
Crocodylian remains from the Miocene of the Fore-Carpathian Basin and its foreland—including the world’s northernmost Neogene crocodylian
MARCIN GÓRKA, JAKUB BŘEZINA, MILAN CHROUST, RAFAŁ KOWALSKI, SERGI LÓPEZ-TORRES, and MATEUSZ TAŁANDA
Górka, M., Březina, J., Chroust, M., Kowalski, R., López-Torres, S., and Tałanda, M. 2025. Crocodylian remains from the Miocene of the Fore-Carpathian Basin and its foreland—including the world’s northernmost Neogene crocodilian. Acta Palaeontologica Polonica 70 (2): 225–251.
The geographic distribution of Crocodylia in Europe throughout the Cenozoic experienced fluctuations in the extension of its northern limit. Whereas crocodylians reached very high latitudes during the early Eocene (78ºN), their northward extension was more moderate during almost the entire Paleogene and the Neogene. Here we reassess previous Early-to-Middle Miocene crocodylian records from the Fore-Carpathian Basin and its foreland, namely from marine limestones of Pińczów (Poland), and Židlochovice (Czechia) and from the new vertebrate site of Szczerców (Poland), currently interpreted as a freshwater paleoenvironment. All crocodylian material from these three sites represents Crocodylia indet. and its possible taxonomic attribution is discussed. The new Szczerców specimen, an osteoderm, represents the world’s northernmost record of a crocodylian from the entire Neogene.
Key words: Crocodylia, Diplocynodon, Gavialosuchus, Tomistoma, Miocene, Central Paratethys, Czechia, Poland.
Marcin Górka [magurka@uw.edu.pl; ORCID: https://orcid.org/0000-0001-7811-249X ], Department of Sedimentary Basins, Faculty of Geology, University of Warsaw, ul. Żwirki i Wigury 93, 02-089 Warsaw, Poland.
Jakub Březina [jbrezina@mzm.cz; ORCID: https://orcid.org/0000-0003-2087-6053 ], Department of Geology and Paleontology, Moravian Museum, Zelný trh 6, 659 37 Brno, Czechia.
Milan Chroust [chroust@twarda.pan.pl; ORCID: https://orcid.org/0000-0003-3301-5578 ], Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818 Warsaw, Poland
Rafał Kowalski [rafal.kowalski@mz.pan.pl; ORCID: https://orcid.org/0000-0002-1472-9930 ], Polish Academy of Sciences Museum of the Earth, al. Na Skarpie 20/26, 00-488 Warsaw, Poland.
Sergi López-Torres [s.lopez-torres@uw.edu.pl; ORCID: https://orcid.org/0000-0002-0046-1013 ] and Mateusz Tałanda [m.talanda@uw.edu.pl; ORCID: https://orcid.org/0000-0003-3358-9539 ], Institute of Evolutionary Biology, Faculty of Biology, University of Warsaw, ul. Żwirki i Wigury 101, 02-089 Warsaw, Poland.
Received 23 June 2024, accepted 19 February 2025, published online 9 May 2025.
Copyright © 2025 M. Górka et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
During the Early and Middle Miocene, distinct climatic events occurred. First, a relatively warm period, known as the Miocene Climatic Optimum (MCO, the last significant warming event), extended throughout ca. 18 and 15 Ma. It was later followed by the Miocene Climatic Transition (MCT), a rapid cooling event between 15 and 13 Ma. Those climatic events inflicted dramatic changes in the distribution of faunal and floral assemblages (Zachos et al. 2001, 2008; Böhme 2003; Kováčová et al. 2011; Kopecká et al. 2022; Scheiner et al. 2023; Vernyhorova et al. 2023). This applies to thermophilic ectothermic vertebrates in particular, crocodylians among them.
In the Early and Middle Miocene, central Europe was home to two different genera of Crocodylia: Gavialosuchus (often referred to as the extant genus Tomistoma) and Diplocynodon. Gavialosuchus was a marine long-snouted tomistomine gavialoid, with members of this group usually having a total body length over 6 m (Antunes 2017). The debate over which genus central European Middle Miocene tomistomines should belong to (either Gavialosuchus or Tomistoma) and their relationships are still unresolved and for the purpose of our study we prefer to use Gavialosuchus, following the original description by Toula and Kail (1885). For more details about the Gavialosuchus or Tomistoma debate see e.g., Nicholl et al. (2020) or Burke et al. (2024). The other crocodylian present during the same time and region, Diplocynodon, was a basal endemic alligatoroid, predominantly freshwater, with a total body length between 1.5 and 3 m (Brochu 1999; Massonne and Böhme 2022; Venczel 2023).
The aim of this paper is a reappraisal of the crocodylian material from the Early-to-Middle Miocene of the Fore-Carpathian Basin and its foreland. The studied material includes crocodylian teeth from the lower Badenian limestones of Pińczów (Poland) and Židlochovice (Czechia). The latter represents an unpublished historical find from the Moravian part of Fore-Carpathian Basin. Additionally, we also describe a newly discovered crocodylian specimen from the Szczerców field of the Bełchatów mine (Poland), representing the world’s northernmost record of crocodylians in the Neogene.
Institutional abbreviations.—KME, Krahuletz Museum, Eggenburg, Austria; LNEG, Museu Geológico, Lisbon, Portugal; MDLCA, Museo sardo di Geologia e Paleontologia “Domenico Lovisato”, University of Cagliari, Italy; MGGC, Collezione di Geologia “Museo Giovanni Capellini”, University of Bologna, Italy; MNHN, Muséum national d’Histoire naturelle, Paris, France; MWGUW, Museum of the Faculty of Geology, University of Warsaw, Poland (Stanisław Józef Thugutt Geological Museum); MZ, Polish Academy of Sciences, Museum of the Earth, Warsaw, Poland; NHMUK, Natural History Museum, London, UK; NMNH, National Museum of Natural History, Mdina, Malta; NMP, National Museum, Prague, Czechia; ÚGV, Department of Geological Sciences, Faculty of Science, Masaryk University, Brno, Czechia.
Other abbreviations.—MCO, Miocene Climatic Optimum; MCT, Miocene Climatic Transition.
Palaeogeography of the Central Paratethys
The Paratethys was a widespread intercontinental bioprovince (Popov et al. 2004; Harzhauser and Piller 2007) situated along the northern margins of the vanishing Tethys Ocean, which eventually transformed into the Mediterranean realm in its western region. Comprising a system of connected basins, the Paratethys stretched for over 4000 kilometres from the North Alpine Foreland Basin to the Aral Sea.
The whole system developed during the Oligocene (Popov et al. 2004; Harzhauser and Piller 2007) and its palaeogeographic pattern profoundly and repeatedly transformed up until the Pliocene. Tectonic events that formed the Alpid orogenic belt caused significant changes in the local and regional palaeogeography. This led not only to the modelling of the individual basins but also to the opening and/or closing of interbasinal connections, shaping the temporary pattern of the Paratethyan system.
The general division of the Paratethys comprises three parts (Fig. 1): Western (Rhône Basin and North Alpine Foreland Basin), Central (Fore-Carpathian Basin with intramontane and back-arc basins: Pannonian, Vienna, etc.), and Eastern, the largest, reaching from the Black Sea (temporally even from Eastern Carpathians) to the Aral Sea (e.g., Popov et al. 2004).
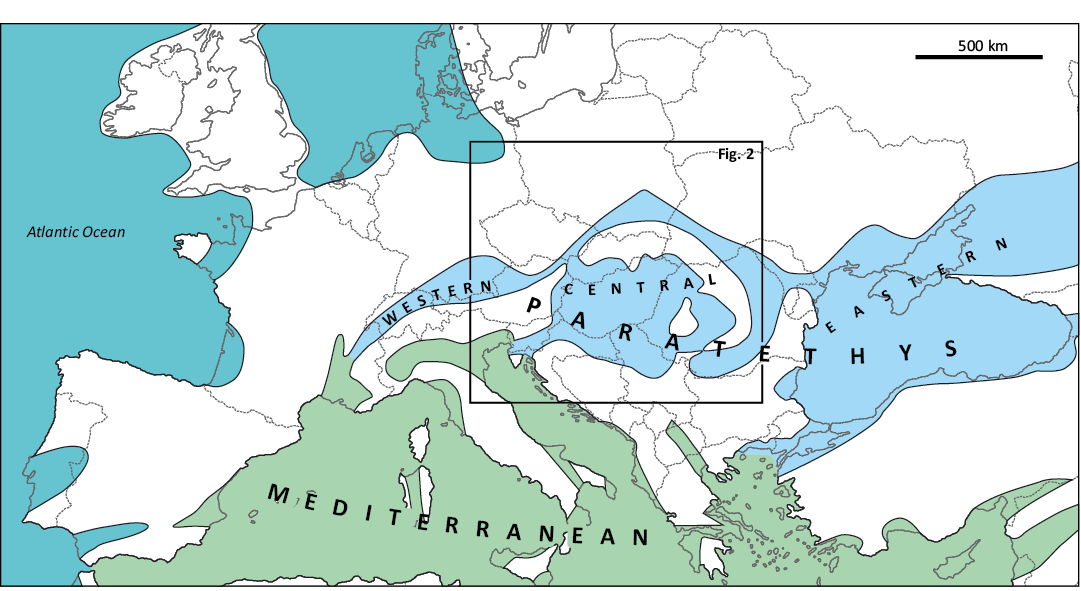
Fig. 1. Generalized palaeogeography of Europe during the Miocene epoch (after Rögl and Steininger 1983). The extent of Paratethyan and Mediterranean realms marked with light-blue and light-green, respectively.
The complex palaeogeographic evolution and large dimensions of these Paratethyan realms resulted in separate regional stratigraphic divisions for each of them (Andreyeva-Grigorovich et al. 1997; Piller et al. 2007; Kováč et al. 2017). The correlation of these divisions with the standard stratigraphic division is locally uncertain but becomes more precise over time (Kováč et al. 2018). Moreover, the opening and closure of interbasin connections caused particular basins (e.g., Fore-Carpathian Basin) to be included into different realms of the Paratethys during its existence (Studencka et al. 1998; Studencka and Jasionowski 2011; Wysocka et al. 2016; Kováč et al. 2018).
During the Early Miocene and until the end of the Badenian (Middle Miocene, approximately mid-Serravallian) the Central Paratethys consisted of two main parts divided by the uplifted mountain range of the Carpathians, the northern fore-arc basin (Fore-Carpathian Basin) and the southern system of back-arc basins (see Figs. 1 and 2). The latter was dominated by the complex Pannonian Basin, which included some smaller sub-basins and was connected with some smaller neighbouring depressions such as the Transylvanian and Vienna basins (Kováč et al. 2007, 2017).
The end of the Early Miocene marked the beginning of a period of increased global temperature that lasted up almost until the end of the Middle Miocene, with its peak during the Langhian age (approx. early Badenian in regional Central Paratethys division), i.e., corresponding to global Miocene Climatic Optimum (Itoigawa 1989, Müller 1996; Piller and Harzhauser 2005; Harzhauser et al. 2007).
The rise of temperature generated by the widely open connections with the Indo-Pacific bioprovince resulted in the Central Paratethys developing a very rich and diverse fauna. In the Fore-Carpathian Basin, a marine fauna comprising taxa indicative of subtropical waters was documented within lower Badenian deposits (Bałuk and Radwański 1977; Radwański 1977, 1985, 2015; Doláková et al. 2008, 2014; Wysocka et al. 2016). Only rare findings of reptiles were reported from the Fore-Carpathian Basin (Roemer 1870; Radwański 1974, 1977, 1985, 2015). This warm period (MCO), interrupted by a relatively short cooling event (MCT; see Hohenegger et al. 2009; Wysocka et al. 2016), lasted almost until the end of the Badenian. After the MCT, warm-temperate invertebrate marine fauna re-appeared (Müller 1996; Riegl and Piller 2000; Radwański et al. 2006; Górka et al. 2012; Hyžný et al. 2012; Pivko et al. 2017; Górka 2018a, b).
The periods of warm and humid climate of the Miocene resulted not only in the development of specific faunules and deposits within the marine basins of Central Paratethys. It also facilitated the periodic development of rich vegetation in the foreland of the Fore-Carpathian Basin. That enabled the formation of coal (lignite) seams, the thickness of which increases significantly in local depressions, often of tectonic origin, along the Ohře/Eger Graben in Czechia, e.g., the Most and Zittau Basins, extending into Poland (Rajchl et al. 2009; Brachaniec et al. 2022) or Bełchatów mines in Kleszczów Graben (Szynkiewicz 2000; Wagner and Matl 2007). Noteworthy, the northern foreland realm has been almost completely separated from the Fore-Carpathian Basin by the Bohemian Massif and the Central Polish Uplands. Between these highland areas a narrow ephemeric passage has occurred sporadically, connecting the northern foreland and the marine basin of the Fore-Carpathian Basin (see Fig. 2).
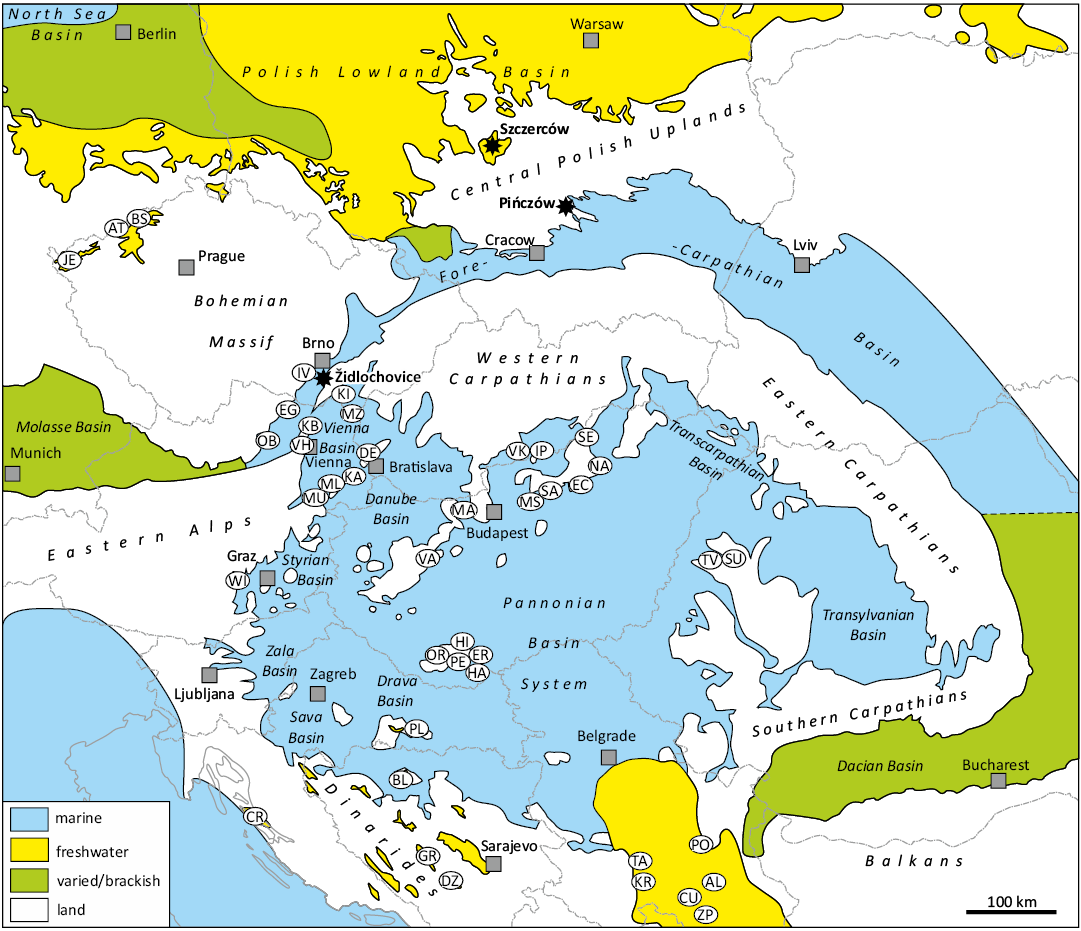
Fig. 2. Prevailing environments of the main palaeogeographic units of the Central Paratethys and adjoining areas in the Early–Middle Miocene time. Crocodylian-bearing sites marked with abbreviations, location of the discussed findings marked with asterisks. Abbreviations: AL, Aleksinac, Serbia; AT, Ahníkov and Tušimice, Czechia; BL, Banja Luka, Bosnia and Herzegovina; BS, Břešťany and Skyřice, Czechia; CR, Crnika beach, Croatia; CU, Čučale, Serbia; DE, Devínska Nová Ves, Slovakia; DZ, Džepi, Bosnia and Herzegovina; EC, Egercsehi, Hungary; EG, Eggenburg, Austria; ER, Erdősmecske, Hungary; GR, Gračanica, Bosnia and Herzegovina; HA, Himesháza, Hungary; HI, Hidas, Hungary; IP, Ipolytarnóc, Hungary; IV, Ivančice, Czechia; JE, Jehličná near Sokolov, Czechia; KA, Kaisersteinbruch, Austria; KB, Korneuburg Basin, Austria; KI, Kienberg, Czechia; KR, Kraljevo, Serbia; MA, Máriahalom, Hungary; ML, Mannersdorf am Leithagebirge, Austria; MS, Mátraszőlős, Hungary; MU, Müllendorf, Austria; MZ, Maustrenk, Austria; NA, Nagyvisnyó, Hungary; OB, Obritzberg, Austria; OR, Orfű-Bános, Hungary; PE, Pécs-Danitzpuszta and Pécsvárad, Hungary; PL, Poljanska, Croatia; PO, Popovac, Serbia; SA, Sámsonháza, Hungary; SE, Serényfalva, Hungary; SU, Subpiatră, Romania; TA, Tavnik, Serbia; TV, Vârciorog and Tăşad, Romania; VA, Várpalota, Hungary; VH, Vienna, Austria; VK, Veľký Krtíš, Slovakia; WI, Wies, Austria; ZP, Žitni Potok, Serbia. Compiled after: Rzehak (1912); Kordos and Solt (1984); Młynarski (1984); Zapfe (1984, 1989); Kordos (1985); Böhme and Ilg (2003); Stefanović and Mijovic (2004); Bulić and Jurišić-Polšak (2009); Jipa and Olariu (2009); Alaburić and Marković (2010); Kováčová et al. (2011); Krstić et al. (2012) ; Gibbard and Lewin (2016); Hír et al. (2016, 2024); Neubauer et al. (2016); Wysocka et al. (2016); Sant et al. (2018); Luján et al. (2019); Vasilyan (2020); Chroust et al. (2021); Botfalvai et al. (2023); Hajek-Tadesse et al. (2023); Weydt et al. (2023); Březina et al. (2024); Sebe et al. (2024); and Harzhauser et al. (2025).
Intense mining of coal deposits gave way for unearthing vertebrate remains. They are quite common in coal basins in the Lusatia region of SE Germany (Leder 2012) and NW Bohemia (Czechia), mainly in the Most Basin (Dvořák et al. 2010; Ekrt et al. 2016; Chroust et al. 2021, 2023). To the contrary, vertebrate remains are very uncommon in the Miocene coal deposits of Poland, where just two sites yielding rather scarce vertebrate material have been discovered so far. One of them is the Bełchatów mine, where remains of fishes and small mammals (mainly rodents) were recorded (for details see Kowalski and Rzebik-Kowalska 2002; Fostowicz-Frelik et al. 2012; Kovalchuk et al. 2020), accompanied by rare findings of proboscideans, artiodactyls, perissodactyls, and carnivores (Kowalski and Kubiak 1993; Van der Made and Kowalski 1996; Nadachowski 2001; Kowalski and Rzebik-Kowalska 2002; Marciszak and Lipecki 2020). The second of the Polish sites, the Turów mine, yielded just one single shell fragment of Testudinoidea indet. (Brachaniec et al. 2022).
A review of Miocene crocodylians from Central Paratethys
Fossil remains of crocodylians of the Miocene age are numerous in Europe, but usually come from the Western Paratethys. This is the case of, for example, fossils from the North Alpine Foreland Basin in Bavaria that are attributed to Diplocynodon sp. and dated as Burdigalian (Böhme 2003, 2010). From the same basin, a partial maxilla originally described as Gavialosuchus cf. gaudensis (Rossmann et al. 1999), currently interpreted as Tomistominae indet. (Nicholl et al. 2020), is known from the Bodensee area with other putative mentions from southern Germany (Sach 2016). The same applies to the more northerly situated areas in Germany (e.g., Mainzer Becken or Hambach mine in North Rhine-Westphalia), where findings of Diplocynodon and/or other (often unidentified) crocodylians are abundant (Ludwig 1877; Schleich 1988; Mörs et al. 2000; Böhme and Ilg 2003; Macaluso et al. 2022).
On the contrary, the crocodylian fossil record from the Central Paratethys is scarcer. In intramontane and back-arc basins of the Central Paratethys, crocodylian remains described as Crocodylia indet. (cf. Diplocynodon sp.) were found in the fossil site of Máriahalom (Mány-Zsambék Basin) in Hungary, the age of which, long considered as Chattian (late Oligocene), has been recently identified as presumably Aquitanian (Kocsis et al. 2023). Slightly younger findings of Burdigalian age are known from the Ipolytarnóc area (remains identified as Gavialis sp. and crocodylian tracks) and Serényfalva (identified as Crocodilus indet.) in northern Hungary (Kordos and Solt 1984; Kordos et al. 2021). Younger remains (Ottnangian; see Fig. 4) are known from Baňa Dolina Mine in Veľký Krtíš in Slovakia; teeth and an osteoderm from this locality were described by Čerňanský et al. (2012) as Crocodila indet.; in the nearby site of Egercsehi in northern Hungary, remains described as Crocodylus indet. of the same age were found (Kordos and Solt 1984). In the locality of Erdősmecske, near the Mecsek Mountains in southern Hungary, remains identified as belonging to the genus Crocodilus were recorded (Kordos and Solt 1984; Kordos 1985); however, their age is uncertain, ranging from Ottnangian to Karpatian (see Fig. 4).
Isolated teeth and one angular bone are described from the Korneuburg Basin in Austria and dated as Karpatian (Böhme 2002). Of Karpatian–Badenian age are findings of Diplocynodon sp. from Orfű-Bános near the Mecsek Mountains (Kordos and Solt 1984; Sebe et al. 2024). Zapfe (1984) reported teeth of Crocodylus sp. and Diplocynodon sp. from the upper lower Badenian of Kaisersteinbruch, Mannersdorf am Leithagebirge, and Müllendorf (Austria) in the southern part of the Vienna Basin. Of the same age is the tooth of Crocodylus sp. from Maustrenk near Zisterdorf, Austria, in the northern part of the Vienna Basin (Zapfe 1984). The tooth of Crocodylia indet. from Kienberg near Mikulov (Czechia) in the Moravian part of Vienna Basin (Březina et al. 2024) is of the latest early to earliest late Badenian age. Diplocynodon ungeri dated to the MN5 mammal unit (Badenian), is known from the Styrian Basin near Wies, Austria (Martin and Gross 2011). The sites of Nagyvisnyó (lower Badenian; Kordos and Solt 1984) and Sámsonháza (middle Badenian; Hír et al. 2016) are situated in northern Hungary, both of which yield crocodylian remains. Of late Badenian age are the localities of Mátraszőlős in northern Hungary (Kordos and Solt 1984), Várpalota near Székesfehérvár (Kordos and Solt 1984; Kordos 1985), and Hidas near the Mecsek Mountains, all of which yield Diplocynodon sp. (Kordos 1985; Hír et al. 2024). The western Romanian site of Subpiatră 2/1 yields Diplocynodon sp. of middle-to-late Badenian age (Venczel 2007; Hír et al. 2016). Isolated teeth tentatively ascribed to Gavialosuchus sp. are also known from the upper Badenian of Devínska Nová Ves in Slovakia (Schlögl and Holec 2004).
Along the southern margin of the Central Paratethys, crocodylian remains are known from the Lower and Middle Miocene of Serbia, predominantly from lacustrine deposits in the south of the country. Remains of Crocodylia indet. are known from the Lower Miocene of Aleksinac and Kraljevo (Jovanović et al. 2002; Vasilyan 2020), while Crocodilus sp. is known from the upper Lower Miocene of Čučale (Stefanović and Mijovic 2004). Other crocodylian-bearing localities are the Middle Miocene site of Žitni Potok (Jovanović et al. 2002; Vasilyan 2020), the middle Badenian site of Tavnik (Krstić et al. 2012), and the famous lower Badenian site at Popovac yielding Gavialosuchus eggenburgensis, Crocodylus moraviensis, and Alligatorinae indet. (Jovanović et al. 2002; Alaburić and Marković 2010; Sant et al. 2018; Vasilyan 2020).
In the south-western margins of the Central Parathethys, the Early Miocene crocodylians are known from Crnika beach on the Croatian island of Pag (Bulić and Jurišić-Polšak 2009) and from Poljanska in the North Croatian Basin (Hajek-Tadesse et al. 2023). From Bosnia and Herzegovina three occurrences are known: Banja Luka of Early-to-Middle Miocene age, the early Badenian Bugojno palaeolake in Gračanica with presumed Diplocynodon sp. (Vasilyan 2020), and the site of Džepi in the Prozor Basin also of early Badenian age (Neubauer et al. 2016).
The youngest certain occurrences of crocodylians in the coeval deposits of the intramontane and back-arc basins of the Central Paratethys are findings of Diplocynodon sp. are of the Sarmatian age. The findings of Diplocynodon sp. from the early Sarmatian are known from the Tăşad, Vârciorog, and Subpiatră 2/2 sites in Romania (Venczel 2007; Hír et al. 2016) while the most recent report of the mid-Sarmatian osteoderm of Crocodylia indet. (? Diplocynodon sp.) from Hernals district of Vienna appears to be the latest known occurrence of the crocodylian in Central Paratethys (Harzhauser et al. 2025).
Even younger known crocodylian-bearing deposits in the Central Paratethys seem to be sands and gravels of the Pannonian Kálla Member exposed in the Pécs-Danitzpuszta, Pécsvárad, and Himesháza sand-pits in eastern Mecsek Mountains, but remains of Diplocynodon sp. in this site were most probably re-deposited from older Miocene deposits as a result of erosion following the Late Miocene uplift of the Mecsek Mountains (Botfalvai et al. 2023; Sebe et al. 2024). The putative younger occurrence of cf. Diplocynodon sp. from the lower Pontian of Götzendorf an der Leitha in Austria (Zapfe 1989) remains unconfirmed (Harzhauser et al. 2025).
Crocodylians are even scarcer in the Fore-Carpathian Basin, however, some particularly interesting findings have been recorded from this region. The Lower Miocene site of Eggenburg in Austria yielded the holotype skull of Gavialosuchus eggenburgensis Toula & Kail, 1885. Another occurrence of a crocodylian is known from the Ottnangian, nearby Ivančice, in Czechia, where remains of the Diplocynodon sp., were found (Rzehak 1912). The Polish crocodylian fossil record in the Fore-Carpathian Basin has been so far limited only to a single tooth attributed to Tomistoma sp. from Pińczów (Radwański 1974, 1977, 1985, 2015; Antunes in Młynarski 1984).
The known record of crocodylian remains from the more distant foreland of Central Paratethys is limited practically to its western region, the NW part of the Bohemian Massif. In this area, Diplocynodon ratelii Pomel, 1847, is known from the Burdigalian (MN3 Mammal Zone) of Most Basin (Luján et al. 2019). The older records from this area come from the vicinity of Most—for example, from Břešťany (Liebus 1936) or Skyřice (Laube 1910; Luján et al. 2019), whereas modern findings were reported from Ahníkov (Chroust et al. 2021) near Kadaň. Further to the south, crocodylian remains were reported from Jehličná of the Sokolov Basin (Redlich 1902; Luján et al. 2019).
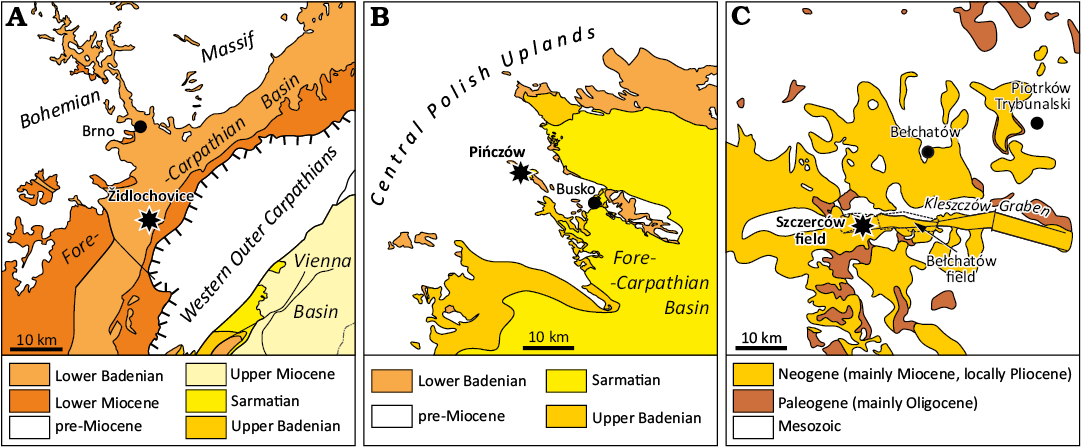
Fig. 3. Simplified geologic maps of the discussed localities. A. Židlochovice. B. Pińczów. C. Szczerców. Compiled after Buday (1963), Danel and Gałązka (2007), Haisig (2011), and Górka (2019).
Geological setting
Židlochovice.—This locality is situated in the southwestern part of Fore-Carpathian Basin (Figs. 2 and 3A) in close proximity to the Western Outer Carpathian nappe. This locality is represented by lower Badenian clays, sands and limestones, which build the northwestern slope of Výhon Hill. It is one of the richest palaeontological localities in Moravia and has consequently attracted palaeontologists and geologists since the 19th century (e.g., Hörnes and Reuss 1870; Rzehak 1880; Procházka 1893). An illustrative section (up to 30 m thick) was exposed by a large former brickyard and became the faciostratotype of the lower Badenian (i.e., Moravian, Upper Lagenidae Zone; see Cicha 1978). This fossil site was later comprehensively studied with the support of data from two boreholes (altogether 21 m thick section) and the section was biostratigraphically correlated with the NN5 Zone, 14.9 to 13.9 Ma (Doláková et al. 2014; see Fig. 4).
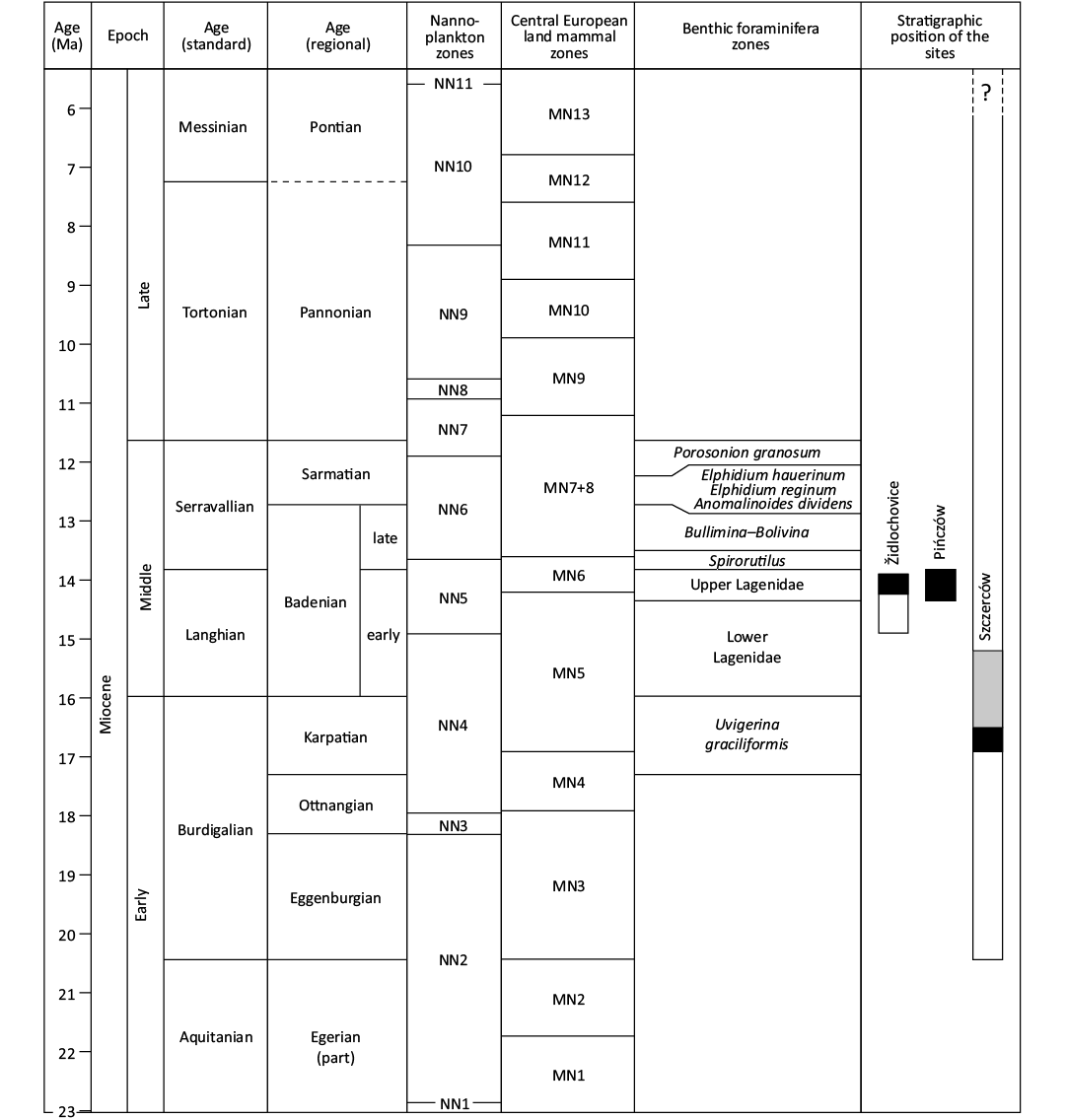
Fig. 4. Stratigraphic chart of the Miocene of the Central Paratethys. The stratigraphic position of the sites discussed in the text is indicated on the right-hand side. Black bands denote the most certain age of the material described in the present study; grey band denotes possible age; white bands correspond to the total stratigraphic range recorded in the sites. Compiled after: Harzhauser and Piller 2004; Kováč et al. 2005, 2007, 2018; Piller et al. 2007; Hohenegger et al. 2009, 2014; Pezelj et al. 2013; Doláková et al. 2014; Neubauer et al. 2015; Luján et al. 2019; Březina et al. 2021; and Ivanov 2022.
The lower Badenian sedimentary sequence of Židlochovice is characteristic by the dominance of pelitic sedimentation of unlaminated calcareous clays (“tegel”), which in the upper part alternate with calcareous sands to sandstones and limestones with dominance of red algae and bryozoans (Cicha 1978; Doláková et al. 2014). The rich flora of red algae and nanoplankton and the rich fauna of foraminifera, radiolarians, bryozoans, ostracods, mollusks, sea urchins, corals, bony fish, sharks and cetaceans indicate a marine, warm to subtropical palaeoenvironment shallowing-upwards from epibathyal/circalittoral (up to 250 m) to shallow (up to 20 m) infralittoral (Brzobohatý 1997; Seko et al. 2012; Doláková et al. 2014).
The original position of the tooth in the section is unknown, however residual algal limestone at the base of the crown of the studied crocodylian tooth suggests that it comes from the upper parts of the section, dominated by red-algal limestones. The limestones indicate shallow marine environment with seagrass meadows representing a shallowing and slight cooling event coeval with the regressive phase of TB 2.4 cycle, dated at 14.2 to 13.6 Ma (Doláková et al. 2014; see also Fig. 4).
Pińczów.—The area is located in the northern marginal zone of the Fore-Carpathian Basin, and the fossiliferous site is situated in one of the quarries (Quarry no. 2; Młynarski 1984) located in the northernmost part of the town of Pińczów (Figs. 2 and 3B), formerly called Nowa Wieś. This quarry has been the source of stone since the Middle Ages but finally became abandoned in the early 2000s.
The whole section is dominated by red-algal (traditionally referred to as “lithothamnian”) organodetrital limestones, so-called “Pińczów limestones”, lithologically similar, e.g., to the Badenian “Leithakalk” in the Vienna Basin (Studencki 1979). The lower part of the section is composed of very-thick bedded, fine-grained limestones passing into coarser variety in the upper part. Frequent accumulations of rhodoliths and 1–2 local horizons of algal-bryozoan marls also occur. The thickness of the exposed section is approximately 15 m (Drewniak 1994; Studencki 1988a) but the total thickness of the Pińczów limestones in the area reaches up to 25 metres (Studencki 1988a).
The site has been known for years for its ubiquitous and rich fossils. The faunal assemblage comprises numerous invertebrate remains: corals, polychaetes, brachiopods, mollusks, crustaceans, and echinoderms (Kowalewski 1930, 1958; Radwański 1969, 1977, 2015; Förster 1979, Studencka and Studencki 1988; Studencki 1988b; Mączyńska 1993). Other remarkable findings from Pińczów limestones include vertebrate remains, such as those of chondrichthyans and teleost fish (Kowalewski 1930; Jerzmańska 1958; Pawłowska 1960; Radwański 1965), as well as marine mammals, whales (Czyżewska and Ryziewicz 1976; Czyżewska 1988; Czyżewska and Radwański 1991) and sirenians (Radwański 2015). The only reptile representative is a single crocodylian tooth reported by Radwański (1974, 1977, 1985, 2015) and assigned to the genus Tomistoma (Antunes in Młynarski 1984). However, aside from a very basic description by Antunes in Młynarski (1984), this tooth has never been fully described in detail nor illustrated.
The composition of the marine fauna indicates a warm, subtropical shallow marine environment of oceanic salinity (Pawłowska 1960; Radwański 1965, 1977, 1985, 2015; Bałuk and Radwański 1977; Czyżewska 1988). The age of the Pińczów limestones and coeval deposits in the Polish part of Fore-Carpathian Basin has been considered for decades as early Tortonian (e.g., Kowalewski 1930, 1958; Czarnocki 1933; Radwański 1969). In the early 1970s it was confirmed that the exact age of the Pińczów limestones is early Badenian (see Fig. 4; Radwański 1973; Martini 1977; Studencki 1979; Górka 2019).
Further studies provided a more detailed stratigraphic position of the Pińczów limestones, they belong to the Upper Lagenidae Zone, i.e., the upper part of the lower Badenian (Studencki 1988a; Studencka 1999; Peryt 2013), and to the NN 5 nannoplankton zone (Martini 1977).
The long-lasting confusion regarding the determination of the age of the discussed deposits resulted in numerous inaccuracies. One of such examples is the case of the crocodylian tooth mentioned above, the age of which has been incorrectly attributed to as “Upper Miocene–Lower Tortonian” by Młynarski (1984). The latter author cited the outdated paper of Radwański (1965) literally, instead of applying the updated age affinity, i.e., early Badenian of the Middle Miocene.
Szczerców.—The site is located in the western mining field (i.e., Szczerców field) of the large lignite mine of Bełchatów, situated in central Poland (Figs. 2 and 3C). The Bełchatów mine opened in the 1970s and since then the thick succession of Miocene deposits including lignite seams has been intensively mined. The larger eastern field (Bełchatów field) is currently being filled with dump deposits, but through the last half of the century its deposits have been intensively palaeontologically studied (for a list of papers see Fostowicz-Frelik et al. 2012; Worobiec and Worobiec 2019; Kovalchuk et al. 2020). To the contrary, the Szczerców field has been operative for a shorter period of time (it has been operating since 1998 and the mining of lignite started in 2009), but its Miocene deposits have not been so intensively studied palaeontologically as the Bełchatów field. Because there is a large similarity in the depositional history between these two fields and the Szczerców field has not been studied as extensively, the geological features of the Bełchatów field could be treated as general benchmarks (see: Wagner and Matl 2007).
The lignite deposits of the Bełchatów mine belong to a thick Miocene succession reaching up to 600 m in thickness that was deposited in a ~30 km-long section of Kleszczów Graben, which in turn is ca. 80 km long and up to 3 km wide (Gotowała and Hałuszczak 2002). The Miocene succession lies on Upper Jurassic, Upper Cretaceous, and, locally, Paleogene rocks (Bielowicz and Morga 2021).
The Neogene succession, best recognised in the Bełchatów field, is traditionally divided in four complexes: (i) subcoal, (ii) coal, (iii) clayey-coal, and (iv) clayey-sandy.
The lowermost (i.e., subcoal) complex lies directly on an older substrate and comprises sands with clay and mud intercalations with thin layers of lignite. The overall thickness of the subcoal complex exceeds 100 m and its age is referred to as Eggenburgian (Szynkiewicz 2000; Worobiec and Worobiec 2022).
The coal complex, with a total thickness exceeding 100 m, is divided into two subcomplexes: non-productive and productive (Pawelec and Bielowicz 2016). The former comprises sands with thin lignite seams. The productive subcomplex is dominated by thick lignite seams, where the thickest (the main seam, D) reaches 40–60 m (Szynkiewicz 2000). In the upper part of the main seam a thick layer of lacustrine chalk (lacustrine limestone) occurs. The rich vertebrate faunal assemblage (referred to as the Beł-C; Stuchlik et al. 1990; Szynkiewicz 2000) was collected there and determined to belong to the MN 4 Mammal Zone (Kowalski 1993b, 1994; Kovalchuk et al. 2020). Above the main seam there are thinner seams, C (the older) and B (the younger, divided into B1 and B2). Below the seam C a thin tuffite layer (so called paratonstein) of FT (i.e., fissure track) age 18.1 ± 1.7 Ma occurs (Burchart et al. 1988). The age of the coal complex is defined as Ottnangian to Karpatian (Worobiec and Worobiec 2022).
Above the seam B, in the topmost part of the coal complex, there is a thick layer of lacustrine chalk where vertebrate fauna was recorded. This faunal assemblage (Beł-B) represents the MN 5 Mammal Zone (Kowalski 1993b; Kovalchuk et al. 2020).
The coal complex is separated from the clayey-coal complex by the distinct erosional surface (Stuchlik et al. 1990; Szynkiewicz 2000). The clayey-coal complex comprises grey-to-greenish clays with thin lignite intercalations, and layers of muds, sand and even gravels. In its lower part a relatively thick lignite seam (seam A) occurs, with tuffite (paratonstein) layer in its top. The fission-track age of paratonstein is 16.5 ± 1.3 Ma (Burchart et al. 1988). In the upper part of the complex some thin lignite seams and layers of lacustrine chalk were recorded. The occurring vertebrates (Beł-A) indicate an MN 9 Mammal Zone (Kowalski 1993a). The age of the clayey-coal complex ranges from late Karpatian to Sarmatian, presumably even Pannonian (Szynkiewicz 2000; Worobiec et al. 2012).
The youngest complex, i.e., clayey-sandy, contains whitish and grey gravels, sands and greenish clays with a total thickness reaching up to a few dozens of metres and with its erosional boundary at the top (Szynkiewicz 2000). The age of that complex is referred to as Pannonian to ?Pliocene (Worobiec et al. 2012).
The succession in the Szczerców field shows a high level of similarity to the Bełchatów field, however there are some significant differences, as presented below.
The characteristics of the subcoal complex corresponds to those of the Bełchatów field—it is composed of sands with two thin layers of lignite and reaches up to 100 m. The age of the complex, by analogy to the Bełchatów field, is presumably Eggenburgian (Wagner and Matl 2007; Pawelec and Bielowicz 2016).
The coal complex is composed of two subcomplexes. The lower, non-productive subcomplex reaches up to 100 m and is composed of fine-grained carbonaceous sands and thin lignite seams. The productive subcomplex is built of one to two thick lignite seams and its thickness ranges from a few metres in the western part up to 140 m in the east (Wagner and Matl 2007; Pawelec and Bielowicz 2016). This subcomplex is more compact when compared to the Bełchatów field and seams B and C known from the Bełchatów field do not appear to have comparable seams in the Szczerców field (see Wagner and Matl 2007: fig. 1). However, the age of the Szczerców coal complex is Ottnagian to Karpatian similarly to Bełchatów field (Wagner and Matl 2007).
Clayey-coal complex is built of clays (locally carbonaceous) with sandy intercalations and layers of xylite coals correlated with lignite seam A known from the Bełchatów field (Wagner and Matl 2007). The thickness of that complex reaches up to 90 m (Pawelec and Bielowicz 2016) and its age by lithostratigraphic analogy to the Bełchatów field should be referred to as Karpatian to Sarmatian, presumably even Pannonian (Wagner and Matl 2007).
The Szczerców clay-sand complex is represented by greenish clays, silts and its maximum thickness reaches 80 m (Pawelec and Bielowicz 2016). The supposed age of the complex is Pannonian to ?Pliocene.
The main difference between the two mining fields is the high percentage of lacustrine chalk deposits in the Szczerców field compared to Bełchatów. The thickness of lacustrine chalk lithosomes in Szczerców reaches up to 90 m and two main levels were recognised there (Wagner and Matl 2007). The older level (II) occurs in the western part of the field in the upper part of the subcoal complex and lower part of the coal complex. It represents MN 4 Mammal Zone (ca. late Ottnangian to early Karpatian). The younger level (I) occupies the upper part of the coal complex and the lower part of the clayey-coal complex. Its age is determined as MN 5–6 mammal zones (late Karpatian to early Badenian). Locally, an additional level (Ia) occurs, belonging to MN 7–8 mammal zones, mainly of Sarmatian age (Wagner and Matl 2007).
The particular lacustrine chalk bed, bearing a crocodylian osteoderm, occurred in the uppermost part of the coal complex, thus it represents the younger level of lacustrine chalk (I). We found there only scarce plant material of Magnolia cf. burseracea (Menzel, 1913) Mai, 1975, Umbelliferopsis molassicus Gregor, 1982, and unidentifiable wood fragments that are not sufficiently informative for environmental reconstruction. However, more information was obtained from the lacustrine chalk from the lower part of the clayey-coal complex, which is slightly younger than the investigated crocodylian specimen. Aside from small fragments of wood and leaves (presumably of Cyperaceae), a relatively rich assemblage of carpological remains was found (ca. 300 specimens representing 14 taxa stored at the Polish Academy of Sciences, Museum of the Earth in Warsaw, Poland). All identified taxa are representatives of different types of wetlands vegetation. The only tree typical of the Cenozoic swamp forests that was recorded is Nyssa disseminata (Ludwig, 1857) Kirchheimer, 1937 (Mai 2000). Remaining woody taxa can be assigned to swamp forests understorey or shrub bogs including Frangula solitaria Gregor, 1977, Lyonia danica Friis, 1985, Myrica ceriferiformoides Bůžek & Holý, 1964, and M. burseracea (Mai 2000; Kowalski 2017). A few herbaceous plants typical of reeds have also been identified, including Cladium oligovasculare Mai in Knobloch, 1978, Dulichium vespiforme C. Reid & E.M. Reid, 1908, Carex sp., Umbelliferopsis molassicus (see Gregor 1982; Mai 2000). Another herbaceous plant from Szczerców, Decodon gibbosus (E.M. Reid, 1920) Nikitin, 1929, could occur in a wide range of non-arborescent types of wetlands (Kvaček and Sakala 1999). Macrophytes are represented only by Potamogeton sp.; this floating leaved genus occurs today in various aquatic habitats (Cook et al. 1974), but in the clayey-coal complex in Szczerców it most likely indicates the presence of open (full-sun), stagnant water (pond or lake) or a slow flowing river. The striking feature of this sample is the apparent lack of Taxodium and Glyptostrobus, two of the most characteristic European Cenozoic swamp forest trees (LePage 2007; Kunzmann et al. 2009), documented also from the neighbouring Bełchatów field (Stuchlik et al. 1990). This indicates that swamp forests may not have been fully developed in Szczerców in the studied time and area. Considering the lithological characteristics of the deposits, the presumed structure and height of vegetation may result from specific water chemistry. Comparable conditions can be observed in modern minerotrophic peatlands (Bedford and Godwin 2003; Mitsch and Gosselink 2015) often populated by narrowly adapted ecological specialists, calciphilic species in particular. However, most species identified are ecological generalists, typical of Neogene coal-producing plant communities. On the other hand, we cannot exclude that some (edaphic) specialists can hide among the few unidentified disseminules (e.g., Potamogeton sp.). Climatic conditions cannot be precisely estimated based on the few fossil taxa found in the samples, because of the very broad preferences of their closest living relatives.
According to Wagner and Matl (2007), who dated the different stratigraphic levels in Szczerców using freshwater malacofauna assemblages as the biostratigraphic indicators, the level that yields the crocodylian osteoderm belongs to MN 5–6 Mammal Zone (upper Karpatian–lower Badenian, i.e., upper Burdigalian–Langhian standard stages). However, because the fission-track age of paratonstein above the lignite seam A in the Bełchatów field is 16.5 ± 1.3 Ma, the age of the discussed lacustrine chalk should be also considered as Karpatian. It is younger than ca. 16.8 Ma (i.e. MN 4/MN 5 transition, see Fig. 4) but likely not younger than 16.5 Ma (black band in Fig. 4). Nevertheless, considering the confidence interval (± 1.3 Myr), it cannot be definitely excluded that the described crocodylian osteoderm may be significantly younger, even as young as 15.2 Ma, and therefore of early Badenian age (marked with a grey band in Fig. 4).
Material and methods
Palaeozoological material.—The studied material comprises two isolated teeth and one dorsal osteoderm.
The studied tooth from Židlochovice comes from the collection of Anthon Rzehak (1855–1923), but he never mentioned it in any of his works, despite correctly recognizing it as a crocodile tooth in his handwritten label. The specimen is housed in the Paleontological collection of the Department of Geological Sciences, Faculty of Science, Masaryk University, Brno under inventory number ÚGV E1093.
The tooth from Pińczów was collected by Prof. Andrzej Radwański (1934–2016) in the 1960s–1970s and is housed at the Museum of the Faculty of Geology, University of Warsaw (Stanisław Józef Thugutt Geological Museum) under inventory number MWGUW/ZI/113/067. The specimen was reported by Radwański (1974, 1977, 1985, 2015), and by Antunes in Młynarski (1984).
The single dorsal osteoderm from the Szczerców mining field was collected by our prospecting team (MG, RK, SL-T) in June 2023 during fieldwork in the Bełchatów lignite mine and is kept at the Museum of the Faculty of Geology, University of Warsaw under inventory number MWGUW/ZI/113/064. The coordinates for the point where the osteoderm was collected are 51°14’00.0”N, 19°10’07.6”E, 61.14 m a.s.l.
The specimens from Pińczów and Židlochovice were supposedly embedded in a limestone matrix, but the exact method of their extraction is unknown (they were most probably extracted mechanically). The specimen from Szczerców was embedded in a lacustrine chalk sample that was picked up from the surface. The osteoderm was partially broken because of the deformation of the surrounding soft matrix. After the sample was softened with water, the osteoderm was manually extracted and glued back.
The fossil material of crocodylians that was used for comparisons is presented in Table 1.
Table 1. The fossil material of crocodylians used for comparisons.
|
Species |
Number |
Material |
Age |
Locality |
Reference |
|
Diplocynodon ratelii |
MNHN Ar888 |
teeth |
Early Miocene |
Artenay, France |
unpublished |
|
Diplocynodon hantoniensis |
NHMUK OR 25166 |
teeth |
late Eocene |
Hordwell Cliff, UK |
Rio et al. 2020: fig. 13A–C |
|
Diplocynodon hantoniensis |
NHMUK OR 30317 |
tooth |
late Eocene |
Hordwell Cliff, UK |
Rio et al. 2020: fig. 13D |
|
Diplocynodon hantoniensis |
NHMUK OR 46434 |
osteoderm |
late Eocene |
Hordwell Cliff, UK |
Rio et al. 2020: fig. 20B |
|
Diplocynodon hantoniensis |
NHMUK R.5214 |
osteoderm |
late Eocene |
Hordwell Cliff, UK |
Rio et al. 2020: fig. 20C |
|
Diplocynodon cf. ratelii |
NMP Pv 11650–62 |
teeth |
Early Miocene |
Ahníkov, Czechia |
Chroust et al. 2021: fig. 6 |
|
Diplocynodon cf. ratelii |
NMP Pv 11663–11668 |
osteoderms |
Early Miocene |
Ahníkov, Czechia |
Chroust et al. 2021: fig. 7 |
|
Gavialosuchus eggenburgensis |
KME F/2368 |
tooth |
Early Miocene |
Eggenburg, Austria |
unpublished |
|
Gavialosuchus eggenburgensis |
KME F/3707 |
tooth |
Early Miocene |
Eggenburg, Austria |
unpublished |
|
Tomistoma calaritanum |
MDLCA 148 |
teeth |
Late Miocene |
Mirrionis (Cagliari), Sardinia, Italy |
Nicholl et al. 2020: fig. 5C |
|
Tomistoma lusitanica |
LNEG 4783 |
teeth |
Early Miocene |
Lisbon, Portugal |
unpublished |
|
Tomistoma lusitanica |
LNEG 4794 |
teeth |
Early Miocene |
Lisbon, Portugal |
unpublished |
|
Tomistoma lusitanica |
LNEG 8709.19 |
teeth |
Early Miocene |
Lisbon, Portugal |
unpublished |
|
Melitosaurus champsoides |
NHMUK PV OR41151 |
snout with teeth |
Early Miocene |
Gozo Island, Malta |
Nicholl et al. 2020: fig. 3 |
|
Tomistoma gaudense |
NMNH-T11228 |
teeth |
Early Miocene |
Gozo Island, Malta |
Nicholl et al. 2020: fig. 5B |
|
Tomistoma calaritanum |
MDLCA 14401 |
osteoderms |
Late Miocene |
Piazza d’Armi |
Zoboli et al. 2019: fig. 8 |
Palaeobotanical material.—For further examination two samples of lacustrine chalk were taken from Szczerców: (i) sample 1b-2023 from the topmost part of the coal complex where the crocodylian remains were found; (ii) sample 1b-2022 from lower part of the clayey-coal complex. The matrix of the samples was dissolved using 8% acetic acid. The residuum was washed on a sieve of a mesh 0.5 mm, then dried, and examined with the use of a stereomicroscope to extract fossil plant remains. The collection of fossil plants is stored at the Polish Academy of Sciences, Museum of the Earth in Warsaw, Poland, under inventory numbers MZ VII/160/5–7, 10, 12–22.
Systematic palaeontology
Order Crocodylia Gmelin, 1789
Crocodylia indet.
Fig. 5.
Material.—ÚGV E1093, tooth from Židlochovice, Czechia, lower Badenian. MWGUW/ZI/113/067, tooth from Pińczów, Poland, lower Badenian. MWGUW/ZI/113/064, osteoderm from Szczerców, Poland, Karpatian–?lower Badenian.
Description.—Teeth: ÚGV E1093 is conical, more slender and more lingually curved when compared to the specimen from Pińczów (MWGUW/ZI/113/067). It displays the crown portion, and the apicobasal length of the tooth is approximately 30 mm (Fig. 5A). However, the apical tip and the root are missing. The tooth is partially infilled by a matrix composed of bioclasts and sand grains. The enamel is generally smooth, containing several cracks, and similar in thickness to the Pińczów specimen. Both mesial and distal carinae are present, one of them is more distinct along the edge but disappears in the proximal part. The surface has faint and diffused longitudinal striations. Incremental bands are not as significant as in the Pińczów specimen. The tooth displays the general pattern of slender curved conical teeth, but it is impossible to determine its precise position in the jaw.
MWGUW/ZI/113/067 (Fig. 5B) is quite robust and displays only the crown portion. The apicobasal length of the tooth is approximately 25 mm. The base is partially covered with calcareous sediment. The enamel is smooth and quite thin. The tooth is conical, stout and slightly blunt. A small lingual curvature and both mesial and distal carinae are present. The tooth itself has faint longitudinal striations and is crossed by incremental bands of grey colour. The tooth does not display the general pattern of premaxillary teeth, which are slender, more conical, and curved. Also, the globular morphotype typical for the posterior dentition of alligatoroids is not displayed either.
Osteoderm: MWGUW/ZI/113/064, a single isolated dorsal osteoderm (Fig. 5C) is partially preserved and lacks the anterior articular facet, as well as the right part of the bone, located laterally to the medial keel. The external surface is covered by subcircular pits and holes, with some degree of merging between them. The pits extend right up to the left lateral and posterior edges. The medial longitudinal keel is present, but its sagittal apex is broken. From an internal view, the surface is smooth, with small foramina, which are accumulated in the posterior edge, where the posterior articular facet is present. In the left posterolateral aspect of the interior surface, close to the edge a pit is present caused by damage of unknown origin.

Fig. 5. The discussed remains of Crocodylia indet. from the Miocene of Fore-Carpathian Basin and its foreland. A. ÚGV E1093, an isolated tooth from the lower Badenian (Langhian) of Židlochovice, Czechia; lingual (A1), mesial/distal (A2), basal (A3), distal/mesial (A4), and labial (A5) views. B. MWGUW/ZI/113/067, an isolated tooth from the lower Badenian (Langhian) of Pińczów, Poland (previously referred to as Tomistoma sp.); labial (B1), mesial/distal (B2), basal (B3), distal/mesial (B4), and lingual (B5) views. C. MWGUW/ZI/113/064, a dorsal osteoderm from Karpatian–?lower Badenian (Burdigalian–?Langhian) of Szczerców mining field, Poland, the world’s northernmost recorded Neogene crocodilian; dorsal (C1) and ventral (C2) views.
Remarks.—Due to the scarcity of the preserved material (i.e., a single osteoderm and two isolated teeth), the possibility of identifying the material with precision is somewhat hindered. Detailed comparisons with remains of Diplocynodon, Gavialosuchus/Tomistoma, and Melitosaurus did not allow us to form a definitive opinion on the generic affiliation. Therefore, we deem the attribution to Crocodylia indet. as sufficient. During the Miocene, the Central Paratethys was home to only two crocodylian genera: Diplocynodon and Gavialosuchus. Whereas the dentition is generally very similar among Crocodylia, a few important differences have been observed. Adult Diplocynodon has much smaller teeth in overall size when compared to adult Gavialosuchus. However, the 4th and 5th maxillary and 3rd and 4th dentary teeth are enlarged in Diplocynodon, and the posterior 9th, 10th or 11th maxillary and 11th or 12th dentary teeth are large as well. In some specimens, teeth are quite large and robust and the crown itself can reach more than 30 mm in apicobasal length (Chroust et al. 2021; Massonne and Böhme 2022). The dental enamel in basal view seems to be generally thicker in Gavialosuchus or Tomistoma compared to Diplocynodon. However, in a closer look, enamel thickness is variable among the teeth. The smoothness of the enamel is typical for Diplocynodon, whereas teeth of Gavialosuchus or Tomistoma contain apicobasal ridges and therefore the surface is rough. When the enamel is worn, gavialoid teeth can resemble those of Diplocynodon (morphotype D in Venczel et al. 2023). Nevertheless, this character could be more tied to taphonomic conditions rather than phylogenetic relations (Christopher A. Brochu, personal communication 2024). The presence of longitudinal striation is more frequent in Diplocynodon than in Gavialosuchus or Tomistoma but this character can also be misleading. The described teeth follow the pattern observed in Diplocynodon; however, the sample does not include any globular teeth, which are present in Diplocynodon and absent in Gavialosuchus, and therefore of higher diagnostic value.
A puzzling issue is the former generic assignment of the tooth MWGUW/ZI/113/067 (Fig. 5B) to Tomistoma. The specimen was identified by Miguel Telles Antunes (see Antunes in Młynarski 1984), after a putative mail correspondence with Andrzej Radwański. In fact, his description was not supported by any image of the specimen and the main argument for its identification as Tomistoma was the size of the specimen (“very big [...] but not yet a gigantic one”; Antunes in Młynarski 1984: 134). Furthermore, Antunes in Młynarski (1984: 135) noted that the tooth was “more delicate [...] than those found so far among [...] this group of reptiles” and suggested that the Pińczów specimen may represent a distinct genus from Tomistoma or Gavialis. Herein, we suggest a series of potential reasons that can explain this confusion.
Firstly, the tooth was found in marine deposits and therefore its affinity to the Tomistoma, which inhabited marine environments, seemed reasonable. Secondly, the Pińczów fossil site was in the past referred to as Tortonian, i.e., Late Miocene in standard (“Mediterranean”) stratigraphy. In the Late Miocene only Tomistominae and true crocodiles (Crocodylidae) were present in Europe (Delfino and Rossi 2013; Nicholl et al. 2020; Delfino et al. 2021; Rio and Mannion 2021). Moreover, in 1984, the presence of the genus Crocodylus in Late Miocene deposits of Europe was still unconfirmed. Thirdly, and we assume this as the strongest argument, Antunes had most probably no opportunity to see the original specimen, only its photographs. Unfortunately, the photographs that were most presumably presented by Andrzej Radwański to Marian Młynarski and/or Miguel Telles Antunes displayed the specimen twice of its natural size and lacked a scale bar (we have recently gained access to the relevant photographs in Andrzej Radwański’s archives). Miguel Telles Antunes most probably assumed the scale of photographs to be 1:1 and thus estimated the length of the tooth at 50 mm. This would be consistent with the phrase “very big” and Antunes’ belief that the tooth belonged to Tomistoma, considering the fact that adult tomistomine teeth are much larger when compared to other crocodylians of similar age and geographic origin.
There are differences also in osteoderms between gavialoids and diplocynodontines. Firstly, osteoderms are much bigger and thicker in adult Gavialosuchus or Tomistoma when compared to adult Diplocynodon. Dorsal osteoderms of Diplocynodon always have a medial keel and in gavialoids is generally absent. A similar pattern was observed in the study of Tomistoma lusitanica (Vianna & Moraes, 1942) (Milan Chroust, personal observation 2024). However, this is not the case of Tomistoma calaritanum osteoderms, in which the keel is partially present (Zoboli et al. 2019). In Gavialosuchus or Tomistoma, dorsal osteoderms are wider than longer, whereas in Diplocynodon are mostly wider or subquadrate, however, this requires further study. Lastly, after comparisons with Tomistoma calaritanum material published in Zoboli et al. (2019), the external surface of dorsal osteoderms in Tomistoma generally has smooth edges without subcircular pits (see Zoboli et al. 2019: fig. 8), whereas in Diplocynodon subcircular pits are more present in the entire external surface, including even the edges of the osteoderm. Therefore, the described dorsal osteoderm is more consistent with the pattern observed in Diplocynodon. However, our sample does not include any ventral bipartite osteoderms, which occur in Diplocynodon but not in Gavialosuchus, and are of higher diagnostic value.
Therefore, all these discussed observations guardedly point to the generic assignment of the presented remains to the genus Diplocynodon. However, due to the scarcity of the material and lack of more diagnostic elements, we attribute the teeth and osteoderm to Crocodylia indet.
Discussion
The northernmost occurrence of crocodylians in the Cenozoic.—In the northern hemisphere, crocodylians currently inhabit tropical and sub-tropical zones ranging from 0–35° latitudes (Stevenson 2019). However, the northern limit of this range is reached only by alligatoroids (American and Asian alligators) and is a result of co-occurring natural conditions together with anthropogenic pressure (see below).
But how was the northernmost record changing throughout the Cenozoic era, especially during the warming periods? According to their physiology, with rising global temperatures, we can assume an expansion of the crocodylian biogeographic range to higher latitudes (Markwick 1998; Rio and Mannion 2021).
In the beginning of the Paleogene, the global distribution of crocodylians was generally similar to that of the Late Cretaceous (Markwick 1998; Puértolas-Pascual et al. 2015; Rio and Mannion 2021). The Danian (early Paleocene) records of Borealosuchus from Ravenscrag Formation in southern Canada occur at almost 52°N (Wu et al. 2001; Lindblad et al. 2022). Danian crocodylomorphs in Europe occurred even more northerly (see Fig. 6A and Table 2)—the range of Thoracosaurus in Denmark and southern Sweden exceeds 55°N (Puértolas-Pascual et al. 2015). In the Selandian (middle Paleocene), the range of North American crocodylians decreased to ca. 49°N (Wu et al. 2001; Lindblad et al. 2022) while European occurrences still exceeded 55°N in Denmark (Puértolas-Pascual et al. 2015). This changed in the Thanetian (late Paleocene) when the northern limit of crocodylians shifted to the south—the northernmost occurrence is known from Jeumont in NE France at 50°18’N (Puértolas-Pascual et al. 2015).
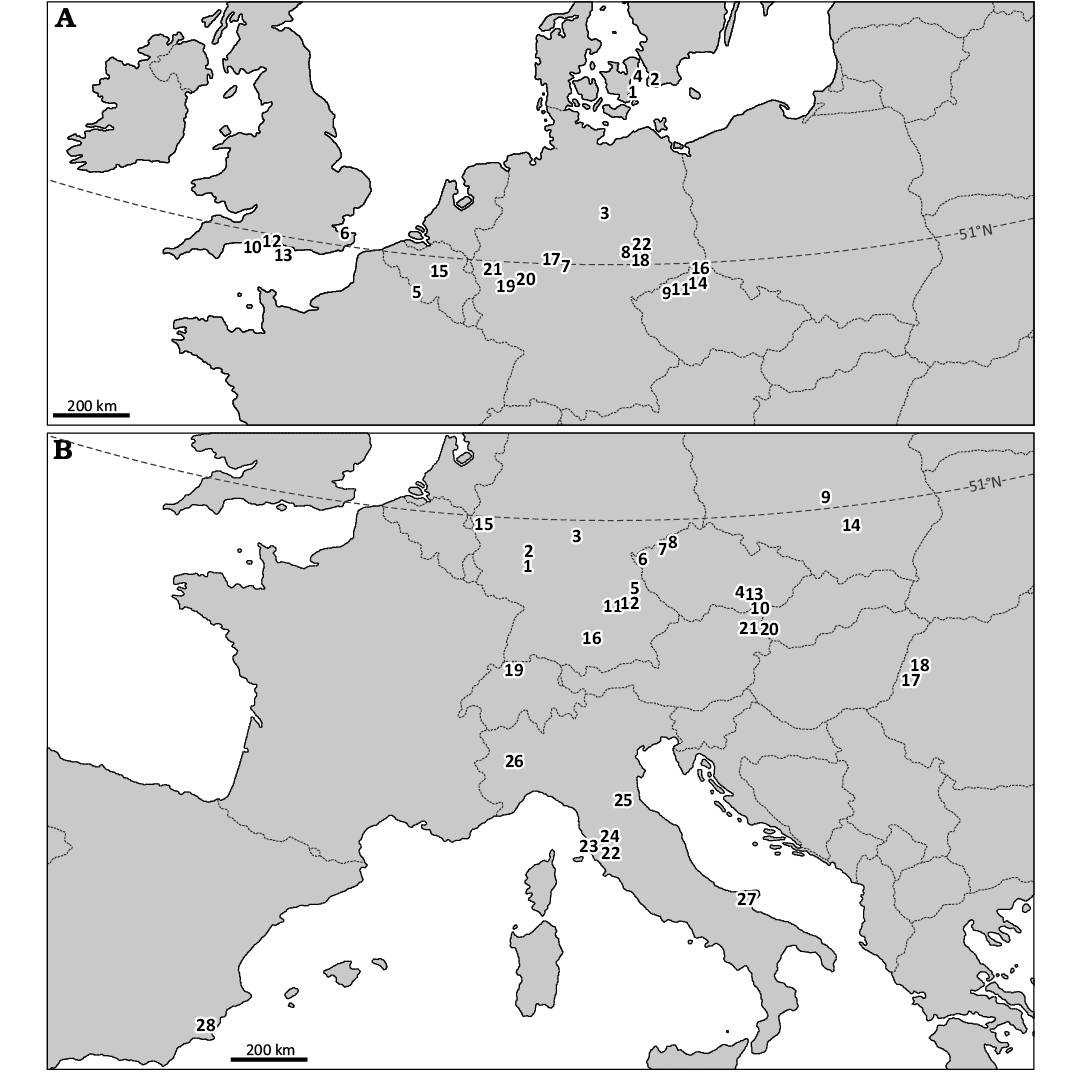
Fig. 6. Northernmost occurrences of crocodylians in Europe in succeeding stages of Paleogene and Neogene. A. Paleogene occurrences. Paleocene: Danian: 1, Faxe Quarry, Denmark; 2, Limhamn quarry near Malmö, Sweden; Selandian: 3, Walbeck near Weferlingen, Germany; 4, Gemmas Allé, Denmark; Thanetian: 5, Jeumont, France. Eocene: Ypresian: 6, Warden Point, Sheppey Island, England; Lutetian: 7, Stolzenbach near Borken, Germany; 8, Geiseltal, Germany; Bartonian: 9, Kučlín, Trupelník Hill, Czechia; 10, Creech Barrow Hill, England, UK; Priabonian: 11, Roudníky, Czechia; 12, Hordle Cliff near Milford on Sea, England, UK. Oligocene: Rupelian: 13, Bouldnor Cliff, Isle of Wight, England, UK; 14, Bechlejovice near Děčín, Czechia; 15, Hoogbutsel near Leuven, Belgium; 16, Seifhennersdorf, Germany; 17, Altenburg III and IV pit-mines, Borken, Germany; 18, Böhlen and Espenheim near Leipzig, Germany; Chattian: 19, Enspel near Marienberg, Germany; 20, Gusternhain, Germany; 21, Rott near Hennef, Germany; 22, Breitenfeld near Leipzig, Germany. B. Neogene occurrences. Miocene: Aquitanian: 1, Mombach and Weisenau, Mainz, Germany; 2, Biebrich and Mainz-Kastel, Wiesbaden, Germany; Aquitanian–Burdigalian: 3, Kaltennordheim, Germany; Burdigalian: 4, Ivančice, South Moravian Region, Czechia; 5, Wackersdorf near Schwandorf, Germany; 6, Jehličná near Sokolov, Czechia; 7, Ahníkov and Tušimice, Czechia; 8, Břešťany and Skyřice, Czechia; Burdigalian–?Langhian: 9, Szczerców, Poland; Langhian: 10, Kienberg near Mikulov, Czechia; 11, Appertshofen and Denkendorf near Ingolstadt, Germany; 12, Undorf near Regensburg, Germany; 13, Židlochovice, Czechia; 14, Pińczów, Poland; 15, Hambach, Germany; ?Langhian–Serravallian: 16, Kirrberg near Balzhausen, Germany; Serravallian: 17, Tășad and Vârciorog, Romania; 18, Subpiatră, Romania; 19, Anwil, Switzerland; 20, Devinská Nová Ves, Slovakia; 21, Vienna, Austria; Tortonian: 22, Casteani and Ribolla, Italy; Messinian: 23, Montebamboli, Italy; 24, Radicondoli, Italy; 25, Cava di Monticino, Italy; 26, Moncucco Torinese, Italy. Miocene–Pliocene transition: latest Messinian–earliest Zanclean: 27, Gargano, Italy. Pliocene: Zanclean: 28, Puerto de la Cadena, Spain. For references see Table 2.
It is striking that from the late Paleocene to the Middle Miocene, i.e., for over 40 Myr, the northern limit of crocodylian distribution in Europe was located at approx. the present-day 51°N parallel (see Figs. 6 and 7 and Table 2). Obviously, the true value of palaeolatitude of this limit was a bit different during the Cenozoic. From ca. 60 Ma to 20 Ma, due to the northward continental drift of central Europe, the true value of palaeolatitude was slowly increasing from ca. 42°N to 46°N and this rate markedly doubled in the Neogene (paleolatitude.org; also see van Hinsbergen et al. 2015). It can thus be estimated that the true palaeolatitude value for the Szczerców site (present-day 51°14’N) in the late Burdigalian (i.e., ca. 16.5 Ma) was approx. 47°N. Similar values can be obtained based on palaeogeographic reconstructions of the Tethyan (Mediterranean) realm and its surroundings (Ricou 1994; Perrin et al. 1998). From this point forward, the present-day coordinates are used unless clearly stated otherwise, with the understanding that the true palaeolatitude values were lower during the early stages of the Cenozoic.
This broad distribution of crocodylians was possible due to the Paleocene greenhouse climate following the Cretaceous climatic optimum with a slowly increasing mean annual temperature. This trend peaked and ended with the Paleocene–Eocene Thermal Maximum, without a significant impact on crocodylian distribution (Zachos et al. 2001, 2008; Mannion et al. 2015; Westerhold et al. 2020).
The warm climate during the early Eocene caused favourable conditions for crocodylians (Markwick 1998; Zachos et al. 2008; Mannion et al. 2015; Westerhold et al. 2020). The Ypresian (early Eocene) is the age of the northernmost recorded range limit of crocodylian distribution, remains of ?Allognathosuchus sp. were found in the Canadian Arctic, at over 78°N in Ellesmere Island, Nunavut (Dawson et al. 1976; Estes and Hutchison 1980) and at over 74°N in Banks Island, Northwest Territories (Eberle et al. 2014). The Asian record of crocodyliforms is known from almost 50°N, in Mongolia (Iijima et al. 2019). The northernmost Ypresian occurrence in Europe is at 51°26’N in Sheppey Island, United Kingdom (Benton and Spencer 1995).
From the Lutetian (early middle Eocene) to the Zanclean (Early Pliocene), i.e., during almost 45 Myr, Europe took the lead as the most favourable area for the northernmost distribution of crocodylians (see Figs. 6, 7 and Table 2). The northernmost record of Lutetian crocodylians is known from Geiseltal in Germany, over 51°N (Hastings and Hellmund 2017). Bartonian and Priabonian (late middle Eocene to late Eocene) occurrences of Diplocynodon sp. exceeding 50°N are known from Czechia (Böhme 2007; Chroust et al. 2019; see Fig. 6A and Table 2), but they are slightly surpassed by findings from southern England (Benton and Spencer 1995).
Table 2. The northernmost occurrences of crocodylians throughout the Cenozoic. Records in bold indicate the world’s northernmost occurrences, underlined records denote northernmost occurrences in Europe, in grey are the records described in this paper.
|
Epoch |
Age |
Locality |
Latitude |
Reference |
|
Holocene/Recent |
Alligator
|
extant, exceptional: Arkansas River, Wichita, Kansas, USA |
37°40’45.0”N |
|
|
extant, constant: Merchants Millpond, North Carolina, USA |
36°26’10.0”N |
|||
|
Alligator sinensis |
fossil: Wangyin site, Yanzhou District, Jining City, Shandong Province, China |
35°27’41.0”N |
||
|
historic: Hongze Lake, Jiangsu Province, China |
33°40’00.0”N |
|||
|
extant: Yangtze River Valley, Anhou Province, China |
31°00’00.0”N |
|||
|
Pleistocene |
Late Pleistocene |
Herculaneum, Missouri, USA |
38°15’26.0”N |
|
|
Jones Spring, Missouri, USA |
38°03’58.5”N |
|||
|
Chibanian |
Yage, Inasa-cho, Hamamatsu City, Shizuoka Prefecture, Honshu, Japan |
34°50’16.0”N |
||
|
Machikane-yama, Toyonaka City, Osaka Prefecture, Honshu, Japan |
34°48’25.0”N |
|||
|
Calabrian |
Nikaryo Syukugawara, Kawasaki City, Kanagawa Prefecture, Honshu, Japan |
35°37’12.6”N |
||
|
Kaminokuchi, Takatsuki City, Osaka Prefecture, Japan |
34°53’10.0”N |
|||
|
Gelasian |
Taga City, Shiga Prefecture, Honshu, Japan |
35°13’33.4”N |
||
|
Hino Town, Kōka City, and Konan City, Shiga Prefecture, Honshu, Japan |
35°01’34.0”N |
|||
|
Pliocene |
Piacenzian |
Kibogaoka, Terasho, and Oki, Kōka City, Shiga Prefecture, Honshu, Japan |
34°57’01.5”N |
|
|
Nomura-cho, Kameyama City, Mie Prefecture, Honshu, Japan |
34°51’01.0”N |
|||
|
Hatamura, Iga City, Mie Prefecture, Honshu, Japan |
34°46’09.0”N |
|||
|
Zanclean |
Puerto de la Cadena, Murcia, Spain |
37°55’09.0”N |
||
|
Gray Fossil Site, Washington County, Tennessee, USA |
36°23’10.0”N |
|||
|
Miocene–Pliocene transition |
latest Messinian–earliest Zanclean |
Gargano, Apulia, Italy |
41°48’00.0”N |
|
|
Miocene |
Messinian |
Moncucco Torinese, Piedmont, Italy |
45°03’36.5”N |
|
|
Cava di Monticino, Brisighella, Emilia-Romagna, Italy |
44°13’29.1”N |
|||
|
Radicondoli, Tuscany, Italy |
43°15’34.0”N |
|||
|
Montebamboli, Tuscany, Italy |
43°04’37.0”N |
|||
|
Tortonian |
Casteani and Ribolla, Tuscany, Italy |
42°59’32.0”N |
||
|
UNSM locality Sh-107, Sheridan County, Nebraska, USA |
42°48’00.0”N |
|||
|
George Sawyer Ranch, Cherry County, Nebraska, USA |
42°41’24.0”N |
|||
|
Serravallian |
Hernals, Vienna, Austria |
48°13’25.0”N |
||
|
Devinská Nová Ves – Sandberg, Bratislava, Slovakia |
48°12’00.0”N |
|||
|
Anwil, Basel-Country, Switzerland |
47°27’00.0”N |
|||
|
Subpiatră, Bihor District, Romania |
47°01’13.0”N |
|||
|
Tăşad and Vârciorog, Bihor District, Romania |
46°59’00.0”N |
|||
|
?Langhian– |
Kirrberg near Balzhausen, Bavaria, Germany |
48°13’37.4”N |
||
|
Langhian |
Hambach Pit-mine, North Rhine-Westphalia, Germany |
50°54’34.6”N |
||
|
Pińczów, Świętokrzyskie Voivodeship, Poland |
50°31’56.0”N |
Radwański 1974, 1977, 1985, 2015; this paper |
||
|
Židlochovice, South Moravian Region, Czechia |
49°02’31.6”N |
this paper |
||
|
Undorf near Regensburg, Bavaria, Germany |
49°01’38.0”N |
|||
|
Denkendorf near Ingolstadt, Bavaria, Germany |
48°55’57.0”N |
|||
|
Appertshofen near Ingolstadt, Bavaria, Germany |
48°52’21.3”N |
|||
|
Kienberg near Mikulov, South Moravian Region, Czechia |
48°48’13.5”N |
|||
|
Burdigalian– |
Szczerców field of Bełchatów pit-mine, Łódź Voivodeship, Poland |
51°14’00.0”N |
this paper |
|
|
Epoch |
Age |
Locality |
Latitude |
Reference |
|
Miocene |
Burdigalian |
Břešťany near Bílina, Ústí nad Labem Region, Czechia |
50°35’00.0”N |
|
|
Marianna Pit-mine, Skyřice near Most, Ústí nad Labem Region, Czechia |
50°29’00.0”N |
|||
|
Tušimice near Chomutov, Ústí nad Labem Region, Czechia |
50°26’00.0”N |
|||
|
Ahníkov I/II-Merkur North Pit-mine, Ústí nad Labem Region, Czechia |
50°26’00.0”N |
|||
|
Friedrich-Anna mine, Jehličná near Sokolov, Karlovy Vary Region, Czechia |
50°12’09.0”N |
|||
|
Wackersdorf near Schwandorf, Bavaria, Germany |
49°18’00.0”N |
|||
|
Ivančice, South Moravian Region, Czechia |
49°07’01.2”N |
|||
|
Aquitanian– |
Kaltennordheim, Thuringia, Germany |
50°37’40.0”N |
||
|
Aquitanian |
Biebrich and Mainz-Kastel, Wiesbaden, Hesse, Germany |
50°03’00.0”N |
||
|
Mombach and Weisenau, Mainz, Rhineland-Palatinate, Germany |
50°01’00.0”N |
|||
|
Oligocene |
Chattian |
Breitenfeld near Leipzig, Saxony, Germany |
51°25’50.0”N |
|
|
Rott near Hennef, North Rhine-Westphalia, Germany |
50°45’15.0”N |
|||
|
Gusternhain, Hesse, Germany |
50°40’10.0”N |
|||
|
Enspel near Marienberg in Westerwald, Rhineland-Palatinate, Germany |
50°37’15.0”N |
|||
|
Rupelian |
Böhlen and Espenheim near Leipzig, Saxony, Germany |
51°11’35.0”N |
||
|
Altenburg III and IV pit-mines, Borken, Hesse, Germany |
51°02’07.0”N |
|||
|
Seifhennersdorf, Saxony, Germany |
50°56’03.5”N |
|||
|
Hoogbutsel near Leuven, Brabant, Belgium |
50°50’46.0”N |
|||
|
Bechlejovice near Děčín, Ústí nad Labem Region, Czechia |
50°45’38.0”N |
|||
|
Bouldnor Cliff, Isle of Wight, England, UK |
50°42’30.0”N |
|||
|
Eocene |
Priabonian |
Hordle Cliff near Milford on Sea, Hampshire, England, UK |
50°43’43.0”N |
|
|
drilling Ru 60, Roudníky, Ústí nad Labem Region, Czechia |
50°39’30.0”N |
|||
|
Calf Creek near Eastend, Saskatchewan, Canada |
49°38’00.0”N |
|||
|
Bartonian |
Creech Barrow Hill, Dorset, England, UK |
50°38’29.5”N |
||
|
Kučlín-Trupelník Hill near Bílina, Ústí nad Labem Region, Czechia |
50°32’17.0”N |
|||
|
Lutetian |
Geiseltal, Sachsen-Anhalt, Germany |
51°20’02.0”N |
||
|
Stolzenbach near Borken, Hesse, Germany |
51°00’36.0”N |
|||
|
Ypresian |
Ellesmere Island, Nunavut, Canada |
78°40’10.0”N |
||
|
Eames River, Aulavik National Park, Banks Island, Canada |
74°10’00.0”N |
|||
|
Warden Point, Sheppey Island, Kent, England, UK |
51°26’00.0”N |
|||
|
Paleocene |
Thanetian |
Jeumont, Maubeuge, Nord Department, France |
50°18’00.0”N |
|
|
Selandian |
Gemmas Allé, Amager, Copenhagen, Denmark |
55°37’41.9”N |
||
|
Walbeck near Weferlingen, Saxony-Anhalt, Germany |
52°17’39.0”N |
|||
|
Danian |
Limhamn quarry near Malmö, Sweden |
55°34’03.1”N |
||
|
Faxe Quarry, Sjælland, Denmark |
55°15’29.8”N |
The cooling event at the transition to the Oligocene caused a decline in crocodylian diversity and distributional range, with only the genus Diplocynodon persisting in central Europe (Martin 2010; Mannion et al. 2015; Chroust et al. 2019; Macaluso et al. 2019; Celis et al. 2020). Other genera like Asiatosuchus, Allognathosuchus, Hassiacosuchus, Boverisuchus or sebecosuchians like Bergisuchus do not appear to survive the Eocene–Oligocene transition (Macaluso et al. 2019).
After this cooling event, the genus Diplocynodon did not change its range significantly. The 51°N parallel again served as a general northern boundary of crocodylian distribution (see Fig. 7). This is not only confirmed by the northernmost records from the Rupelian (lower Oligocene) of Böhlen and Espenheim near Leipzig (Karl 1990, 2007) and of Borken (Karl 2007; Karl and Müller 2008; Rio et al. 2020) in Germany (all slightly exceeding 51°N) but this is even further supported by a series of other Rupelian records approaching 51.0°N from Czechia, Belgium, England, and Germany (Smith 2003; Böhme 2007; Wolff et al. 2007; Chroust et al. 2019; see Fig. 6A and Table 2). This distribution pattern was generally similar in the Chattian (upper Oligocene) although the northernmost occurrences were solely reported from Germany, with the most northerly located site of Breitenfeld (51°25’50”N) near Leipzig (Karl 1991, 2007). Other German occurrences (see Fig. 6A and Table 2), almost reaching 50°N, are in the western part of the country (Ludwig 1877; Böhme and Lang 1991; Mörs 2002; Mayr and Poschmann 2009).
The Miocene recorded the highest peak of crocodylian radiation, particularly in the southern hemisphere (Celis et al. 2020). In the northern hemisphere, while North American and Asian alligatoroids did not reach 50°N during the Miocene (Li and Wang 1987; Brochu 1999; Rio and Mannion 2021), crocodylians did so in Europe, even exceeding 51°N (Szczerców).
In the beginning of the Miocene, tomistomines and diplocynodontines were present in central Europe (Böhme 2003; Luján et al. 2019; Chroust et al. 2021). During the Miocene Climatic Optimum, temperatures were more favourable again for crocodylians, which allowed them to expand their distribution northward. Indeed, diplocynodontines spread across western and central Europe, while tomistomines were located mostly in the Mediterranean area (Böhme 2003; Martin and Gross 2011; Nicholl et al. 2020). However, Gavialosuchus eggenburgensis from Eggenburg (Austria) dated to the Burdigalian, is an exception and reached 48°64’N and therefore represents the northernmost Neogene record of tomistomines (Toula and Kail 1885). The northernmost record of diplocynodontines during the Aquitanian comes from the Mainz/Wiesbaden area at ca. 50°N (Ludwig 1877; Schleich 1988). Younger, Burdigalian northernmost findings are relatively common in the Most Basin, NW Czechia, at ca. 50°30’N (Laube 1901, 1910; Liebus 1936; Luján et al. 2019; Chroust et al. 2021). However, these occurrences are slightly surpassed by an old record attributed to the Early Miocene age from Kaltennordheim, central Germany (Herbst 1857). Another Lower Miocene occurrence of supposed Burdigalian age is the present record of the osteoderm from Szczerców (51°14’00”N), the world’s northernmost record of a Neogene crocodylian (see Fig. 6B and Table 2). The northern range limit of crocodylians in Europe then reached the 51°N parallel and persisted through the Langhian, with the northernmost records from the coal deposits of Hambach, western Germany (Mörs et al. 2000; Macaluso et al. 2022) and from the littoral facies of Pińczów, Poland (Radwański 1974, 1977, 1985, 2015; Antunes in Młynarski 1984; see Figs. 3B, 5B, 6B, and Table 2). The area of distribution of crocodylians in Europe during the Early Miocene and the Langhian (i.e., MCO) was comparable to that of the late Paleocene through the Oligocene (Figs. 6, 7, and Table 2).
After the cooling event of the Miocene Climatic Transition the diversity and spatial distribution of crocodylians in the the northern hemisphere started to decline gradually and never exceeded the former latitudinal values (Mannion et al. 2015). Although in the Serravallian the northernmost occurrences were still located in central Europe, the northern limit of the range barely exceeded 48°N, known localities (see Figs. 6B, 7 and Table 2) are Kirrberg near Balzhausen, Germany (latest Langhian?–early Serravallian age; Böhme 2003), Devinská Nová Ves—Sandberg, Slovakia (Schlögl and Holec 2004; Pivko et al. 2017), and Hernals district of Vienna (Harzhauser et al. 2025)—the latter two of confirmed Serravallian age. In the Late Miocene the northern range limit of crocodylians shifted to the south, and the Tortonian northernmost records are known from the Italian localities of Casteani and Ribolla (ca. 43°N; Kotsakis et al. 2004; Delfino and Rook 2008), but they are closely followed by North American occurrences from Nebraska (Mook 1946; Markwick 1994; Whiting and Head 2020). In the Messinian, a short-lasting northward shift can be observed when crocodylians returned to northern Italy (Moncucco Torinese, ca. 45°N; Colombero et al. 2017). Generally, European tomistomines and diplocynodontines probably went extinct during the Late Miocene (Böhme 2003; Luján et al. 2019; Nicholl et al. 2020) and in southern Europe they were gradually replaced by the extant genus Crocodylus (Delfino et al. 2007; Delfino and Rook 2008; Delfino and Rossi 2013; Delfino et al. 2021). The latter is reported from the uppermost Miocene and Pliocene of southern Europe; however, it never reached higher latitudes than its predecessors and was apparently confined to the coastal areas or areas somehow linked to the coast. From the Miocene–Pliocene transition (Messinian–Zanclean) a crocodylian record is known from Gargano, Italy (almost 41°N; Kotsakis et al. 2004; Delfino et al. 2007, 2021). Unidentified teeth from Puerto de la Cadena, Spain (Piñero et al. 2017) located at almost 38ºN (Fig. 6B and Table 2) are the world’s northernmost crocodylian record of Zanclean age and represent the last known occurrence of crocodylians in Europe.
For the late Pliocene and almost the entire Pleistocene the centre of northernmost crocodylian occurrences shifted to Japan (Aoki 1983; Nojima and Itoigawa 2017; Iijima et al. 2018) and reached ca. 35°N for almost 3.5 Myr (see Fig. 7 and Table 2). This limit was exceeded by Alligator mississippiensis in Missouri, USA (ca. 38°N) only in the latest Pleistocene (Olson 1940; Mehl 1962; Preston 1979; Haynes 1985).
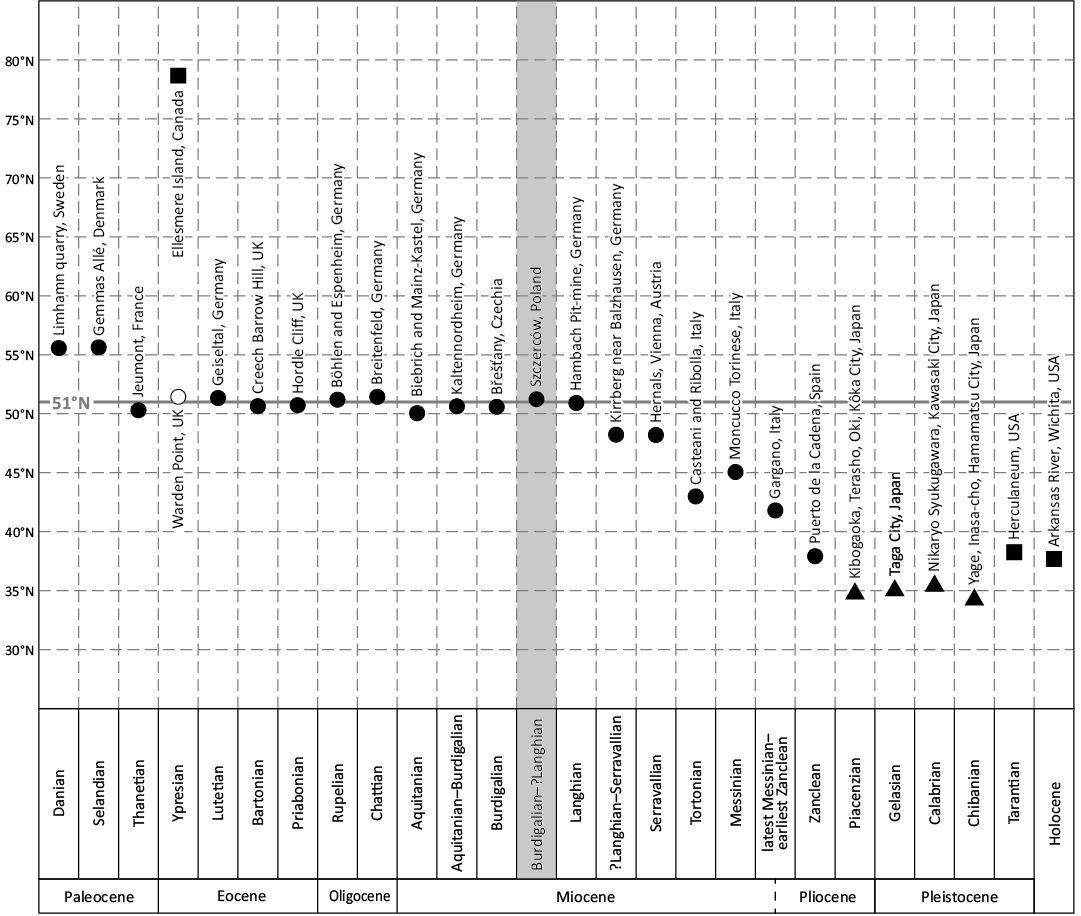
Fig. 7. The northermost occurrences of crocodylians throughout the Cenozoic. Records with black symbols indicate the world’s northernmost occurrences. Occurrences in Europe marked with circles, in North America with squares, in eastern Asia with triangles. Note the relatively stable location of the northern limit of crocodylian distribution in Europe at approx. the present-day 51°N parallel throughout the Thanetian to Langhian timespan. For references see Table 2.
In the Holocene, the northernmost limit of crocodylians, represented by alligators A. mississippiensis and A. sinensis moved southward, to a large extent due to human activity (Thorbjarnarson et al. 2002; Gardner et al. 2016; Pan et al. 2019).
The current northernmost limit of the natural range of crocodylians belongs to Alligator mississippiensis, which reaches the north-eastern coastal area of North Carolina, up to Albemarle Sound (Kellogg 1929; Whiting et al. 2016), locally even reaching ca. 36°25’N (Merchants Millpond; Gardner et al. 2016: 545, fig. 1). However, there are exceptional historical reports of individuals reaching as far as 38°N, e.g., Arkansas River in Wichita, Kansas (Cragin 1885).
The range of A. sinensis decreased gradually. Fossil remains from the Neolithic of the Wangyin site, Shandong Province in China were located at ca. 35°42’N, i.e., similar to the Pleistocene range of crocodylians in Japan (Iijima et al. 2016). In historic times the northern limit of this range retracted to 33°40’N (Hongze Lake, Jiangsu Province in China; Thorbjarnarson and Wang 1999; Iijima et al. 2016). The species was recently at the verge of extinction and its range shrunk to just a few water bodies in Yangtze River Valley, Anhou Province in China at ca. 31°N (Thorbjarnarson et al. 2002; Iijima et al. 2016).
Like most of the ectothermic fauna, crocodylians are sensitive to climatic changes and therefore serve as a climatic indicator (Markwick 1998; Böhme 2003). The crucial limiting factor for their distribution, aside from the humidity, is the minimal mean annual temperature. Most of the extant taxa live in temperatures above the freezing point, however, alligators are better adapted to cold, and the short-time freezing is not limiting for them. Alligator mississippiensis lives in an environment with a mean annual temperature of 16.4 °C and a minimal mean annual temperature of 1.7 °C, whereas Alligator sinensis lives in even colder conditions, with a mean annual temperature of 15.7 °C and a minimal mean annual temperature of -1.7 °C (Haller-Probst 1997). Alligator mississippiensis is often spotted in frozen waters, where only the distal snout is kept above the frozen water surface level for breathing (Brisbin et al. 1982). The occurrence of a crocodylian in Szczerców may suggest subtropical or warm-temperate conditions, with minimal mean annual temperature not falling below freezing point at ca. 47° northern latitude in the late Burdigalian of Europe (present-day 51°N). For more precise palaeoclimatic reconstruction, a more diversified fossil sample is needed.
Environmental requirements of Diplocynodon and possible dispersal routes.—Despite the presented material not being able to be classified at a lower taxonomic level, it brings new information about crocodylian biogeography and potential environmental requirements. Only two crocodylian genera are known from the Central Parathetys area in the Middle Miocene, with these two taxa being traditionally split between two environmental settings: Diplocynodon from freshwater environments and Gavialosuchus inhabiting marine settings. The Crocodylia indet. material described here (teeth and osteoderm) is more reminiscent of Diplocynodon than Gavialosuchus, and therefore the possibility of Diplocynodon inhabiting a marine setting is worth discussing. In fact, Diplocynodon occurrences in other marine deposits of the Parathetys Sea have been a topic of discussion in the context of its saltwater tolerance (Sabău et al. 2021; Venczel and Codrea 2022; Kocsis et al. 2023; Venczel 2023). Extant true crocodiles (Crocodylidae) have lingual glands to help them exclude the salt and therefore persist in hyperosmotic conditions across the seas. Alligators and caimans (Alligatoridae) lack lingual glands and therefore their presence in saltwater conditions is limited (Taplin et al. 1982; Taplin and Grigg 1989; Brochu 1999; Stout 2024). However, Alligator mississippiensis or Caiman latirostris could temporarily survive in a hyperosmotic environment (Taplin and Grigg 1993) and surprisingly, A. mississippiensis has been observed offshore in the Gulf of Mexico, 63 km distance from the mainland (Elsey 2005). Several examples of fossil alligatoroids (including the genus Diplocynodon) have been found in coastal deposits, and alligatoroids have also been suggested to disperse between continents throughout the Cenozoic (Stout 2024). Here, the described material is identified as Crocodylia indet. and comes from both freshwater and marine deposits. Based on traditional interpretations, the freshwater material (the osteoderm from Szczerców) could be arguably attributed to Diplocynodon, while the marine material (teeth from Pińczów and Židlochovice) could be linked to Gavialosuchus. After our examination of the material, all three specimens tentatively resemble Diplocynodon and therefore, the environmental requirements of the genus Diplocynodon are further discussed here. The Pińczów and Židlochovice sites represent shallow marine ecosystems rich in faunal assemblages typical of sub-tropical shallow waters of oceanic salinity. The preserved crocodylian material is fragmentary (teeth only), and therefore, transport from land and later re-deposition into marine deposits is possible. In both of these sites the mentioned remains are the only documented possible terrestrial elements among otherwise a very rich marine fauna. Although the terrestrial fauna at these sites is minor, at least an occasional marine lifestyle cannot be excluded for Diplocynodon as the hypothetical representative of both teeth from the discussed localities. Such an assumption is supported by the presence of Diplocynodon kochi in marine deposits from Romania (Venczel and Codrea 2022). A similar case has been described from the Máriahalom fossil site in Hungary, where dozens of teeth were tentatively assigned to the genus Diplocynodon. The isotopic composition of the dental enamel of the Máriahalom teeth is consistent with a brackish water environment (Kocsis et al. 2023). As such, if our hypothetical attribution to the genus Diplocynodon is correct, the herein presented material is consistent with the presence of this genus in a definite freshwater environment (Szczerców) but also supports its putative presence in marine environments (Pińczów and Židlochovice).
Therefore, the described findings: (i) contemplate the possibility of an at least partial marine lifestyle of Diplocynodon, (ii) throw into question the simplified division (and often diagnosis) of two discussed genera in relation to the environment, i.e., Gavialosuchus as saltwater and Diplocynodon as freshwater dwellers. This also assumes the potential coexistence of these two genera in particular environmental niches. Both extant A. mississippiensis and Crocodylus acutus also inhabit the coastal area of the southern tip of the Florida peninsula (Mazzotti et al. 2007, 2009). A similar sympatric coexistence was found in northern Australia, where Crocodylus porosus and Crocodylus johnstoni share the tidal rivers (Webb et al. 1983). In northern India, Gavialis gangeticus and Crocodylus palustris also cohabit in the same areas (Choudhary et al. 2018). Also, the current distributions of Tomistoma schlegelii and Crocodylus siamensis overlap in Mesangat Lake on Kalimantan Island, Indonesia (Staniewicz et al. 2018). The same fit for the fossil forms, for example, the coexistence of Late Miocene alligatoroid and gavialoid taxa is highlighted by the unique degree of crocodylian sympatry observed in the Urumaco Formation in Venezuela (Scheyer et al. 2013), similar to that at La Venta in Colombia (Wilson and Parker 2023). And of course, the European Eocene sites like Messel and Geisetal are also famous for the high niche partitioning of crocodylians (Morlo et al. 2004; Hastings and Hellmund 2017; Brochu and Miller-Camp 2018).
For the fossil forms here discussed, a different diet is presumed, where Gavialosuchus is a specialized ichthyophagous taxon (Antunes 2017), and Diplocynodon has a more generalized diet, comprising small mammals and herpetofauna like amphibians, snakes or turtles (Hastings and Hellmund 2017). A few findings of the coexistence of those two taxa exist in the literature but have never been solidly demonstrated; for instance, the material from German Molasse Basin in Ulm (Heizmann et al. 1989) or Portuguese Tagus Basin near Lisbon (Antunes 1994). In the case of the German and Austrian material, the coexistence has not been confirmed (Böhme 2003: supplement 1). However, to reveal these trophic relations, more fossil evidence is required.
The possible presence of Diplocynodon in the Szczerców field was undoubtedly allowed due to the freshwater environment in the Kleszczów Graben. The site was located in the southern part of the Polish Lowland Basin and was included in the catchment area of the North Sea Basin during the considered timespan of Early-to-Middle Miocene. The whole area was then most probably included within the fluvial system of the hypothetical Eridanos river flowing from Scandinavia, through the region of the modern Baltic Sea, northern Poland, northern Germany, and Denmark to the North Sea Basin (see Gibbard and Lewin 2016). Noteworthy, similar was the situation in the Most Basin in Czechia, drained by a river flowing NW, i.e., to the North Sea Basin (“Central River”; Mach et al. 2014: fig. 6). Both of these areas where Diplocynodon presence was recorded were thus included within the large system of fluvial/lacustrine/swamp environments reaching as far as NW Germany and including also the site of Hambach. As such, this vast freshwater domain was the most probable route of dispersal of Diplocynodon across inland central Europe, situated away from marine basins of the Paratethys.
In the context of various findings of Diplocynodon in central Europe, the dispersal routes of this genus cannot be restricted to freshwater basins only and seashore pathways should also be taken into consideration. This possibly applies to the specimens from Pińczów and Židlochovice, if they are indeed Diplocynodon. While the presence of rivers along the shores of the Fore-Carpathian Basin is obvious, they could not provide significant connections (i.e., dispersal routes) with the predominantly freshwater Polish Lowland Basin (or North Sea Basin in general) due to morphological barriers such as the Bohemian Massif and Central Polish Uplands. This means that putative saltwater tolerance of Diplocynodon is potentially an important factor that should be taken into consideration when reconstructing possible routes of dispersal of the representatives of this genus.
Conclusions
The current study presents records of Crocodylia indet. from the Karpatian (upper Lower Miocene) and lower Badenian (lower Middle Miocene) of the Fore-Carpathian Basin and its foreland, as the described material is either new or has been previously assigned to the gavialoid genus Tomistoma. A new specimen is described from the site of Szczerców, Poland, which was previously not known for yielding any vertebrate remains. A thorough compilation of the northernmost occurrences of these reptiles during the Cenozoic shows that the material from Szczerców is also the world’s northernmost record of a crocodylian from the entire Neogene. The palaeoenvironment reconstruction of Szczerców points towards this locality being a freshwater swamp, but other sites are reconstructed as being shallow marine. The taxonomic attribution of crocodylian remains together with their plausible ecology is discussed.
The description of a crocodylian from Szczerców opens the possibility to explore new vertebrate findings in the Middle Miocene of central Poland, and it specifically helps better understand the composition of the inland fauna in the north of the Central Paratethyan coast. The study of this region could help us establish biogeographical links between southwestern and eastern localities and draw the bigger picture of Eurasian faunal exchanges.
Acknowledgements
The authors are greatly indebted to the management and geologic staff of PGE GiEK KWB Bełchatów (Poland) for their supportive attitude and guidance during the reconnaissance fieldworks. We are grateful to Urszula Radwańska (Faculty of Geology, University of Warsaw, Poland), who helped us find old documentation and gave us access to the Pińczów material, which was part of the collection of the late Professor Andrzej Radwański. The cordial support and information on crocodylian occurrences in Central Paratethys from the participants of 10th NCSEE in Podčetrtek, Slovenia, particularly Valentina Hajek-Tadesse (Croatian Geological Survey, Zagreb, Croatia), Krešimir Krizmanić (INA d.d., Zagreb, Croatia), and Krisztina Sebe (Hungarian Natural History Museum, Budapest, Hungary) is highly appreciated. We deeply thank Víctor López-Rojas (Universidade Nova de Lisboa, Lisbon, Portugal) for taking additional pictures of Tomistoma lusitanica and Museu Geológico de Lisboa (LNEG) for providing the material. We equally extend our gratitude to Boris Ekrt (National Museum, Prague, Czechia) and Ewan Bodenham (University College, London, UK) for taking additional pictures of the genus Diplocynodon. For further discussion, we thank Christopher A. Brochu (University of Iowa, Iowa City, USA) and Phillip Mannion (University College, London, UK). Finally, we thank the editor Daniel Barta (Oklahoma State University, Tahlequah, USA) and reviewers Ane de Celis (National University of Distance Education, Madrid, Spain) and Massimo Delfino (University of Torino, Italy), whose comments highly improved this manuscript. This study was funded by Faculty of Geology, University of Warsaw (grant UW no. 501-D113-01-1130202) to Marcin Górka, an Arthur James Boucot Research Grant (Paleontological Society) to Sergi López-Torres, National Science Centre (NCN) grant (2022/47/B/ST10/02686) to Mateusz Tałanda, and by institutional support of long-term conceptual development of research institutions provided by the Ministry of Culture of the Czech Republic (ref. MK000094862) to Jakub Březina.
Editor: Daniel Barta.
References
Alaburić, S. and Marković, Z. 2010. New records of mammal remains from the Miocene sediments of the cement mine in Popovac (Serbia). Bulletin of the Natural History Museum 3: 95–103.
Andreyeva-Grigorovich, A.S., Kulchytsky, Y.O., Gruzman, A.D., Lozynyak, P.Y., Petrashkevich, M.Y., Portnyagina, L.O., Ivanina, A.V., Smirnov, S.E., Trofimovich, N.A., Savitskaya, N.A., and Shvareva, N.J. 1997. Regional stratigraphic scheme of Neogene formations of the Central Paratethys in the Ukraine. Geologica Carpathica 48: 123–136.
Antunes, M.T. 1994. On Western Europe Miocene Gavials (Crocodylia) their paleogeography, migrations and climatic significance. Comunicações Instituto Geológico e Mineiro 80: 57–69.
Antunes, M.T. 2017. Huge Miocene crocodilians from Western Europe: predation, comparisons with the “false gharial” and size. Anuário do Instituto de Geociências – UFRJ 40 (3): 117–130. Crossref
Aoki, R. 1983. A new generic allocation of Tomistoma machikanense, a fossil crocodilian from the Pleistocene of Japan. Copeia 1: 89–95. Crossref
Bałuk, W. and Radwański, A. 1977. Organic communities and facies development of the Korytnica Basin (Middle Miocene; Holy Cross Mountains, Central Poland). Acta Geologica Polonica 27: 85–123.
Bedford, B.L. and Godwin K.S. 2003. Fens of the United States: distribution, characteristics, and scientific connection versus legal isolation. Wetlands 23 (3): 608–629. Crossref
Berg, D.E. 1966. Die Krokodile, insbesondere Asiatosuchus und aff. Sebecus?, aus dem Eozan von Messel bei Darmstadt/Hessen. Abhandlungen des Hessischen Landesamtes fur Bodenforschung 52: 1–105.
Benton, M.J. and Spencer, P.S. 1995. Fossil Reptiles of Great Britain. Geological Conservation Review Series no. 10. xii+386 pp. London. Crossref
Bielowicz, B. and Morga, R. 2021. Micro-Raman spectroscopy of selected macerals of the huminite group: An example from the Szczerców lignite deposit (Central Poland). Energies 14: 281. Crossref
Böhme, M. 2002. Lower vertebrates (Teleostei, Amphibia, Sauria) from the Karpatian of the Korneuburg Basin—palaeoecological, environmental and palaeoclimatical implications. Beiträge zur Paläontologie 27: 339–353.
Böhme, M. 2003. Miocene climatic optimum: evidence from ectothermic vertebrates of Central Europe. Palaeogeography, Palaeoclimatology, Palaeoecology 195: 389–401. Crossref
Böhme, M. 2007. Revision of the cyprinids from the Early Oligocene of the České Středohoří Mountains, and the phylogenetic relationships of Protothymallus Laube 1901 (Teleostei, Cyprinidae, Gobioninae). Acta Musei Nationalis Pragae Series B Historia Naturalis 63: 177–196.
Böhme, M. 2010. Ectothermic vertebrates (Actinopterygii, Allocaudata, Urodela, Anura, Crocodylia, Squamata) from the Miocene of Sandelzhausen (Germany, Bavaria) and their implications for environment reconstruction and palaeoclimate. Palaontologische Zeitschrift 84: 3–41. Crossref
Böhme, M. and Ilg, A. 2003. fosFARbase, https://www.fosfarbase.org/ (accessed 1.02.2024)
Böhme, W. and Lang, M. 1991. The reptilian fauna of the Late Oligocene locality Rott near Bonn (Germany) with special reference to the taxonomic assignment of ‘Lacerta’ rottensis von Meyer, 1856. Neues Jahrbuch für Geologie und Paläontologie. Monatshefte 1991 (9): 515–525. Crossref
Botfalvai, G., Kocsis, L., Szabó, M., Király, E., and Sebe, K. 2023. Preliminarily report on rare earth element taphonomy of a Miocene mixed age fossil vertebrate assemblage (Pécs-Danitzpuszta, Mecsek Mts., Hungary): uptake mechanism and possible separation of palaeocommunities. Historical Biology 35: 498–517. Crossref
Brachaniec, T., Środek, D., Surmik, D., Niedźwiedzki, R., Georgalis, G.L., Płachno, B.J., Duda, P., Lukeneder, A., Gorzelak, P., and Salamon M.A. 2022. Comparative actualistic study hints at origins of alleged Miocene coprolites of Poland. PeerJ 10: e13652. Crossref
Březina, J., Alba, D.M., Ivanov, M., Hanáček, M., and Luján, Á.H. 2021. A middle Miocene vertebrate assemblage from the Czech part of the Vienna Basin: Implications for the paleoenvironments of the Central Paratethys. Palaeogeography, Palaeoclimatology, Palaeoecology 575: 110473. Crossref
Březina, J., Chroust, M., Szczygielski, T., and Hrazdil, V. 2024. Miocene crocodylian and marine turtle from Kienberg: First record from the Moravian part of the Vienna Basin. Geologica Carpathica 75: 329–338. Crossref
Brisbin, I.L., Standora, E.A., and Vargo, M.J. 1982. Body temperatures and behavior of American alligators during cold winter weather. American Midland Naturalist 107: 209–218. Crossref
Brochu, C.A. 1999. Phylogenetics, taxonomy, and historical biogeography of Alligatoroidea. Journal of Vertebrate Paleontology 19: 9–100. Crossref
Brochu, C.A. and Miller-Camp, J. 2018. Crocodyliforms—large-bodied Carnivores. In: K.T. Smith, S.F. Schaal, and J. Habersetzer (eds.), Messel—An Ancient Greenhouse Ecosystem, 59–168. Schweizerbart Science Publishers, Frankfurt am Main.
Brzobohatý, R. 1997. Paleobathymetrie spodního badenu karpatské předhlubně na Moravě z pohledu otolitových faun. In: Š. Hladilová (ed.), Sborník příspěvků – Dynamika vztahů marinního a kontinentálního prostředí, 37–45. Masaryk University, Brno.
Buday, T. (ed.) 1963. Geologická mapa ČSSR, mapa předčtvrtohorních útvarů, 1:200 000, M-33-XXXIX, list Brno. Ústřední ústav geologický, Prague.
Bulić, J. and Jurišić-Polšak, Z. 2009. Macropalaeontology and stratigraphy of lacustrine Miocene deposits at Crnika beach on the Island of Pag (Croatia). Geologica Croatica 62: 135–156. Crossref
Burchart, J., Kasza, L., and Lorenc, S. 1988. Fission-track zircon dating of tuffitic intercalations (Tonstein) in the Brown-Coal Mine “Bełchatów”. Bulletin of the Polish Academy of Sciences, Earth Sciences 36: 281–286.
Burke, P.M.J., Nicholl, C.S.C., Pittard, B.E., Sallam, H., and Mannion, P.D. 2024. The anatomy and taxonomy of the North African Early Miocene crocodylian ‘Tomistoma’ dowsoni and the phylogenetic relationships of gavialoids. Journal of Systematic Palaeontology 22 (1): 2384548. Crossref
Bůžek, Č. and Holý, F. 1964. Small-sized plant remains from the coal formation of the Chomutov-Most-Teplice Basin. Sborník geologických věd, Paleontologie 4: 105–138.
Capellini, G. 1890. Sul coccodrilliano garialoide (Tomistoma calaritanus) scoperto nella collina di Cagliari nel MDCCCLXVIII. Atti della Reale Accademia dei Lincei, Memorie della Classe di Scienze Fisiche, Matematiche e Naturali 286: 507–533.
Celis, A.D. de, Narváez, I., and Ortega, F. 2020. Spatiotemporal palaeodiversity patterns of modern crocodiles (Crocodyliformes: Eusuchia). Zoological Journal of the Linnean Society 189: 635–656. Crossref
Čerňanský, A., Tóth, C., and Šurka, J. 2012. Nálezy krokodíla a korytnačiek zo spodného miocénu lokality Baňa Dolina vo Veľkom Krtíši (Slovensko). Acta Geologica Slovaca 4: 113–123.
Choudhary, S., Choudhury, B.C., and Gopi, G.V. 2018. Spatio‐temporal partitioning between two sympatric crocodilians (Gavialis gangeticus & Crocodylus palustris) in Katarniaghat Wildlife Sanctuary, India. Aquatic Conservation: Marine and Freshwater Ecosystems 28: 1067–1076.
Chroust, M., Mazuch, M., and Luján, À.H. 2019. New crocodilian material from the Eocene–Oligocene transition of the NW Bohemia (Czech Republic): An updated fossil record in Central Europe during the Grande Coupure. Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen 293: 73–82. Crossref
Chroust, M., Mazuch, M., Ivanov, M., Ekrt, B., and Luján, À.H. 2021. First remains of Diplocynodon cf. ratelii from the early Miocene sites of Ahníkov (Most Basin, Czech Republic). Bulletin of Geosciences 96: 123–138. Crossref
Chroust, M., Mazuch, M., Ivanov, M., Alba, D.M., and Luján, À.H. 2023. Redescription of the soft-shell turtle Rafetus bohemicus (Testudines, Trionychidae) from the Early Miocene of Czechia. PeerJ 11: e15658. Crossref
Cicha, I. 1978. Faziostratotypus: Židlochovice. In: A. Papp, I. Cicha, J. Seneš, and F. Steininger (eds.), Chronostratigraphie und Neostratotypen, Miozän der Zentralen Parathetys, M4 Badenien. [Chronostratigraphy and Neostratotypes, Miocene of the Central Paratethys, M4 Badenian], 168–169. Slovenská akadémia vied, Bratislava.
Colombero, S., Alba, D.M., D’Amico, C., Delfino, M., Esu, D., Giuntelli, P., Harzhauser, M., Mazza, P.P.A., Mosca, M., Neubauer, T.A., Pavia, G., Pavia, M., Villa, A., and Carnevale, G. 2017. Late Messinian mollusks and vertebrates from Moncucco Torinese, north-western Italy. Paleoecological and paleoclimatological implications. Palaeontologia Electronica 20.1.10A: 1–66. Crossref
Cook, C.D.K., Gut, B.J., Rix, E.M., and Schneller, J. 1974. Water Plants of the World: A Manual for the Identification of the Genera of Freshwater Macrophytes. 561 pp. Springer, Dordrecht.
Cragin, F.W. 1885. Editorial notes. Bulletin of the Washburn College Laboratory of Natural History 1 (3): 111.
Czarnocki, J. 1933. Przewodnie rysy stratygrafii i paleogeografii miocenu w południowej Polsce. Posiedzenia Naukowe Państwowego Instytutu Geologicznego 36: 16–25.
Czyżewska, T. 1988. Natural endocranial casts of the whales Pinocetus polonicus Czyżewska & Ryziewicz, 1976, from the Pińczów Limestones (Middle Miocene; southern slopes of the Holy Cross Mountains, Central Poland). Acta Geologica Polonica 38: 45–49.
Czyżewska, T. and Radwański, A. 1991. Middle Miocene (Badenian) delphinid and phocoenid remains from the Fore-Carpathian Depression in southern Poland. Acta Geologica Polonica 41: 183–191.
Czyżewska, T. and Ryziewicz, Z. 1976. Pinocetus polonicus gen. n., sp. n. (Cetacea) from the Miocene limestones of Pińczów, Poland. Acta Palaeontologica Polonica 21: 259–291.
Danel, W. and Gałązka, D. 2007. Mapa geologiczna Polski bez utworów czwartorzędowych, 1:200000, arkusz Łódź. Państwowy Instytut Geologiczny, Warszawa.
Dawson, M.R., West, R.M., Langston, Jr., W., and Hutchison, J.H. 1976. Paleogene terrestrial vertebrates: northernmost occurrence, Ellesmere Island, Canada. Science 192: 781–782. Crossref
Delfino, M. and Rook, L. 2008. African crocodylians in the late Neogene of Europe. A revision of Crocodylus bambolii Ristori, 1890. Journal of Paleontology 82: 336–343. Crossref
Delfino, M. and Rossi, M.A. 2013. Fossil crocodylid remains from Scontrone (Tortonian, Southern Italy) and the late Neogene Mediterranean biogeography of crocodylians. Geobios 46: 25–31. Crossref
Delfino, M., Böhme, M., and Rook, L. 2007. First European evidence for transcontinental dispersal of Crocodylus (late Neogene of southern Italy). Zoological Journal of the Linnean Society 149: 293–307. Crossref
Delfino, M., Luján, À.H., Abella, J., Alba, D.M., Böhme, M., Pérez-Ramos, A., Tschopp, E., Morales, J., and Montoya, P. 2021. Late Miocene remains from Venta del Moro (Iberian Peninsula) provide further insights on the dispersal of crocodiles across the late Miocene Tethys. Journal of Paleontology 95: 184–192. Crossref
Doláková, N., Brzobohatý, R., Hladilová, Š., and Nehyba, S. 2008. The red-algal facies of the lower Badenian limestones of the Carpathian Foredeep in Moravia (Czech Republic). Geologica Carpathica 59 (2): 133–146.
Doláková, N., Holcová, K., Nehyba, S., Hladilová, Š., Brzobohatý, R., Zágoršek, K., Hrabovský, J., Seko, M., and Utescher, T. 2014. The Badenian parastratotype at Židlochovice from the perspective of the multiproxy study. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 271 (2): 169–201. Crossref
Drewniak, A. 1994. Coralline algae from the Pinczów Limestones (Middle Miocene; southern slopes of the Holy Cross Mountains, Central Poland) as environmental indicators. Acta Geologica Polonica 44: 117–135.
Dvořák, Z., Mach, K., Prokop, J., and Knor, S. 2010. Třetihorní Fauna Severočeské Hnědouhelné Pánve. 178 pp. Granit, Praha.
Eberle, J.J., Gottfried, M.D., Hutchison, J.H., and Brochu, C.A. 2014. First record of Eocene bony fishes and crocodyliforms from Canada’s Western Arctic. PLoS ONE 9 (5): e96079. Crossref
Ekrt, B., Mikuláš, R., Wagner, J., Čermák, S., Procházková, K., Kadlecová, E., and Fejfar, O. 2016. New contribution to the palaeoichnology and taphonomy of the Ahníkov fossil site, Early Miocene, Most Basin (The Czech Republic). Fossil Imprint 72: 10–13. Crossref
Elsey, R.M. 2005. Unusual offshore occurrence of an American alligator. Southeastern Naturalist 4: 533–536. Crossref
Estes, R. and Hutchison, J.H. 1980. Eocene lower vertebrates from Ellesmere Island, Canadian Arctic Archipelago. Palaeogeography, Palaeoclimatology, Palaeoecology 30: 325–347. Crossref
Fostowicz-Frelik, Ł., Nadachowski, A., and Kowalewska-Groszkowska, M. 2012. New data on the Miocene stem lagomorph Eurolagus fontannesi, and its northernmost record. Acta Palaeontologica Polonica 57:1–20. Crossref
Förster, R. 1979. Decapod crustaceans from the Middle Miocene (Badenian) deposits of Southern Poland. Acta Geologica Polonica 29: 89–106.
Friis, E.M. 1985. Angiosperm fruits and seeds from the Middle Miocene of Jutland (Denmark). Biologiske Skrifter 24 (3): 1–165.
Gardner, B., Garner, L.A., Cobb, D.T., and Moorman, C.E. 2016. Factors affecting occupancy and abundance of American alligators at the northern extent of their range. Journal of Herpetology 50: 541–547. Crossref
Gibbard, P.L. and Lewin, J. 2016. Filling the North Sea Basin: Cenozoic sediment sources and river styles. Geologica Belgica 19: 201–217. Crossref
Gmelin, J.F. 1789. Caroli a Linné, equitis aurati de stella polari, …Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima tertia, aucta, reformata. Tom. 1 Pars 3, 1033–1516. Georg Emanuel Beer, Leipzig.
Gotowała, R. and Hałuszczak, A. 2002. The Late Alpine structural development of the Kleszczów Graben (Central Poland) as a result of reactivation of the pre-existing, regional dislocations. EGU Stephan Mueller Special Publication Series 1: 137–150. Crossref
Górka, M. 2018a. Badenian (middle Miocene) decapod crustaceans from western Ukraine, with remarks on eco-taphonomy, palaeoecology and biogeography. Acta Geologica Polonica 8: 511–535.
Górka, M. 2018b. Late Badenian zooxanthellate corals of the Medobory Hills (western Ukraine) and their environmental significance. Annales Societatis Geologorum Poloniae 88: 243–256. Crossref
Górka, M. 2019. An overview of the Badenian and Sarmatian succession in the northern margin of the Carpathian Foredeep Basin. In: B. Studencka (ed.), 8th International Workshop Neogene of Central and South-Eastern Europe. Abstract Volume, Field Trip Guidebook, 27th – 31st May, 2019, Chęciny, Poland, 91–97. Faculty of Geology, University of Warsaw, Warsaw.
Górka, M., Studencka, B., Jasionowski, M., Hara, U., Wysocka, A., and Poberezhskyy, A. 2012. The Medobory Hills (Ukraine): Middle Miocene reef systems in the Paratethys, their biological diversity and lithofacies. Biuletyn Państwowego Instytutu Geologicznego 449: 147–174.
Gregor, H.-J. 1977. Subtropische Elemente im europäischen Tertiär II (Fruktifikationen) Die Gattungen Quercus (Fagaceae), Myristicacarpum (Myristicaceae), Brueckelholzia und Cyclea (Menispermaceae), Disanthus und Fortunearia (Hamamelidaceae), Frangula (Rhamnaceae), Palaeocayratia (Vitaceae) und Acanthopanax (Araliaceae). Paläontologische Zeitschrift 51 (3–4): 199–226. Crossref
Gregor, H.-J. 1982. Die jungtertiären Floren Süddeutschlands: Paläokarpologie, Phytostratigraphie, Paläoökologie, Paläoklimatologie. 278 pp. Ferdinand Enke, Stuttgart.
Haisig, J. 2011. Mapa geologiczna Polski bez utworów czwartorzędowych, 1:200000, arkusz Częstochowa. Państwowy Instytut Geologiczny, Warszawa.
Hajek-Tadesse, V., Wacha, L., Horvat, M., Galović, I., Bakrač, K., Grizelj, A., Mandic, O., and Reichenbacher, B. 2023. New evidence for Early Miocene palaeoenvironmental changes in the North Croatian Basin: Insights implicated by microfossil assemblages. Geobios 77: 1–25. Crossref
Haller-Probst, M.S. 1997. Die Verbreitung der Reptilia in den Klimazonen der Erde, unter Beruecksichtigung kaenozoischer Vorkommen Europas. Courier Forschungsinstitut Senckenberg 203: 1–67.
Harzhauser, M. and Piller, W. 2004: Integrated stratigraphy of the Sarmatian (Upper Middle Miocene) in the western Central Paratethys. Stratigraphy 1 (1): 65–86. Crossref
Harzhauser, M. and Piller, W.E. 2007. Benchmark data of a changing sea —Palaeogeography, Palaeobiogeography and events in the Central Paratethys during the Miocene. Palaeogeography, Palaeoclimatology, Palaeoecology 253: 8–31. Crossref
Harzhauser, M., Kroh, A., Mandic, O., Piller, W.E., Göhlich, U., Reuter, M., and Berning, B. 2007. Biogeographic responses to geodynamics: A key study all around the Oligo-Miocene Tethyan Seaway. Zoologischer Anzeiger 246: 241–256.
Harzhauser, M., Göhlich, U.B., Gross, M., and Vasilyan, D. 2025. The last crocodylian in Central Europe? A new occurrence from the late Middle Miocene of the Vienna Basin (Austria). Historical Biology, 1–8 [available online, https://doi.org/10.1080/08912963.2025.2466048]. Crossref
Hastings, A.K. and Hellmund, M. 2017. Evidence for prey preference partitioning in the middle Eocene high-diversity crocodylian assemblage of the Geiseltal-Fossillagerstätte, Germany utilizing skull shape analysis. Geological Magazine 154: 119–146. Crossref
Haynes, C.V., Jr. 1985. Mastodon-bearing springs and Late Quaternary geochronology of the lower Pomme de Terre Valley, Missouri. Geological Society of America Special Paper 204: 1–35. Crossref
Heizmann, E.P.J., Bloos, G., Böttcher, R., Werner, J., and Ziegler, R. 1989. Ulm-Westtangente und Ulm-Uniklinik: Zwei neue Wirbeltier-Faunen aus der Unteren Süßwasser-Molasse (Untermiozän) von Ulm (Baden-Württemberg). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie un Paläontologie) 153: 1–14.
Herbst, G. 1857. Braunkohlen-Gebirge mit Folliculites Kaltennordheimensis und Acerotherium. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1857: 58–59.
Hír, J., Venczel, M., Codrea, V., Angelone, C., Van den Hoek Ostende, L.W., Kirscher, U., and Prieto, J. 2016. Badenian and Sarmatian s.str. from the Carpathian area: Overview and ongoing research on Hungarian and Romanian small vertebrate evolution. Comptes Rendus Palevol 15: 863–875. Crossref
Hír, J., Venczel, M., and Sebe, K. 2024. Middle Miocene (late Badenian) microvertebrates from Hidas, SW Hungary. Geologia Croatica 77: 15–28. Crossref
Hohenegger, J., Ćorić, S., and Wagreich, M. 2014: Timing of the Middle Miocene Badenian Stage of the Central Paratethys. Geologica Carpathica 65: 55–66. Crossref
Hohenegger, J., Ćorić, S., Khatun, M., Pervesler, P., Rögl, F., Rupp, C., Selge, A., Uchman, A., and Wagreich, M. 2009. Cyclostratigraphic dating in the lower Badenian (Middle Miocene) of the Vienna Basin (Austria): the Baden-Sooss core. International Journal of Earth Sciences 98: 915–930. Crossref
Holman, J.A. 1972. Herpetofauna of the Calf Creek Local Fauna (Lower Oligocene: Cypress Hills Formation) of Saskatchewan. Canadian Journal of Earth Sciences 9: 1612–1631. Crossref
Hörnes, M. and Reuss, A.E. 1870. Die Fossilen Mollusken des Tertiaer-Beckens von Wien. II. Band: Bivalven. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 4: 1–479.
Hyžný, M., Hudáčková, N., Biskupič, R., Rybár, R., Fuksi, T., Halásová, E., Zágoršek, K., Jamrich, M., and Ledvák, P. 2012. Devínska Kobyla —a window into the Middle Miocene shallow-water marine environments of the Central Paratethys (Vienna Basin, Slovakia). Acta Geologica Slovaca 4: 95–111.
Iijima, M., Takahashi, K., and Kobayashi, Y. 2016. The oldest record of Alligator sinensis from the Late Pliocene of Western Japan, and its biogeographic implication. Journal of Asian Earth Sciences 124: 94–101. Crossref
Iijima, M., Momohara, A., Kobayashi, Y., Hayashi, S., Ikeda, T., Taruno, H., Watanabe, K., Tanimoto, M., and Furui, S. 2018. Toyotamaphimeia cf. machikanensis (Crocodylia, Tomistominae) from the Middle Pleistocene of Osaka, Japan, and crocodylian survivorship through the Pliocene–Pleistocene climatic oscillations. Palaeogeography, Palaeoclimatology, Palaeoecology 496: 346–360. Crossref
Iijima, M., Tsubamoto, T., Tsogtbaatar, K., Chinzorig, T., and Baasankhuu, S. 2019. Discovery of a crocodyliform tooth from the upper Eocene Ergilin Dzo Formation, Mongolia. Acta Palaeontologica Polonica 64: 775–778. Crossref
Itoigawa, J. 1989. Tropical spike in Early Middle Miocene (ca. 16 Ma) of southwest Japan. In: L. Gengwu, R. Tsuchi, and L. Qibin (eds.), Proceedings of International Symposium on Pacific Neogene Continental and Marine Events, National Working Group of China for IGCP-246, 19–26. Nanjing University Press, Nanjing.
Ivanov, M. 2022. Miocene snakes of Eurasia: A review of the evolution of snake communities. In: D.J. Gower and H. Zaher (eds.), The Origin and Early Evolutionary History of Snakes. Systematics Association Special Volume Series, 85–110. Cambridge University Press, Cambridge. Crossref
Jerzmańska, A. 1958. Scorpaena ensiger (Jordan & Gilbert) z miocenu Pińczowa (Scorpaena ensiger (Jordan & Gilbert) from the Miocene of Pińczów, Poland). Acta Palaeontologica Polonica 3: 151–159.
Jipa, D.C. and Olariu, C. 2009. Dacian Basin. Depositional architecture and sedimentary history of a Paratethys Sea. Geo-Eco-Marina. Special Publication 3: 1–264.
Jovanović, M., Ðurić, D., and Marković, Z. 2002. Tertiary reptiles of the central part of the Balkan peninsula. Biota 3: 67–75.
Karl, H.-V. 1990. Fossile Krokodilreste (Reptilia, Crocodylia) aus dem marinen Mitteloligozän des Weisselsterbeckens. Mauritiana (Altenburg) 12: 483–488.
Karl, H.-V. 1991. Über einen weiteren fossilen Krokodilrest aus dem sächsischen Oligozän. Mauritiana (Altenburg) 13: 247–248.
Karl, H.-V. 2007. The fossil reptiles (Reptilia: Chelonii, Crocodylia) from the marine Early Oligocene of the Weisselster Basin (Central Germany:Saxonia). Studia Geologica Salmanticensia 43: 25–66.
Karl, H.-V. and Müller, A. 2008. New fossil reptil material (Reptilia: Chelonii, Crocodylia) from the Lower Oligocene of Borken (Central Germany: Hesse). Studia Geologica Salmanticensia 44: 41–58.
Keenan, S.W. and Engel, A.S. 2017. Reconstructing diagenetic conditions of bone at the Gray Fossil Site, Tennessee, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 471: 48–57. Crossref
Kellogg, R. 1929. The habits and economic importance of alligators. Technical Bulletin 147: 1–36.
Kirchheimer, F. 1937. Paläobotanische Beiträge zur Kenntnis des Alters deutscher Braunkohlenschichten II: Die braunkohlenführenden Tone von Siegburg (Rheinland) und Kamenz (Oberlausitz). Braunkohle: Zeitschrift für Gewinnung und Verwertung der Braunkohle 36: 893–931.
Knobloch, E. 1978. Die untermiozäne Flora von Šafov in Südmähren. Vĕstnik Ústředniho ústavu geologického 53 (3): 153–162.
Kocsis, L., Rabi, M., Ulianov, A., Cipriani, A., Farkas, I.M., and Botfalvai, G. 2023. Geochemical investigation of the mixed Máriahalom vertebrate fauna at the Paleogene–Neogene boundary in the Central Paratethys: environmental conditions and age constrain. Swiss Journal of Palaeontology 142 (17): 1–21. Crossref
Kopecká, J., Holcová, K., Brlek, M., Scheiner, F., Ackerman, L., Rejšek, J., Milovský, R., Baranyi, V., Gaynor, S., Galović, I., Brčić, V., Belak, M., and Bakrač, K. 2022. A case study of paleoenvironmental interactions during the Miocene Climate Optimum in southwestern Paratethys. Global and Planetary Change 211: 103784. Crossref
Kordos, L. 1985. A magyarországi eggenburgi – szarmata képződmények szárazföldi gerinces maradványai, biozonációja és rétegtani korrelációja. Magyar Állami Földtani Intézet évi jelentése az 1983 évről: 157–166.
Kordos, L. and Solt, P. 1984 A magyarországi miocén tengeri gerinces faunaszintek vázlata. Magyar Állami Földtani Intézet évi jelentése az 1982 évről: 347–353.
Kordos, L., Mészáros, I., and Szarvas, I. 2021. Tracking a “Prehistoric Pompeii,” Rhinoland and Crocodilia: New discoveries and interpretations of Ipolytarnóc (Hungary) Lower Miocene track site. Geoconservation Research 4: 621–634.
Kotsakis, T., Delfino, M., and Piras, P. 2004. Italian Cenozoic crocodilians: taxa, timing and palaeobiogeographic implications. Palaeogeography, Palaeoclimatology, Palaeoecology 210: 67–87.
Kováč, M., Andreyeva-Grigorovich, A., Bajraktarević, Z., Brzobohatý, R., Filipescu, S., Fodor, L., Harzhauser, M., Nagymarosy, A., Oszczypko, N., Pavelić, D., Rogl, F., Saftić, B., Sliva, Ľ., and Studencka, B. 2007. Badenian evolution of the Central Paratethys Sea: paleogeography, climate and eustatic sea-level changes. Geologica Carpathica 58: 579–606.
Kováč, M., Fordinál, K., Grigorovich, A.S.A., Halásová, E., Hudáčková, N., Joniak, P., Pipík, R., Sabol, M., Kováčová, M., and Sliva, Ľ. 2005. Západokarpatské fosílne ekosystémy a ich vzťah k paleoprostrediu v kontexte neogénneho vývoja eurázijského kontynentu. Geologické práce, Správy 111: 61–121.
Kováč, M., Halásová, E., Hudáčková, N., Holcová, K., Hyžný, M., Jamrich, M., and Ruman, A. 2018. Towards better correlation of the Central Paratethys regional time scale with the standard geological time scale of the Miocene Epoch. Geologica Carpathica 69: 283–300. Crossref
Kováč, M., Hudáčková, N., Halásová, E., Kováčová, M., Holcová, K., Oszczypko-Clowes, M., Báldi, K., Less, G., Nagymarosy, A., Ruman, A., Klučiar, T., and Jamrich, M. 2017. The Central Paratethys palaeoceanography: a water circulation model based on microfossil proxies, climate, and changes of depositional environment. Acta Geologica Slovaca 9: 75–114.
Kováčová, M., Doláková, N., and Kováč, M. 2011. Miocene vegetation pattern and climate change in the northwestern Central Paratethys domain (Czech and Slovak Republic). Geologica Carpathica 62: 251–266. Crossref
Kovalchuk, O., Nadachowski, A., Świdnicka, E., and Stefaniak, K. 2020. Fishes from the Miocene lacustrine sequence of Bełchatów (Poland). Historical Biology 32: 1011–1018. Crossref
Kowalewski, K. 1930. Stratygrafia miocenu okolic Korytnicy w porównaniu z trzeciorzędem pozostałych obszarów Gór Świętokrzyskich. Sprawozdania Państwowego Instytutu Geologicznego 6 (1): 1–211.
Kowalewski, K. 1958. Stratygrafia miocenu południowej Polski ze szczególnym uwzględnieniem południowego obrzeżenia Gór Świętokrzyskich. Geological Quarterly 2: 3–34.
Kowalski, K. 1993a. Microtocricetus molassicus Fahlbusch and Mayr, 1975 (Rodentia, Mammalia) from the Miocene of Bełchatów (Poland). Acta Zoologica Cracoviensia 36: 251–258.
Kowalski, K. 1993b. Neocometes Schaub and Zapfe, 1953 (Rodentia, Mammalia) from the Miocene of Bełchatów (Poland). Acta Zoologica Cracoviensia 36: 259–265.
Kowalski, K. 1994. Evolution of Anomalomys Gaillard, 1900 (Rodentia, Mammalia) in the Miocene of Poland. Acta Zoologica Cracoviensia 37: 163–176.
Kowalski, K. and Rzebik-Kowalska, B. 2002. Paleoecology of the Miocene fossil mammal fauna from Bełchatów (Poland). Acta Theriologica 47 (Supplement 1): 115–126. Crossref
Kowalski, K. and Kubiak, H. 1993. Gomphotherium angustidens (Cuvier, 1806) (Proboscidea, Mammalia) from the Miocene of Bełchatów and the Proboscidean Datum in Poland. Acta Zoologica Cracoviensia 36: 275–280.
Kowalski, R. 2017. Miocene carpological floras of the Konin region (Central Poland). Acta Palaeobotanica 57: 39–100. Crossref
Krstić, N., Savić, Lj., and Jovanović, G., 2012. The Neogene Lakes on the Balkan Land. Annales géologiques de la Peninsule Balkanique 73: 37–60. Crossref
Kunzmann, L., Kvaček, Z., Mai, D.H., and Walther, H. 2009. The genus Taxodium (Cupressaceae) in the Palaeogene and Neogene of Central Europe. Review of Palaeobotany and Palynology 153: 153–183. Crossref
Kvaček, Z. and Sakala, J. 1999. Twig with attached leaves, fruits and seeds of Decodon (Lythraceae) from the Lower Miocene of northern Bohemia, and implications for the identification of detached leaves and seeds. Review of Palaeobotany and Palynology 107: 201–222. Crossref
Laube, C.G. 1901. Synopsis der Wirbeltierfauna der böhmischen Braunkohlenformation und Beschreibung neuer, oder bisher unvollständig bekannter Arten. Abhandlungen des Deutschen Naturwissenschaftlich-Medizinischen Vereins für Böhmen “Lotos” 2 (4): 1–76.
Laube, C.G. 1910. Vogel- und Reptilienreste ans der Braunkohle von Skiritz bei Brüx. Lotos 58: 120–121.
Leder, R.M. 2012. Die Fauna der Lausitz im Miozän. In: R.M. Leder (ed.), Tropenparadies Lausitz? Klimawandel im Tertiär. Begleitband zur Sonderausstellung “Tropenparadies Lausitz? Klimawandel im Tertiär”, 152–177. Museum der Westlausitz, Kamenz.
LePage, B.A. 2007. The taxonomy and biogeographic history of Glyptostrobus Endlicher (Cupressaceae). Bulletin of the Peabody Museum of Natural History 48: 359–426. Crossref
Li, J. and Wang, B. 1987. A new species of Alligator from Shanwang, Shandong. Vertebrata PalAsiatica 25: 199–207.
Liebus, A. 1936. Krokodilreste aus den tertiären Tonen von Preschen. Lotos 84: 94–97.
Lindblad, K.T., Moreno-Bernal, J.W., McKellar, R.C., and Velez, M.I. 2022. The Northern Crocodile: first report of Borealosuchus (Eusuchia; Crocodylia) from Saskatchewan’s lower Ravenscrag Formation (earliest Paleocene) with implications for biogeography. Canadian Journal of Earth Sciences 59: 623–638. Crossref
Ludwig, R. 1857. Fossile Pflanzen aus der jüngsten Wetterauer Braunkohle. Palaeontographica 5 (3–4): 81–110.
Ludwig, R. 1877. Fossile Crocodiliden aus der Tertiärformation des Mainzer Beckens. Palaeontographica – Supplementbände 3 (4–5): 1–52.
Luján, À.H., Chroust, M., Čerňanský, A., Fortuny, J., Mazuch, M., and Ivanov, M. 2019. First record of Diplocynodon ratelii Pomel, 1847 from the early Miocene site of Tušimice (Most Basin, Northwest Bohemia, Czech Republic). Comptes Rendus Palevol 18: 877–889. Crossref
Macaluso, L., Martin, J.E., Favero, L.D., and Delfino, M. 2019. Revision of the crocodilians from the Oligocene of Monteviale, Italy, and the diversity of European eusuchians across the Eocene–Oligocene boundary. Journal of Vertebrate Paleontology 39: 1–13. Crossref
Macaluso, L., Villa, A., and Mörs, T. 2022. A new proteid salamander (Urodela, Proteidae) from the middle Miocene of Hambach (Germany) and implications for the evolution of the family. Palaeontology 65 (1): e12585.
Mach, K., Teodoridis, V., Grygar, T.M., Kvaček, Z., Suhr, P., and Standke, G. 2014. An evaluation of palaeogeography and palaeoecology in the Most Basin (Czech Republic) and Saxony (Germany) from the late Oligocene to the early Miocene. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 272: 13–45. Crossref
Mai, D.H. 1975. Beiträge zur Bestimmung und Nomenklatur fossiler Magnolien. Feddes Repertorium 86 (9–10): 559–578. Crossref
Mai, D.H. 2000. Die untermiozänen Floren aus der Spremberger Folge und dem 2. Flözhorizont in der Lausitz. IV: Fundstellen und Paläobiologie. Palaeontographica Abteilung B 254: 65–176. Crossref
Mannion, P.D., Benson, R.B.J., Carrano, M.T., Tennant, J.P., Judd, J., and Butler, R.J. 2015. Climate constrains the evolutionary history and biodiversity of crocodylians. Nature Communications 6: 1–9. Crossref
Marciszak, A. and Lipecki, G. 2020. The history of bears (Ursidae, Carnivora, Mammalia) from Silesia (southern Poland) and the neighbouring areas. Geological Quarterly 64: 876–897. Crossref
Markwick, P.J. 1994. “Equability,” continentality, and Tertiary “climate”: the crocodilian perspective. Geology 22: 613–616. Crossref
Markwick, P.J. 1998. Fossil crocodilians as indicators of Late Cretaceous and Cenozoic climates: implications for using palaeontological data in reconstructing palaeoclimate. Palaeogeography, Palaeoclimatology, Palaeoecology 137: 205–271. Crossref
Martin, J.E. 2010. A new species of Diplocynodon (Crocodylia, Alligatoroidea) from the Late Eocene of the Massif Central, France, and the evolution of the genus in the climatic context of the Late Palaeogene. Geological Magazine 147: 596–610. Crossref
Martin, J.E. and Gross, M. 2011. Taxonomic clarification of Diplocynodon Pomel, 1847 (Crocodilia) from the Miocene of Styria, Austria. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 261: 177–193. Crossref
Martini, E. 1977. Calcareous nannoplankton from the Korytnica basin (Middle Miocene; Holy Cross Mountains, Poland). Acta Geologica Polonica 27: 125–133.
Massonne, T. and Böhme, M. 2022. Re-evaluation of the morphology and phylogeny of Diplocynodon levantinicum Huene & Nikoloff, 1963 and the stratigraphic age of the West Maritsa coal field (Upper Thrace Basin, Bulgaria). PeerJ 10: e14167. Crossref
Mayr, G. and Poschmann, M. 2009. A loon leg (Aves, Gaviidae) with crocodilian tooth from the Late Oligocene of Germany. Waterbirds 32: 468–471. Crossref
Mazzotti, F.J., Best, G.R., Brandt, L.A., Cherkiss, M.S., Jeffery, B.M., and Rice, K.G. 2009. Alligators and crocodiles as indicators for restoration of Everglades ecosystems. Ecological Indicators 9 (6) Supplement: S137–S149. Crossref
Mazzotti, F.J., Brandt, L.A., Moler, P., and Cherkiss, M.S. 2007. American crocodile (Crocodylus acutus) in Florida: recommendations for endangered species recovery and ecosystem restoration. Journal of Herpetology 41: 122–132. Crossref
Mączyńska, S. 1993. Echinoids from the Pińczów Limestones (Middle Miocene; Holy Cross Mountains, Central Poland). Acta Geologica Polonica 43: 103–114.
Mehl, M.G. 1962. Missouri’s ice age animals. Educational series—Missouri Geological Survey and Water Resources 1: 1–104.
Menzel, P. 1913. Beitrage zur Flora der niederrheinischen Braunkohlenformation. Jahrbuch der Preussischen Geologischen Landesanstalt zu Berlin 46 (1): 1–98.
Mitsch, W.J. and Gosselink, J.G. 2015. Wetlands. Fifth Edition. 736 pp. John Wiley & Sons, Inc., Hoboken.
Młynarski, M. 1984. Notes on the amphibian and reptilian fauna of the Polish Miocene. Acta Zoologica Cracoviensia 27 (8): 127–148.
Mook, C.C. 1946. A new Pliocene alligator from Nebraska. American Museum Novitates 1311: 1–12.
Morlo, M., Schaal, S., Mayr, G., and Seiffert, C. 2004. An annotated taxonomic list of the Middle Eocene (MP11) Vertebrata of Messel. Courier Forschungsinstitut Senckenberg 252: 95–108.
Mörs, T. 2002. Biostratigraphy and paleoecology of continental Tertiary vertebrate faunas in the Lower Rhine Embayment (NW-Germany). Netherlands Journal of Geosciences/Geologie en Mijnbouw 81: 177–183. Crossref
Mörs, T., von der Hocht, F., and Wutzler, B. 2000. Die erste Wirbeltierfauna aus der miozänen Braunkohle der Niederrheinischen Bucht (Ville-Schichten, Tagebau Hambach). Paläontologische Zeitschrift 74 (1/2): 145–170. Crossref
Müller, P. 1996. Middle Miocene decapod Crustacea from southern Poland. Prace Muzeum Ziemi 43: 3–16.
Nadachowski, A. 2001. New important Neogene and Pleistocene mammal assemblages from Poland. Bollettino della Società Paleontologica Italiana 40: 243–248.
Neubauer, T.A., Georgopoulou, E., Kroh, A., Harzhauser, M, Mandic, O., and Esu, D. 2015. Synopsis of European Neogene freshwater gastropod localities: updated stratigraphy and geography. Palaeontologia Electronica 18.1.3T: 1–7. Crossref
Neubauer, T.A., Mandic, O., and Harzhauser, M. 2016. The early middle Miocene lacustrine gastropod fauna of Džepi, Bosnia and Herzegovina (Dinaride Lake System): high endemism in a small space. Bulletin of Geosciences 91: 271–296. Crossref
Nicholl, C.S.C., Rio, J.P., Mannion, P.D., and Delfino, M. 2020. A re-examination of the anatomy and systematics of the tomistomine crocodylians from the Miocene of Italy and Malta. Journal of Systematic Palaeontology 18: 1853–1889. Crossref
Nikitin, P.A. 1929. The systematic position of the fossil genus Diclidocarya. Journal of Botany 67: 33–38.
Nojima, K. and Itoigawa, J., 2017. Tomistominae gen. et sp. indet. (Crocodylia: Crocodylidae) from the Lower Yage Formation (Middle Pleistocene) in Hamamatsu City, Shizuoka Prefecture, Japan. Bulletin of the Mizunami Fossil Museum 43: 35–46.
Olson, E.C. 1940. A late Pleistocene fauna from Herculaneum, Missouri. The Journal of Geology 48: 32–57. Crossref
Pan, T., Wang, H., Duan, S., Ali, I., Yan, P., Cai, R., Wang, M., Zhang, J., Zhang, H., Zhang, B., and Wu, X. 2019. Historical population decline and habitat loss in a critically endangered species, the Chinese alligator (Alligator sinensis). Global Ecology and Conservation 20: e00692. Crossref
Pawelec, S. and Bielowicz, B. 2016. Petrographic composition of lignite from the Szczerców deposit, Polish Lowlands. Contemporary Trends in Geoscience 5 (2): 92–103. Crossref
Pawłowska, K. 1960. Szczątki ryb z wapieni mioceńskich Pińczowa. Acta Geologica Polonica 5: 421–432.
Perrin, C., Plaziat, J.-C., and Rosen, B.R. 1998. Miocene coral reefs and reef corals of the south-western Gulf of Suez and north-western Red Sea: distribution, diversity and regional environmental controls. In: B.H. Purser and D.W.J. Bosence (eds.), Sedimentation and Tectonics of Rift Basins: Red Sea—Gulf of Aden, 296–319. Springer, Dordrecht. Crossref
Peryt, D. 2013.Foraminiferal
record of the Middle Miocene climate transition prior to the Badenian
salinity crisis in the Polish Carpathian Foredeep Basin (Central
Paratethys). Geological Quarterly
57: 141–164. Crossref
Pezelj, Đ., Mandic, O., and Ćorić, S. 2013. Paleoenvironmental dynamics in the southern Pannonian Basin during initial Middle Miocene marine flooding. Geologica Carpathica 64: 81–100.
Piller, W.E. and Harzhauser, M. 2005. The myth of the brackish Sarmatian Sea. Terra Nova 17: 450–455. Crossref
Piller, W.E., Harzhauser, M., and Mandic, O. 2007. Miocene Central Paratethys stratigraphy—current status and future directions. Stratigraphy 4: 151–168. Crossref
Piñero, P., Agustí, J., Oms, O., Fierro, I., Montoya, P., Mansino, S., Ruiz-Sánchez, F., Alba, D.M., Alberdi, M.T., Blain, H.-A., Laplana, C., Van der Made, J., Mazo, A.V., Morales, J., Murelaga, X., Pérez-García, A., Pérez-Valera, F., Pérez-Valera, J.A., Sevilla, P., Soria, J.M., and Romero, G. 2017. Early Pliocene continental vertebrate fauna at Puerto de la Cadena (SE Spain) and its bearing on the marine-continental correlation of the Late Neogene of Eastern Betics. Palaeogeography, Palaeoclimatology, Palaeoecology 479: 102–114. Crossref
Pivko, D., Hudáčková, N., Hrabovský, J., Sládek, I., and Ruman, A., 2017. Palaeoecology and sedimentology of the Miocene marine and terrestrial deposits in the “Medieval Quarry” on Devínska Kobyla Hill (Vienna Basin). Geological Quarterly 61: 549–568. Crossref
Pomel, A. 1847. Note sur des animaux fossiles découverts dans le département de l’Allier. Bulletin de la Société géologique de France 4: 378–385.
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Shcherba, I.G., and Kováč, M. (eds.) 2004. Lithological-paleogeographic maps of Paratethys. 10 Maps Late Eocene to Pliocene. Courier Forschungsinstitut Senckenberg 250: 1–46.
Preston, R.E. 1979. Late Pleistocene cold-blooded vertebrate faunas from the mid-continental United States. I. Reptilia; Testudines, Crocodilia. Papers on Paleontology 19: 1–53.
Procházka, V.J. 1893. Miocaen židlochovický na Moravě a jeho zvířena. Rozpravy České akademie císaře Františka Josefa pro vědy, slovesnost a umění. Třída II Mathematicko-přírodnická 2 (24): 1–90.
Puértolas-Pascual, E., Blanco, A., Brochu, C.A., and Canudo, J.I. 2015. Review of the Late Cretaceous–early Paleogene crocodylomorphs of Europe: Extinction patterns across the K-Pg boundary. Cretaceous Research 57: 565–590. Crossref
Radwański, A. 1965. A contribution to the knowledge of Miocene Elasmobranchii from Pińczów (Poland). Acta Palaeontologica Polonica 10: 267–276.
Radwański, A. 1969. Transgresja dolnego tortonu na południowych stokach Gór Świętokrzyskich (strefa zatok i ich przedpola) (Lower Tortonian transgression onto the southern slopes of the Holy Cross Mts.) Acta Geologica Polonica 19: 1–164.
Radwański, A. 1973. Transgresja dolnego tortonu na południowo-wschodnich i wschodnich stokach Gór Świętokrzyskich. Acta Geologica Polonica 23: 375–434.
Radwański, A. 1974. Fore-Carpathian Depression. In: G. Larsen (ed.), Geologisk Ekskursion til Polen, September 1973, I. Kollokvie – Rapport, 94–132. Geologisk Institut, Aarhus Universitet, Aarhus.
Radwański, A. 1977. Neogen. In: H. Makowski (ed.), Geologia historyczna, 731–770. Wydawnictwa Geologiczne, Warszawa.
Radwański, A. 1985. Fore-Carpathian Depression. In: Z. Bełka, B.A. Matyja, and A. Radwański (eds.), Field-guide to the Geological Excursion to Poland, 79–110. Institute of Geology, University of Warsaw, Warsaw.
Radwański, A. 2015. Neogene. In: S. Skompski and A. Żylińska (eds.), The Holy Cross Mountains—25 Journeys Through Earth History, 27–29. Faculty of Geology, University of Warsaw, Warsaw.
Radwański, A., Górka, M., and Wysocka, A. 2006. Middle Miocene coralgal facies at Maksymivka near Ternopil (Ukraine): A preliminary account. Acta Geologica Polonica 56: 89–103.
Rajchl, M., Uličný, D., Grygar, R., and Mach, K. 2009. Evolution of basin architecture in an incipient continental rift: the Cenozoic Most Basin, Eger Graben (Central Europe). Basin Research 2: 269–294. Crossref
Redlich, K.A. 1902. Wirbelthierreste aus der böhmischen Braunkohlenformation. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt 52: 135–140.
Reid, C. and Reid, E.M. 1908. On Dulichium vespiforme sp. nov. from the brick-earth of Tegelen. Proceedings of the Section of Science Koninklijke Akademie van Wetenschappen te Amsterdam 10 (2): 860–861.
Reid, E.M. 1920. Recherches sur quelques graines pliocènes de Pont-de-Gail (Cantal). Bulletin de la Société Géologique de France, Quatrième série 20 (1–3): 48–87
Ricou, L.-E. 1994. Tethys reconstructed: plates, continental fragments and their boundaries since 260 Ma from Central America to South-eastern Asia. Geodinamica Acta 7 (4): 169–218. Crossref
Riegl, B. and Piller, W.E. 2000. Biostromal coral facies—a Miocene example from the Leitha Limestone (Austria) and its actualistic interpretation. Palaios 15: 399–413. Crossref
Rio, J.P. and Mannion, P.D. 2021. Phylogenetic analysis of a new morphological dataset elucidates the evolutionary history of Crocodylia and resolves the long-standing gharial problem. PeerJ 9: e12094. Crossref
Rio, J.P., Mannion, P.D., Tschopp, E., Jeremy, E., and Delfino, M. 2020. Reappraisal of the morphology and phylogenetic relationships of the alligatoroid crocodylian Diplocynodon hantoniensis from the late Eocene of the United Kingdom. Zoological Journal of the Linnean Society 188: 579–629. Crossref
Roemer, F. 1870. Geologie von Oberschlesien. 587 pp. Nischkowsky, Breslau.
Rossmann, T., Berg, D.E., and Salisbury, S. 1999. Studies on Cenozoic crocodiles: 3. Gavialosuchus cf. gaudensis (Eusuchia: Tomistomidae) from the Lower Miocene of south Germany. Neues Jahrbuch für Geologie und Paläontologie. Monatshefte 1999 (6): 321–330. Crossref
Rögl, F. and Steininger, F.F. 1983. Vom Zerfall der Tethys zu Mediterran und Paratethys. Die neogene Paläogeographie und Palinspastik des zirkum-mediterranen Raumes. Annalen des Naturhistorischen Museums in Wien 85/A: 135–163.
Rzehak, A. 1880. Ueber die Gliederung und Verbreitung der älteren Mediterranstufe in der Umgebung von Gr. Seelowitz in Mähren. Verhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 16, 300–303.
Rzehak A. 1912. Beitrag zur Kenntnis der Oncophoraschichten Mährens. Verhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 15, 344–347.
Sabău, I., Venczel, M., Codrea, V.A., and Bordeianu, M. 2021. Diplocynodon: a salt water Eocene crocodile from Transylvania? North-Western Journal of Zoology 17: 117–121.
Sach, V.J. 2016. Fossilienkatalog der Miozän-Molasse in Südwestdeutschland (Obere Süßwassermolasse, Brackwassermolasse, Obere Meeresmolasse, Untere Süßwassermolasse). Documenta Naturae SB 70: 1–115.
Sant, K., Mandic, O., Rundić, L., Kuiper, K.F., and Krijgsman, W. 2018. Age and evolution of the Serbian Lake System: integrated results from Middle Miocene Lake Popovac. Newsletters on Stratigraphy 51: 117–143. Crossref
Scheiner, F., Havelcová, M., Holcová, K., Doláková, N., Nehyba, S., Ackerman, L., Trubač, J., Hladilová, Š., Rejšek, J., and Utescher, T. 2023. Evolution of palaeoclimate, palaeoenvironment and vegetation in Central Europe during the Miocene Climate Optimum. Palaeogeography, Palaeoclimatology, Palaeoecology 611: 111364. Crossref
Scheyer, T.M., Aguilera, O.A., Delfino, M., Fortier, D.C., Carlini, A.A., Sánchez, R., Carrillo-Briceño, J.D., Quiroz, L., and Sánchez-Villagra, M.R. 2013. Crocodylian diversity peak and extinction in the late Cenozoic of the northern Neotropics. Nature Communications 4: 1–9. Crossref
Schleich, H.H. 1994. Neue Reptilfunde aus dem Tertiär Deutschlands. 13. Schildkröten und Krokodilreste aus der eozänen Braunkohle des Untertagebaues Stolzenbach bei Borken (Hessen). Courier Forschungsinstitut Senckenberg 173: 79–101.
Schleich, H.H. 1988. Paläoherpetologische Materialien und Faunenspektren aus dem Kalktertiär des Mainzer Beckens (Oberoligozän-Untermiozän). Geologisches Jahrbuch A 110: 289–306.
Schlögl, J. and Holec, P. 2004. Crocodile remains from the Middle Miocene (late Badenian) of the Vienna Basin (Sandberg, Western Slovakia). Slovak Geological Magazine 10: 293–297.
Sebe, K., Szabó, M., Szentesi, Z., Pandolfi, L., Jankó, N., and Magyar, I. 2024. Fossils from the Upper Miocene (Pannonian) sands of the Pécsvárad sand pit (Eastern Mecsek Mts., SW Hungary). Geoloski anali Balkanskoga poluostrva 85: 49–71. Crossref
Seko, M., Pipík, R., and Doláková, N. 2012. Early Badenian ostracod assemblage of the stratotype Židlochovice (Carpathian Foredeep, Czech Republic). Central European Journal of Geosciences 4: 111–125. Crossref
Smith, R. 2003. Les vertébrés terrestres de l’Oligocène inférieur de Belgique (Formation de Borgloon, MP 21): inventaire et interprétation des données actuelles. Coloquios de Paleontología Extra 1: 647–657.
Staniewicz, A., Behler, N., Dharmasyah, S., and Jones, G. 2018. Niche partitioning between juvenile sympatric crocodilians in Mesangat Lake, East Kalimantan, Indonesia. Raffles Bulletin of Zoology 66: 528–537.
Stefanović, I. and Mijovic, D. 2004. The Middle Miocene fossil site Prebreza in southern Serbia. Polish Geological Institute Special Papers 13: 209–212.
Stevenson, C. 2019. Crocodiles of the World: A Complete Guide to Alligators, Caimans, Crocodiles and Gharials. 288 pp. New Holland Publishers, London.
Stout, J.B. 2024. Osmoregulation in Alligatoroidea: shifting the paradigm untethers biogeographic questions. Historical Biology, 1–6 [available online, https://doi.org/10.1080/08912963.2024.2379029]. Crossref
Stuchlik, L., Szynkiewicz, A., Łańcucka-Środoniowa, M., and Zastawniak, E. 1990. Wyniki dotychczasowych badań paleobotanicznych trzeciorzędowych węgli brunatnych złoża “Bełchatów”. Acta Palaeobotanica 30: 259–305.
Studencka, B. 1999. Remarks on Miocene bivalve zonation in the Polish part of the Carpathian Foredeep. Geological Quarterly 43: 467–477.
Studencka, B. and Jasionowski, M. 2011. Bivalves from the Middle Miocene reefs of Poland and Ukraine: A new approach to Badenian/Sarmatian boundary in the Paratethys. Acta Geologica Polonica 61: 79–114.
Studencka, B. and Studencki, W. 1988. Middle Miocene (Badenian) bivalves from the carbonate deposits of the Wójcza-Pińczów Range (southern slopes of the Holy Cross Mountains, Central Poland). Acta Geologica Polonica 38: 1–44.
Studencka, B., Gontsharova, I.A., and Popov, S.V. 1998. The bivalve fauna as a basis for reconstruction of the Middle Miocene history of the Paratethys. Acta Geologica Polonica 48: 285–342.
Studencki, W. 1979. Sedimentation of algal limestones from Busko-Spa environs (Middle Miocene, Central Poland). Palaeogeography, Palaeoclimatology, Palaeoecology 27: 155–165. Crossref
Studencki, W. 1988a. Facies and sedimentary environment of the Pinczow Limestones (Middle Miocene; Holy Cross Mountains, Central Poland). Facies 18: 1–26. Crossref
Studencki, W. 1988b. Red algae from the Pińczów Limestones (Middle Miocene; Świętokrzyskie Mountains, Central Poland). Acta Palaeontologica Polonica 33: 3–57.
Szynkiewicz, A. 2000. Age of the brown coal deposits from the Bełchatów lignite mine (Central Poland). Przegląd Geologiczny 48 (11): 1038–1044.
Taplin, L.E. and Grigg, G.C. 1989. Historical zoogeography of the eusuchian crocodilians: a physiological perspective. American Zoologist 29: 885–901. Crossref
Taplin, L.E. and Grigg, G.C. 1993. Morphology and physiology of the Crocodylia. In: C.G. Glasby, J. Ross, and P.L. Beesley (eds.), Fauna of Australia Vol. 2A: Amphibia and Reptilia, 326–336. AGPS Canberra, Canberra.
Taplin, L.E., Grigg, G.C., Harlow, P., Ellis, T.M., and Dunson, W.A. 1982. Lingual salt glands in Crocodylus acutus and C. johnstoni and their absence from Alligator mississipiensis and Caiman crocodilus. Journal of Comparative Physiology 149 (1): 43–47.
Thorbjarnarson, J. and Wang, X. 1999. The conservation status of the Chinese alligator. Oryx 33: 152–159. Crossref
Thorbjarnarson, J., Wang, X., Ming, S., He, L., Ding, Y., Wu, Y., and McMurry, S.T. 2002. Wild populations of the Chinese alligator approach extinction. Biological Conservation 103: 93–102. Crossref
Toula, F. and Kail, J.A. 1885. Über einen Krokodil-Schädel aus den Tertiärablagerungen von Eggenburg in Niederösterreich: eine paläontologische Studie. Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschatflliche Klasse 50: 299–335.
Van der Made, J. and Kowalski, K. 1996. Suidae (Artiodactyla, Mammalia) from the Miocene of Bełchatów in Poland. Acta Zoologica Cracoviensia 39: 305–310.
van Hinsbergen, D.J.J., de Groot, L.V., van Schaik, S.J., Spakman, W., Bijl, P.K., Sluijs A, Langereis, C.G., and Brinkhuis, H. 2015. A Paleolatitude calculator for paleoclimate studies. PLoS ONE 10 (6): e0126946. Crossref
Vasilyan, D. 2020. Fish, amphibian and reptilian assemblage from the middle Miocene locality Gračanica-Bugojno palaeolake, Bosnia and Herzegovina. Palaeobiodiversity and Palaeoenvironments 100: 437–455. Crossref
Venczel, M. 2007. Late Middle Miocene amphibians and reptiles from Subpiatrǎ (Bihor district, Romania). Nymphaea, Folia naturae Bihariae 34: 39–66.
Venczel, M. 2023. Updating the fossil record of the alligatoroid crocodylian Diplocynodon from the late Eocene of Transylvanian Basin. Frontiers in Amphibian and Reptile Science 1: 1217025. Crossref
Venczel, M. and Codrea, V.A. 2022. A new late Eocene alligatoroid crocodyliform from Transylvania. Comptes Rendus Palevol 21: 411–429. Crossref
Venczel, M., Codrea, V.A., and Trif, N. 2023. Eocene gavialoid teeth from southern Transylvania with notes on the diversity of Paleogene crocodilians from Romania. North-Western Journal of Zoology 19: 71–79.
Vernyhorova, Y.V., Holcová, K., Doláková, N., Reichenbacher, B., Scheiner, F., Ackerman, L., Rejšek, J., Bortoli, L.D., Trubač, J., and Utescher, T. 2023. The Miocene Climatic Optimum at the interface of epicontinental sea and large continent: A case study from the Middle Miocene of the Eastern Paratethys. Marine Micropaleontology 181: 102231. Crossref
Vianna, A. and Moraes, A. 1942. Sôbre um Crâneo de Crocodilo Fóssil encontrado no Mioceno de Lisboa. Boletim da Sociedade Portuguesa de Ciências Naturais 13 (Suplemento 3): 542–546.
Villa, M., Carnevale, G., Pavia, M., Rook, L., Sami, M., Szyndlar, Z., and Delfino, M. 2021. An overview on the late Miocene vertebrates from the fissure fillings of Monticino Quarry (Brisighella, Italy), with new data on non-mammalian taxa. Rivista Italiana di Paleontologia e Stratigrafia 127: 297–354.
Wagner, M. and Matl, K. 2007. Stratigraphy of lacustrine chalk from the “Szczerców” lignite deposit. Geologia 33: 289–315.
Webb, G., Manolis, S., and Sack, G. 1983. Crocodylus johnstoni and C. porosus coexisting in a tidal river. Wildlife Research 10: 639–650. Crossref
Westerhold, T., Marwan, N., Drury, A.J., Liebrand, D., Agnini, C., Anagnostou, E., Barnet, J.S.K., Bohaty, S.M., Vleeschouwer, D.D., Florindo, F., Frederichs, T., Hodell, D.A., Holbourn, A.E., Kroon, D., Lauretano, V., Littler, K., Lourens, L.J., Lyle, M., Pälike, H., Röhl, U., Tian, J., Wilkens, R.H., Wilson, P.A., and Zachos, J.C. 2020. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 369: 1383–1387. Crossref
Weydt, L.M., Agemar, T., Bär, K., Erb, M., Dobrzinski, N., Mantei, N., Moeck, I., Singh, M., Sperlich, S., van der Vaart, J., Weber, J and Sass, I. 2023. Assessment for medium-depth geothermal energy utilization in Germany—first results from the ArtemIS project. Geothermal Resources Council Transactions 47: 1640–1650.
Whiting, E.T. and Head, J.J. 2020. Neogene crocodylians from the Central Great Plains: spatiotemporal histories and relationships with regional climatic and environmental changes. Journal of Vertebrate Paleontology 40 (1): e1764007. Crossref
Whiting, E.T., Steadman, D.W., and Vliet, K.A. 2016. Cranial polymorphism and systematics of Miocene and living Alligator in North America. Journal of Herpetology 50: 306–315. Crossref
Wilson, O.E. and Parker, A.K. 2023. Low predator competition indicates occupation of macro-predatory niches by giant Miocene reptiles at La Venta, Colombia. Palaeogeography, Palaeoclimatology, Palaeoecology 632: 111843. Crossref
Wolff, E.D.S., Fowler, D.W., and Bonde, J.W. 2007. A possible case of necrotizing dermatitis in the crocodylian Diplocynodon, from the Oligocene of the Isle of Wight, United Kingdom. Historical Biology 19: 203–207. Crossref
Worobiec, G., Worobiec, E., and Szynkiewicz, A. 2012. Plant assemblage from the Upper Miocene deposits of the Bełchatów Lignite Mine (Central Poland). Acta Palaeobotanica 52: 369–413.
Worobiec, E. and Worobiec, G. 2022. Palynoflora and palaeoenvironment of the early Miocene palaeolake from the Bełchatów mine, central Poland. Geological Quarterly 66 (4): 32. Crossref
Worobiec, G. and Worobiec, E. 2019. Wetland vegetation from the Miocene deposits of the Bełchatów Lignite Mine (Central Poland). Palaeontologia Electronica 22.3.63: 1–38. Crossref
Wu, X.C., Brinkman, D.B., Fox, R.C., 2001: A new crocodylian (Archosauria) from the basal Paleocene of the Red Deer River Valley, southern Alberta. Canadian Journal of Earth Sciences 38: 1689–1704. Crossref
Wysocka, A., Radwański, A., Górka, M., Bąbel, M., Radwańska, U., and Złotnik, M. 2016. The Middle Miocene of the Fore-Carpathian Basin (Poland, Ukraine and Moldova). Acta Geologica Polonica 66: 351–401. Crossref
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K. 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292: 686–693. Crossref
Zachos, J.C., Dickens, G.R., and Zeebe, R.E. 2008. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 451: 279–283. Crossref
Zapfe, H. 1984. Krokodile im Mittelmiozän des Wiener Beckens. Sitzungsberichte der Akademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse 193: 161–169.
Zapfe, H. 1989. Pongidenzähne (Primates) aus dem Pontien von Götzendorf, Niederösterreich. Sitzungsberichte der Akademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse 197: 423–450.
Zoboli, D., Sanciu, L., Pillola, G.L., and Delfino, M. 2019. An overview of the crocodylian fossil record from Sardinia (Italy). Annales de Paléontologie 105: 123–137. Crossref
Acta Palaeontol. Pol. 70 (2): 225–251, 2025
https://doi.org/10.4202/app.01194.2024