
The first description of dinosaurian eggshell from the Maastrichtian Lance Formation, Wyoming, North America
TOM T.P. VAN DER LINDEN, JONATHAN J.W. WALLAARD, MAARTEN DE RIJKE, and RENÉ H.B. FRAAIJE
Van der Linden, T.T.P., Wallaard, J.J.W., de Rijke, M., and Fraaije, R.H.B. 2025. The first description of dinosaurian eggshell from the Maastrichtian Lance Formation, Wyoming, North America. Acta Palaeontologica Polonica 70 (3): 573–579.
Although numerous fossils have been excavated and described from the Upper Cretaceous (Maastrichtian) Lance Formation, eggs and eggshell remains are rare and have yet to be described in detail. Here, we provide the first description of an eggshell found in the Lance Formation in eastern Wyoming. The eggshell can be attributed to the oofamily Ovaloolithidae, and is most comparable to the oogenus Ovaloolithus based on the smooth outer surface, the eggshell thickness, the closely packed shell units, and the presence of two layers. The eggshell can tentatively be ascribed to either ornithopod or non-avian theropod dinosaurs. This new eggshell expands our understanding of the geographical distribution of ovaloolithid ootaxa throughout the uppermost Cretaceous of North America, having been previously described from the Maastrichtian North Horn Formation of Utah.
Key words: Ovaloolithidae, Dinosauria, eggs, Maastrichtian, Lance Formation, Wyoming.
Tom T.P. van der Linden [tppaleo@gmail.com; ORCID: https://orcid.org/0000-0002-7898-3292 ], Jonathan J.W. Wallaard [curator@oertijdmuseum.nl; ORCID: https://orcid.org/0000-0002-7752-1598 ], Maarten de Rijke [maarten@oertijdmuseum.nl; ORCID: https://orcid.org/0000-0003-4652-3782 ], and René H.B. Fraaije [directie@oertijdmuseum.nl; ORCID: https://orcid.org/0000-0002-3465-1093 ], Oertijdmuseum, Bosscheweg 80, 5283 WB, Boxtel, The Netherlands.
Received 23 September 2024, accepted 9 July 2025, published online 18 September 2025.
Copyright © 2025 T.P. van der Linden et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Dinosaur eggs and eggshell remains are important body fossils (e.g., Bertling et al. 2006, 2022; Lawver and Jackson 2014), as they provide information on reproductive behavior (e.g., Sellés and Galobart 2016), nesting conditions (e.g., Tanaka et al. 2018), and nesting environments of dinosaurs (He et al. 2019, 2020). In addition, dinosaur eggs inform on the faunal biodiversity of past ecosystems, especially when skeletal remains are lacking (e.g., Tanaka et al. 2016; Uematsu et al. 2023). On the North American continent, fossiliferous Cretaceous deposits are common, including formations preserving egg and eggshell material (e.g., Carpenter 1982; Hirsch and Quinn 1990; Zelenitsky and Hills 1997; Bray 1999; Zelenitsky and Therrien 2008; Jackson and Varricchio 2010, 2016, 2017; Zelenitsky et al. 2017a, b; Voris et al. 2018).
However, two Upper Cretaceous North American strata, the Hell Creek Formation and the Lance Formation, have yielded few egg remains (Carpenter 1982; Jackson and Varricchio 2016). Both the Hell Creek and the Lance formations have produced numerous skeletal remains (e.g., Lehman et al. 2006; Brown et al. 2022; Ponstein et al. 2024), however, temporally equivalent formations, such as the Willow Creek and St. Mary River formations in Canada, have yielded both skeletal and oological remains (Brown et al. 2015; Zelenitsky et al. 2017a; Voris et al. 2018). The lack of egg remains in the Hell Creek and Lance formations has been attributed to soil drainage and soil pH (Carpenter 1982). Relatively recently, the first oological remains from the Hell Creek Formation have been described (Jackson and Varricchio 2016), but eggshell remains from the Lance Formation remain undescribed. Here, we provide the first description of an eggshell fragment from the Lance Formation of Wyoming.
Institutional abbreviation.—MAB, Oertijdmuseum, Boxtel, the Netherlands.
Other abbreviations.—CL, continuous layer; ML, mammillary layer.
Material and methods
The eggshell (MAB16636) was recovered by RHBF (4th author) in 2022 during sieving of a prolific microsite in the Lance Formation located near Lusk in eastern Wyoming, USA (Fig. 1). The exact location remains undisclosed due to the vulnerability of the site. Detailed locality information can be requested from the authors. This microsite has yielded remains of mammals, fish, turtles, crocodiles, and dinosaurs. A single radial section of MAB16636 was made, which was subsequently scanned using a Zeiss Axio Scan.Z1 v1.0, using 10× and 20× objective lenses. An Axio Scan can be used for automatic imaging to create a virtual slide of histological sections of fossil eggshell. Zeiss Zen Lite (Blue edition) version 3.2 software was used to subsequently photograph and measure the eggshell. Scanning electron micrographs were made with a JEOL Neoscope JCM-6000 tabletop SEM and a ZEISS EVO 15 SEM.
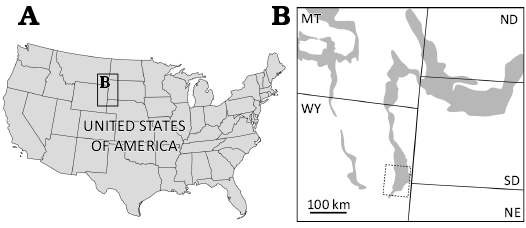
Fig. 1. Map of United States of America (A) with locality of the eggshell in eastern Wyoming (B). Lance Formation outcrops indicated in grey.
Systematic palaeontology
Oofamily Ovaloolithidae Mikhailov, 1991
Oogenus Ovaloolithus Zhao, 1979
Type species: Ovaloolithus chinkangkouensis Zhao & Jiang, 1974 amended Zhao, 1979. Laiyang Basin, China; Jingangkou Formation, Coniacian–Campanian.
Ovaloolithus oosp.
Figs. 2, 3.
Material.—The eggshell (MAB16636) from the Upper Cretaceous, Lance Formation, USA, eastern Wyoming, near Lusk.
Description.—The eggshell fragment is small (<1 cm2), with a smooth outer surface (Fig. 2A). The minimum thickness of the eggshell is 1.1 mm, as the fragment has suffered minor abrasion, particularly on the inner surface. The eggshell consists of two layers: a mammillary and a continuous (Fig. 2B1). The ML extends to about half the thickness of the eggshell, with a ratio of 1:1 between the ML and CL (Fig. 2B1). A relatively distinct contact between the ML and CL is accentuated by a brown horizontal line (Fig. 2B1). The ML consists of closely packed shell units (Fig. 3A1, A4) that are wedge-like with a radial-tabular ultrastructure. Only the ML shows vertical boundaries between adjacent shell units—these are lost in the CL such that the shell units become indistinguishable from each other. The CL shows considerable brown horizontal lineation. The SEM microphotographs (Fig. 3A5) show cleavage of the eggshell in some parts where the brown lineation is visible under normal and polarized light. Faint horizontal accretion lines are visible in both the ML and CL (Fig. 2). Part of a pore can be seen in radial section (Fig. 2B1). The pore is circular, and appears oriented obliquely. However, SEM microphotographs (Fig. 3A3) confirm that the pores in MAB16636 are vertically oriented, with slightly undular canal walls, possibly affected by infill.

Fig. 2. Eggshell MAB16636 from the Maastrichtian Lance Formation, near Lusk, eastern Wyoming, USA. A. Outer (A1) and inner (A2) surface morphology. B. Radial section of eggshell, in normal (B1) and polarized (B2) light. Note the pore (white arrow) near the transition between the mammillary layer (ML) and continuous layer (CL).
Remarks.—The abovementioned characteristics correspond to the description of the angustispherulitic morphotype, and in combination with the smooth outer surface and vertical pores, MAB16636 can be placed within Ovaloolithidae (Mikhailov 1991, 1997). Two oogenera are known in this oofamily: Ovaloolithus Zhao, 1979, and Minioolithus Wu et al., 2024. The former oogenus contains twelve oospecies, we herein disregard ?Ovaloolithus weiqiaoensis Yu, 1998, and ?Ovaloolithus huangtulingensis Yu, 1998, due to their insufficient description and uncertain affinities (Zhao et al. 2015), distributed in the Upper Cretaceous outcrops of China, Mongolia, and USA. Minioolithus is a recently erected oogenus from the Tangbian Formation (Santonian–Campanian) of China (Wu et al. 2024). MAB16636 can easily be distinguished from Minioolithus, as this ootaxon shows distinct ornamentation and is very thin for an ovaloolithid oospecies (0.13–0.35 mm). The characteristics of MAB16636, therefore, better fit Ovaloolithus sensu Mikhailov (1997).
Two oospecies of Ovaloolithus are known from USA: Ovaloolithus utahensis Bray, 1999, and Ovaloolithus tenuisus Bray, 1999. Both are from the Maastrichtian North Horn Formation of Utah. MAB16636 can be distinguished from both Ovaloolithus utahensis (1.42–1.68 mm) and Ovaloolithus tenuisus (0.43–0.58 mm) based on eggshell thickness. Similarly, MAB16636 can be distinguished from the following Asian oospecies due to eggshell thickness: Ovaloolithus chinkangkouensis (Zhao & Jiang, 1974) (2.60 mm in Zhao et al. 2015), Ovaloolithus tristriatus (Zhao & Jiang, 1974) (2.40 mm in Zhao et al. 2015), Ovaloolithus mixtistriatus (Zhao & Jiang, 1974) (2.50 mm in Zhao et al. 2015), Ovaloolithus monostriatus (Zhao & Jiang, 1974) (1.50 mm in Zhao et al. 2015), Ovaloolithus (Pinnatoolithus) shitangensis (Fang et al., 2009) (1.80–2.60 mm in Zhao et al. 2015), Ovaloolithus (Pinnatoolithus) nanxiongensis (Fang et al., 2009) (1.70 mm in Zhao et al. 2015), Ovaloolithus (Pinnatoolithus) sangequanensis (Fang et al., 2009) (3.00 mm in Zhao et al. 2015), and Ovaloolithus turpanensis Zhang & Wang, 2010 (1.88 mm in Zhang and Wang 2010). Only two ovaloolithid oospecies show overlap with MAB16636 in eggshell thickness: Ovaloolithus laminadermus (Zhao & Jiang, 1974) (0.60–1.20 mm in Zhao and Jiang 1974; 0.90 mm in Zhao et al. 2015) and Ovaloolithus dinornithoides Mikhailov, 1994 (1.10–1.80 mm in Mikhailov 1994). However, direct comparisons of MAB16636 with other ovaloolithid oospecies solely based on eggshell thickness are problematic and exemplified by Mikhailov (1994), who points out that for Ovaloolithus dinornithoides, the full eggshell range is 1.1–1.8 mm, but that the standard ranges are 1.5–1.8 mm and 1.3–1.6 mm, respectively. For Ovaloolithus chinkangkouensis, Mikhailov (1994) adds similar variation in eggshell thickness ranges, as the standard range for this oospecies is 2.2–3.0 mm, whereas the full range within a single egg is 1.4–3.0 mm. Therefore, for most ovaloolithid oospecies, the full ranges of eggshell thickness remain unexplored and difficult to compare to MAB16636. In addition, MAB16636 only represents a small fraction of the full egg. A thickness range cannot be established for this specimen.
MAB16636 can be further distinguished from all Asian ovaloolithid oospecies based on the lack of ornamentation (Young 1954; Zhao and Jiang 1974; Mikhailov 1994; Fang et al. 2009; Zhang and Wang 2010; Zhao et al. 2015), but Wu et al. (2024: table 1) report for most ovaloolithid oospecies that the ornamentation is unknown. However, Young (1954), Zhao and Jiang (1974), Mikhailov (1994), and the radial sections in Zhao et al. (2015) clarify the presence of weak to strong nodose/sagenotuberculate ornamentation in the majority of ovaloolithid oospecies. An exception appears to be Ovaloolithus (Pinnatoolithus) nanxiongensis, which Fang et al. (2009) report to be relatively smooth and not very undulating in thin section. However, we would argue that the description does not match the figured thin section by Fang et al. (2009), which shows a significantly more undulating outer surface than MAB16636, Ovaloolithus utahensis, or Ovaloolithus tenuisus (Bray 1999). Additionally, Ovaloolithus dinornithoides consists of a complete egg that shows variation across the eggshell surface, with the majority of eggshell not expressing any ornamentation (Mikhailov 1994). Significant variation in the presence or absence of ornamentation is therefore to be expected within Ovaloolithidae.
Most ovaloolithid oospecies, including MAB16636, show a ratio of 1:1 between the ML and CL. Exceptions include Ovaloolithus (Pinnatoolithus) shitangensis (2:5, Fang et al. 2009), Ovaloolithus (Pinnatoolithus) nanxiongensis (1:3, Fang et al. 2009), Ovaloolithus (Pinnatoolithus) sangequanensis (1:3, Fang et al. 2009), and Ovaloolithus turpanensis (1:3, Zhang and Wang 2010). The variation in this ratio throughout the entire eggshell, however, remains unexplored for this oofamily.
Although weakly ornamented, Ovaloolithus laminadermus and Ovaloolithus dinornithoides are most comparable to MAB16636. Comparisons are hindered by the limited description of the ultrastructure of most ovaloolithid oospecies, as they are mainly differentiated by the horizontal lineation present throughout the eggshell. Wang et al. (2022) determined that this lineation is unsuitable for distinguishing ovaloolithid oospecies. These lines are numerous in MAB16636, and at least some are the result of weathering based on the SEM images (Fig. 3A5), although the majority of the lineation may in fact be thermally matured organic matter (Choi et al. 2021, 2025). If these lines are dependent on preservation and variable within a single egg or clutch (Wang et al. 2022), then the remaining characteristics of most ovaloolithid oospecies overlap (see Wu et al. 2024: table 1). Therefore, we propose that this oofamily is in need of revision, in accordance with Wang et al. (2022).
We argue that MAB16636 cannot be assigned to a specific oospecies within Ovaloolithidae, as (i) thickness ranges within the oofamily have not been thoroughly explored, (ii) ornamentation can vary significantly within a single egg, and (iii) diagnostic lineation may depend on preservation and vary considerably, leading to considerable variation within this oofamily that complicates oospecies assignment. MAB16636 consists of only a single eggshell fragment, which we assign to Ovaloolithus oosp., and not to a specific oospecies within Ovaloolithidae.
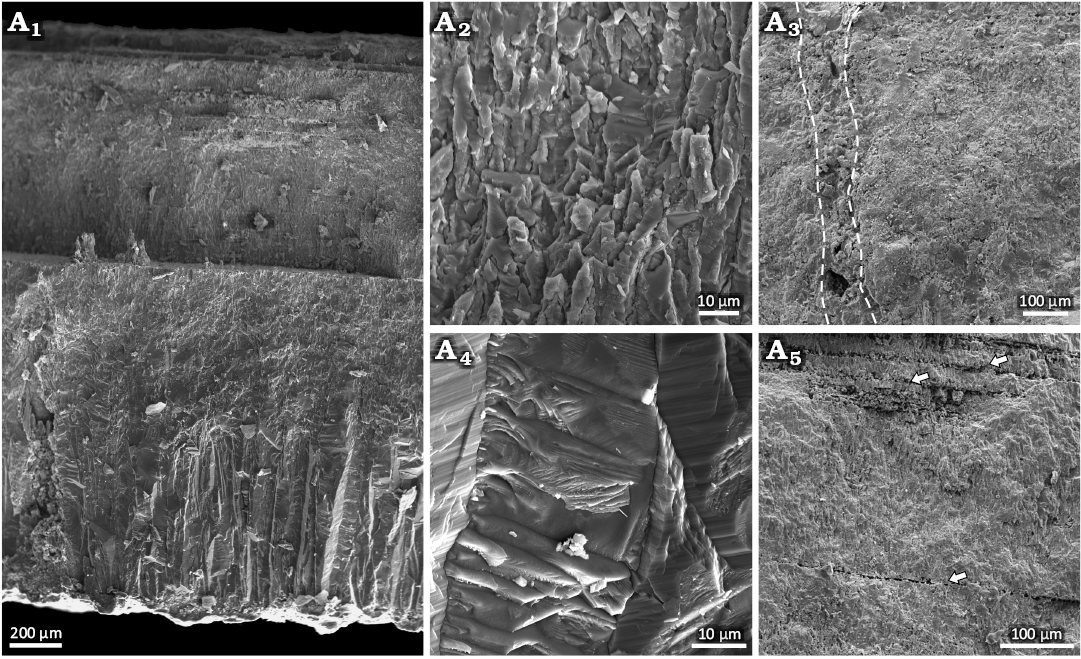
Fig. 3. SEM microphotographs of eggshell (MAB16636) from the Maastrichtian Lance Formation, near Lusk, eastern Wyoming, USA, including an overview of the eggshell (A1); A2, close-up of the continuous layer (CL); A3, the CL and a pore highlighted by a dashed line; A4, the wedged structure of the mammillary layer (ML); A5, the cleaved layers near the outer surface, highlighted by arrows. Note that in A1, one of the cleavage planes caused an uneven break, such that part of the middle of the eggshell became exposed.
Discussion
Late-stage embryonic remains are the primary and most trustworthy source for inferring links between parataxonomy and taxonomy. Few dinosaurian taxa have been confidently attributed to ootaxa (Choi et al. 2022). There are no ovaloolithid eggs known preserving embryonic remains, such that an unambiguous link with a dinosaur taxon cannot be made. Mikhailov (1997) describes ovaloolithid eggs as most reminiscent of spheroolithid eggs, that are ascribed to ornithopod dinosaurs (Moreno-Azanza et al. 2014; Van der Linden et al. 2024), with the former similarly interpreted as ornithopod eggs. This is further supported by several stratigraphic layers in Mongolia which yield only ovaloolithid eggs and lambeosaurine ornithopods (Mikhailov 1997). However, lambeosaurine hadrosaurs are effectively unknown from the Lance Formation (see Wegwesier et al. 2003 and Rolleri et al. 2020 for possible exceptions) or the contemporaneous Hell Creek Formation. Therefore, ovaloolithid eggs could be, similar to spheroolithid eggs, taxonomically more inclusive, and thus may encompass all of Ornithopoda, or even Ornithischia. The latter suggestion may be supported by Sochava (1972), who describes ovaloolithid eggshell in association with embryonic metatarsals of a ceratopsian dinosaur similar to Leptoceratops or Protoceratops. However, recent description of a Protoceratops clutch revealed that ceratopsian eggs consist of non-biomineralized, soft eggshell (Norell et al. 2020).
Recently, Wu et al. (2024) argued for a different taxonomic link, and instead proposed a non-avian theropod affinity for Ovaloolithidae based on the eggshell characteristics and a novel phylogenetic analysis of dinosaur eggs. Non-avian theropod remains are common in the Lance Formation (Brown et al. 2022). Caenagnathids (Pu et al. 2017), alvarezsaurids (Agnolin et al. 2012), dromaeosaurids (Grellet-Tinner and Makovicky 2006), and troodontids (Varricchio et al. 2002) are known to have produced significantly different eggs from Ovaloolithidae. No eggs unambiguously attributed to ornithomimosaurs or tyrannosaurs are known, such that ovaloolithid eggs may be parataxonomically linked to either group. Due to the lack of embryonic remains associated with ovaloolithid eggs, MAB16636 is herein tentatively referred to either ornithopods or non-avian theropods until unambiguous evidence is presented.
The ambiguous link between ovaloolithid eggs and a particular non-avian dinosaur prevents clear biogeographic implications. A similar claim as made by Simon et al. (2019) regarding the presence of a giant oviraptorosaur in both North America and Asia based on the presence of Macroelongatoolithus cannot be made. However, some tentative but interesting observations can be made. The North American record of Ovaloolithidae, albeit sparce, is restricted to the Maastrichtian (Bray 1999; this study). However, in Asia, ovaloolithid eggs are known from possibly the Coniacian till the Maastrichtian (Wu et al. 2024), with a particularly rich record in the Campanian–Maastrichtian (Wan et al. 2007; Wang et al. 2022; Zhang et al. 2023; Wu et al. 2024). It, therefore, can be hypothesized that the parental taxon of ovaloolithid eggs migrated from Asia into Laramidia prior to the Maastrichtian.
Morrison et al. (2025) recently recovered that Tyrannosaurus possibly originated from a pre-Maastrichtian Asian ancestor, such that a similar dispersal route is postulated for the origins of Tyrannosaurini as hypothesized here for Ovaloolithidae. This could be interpreted as additional, albeit highly speculative, evidence that ovaloolithid eggs may be linked with tyrannosaurine theropods, given the similar dispersal route and timing. The Lance and Hell Creek formations include Ornithomimus and/or Struthiomimus, with the exact identifications of the specimens still unresolved (Stein 2019). Testing the link of ovaloolithid eggs to these ornithomimosaurs from a biogeographic perspective is hampered by the poor resolution of ornithomimid phylogenies (e.g., Tsogtbaatar et al. 2017; Serrano-Brañas et al. 2025). Therefore, no clear dispersal origin can be reconstructed, such that no tentative additional evidence can be provided for a simultaneous dispersal event of ovaloolithid eggs, through the parental taxon, and ornithomimosaurs, linking them to each other. A saurolophine hadrosaurid link is, similar to the tyrannosaur hypothesis, a possibility for ovaloolithid eggs based on the origins of Edmontosaurus annectens. Edmontosaurini most likely originated in Asia as well, such that a similar dispersal route can be postulated (Xing et al. 2017; Alarcón-Muñoz et al. 2023). However, as aforementioned, a theropod origin is better supported for ovaloolithid eggs (Wu et al. 2024). Therefore, the link between ovaloolithid eggs and saurolophine hadrosaurs is not well supported. This is evidenced by the differing eggshell characteristics of saurolophine eggs, as exemplified by Maiasaura (Hirsch and Quinn 1990; Zhu et al. 2022) and potentially Saurolophus (Dewaele et al. 2015).
Although Carpenter (1982) reported the presence of eggshell from the Lance Formation, none of the material was parataxonomically identified. Jackson and Varricchio (2016) identified eggshell material from the Hell Creek Formation, including theropod, ornithopod (spheroolithid), turtle, and crocodile egg material. As more eggshell material is known from the Lance Formation than reported in the present study, see Carpenter (1982), future studies should focus on the oological diversity present in this formation.
Conclusions
The first eggshell from the Maastrichtian Lance Formation is described here, and is most comparable to the oofamily Ovaloolithidae, and is assigned here to Ovaloolithus oosp. Although no unambiguous link between ovaloolithid eggs and a dinosaurian parent exist, we tentatively assign the eggshell fragment to either ornithopod or non-avian theropod dinosaurs. As eggshell material is very scarce from both the Hell Creek and Lance formations, this new eggshell from Wyoming adds significantly to the description of Maastrichtian oological material from North America.
Acknowledgements
The authors thank the ranchers for their hospitality to conduct fieldwork on their land. We thank Jean-Pierre Cavigelli (Tate Museum, Casper, USA) for aiding the fieldwork. We thank Leonard Bik and Richard Wessels (Utrecht University, the Netherlands) for producing and scanning the microslides. We thank Wilma Wessels, Ivo van der Beek and Eric W.G. Hellebrand (all Utrecht University, the Netherlands) for aiding with scanning electron microscopy at. Finally, we thank Darla K. Zelenitsky (University of Calgary, Calgary, Canada), Cassius Morrison (University College London, London, UK) and the anonymous reviewers for their detailed, helpful and constructive feedback on earlier versions of this manuscript.
Editor: Daniel Barta
References
Agnolin, F.L., Powell, J.E., Novas, F.E., and Kundrát, M. 2012. New alvarezsaurid (Dinosauria, Theropoda) from uppermost Cretaceous of north-western Patagonia with associated eggs. Cretaceous Research 35: 33–56. Crossref
Alarcón-Muñoz, J., Vargas, A.O., Püschel, H.P., Soto-Acuña, S., Manríquez, L., Leppe, M., Kaluza, J., Milla, V., Gutstein, C.S., Palma-Liberona, J., Stinnesbeck, W., Frey, E., Pino, J.P., Bajor, D., Núñez, E., Ortiz, H., Rubilar-Rogers, D., and Cruzado-Caballero, P. 2023. Relict duck-billed dinosaurs survived into the last age of the dinosaurs in subantarctic Chile. Science Advances 9: eadg2456. Crossref
Bertling, M., Braddy, S.J., Bromley, R.G., Demathieu, G.R., Genise, J., Mikuláš, R., Nielsen, J.K., Nielsen, K.S.S., Rindsberg, A.K., Schlirf, M., and Uchman, A. 2006. Names for trace fossils: a uniform approach. Lethaia 39: 265–286. Crossref
Bertling, M., Buatois, L.A., Knaust, D., Laing, B., Mángano, M.G., Meyer, N., Mikuláš, R., Minter, N.J., Neumann, C., Rindsberg, A.K., Uchman, A., and Wisshak, M. 2022. Names for trace fossils 2.0: theory and practice in ichnotaxonomy. Lethaia 55: 1–19. Crossref
Bray, E.S. 1999. Eggs and eggshell from the Upper Cretaceous North Horn Formation, central Utah. In: D.D. Gilette (ed.), Vertebrate paleontology in Utah, 361–375. Utah Geological Survey, Salt Lake City.
Brown, C.M., Campione, N.E., Wilson Mantilla, G.P., and Evans, D.C. 2022. Size-driven preservational and macroecological biases in the latest Maastrichtian terrestrial vertebrate assemblages of North America. Paleobiology 48: 210–238. Crossref
Brown, C.M., Ryan, M.J., and Evans, D.C. 2015. A census of Canadian dinosaurs: more than a century of discovery. In: O.R.P. Bininda-Emonds, G.L. Powell, H.A. Jamniczky, A.M. Bauer, and J. Theodor (eds.), All Animals are Interesting, 151–209. BIS-Verlag der Carl von Ossietzky Universität, Oldenburg.
Carpenter, K. 1982. Baby dinosaurs from the Late Cretaceous Lance and Hell Creek formations and a description of a new species of theropod. Contributions to Geology, University of Wyoming 20: 123–134.
Choi, S., Barta, D.E., Moreno‐Azanza, M., Kim, N., Shaw, C.A., and Varricchio, D.J. 2022. Microstructural description of the maniraptoran egg Protoceratopsidovum. Papers in Palaeontology 8: e1430. Crossref
Choi, S., Park, Y., Kweon, J.J., Kim, S., Jung, H., Lee, S.K., and Lee, Y.N. 2021. Fossil eggshells of amniotes as a paleothermometry tool. Palaeogeography, Palaeoclimatology, Palaeoecology 571: 110376. Crossref
Choi, S., Zhang, S., Kim, N.H., Kweon, J.J., Tanaka, K., Kubota, K., Lee, Y.N., Xie, J., Paik, I., and Lee, S.K. 2025. Thermal maturity and colors of Cretaceous East Asian fossil eggs. Sedimentary Geology 481: 106855. Crossref
Dewaele, L., Tsogtbaatar, K., Barsbold, R., Garcia, G., Stein, K., Escuillié, F., and Godefroit, P. 2015. Perinatal specimens of Saurolophus angustirostris (Dinosauria: Hadrosauridae), from the Upper Cretaceous of Mongolia. PLoS One 10: e0138806. Crossref
Fang, X.S., Li, P.X., Zhang, Z.J., Zhang, X.Q., Lin, Y.L., Guo, S.B., Cheng, Y.M., Li, Z.Y., Zhang, X.J., and Cheng, Z. W. 2009. Cretaceous strata in Nanxiong Basin of Guangdong and the evolution from the dinosaur egg to the bird egg. Acta Geoscientica Sinica 30: 167–186.
Grellet-Tinner, G. and Makovicky, P. 2006. A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications. Canadian Journal of Earth Sciences 4: 705–719. Crossref
He, Q., Jiang, Q., Xing, L., Zhang, S., Pang, W., Hu, H., Lu, S., and Yin, Q. 2019. Geochemical characteristics of newly discovered Elongatoolithidae eggs from the Upper Cretaceous of Jiangxi Province, southern China: Palaeoenvironmental and palaeoclimatic inferences. Cretaceous Research 99: 352–364. Crossref
He, Q., Yang, S., Jia, S., Xu, L., Xing, L., Gao, D., Liu, D., Gao, Y., and Zheng, Y. 2020. Trace element and isotope geochemistry of macroelongatoolithid eggs as an indicator of palaeoenvironmental reconstruction from the Late Cretaceous Xixia Basin, China. Cretaceous Research 109: 104373. Crossref
Hirsch, K.F. and Quinn, B. 1990. Eggs and eggshell fragments from the Upper Cretaceous Two Medicine Formation of Montana. Journal of Vertebrate Paleontology 10: 491–511. Crossref
Jackson, F.D. and Varricchio, D.J. 2010. Fossil eggs and eggshell from the lowermost Two Medicine Formation of western Montana, Sevenmile Hill locality. Journal of Vertebrate Paleontology 30: 1142–1156. Crossref
Jackson, F.D. and Varricchio, D.J. 2016. Fossil egg and eggshells from the Upper Cretaceous Hell Creek Formation, Montana. Journal of Vertebrate Paleontology 36: e1185432. Crossref
Jackson, F.D. and Varricchio, D.J. 2017. Paleoecological implications of two closely associated egg types from the Upper Cretaceous St. Mary River Formation, Montana. Cretaceous Research 79: 182–190. Crossref
Lawver, D.R. and Jackson, F.D. 2014. A review of the fossil record of turtle reproduction: eggs, embryos, nests and copulating pairs. Bulletin of the Peabody Museum of Natural History 55: 215–236. Crossref
Lehman, T.M., Mcdowell, F.W., and Connelly, J.N. 2006. First isotopic (U-Pb) age for the Late Cretaceous Alamosaurus vertebrate fauna of west Texas, and its significance as a link between two faunal provinces. Journal of Vertebrate Paleontology 26: 922–928. Crossref
Mikhailov, K.E. 1991. Classification of fossil eggshells of amniotic vertebrates. Acta Palaeontologica Polonica 36: 193–238.
Mikhailov, K.E. 1994. Eggs of sauropod and ornithopod dinosaurs from the Cretaceous deposits of Mongolia. Paleontological Journal 28: 141–159.
Mikhailov, K.E. 1997. Fossil and recent eggshell in amniotic vertebrates: fine structure, comparative morphology and classification. Special Papers in Palaeontology 56: 1–80.
Moreno-Azanza, M., Canudo, J.I., and Gasca, J.M. 2014. Spheroolithid eggshells in the Lower Cretaceous of Europe. Implications for eggshell evolution in ornithischian dinosaurs. Cretaceous Research 51: 75–87. Crossref
Morrison, C., Scherer, C.R., O’Callaghan, E.V., Layton, C., Boisvert, C., Rolando, M.A., Durrant, L., Salas, P., Allain, S.J.R., and Gascoigne, S.J. 2025. Rise of the king: Gondwanan origins and evolution of megaraptoran dinosaurs. Royal Society Open Science 12: 242238. Crossref
Norell, M.A., Wiemann, J., Fabbri, M., Yu, C., Marsicano, C.A., Moore-Nall, A., Varricchio, D.J., Pol, D., and Zelenitsky, D.K. 2020. The first dinosaur egg was soft. Nature 583: 406–410. Crossref
Ponstein, J., Wallaard, J.J.W., de Rijke, M., and Fraaije, R.H.B. 2024. The first cranium of Axestemys infernalis (Testudines: Trionychidae) from the Lance Formation of Wyoming, USA; an updated description and phylogenetic analysis. Palaeontologia Electronica 27: a31. Crossref
Pu, H., Zelenitsky, D.K., Lü, J., Currie, P.J., Carpenter, K., Xu, L., Koppelhus, E.B., Jia, S., Xiao, L., Chuang, H., Li, T., Kundrát, M., and Shen, C. 2017. Perinate and eggs of a giant caenagnathid dinosaur from the Late Cretaceous of central China. Nature Communications 8: 14952. Crossref
Rolleri, L., Gates, T.A., and Zanno, L.E. 2020. A possible lambeosaurine (Hadrosauridae Dinosauria) humerus from the Late Maastrichtian Hell Creek Formation of South Dakota. The Society of Vertebrate Paleontology 80th Annual Meeting, Virtual 2020, October, 12–16, 2020, Meeting Program and Abstracts, 284–285.
Sellés, A.G. and Galobart, À. 2016. Reassessing the endemic European Upper Cretaceous dinosaur egg Cairanoolithus. Historical Biology 28: 583–596. Crossref
Serrano-Brañas, C.I., Espinosa-Chávez, B., de León-Dávila, C., Maccracken, S.A., Barrera-Guevara, D., Torres-Rodríguez, E., and Prieto-Márquez, A. 2025. A long-handed new ornithomimid dinosaur from the Campanian (Upper Cretaceous) Cerro del Pueblo Formation, Coahuila, Mexico. Cretaceous Research 169: 106087. Crossref
Simon, D.J., Varricchio, D.J., Jin, X., and Robison, S.F. 2019. Microstructural overlap of Macroelongatoolithus eggs from Asia and North America expands the occurrence of colossal oviraptorosaurs. Journal of Vertebrate Paleontology 38: e1553046. Crossref
Sochava, A. 1972. The skeleton of an embryo in a dinosaur egg. Paleontological Journal 4: 527–531.
Stein, W.W. 2019. Taking count: a census of dinosaur fossils recovered from the Hell Creek and Lance Formations (Maastrichtian). Journal of Paleontological Sciences 8: 1–42.
Tanaka, K., Zelenitsky, D.K., Saegusa, H., Ikeda, T., DeBuhr, C.L., and Therrien, F. 2016. Dinosaur eggshell assemblage from Japan reveals unknown diversity of small theropods. Cretaceous Research 57: 350–363. Crossref
Tanaka, K., Zelenitsky, D.K., Therrien, F., and Kobayashi, Y. 2018. Nest substrate reflects incubation style in extant archosaurs with implications for dinosaur nesting habits. Scientific Reports 8: 3170. Crossref
Tsogtbaatar, C., Kobayashi, Y., Tsogtbaatar, K., Currie, P.J., Watabe, M., and Barsbold, R. 2017. First ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia. Scientific Reports 7: 5835. Crossref
Uematsu, R., Tanaka, K., Kozu, S., Isaji, S., and Shimojima, S. 2023. Fossil eggshells from the Early Cretaceous Okurodani Formation, northern central Japan. Historical Biology 35: 2396–2407. Crossref
Van der Linden, T.T.P., Zelenitsky, D.K., Fraaije, R.H.B., Garcia, G., Valentin, X., Holwerda, F.M., and Schulp, A.S. 2024. The first hadrosauroid eggshell from the Aix-en-Provence Basin (late Maastrichtian) of France. Historical Biology 37 (6): 1435–1442. Crossref
Varricchio, D.J., Horner, J.R., and Jackson, F.D. 2002. Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus. Journal of Vertebrate Paleontology 22: 564–576. Crossref
Voris, J.T., Zelenitsky, D.K., Therrien, F., and Tanaka, K. 2018. Dinosaur eggshells from the lower Maastrichtian St. Mary River Formation of southern Alberta, Canada. Canadian Journal of Earth Sciences 55: 272–282. Crossref
Wan, X.Q., Chen, P.J., and Wei, M.J. 2007. The Cretaceous System in China. Acta Geologica Sinica 81: 957–983. Crossref
Wang, Q., Xing, H., Shi, H.T., Fang, K.Y., Zhu, X.F., Zhou, M.X., and Wang, X.L. 2022. First discovery of dinosaur eggs in Nanhu Gebi of Hami, Xinjiang, China. Vertebrata PalAsiatica 60: 324–327.
Wegweiser, M.D., Breithaupt, B.H., Matthews, N.A., Sheffield, J.W., and Skinner, E.S. 2004. Quo vadis? Paleoenvironmental and diagenetic constraints on Late Cretaceous dinosaur skin from western North America. The Sedimentary Record 2: 4–8. Crossref
Wu, R., Lou, F., Yu, J., Xue, Y., Zhang, S., Yang, L., Qui, W., Wang, H., and Han, F. 2024. The smallest known complete dinosaur fossil eggs from the Upper Cretaceous of South China. Historical Biology: 1–10. Crossref
Xing, H., Mallon, J.C., and Currie, M.L. 2017. Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLOS ONE 12: e0175253. Crossref
Young, C. 1954. Fossil Reptilian Eggs from Laiyang, Shantung, China. Acta Paleontologica Sinica 2: 371–388.
Yu, X.Q. 1998. Characteristics of dinosaur fossils from southern Anhui and their significance for stratigraphic division. Regional Geology of China 17: 278–284.
Zelenitsky, D.K. and Hills, L.V. 1997. Normal and pathological eggshells of Spheroolithus albertensis, oosp. nov., from the Oldman Formation (Judith River Group, late Campanian), southern Alberta. Journal of Vertebrate Paleontology 17: 167–171. Crossref
Zelenitsky, D.K. and Therrien, F. 2008. Unique maniraptoran egg clutch from the Upper Cretaceous Two Medicine Formation of Montana reveals theropod nesting behaviour. Palaeontology 51: 1253–1259. Crossref
Zelenitsky, D.K., Therrien, F., Tanaka, K., Currie, P.J., and DeBuhr, C.L. 2017a. Latest Cretaceous eggshell assemblage from the Willow Creek Formation (upper Maastrichtian–lower Paleocene) of Alberta, Canada, reveals higher dinosaur diversity than represented by skeletal remains. Canadian Journal of Earth Sciences 54: 134–140. Crossref
Zelenitsky, D.K., Therrien, F., Tanaka, K., Kobayashi, Y., and DeBuhr, C.L. 2017b. Dinosaur eggshells from the Santonian Milk River Formation of Alberta, Canada. Cretaceous Research 74: 181–187. Crossref
Zhang, J., Wang, X., Jiang, S., and Li, G. 2023. The postcranial anatomy of the saurolophine hadrosaurid Laiyangosaurus youngi from the Upper Cretaceous of Laiyang, Shandong, China. The Anatomical Record (Special Issue): 1–17. Crossref
Zhang, S.K. and Wang, Q. 2010. A new oospecies of ovaloolithids from Turpan Basin in Xinjiang, China. Vertebrata Palasiatica 48: 71–75.
Zhao, Z.K. 1979. The advancement of researchers on the dinosaurian eggs in China. In: IVPP and NIGP (eds.), Mesozoic and Cenozoic Red Beds in Southern China, 330–340. Science Press, Beijing.
Zhao, Z.K. and Jiang, Y.K. 1974. Microscopic studies on the dinosaurian egg-shells from Laiyang, Shantung Province. Scientia Sinica 17: 73–83.
Zhao, Z.K., Wang, Q., and Zhang, S.K. 2015. Dinosaur eggs [in Chinese]. Palaeovertebrata Sinica 2 (7): 1–172.
Zhu, X., Wang, Q., and Wang, X. 2022. Restudy of the original and new materials of Stromatoolithus pinglingensis and discussion on some Spheroolithidae eggs. Historical Biology 34: 283–297. Crossref
Acta Palaeontol. Pol. 70 (3): 573–579, 2025
https://doi.org/10.4202/app.01211.2024