. The Conchocele sp. specimens figured in Mazumdar et al. (2019: fig. 3d) and Dewangan et al. (2021: fig. 5b) are probably the species as the Thyasiridae gen. et sp. indet. in Sangodkar et al. (2022: fig. 3d). These specimens have a slightly different shape to our Conchocele sp. in having a less projecting anterior margin and a deeper posterior sulcus. However, these differences may be ontogenetic, as the fossil specimens are considerably smaller than the modern figured ones. Mazumdar et al. (2019: fig. 3f) identify a ribbed pectinoid bivalve from the modern Krishna-Godavari Basin seeps as belonging to the Pectinidae. He et al. (2023) identify this as Catillopecten. This species could be conspecific with our Parvamussium aff. scitulum, although the image of the modern taxon is not good enough to make a definitive comparison. Gastropods are also present in the modern Krishna-Godavari Basin seep fauna. The limpets shown in Mazumdar et al. (2019: fig. 3g, h) could be conspecific with our Paralepetopsis bathyalus Hoffman & Little sp. nov. The “Phenocolepadidae” in Sangodkar et al. (2022: fig. 3m), which He et al. (2023) identify as Bathyacmaea, is a more questionable comparison to either of our fossil limpet species.
Molluscan taxa in the fossil seep assemblage that cannot be easily compared to taxon descriptions or figures in Mazumdar et al. (2019), Dewangan et al. (2021) or Sangodkar et al. (2022) are the gastropods Mesopelex godavariensis Hoffman & Little sp. nov., Fissurellidae gen. et sp. indet., Anatoma sahlingi Hoffman & Little sp. nov., Seguenzioidea gen. et sp. indet., Cirsonella aperta Hoffman & Little sp. nov., Dikoleps? magnarota Hoffman & Little sp. nov., Alvania axistriata Hoffman & Little sp. nov., Benthonellania sp., Columbellidae gen. et sp. indet., Conoidea gen. et sp. indet., Eulimella sp. 1, Eulimella sp. 2, Odostomia sp., and Cylichnidae? gen. et sp. indet., the bivalves Ledella favus Hoffman & Little sp. nov., Yoldiella umbostriata Hoffman & Little sp. nov., Propeamussiidae? gen. et sp. indet., Vesicomya prashadi Hoffman & Little sp. nov., Pliocardia cf. solidissima (Prashad, 1932), Callogonia cf. leeana (Dall, 1889a), and Cuspidariidae? gen. et sp. indet., and the scaphopod Cadulus chuni Jaeckel, 1932. Most of these are small taxa, so may have been overlooked in the samples of Mazumdar et al. (2019), Dewangan et al. (2021), and Sangodkar et al. (2022), or were present, and simply not reported in these publications. Alternatively, these fossil taxa are not present in the modern Krishna-Godavari Basin seep communities. Conversely, some of the gastropod taxa figured in Mazumdar et al. (2019), Dewangan et al. (2021), and Sangodkar et al. (2022) are not present in the fossil seep assemblage: neritids (Mazumdar et al. 2019: fig. 3j), “Provannidae” (Mazumdar et al. 2019: fig. 3i; identified as Phymorhynchus in He et al. 2023), and neither of the gastropods figured in Sangodkar et al. (2022: fig. 3o) as “gastropod unidentified spp.”, which appear to be a buccinid (image on right in fig. 3o) and provannids (three left images in fig. 3o). The latter are also identified as Provanna by He et al. (2023). The absence of Provannidae from the fossil seep assemblage is interesting, as this is one of the most characteristic and species-rich taxon in modern and many fossil seep communities (e.g., Warén and Bouchet 2001; Sasaki et al. 2010; Betters and Cordes 2023).
In the non-molluscan elements of the modern and fossil Krishna-Godavari Basin seep faunas there are also differences and similarities. Shared taxa include echinoids, which are present in the fossil Krishna-Godavari Basin assemblage as very occasional isolated test ossicles, and serpulids, the tubes of which are very commonly found attached onto bivalve shells and neolepadid barnacle plates in the fossil material (Figs. 2, 15A1), and are figured attached onto bivalve shells in the modern communities (Dewangan et al. 2021: fig. 5d). This is also the case for neolepadids in the modern seep communities (Mazumdar et al. 2019: fig. 3q; Dewangan et al. 2021: fig. 5a). These modern neolepadids have been identified as Neolepas sp. in Mazumdar et al. (2019), Dewangan et al. (2021), and Peketi et al. (2022), and Leucolepas in He et al. (2023). It is possible that these are conspecific with Ashinkailepas indica Gale in Gale et al., 2020, the isolated plates of which are very common elements in most of NGHP-01-12A core samples and from the 15.54 mbsf level in core NGHP-01-10D (Table 1).
The modern Krishna-Godavari Basin seep communities include soft bodied taxa that would not be expected to be present in a fossil assemblage for purely taphonomic reasons, such as non-tubicolous polychaete worms (Mazumdar et al. 2019: fig. 3l, m; Sangodkar et al. 2022: fig. 3f, g). This may also pertain to taxa with lightly mineralized exoskeletons, such as decapod crustaceans (Mazumdar et al. 2019: fig. 3n, o; Sangodkar et al. 2022: fig. 3e, n), which are not present in the fossil assemblage. Siboglinid tubeworms are absent from the fossil Krishna-Godavari Basin seep assemblage. Whilst this is expected for the soft tissues of these worms, the chitin-rich tubes are known to get replaced in modern seep sites by carbonates (Haas et al. 2009), and have a rich fossil record (Georgieva et al. 2019). It is possible that some of the tubular carbonates present in Krishna-Godavari Basin sediment cores reported in Kocherla et al. (2015) and Mazumdar et al. (2019) represent replaced siboglinid tubes. One more taxon not present in the fossil seep assemblage is the Ophiuroidea (Mazumdar et al. 2019: fig. 3p; Sangodkar et al. 2022: fig. 3k).
The Mannar Basin seep community includes siboglinids (as Lamellibrachia sp., Mazumdar et al. 2021: fig. 2), munidopsids (Mazumdar et al. 2021: fig. 4a–d), and gastropods and bivalves. Some of the illustrated siboglinid tubes have neolepadid specimens attached to them, identified as Neolepas sp. (Mazumdar et al. 2021: fig. 2c, d). The gastropods are large buccinoid species (Mazumdar et al. 2021: fig. 4e); the bivalves are bathymodiolins and vesicomyids, identified as Calyptogena sp. (Mazumdar et al. 2021: fig. 4f). Based on the external morphology of the illustrated specimen, this latter taxon looks similar to the large vesicomyids from the modern Krishna-Godavari Basin seep communities, but not elongated enough to be conspecific with our Archivesica cf. kawamurai specimens.
Some of the taxa characteristic of deep-water seep communities are shared between the Krishna-Godavari Basin fossil seep assemblage and the modern seeps elsewhere in the Indian Ocean, at least at higher taxonomic levels. In the seeps off Indonesia shared taxa include serpulids and mytilids in the Semeulue seep community off northern Sumatra, vesicomyids, thyasirids and Acharax at the Snail Hill seep in the Sunda Arc, and Archivesica garuda (Okutani & Soh, 2005) at the seep off eastern Java. The latter species is not the same as our Archivesica cf. kawamurai, which is interesting, as Archivesica kawamurai is a species common in modern and fossil (Pliocene) seeps from Japan (Amano and Kiel 2010), geographically considerably further away from India than the Indonesian seeps. There are also shared higher taxa between the Krishna-Godavari Basin fossil seep assemblage and the modern seeps on the Makran accretionary margin below 1000 m water depth. These are neolepadids, bathymodiolins, Archarax, and Calyptogena makranensis Krylova & Sahling, 2006.
The non-obligate molluscan taxa in the Krishna-Godavari Basin fossil seep assemblage comprises typical representatives of taxa found in upper bathyal zone sediments globally, some already known from around the Indian continental margin (e.g., Conidae, Acharax, Ledella, Yoldiella, Propeamussidae, Vesicomya), and some that where previously not recorded from this area (e.g., Mesopelex, Cirsonella, Cadulus, and possibly Dikoleps). However, the latter observation may reflect the general poor state of knowledge of Indian deep water molluscs. For example, the compilation in Rajendra et al. (2021; Table 1) lists only 25 deep sea bivalve, gastropod and scaphopod species.
In summary, the Upper Pleistocene seep assemblage was structured ecologically and taxonomically very like modern deep water seep communities, from offshore India and elsewhere in the Indian Ocean in containing a small number obligate taxa, most of which likely had chemosymbionts (e.g., Acharax, Gigantidas, Pliocardiinae, Conchocele, Ashinkailepas), as well as a greater diversity of morphologically smaller species that represent common deep-sea taxa, with a wide variety of palaeoecologies (Table 2). However, pending taxonomic work on the modern seep fauna, the significance of the fossil seep faunas in the Krishna-Godavari Basin for understanding community evolution and palaeobiogeography in hydrocarbon seep environments in the Indian Ocean will be limited. What is certain is that many more seep communities are to be identified in the Indian Ocean, both modern and fossil. In the Recent this is evidenced, for example, by the occurrence of probable seep obligate pliocardiins in deep water off Bali (Pliocardia solidissima [Prashad, 1932] at SIBOGA sta. 18; Prashad 1932). Additional fossil evidence comes from the presence of the shells of large bathymodiolins and pliocardiins, together with MDAC, in horizons of sediment cores both younger and older than the Upper Pleistocene, in the Krishna-Godavari Basin and Mannar Basin (e.g., Teichert et al. 2014; Mazumdar et al. 2019, 2021; Kocherla et al. 2015).
Table 2. Suggested palaeoecological determinations for bivalve, gastropod, and scaphopod taxa from cores NGHP-01-12A and NGHP-01-10D. CSV version of this table is available in SOM 3.
|
Taxon |
Occurrence |
Palaeoecology |
|
Paralepetopsis bathyalus Hoffman & Little sp. nov. |
obligate |
epifaunal grazer |
|
Mesopelex godavariensis Hoffman & Little sp. nov. |
facultative |
epifaunal grazer |
|
Fissurellidae gen. et sp. indet. |
facultative |
spongivore |
|
Anatoma bengalensis Hoffman & Little sp. nov. |
?obligate |
epifaunal grazer |
|
Seguenzioidea gen. et sp. indet. |
facultative |
epifaunal grazer |
|
Cirsonella aperta Hoffman & Little sp. nov. |
facultative |
epifaunal grazer |
|
Dikoleps? magnarota Hoffman & Little sp. nov. |
facultative |
epifaunal grazer |
|
Alvania axistriatus Hoffman & Little sp. nov. |
facultative |
epifaunal grazer |
|
Benthonellania sp. |
facultative |
epifaunal grazer |
|
Columbellidae gen. et sp. indet. |
facultative |
epifaunal predator/scavenger |
|
Conoidea gen. et sp. indet. |
facultative |
epifaunal predator/scavenger |
|
Eulimella sp. 1 |
facultative |
epifaunal parasite |
|
Eulimella sp. 2 |
facultative |
epifaunal parasite |
|
Odostomia sp. |
facultative |
epifaunal parasite |
|
Cylichnidae? gen. et sp. indet. |
facultative |
infaunal predator/scavenger |
|
Acharax sp. |
facultative |
infaunal chemosymbiotic |
|
Ledella favus Hoffman & Little sp. nov. |
facultative |
infaunal deposit feeder |
|
Yoldiella umbostriatus Hoffman & Little sp. nov. |
facultative |
infaunal deposit feeder |
|
Gigantidas cf. platifrons |
obligate |
semi-infaunal chemosymbiotic |
|
Parvamussium? aff. scitulum |
facultative |
epifaunal microcarnivore |
|
Propeamussiidae? gen. et sp. indet. |
facultative |
epifaunal microcarnivore |
|
Conchocele sp. |
obligate |
infaunal chemosymbiotic detritivore |
|
Vesicomya prashadi Hoffman & Little sp. nov. |
facultative |
infaunal detritivore |
|
Pliocardia cf. solidissima |
obligate |
infaunal chemosymbiotic |
|
Callogonia cf. leeana |
obligate |
infaunal chemosymbiotic |
|
Archivesica cf. kawamurai |
obligate |
semi-infaunal chemosymbiotic |
|
Cuspidariidae? gen. et sp. indet. |
facultative |
epifaunal microcarnivore |
|
Dentaliidae? gen. et sp. indet. |
facultative |
semi-infaunal microcarnivore |
|
Cadulus chuni |
facultative |
semi-infaunal microcarnivore |
Conclusions
The here described mollusc fauna of a bathyal Upper Pleistocene fossil methane seep deposit from between 15.3 mbsf and 17 mbsf in cores NGHP-01-12A and NGHP-01-10D in the Krishna-Godavari Basin comprises 29 taxa: 15 gastropods, 12 bivalves, and two scaphopods. Of these nine are new species: six gastropods and three bivalves. Six are likely to have had chemosymbionts, representing 21% of the diversity. The majority of these taxa were likely to have been obligate to seeps, as was probably the case for two of the new gastropod species. The rest of the mollusc taxa have living relatives common in bathyal habitats and were thus most likely facultative or “background” fauna in the fossil seep assemblage. The presence of a bathymodiolin mussel shell fragment from 42 mbsf level in Core NGHP-01-12A indicates that methane seep communities were also present sometime earlier in the Krishna-Godavari Basin. The molluscan assemblage from the Upper Pleistocene Seep deposit shares some taxa with recent seep communities from the east coast of India and elsewhere in the Indian Ocean, as well as with fossils from other sediment cores from the Krishna-Godavari Basin. The extent of taxonomic similarities between these Indian Ocean seep faunas awaits further systematic work.
Acknowledgements
We thank the following people for help and advice with identifications of various taxa in the fossil material: Elena Krylova (Shirshov Institute of Oceanology, Moscow, Russia), Chong Chen (JAMSTEC, Yokosuka, Japan), Julia Sigwart (Senckenberg, Frankfurt, Germany), Elizabeth Harper (University of Cambridge, Cambridge, UK), Leonardo Santos de Souza (Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil), Helmut Zibrowius (Marseille, France), Giuliana Panieri (Arctic University of Norway in Tromsø, Tromsø, Norway), Deborah Wall-Palmer and Harry ten Hove (both Naturalis Biodiversity Center, Leiden, Netherlands), Olev Vinn (University of Tartu, Tartu, Estonia), and Max Wisshak (Senckenberg am Meer, Wilhelmshaven, Germany). The authors wish to thank those that contributed to the success of the National Gas Hydrate Program Expedition 01 (NGHP-01). NGHP-01 was planned and managed through collaboration between the Directorate General of Hydrocarbons (DGH) under the Ministry of Petroleum and Natural Gas (India), the U.S. Geological Survey (USGS), and the Consortium for Scientific Methane Hydrate Investigations (CSMHI) led by Overseas Drilling Limited (ODL) and FUGRO McClelland Marine Geosciences (FUGRO). The platform for the drilling operation was the research drill ship JOIDES Resolution, operated by ODL. Much of the drilling/coring equipment used was provided by the Integrated Ocean Drilling Program (IODP) through a loan agreement with the US National Science Foundation. Wireline pressure coring systems and supporting laboratories were provided by IODP/Texas A&M University (TAMU), FUGRO, USGS, U.S. Department of Energy (USDOE) and HYACINTH/GeoTek. Downhole logging operational and technical support was provided by Lamont-Doherty Earth Observatory (LDEO) of Columbia University. The financial support for the NGHP-01, from the Oil Industry Development Board, Oil and Natural Gas Corporation Ltd., GAIL (India) Ltd. and Oil India Ltd. is gratefully acknowledged. We also acknowledge the support extended by all the participating organizations of the NGHP: MoP&NG, DGH, ONGC, GAIL, OIL, NIO, NIOT, and RIL. We greatly appreciate the efforts of the NGHP-01 shipboard scientific party for the collection and initial interpretation of these cores. We thank Graham Oliver (National Museum of Wales, Cardiff, UK) and an anonymous reviewer for constructive comments on an earlier version of the paper.
Editor: Krzysztof Hryniewicz
References
Abbott, R.T. 1954. American Seashells. xiv + 541 pp. Van Nostrand, New York. Crossref
Adams, A. 1864. On the species of Nearea found in the seas of Japan. Annals and Magazine of Natural History Ser. 3, 13 (75): 206–209. Crossref
Adams, Н. and Adams, A. 1854–1858. The Genera of Recent Mollusca Arranged According to Their Organisation, Volume 2. 1–92 (1854), 93–284 (1855), 285–412 (1856), 413–540 (1857), 541–661 (1858). Van Voorst, London.
Allen, J.A. 1993. A new deep-water hydrothermal species of Nuculana (Bivalvia: Protobranchia) from the Guaymas Basin. Malacologia 35 (1): 141–151.
Amano K. and Kiel S. 2010. Taxonomy and distribution of fossil Archivesica (Bivalvia: Vesicomyidae) in Japan. The Nautilus 124 (4): 155–165.
Angas, G.F. 1877. A further list of additional species of marine mollusca to be included in the fauna of Port Jackson and the adjacent coasts of New South Wales. Proceedings of the Zoological Society of London 1877 (2): 178–194.
Baba, K. and Williams, A.B. 1998. New Galatheoidea (Crustacea, Decapoda, Anomura) from hydrothermal systems in the West Pacific Ocean Bismarck Archipelago and Okinawa Trough. Zoosystema 20: 143–156. Crossref
Badesab, F., Dewangan, P., and Gaikwad, V. 2020. Magnetic mineral diagenesis in a newly discovered active cold seep site in the Bay of Bengal. Frontiers in Earth Science 8: 592557. Crossref
Beck, L.A. 1996. Morphology and anatomy of new species of neolepetopsid, acmaeid, fissurellid and pyropeltid limpets from Edison Seamount off Lihir Islands (West Pacific). (Gastropoda: Prosobranchia: Neolepetopsidae, Acmaeidae, Fissurellidae, Pyropeltidae). Archiv für Molluskenkunde 125 (1–2): 87–103. Crossref
Beets, C. 1942. Beiträge zur Kenntniss der Angeblich Oberoligocänen Mollusken-Fauna der Insel Buton, Niederländisch-Ostindlen. Leidsche Geologische Mededeelingen 13 (255): 256–328.
Bellardi, L. 1875. Novae pleurotomidarum Pedimonti et Liguriae fossilium: dispositionis prodromus. Bollettino della Societa Malacologica Italiana 1: 16–24.
Betters, M.J. and Cordes, E.E. 2023. New records of Provanna (Gastropoda, Provannidae) from the Costa Rica Margin and an identification key for the genus. ZooKeys 1189: 1–32. Crossref
Bohrmann, G. 2008. Report and preliminary results of R/V Meteor cruise M74/3, Fujairah-Male, 30 October – 28 November 2007. Cold seeps of the Makran Subduction Zone (Continental margin of Pakistan), Berichte Fachbereich Geowissenschaft Universität Bremen 266: 1–161.
Bouchet P. and Warén A. 1993. Revision of the Northeast Atlantic bathyal and abyssal Mesogastropoda. Bollettino Malacologico Supplemento 3: 579–840. Crossref
Breton, G., Wisshak, M., Néraudeau, D., and Morel, N. 2017. Parasitic gastropod bioerosion trace fossil on Cenomanian oysters from Le Mans, France and its ichnologic and taphonomic context. Acta Palaeontologica Polonica 62: 45–57. Crossref
Bromley, R.G. 1981. Concepts in ichnotaxonomy illustrated by small round holes in shells. Acta geológica hispánica 16: 55–64.
Bronn, H.G. 1831. Italiens Tertiär-Gebilde und deren organische Einschlüsse. xii + 176 pp. Karl Groos, Heidelberg. Crossref
Bronn, H.G. 1862. Die Klassen und Ordnungen der Weichthiere (Malacozoa), wissenschaftlich dargestellt in Wort und Bild. Dritten Bandes erste Abtheilung. Kopflose Weichthiere (Malacozoa Acephala) Weichthiere. 518 pp. Wintersche Verlagshandlung, Leipzig. Crossref
Beurlen, K. 1944. Beiträge zur Stammesgeschichte der Muscheln. Mathematisch-Naturwissenschaftlichen Abteilung der Bayerischen Akademie der Wissenschaften zu München, Sitzungsberichte. 1944(1–2):133–145.
Burmeister, H. 1837. Handbuch der Naturgeschichte. Zum Gebrauch bei Vorlesungen. Zweite Abteilung: Zoologie. xii + 858 pp. T.C.F. Enslin, Berlin. Crossref
Carter, J.G., Campbell, D.C., and Campbell, M.R. 2000. Cladistic perspectives on early bivalve evolution. In: E. M. Harper, J.D. Taylor, and J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia. The Geological Society of London, Special Publication 177: 47–79. Crossref
Chen, C., Okutani, T., Liang, Q., and Qiu, J.-W. 2018. A noteworthy new species of the family Vesicomyidae from the South China Sea (Bivalvia: Glossoidea). Venus 76 (1–4): 29–37.
Chen, C, Zhong, Z, Qiu, J.-W., and Sun, J. 2023. A new Paralepetopsis limpet from a South China Sea seep hints at a paraphyletic Neolepetopsidae. Zoological Studies 62: e26.
Children, J.G. 1834. Mollusca. In: Synopsis of the Contents of the British Museum, 28th Edition, 88–118. British Museum, London.
Clark, W. 1851. On the classification of the British marine testaceous mollusca. The Annals and Magazine of Natural History 7 (2): 469–481.
Chistikov S.D. 1979. Phylogenetic relations of the Scaphopoda [in Russian]. In: I.M. Likharev (ed.), Molliuski, osnovnye rezul’taty ih izučeniâ, 20–22. Nauka, Leningrad.
Collett, T.S., Boswell, R., Cochran, J.R., Kumar, P., Lall, M., Mazumdar, A., Venkata Ramana, M., Ramprasad, T, Riedel, M., Sain, K., Vasant Sathe, A., and Vishwanath, K. 2014. NGHP Expedition 01 Scientific Party. Geologic implications of gas hydrates in the offshore of India: Results of the National Gas Hydrate Program Expedition 01. Marine and Petroleum Geology 58: 3e28. Crossref
Collett, T., Riedel, M., Cochran, J., Boswell, R., Presley, J., Kumar, P., Sathe, A., Sethi, A., Lall, M., and NGHP Expedition Scientists. 2015. Indian National Gas Hydrate Program Expedition 01 Report. Scientific Investigations Report 2012–5054. U.S. Geological Survey, Reston. Crossref
Conrad, T.A. 1849. Appendix I. Descriptions of fossils. In: J.D. Dana (ed.), Geology. With a Folio Atlas of twenty-one Plates. United States Exploring Expedition during the Years 1838, 1839, 1840, 1841, (1842) Under the Command of Charles Wilkes, U.S.N., 723–729. Putnam, New York.
Cordes, E.E., Cunha, M.R., Galeron, J., Mora, C., Olu‐Le Roy, K., Sibuet, M., Van Gaever, S., Vanreusel, A., and Levin, L.A. 2010. The influence of geological, geochemical, and biogenic habitat heterogeneity on seep biodiversity. Marine Ecology 31 (1): 51–65. Crossref
Cosel, R. von and Marshall, B.A. 2003. Two new species of large mussels (Bivalvia: Mytilidae) from active submarine volcanoes and a cold seep off the eastern North Island of New Zealand, with description of a new genus. The Nautilus 117 (2): 31–46.
Cosel, R. von and Olu, K. 2009. Large Vesicomyidae (Mollusca: Bivalvia) from cold seeps in the Gulf of Guinea off the coasts of Gabon, Congo and northern Angola. Deep Sea Research Part II: Topical Studies in Oceanography 56: 2350–2379. Crossref
Cosel, R. von and Salas, C. 2001. Vesicomyidae (Mollusca: Bivalvia) of the genera Vesicomya, Waisiuconcha, Isorropodon and Callogonia in the Eastern Atlantic and the Mediterranean. Sarsia 86: 333–366. Crossref
Cox, L.R. 1960. Thoughts on the classification of the Gastropoda. Proceedings of the Malacological Society of London 33 (6): 239–261.
Cuvier, G.L.C.F.D. 1795. Second Mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des Mollusques et de leur division en ordre, lu à la société d’Histoire Naturelle de Paris, le 11 prairial an troisième. Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts 2: 433–449.
, W.H. 1886. Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Gulf of Mexico (1877–78) and in the Carribean Sea (1879–80), by the U.S. Coast Survey steamer “Blake”, Lieut.-Commander C.D. Sigsbee, U.S.N. and Commander J.R. Bartlett, U.S.N. commanding. XXIX. Report on the Mollusca. Part 1, Brachiopoda and Pelecypoda. Bulletin of the Museum of Comparative Zoölogy at Harvard College 12 (6): 171–318.
Dall, W.H. 1889a. Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Gulf of Mexico (1877–78) and in the Caribbean Sea (1879–80), by the U.S. Coast Survey Steamer “Blake”, Lieut.-Commander C.D. Sigsbee, U.S.N., and Commander J.R. Bartlett, U.S.N., commanding. XXIX. Report on the Mollusca. Part 2, Gastropoda and Scaphopoda. Bulletin of the Museum of Comparative Zoology at Harvard College 18: 1–492.
Dall, W.H. 1889b. On the hinge of pelecypods and its development, with an attempt toward a better subdivision of the group. American Journal of Science (Series 3) 38: 445–462. Crossref
Dall, W.H. 1891. On some new or interesting west American shells obtained from the dredgings of the U.S. Fish Commission steamer Albatross in 1888, and from other sources. Proceedings of the United States National Museum 14: 173–191. Crossref
Dall, W.H. 1896. Diagnoses of new species of mollusks from the west coast of America. Proceedings of the United States National Museum. 18 (1034): 7–20. Crossref
Dall, W.H. 1900. Contributions to the Tertiary fauna of Florida, with especial reference to the Silex beds of Tampa and the Pliocene beds of the Caloosahatchie River, including in many cases a complete revision of the generic groups treated of and their American Tertiary species. Part V. Teleodesmacea: Solen to Diplodonta. Transactions of the Wagner Free Institute of Science of Philadelphia 3 (5): 949–1218, pls. 36–47.
Dall, W.H. 1903. Contributions to the Tertiary of Florida with especial reference to the Silex beds of Tampa and the Pliocene beds of the Caloosahatchie River, including in many cases a complete revision of the generic groups treated of and their American Tertiary species. Part VI. Concluding the work. Transactions of the Wagner Free Institute of Science of Philadelphia 3 (6): 1219–1654. Crossref
Dall, W.H. 1908a. A revision of the Solenomyacidae. The Nautilus. 22 (1): 1–2.
Dall, W.H. 1908b. Reports on the dredging operations off the west coast of Central America to the Galapagos, to the west coast of Mexico, and in the Gulf of California, in charge of Alexander Agassiz, carried on by the U.S. Fish Commission steamer “Albatross,” during 1891, Lieut.-Commander Z.L. Tanner, U.S.N., commanding. XXXVII. Reports on the scientific results of the expedition to the eastern tropical Pacific, in charge of Alexander Agassiz, by the U.S. Fish Commission steamer “Albatross”, from October, 1904 to March, 1905, Lieut.-Commander L.M. Garrett, U.S.N., commanding. XIV. The Mollusca and Brachiopoda. Bulletin of the Museum of Comparative Zoology 43 (6): 205–487.
Dall, W.H. and Simpson, C.T. 1901. The Mollusca of Porto Rico. Bulletin of the United States Fish Commission 20: 351–524.
Dautzenberg, P. and Fischer, H. 1896. Dragages effectués par l’Hirondelle et par la Princesse Alice 1888–1895. Mollusques Gastéropodes. Mémoires de la Société Zoologique de France 9: 395–498.
De Gibert, J.M., Domènech, R., and Martinell, J. 2007. Bioerosion in shell beds from the Pliocene Roussillon Basin, France: Implications for the (macro) bioerosion ichnofacies model. Acta Palaeontologica Polonica 52: 783–798.
Dewangan, P., Sriram, G., Kumar, A., Mazumdar, A., Peketi, A., Mahale, V., Reddy, S.S.C., and Babu, A. 2021. Widespread occurrence of methane seeps in deep-water regions of Krishna-Godavari basin, Bay of Bengal. Marine and Petroleum Geology 124: 104783. Crossref
Dijkstra, H.H. and Janssen, R. 2013. Bathyal and abyssal Pectinoidea from the Red Sea and Gulf of Aden (Bivalvia: Propeamussiidae, Entoliidae, Pectinidae). Archiv für Molluskenkunde 142 (2): 181–214. Crossref
Dijkstra, H.H. and Maestrati, P. 2015. Pectinoidea (Bivalvia: Propeamussiidae and Cyclochlamydidae) from the southwestern Indian Ocean. African Invertebrates 56 (3): 585–628. Crossref
Dubilier, N., Bergin, C., and Lott, C. 2008. Symbiotic diversity in marine animals: the art of harnessing chemosynthesis. Nature Reviews Microbiology 6 (10): 725–740. Crossref
Férussac, A.E.J.P.F. d’Audebard de 1822. Tableaux systématiques des animaux mollusques classés en familles naturelles, dans lesquels on a établi la concordance de tous les systèmes; suivis d’un prodrome général pour tous les mollusques terrestres ou fluviatiles, vivants ou fossiles. 111 pp. J.B. Baillière, Paris. Crossref
Fischer, P. (1880–1887). Manuel de conchyliologie et de paléontologie conchyliologique, ou histoire naturelle des mollusques vivants et fossiles suivi d’un Appendice sur les Brachiopodes par D.P. Oehlert. Avec 23 planches contenant 600 figures dessinées par S.P. Woodward. Published in 11 parts (fascicules) [1: 1–112, 21 September 1880; 2: 113–192, 16 March 1881; 3: 193–304, 28 July 1881; 4: 305–416, 5 May 1882; 5: 417–512, 21 February 1883; 6: 513–608, 20 December 1883; 7: 609–688, 30 June 1884; 8: 689–784, 29 January 1885; 9: 785–896, 31 August 1885; 10: 897–1008, 30 April 1886; 11: 1009–1369, 15 June 1887]. F. Savy, Paris. Crossref
Fleming, J. 1813. Conchology. In: D. Brewster (ed.), The Edinburgh Encyclopaedia, Vol. 7, 55–107. William Blackwood, Edinburg.
Fleming, J. 1822. The Philosophy of Zoology: or a General View of the Structure, Functions and Classification of Animals, Vol. 2. 618 pp. Archibald Constable & Co, Edinburgh. Crossref
Fleming, J. 1825. On the British testaceous annelids. The Edinburgh Philosophical Journal 12 (24): 238–248.
Fleming, J. 1828. A History of British Animals, Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca, and Radiata of the United Kingdom; Including the Indigenous, Extirpated, and Extinct Kinds, Together with Periodical and Occasional Visitants. 565 pp. Bell & Bradfute, Edinburgh. Crossref
Flores, J.A., Johnson, J.E., Mejia-Molina, A.E., Álvarez, M.C., Sierro, F.J., Singh, S.D., Mahanti, S., and Giosan, L. 2014. Sedimentation rates from calcareous nannofossil and planktonic foraminifera biostratigraphy in the Andaman Sea, northern Bay of Bengal, and eastern Arabian Sea. Marine and Petroleum Geology 58: 425–437. Crossref
Forbes, E. and M’Andrew, [R.]. 1846. Notes on some new and rare animals observed during cruises in the British seas, since the last meeting. The Athenaeum: Journal of Literature, Science, the Fine Arts 3052: 1027.
Frauenfeld, G. von. 1867. Mollusken. In: C. Felder, R. Felder, and A.F. Rogenhofer (eds.),, Reise der österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859 unter den Befehlen des Commodore B. von Wüllerstorf-Urbair. Zoologischer Theil. Zweiter Band. Dritte Abtheilung, 1–16. Karl Gerold’s Sohn, Wien.
Fujikura, K., Kojima, S., Tamaki, K., Maki, Y., Hunt, J., and Okutani, T. 1999. The deepest chemosynthesis-based community yet discovered from the hadal zone, 7326 m deep, in the Japan Trench. Marine Ecology Progress Series 190: 17–26. Crossref
Gabb, W.M. 1866. Cretaceous and Tertiary fossils. Geological Survey of California. Palaeontology 2: 1–38.
Gale A.S., Little C.T.S, Johnson J.E., and Giosan L. 2020. A new neolepadid cirripede from a Pleistocene cold seep, Krishna-Godavari Basin, offshore India. Acta Palaeontologica Polonica 65 (2): 351–362. Crossref
Geiger, D.L. 2012. Monograph of the little slit shells. Volume 2. Anatomidae, Larocheidae, Depressizonidae, Sutilizonidae, Temnocinclidae. Santa Barbara Museum of Natural History Monographs 7: 729–1291.
Georgieva, M.N., Little, C.T.S., Watson, J.S., Sephton, M.A., Ball, A.D., and Glover, A.G. 2019. Identification of fossil worm tubes from Phanerozoic hydrothermal vents and cold seeps. Journal of Systematic Palaeontology 17: 287–329. Crossref
Gofas, S. and Warén, A. 1982. Taxonomie de quelque espèces du genre Alvania (Mollusca, Gastropoda) des côtes Ibériques et Marocaines. Bollettino Malacologico 18 (1–4): 1–16.
Gonsalves, M.J., Tiwari, S., Padate, V.P., Trivedi, J.N., Samuel, V.D., and Viswanathan, C. 2022. First record of galatheid squat lobster, Shinkaia crosnieri Baba & Williams, 1998 (Decapoda: Galatheoidea) from cold-seep environment of the Indian Ocean. Thalassas: An International Journal of Marine Sciences 38: 1245–1252. Crossref
Gray, J.E. 1840. Shells of molluscous animals. Synopsis of the Contents of the British Museum 42: 105–152.
Gray, J.E. 1847. A list of the genera of Recent Mollusca, their synonyma and types. Proceedings of the Zoological Society of London 15 (178): 129–219.
Gray, J.E. 1854. A revision of the arrangement of the families of bivalve shells (Conchifera). Annals and Magazine of Natural History 13 (77): 408–418. Crossref
Haas, A., Little, C.T.S., Sahling, H., Bohrmann, G., Himmler, T., and Peckmann, P. 2009. Mineralization of vestimentiferan tubes at methane seeps on the Congo deep sea fan. Deep-Sea Research 56: 283–293. Crossref
Habe, T. 1951. Scissurellidae in Japan. In: T. Kuroda (ed.), Illustrated CatClogue of Japanese Shells 1 (11): 65–69.
Hansen, J., Hoff, U., Sztybor, K., and Rasmussen, T.I. 2017. Taxonomy and palaeoecology of two Late Pleistocene species of vesicomyid bivalves from cold methane seeps at Svalbard (79°N). Journal of Molluscan Studies 83: 270–279. Crossref
Hashimoto, J. and Okutani, T. 1994. Four new mytilid mussels associated with deep sea chemosynthetic communities around Japan. Venus 53 (2): 61–83.
He, X., Xu, T., Chen, C., Xu, L., Yi-Xuan, L., Zhaoyan, Z., Xinyu, G.,Yi-Tao, L., Yi, L., Guoyong, Y., Yanan, S., Jian-Wen, Q., Pei-Yuan, Q., and Jin, S. 2023. Same (sea) bed different dreams: Biological community structure of the Haima seep reveals distinct biogeographic affinities. The Innovation Geoscience 1 (2): 100019. Crossref
Herbert, D.G. 2024. The Vetigastropoda (Mollusca) of Walters Shoal, with descriptions of two new genera and thirty new species. European Journal of Taxonomy 923: 1–119. Crossref
Hickman, C.S. 1983. Radular patterns, systematics, diversity, and ecology of deep-sea limpets. The Veliger 26: 73–92.
Hinds, R.B. 1843. Descriptions of new species of Nucula, from the collections of Sir Edward Belcher, C.B., and Hugh Cuming, Esq. Proceedings of the Zoological Society of London 11: 97–101.
Hoffman, L., Gofas, S., and Freiwald, A. 2020. A large biodiversity of “skeneimorph” (Gastropoda: Vetigastropoda) species from the South Azorean Seamount Chain, with the description of seventeen new species. Iberus 38 (Supplement 9): 1–82.
Hoffman, L., Kniesz, K., Martínez Arbizu, P., and Kihara, T.C. 2022. Abyssal vent field habitats along plate margins in the Central Indian Ocean yield new species in the genus Anatoma (Vetigastropoda: Anatomidae). European Journal of Taxonomy 826: 135–162. Crossref
Høisaeter, T. 1968. Taxonomic notes on the North-European species of Cyclostrema sensu Jeffreys, 1863 (Prosobranchia: Diotocardia). Sarsia 33: 43–58. Crossref
Huber, M. 2010. Compendium of Bivalves. A Full-color Guide to 3,300 of the World’s Marine Bivalves. A Status on Bivalvia After 250 Years of Research. 901 pp. ConchBooks, Harxheim.
Imhoff, J.F., Sahling, H., Süling, J, and Kath, T. 2003. 16S rDNA-based phylogeny of sulphur-oxidising bacterial endosymbionts in marine bivalves from cold-seep habitats. Marine Ecology Progress Series 249: 39–51. Crossref
Jaeckel, S.H. 1932. Nachtrag zu den Scaphopoden der Valdivia-Expedition. In: C. Chun (ed.), Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition 1898–1899 21 (2): 301–315.
Jeffreys, J.G. 1847. Additional notices on British shells. Annals and Magazine of Natural History 20: 16–19. Crossref
Jeffreys, J.G. 1870. Norwegian Mollusca. Annals and Magazine of Natural History 5 (4): 438–448. Crossref
Johnson, J.E., Phillips, S.C., Clyde, W.C., Giosan, L., and Torres, M.E. 2021. Isolating detrital and diagenetic signals in magnetic susceptibility records from methane-bearing marine sediments. Geochemistry, Geophysics, Geosystems 22 (9): e2021GC009867. Crossref
Johnson, J.E., Phillips, S.C., Torres, M.E., Piñero, E., Rose, K.K., and Giosan, L. 2014. Influence of total organic carbon deposition on the inventory of gas hydrate in the Indian continental margins. Marine and Petroleum Geology 58: 406–424. Crossref
Joshi, R.K., Mazumdar, A., Peketi, A., Ramamurty, P.B., Naik, B.G., Kocherla, M., Carvalho, M.A., Mahalakshmi, P., Dewangan, P., and Ramana, M.V. 2014. Gas hydrate destabilization and methane release events in the Krishna-Godavari Basin, Bay of Bengal. Marine and Petroleum Geology 58: 476–489. Crossref
Joye, S.B. 2020. The geology and biogeochemistry of hydrocarbon seeps. Annual Review of Earth and Planetary Sciences 48: 205–231. Crossref
Kiel, S. 2006. New records and species of molluscs from Tertiary cold-seep carbonates in Washington State, USA. Journal of Paleontology 80: 121–137. Crossref
Kiel, S., Aguilar, Y.M., and Kase, T. 2020. Mollusks from Pliocene and Pleistocene seep deposits in Leyte, Philippines. Acta Palaeontologica Polonica 65: 589–627. Crossref
Kiel, S., Birgel, D., Peckmann, J., and Wang, S.-W. 2024. An overview of Miocene to Pleistocene methane-seep faunas from Taiwan: Paleoecology and paleobiogeographic implications. Journal of Asian Earth Sciences 266: 106119. Crossref
Knudsen, J. 1970. The systematics and biology of abyssal and hadal Bivalvia. Galathea Reports 11: 1–241.
Kocherla, M., Teichert, B.M.A., Pillai, S., Satyanarayanan, M., Ramamurty, P.B., Patil, D.J., and Rao, A.N. 2015. Formation of methane-related authigenic carbonates in a highly dynamic biogeochemical system in the Krishna-Godavari Basin, Bay of Bengal. Marine and Petroleum Geology 64: 324–333. Crossref
Kojima, S., Fujikura, K., and Okutani, T. 2004. Multiple trans-Pacific migrations of deep-sea vent/seep-endemic bivalves in the family Vesicomyidae. Molecular Phylogenetics and Evolution 32: 396–406. Crossref
Krylova, E.M. and Sahling, H. 2006. Recent bivalve molluscs of the genus Calyptogena (Vesicomyidae). Journal of Molluscan Studies 72: 359–395. Crossref
Krylova, E.M. and Sahling, H. 2010. Vesicomyidae (Bivalvia): current taxonomy and distribution. PloSONE 5 (4): e9957. Crossref
Krylova, E.M., Sahling, H., and Borowski, C. 2018. Resolving the status of the families Vesicomyidae and Kelliellidae (Bivalvia: Venerida), with notes on their ecology. Journal of Molluscan Studies 84: 69–91. Crossref
Kuroda, T. 1943. Akebiconcha, a new pelecypod genus. Venus 13: 14–18.
Leifer, I. 2019. A synthesis review of emissions and fates for the Coal Oil Point marine hydrocarbon seep field and California marine seepage. Geofluids 2019: 1–48. Crossref
Levin, LA. 2005. Ecology of cold seep sediments: interactions of fauna with flow, chemistry and microbes. Oceanography and Marine Biology 43: 1–46. Crossref
Lin, Y.T., Kiel, S., Xu, T., and Qiu, J.W. 2022. Phylogenetic placement, morphology and gill-associated bacteria of a new genus and species of deep-sea mussel (Mytilidae: Bathymodiolinae) from the South China Sea. Deep Sea Research Part I: Oceanographic Research Papers 190: 103894. Crossref
Lindberg, D.R. 1986. Radular evolution in the Patellogastropoda. American Malacological Bulletin 4: 115.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Lorenson, T.D. and Collett, T.S. 2018. National Gas Hydrate Program Expedition 01 offshore India; gas hydrate systems as revealed by hydrocarbon gas geochemistry. Marine and Petroleum Geology 92: 477–492. Crossref
Lorion, J., Kiel, S., Faure, B., Kawato, M., Ho, S.Y., Marshall, B., Tsuchida, S., Miyazaki, J.I., and Fujiwara, Y. 2013. Adaptive radiation of chemosymbiotic deep-sea mussels. Proceedings of the Royal Society B: Biological Sciences 280: 20131243. Crossref
Lovén, S.L. 1846. Index Molluscorum litora Scandinaviae occidentalia habitantium. Öfversigt af Kongliga Vetenskaps Akademiens Förhandlingar 1846: 134–160, 182–204. Crossref
Lozouet, P. 1990. Benthonellania, nouveau genre de Rissoidae (Gastropoda, Prosobranchia) du bathyal atlantique. Bulletin du Muséum National d’Histoire Naturelle (section A Zoologie) 12 (4): 313–328. Crossref
Marshall, B.A. 1986. Recent and Tertiary Cocculinidae and Pseudococculinidae (Mollusca: Gastropoda) from New Zealand and New South Wales, New Zealand. Journal of Zoology 12 (4): 505–546. Crossref
Mazumdar, A., Dewangan, P., Joäo, H.M., Peketi, A., Khosla, V.R., Kocherla, M., Badesab, F.K., Joshi, R.K., Roxanne, P., Ramamurty, P.B., and Karisiddaiah, S.M. 2009. Evidence of paleo-cold seep activity from the Bay of Bengal, offshore India. Geochemistry, Geophysics, Geosystems 10 (6): 1–15. Crossref
Mazumdar, A., Dewangan, P., Peketi, A., Badesaab, F., Sadique, M., Sivan, K., Mathai, J., Ghosh, A., Zatale, A., Pillutla, S.P.K., and Uma, C. 2021. The first record of the genus Lamellibrachia (Siboglinidae) tubeworm along with associated organisms in a chemosynthetic ecosystem from the Indian Ocean: A report from the Cauvery-Mannar Basin. Journal of Earth System Science 130 (2): 1–10. Crossref
Mazumdar, A., Dewangan, P., Peketi, A., Gullapalli, S., Kalpana, M.S., Naik, G.P., Shetty, D., Pujari, S., Pillutla, S.P.K., Gaikwad, V.V., and Nazareth, D. 2019. The first record of active methane (cold) seep ecosystem associated with shallow methane hydrate from the Indian EEZ. Journal of Earth System Science 128: 1–8. Crossref
Melvill, J.C. 1893. Descriptions of twenty-five new species of marine shells from Bombay collected by Alexander Abercrombie, Esq. Memoirs and Proceedings of the Manchester Literary and Philosophical Society 7: 52–67.
McLean, J.H. 1989. New slit-limpets (Scissurellacea and Fissurellacea) from hydrothermal vents. Part 1: Systematic descriptions and comparisons based on shell and radular characters. Contributions in Science, Natural History Museum of Los Angeles County 407: 1–29. Crossref
McLean, J.H. 1990. Neolepetopsidae, a new docoglossate limpet family from hydrothermal vents and its relevance to patellogastropod evolution. Journal of Zoology 222 (3): 485–528.
McLean, J.H. 1991. Four new pseudococculinid limpets collected by the deep-submersible Alvin in the eastern Pacific. The Veliger 34 (1): 38–47.
McLean, J.H. 2008. Three New Species of the Family Neolepetopsidae (Patellogastropoda) from Hydrothermal Vents and Whale Falls in the Northeastern Pacific. Journal of Shellfish Research 27 (1): 15–20. Crossref
MolluscaBase eds. 2024. MolluscaBase. Gigantidas Cosel & B.A. Marshall, 2003 and other genera. Accessed through: World Register of Marine Species at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=391122 on 2024-02-01
Montagu, G. 1803. Testacea Britannica or Natural History of British Shells, Marine, Land, and Fresh-water, Including the Most Minute: Systematically Arranged and Embellished with Figures. Vol. 1, i–xxxvii + 1–291; Vol. 2, 293–606. J. White, London. Crossref
Monterosato, T.A. di 1872. Notizie intorno alle conchiglie fossili di Monte Pellegrino e Ficarazzi. Ufficio Tipografico Michele Amenta. 44 pp. Printed for the Author, Palermo. Crossref
Monterosato, T.A. di. 1874. Recherches conchyliologiques, effectuées au Cap Santo Vito, en Sicile. Journal de Conchyliologie 22 (3): 243–282.
Neulinger, S.C., Sahling, H., Süling, J., and Imhoff, J.F. 2006. Presence of two phylogenetically distinct groups in the deep-sea mussel Acharax (Mollusca: Bivalvia: Solemyidae). Marine Ecology Progress Series 312: 161–168. Crossref
Okutani, T. 1964. Report on the archibenthal and abyssal gastropod Mollusca mainly collected from Sagami Bay and adjacent waters by the R. V. Soyo-Maru during the years 1955–1963. Journal of the Faculty of Science, University of Tokyo, Section II 15: 371–447.
Okutani, T. and Soh, W. 2005. Calyptogena (Archivesica) garuda, a magnificent new species of vesicomyid bivalve from the Java Trench, Indonesia. Venus 64 (1–2): 23–29.
Okutani T., Fujikura K., and Kojima S. 2000. New taxa and review of vesicomyid bivalves collected from the northwest Pacific by deep-sea research systems of Japan Marine Science and Technology Center. Venus 59 (2): 83–101.
Oliver P.G. and Frey M.A. 2014. Ascetoaxinus quatsinoensis sp. et gen. nov. (Bivalvia: Thyasiroidea) from Vancouver Island, with notes on Conchocele Gabb, 1866, and Channelaxinus Valentich-Scott & Coan, 2012. Zootaxa 3869 (4): 452–468. Crossref
Oliver, P.G., Gonsalves, M.-J., Deepak, S., and Garzia, M. 2024. Gigantidas niobengalensis sp. nov. (Bivalvia: Mytiloidea) from a methane seep in the Krishna-Godavari Basin, Bay of Bengal. Journal of Conchology 45: 221–233. Crossref
Peketi, A., Mazumdar, A., Sawant, B., Manaskanya, A., and Zatale, A. 2022. Biogeochemistry and trophic structure of a cold seep ecosystem, offshore Krishna-Godavari basin (east coast of India). Marine and Petroleum Geology 138: 105542. Crossref
Pelseneer, P. 1889. Sur la classification phylogenetique de pelecypods. Bulletin Scientifique de la France et de la Belgique 20: 27–52.
Pereira, O.S., Gonzalez, J., Mendoza, G.F., Le, J., Coscino, C.L., Lee, R.W., Cortés, J., Cordes, E.E., and Levin, L.A. 2021. The dynamic influence of methane seepage on macrofauna inhabiting authigenic carbonates. Ecosphere 12 (10): e03744. Crossref
Philippi, R.A. 1844. Enumeratio molluscorum Siciliae cum viventium tum in tellure tertiaria fossilium, quae in itinere suo observavit. Vol. 2. 303 pp. Eduard Anton, Halle.
Phillips, S.C., Johnson, J.E., Underwood, M.B., Guo, J., Giosan, L., and Rose, K. 2014. Long-timescale variation in bulk and clay mineral composition of Indian continental margin sediments in the Bay of Bengal, Arabian Sea, and Andaman Sea. Marine and Petroleum Geology 58: 117–138. Crossref
Prashad, B. 1932. The Lamellibranchia of the Siboga expedition. Systematic part II (exclusive of the Pectinidae). Siboga Expeditie 53c: 1–351.
Rafinesque, C.S. 1815. Analyse de la nature ou Tableau de l’univers et des corps organisés. 224 pp. Self-published, Palermo. Crossref
Rajendra, S., Mondal, T., and Raghunathan, C. 2021. Chapter 33 Mollusca. In: K. Chandra, C. Raghunathan, H.U.K. Pillai, P. Jasmine, and T. Mondal (eds.), Deep Sea Faunal Diversity in India, 471–482. Zoological Survey of India, Kolkata.
Ravinesh, R., Oliver, P.G., and Saravanane, N. 2024. Ascetoaxinus ravichandrani sp. nov., a new species of deep-water Thyasiridae (Mollusca: Bivalvia) from the northern Indian Ocean. Journal of Conchology 45: 125–134.
Risso, A. 1826. Histoire naturelle des principales productions de l’Europe méridionale et particulièrement de celles des environs de Nice et des Alpes maritimes: tome cinquième. F.-G. 400 pp. Levrault, Paris. Crossref
Römer, M., Sahling, H., Pape, T., Bohrmann, G., and Spieß, V. 2012. Quantification of gas bubble emissions from submarine hydrocarbon seeps at the Makran continental margin (offshore Pakistan). Journal of Geophysical Research: Oceans 117: C10015. Crossref
Sacco, F. 1897. I Molluschi dei terreni terziarii del Piemonte e della Liguria, Parte 24. Bollettino dei Musei di Zoologia ed Anatomia Comparata della R. Università di Torino 12 (298): 101–102.
Sahling, H., Galkin, S.V., Salyuk, A., Greinert, J., Foerstel, H., Piepenburg, D., and Suess, E. 2003. Depth-related structure and ecological significance of cold-seep communities—a case study from the Sea of Okhotsk. Deep Sea Research Part I: Oceanographic Research Papers 50 (12): 1391–1409. Crossref
Salvini-Plaven, L. 1980. A reconsideration of systematic in the Mollusca (Phylogeny and higher classification). Malacologia 19 (2): 249–278.
Sangodkar, N., Gonsalves, M.J., and Nazareth, D.R. 2022. Macrofaunal distribution, diversity, and its ecological interaction at the cold seep site of Krishna-Godavari Basin, East Coast of India. Microbial Ecology 85 (1): 61–75. Crossref
Sasaki, T., Okutani, T., and Fujikura, K. 2005. Molluscs from hydrothermal vents and cold seeps in Japan: a review of taxa recorded in twenty recent years (1984–2004). Venus 64 (3–4): 87–133.
Sasaki, T., Warén, A., Kano, Y., Okutani, T., and Fujikura, K. 2010. Gastropods from recent hot vents and cold seeps: systematics, diversity and life strategies. In: S. Kiel (ed.), The Vent and Seep Biota: Aspects from Microbes to Ecosystems, 169–254. Springer, Dordrecht. Crossref
Scacchi, A. 1835. Notizie intorno alle conchiglie ed a’ zoofiti fossili che si trovano nelle vicinanze di Gravina in Puglia. Annali Civili del Regno delle Due Sicilie 6: 75–84; 7: 5–18.
Scarabino, V. 1995. Scaphopoda of the tropical Pacific and Indian Oceans, with description of 3 new genera and 42 new species. Mémoires du Muséum national d’Histoire naturelle. Série A, Zoologie 167: 189–379.
Sibuet, M. and Olu, K. 1998. Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins. Deep Sea Research Part II: Topical Studies in Oceanography 45 (1–3): 517–567. Crossref
Siegert, M., Krager, M., and Teichert, B. 2011. Anaerobic oxidation of methane at a marine methane seep in a forearc sediment basin off Sumatra, Indian Ocean. Frontiers in Microbiology 2: 1–16. Crossref
Smith, E.A. 1885. Report on the Lamellibranchiata collected by HMS Challenger during the years 1873–76. Report on the scientific results of the voyage of H.M.S. Challenger during the years 1873–76. Zoology 13: 1–341.
Smith, E.A. 1904. Natural history notes from H.M. Indian Marine Survey Steamer “Investigator”, Commander T.H. Heming, R.N., ser. 3 (1). On Mollusca from the Bay of Bengal and the Arabian Sea. Annals and Magazine of Natural History, Series 7 14: 1–14. Crossref
Southward, E.C., Schulze, A., and Tunnicliffe, V. 2002. Vestimentiferans (Pogonophora) in the Pacific and Indian Oceans: a new genus from Lihir Island (Papua New Guinea) and the Java Trench, with the first report of Arcovestia ivanovi from the North Fiji Basin. Journal of Natural History 36 (10): 1179–1197. Crossref
Starobogatov Y.I. 1974. Xenoconchias and their bearing on the phylogeny and systematics of some molluscan classes [in Russian]. Paleontologičeskij žurnal 1974 (1): 3–18 [Translated in Paleontological Journal of the American Geological Institute 8 (1): 1–13].
Stoliczka, F. 1868. Cretaceous Fauna of Southern India. II: Gastropoda (Prosocephala). Memoirs of the Geological Survey of India, Calcutta 5 (2), 1–497.
Sturany, R. 1896. Zoologische Ergebnisse VII. Mollusken I (Prosobranchier und Opisthobranchier; Scaphopoden; Lamellibranchier) gesammelt von S.M. Schiff “Pola” 1890–1894. Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematische-Naturwissenschaftlischen Classe 63: 1–36.
Suárez-Mozo, N.Y., Valentich-Scott, P., and Hendrickx, M.E. 2019. Deep-water bivalves from the oxygen minimum zone area off the western Peninsula of Baja California, Mexico. Molluscan Research 39: 99–109. Crossref
Swainson, W. 1840. A Treatise on Malacology; or the Natural Classification of Shells and Shell-fish. 419 pp. Longman, London. Crossref
Tan, K.S. 2021. Deep-sea Protobranchia (Bivalvia) from the SJADES biodiversity cruise, with a description of a new species of Propeleda (Nuculanidae). Raffles Bulletin of Zoology (Supplement) 36: 346–385.
Teichert, B.M.A., Johnson, J.E., Solomon, E.A., Giosan, L., Rose, K., Kocherla, M., Connolly, E.C., and Torres, M.E. 2014. Composition and origin of authigenic carbonates in the Krishna-Godavari and Mahanadi Basins, eastern continental margin of India. Marine and Petroleum Geology 58: 438–460. Crossref
Tenison Woods, J.E. 1877. On some new Tasmanian marine shells (Second series). Papers and Proceedings and Report of the Royal Society of Tasmania 1876: 131–159. Crossref
Termier, G. and Termier, H. 1950. Paléontologie marocaine. 2, Invertébrés de l’ère primaire. Fascicule 3, Mollusques. 246 pp. Hermann, Paris.
Verrill, A.E. 1884. Notice of the remarkable marine fauna occupying the outer banks off the southern coast of New England, No. 9. The American Journal of Science, Series 3 28 (165): 213–220. Crossref
Verrill, A.E. and Bush, K.J. 1897. Revision of the genera of Ledidae and Nuculidae of the Atlantic coast of the United States. The American Journal of Science, Series 4 3: 51–63. Crossref
Voigt, E. 1977. On grazing traces produced by the radula of fossil and Recent gastropods and chitons. In: T.P. Crimes and J.C. Harper (eds.), Trace Fossils 2, 335–346. Seel House Press, Liverpool.
Warén, A. 1978. The taxonomy of some North Atlantic species referred to Ledella and Yoldiella (Bivalvia). Sarsia 63 (4): 213–219. Crossref
Warén, A. 1992. New and little known “Skeneimorph” gastropods from the Mediterranean Sea and the adjacent Atlantic Ocean. Bollettino Malacologico 27 (10–12): 149–248.
Warén, A. and Bouchet, P. 2001. Gastropoda and Monoplacophora from hydrothermal vents and seeps; new taxa and records. The Veliger 44: 116–231.
Warén, A. and Bouchet, P. 2009. New gastropods from deep-sea hydrocarbon seeps off West Africa. Zoologica Scripta 22: 1–90.
Wei, J., Wu, T., Deng, X., Haider, S.W., Kahkashan, S., and Yang, S. 2021. Seafloor methane emission on the Makran continental margin. Science of The Total Environment 801: 149772. Crossref
Wenz, W., 1938–1944. Gastropoda. Teil 1: Allgemeiner Teil und Prosobranchia. In: O.H. Schindewolf (ed.), Handbuch der Paläozoologie, Band 6. [1, 1–240, March 1938; 3, 241–480, October 1938; 4, 481–720, July 1939; 6, 721–960, August 1940; 7, 961–1200, October 1941; 8, 1201–1506, October 1943; 9, 1507–1639 + i–xii , November 1944. Bornträger, Berlin.
Wiedicke, M., Sahling, H., Delisle, G., Faber, E., Neben, S., Beiersdorf, H., Marchig, V., Weiss, W., Von Mirbach, N., and Afiat, A. 2002. Characteristics of an active vent in the fore-arc basin of the Sunda Arc, Indonesia. Marine Geology 184 (1–2): 121–141. Crossref
Wiedicke-Hombach, M., Ardhayastuti, S., Bruns, A., Delisle, G., Goergens, R., Hermawan, T., Kanamatsu, T., Lückge, A., Mohtadi, M., Mühr, P., Rahadyan, T., Riyadi, A., Rühlemann, C., Schippers, A., Schlömer, S., Taufik, M.,Teichert, B.,Vink, A., Weiss, W., Wijaya, H., Wöhrl, C., Zeibig, M., and Zoch, D. 2006. SUMATRA—The Hydrocarbon System of the Sumatra Forearc–SO189-2 Cruise Report. Bundesanstalt für Geowissenschaften und Rohstoffe, Hannover.
Woodring, W.P. 1925. Miocene mollusks from Bowden Jamaica. Pelecypods and scaphopods. Carnegie Institution of Washington Publication 366: 1–222. Crossref
Woodward, S.P. 1859. On a new species of mollusk of the genus Scissurella D’Orbigny. Proceedings of the Zoological Society of London 27 (392–405): 202–204.
Xu, T., Feng, D., Tao, J., and Qiu, J.-W. 2019. A new species of deep-sea mussel (Bivalvia: Mytilidae: Gigantidas) from the South China Sea: Morphology, phylogenetic position, and gill-associated microbes. Deep Sea Research Part I: Oceanographic Research Papers 146: 79–90. Crossref
Yang, M., Li, B., Gan, Z., Dong, D., and Li, X. 2024 A new chemosymbiotic bivalve species of the genus Acharax Dall, 1908 (Bivalvia, Solemyida, Solemyidae) from the Haima cold seep of the South China Sea. ZooKeys 1198: 185–192. Crossref
Zanzerl, H., Salvo, F., Jones, S.W., and Dufour, S.C. 2019. Feeding strategies in symbiotic and asymbiotic thyasirid bivalves. Journal of Sea Research 145: 16–23. Crossref
Zettler, M.L. and Hoffman, L. 2021. A wide distribution of Waisiuconcha haeckeli (Bivalvia: Vesicomyidae) in the eastern Atlantic Ocean. Miscellanea Malacologica 9 (1): 1–3.
Zhang, K., Sun, J., Xu, T., Qiu, J.-W., and Qian, P.-Y. 2021. Phylogenetic relationships and adaptation in deep-sea mussels: Insights from mitochondrial genomes. International Journal Molecular Science 22 (4): 1900. Crossref
Acta Palaeontol. Pol. 70 (3): 443–477, 2025
https://doi.org/10.4202/app.01179.2024