
New Early Jurassic thylacocephalan assemblage from the Western Carpathians in Slovakia
ALEXIS GERBE, MATÚŠ HYŽNÝ, and JÁN SCHLÖGL
Gerbe, A., Hyžný, M., and Schlögl, J. 2025. New Early Jurassic thylacocephalan assemblage from the Western Carpathians in Slovakia. Acta Palaeontologica Polonica 70 (3): 557–571.
Thylacocephalans (Euarthropoda: Pancrustacea) are extinct marine pancrustaceans characterised by a folded shield, with fossils known from the Upper Ordovician to the Upper Cretaceous, reaching a diversity peak in the Triassic. We described two new thylacocephalan taxa, Dollocaris toarcica sp. nov. and Zazrivacaris jodorowskyi gen. et sp. nov., from the lower Toarcian, Lower Jurassic black shales of the “Hrýzeň beds” of the Šariš Unit (Grajcarek Succession) exposed in Zázrivá, Slovakia. Thylacocephalans are an accessory element of an euarthropod association, dominated by dendrobranchiate shrimps. This occurrence represents not only the first record of thylacocephalans from Slovakia but also their first formal description from the Toarcian. Dollocaris toarcica sp. nov. is the third described species of its genus, previously known from the Middle Jurassic (Callovian) of France and Upper Jurassic (Tithonian) of Germany, expanding the stratigraphic span of the genus to the Lower Jurassic. The newly described genus Zazrivacaris shows morphological similarities with Mayrocaris, yet another genus known from the Upper Jurassic (Tithonian) of Germany. Considering current knowledge, we present a review of all known micro- and macro-structural patterns present on thylacocephalan shields. These structures appear to be unique to each genus and/or species and are therefore important for taxonomic evaluation.
Key words: Arthropoda, Thylacocephala, Dollocaris, Zazrivacaris, Lower Toarcian, Jurassic, Slovakia, Carpathians.
Alexis Gerbe [alexis.gerbe@palaeo-evo-devo.info; ORCID: https://orcid.org/0009-0002-0609-2391 ], Université Claude Bernard Lyon 1, 43 boulevard du 11 Novembre 1918, 69622 Villeurbanne cedex, France. Ludwig-Maximilians-Universität München, Biocenter, Großhaderner Str. 2, 82152 Planegg-Martinsried, Germany.
Matúš Hyžný [matus.hyzny@uniba.sk; ORCID: https://orcid.org/0000-0002-8960-2846 ], Department of Geology and Paleontology, Faculty of Natural Sciences, Comenius University, Mlynská dolina, Ilkovičova 6, SK-842 15 Bratislava, Slovakia; Earth Science Institute, Slovak Academy of Sciences, Dúbravská cesta 9, P.O. Box 106, 840 05 Bratislava, Slovakia.
Ján Schlögl [jan.schlogl@uniba.sk; ORCID: https://orcid.org/0000-0001-7411-4324 ], Department of Geology and Paleontology, Faculty of Natural Sciences, Comenius University, Mlynská dolina, Ilkovičova 6, SK-842 15 Bratislava, Slovakia.
Received 18 June 2024, accepted 1 July 2025, published online 10 September 2025.
Copyright © 2025 A. Gerbe et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Thylacocephalans are among the most intriguing group of extinct marine pancrustaceans. They are characterised by a folded shield enclosing a major part of the body. Three pairs of sub-chelate raptorial appendages and large compound eyes suggest that at least some of these animals were likely predators (Schram et al. 1999; Vannier et al. 2006, 2016; Charbonnier et al. 2010, 2017; Broda and Zatoń 2017; Jobbins et al. 2020). Fossils of thylacocephalans have been known since the 19th century. Firstly, they were interpreted as representatives of Phyllocarida Packard, 1879 (Meek and Worthen 1868), Stomatopoda Latreille, 1816 (Hilgendorf 1885) or Mysida Haworth, 1825 (Van Straelen 1923). Discoveries made at the end of the 20th century, especially from Italy (Arduini et al. 1980; Pinna et al. 1982, 1985), Australia (Briggs and Rolfe 1983), France (Secretan and Riou 1983; Secretan 1985), and Germany (Polz 1994) helped to understand the unique systematic position of thylacocephalans among known arthropod groups and recognise them as probable representatives of the clade Pancrustacea Zrzavý & Štys, 1997 (Schram 2014; Schram and Koenemann 2022). A few decades later, our knowledge of these animals has significantly improved thanks to descriptions of soft part morphology and anatomy of their bodies (Charbonnier et al. 2010; Haug et al. 2014; Vannier et al. 2016; Jobbins et al. 2020; Laville et al. 2021a, 2023a), allowing for evaluation of taxonomic relationships of Thylacocephala within euarthropods (e.g., Haug et al. 2014; Broda and Zatoń 2017; Laville et al. 2021b). In this respect, every new occurrence helps to add much needed data for understanding the evolution and diversity of these animals.
The known fossil record of Thylacocephala includes 70 species (Table 1) and spans from the Upper Ordovician (Van Roy et al. 2021) to the Upper Cretaceous (Hilgendorf 1885; Roger 1946; Schram et al. 1999; Lange et al. 2001; Charbonnier et al. 2017), reaching a diversity peak in the Triassic (Charbonnier and Brayard 2019). Thylacocephalan fossils have been found throughout the world, including Europe (e.g., Van Straelen 1923; Glaessner 1931; Arduini et al. 1980; Charbonnier et al. 2010; Broda and Zatoń 2017; Rak et al. 2018; Braig et al. 2019), Asia (Dames 1886; Roger 1946; Ehiro et al. 2015, 2019; Charbonnier et al. 2017; Ji et al. 2017, 2021; Ehiro and Kano 2024), Africa (Arduini 1990; Jobbins et al. 2020; Laville et al. 2021c), North America (e.g., Schram 1990; Hegna et al. 2014; Charbonnier and Brayard 2019; Laville et al. 2021a), and Australia (Briggs and Rolfe 1983; Haig et al. 2015).
The occurrence of Thylacocephala from the Lower Jurassic (Toarcian) of Slovakia reported herein not only represents the first report of these animals from the respective country but also the first formal description of Thylacocephala from the Toarcian. Previous report on thylacocephalans from the Toarcian by Williams et al. (2015) is limited to a mere statement without taxonomic evaluation or figures. Our contribution also refines the classification and morphological characterization of the shield structures of thylacocephalans, serving as a foundation for future studies on shield structures.
Institutional abbreviations.—SNM Z, Natural History Museum of Slovak National Museum in Bratislava, Slovakia.
Other abbreviations.—Aad, anterodorsal angle; Aav, anteroventral angle; Apd, posterodorsal angle; Apv, posteroventral angle; Ha, anterior shield height; Hmax, maximum shield height; Hp, posterior shield height; L, length; Ls, maximum shield length.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in Zoobank: urn:lsid:zoobank.org:pub:1110D1FC-DDC7-42D6-84C8-5AE1446E4ED3.
Geological setting
The material forming the basis of this study was collected from a natural outcrop on the root of the Hrýzeň Hill (GPS N49.283299°, E19.175561°; Zázrivá district, Slovakia; Fig. 1). The Hrýzeň Hill is a narrow ridge on the left side of the Kozinský potok Brook Valley (Kozinská settlement), ca. 800 m east-northeast of the Zázrivá Centre and 300 m east of the crossroad Končitá-Havrania-Kozinská.
The ridge is predominantly formed by grey spotted marly limestones (Fleckenmergel-Fleckenkalk facies) that most probably belong to the Lower–Middle Jurassic Krempachy Marl Formation (equivalent of the Allgäu Formation). In the middle part of the ridge, there are signs of old mining attempts in the Toarcian black, brown weathered, Mn-bearing shales. In addition, clasts of micaceous quarzitic sandstones and black crinoidal limestones occur in the debris, possibly representing allodapic limestones from the upper part of the Middle Jurassic Szlachtowa Formation. The whole succession has been tentatively affiliated with the Šariš Unit (Grajcarek Succession) by Plašienka et al. (2021).
The black shale sequence at the washed-out bank of the Kozinský potok Brook was studied by Schlögl et al. (2012) and later in more detail by Suan et al. (2018). The studied section is 36 meters thick and is in a stratigraphically reversed position, with a dip of strata oriented to the SW. The lower and middle parts of the section are essentially composed of dark grey to black, finely laminated shales (black shales or “schistes-carton”), locally interrupted by dark grey laminated siltstone-mudstone beds. Black shales are locally rich in macrofauna, including ammonites, soft bodied cephalopods (Schlögl et al. 2012), bivalves, arthropods, echinoids, and fish remains. Marls in the upper part of the succession are grey in colour. Based on ammonites and calcareous nannofossils, the early Toarcian age was proposed for this sequence and was related to the Toarcian Oceanic Anoxic Event (T-OAE). Informally, this sequence is designated as the “Hrýzeň beds” (Plašienka et al. 2021: fig. 2.6). The locality records organic-rich sedimentation at the margin of NW-European shelf (Suan et al. 2018).
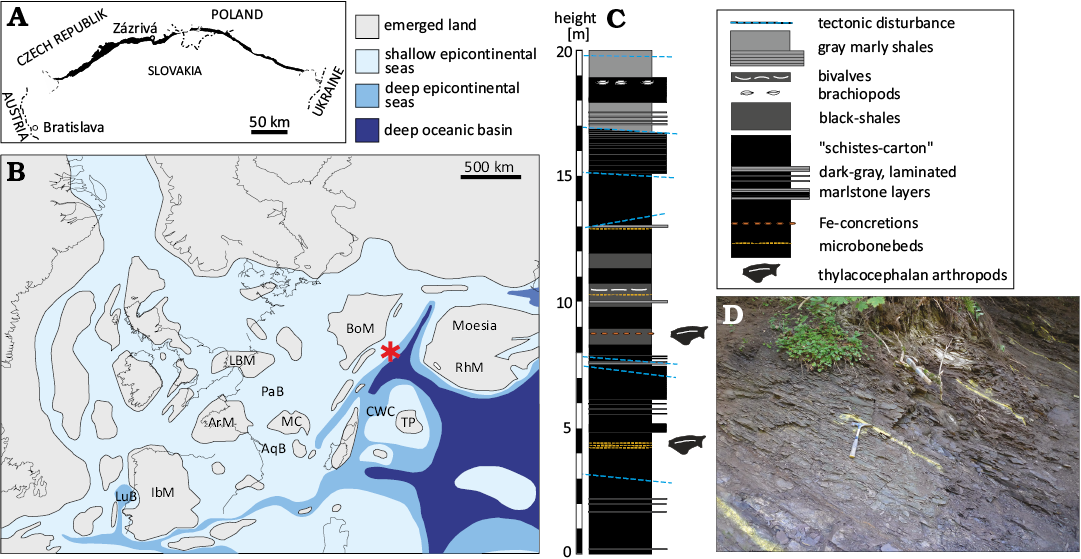
Fig. 1. A. Geographical location of the Zázrivá village. Dark area represents remains of the Pieniny Klippen Basin. B. Palaeogeographical position of the Hrýzeň section (asterisk) during Toarcian. AqB, Aquitaine Basin; ArM, Armorican Massif; BoM, Bohemian Massif; CWC, Central Western Carpathians; IbM, Iberian Massif; LBM, London-Brabant Massif; LuB, Lusitanian Basin; MC, Massif Central; PaB, Paris Basin; RhM, Rhodope Massif; TP, Tisza Plate. Map modified after Thierry (2000) and Dera et al. (2010). C. Middle part of the Hrýzeň section with the distribution of thylacocephalan fossils. D. The field photo showing typical black shale lithofacies bearing the crustacean remains. Level 9 marked by hammer, the succession is in overturned position, so the numbering descents towards the top of the outcrop.
Ammonites and calcareous nannofossils provide the basis for the biostratigraphy of the black shales (Suan et al. 2018). The co-occurrence of scarce Eleganticeras elegans Sowerby, 1815, and Eleganticeras exaratum Young & Bird, 1828, together with common Hildaites ex. gr. murleyi–levisoni in the middle part of the section, indicates the lower Toarcian Exaratum Subzone of the Serpentinum Zone. Overlying grey marls are poor in ammonites; however, Hildaites cf. subserpentinus and Harpoceras ex. gr. falciferum indicate the Falciferum Subzone of the late Serpentinum Zone. The first occurrence (FO) of calcareous nannofossil Carinolithus superbus Deflandre, 1954, marks the base of the NJ6 zone and the FO of Discorhabdus striatus Rood et al., 1971 characterises the base of the NJ7 zone (Bown and Cooper 1998; Mattioli and Erba 1999). Thylacocephalan pancrustaceans occur in two stratigraphic intervals, an older one between 4.1 m and 4.33 m and a slightly younger one between 8.4 m and 10.4 m (Fig. 1C). Zazrivacaris jodorowskyi gen. et sp. nov. comes only from the younger interval, whereas Dollocaris toarcica sp. nov. comes from both intervals. While only two specimens of D. toarcica sp. nov. were recovered from the 4.1–4.33 m interval, the vast majority of the thylacocephalan material, including both newly identified species, originates from the 8.4–10.4 m interval. The deposition of the intervals with thylacocephalans took place under sulphidic and anoxic conditions (euxinia) interrupted by brief events of improved oxygenation (Suan et al. 2018).
Material and methods
Fossils were collected during several field campaigns organized by the Department of Geology and Palaeontology, Faculty of Natural Sciences, Comenius University in Bratislava. The material consists of 18 specimens representing laterally flattened near complete shields, or fragments preserved in black shales. The specimens were further prepared under a stereomicroscope using a vibrating needle and later also preparatory needles.
Samples have been observed and documented in detail with a Leica DVM6 digital microscope with two different objectives: a PLANAPO FOV 12.55 and a PLANAPO FOV 43.75 with a polarized light. Additional documentation was done by Leica M205 C with a PLANAPO 0.63. An attempt to document fossils using the UV light was unsuccessful; the specimens from the studied locality do not exhibit UV fluorescence.
The morphological terms adopted in descriptions follow Laville et al. (2021b) and are provided in Fig. 2A. We refrained from using “anterior spine”; instead, we used the term “rostrum” which is widely used in descriptions of thylacocephalan shield (e.g., Charbonnier et al. 2017; Rak et al. 2018; Laville et al. 2021c). Measurements were made digitally with ImageJ and follow Laville et al. (2021b). Because the nature of the shield structures of D. toarcica sp. nov. is uncertain (ornamentation vs. integumental hemolymphatic circulatory network), we are using the term “shield structure” rather than “shield ornamentation”.
The material presented herein is deposited in the Natural History Museum of Slovak National Museum in Bratislava, Slovakia.
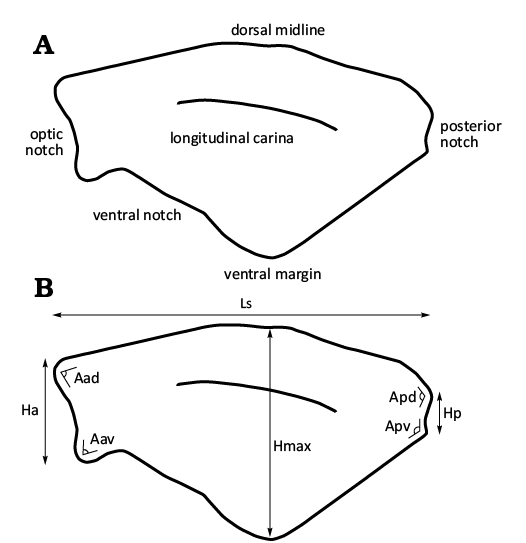
Fig. 2. Dollocaris toarcica sp. nov., shield structure scheme with terminology (A), angles and measurements (B). Not to scale.
Systematic palaeontology
Euarthropoda Lankester, 1904
Thylacocephala Pinna et al., 1982
Genus Dollocaris Van Straelen, 1923
Type species: Dollocaris ingens Van Straelen, 1923, by monotypy; Callovian, Middle Jurassic, France (la Voulte-sur-Rhône).
Original diagnosis (Secretan and Riou 1983).—The shield covers almost the entire body, except for a caudal mass that slightly protrudes posteriorly. It is approximately twice as long as it is wide, with a height roughly equal to its width. The lower lateral margins fold strongly over the sternal side. The shield is bordered by a marginal groove along its entire outline.
A longitudinal dorsal median groove extends along its full length. A short, pointed rostrum, slightly curved downward, extends from a medio-longitudinal ridge, which becomes more elevated towards the posterior before thinning abruptly and ending in a sudden step at the posterior third of the shield length. On either side of the rostrum, a very large notch in the anterior margin, terminating below in a strong spine, accommodates an extremely large, apparently sessile compound eye, which arises from a broad peduncle located beneath the shield. A transverse longitudinal ridge marks the shield laterally. A strong oblong protuberance elevates its lower region.
On either side of a short posterior median dorsal spine, a notch, less pronounced than that of the anterior margin, allows the caudal mass of the body to extend beyond the shield. The body consists of several differentiated tagmata. The anteroventral margin of the shield is the point of emergence for two categories of appendages. The first category consists of short appendages, presumed to be cephalic. The second category, presumed to be thoracic, consists of long appendages that increase in size posteriorly, bend forward, and are laterally superimposed. The last three are believed to end in spiniform pincers. A branchial chamber extends above these appendages under the shield, containing large, lamellar gills, with at least eight pairs.
The posteroventral margin is the point of origin for filamentous appendages emerging from strong, aligned plates that continue onto the caudal mass protruding beyond the shield. The posterior end of the body is known only by the presence of a short, apparently unpaired spine and indistinct foliaceous scales. No traces of a furca have been observed. [Translated from French from Secretan and Riou 1983: 78–79.]
Emended diagnosis.—Shield outline pentagonal to hexagonal in lateral view, approximately twice as long as hight, with median carina; optic notch higher than posterior notch; rostrum, if present, sharp and slightly curved downward; anterior midline convex; posterior notch round and concave; posterodorsal and posteroventral corners sharp; posteroventral margin descending about halfway up the shield; anteroventral margin ascending anteriorly; ventral margin with more- or less-pronounced ventral notch.
Remarks.—Ventral notch observed in the material from Zázrivá are unique to two genera: Dollocaris Van Straelen, 1923, and Mayrocaris Polz, 1994. Dollocaris differs from Mayrocaris in several aspects: its shield is almost twice as long as it is high, with a concave posterior notch and with a ventral margin, which is less curved anteriorly than in Mayrocaris. The new species presented further below exhibits a set of characters suggesting its attribution to Dollocaris.
Dollocaris can be distinguished from other thylacocephalan genera by several distinct shield features. Dollocaris possesses a ventral notch, which is missing in Ankitokazocaris Arduini, 1990, Clausocaris Oppenheim, 1888, Concavicaris Rolfe, 1961, Kilianicaris Van Straelen, 1923, Ligulacaris Charbonnier & Brayard, 2019, Ostenocaris Arduini et al., 1980, Paraostenia Secretan, 1985, and Yangzicaris Shen, 1983. Moreover, Dollocaris exhibits a less pronounced rostrum than that in Falcatacaris Braig et al., 2019, Microcaris Pinna, 1974, and Ferrecaris Calzada &Mañé, 1993.
Stratigraphic and geographic range.—Toarcian to Tithonian, Lower Jurassic to Upper Jurassic; France, Germany, Slovakia.
Dollocaris toarcica sp. nov.
Figs. 3, 4.
Zoobank LSID: urn:lsid:zoobank.org:act:31AF80D8-9941-4396-A1A7- 71AD192F12CF.
Etymology: In reference to the stratigraphic occurrence (Toarcian stage of the Lower Jurassic) of the species.
Type material: Holotype SNM Z 41680, complete shield. Paratypes SNM Z 41034–41041, 41683, nine shields in various states of completeness, from the type locality and horizon.
Type locality: The Hrýzeň Hill, Zázrivá district, Slovakia.
Type horizon: Serpentinum Zone (Exaratum Subzone), lower Toarcian, Lower Jurassic.
Material.—Type material and SNM Z 41042–41046, 41682, six shields of fragmentary nature, all from the type locality and horizon.
Diagnosis.—Shield outline hexagonal in lateral view; anteroventral margin with ventral notch extending from anteroventral corner to posteroventral margin; optic notch concave without rostrum; anteroventral corner rounded; posterior notch small and concave.
Description.—Shield outline hexagonal in lateral view, longer than height (Ls/Hmax mean ratio: 1.99; min: 1.76; max: 2.23). Optic notch large and not well pronounced (Ha: mean 4.57 mm; min 3.70 mm; max 5.76 mm). Anteroventral corner rounded with mean angle 57° (min 41.88°; max 70.33°). Anteroventral margin with well-developed square-shaped ventral notch. Posteroventral margin slightly concave and ascending posteriorly. Posteroventral corner sharp (Apv: mean 105.95°; min 93.28°; max 114.311°). Small posterior notch (Hp: mean 1.83 mm; min 1.16 mm; max 2.30 mm) rounded and concave. Posterodorsal corner sharp (mean 111.31°; min 83.81°; max 133.80°). Dorsal midline convex. Anterodorsal corner with mean angle 56° (min 41.89°; max 71.55°). Rounded muscle scar located at mid-length close to ventral margin. Shield with longitudinal carina taking approximately two thirds of shield length. Presence of curved ridges arranged largely sub-horizontally that could be interpreted as an integumental hemolymphatic circulatory network (see Discussion). Anteriorly, ridges converging at centre of shield; posteriorly, ridges converging to ventral margin; ventrally, ridges parallel with ventral margin. On dorsal part, near to carina, ridges end on other ridges. On ventral part, some ridges start in middle part of shield, near to carina.
Remarks.—Dollocaris toarcica sp. nov. differs from its congeners, Dollocaris ingens Van Straelen, 1923, and Dollocaris michelorum Polz, 2001, in several aspects. The shield of both previously described congeners are pentagonal and possess a well-developed rostrum, which is hexagonal, and the rostrum is absent in the species described herein. The anterior and posterior notches are less pronounced in D. toarcica sp. nov. than in D. michelorum and D. ingens. The latter shows an abrupt drop (i.e., dorsal carina) in the posterior third of the shield’s length that is absent in the new species. The ventral notch in D. michelorum is very slightly pronounced.
Clearly recognizable appendages and eyes are not preserved in the studied material of Dollocaris toarcica sp. nov., thus, these features cannot be readily compared with those in D. ingens. Nevertheless, some specimens of D. toarcica sp. nov. exhibit poorly preserved structures close to the anterior and posterior notches (SNM Z 41035, Fig. 3B) which could represent remains of anterior raptorial and posterior trunk appendages.

Fig. 3. Thylacocephalan arthropod Dollocaris toarcica sp. nov. from the lower Toarcian, Lower Jurassic of the Hrýzeň section, Zázrivá, Slovakia. A. Holotype SNM Z 41680, photograph (A1), line drawing (A2). B. Paratype SNM Z 41035; general view (B1), close-up pictures of potential anterior trunk appendages (B2), close-up pictures of potential posterior trunk appendages (B3). C. Paratype SNM Z 41038. D. Paratype SNM Z 41043. E. Paratype SNM Z 41036. F. Paratype SNM Z 41046. G. Paratype SNM Z 41683. Arrows indicate anterior (B2, G) and postarior (B3) possible trunk appendages.
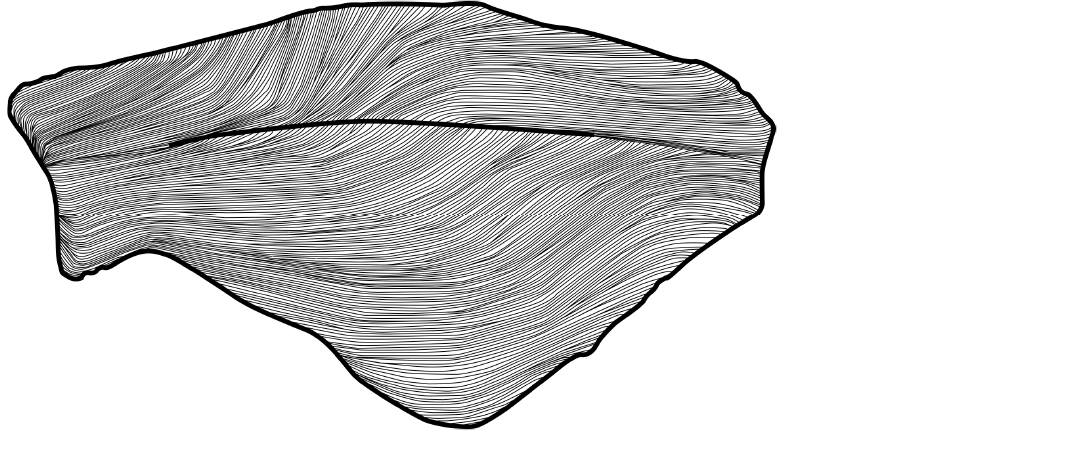
Fig. 4. Thylacocephalan arthropod Dollocaris toarcica sp. nov., macro-structures scheme of subparallel lines. Not to scale.
Present material is the first known occurrence of Dollocaris from Slovakia, while all previously reported occurrences are known from France (Van Straelen 1923; Secretan and Riou 1983; Secretan 1985; Charbonnier et al. 2010; Vannier et al. 2016) and Germany (Polz 2001; Laville et al. 2021b). Dollocaris ingens and D. michelorum are known from the Middle Jurassic (Callovian) and Upper Jurassic (Tithonian), respectively; thus, D. toarcica sp. nov. from the Lower Jurassic (Toarcian) of Slovakia is the oldest occurrence of the genus and represents a significant extension of its known geographic range and stratigraphic span.
Stratigraphic and geographic range.—Type horizon and locality only.
Genus Zazrivacaris nov.
Zoobank LSID: urn:lsid:zoobank.org:act:4411883A-DE40-4C8D-BF92- 683A23E30FE1.
Type species: Zazrivacaris jodorowskyi sp. nov., by monotypy; Toarcian, Lower Jurassic, Zázrivá, Slovakia.
Etymology: In reference to the settlement of Zázrivá, from where the type material originates. Gender of the genus: feminine.
Diagnosis.—As for the type species (by monotypy).
Remarks.—Zazrivacaris gen. nov. can be distinguished from Ankitokazocaris, Falcatacaris, Kilianicaris, Microcaris, Ferrecaris, and Concavicaris by the absence of a rostrum. The newly erected genus differs from Clausocaris by the overall morphology of the shield, which is cone-shaped in Clausocaris, while it is subtrapezoid in Zazrivacaris gen. nov. Paraostenia and Ostenocaris differ from Zazrivacaris gen. nov. by the rounded anteroventral and posterovental corner and by the concave anterior margin, respectively. Austriocaris Glaessner, 1931, and Paraclausocaris Laville et al., 2023a, differ from the new genus by an outline of the optic notch, which is concave and asymmetric. In Austriocaris, the optic notch is restricted to the ventral portion of the anterior margin, whereas in Zazrivacaris gen. nov. it extends along both the ventral and dorsal margins. Additionally, the dorsal midline in Austriocaris is straight, while in Zazrivacaris gen. nov. the posterior third exhibits a slight downward slope. Paraclausocaris is further distinguished by the presence of small sinuous ridges on the carapace shield, a feature absents in the new genus. Zazrivacaris gen. nov. differs from Dollocaris by the morphology of the ventral and dorsal margins, as well as the absence of a ventral notch. The overall morphology of the shield of Zazrivacaris gen. nov. is similar to Mayrocaris, indicating a possible close relationship between the two genera. However, the absence of a ventral notch, rounded and concave posterior notch and the anteroventral angle with rounded outline support the erection of a new genus.
Stratigraphic and geographic range.—Type locality and horizon only.
Zazrivacaris jodorowskyi sp. nov.
Fig. 5.
Zoobank LSID: urn:lsid:zoobank.org:act:96376AC1-D08A-4E72-9A16- 52CAA3A3BDA8.
Etymology: In honour of Alejandro Jodorowsky (born 17.02.1929), a Chilean and French avant-garde filmmaker and comic book writer, whose surreal imagery is reminiscent of uncommon morphology of thylacocephalan pancrustaceans.
Type material: Holotype SNM Z 41047, near-complete shield. Paratype SNM Z 41681, one fragmented shield, from the type locality and horizon.
Type locality: The Hrýzeň Hill, Zázrivá district, Slovakia.
Type horizon: Serpentinum Zone (Exaratum Subzone), lower Toarcian, Lower Jurassic.
Diagnosis.—Shield subtrapezoid in lateral view, optic notch concave, asymmetric and not clearly delineated; two thirds of dorsal midline straight, last third descending slightly posteriorly; posterior notch symmetrical and concave; rostrum absent; posteroventral margin straight and descending at 39° to central part of shield, from here anteroventral margin straight rising at 37° anteriorly. Large raptorial appendage as long as shield length.
Description.—Shield small, with subtrapezoid outline in lateral view (Ls 8.8 mm; Hmax 6.2 mm; Ls/Hmax ratio 1.4); asymmetrical optic notch (Ha 3.97 mm) taking up almost two thirds of maximum shield height, absence of rostrum. Anterodorsal angle sharp (Aad 89.26°). First two thirds of dorsal midline straight, last third descending slightly posteriorly. Posterodorsal angle sharp (Apd 81.64°). Posterior notch symmetrical and concave (Hp 2.60 mm). Posteroventral angle sharp (Apv 108.19°), posteroventral margin straight and descending at 39° anteriorly. Anteroventral margin rising to 37° anteriorly. Muscle scar very large. Potential location where appendages were attached visible above muscle scar. Antero ventral angle sharp (Aav 103.79°). Raptorial appendage poorly preserved (L 8.78 mm), indistinct delimitation between each appendage element.

Fig. 5. Thylacocephalan arthropod Zazrivacaris jodorowskyi gen. et sp. nov. holotype (SNM Z 41047) from the lower Toarcian, Lower Jurassic of the Hrýzeň section, Zázrivá, Slovakia. Photograph (A1), line drawing (A2), close-up picture of the shied (A3).
Remarks.—The asymmetric and concave optic notch, symmetric and concave posterior notch, two thirds of dorsal midline straight, last third descending slightly posteriorly, the ventral margin subdivided into an anterior sub-horizontal part, and a posterior part slightly descending anteroventrally, absence of rostrum and the large appendages represent a unique combination of characters warranting a designation of a new genus. The general morphology of the shield is somewhat similar to Ankitokazocaris chaohuensis Ji et al., 2017, from the Triassic of China. However, our specimen presents Ls/Hmax ratio of 1.4, while it is 2.0 in A. chaohuensis (Ji et al. 2017). The anteroventral and the posteroventral margins of our specimen are of approximately the same size (anteroventral margin: 4.5 mm, posteroventral margin: 4.7 mm), while A. chaohuensis exhibits a shorter posteroventral margin (Ji et al. 2017). In our material, anterior and medial ventral margins form an angle of 107°, while in A. chaohuensis the angle is 125–145° (Ji et al. 2017). Moreover, Zazrivacaris jodorowskyi gen. et sp. nov. has a concave posterior notch, while A. chaohuensis has a straight posterior margin.
Another morphologically similar species is Mayrocaris bucculata Polz, 1994, which, however, differs in the presence of ventral notch and sub-quadratic posterior notch. The mean value of the Apd in M. bucculata (Apd 81.68°; Laville et al. 2021b) is similar to the mean value in Zazrivacaris jodorowskyi gen. et sp. nov (Apd 81.64°). The Hp is 2.6 mm in both species (Laville et al. 2021b; this study). The Ls/Hmax ratio is also quite similar, the mean ratio is 1.6 mm in M. bucculata (Laville et al. 2021b) and 1.4 in Z. jodorowskyi sp. nov. However, Ha (3.97°), Apv (89°) and Aav (103°) of the new species differ significantly from M. bucculata (Ha 5.9°; Apv 66.15°; Aav 114.3°) (Laville et al. 2021b).
Zazrivacaris jodorowskyi gen. et sp. nov. differs from co-occurring Dollocaris toarcica sp. nov. by several features, including the shield outline, outline of the optic notch, absence of a ventral notch and a faint posterior notch. Additionally, the Ls/Hmax ratio is 1.4 for Z. jodorowskyi sp. nov., while it is 2.0 in D. toarcica sp. nov. The posterior notch is wider, and the optic notch is smaller in Z. jodorowskyi sp. nov. than in D. toarcica sp. nov..
Stratigraphic and geographic range.—Type locality and horizon only.
Discussion
Anatomy of Zazrivacaris jodowskyi gen. et sp. nov.—The holotype of Zazrivacaris jodorowskyi sp. nov. displays three structures situated above the muscle scar. Although their precise function remains uncertain, their number and general morphology suggest that they represent attachment sites for thoracic appendages, reminiscent of those observed in some shrimps. Interestingly, similar structures have been observed in shrimp specimens from the same assemblage as the thylacocephalans described herein (AG, unpublished data). Moreover, the appendages of Mayrocaris bucculata (i.e., a taxon potentially closely related to Zazrivacaris jodorowskyi gen. et sp. nov.) appear to extend at least halfway up the optic notch (see Laville et al. 2021b: figs. 4b, c, f, g; 6b, d; 8d, e; 11c; 12c; 16). The position and morphology of the structures observed in Zazrivacaris jodorowskyi gen. et sp. nov. may also suggest a possible interpretation as gills. In thylacocephalans, gills were typically laminar and located in the anterior to mid-body region, just above the raptorial trunk appendages (e.g., Secretan and Riou 1983; Laville et al. 2023b). However, only three putative gills are visible in the specimen, whereas up to eight pairs are usually reported (Secretan and Riou 1983). Given the preservation state of the material, the identity and function of these structures remain uncertain.
Morphological variations.—Because majority of studied specimens of Dollocaris toarcica sp. nov. represents incomplete shields, not all measurements are available for every specimen (for measurements, see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app70-Gerbe_etal_SOM.pdf). Due to this limited dataset, statistical analyses could not be performed. The measured values of Hp, Aad, and Apd exhibit a large variability, Hp 1.15–2.29 mm; Aad 41.88–71.55° and Apd 83.81–133.808°. The values of Hmax, Ha, Aav, and Apv also vary, but to a lesser extent. Most of the maximum values were recorded for SNM Z 41680, while most of the minimum values were measured on SNM Z 41043. Dimorphism among thylacocephalans is still largely unknown. Laville et al. (2021b) observed a bimodal distribution for Aad, which might be linked to dimorphism for Mayrocaris bucculata, although the connection remains somewhat unclear. Charbonnier et al. (2017) hypothesised that morphological variation in size could also be linked to sexual dimorphism but have not been able to demonstrate this due to our limited knowledge of thylacocephalan biology. Briggs and Rolfe (1983) suggested a potential sexual dimorphism for Concavicaris milesi Briggs & Rolfe, 1983, and Concavicaris playfordi Briggs & Rolfe, 1893. The small sample size and lack of statistical analyses prevent us from reaching a definitive conclusion about the presence of dimorphism in the newly described material from Zázrivá.
Shield features of Zazrivacaris jodorowskyi gen. et sp. nov. are characterized by a short shield and a large optic notch occupying almost the entire height of the shield. These features are associated with juveniles (Briggs and Rolfe 1983; Lange et al. 2001). Charbonnier et al. (2010) presented juveniles of Dollocaris ingens based on the morphological size variation. In addition, Laville et al. (2023a) suggested, based on a morphometric analysis, that juveniles of Paraostenia voultensis Secretan, 1985, may be represented by smaller absolute size, square-shaped shield, posterior margin tilted and no field of punctuactions; instead of adult representatives that showed larger absolute size, trapezoidal-shaped shield, vertical posterior margin and a field of punctuations. However, since Z. jodorowskyi gen. et sp. nov. is known from only two specimens, investigating possible ontogenetic changes is not possible.
Shield structures.—In Thylacocephala, shield surface structures are important features for taxonomic evaluation which have been used as diagnostic characters on lower (e.g., Rak et al. 2018) and also higher taxonomic levels (e.g., Charbonnier et al. 2017). Two main types of surface structures have been recognized: micro-structures and macro-structures (Figs. 6, 7; see also Rak et al. 2018; Broda et al. 2020). The micro-structure consists of the morphology of the cells (often called micro-ornamentation) that have been divided into three types: linear, polygonal or scale-like (Broda et al. 2020). Several other features are considered as surface micro-structures such as lattice organs, setal lumina, grooves and tube-like structures (Fig. 6; Lange and Schram 2002; Broda et al. 2020). The surface macro-structures have not been investigated or exhaustively synthetised as micro-structures. However, different patterns in macro-structures are observable: ridges (often called terraces, lirae, lines or ribs) (e.g., Calzada and Mañé 1993; Ehiro et al. 2015; Rak et al. 2018; this study), pore(s) (e.g., Charbonnier et al. 2017; Schram and Koenemann 2022; Laville et al. 2023a), carinae (e.g., Briggs and Rolfe 1983; Ji et al. 2017; Charbonnier and Brayard 2019), punctuations (e.g., Charbonnier et al. 2017; Laville et al. 2023a, 2024b), furrow(s) (e.g., Laville et al. 2021c, 2024b), tubercles (e.g., Laville et al. 2024a, b), grooves, depressions (e.g., Laville et al. 2021c, 2024b), pits (e.g., Briggs and Rolfe 1963; Charbonnier et al. 2017), serrations (Laville et al. 2023a), humps (Laville et al. 2023a) or a combination of multiple types, for instance ridges plus pores (e.g., Charbonnier et al. 2017; Laville et al. 2021b; Fig. 7). The macro-structures are defined by the type of structures (punctuations, carinae, ridges and/or other macro-structures), the quantity/density of structures, and their overall pattern/orientation in respect to main shield axes. As far as we know, ridges can be oriented vertically (e.g., Ehiro et al. 2015), obliquely (Charbonnier and Brayard 2019) or following a more complex pattern (e.g., Rak et al. 2018; this study). Each thylacocephalan species discovered so far exhibits a unique macro-structures (if present) pattern which can therefore be used for an assignment on the species level. It is of note that terminology describing surface micro- and macro-structure features is not uniform across published contributions; the same morphological features are described using various terms. For instance, ridges have been called lirae (Rak et al. 2018; Broda et al. 2020), vertical ridges (Laville et al. 2021c), rugations (Charbonnier et al. 2017), terraces (Briggs and Rolfe 1963) or ribs (Ehiro et al. 2015). Several terms have been introduced without precise definitions, resulting in different terms being used for similar or maybe identical surface structures, such as punctuations, pores, and pits (e.g., Briggs and Rolfe 1983; Charbonnier et al. 2017; Ehiro et al. 2019; Laville et al. 2023a), or grooves, depressions, and furrows (e.g., Charbonnier and Brayard 2019; Jobbins et al. 2020; Laville et al. 2023a). It should be noted that we focused our synthesis exclusively on the surface structures. Therefore, structures found along the shield margins are not included such as pores (e.g., Laville et al. 2021a), serrations (e.g., Arduini and Brasca 1984; Arduini 1992), spines (e.g., Ji and Tintori 2024; Laville et al. 2024a), or square-shaped tooth-like structures (e.g., Braig et al. 2019).

Fig. 6. Line drawings of all surface shield micro-structures known to date. A–C, E, based on Broda et al. (2020); D, based on Lange and Schram (2002).
In respect to current literature data, stratigraphically oldest thylacocephalans from the Ordovician and Silurian have smooth shields (Van Roy et al. 2021; Haug et al. 2014, respectively), while the first occurrence of thylacocephalans bearing shield structures is dated to Devonian (Chlupáč 1963; Briggs and Rolfe 1983; Broda et al. 2020). Ridges seem more often present in Triassic thylacocephalans (e.g., Arduini 1990; Ehiro and Kano 2024; Laville et al. 2024a), whereas pores appear more common from Middle Jurassic to Late Cretaceous species (Dames 1886; Roger 1946; Lange et al. 2001; Charbonnier et al. 2017; Laville et al. 2023a) (Table 1). The smooth shield of the Ordovician specimens may be due to misidentification, as the material reported by Van Roy et al. (2021) may not belong to thylacocephalans. Indeed, the characteristic appendages are not visible. As for the only known Silurian specimen, it represents an insufficiently preserved shield, preventing any assessment of potential surface structures. Interestingly, only the species of Paraostenia Secretan, 1985, present furrows on their shield and might be a specific structure to Paraostenia. The “furrow and ridge” structure described in Ankitokazocaris bandoi may correspond to a lateral carina that is poorly preserved. Nevertheless, the fossil record of thylacocephalans is still patchy and major conclusions cannot be drawn yet.

Fig. 7. Line drawings of all surface shield macro-structures known to date. A. Ankitokazocaris triassica. B. Dollocaris toarcica sp. nov. C. Paraconcavicaris viktoryni. D. Ankitokazocaris utatsuensis. E. Ligulacaris parisiana. F. Thylacocephalus cymolopos. G. Mayrocaris bucculata.
Table 1. Species of Thylacocephala described to date with notion on the presence/absence of micro-/macro-structures. – indicates a true absence; × indicates that shield structures have not been investigated.
|
Species |
Micro-structures |
Macro-structures |
Age |
Country |
References |
|
Bohemiacaris libori Van Roy et al., 2021 |
× |
– |
Late Ordovician |
Czech Republic |
|
|
Pseudoprotozoea irenae Van Roy et al., 2021 |
× |
– |
Late Ordovician |
Czech Republic |
|
|
Ainiktozoon loganense Scourfield, 1937 |
× |
– |
Llandovery |
Scotland |
|
|
Thylacares brandonensis Haug et al., 2014 |
× |
× |
Llandovery |
USA |
|
|
Concavicaris desiderata (Barrande, 1872) |
× |
– |
Early Devonian |
Czech Republic |
|
|
Concavicaris sinuata (Meek & Worthen, 1868) |
scale |
– |
Pennsylvanian |
USA |
Meek and Worthen 1868; Zangerl and Richardson 1963; Briggs and Rolfe 1983; Schram 1990; Rak et al. 2018; Broda et al. 2020 |
|
Concavicaris campi Briggs & Rolfe, 1983 |
tube-like |
carina(e) |
Late Devonian |
Australia |
|
|
Concavicaris elytroides (Meek, 1812) |
– |
ridges/carina(e) |
Late Devonian–Mississippian |
USA |
|
|
Concavicaris glenisteri Briggs & Rolfe, 1983 |
tube-like |
carina(e)/pits |
Late Devonian |
Australia |
|
|
Concavicaris incola Chlupáč, 1963 |
scale |
– |
Late Devonian |
Czech Republic |
|
|
Concavicaris martinae Broda et al., 2020 |
polygonal/setal lumina/grooves |
carina(e) |
Late Devonian |
Czech Republic |
|
|
Concavicaris milesi Briggs & Rolfe, 1983 |
scale/ |
ridges/carina(e) |
Late Devonian |
Australia |
|
|
Concavicaris pikae Zatoń et al., 2014 |
polygonal/setal lumina/tube-like |
carina(e) |
Late Devonian |
Poland |
|
|
Concavicaris playfordi Briggs & Rolfe, 1983 |
polygonal |
carina(e)/pits |
Late Devonian |
Australia |
|
|
Concavicaris submarinus Jobbins et al., 2020 |
polygonal |
depression(s) |
Late Devonian |
Morocco |
|
|
Concavicaris woodfordi (Cooper, 1932) |
× |
depression(s) |
Late Devonian |
USA |
|
|
Concavicaris woodruffensis Broda et al., 2020 |
polygonal/setal lumina |
carina(e) |
Late Devonian |
USA |
|
|
Harrycaris whittingtoni Briggs and Rolfe, 1983 |
scale/ |
ridges/ |
Late Devonian |
Australia |
|
|
Suttropocaris bottkei Koch et al., 2003 |
× |
carina(e) |
Late Devonian |
Germany |
|
|
Concavicaris bradleyi (Meek, 1871) |
polygonal |
carina(e) |
Mississippian |
USA |
|
|
Paraconcavicaris viktoryni Rak et al., 2018 |
linear |
ridges/carina(e) |
Mississippian |
Czech Republic |
|
|
Concavicaris georgeorum Schram, 1990 |
× |
carina(e) |
Pennsylvanian |
USA |
|
|
Convexicaris mazonensis Schram, 1990 |
× |
carina(e) |
Pennsylvanian |
USA |
|
|
Eodollocaris keithflinti Laville et al., 2021a |
× |
tubercules |
Pennsylvanian |
USA |
|
|
Ankitokazocaris acutirostris Arduini, 1990 |
linear |
pore(s)/tubercules/Groove(s) |
Early Triassic |
Madagascar |
|
|
Ankitokazocaris bandoi Ehiro et al., 2015 |
× |
carina(e) |
Early Triassic |
Japan |
|
|
Ankitokazocaris chaohuensis Ji et al., 2017 |
× |
carina(e) |
Early Triassic |
China, Japan |
|
|
Ankitokazocaris parva (Ehiro et al., 2019) |
× |
pits |
Early Triassic |
Japan |
|
|
Ankitokazocaris triassica (Charbonnier & Brayard, 2019) |
× |
ridges/carina(e)/groove(s) |
Early Triassic |
USA |
|
|
Diplacanthocaris chaohuensis Ji et al., 2021 |
× |
– |
Early Triassic |
China |
|
|
Ankitokazocaris utatsuensis Ehiro et al., 2015 |
× |
ridges |
Early Triassic |
Japan |
|
|
Ligulacaris parisiana Charbonnier & Brayard, 2019 |
× |
carina(e) |
Early Triassic |
USA |
|
|
Microcaris rectilineatus Ji et al., 2021 |
× |
ridges/carina(e) |
Early Triassic |
China |
|
|
Paraostenia ambatolokobensis (Arduini, 1990) |
× |
ridges/carina(e)/furrow(s) |
Early Triassic |
Madagascar |
|
|
Parisicaris naoyai Ehiro & Kano, 2024 |
× |
ridges |
Early Triassic |
Japan |
|
|
Yangzicaris xiangxiensis Shen, 1983 |
× |
carina(e)/tubercules |
Middle Triassic |
China |
|
|
Species |
Micro-structures |
Macro-structures |
Age |
Country |
References |
|
Ferrecaris magransi Calzada & Mañé, 1993 |
× |
ridges |
Middle Triassic |
Spain |
|
|
Atropicaris lintveri Laville et al., 2024a |
× |
ridges/carina(e)/tubercules |
Middle Triassic |
Slovenia |
|
|
Kamnikaris cemazevkaensis Laville et al., 2024a |
× |
ridges |
Middle Triassic |
Slovenia |
|
|
Ankitokazocaris lariensis Ji & Tintori, 2024 |
× |
carina(e) |
Middle Triassic |
Italy |
|
|
Stoppanicaris grignaensis Ji & Tintori, 2024 |
× |
– |
Middle Triassic |
Italy |
|
|
Microcaris minuta Pinna, 1974 |
× |
ridges/carina(e) |
Late Triassic |
Italy |
Pinna 1974, 1976; Arduini 1988; Dalla Vecchia 1993; Dalla Vecchia and Muscio 1990 |
|
Microcaris ajdovskae Laville et al., 2024b |
× |
ridges |
Late Triassic |
Slovenia |
|
|
Paraostenia striata (Glaessner, 1931) |
× |
ridges/tubercules/furrow(s)/ |
Late Triassic |
Austria |
|
|
Austriocaris carinata Glaessner, 1931 |
× |
carina(e)/ |
Late Triassic |
Austria |
Glaessner 1931; Forchielli and Pervesler 2013; Laville et al. 2024b |
|
Clausocaris pinnai Arduini, 1992 |
× |
– |
Late Triassic |
Italy |
|
|
Atropicaris rostrata Arduini & Brasca, 1984 |
× |
ridges/carina(e) |
Late Triassic |
Italy |
|
|
Dollocaris toarcica sp. nov. |
– |
ridges/carina(e) |
Early Jurassic |
Slovakia |
this study |
|
Zazrivacaris jodorowskyi gen. et sp. nov. |
– |
– |
Early Jurassic |
Slovakia |
this study |
|
Ostenocaris cypriformis Arduini et al., 1980 |
× |
carina(e) |
Early Jurassic |
Italy |
|
|
Austriocaris secretanae Laville et al., 2023a |
× |
ridges/carina(e)/pore(s) |
Middle Jurassic |
France |
|
|
Dollocaris ingens Van Straelen, 1923 |
lattice organs? |
ridges/carina(e) |
Middle Jurassic |
France |
Van Straelen 1923; Secretan and Riou 1983; Secretan 1985; Charbonnier et al. 2010; Vannier et al. 2016 |
|
Kilianicaris lerichei Van Straelen, 1923 |
× |
carina(e)/pore(s)/ |
Middle Jurassic |
France |
Van Straelen 1923; Secretan and Riou 1983; Secretan 1985; Charbonnier et al. 2020; Laville et al. 2023a |
|
Ostenocaris ribeti (Secretan, 1985) |
× |
carina(e)/tubercules |
Middle Jurassic |
France |
|
|
Paraclausocaris harpa Laville et al., 2023a |
× |
ridges/carina(e)pore(s)/serrations/depression(s)/groove(s)/hump |
Middle Jurassic |
France |
|
|
Paraostenia voultensis Secretan, 1985 |
× |
carina(e)/pore(s)/punctuations/ |
Middle Jurassic |
France |
|
|
Clausocaris lithographica Oppenheim, 1888 |
× |
– |
Late Jurassic |
Germany |
Polz 1989, 1990, 1992; Haug et al. 2014; Laville et al. 2021b |
|
Dollocaris michelorum Polz, 2001 |
× |
ridges/carina(e) |
Late Jurassic |
Germany |
|
|
Falcatacaris bastelbergeri Braig et al., 2019 |
× |
carina(e) |
Late Jurassic |
Germany |
|
|
Mayrocaris bucculata Polz, 1994 |
– |
ridges/carina(e)/pore(s) |
Late Jurassic |
Germany |
|
|
Polzia eldoctorensis Hegna et al., 2014 |
× |
– |
mid-Cretaceous |
USA |
|
|
Victoriacaris muhiensis Hegna et al., 2014 |
× |
– |
mid-Cretaceous |
USA |
|
|
Globulocaris garassinoi Charbonnier et al., 2017 |
× |
– |
mid-Cretaceous |
Lebanon |
|
|
Paradollocaris vannieri Charbonnier in Charbonnier et al., 2017 |
× |
carina(e)/ |
mid-Cretaceous |
Lebanon |
|
|
Thylacocaris schrami Audo & Charbonnier in Charbonnier et al., 2017 |
× |
punctuations |
mid-Cretaceous |
Lebanon |
|
|
Hamaticaris damesi (Roger, 1946) |
lattice organs/tube-like |
carina(e)/ |
Late Cretaceous |
Lebanon |
Schram et al. 1999; Lange and Schram 2002; Charbonnier et al. 2017; Broda et al. 2020 |
|
Keelicaris deborae Teruzzi & Charbonnier in Charbonnier et al., 2017 |
× |
ridges/carina(e)/punctuations |
Late Cretaceous |
Lebanon |
|
|
Protozoea hilgendorfi Dames, 1886 |
lattice organs/tube-like |
pore(s) |
Late Cretaceous |
Lebanon |
Schram et al. 1999; Lange and Schram 2002; Charbonnier et al. 2017; Broda et al. 2020 |
|
Pseuderichthus cretaceus Dames, 1886 |
× |
pore(s) |
Late Cretaceous |
Lebanon |
Schram et al. 1999; Lange and Schram 2002; Charbonnier et al. 2017 |
|
Thylacocephalus cymolopos Lange et al., 2001 |
× |
pore(s) |
Late Cretaceous |
Lebanon |
Not all thylacocephalan taxa exhibit some type of shield structures. Nevertheless, the absence of micro-structure can be caused by exfoliation of the external, ornamented cuticle layers (Broda et al. 2015, 2020) and can therefore represent a taphonomic artefact. Moreover, several taxa were examined in earlier studies (e.g., Glassner 1931; Arduini and Brasca 1984; Secretan and Riou 1983; Polz 2001; Laville et al. 2021a; Ehiro and Kano 2024), in which authors did not specifically address micro-structures. The presence of structures on the shield are important features for taxonomic evaluation and could be linked with the mode of life of the thylacocephalans.
Similar ridges found in Dollocaris toarcica sp. nov. have been described on the inner layer of the shield in other thylacocephalans such as D. ingens (Secretan & Riou, 1983; Vannier et al. 2016), Paraostenia ambatolokobensis (Arduini, 1990) (Laville et al. 2021c), Concavicaris rostellata Rolfe, 1969 (species not formally described); Concavicaris elytroides Meek, 1872 (Briggs and Rolfe 1983) and the possible thylacocephalan Decoracaris hildebrandi Briggs et al., 2015. Vannier et al. (2016) interpreted these macro-structures as an integumentary hemolymphatic circulatory network, given their resemblance to those observed in certain myodocopans.
Thylacocephalan association.—The presence of Dollocaris and Zazrivacaris gen. nov. (possibly related to Mayrocaris) appears to be close to other Jurassic assemblages. From the Callovian La Voulte-sur-Rhône and Tithonian Solnhofen Lagerstätten, associations of Dollocaris and Mayrocaris have been reported (Polz 1994, 1997, 2001; Laville et al. 2023a). This observation is not surprising, because all these localities were once parts of the same marine basin, the Tethys Ocean.
Mode of life.—Thylacocephalans with and without structures have been found in diverse sedimentary settings, including carbonates (e.g., Lange et al. 2001; Ehiro et al. 2015; Charbonnier et al. 2017; Rak et al. 2018; Charbonnier and Brayard 2019), siliciclastic rocks (e.g., Glaessner 1931; Van Roy et al. 2021; this study), or concretions of various kind (e.g., Schram 1990; Briggs and Rolfe 1983; Jobbins et al. 2020; Laville et al. 2021a). The shales with Dollocaris toarcica sp. nov. originated under sulphidic and anoxic conditions interrupted by short events of improved oxygenation (Suan et al. 2018). The black shales at Zázrivá contain a decapod pancrustacean assemblage similar to that of the La Voulte-sur-Rhône Lagerstätte (Charbonnier et al. 2010). The presence of polychelidan lobsters (AG unpublished data) suggests a deep-sea environment. It is likely that the newly described thylacocephalans from this setting were benthic or nectobenthic and were probably killed during dysoxic or anoxic events. Unfortunately, only isolated shields of D. toarcica sp. nov. have been found; no preserved appendages were associated with shields. The black shales at Zázrivá delivered decapod pancrustaceans with preserved appendages. This may suggest that empty thylacocephalan shields were transported to their place of burial, although given the type of sedimentation the transport likely was not long.
Conclusions
The occurrence of Thylacocephala from the Toarcian, Lower Jurassic of Slovakia reported herein is the first report of this group of euarthropods from the respective country. The assemblage from the Zázrivá locality consists of two taxa, Dollocaris toarcica sp. nov. and Zazrivacaris jodorowskyi gen. et sp. nov., and represents the only formally described Thylacocephala from the Toarcian stage. The newly described species D. toarcica sp. nov. is the third described species of the genus Dollocaris, previously known from the Middle Jurassic (Callovian) of France and Upper Jurassic (Tithonian) of Germany, expanding the stratigraphic span of the genus to the Lower Jurassic. Zazrivacaris is a newly described genus, presenting unique combination of characters. Additionally, we examined the micro- and macrostructures of the thylacocephalan shield to better understand their macroevolution. However, due to preservation constraints and historical biases, no definitive conclusions can be drawn. Future studies utilizing advanced imaging technologies and/or newly discovered material will undoubtedly offer valuable insights into the evolutionary patterns of these structures.
Acknowledgements
Barbara Zahradníková (SNM) and Jozef Klembara (Comenius University, Bratislava, Slovakia) are thanked for access to the documentation facilities with the Leica DVM6 and Leica M205 C digital microscopes, respectively. We thank Thomas Laville (Ludwig-Maximilians-Universität, Munich, Germany) and Sylvain Charbonnier (Muséum national d’Histoire naturelle, Paris, France) for their helpful reviews. The research was supported by the Slovak Agency for Research and Development (APVV-22-0523) and the Slovak Scientific Grant Agency (VEGA 2/0106/23).
Editor: Andrzej Kaim
References
Alessandrello, A., Arduini, P., Pinna, G., and Teruzzi, G. 1991. New observations on the Thylacocephala (Arthropoda, Crustacea). In: A.M. Simonetta and S. Conway Morris (eds.), The Early Evolution of Metazoa and the Significance of Problematic Taxa, 245–251. Cambridge University Press, Cambridge.
Arduini, P. 1988. Microcaris and Atropicaris, two genera of the class Thylacocephala. Atti della Società Italiana di Scienze Naturalie del Museo Civico di Storia Naturale di Milano 129: 159–163.
Arduini, P. 1990. Studies on Permo-Trias of Madagascar. 4. Thylacocephala from Lower Trias of Madagascar. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano 131 (9): 197–204.
Arduini, P. 1992. Clausocaris pinnai n. sp., (Order Clausocarida nov.), thylacocephalan crustacean from the Norian of the Preone Valley (Udine, N. Italy) and morphological considerations on Thylacocephala. Atti della Società italiana di Scienze naturali e del Museo civico di Storia naturale di Milano 132 (21): 265–272.
Arduini, P. and Brasca, A. 1984. Atropicaris: nuovo genere della classe Thylacocephala. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 125 (1–2): 87–93.
Arduini, P., Pinna, G., and Teruzzi. G. 1980. A new and unusual Lower Jurassic cirriped from Osteno in Lombardy: Ostenia cypriformis n. g. n. sp. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 121: 360–370.
Barrande, J. 1872. Système silurien du centre de la Bohème. 1ère partie. Recherches paléontologiques. Supplément au Volume 1. Trilobites, crustacés divers et poissons. 647 pp. The Author, Prague.
Braig, F., Haug, J.T., Schädel, M., and Haug, C. 2019. A new thylacocephalan crustacean from the Upper Jurassic lithographic limestones of southern Germany and the diversity of Thylacocephala. Palaeodiversity 12 (1): 69–87. Crossref
Briggs, D.E.G. and Rolfe, W.D.I. 1983. New Concavicarida (new Order: ?Crustacea) from the Upper Devonian of Gogo, Western Australia, and the palaeoecology and affinities of the group. Special Papers in Palaeontology 30: 249–276.
Briggs, D.E.G., Liu, H.P., McKay, R.M., and Witzke, B.J. 2015. Bivalved arthropods from the Middle Ordovician Winneshiek Lagerstätte, Iowa, USA. Journal of Paleontology 89: 991–1006. Crossref
Bown, P.R. and Cooper, M.K.E. 1998. Jurassic. In: P.R. Bown (ed.), Calcareous Nannofossil Biostratigraphy, 34–85. Kluwer Academic Publishers, London. Crossref
Broda, K. and Zatoń, M. 2017. A set of possible sensory system preserved in cuticle of Late Devonian thylacocephalan arthropods from Poland. Historical Biology 29: 1045–1055. Crossref
Broda, K., Rak, Š., and Hegna, T.A. 2020. Do the clothes make the thylacocephalan? A detailed study of Concavicarididae and Protozoeidae (?Crustacea, Thylacocephala) carapace micro-ornamentation. Journal of Systematic Palaeontology 18: 911–930. Crossref
Broda, K., Wolny, M., and Zatoń, M. 2015. Palaeobiological significance of damaged and fragmented thylacocephalan carapaces from the Upper Devonian of Poland. Proceedings of the Geologists’ Association 126: 589–598. Crossref
Calzada, S. and Mañé, R. 1993. Primera cita de un Tilacocéfalo (Crustacea) (Ferrecaris n. gen. magransi n. sp.) en el Ladiniense español. Trabajos del Museo Geológico del Seminario de Barcelona 224: 13–17.
Charbonnier, S. and Brayard, A. 2019. New thylacocephalans from the Early Triassic Paris Biota (Bear Lake County, Idaho, USA). Geobios 54: 37–43. Crossref
Charbonnier, S., Teruzzi, G., Audo, D., Lasseron, M., Haug, C., and Haug, J.T. 2017. New thylacocephalans from the Cretaceous Lagerstätten of Lebanon. Bulletin de la Société géologique de France 188 (3): art. 19. Crossref
Charbonnier, S., Vannier, J., Hantzpergue, P., and Gaillard, C. 2010. Ecological significance of the arthropod fauna from the Jurassic (Callovian) La Voulte Lagerstätte. Acta Palaeontologica Polonica 55: 111–132. Crossref
Chlupáč, I. 1963. Phyllocarid crustaceans from the Silurian and Devonian of Czechoslovakia. Palaeontology 6: 97–118.
Cooper, C.L. 1932. A crustacean fauna from the Woodford Formation of Oklahoma. Journal of Paleontology 6: 346–352.
Dalla Vecchia, F.M. 1993. Segnalazione di crostacei nell’Unità Fonte Santa (Triassico Sup.) presso Filettino (Lazio, Italia). Gortania 14: 59–69.
Dalla Vecchia, F.M. 2012. Il Friuli 215 milioni di anni fa. Gli straordinari fossili di Preone: finestra su di un mondo scomparso. 224 pp. Comune di Preone, Preone.
Dalla Vecchia, F.M. and Muscio, G. 1990. Occurrence of Thylacocephala (Arthropoda, Crustacea) from the Upper Triassic of Carnic Prealps (NE Italy). Bollettino della Società-Paleontologica Italiana 29: 39–42.
Dames, W. 1886. Ueber einige Crustaceen aus den Kreideablagerungen des Libanon. Zeitschrift der Deutschen Geologischen Gesellschaft 38: 551–575.
Deflandre, G. 1954. Sur les coccolithophoridés fossiles en général et sur quelques formes nouvelles ou peu connues. Comptes Rendus de l’Académie des Sciences de Paris 239: 171–173.
Dera, G., Neige, P., Dommergues, J.-L., Fara, E., Laffont, R., and Pellenard, P. 2010. High-resolution dynamics of Early Jurassic marine extinctions: the case of Pliensbachian–Toarcian ammonites (Cephalopoda). Journal of the Geological Society London 167: 21–33. Crossref
Ehiro, M. and Kano, H. 2024. A new species of Parisicaris (Microcarididae, Thylacocephala) from the upper Olenekian (Lower Triassic) Osawa Formation in the South Kitakami Belt, Northeast Japan. Bulletin of the Tohoku University Museum 23: 23–34.
Ehiro, M., Sasaki, O., Kano, H., and Nagase, T. 2019. Additional thylacocephalans (Arthropoda) from the Lower Triassic (upper Olenekian) Osawa Formation of the South Kitakami Belt, Northeast Japan. Palaeoworld 28: 320–333. Crossref
Ehiro, M., Sasaki, O., Kano, H., Nemoto, J., and Kato, H. 2015. Thylacocephala (Arthropoda) from the Lower Triassic of the South Kitakami Belt, Northeast Japan. Paleontological Research 19: 269–282. Crossref
Forchielli, A. and Pervewsler, P. 2013. Phosphatic cuticle in thylacocephalans: a taphonomic case study of Austriocaris (Arthropoda, Thylacocephala) from the fossil largerstätte Polzberg (Reingraben Shales, Carnian, Upper Triassic, Lower Austria). Austrian Journal of Earth Sciences 106: 46–61.
Glaessner, M.F. 1931. Eine Crustaceenfauna aus den Lunzer Schichten Niederösterreichs. Jahrbuch der Geologischen Bundesanstalt 81: 467–486.
Haig, D.W., Martin, S.K., Mory, A.J., McLoughlin, S., Backhouse, J., Berrell, R.W., Kear, B.P., Hall, R., Foster, C.B., Shi, G.R., and Bevan, J.C. 2015. Early Triassic (early Olenekian) life in the interior of East Gondwana: mixed marine-terrestrial biota from the Kockatea Shale, Western Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 417: 511–533. Crossref
Haug, C., Briggs, D.E.G., Mikulic, D.G., Kluessendorf, J., and Haug, J.T. 2014. The implications of a Silurian and other thylacocephalan crustaceans for the functional morphology and systematic affinities of the group. BMC Evolutionary Biology 14: 159. Crossref
Haworth, A.H. 1825. A new binary arrangement of the macrurous Crustacea. Philosophical Magazine and Journal 65 (323): 183–184. Crossref
Hegna, T.A., Vega, F.J., and González-Rodríguez, K.A. 2014. First Mesozoic thylacocephalans (Arthropoda, ?Crustacea; Cretaceous) in the Western Hemisphere: new discoveries from the Muhi Quarry Lagerstätte. Journal of Paleontology 88: 606–616. Crossref
Hilgendorf, F. 1885. Ueber cretacische Squilliden-Larven vom Libanon. Sitzungs-Berichte der Gesellschaft naturforschender Freunde zu Berlin 1885: 184–185.
Ji, C. and Tintori, A. 2024. New Material of Thylacocephala from the Early Ladinian (Middle Triassic) of Northern Grigna (Lecco, Lombardy, Northern Italy). Diversity 16 (11): 677. Crossref
Ji, C., Tintori, A., Jiang, D., and Motani, R. 2017. New species of Thylacocephala (Arthropoda) from the Spathian (Lower Triassic) of Chaohu, Anhui Province of China. PalZ 91: 171–184. Crossref
Ji, C., Tintori, A., Jiang, D., Motani, R., and Confortini, F. 2021. New Thylacocephala (Crustacea) assemblage from the Spathian (Lower Triassic) of Majiashan (Chaohu, Anhui Province, South China). Journal of Paleontology 95: 305–319. Crossref
Jobbins, M., Haug, C., and Klug, C. 2020. First African thylacocephalans from the Famennian of Morocco and their role in Late Devonian food webs. Scientific Reports 10: 5129. Crossref
Koch, L., Gröning, E., and Brauckmann, C. 2003. Suttropcarididae n. fam. (Phyllocarida, Crustacea) aus dem Ober-Devon des Sauerlandes (Rheinisches Schiefergebirge, Deutschland). Neues Jahrbuch für Geologie und Paläontologie Monatshefte 2003 (7): 415–427. Crossref
Lange, S. and Schram, F.R. 2002. Possible lattice organs in Cretaceous Thylacocephala. Contributions to Zoology 71: 159–169. Crossref
Lange, S., Schram, F.R., Steeman, F.A., and Hof, C.H.J. 2001. New genus and species from the Cretaceous of Lebanon links the Thylacocephala to the Crustacea. Palaeontology 44: 905–912. Crossref
Latreille, P.A. 1816, Les crustacés, les arachnides et les insectes. In: G. Cuvier (ed.), Le règne animal distribué d’après son organisation, pour servir de base à l’histoire naturelle des animaux et d’introduction à l’anatomie comparée, Volume 3, i–xxix+1–653. Déterville, Paris.
Laville, T., Forel, M.-B., and Charbonnier, S. 2023a. Re-appraisal of thylacocephalans (Euarthropoda, Thylacocephala) from the Jurassic La Voulte-sur-Rhône Lagerstätte. European Journal of Taxonomy 898: 1–61. Crossref
Laville, T., Haug, J.T., and Haug, C. 2021a. New species of Thylacocephala, Eodollocaris keithflinti n. gen., n. sp., from the Mazon Creek Lagerstätte, Illinois, United States (c. 307 Ma) and redescription of other Mazon Creek thylacocephalans. Geodiversitas 43 (10): 295–310. Crossref
Laville, T., Haug, C., Haug, J.T., Forel, M.-B., and Charbonnier, S. 2021b. Morphology and anatomy of the Late Jurassic Mayrocaris bucculata (Eucrustacea?, Thylacocephala) with comments on the tagmosis of Thylacocephala. Journal of Systematic Palaeontology 19: 289–320. Crossref
Laville, T., Hegna, T., Forel, M.-B., Darroch, S., and Charbonnier, S. 2023b. New look at Concavicaris woodfordi (Euarthropoda: Pancrustacea?) using micro-computed tomography. Palaeontologia Electronica 26: a1. Crossref
Laville, T., Hitij, T., Forel, M.-B., Gašparič, R., Žalohar, J., and Charbonnier, S. 2024a. Middle and Late Triassic thylacocephalans from Slovenia: implications for their diversification. Bulletin of Geosciences 99: 219–253. Crossref
Laville, T., Lukeneder, P., and Lukeneder, A. 2024b. Revision of the thylacocephalan biota from the Upper Triassic Polzberg Konservat-Lagerstätte, Austria. Acta Palaeontologica Polonica 69: 713–730. Crossref
Laville, T., Smith, Christopher, P.A., Forel, M.-B., Brayard, A., and Charbonnier, S. 2021c. Review of Early Triassic Thylacocephala. Rivista Italiana di Paleontologia e Stratigrafia 127: 73–101.
Mattioli, E. and Erba, E. 1999. Synthesis of calcareous nannofossil events in Tethyan Lower and Middle Jurassic successions. Rivista Italiana di Paleontologia e Stratigrafia 105: 343–376.
Meek, F.B. 1871. Descriptions of New Western Palæozoic fossils, mainly from the Cincinnati group of the Lower Silurian Series of Ohio. Proceedings of the Academy of Natural Sciences of Philadelphia 23: 308–336.
Meek, F.B. 1875. A report on some of the invertebrate fossils of the Waverly Group and Coal Measures of Ohio. Report of the Geological Survey of Ohio 2 (2): 269–347.
Meek, F.B. 1872. Descriptions of new western Paleozoic fossils, mainly from the Cincinnati Group of the Lower Silurian Series of Ohio. Proceedings of the Academy of Natural Sciences of Philadelphia 24: 308–336.
Meek, F.B. and Worthen, A.H. 1868. Preliminary notice of a scorpion, an Eurypterus? and other fossils from the Coal Measures of Illinois. American Journal of Science Series 2 46: 19–28. Crossref
Oppenheim, P. 1888. Neue Crustaceenlarven aus dem lithographischen Schiefer Bayerns. Zeitschrift der Deutschen Geologischen Gesellschaft 40: 709–719.
Packard, A.S., Jr. 1879. The nebaliad Crustacea as types of a new order. American Naturalist 13: 128. Crossref
Plašienka, D., Aubrecht, R., Bezák, V., Bielik, M., Broska, I., Bučová, J., Fekete, K., Gaži, P., Gedl, P., Golej, M., Halásová, E., Hók, J., Hrdlička, M., Jamrich, M., Józsa, Š., Klanica, R., Konečný, P., Kubiš, M., Madarás, J., Majcin, D., Marko, F., Molčan Matejová, M., Potočný, T., Schlögl, J., Soták, J., Suan, G., Šamajová, L., Šimonová, V., Teťák, F., Vozár, J., and Vozár, J. 2021. Structure, composition and Tectonic Evolution of the Pieniny Klippen Belt-Central Western Carpathian Contiguous Zone (Kysuce and Orava Regions, NW Slovakia). 148 pp. Comenius University, Bratislava.
Pinna, G. 1974. I crostacei della fauna Triassica di Cene in Val Seriana (Bergamo). Memoria della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 21: 5–34.
Pinna, G. 1976. I crostacei Triassici dell’alta Valvestino (Brescia). Natura Bresciana 13: 33–42.
Pinna, G., Arduini, P., Pesarini, C., and Teruzzi, G. 1982. Thylacocephala: una nuova classe di crostacei fossili. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano 123: 469–482.
Pinna, G., Arduini, P., Pesarini, C., and Teruzzi, G. 1985. Some controversial aspects of the morphology and anatomy of Ostenocaris cypriformis (Crustacea, Thylacocephala). Earth and Environmental Science Transactions of The Royal Society of Edinburgh 76: 373–379. Crossref
Polz, H. 1989. Clausocaris nom. nov. pro Clausia Oppenheim, 1888. Archaeopteryx 7: 73.
Polz, H. 1990. Clausocaris lithographica (?Crustacea, Thylacocephala). Archaeopteryx 8: 93–109.
Polz, H. 1992. Zur Lebensweise der Thylacocephala (?Crustacea). Archaeopteryx 10: 1–12.
Polz, H. 1994. Mayrocaris bucculata gen. nov. sp. nov. (Thylacocephala, Conchyliocarida) aus den Solnhofener Plattenkalk. Archaeopteryx 12: 35–44.
Polz, H. 1997. Der Carapax von Mayrocaris bucculata (Thylacocephala, Conchyliocarida). Archaeopteryx 15: 59–71.
Polz, H. 2001. Dollocaris michelorum, sp. nov. (Thylacocephala, Concavicarida) aus den Solnhofener Plattenkalken. Archaeopteryx 19: 45–55.
Rak, Š., Broda, K., and Kumpan, T. 2018. First Carboniferous thylacocephalan from Europe and its significance for the understanding of functional morphology of Concavicarididae Schram, 2014. Crustaceana 91: 265–285. Crossref
Roger J. 1946. Invertébrés des couches à poissons du Crétacé supérieur du Liban. Mémoires de la Société géologique de France (Nouvelle série) 51: 5–92. Crossref
Rolfe, W.D.I. 1961. Concavicaris and Quasicaris, substitute names for Colpocaris Meek, 1872, and Pterocaris Barrande, 1872. Journal of Paleontology 35: 12–43.
Rolfe W.D.I. 1969. Phyllocarida. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part R, Arthropoda 4, 1, R296–R331. Geological Society of America & University of Kansas Press, Lawrence.
Rood, A.P., Hay, W.W., and Barnard, T. 1971. Calcareous nannoplankton and biostratigraphy of Late Cretaceous sediments in northeastern Mississippi and western Alabama. Bulletins of American Paleontology 61 (269): 1–79.
Schlögl, J., Košťák, M., and Hyžný, M. 2012. First record of a gladius-bearing coeloid Teudopsis bollensis Voltz (Cephalopoda, Coeloidea) in the Toarcian of the Western Carpathians (Slovakia). Paläontologisches Zeitschrift 86: 367–375. Crossref
Schram, F.R. 1990. On Mazon Creek Thylacocephala. Proceedings of the San Diego Society of Natural History 3: 1–16.
Schram, F.R. 2014. Family level classification within Thylacocephala, with comments on their evolution and possible relationships. Crustaceana 87: 340–363. Crossref
Schram, F.R. and Koenemann, S. 2022. Evolution and Phylogeny of Pancrustacea: A Story of Scientific Method. 827 pp. Oxford University Press, Oxford. Crossref
Schram, F.R., Hof, C.H.J., and Steeman, F.A. 1999. Thylacocephala (Arthropoda: Crustacea?) from the Cretaceous of Lebanon and implications for thylacocephalan systematics. Palaeontology 42: 769–797. Crossref
Scourfield, D.J. 1937. An anomalous fossil organism, possibly a new type of chordate, from the Upper Silurian of Lesmahagow, Lanarkshire—Ainiktozoon loganense, gen. et sp. nov. Proceedings of the Royal Society B 121 (825): 533–547. Crossref
Secretan, S. 1985. Conchyliocarida, a class of fossil crustaceans: relationships to Malacostraca and postulated behaviour. Transactions of the Royal Society of Edinburgh 76: 381–389. Crossref
Secretan, S. and Riou, B. 1983. Un groupe énigmatique de Crustacés: ses représentants du Callovien de la Voulte-sur-Rhône (France). Annales de Paléontologie 69 (4): 59–97.
Shen, Y. 1983. A new genus Yangzicaris (Phyllocarids) in the Middle Triassic of China. Acta Palaeontologica Sinica 22: 346–354.
Sowerby, J. 1815. The mineral conchology of Great Britain; or colored figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the Earth. Parts XV to XVIII. The Mineral Conchology of Great Britain 1 (15–18): 179–234.
Suan, G., Schöllhorn, I., Schlögl, J., Segit, T., Mattioli, E., Lécuyer, C., and Fourel, F. 2018. Euxinic conditions and high sulfur burial near the European shelf margin (Pieniny Klippen Belt, Slovakia) during the Toarcian oceanic anoxic event. Global and Planetary Change 170: 246–259. Crossref
Thierry, J. et al. (40 co-authors). 2000. Middle Toarcian. In: J. Dercourt, M. Gaetani, B. Vrielynck, E. Barrier, B. Biju-Duval, M.F. Brunet, J.P. Cadet, S. Crasquin, and M. Sandulescu (eds.), Atlas Peri-Tethys, Paleogeographical Maps. Map 8. CCGM/CGMW, Paris.
Van der Brugghen, W., Schram, F.R., and Martill. D. 1997. The fossil Ainiktozoon is an arthropod. Nature 385: 589–590. Crossref
Vannier, J., Huang, D., Charbonnier, S., Xiu-Qiang, W., and Chen, J.-Y. 2006. The Early Cambrian origin of thylacocephalan arthropods. Acta Palaeontologica Polonica 51: 201–214.
Vannier, J., Schoenemann, B., Gillot, T., Charbonnier, S., and Clarkson, E. 2016. Exceptional preservation of eye structure in arthropod visual predators from the Middle Jurassic. Nature Communications 7: 10320. Crossref
Van Roy, P., Rak, Š., Budil, P., and Fatka, O. 2021. Upper Ordovician Thylacocephala (Euarthropoda, Eucrustacea) from Bohemia indicate early ecological differentiation. Papers in Palaeontology 7: 1727–1751. Crossref
Van Straelen, V. 1923. Les Mysidacés du Callovien de la Voulte-sur-Rhône (Ardèche). Bulletin de la Société Géologique de France, Notes et Mémoires 23: 431–439.
Williams, M., Benton, M.J., and Ross, A. 2015. The Strawberry Bank Lagerstätte reveals insights into early Jurassic life. Journal of the Geological Society 172: 683–692. Crossref
Young, G. and Bird, J. 1828. A Geological Survey of the Yorkshire Coast. 367 pp. Clark, Whitby.
Zangerl, R. and Richardson, E.S., Jr. (eds.) 1963, The paleoecological history of two Pennsylvanian black shales. Fieldiana, Geology Memoirs 4: 1–352. Crossref
Zatoń, M., Filipiak, P., Rakociński, M., and Krawczyński, W. 2014. Kowala Lagerstätte: Late Devonian arthropods and non-biomineralized algae from Poland. Lethaia 47: 352–364. Crossref
Zrzavý, J. and Štys, P. 1997. The basic body plan of arthropods: Insights from evolutionary morphology and developmental biology. Journal of Evolutionary Biology 10: 353–367. Crossref
Acta Palaeontol. Pol. 70 (3): 557–571, 2025
https://doi.org/10.4202/app.01184.2024