
Teeth from the Middle Jurassic of Morocco reveal the oldest turiasaurian sauropods from Africa
D. CARY WOODRUFF, PAUL M. BARRETT, DRISS OUARHACHE, ABDESSALAM EL KHANCHOUFI, KHADIJA BOUMIR, KAWTAR ECH-CHARAY, AHMED OUSSOU, RICHARD J. BUTLER, SIMON WILLS, LUKE MEADE, MIKE SMITH, and SUSANNAH C.R. MAIDMENT
Woodruff, D.C., Barrett, P.M., Ouarhache, D., El Khanchoufi, A., Boumir, K., Ech-Charay, K., Oussou, A., Butler, R.J., Wills, S., Meade, L., Smith, M., and Maidment, S.C.R. 2025. Teeth from the Middle Jurassic of Morocco reveal the oldest turiasaurian sauropods from Africa. Acta Palaeontologica Polonica 70 (3): 411–420.
Readily identifiable based on their large, “spatulate” teeth with diagnostic “heart”-shaped crowns, turiasaurians are non-neosauropodan eusauropods known from varied Jurassic and Cretaceous formations across Laurasia and Gondwana. Recently, three teeth with turiasaurian features were collected from the Middle Jurassic El Mers III Formation in the Middle Atlas Mountains of north-central Morocco. Although these teeth are superficially similar to those of the Late Jurassic Turiasaurus riodevensis from Spain, the absence of rounded denticles presence of a prominently peaked apex and a mesially flared margin, differ from other known turiasaurians. Turiasaurians have not previously been described from the El Mers III Formation, and the only named sauropod from the El Mers Group, which lacks preserved teeth, is the dubious taxon “Cetiosaurus mogrebiensis”. Due to lack of overlapping material and its lack of clear diagnostic characters, we refrain from referring these teeth to the latter, and identify them as Turiasauria indeterminate instead. These teeth represent the first definitive turiasaurian remains from Morocco, as well as the geologically oldest occurrence of Turiasauria from mainland Africa.
Key words: Dinosauria, Sauropoda, Turiasauria, teeth, Middle Jurassic, Morocco.
D. Cary Woodruff [sauropod4@gmail.com; ORCID: https://orcid.org/0000-0002-3728-134X ], Phillip and Patricia Frost Museum of Science, Miami, FL, USA; Museum of the Rockies, Bozeman, MT, USA.
Paul M. Barrett [p.barrett@nhm.ac.uk; ORCID: https://orcid.org/0000-0003-0412-3000 ], Fossil Reptiles, Amphibians, and Birds Section, Natural History Museum, Cromwell Road, London, UK; Evolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa.
Driss Ouarhache [driss.ouarhache@usmba.ac.ma; ORCID: https://orcid.org/0000-0001-5941-0977 ], Abdessalam El Khanchoufi [abdessalam.elkhanchoufi@usmba.ac.ma; ORCID: https://orcid.org/0000-0003-2664-267X ], Khadija Boumir[khadija.boumir@usmba.ac.ma; ORCID: https://orcid.org/0000-0003-4837-5410 ], Kawtar Ech-Charay [kawtar.echcharay@usmba.ac.ma; ORCID: https://orcid.org/0000-0003-4421-4214 ], and Ahmed Oussou [ahmed.oussou@usmba.ac.ma; ORCID: https://orcid.org/0000-0001-8016-9735 ], GERA Laboratory, Faculty of Sciences Dhar El Mahraz, Université Sidi Mohamed Ben Abdellah, Fez, Morocco.
Richard J. Butler [r.butler.1@bham.ac.uk; ORCID: https://orcid.org/0000-0003-2136-7541 ] and Luke Meade [l.e.meade@bham.ac.uk; ORCID: https://orcid.org/0000-0001-7829-5193 ], School of Geography, Earth & Environmental Sciences, University of Birmingham, Edgbaston, Birmingham, UK.
Simon Wills [s.wills@nhm.ac.uk; ORCID: https://orcid.org/0000-0002-2012-1016] and Mike Smith [Mike.Smith@nhm.ac.uk; ORCID: https://orcid.org/0009-0007-0049-9162 ], Fossil Reptiles, Amphibians, and Birds Section, Natural History Museum, Cromwell Road, London, UK.
Susannah C.R. Maidment [susannah.maidment@nhm.ac.uk; ORCID: https://orcid.org/0000-0002-7741-2500 ], Fossil Reptiles, Amphibians, and Birds Section, Natural History Museum, Cromwell Road, South Kensington, London SW7 5BD, UK; School of Geography, Earth & Environmental Sciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK.
Received 10 October 2024, accepted 17 June 2025, published online 7 August 2025.
Copyright © 2025 D.C. Woodruff et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Turiasauria is a clade of large-bodied non-neosauropod eusauropods ranging from the Early or Middle Jurassic to the Early Cretaceous (Royo-Torres et al. 2006; Mocho et al. 2016; Mannion 2019; Royo-Torres et al. 2021). First identified from the Iberian Peninsula (Royo-Torres et al. 2006) and originally restricted to former Laurasian continents (Britt et al. 2017; Royo-Torres et al. 2017), Turiasauria is now known from various locations in Africa (Mocho et al. 2016; Mannion et al. 2019; Royo-Torres et al. 2021), India (Sharma et al. 2022) and perhaps Argentina (Royo-Torres et al. 2021; Milàn and Mateus 2024). Recent identification of an Early Jurassic putative turiasaurian tooth from Denmark (Milàn and Mateus 2024) indicates that the group may have originated by that time, achieving a wider distribution by the Middle Jurassic (Royo-Torres et al. 2021; Scharma et al. 2022).
Until now, the African record of Turiasauria has consisted of two named taxa from the Middle Jurassic Isalo III Formation of Madagascar (Narindasaurus, Royo-Torres et al. 2021) and the Upper Jurassic Tendaguru Formation of Tanzania (Tendaguria, Mannion et al. 2019), although isolated teeth from the Middle Jurassic of Niger and Madagascar and the Early Cretaceous of Libya have also been referred to the clade (Mocho et al. 2016). Xing et al. (2015) suggested that the Middle Jurassic Moroccan sauropod Atlasaurus formed a clade with the turiasaurians Losillasaurus and Turiasaurus, but this result has not been replicated, and Atlasaurus is more commonly thought to be a brachiosaurid (e.g., Mannion et al. 2019; Royo-Torres et al. 2021). Here, we report three associated sauropod teeth from the Middle Jurassic El Mers III Formation of the Middle Atlas Mountains, Morocco, which represent the earliest definitive record of turiasaurians in Africa.
The Bathonian El Mers III Formation, which crops out in exposures around the town of Boulemane in the Middle Atlas Mountains, is becoming increasingly important for understanding the radiation of dinosaurs during the Middle Jurassic. Dinosaur remains from the formation include the world’s oldest ankylosaur, and the first from Africa, Spicomellus afer (Maidment et al. 2021), the early stegosaurs Adratiklit boulahfa (Maidment et al. 2020) and Thyreosaurus atlasicus (Zafaty et al. 2024), and the world’s oldest cerapodan ornithischian (Maidment et al. 2025). The formation therefore hosts some of the earliest representatives of several major ornithischian dinosaur clades, shedding light on dinosaur diversifications and radiations in the aftermath of the global environmental perturbations that marked the end of the Early Jurassic (Cabreira et al. 2016; Reolid et al. 2022; Maidment et al. 2025).
Institutional abbreviations.—FHPR, Utah Field House of Natural History State Park Museum, Vernal, Utah, USA; MAP, Museo Aragonés de Paleontología, Teruel, Spain; ML, Museu da Lourinhã, Lourinhã, Portugal; MNHN, Muséum national d’Histoire naturelle, Paris, France; NHMD, Natural History Museum of Denmark, Copenhagen, Denmark; RAJ, Geological Survey of India, Kolkata, India; UMNH Natural History Museum of Utah, Salt Lake City, Utah, USA; USMBA, Université Sidi Moha med Ben Abdellah, Fez, Morocco.
Material and methods
The specimens described herein are three teeth (USMBA 002–004) that were surface collected from the Bathonian El Mers III Formation at the Boulahfa Plain, near Boulemane in the Middle Atlas Mountains, Morocco (see Maidment et al. 2020 for locality data). On the Boulahfa Plain, the El Mers III Formation is exposed in erosive badlands formed by frequent flash flood events. This flash flooding transports large amounts of sediment, and in situ bones are frequently exposed and re-buried by these bulk transport events. The teeth were surface-collected by a local farmer in close proximity to large limb and other elements that were subsequently re-buried under at least 5 m of sediment during a flash-flooding event before excavation was possible. The El Mers III Formation at the Boulahfa Plain is composed of variegated mudstones and sparse, laterally discontinuous sandstones, and is predominantly greenish in its lower parts and purple-reddish in its upper parts (Maidment et al. 2020). The zone from which most of the fossils appear to be derived is close to the top of the greenish section and is around 20 m thick (Maidment et al. 2020: fig. 3). The site where the teeth were collected, which we refer to as the “Big Flood Quarry”, is located approximately 15 m below the colour-change from predominantly green to predominantly purple-red mudstones.
Teeth were photographed and measured using standard documentation techniques. Tooth anatomical terminology follows that of Smith and Dodson (2003) and Hendrickx et al. (2015).
Results
Description.—According to Royo-Torres et al. (2021), isolated turiasaur teeth can be assigned approximate locations in the skull based on the orientation of the crowns. In premaxillary and maxillary teeth, in mesial or distal orientation, the crown is curved lingually. In dentary teeth, the crown is angled “labially” (Royo-Torres et al. 2021), although it would be more accurate to describe them as straight, or apically oriented, as the crown apex is not labially inclined. Applying these criteria, USMBA 002 and 004 are from the premaxilla or maxilla, and from the distal orientation of the crown apices, likely from the right side. Because of the large wear facets, tooth USMBA 003 is more challenging to place. In mesial and distal orientation, the undulating crown does not appear to curve strongly lingually, suggesting that this may be a dentary tooth, from the right side also. Measurements of the teeth are provided in Table 1.
Table 1. Measurements (in mm) of the three Boulahfa Plain sauropod teeth. Teeth labelled 1–3 for convenience. * including root; ** heavily worn; e estimate due to breakage. a, b, and c denote the formulae for the Slenderness and Compression Index; i.e., for Slenderness Index, max crown apicobasal height is divided by the max crown mesiodistal width.
| |
USMBA 002 |
USMBA 003 |
USMBA 004 |
|
Total tooth length* |
64.7 |
53.0 |
71.3 |
|
Max crown apicobasal height (a) |
31.3 |
23.4** |
46.6 |
|
Max crown mesiodistal width (b) |
18.5 |
15.6** |
28.6 |
|
Basal crown mesiodistal width |
15.8 |
13.3 |
16.9 |
|
Max crown labiolingual width (c) |
12.6 |
9.4 |
13.0 |
|
Slenderness Index (a/b) |
1.69 |
1.56e |
1.63 |
|
Compression Index (c/b) |
0.68 |
0.60e |
0.45 |
All three teeth include a substantial portion of the root and two of the crowns are well-preserved (Fig. 1). The crowns of USMBA 002 and 003 are truncated by wear facets, heavily in the case of USMBA 003, but USMBA 004 is unworn. The crown surfaces have been polished by wind and/or water action, but the enamel texture is still clearly visible on tooth USMBA 004 and is partially visible in patches on USMBA 002 and 003. USMBA 002 and 003 are consistent in morphology and are described together for convenience, whereas USMBA 004 differs in shape and is described separately. Based on their relative sizes and different degrees of crown expansion, it seems likely that teeth USMBA 002 and 003 are from the distal ends of the tooth row, whereas tooth USMBA 004 was likely more anteriorly or centrally located.

Fig. 1. The three turiasaurian sauropod teeth from Boulahfa Plain, Morocco, Middle Jurassic. A. USMBA 002. B. USMBA 003. C. USMBA 004. Teeth in lingual (A1–C1), mesial (A2–C2), labial (A3–C3), distal (A4–C4), apical (A5–C5), and basal (A6–C6) views.
In labial view, the tooth crowns of USMBA 002 and 003 exhibit slight mesiodistal expansion with respect to the root. Maximum mesiodistal expansion occurs at a point only a few millimetres apical to the root / crown boundary and is developed to similar degrees either side of the crown long axis. Apical to the maximum diameter, the crown tapers to a bluntly rounded terminus. The mesial crown margin is smoothly and gently convex apicobasally, whereas the distal margin is sinuous, being slightly convex basally, but more strongly concave apically. As a result, the crowns are asymmetrical in labial view, and the tooth apex is recurved distally. Overall, these features give the crowns a narrow, heart-shaped outline. The labial surface is strongly convex mesiodistally with a slight break-in-slope near to the distal margin due to the presence of an incipient distal groove. Most of the surface is smooth, but small unpolished patches wear “wrinkled” enamel showing evidence of connected, anastomosing ridges.
In lingual view, the crowns are gently concave mesiodistally and this, in combination with the labial convexity, gives them a strongly D-shaped transverse cross-section. The central part of the lingual convexity is occupied by a low eminence, in a position equivalent to the lingual ridge of many other sauropods. This feature extends apically toward the crown tip and divides the lingual surface into two subequal parts.
In mesial or distal view, the labial crown surfaces are strongly convex apicobasally, whereas the lingual surfaces are strongly concave. The crown apex is lingually inclined.
As preserved, the crown margins of USMBA 002 and 003 lack denticles. The tip of USMBA 002 is truncated by a narrow, high-angled, elliptical wear facet that is apicobasally longer than transversely wide (width is c. 1/2 the length) and extends along the mesial crown margin for a short distance. A very narrow, apicobasally elongate (width is c. 1/3 the length) wear facet is present on the distal margin but does not extend apically to merge with the mesial facet. USMBA 003 is more heavily worn, with large, high-angled, apicobasally ovoid wear facets that have truncated the entire mesial and distal margins and that are confluent around the crown apex. However, although extensive, the wear has not led to the development of the distinctive “shoulders” seen in some other worn sauropod teeth (e.g., Camarasaurus spp.; Upchurch and Barrett 2000). In both cases, the wear facets reveal that the enamel is thin and symmetrically distributed around the tooth crown.
USMBA 004 is similar to the other teeth in many respects, but its crown is much more strongly expanded mesiodistally with respect to the root, in labial view, and the curvature of its mesial and distal margins is correspondingly greater, giving it a much more obviously heart-shaped outline. The expansion is more strongly asymmetrical in this tooth, with most occurring mesial to the crown long-axis. The labial surface is strongly convex mesiodistally, due to the presence of a prominent, centrally positioned, apicobasally extending swelling, which is defined both mesially and distally by incipient grooves. In lingual view, an incipient lingual ridge is present, but is limited to the apical-most part of the tooth. Wear facets and denticles are both absent. The crown surface is better preserved than in USMBA 002 and 003 and an anastomosing pattern of enamel wrinkles covers most of the crown surface.
The slenderness index (SI) of the crowns, the ratio of apicobasal length to mesiodistal width (Upchurch 1998), ranges from 1.56 (USMBA 003, although this is likely an underestimate due to heavy wear), through 1.63 (USMBA 004) to a maximum of 1.69 (USMBA 002). A second ratio, the compression index (CI), defined as crown maximum labiolingual width divided by crown mesiodistal width (Díez Días et al. 2013), varies from 0.45 (USMBA 004), through ~0.6 (USMBA 003) to 0.68 (USMBA 002).
In all three teeth, the root has a sub-circular, cylindrical cross-section and is broken basally to expose the pulp cavity.
Comparisons.—The three teeth from the Boulahfa Plain, especially USMBA 002 and 004, are strikingly similar to previously documented turiasaurian teeth. The elongate, tapered crown apex present in USMBA 004, is likewise seen in Losillasaurus giganteus (Royo-Torres et al. 2021), Turiasaurus riodevensis (Royo-Torres and Upchurch 2012), isolated upper Kimmeridgian to basal Tithonian teeth from Portugal (Mocho et al. 2016), and an isolated Pliensbachian tooth from Denmark (Milàn and Mateus 2024). The mesiodistal widths of the crowns, especially the degree of mesodistal expansion of USMBA 002 and the mesiodistal expansion along USMBA 004 is similarly seen in L. giganteus (from Royo-Torres et al. 2006), T. riodevensis (from Royo-Torres and Upchurch 2012), isolated teeth from the upper Kimmeridgian to basal Tithonian of Portugal (Mocho et al. 2016), an isolated Bathonian tooth from India (Sharma et al. 2022), and the isolated Pliensbachian tooth from Denmark (Milàn and Mateus 2024).
In terms of their overall morphology, the Boulahfa Plain teeth are most similar to those from T. riodevensis. By contrast, many of the maxillary teeth in L. giganteus have a stronger degree of apex distal curvature, and its mesial and distal wear facets are almost shelf-like, with some facets having peaked apices themselves. These features are not observed in the Boulahfa Plain teeth. Unlike in either L. giganteus or T. riodevensis, there are no rounded denticles along the mesial apex boundary. As in T. riodevensis, the apex of the crown of USMBA 004 is relatively apicobasally straight with a low degree of apical curvature (Royo-Torres and Upchurch 2012). The labial groove on the mesial side has a c. 4 mm depressed region, and along with the convex mesial edge, the crown morphology of USMBA 004 is much more mesiodistally “flared” than known turiasaurian examples (i.e., T. riodevensis, L. giganteus; Fig. 2).

Fig. 2. Comparisons to other known or suspected turiasaurs. A. Premaxillary and maxillary (top row MAP-6013–6018), and dentary (bottom row MAP-6032–6038) teeth of Losillasaurus giganteus Casanovas et al., 2001, from the Late Jurassic of Spain (from Royo-Torres et al. 2021). B. Teeth of Turiasaurus riodevensis Royo Torres et al., 2006, from the Late Jurassic of Spain (from Royo-Torres and Upchurch, 2012). C. Isolated Turiasauria-like teeth from the Upper Jurassic (Upper Kimmeridgian-basal Tithonian) of Portugal (from Mocho et al. 2016). D. Premaxillary, maxillary, and dentary teeth of Moabosaurus utahensis Britt et al., 2017, from the Early Cretaceous of the United States (from Britt et al. 2017). E. Premaxillary and maxillary teeth (MNHN MAJ 423) of Narindasaurus thevenini Royo-Torres et al., 2021, from the Middle Jurassic of Madagascar (from Royo-Torres et al. 2021). F. Turiasaurian tooth (NHMD 1185136) from the Lower Jurassic (Pliensbachian) of Denmark (from Milàn and Mateus 2024). G. Premaxillary-maxillary tooth (UMNH.VP.26004) of Mierasaurus bobyoungi Royo-Torres et al., 2017, from the Early Cretaceous of USA (from Royo-Torres et al. 2017). H. Turiasaurian tooth (RAJ/JAIS/CVQS001) from the Middle Jurassic (Bathonian) of India (from Sharma et al. 2022). I. Tooth (ML 368) of Zby atlanticus Mateus et al., 2014, from the Late Jurassic of Portugal (from Mateus et al. 2014). J. Tooth of the possible turiasaur Oplosaurus armatus Gervais, 1852, from the Early Cretaceous of England (from Mocho et al. 2016). K. Drawing of a turiasaur-like tooth from Jobaria tiguidensis Sereno et al., 1999, from the Middle Jurassic of Niger (from Sereno et al. 1994). L. Turiasauria-like tooth (FHPR 18687.2) associated with postcranial remains of Haplocanthosaurus sp. from the Late Jurassic of USA. Photo by J. Foster. Tooth orientations: 1, lingual; 2, labial; 3, mesial; 4, apical; 5, distal; 6, basal. Not to scale.
Discussion
Systematic position.—The Boulahfa Plain teeth possess a series of diagnostic features that permit referral to Eusauropoda, including: tooth crowns with D-shaped cross-sections; a lingual concavity bearing an incipient lingual ridge; and high-angled mesial and distal wear facets (Upchurch 1998; Wilson and Sereno 1998; Wilson 2002; Upchurch et al. 2004). The presence of incipient mesial and distal labial grooves, as well as extensive areas of anastomosing enamel wrinkles, are also consistent with eusauropod affinities (Upchurch 1998; Wilson and Sereno 1998; Wilson 2002; Upchurch et al. 2004) although these features first appear in the early diverging non-eusauropod Pulanesaura eocollum (McPhee and Choiniere 2018), which is close to sauropod origins.
Among eusauropods, heart-shaped crowns in labial/lingual view are characteristic of Turiasauria, enabling referral of the Boulahfa Plain teeth to this clade (Royo-Torres et al. 2006; Mocho et al. 2016). A second possible turiasaurian synapomorphy, the presence of elongate apicobasal grooves along the root (Royo-Torres et al. 2021), is not present in any of the teeth from the Boulahfa Plain, although the root of USMBA 004 possesses ambiguous traces of what might be grooves. However, it is possible that this feature diagnoses a subset of later-occurring turiasaurians and was absent in earlier diverging taxa.
Several other features of the Boulahfa Plain teeth have systematic significance. Their low SI values (1.56–1.69) exclude them from Titanosauria and Diplodocoidea (which have SI >4.0; e.g., Upchurch 1998; Upchurch et al. 2004). The CI values (0.45–0.68) for the Boulahfa Plain teeth are lower than those reported for titanosaurians (~0.7–1.1; Díez Díaz et al. 2013; Holwerda et al. 2018). The absence of denticles (in USMBA 004) is consistent with referral to Eusauropoda (e.g., Upchurch 1998) but does not help to classify the teeth more precisely, as denticles are retained in multiple non-diplodocoid and non-titanosaurian lineages, such as mamenchisaurids and early diverging macronarians (e.g., Wilson and Sereno 1998; Upchurch et al. 2004), suggesting substantial homoplasy in denticle loss.
The Moroccan sauropod record.—Named sauropod dinosaurs from Morocco consist of the Early Jurassic vulcanodontid Tazoudasaurus naimi (Allain et al. 2004), the Middle Jurassic taxa Atlasaurus imelakei (Monbaron et al. 1999; Xing et al. 2015; Royo-Torres et al. 2021) and “Cetiosaurus mogrebiensis” (de Lapparent 1955), and the early Late Cretaceous rebbachisaurid Rebbachisaurus garasbae (Lavocat 1954; Wilson and Allain 2015). However, indeterminate Early to Middle Jurassic and Late Cretaceous teeth (Holwerda et al. 2018), postcranial material (Pereda-Suberbiola et al. 2004; Mannion and Barrett 2013; Ibrahim et al. 2016; Nicholl et al. 2018; Holwerda 2020) and trackways (Termier 1942; Dutuit and Ouazzou 1980; Jenny et al. 1981; Hadri et al. 2007; Belvedere 2008; Marty et al. 2010; Enniouar et al. 2014; Lallensack et al. 2018; Oukassou et al. 2019; Boutakiout et al. 2020; Oussou et al. 2023) are also documented.
Although the teeth from the Boulahfa Plain are referrable to Turiasauria, it is useful to compare them to the teeth associated with other named Moroccan taxa. They can be distinguished from those of Tazoudasaurus naimi, as the latter has almost symmetrical tooth crowns in labial view, which lack the heart-shaped outline of turiasaurians (Allain and Aquesbi 2008). Moreover, the teeth of T. naimi bear marginal denticles (Allain and Aquesbi 2008). The holotype of Rebbachisaurus garasbae does not include teeth (Lavocat 1954; Wilson and Allain 2015) but isolated teeth from the early Late Cretaceous of Morocco have been referred to Rebbachisauridae (e.g., Holwerda et al. 2018). These referred teeth are elongate, peg-like and cylindrical, differing substantially from those of turiasaurians. Of perhaps more relevance, the Middle Jurassic taxon Atlasaurus imelakei has been recovered as a turiasaurian in some recent analyses (Xing et al. 2015) and its holotype includes teeth (Monbaron et al. 1999: fig. 1f, g). In A. imelakei the crowns are mesiodistally narrow, lack a heart-shaped outline and some possess denticles, suggesting that the teeth from the Boulahfa Plain are a distinct taxon, although they are near contemporaries. Moreover, A. imelakei is usually recovered as an early diverging macronarian in most analyses (e.g., Monbaron et al. 1999; Royo-Torres et al. 2021) so it is not closely related to turiasaurians.
“Cetiosaurus mogrebiensis”, is a problematic taxon of uncertain phylogenetic affinity that is currently considered to be a nomen dubium (Upchurch et al. 2004). Named by Albert-Félix de Lapparent in 1955, this species was described from three partial skeletons collected from different localities within a c. 3 km radius of the town of El Mers, which is about 20 km northeast of Boulemane. No teeth are known for “Cetiosaurus mogrebiensis”, so it is not possible to compare the specimens described herein directly. Although de Lapparent (1955) noted similarities between “C. mogrebiensis” and the “cetiosaurids” Bothriospondylus spp. and C. oxoniensis, and Läng and Mahammed (2010) recovered it within a polytomy of “cetiosaurid” eusauropods, Upchurch and Martin (2003) noted that comparisons to “cetiosaurids” were untenable due to the absence of phylogenetically informative characters in the material of “C. mogrebiensis” and several notable anatomical differences between the latter and C. oxoniensis. As a result, they noted that the Moroccan and UK species were unlikely to be congeneric (Upchurch and Martin 2003; Upchurch et al. 2004). Redescription of the “C. mogrebiensis” material is needed to determine its systematic position and to test its taxonomic validity. If “C. mogrebiensis” is found to be a turiasaur, the Boulahfa Plain teeth could conceivably be referred to it (despite lack of overlapping material), but in the absence of any such evidence, we refrain from making any referrals herein.
The oldest turiasaur from Africa.—Occurrences of Turiasauria in both Gondwana and Laurasia indicate that the clade achieved a wide distribution by Bathonian times, as shown by their presence in the Middle Jurassic of India (Sharma et al. 2022), Madagascar (Royo-Torres et al. 2021), Morocco (this paper) and the UK (Barrett 2006, 2021; Royo-Torres et al. 2021; Fig. 3). Turiasaurian taxa are better known from the Upper Jurassic, with representatives from Portugal (Zby), Spain (Losillasaurus, Turiasaurus), Switzerland (Amanzia), and Tanzania (Tendaguria), and the clade persists into the Lower Cretaceous (Moabsaurus, Mierasaurus), at least in North America and Europe (e.g., Britt et al. 2017; Royo-Torres et al. 2017; Mannion 2019; Fig. 3). If a tooth from the Hasle Formation of Denmark is not allochthonous, then the record of Turiasauria begins in the Early Jurassic (Pliensbachian), c. 17 million years earlier than thought previously (Milàn and Mateus 2024). Additionally, both Royo-Torres et al. (2021) and Milàn and Mateus (2024) suggested that the teeth of Bagualia alba from the Toarcian of Argentina (Pol et al. 2020: fig. 2f) were similar to those from turiasaurians, which would suggest a global radiation of the clade by the mid- to late Early Jurassic. However, Bagulia has not been recovered as a turiasaurian in any phylogenetic analysis and it is usually regarded as an early diverging eusauropod (Pol et al. 2020; Gomez et al. 2021, 2024). Moreover, most of the teeth referred to this taxon (e.g., Pol et al. 2020: fig. 2g) are much more similar in shape to those of other early sauropods, such as Tazoudasaurus and Pulanesaura, with relatively narrow, denticulate crowns. Although one figured tooth of Bagualia is superficially heart-shaped, this morphology might be due to differential wear of the crown margins (such as variable wear within the tooth row; Holwerda et al. 2015; Wiersma and Sander 2017; Régent et al. 2024) rather than representing its original shape.
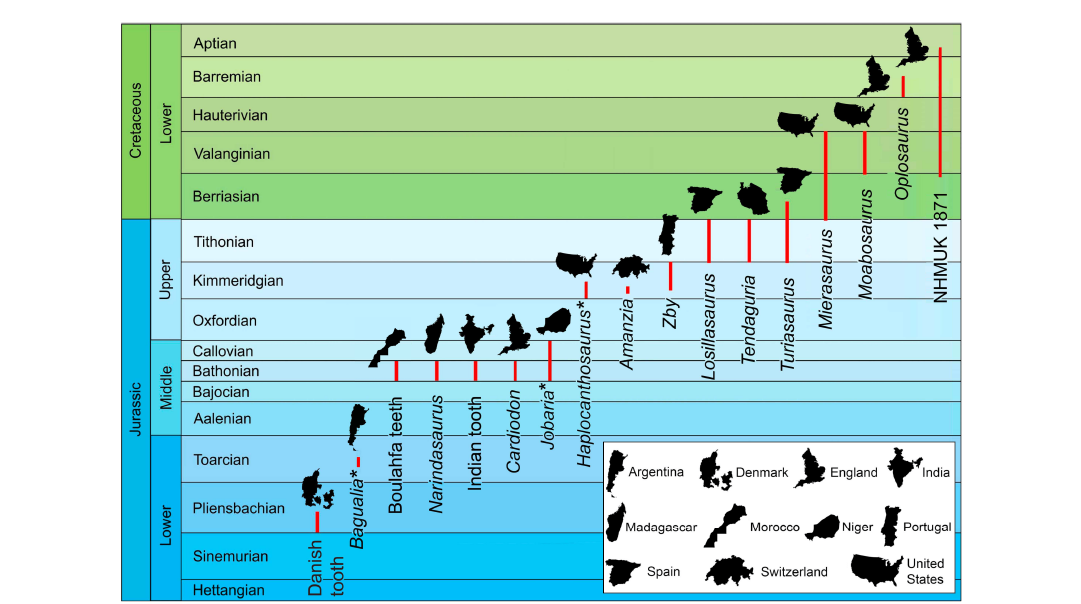
Fig. 3. Temporal and geographic distribution of Turiasauria. Modified after Milàn and Mateus (2024). Debated or questionable taxa with turiasaur-like teeth denoted with *.
Isolated teeth from the Middle Jurassic of Niger (de Lapparent 1960) and Madagascar (Läng 2008), as well as the Lower Cretaceous of Libya (Le Loeuff et al. 2010), have been referred to Turiasauria (Mocho et al. 2016). However, in the case of the tooth from Niger, the apical part of the crown lacks the distinctive strongly convex distal margin that gives turiasaurians their distinctive heart-shaped morphology (Mocho et al. 2016: fig. 10f), while the teeth from Madagascar (Mocho et al. 2016: fig. 10g) and Libya (Le Loeuff et al. 2010: fig. 5) are too incomplete to determine if they were originally heart-shaped or not; although the Libyan teeth possess the synapomorphic apicobasal grooves (Royo-Torres 2021). As a consequence, all these referrals should be regarded with caution, with Mannion (2019) suggesting that these earlier reports of putative African “turiasaurian” teeth should be regarded as Sauropoda indet. (see Poropat et al. 2022 for a review of the taxonomic interpretations of this tooth).
Sereno et al. (1999) noted that the teeth of Jobaria tiguidensis are spatulate and variably denticulated along their mesial and distal margins (Sereno et al. 1994: fig. 4); and while the cross-sectional morphology of the crown was not figured nor discussed, the labial/lingual morphology is strongly reminiscent of Losillasaurus, Turiasaurus, and other turiasaurs (Fig. 2). However, while the teeth of Jobaria appear to be similar to those of turiasaurs, and the prefrontal and quadratojugal of Jobaria has been noted to be similar to Turiasaurus (Royo-Torres and Upchurch 2012), no phylogenetic analyses have recovered Jobaria within Turiasauria (Upchurch et al. 2004; Remes et al. 2009; Chure et al. 2010). Likewise, teeth bearing turiasaur-like morphologies are associated with new specimens of Haplocanthosaurus sp., which could potentially challenge the traditional phylogenetic position of this taxon (DCW and John Foster, personal observation 2024). However, as with Jobaria, no phylogenetic analyses have recovered Haplocanthosaurus within Turiasauria. Potentially, convergence could explain the dental similarities in these taxa, and examinations using dental indices (e.g., Frauenfelder et al. 2024) might help to elucidate their taxonomic position in the future.
By contrast, the Boulahfa Plain teeth show clear evidence of turiasaurian affinities and thus represent the oldest record of the clade from mainland Africa, as well as the first definitive record of the clade from North Africa. They are close in age to Narindasaurus from the Isalo III Formation of Madagascar (Middle Jurassic, Royo-Torres et al. 2021) and pre-date the Late Jurassic (Tithonian) record of Tendaguria from Tanzania (Mannion et al. 2019). The Boulahfa Plain teeth expand the geographic range of the clade during the Middle Jurassic, and together with the material from Madagascar and the possible record from the Pliensbachian of Denmark (Milàn and Mateus 2024) they imply that the initial spread of the clade occurred at some point in the late Early Jurassic.
Conclusions
The three Boulahfa Plain sauropod teeth represent the first definitive turiasaurian remains from Morocco, as well as earliest record of Turiasauria from mainland Africa. As previously demonstrated by the co-occurring ankylosaur Spicomellus afer and the stegosaurs Adratiklit boulahfa and Thyreosaurus atlasicus the Middle Jurassic El Mers III Formation of Morocco is vitally important for documenting the early radiation and biogeographic history of many dinosaurian clades.
Although palaeontological exploration in the El Mers III Formation is in its early stages, so far, its dinosaur fauna consists of two stegosaurs, an ankylosaur, an early-diverging cerapodan, and a turiasaur. A similar faunal composition is observed in various Late Jurassic faunas globally, for example: from the Tendaguru Formation of southeastern Tanzania, from the Lourinhã Formation of western Portugal, possibly even from the Morrison Formation of the western United States, and from the Villar del Arzobispo Formation in eastern Spain (Campos-Soto et al. 2019). Middle Jurassic terrestrial faunas are globally poorly represented (Maidment et al. 2020), making the fauna of the El Mers III Formation critical to our understanding of the establishment of these famous Late Jurassic faunas.
Acknowledgements
Rafael Royo-Torres (University of Zaragoza, Spain), Alessandro Chiarenza (University College London, UK), and Archana Sharma (Geological Survey of India, Kolkata, India) provided relevant literature, Rafael Royo-Torres (University of Zaragoza, Teruel, Spain) and Pedro Mocho (Instituto Dom Luiz Lisbon, Portugal) additionally provided their turiasaurian guidance and expertise. Finally, thanks to handling editor Daniel Barta (Oklahoma State University, Tahlequah, USA), Femke Holwerda (Utrecht University, The Netherlands), and an anonymous reviewer for their constructive comments. Thanks to the donors of Frost Science’s Paleontology Department which enabled DCW to participate in the joint expeditions of the NHM, USMBA, and University of Birmingham. Funding for these expeditions is provided by the British Institute of Libyan and North African Studies and the NHM’s Science Investment Fund.
Editor: Daniel Barta
References
Allain, R. and Aquesbi, N. 2008. Anatomy and phylogenetic relationships of Tazoudasaurus naimi (Dinosauria, Sauropoda) from the late Early Jurassic of Morocco. Geodiversitas 30 (2): 345–424.
Allain, R., Aquesbi, N., Dejax, J., Meyer, C., Monbaron, M., Montenat, C., Richir, P., Rochdy, M., Russell, D., and Taquet, P. 2004. A basal sauropod dinosaur from the Early Jurassic of Morocco. Comptes Rendus Palevol 3 (3): 199–208. Crossref
Barrett, P.M. 2006. A sauropod dinosaur tooth from the Middle Jurassic of Skye, Scotland. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 97 (1) 25–29. Crossref
Barrett, P.M. 2021. Dinosaur material from the Lower Greensand Group of Upware, Cambridgeshire, and the age of ‘Wealden’ vertebrates from the ‘Bedfordshire Straits’. Proceedings of the Geologists’ Association 132 (2021): 497–505. Crossref
Belvedere, M. 2008. Ichnological Researches on the Upper Jurassic Dinosaur Tracks in the Iouaridène Area (Demnat, Central High-Atlas, Morocco). 130 pp. Ph.D. Dissertation, Universita Degli Studi di Padova, Padua.
Boutakiout, M., Masrour, M., and Pérez-Lorente, F. 2020. New sauropod morphotype definition in the oriental section 0F Imilchil megatracksite, high atlas (Morocco). Journal of African Earth Sciences 161: 103664. Crossref
Britt, B.B., Scheetz, R.D., Whiting, M.F., and Wilhite, D.R. 2017. Moabosaurus utahensis, n. gen., n. sp., a new sauropod from the Early Cretaceous (Aptian) of North America. Contributions from the Museum of Paleontology, University of Michigan 32: 189–243.
Cabreira, S.F., Kellner, A.W.A., Dias-da-Silva, S., da Silva, L.R., Bronzati, M., de Almeida Marsola, J.C., Müller, R.T., de Souza Bittencourt, J., Batista, B.J.A., Raugust, T., and Carrilho, R. 2016. A unique Late Triassic dinosauromorph assemblage reveals dinosaur ancestral anatomy and diet. Current Biology 26 (22): 3090–3095. Crossref
Campos-Soto, S., Benito, M.I., Cobos, A., Caus, E., Quijada, I.E., Suarez-Gonzalez, P., Mas, R., Royo-Torres, R., and Alcalá, L. 2019. Revisiting the age and palaeoenvironments of the Upper Jurassic–Lower Cretaceous? Dinosaur-bearing sedimentary record of eastern Spain: implications for Iberian palaeogeography. Journal of Iberian Geology 45: 471–510. Crossref
Casanovas, M.L., Santafé, J.V., and Sanz, J.L. 2001. Losillasaurus giganteus, a new sauropod from the transitional Jurassic–Cretaceous of the Los Serranos basin (Valencia, Spain). Paleontologia y Evolució 32: 99–122.
Chure, D., Britt, B.B., Whitlock, J.A., and Wilson, J.A. 2010. First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition. Naturwissenschaften 97: 379–391. Crossref
Díez Díaz, V., Tortosa, T., and Le Loeuff, J. 2013. Sauropod diversity in the Late Cretaceous of southwestern Europe: the lessons of odontology. Annales de Paléontologie 99 (2): 119–129. Crossref
Dutuit, J.-M., Ouazzou, A. 1980. Découverte dune piste de Dinosaure sauropode sur le site dempreintes de Demnat (Haut-Atlas marocain). Mémoires de la Société géologique de France (Nouvelle Série) 139: 95–102.
Enniouar, A., Lagnaoui, A., and Habib, A. 2014. A Middle Jurassic sauropod tracksite in the Argana Basin, Western High Atlas, Morocco: an example of paleoichnological heritage for sustainable geotourism. Proceedings of the Geologists’ Association 125 (1): 114–119. Crossref
Frauenfelder, T.G., Birch, S.A., Bell, P.R., and Campione, N.E. 2024. Revealing the use of dental indices to infer taxonomic variation in sauropod dinosaurs. Palaeontology 67: e12725. Crossref
Gervais, P. 1852. Zoologie et paléontologie françaises (animaux vertébrés): ou nouvelles recherches sur les animaux vivants et fossiles de la France Vol. 3. 174 pp. Arthus Bertrand, Paris.
Gomez, K.L., Carballido, J.L., and Pol, D. 2021. The axial skeleton of Bagualia alba (Dinosauria: Eusauropoda) from the Early Jurassic of Patagonia. Palaeontologica Electronica 24: a37. Crossref
Gomez, K.L., Carballido, J.L., and Pol, D. 2024. Cranial anatomy of Bagualia alba (Dinosauria, Eusauropoda) from the Early Jurassic of Patagonia and the implications for sauropod cranial evolution. Journal of Systematic Palaeontology 22 (1): 2400471. Crossref
Hadri, M., Boutakiout, M., and Pérez-Lorente, F. 2007. Nuevos yacimientos de icnitas de dinosaurios carixiensis. Sur del Alto Atlas Central (Goulmima. Marruecos). Geogaceta 41: 107–110.
Hendrickx, C., Mateus, O., and Araújo, R. 2015. A proposed terminology of theropod teeth (Dinosauria, Saurischia). Journal of Vertebrate Paleontology 35 (5): e982797. Crossref
Holwerda, F.M. 2020. Sauropod dinosaur fossils from the Kem Kem and extended ‘Continental Intercalaire’ of North Africa: A review. Journal of African Earth Sciences 163: 103738. Crossref
Holwerda, F.M., Díez Díaz, V., Blanco, A., Montie, R., and Reumer, J.W.F. 2018. Late Cretaceous sauropod tooth morphotypes may provide supporting evidence for faunal connections between North Africa and Southern Europe. Peer J 6: e5925. Crossref
Holwerda, F.M., Pol, D., and Rauhut, O.W.M. 2015. Using dental enamel wrinkling to define sauropod tooth morphotypes from the Cañadón Asfalto Formation, Patagonia, Argentina. PLoS ONE 10 (2): e0118100. Crossref
Ibrahim, N., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D.M., Gorscak, E., and Lamanna, M.C. 2016. Evidence of a derived titanosaurian (Dinosauria, Sauropoda) in the “Kem Kem beds” of Morocco, with comments on sauropod paleoecology in the Cretaceous of Africa. Cretaceous Period: Biotic Diversity and Biogeography. New Mexico Museum of Natural History and Science Bulletin 71: 149–159.
Jenny, J., Le Marrec, A., and Monbaron, M. 1981. Les couches rouges du Jurassique moyen du Haut Atlas central (Maroc); Correlations lithostratigraphiques, elements de datations et cadre tectono-sedimentaire. Bulletin de la Société géologique de France 7 (6): 627–640. Crossref
Lallensack, J.N., Ishigaki, S., Lagnaoui, A., Buchwitz, M., and Wings, O. 2018. Forelimb orientation and locomotion of sauropod dinosaurs: insights from the? Middle Jurassic Tafaytour tracksites (Argana Basin, Morocco). Journal of Vertebrate Paleontology 38 (5): e1512501. Crossref
Lapparent, A.F. de 1955. Étude paléontologique des vertébrés du Jurassique d’El Mers (Moyen Atlas). Notes et Mémoires du Service Géologique du Maroc 124: 1–36.
Lapparent, A.F. de 1960. Les dinosauriens du “Continental Intercalaire” du Sahara Central. Mémoires de la Société Géologique de France (Nouvelle Série) 88A: 1–56.
Lavocat, R. 1954. Sur les dinosauriens du Continental Intercalaire des Kem-Kem de la Daoura. Comptes Rendus 19th International Geological Congress 1952 (1): 65–68.
Läng, E. 2008. Les cétiosaures (Dinosauria, Sauropoda) et les sauropods du Jurassique moyen: révision systématique, nouvelles découvertes et implications phylogénétiques. Ph.D. Dissertation, Muséum National d’Histoire Naturelle, Paris.
Läng, E. and Mahammed, F. 2010. New anatomical data and phylogenetic relationships of Chebsaurus algeriensis (Dinosauria, Sauropoda) from the Middle Jurassic of Algeria. Historical Biology 22 (1–3): 142–164. Crossref
Le Loeuff, J., Métais, E., Dutheil, D.B., Rubino. J.L., Buffetaut, E., Lafont, F., Cavin, L., Moreau, F., Tong, H., Blanpied, C., and Sbeta, A. 2010. An Early Cretaceous vertebrate assemblage from the Cabao Formation of NW Libya. Geological Magazine 147: 750–759. Crossref
Maidment, S.C.R, Raven, T.J., Ouarhache, D., and Barrett, P.M. 2020. North Africa’s first stegosaur: implications for Gondwanan thyreophoran dinosaur diversity. Gondwana Research 77: 82–97. Crossref
Maidment, S.C.R, Strachan, S.J., Ouarhache, D., Scheyer, T.M., Brown, E.E., Fernandez, V., Johanson, Z., Raven, T.J., and Barrett, P.M. 2021. Bizarre dermal armour suggests the first African ankylosaur. Nature Ecology & Evolution 5: 1576–1581. Crossref
Maidment, S.C.R., Ouarhache, D., Butler R.J., Boumir K., Oussou A., Ech-charay K., El Khanchoufi, A., and Barrett, P.M. 2025. The world’s oldest cerapodan ornithischian dinosaur from the Middle Jurassic of Morocco. Royal Society Open Science 12: 241624. Crossref
Mannion, P.D. 2019. A turiasaurian sauropod dinosaur from the Early Cretaceous Wealden Supergroup of the United Kingdom. PeerJ 7: e6348. Crossref
Mannion, P.D. and Barrett, P.M. 2013. Additions to the sauropod dinosaur fauna of the Cenomanian (early Late Cretaceous) Kem Kem beds of Morocco: palaeobiogeographical implications of the mid-Cretaceous African sauropod fossil record. Cretaceous Research 45: 49–59. Crossref
Mannion, P.D., Upchurch, P., Schwarz, D., and Wings, O. 2019. Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: phylogenetic and biogeographic implications for eusauropod dinosaur evolution. Zoological Journal of the Linnean Society 185: 784–909. Crossref
Marty, D., Belvedere, M., Meyer, C.A., Mietto, P., Paratte, G., Lovis, C., and Thuering, B. 2010. Comparative analysis of Late Jurassic sauropod trackways from the Jura Mountains (NW Switzerland) and the central High Atlas Mountains (Morocco): implications for sauropod ichnotaxonomy. Historical Biology 22: 109–133. Crossref
Mateus, O., Mannion, P.D., and Upchurch, P. 2014. Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology 34: 618–634. Crossref
McPhee, B.W. and Choiniere, J.N. 2018. The osteology of Pulanesaura eocollum: implications for the inclusivity of Sauropoda (Dinosauria). Zoological Journal of the Linnean Society 182: 830–861. Crossref
Milàn, J. and Mateus, O. 2024. A Turiasaurian (Dinosauria, Sauropoda) tooth from the Pliensbachian Hasle Formation of Bornholm, Denmark, shows an Early Jurassic origin of the Turiasauria. Diversity 16 (1): 12. Crossref
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., Silva, B., and Ortega, F. 2016. Turiasauria-like teeth from the Upper Jurassic of the Lusitanian Basin, Portugal. Historical Biology 28: 861–880. Crossref
Monbaron, M., Russell, D.A., and Taquet, P. 1999. Atlasaurus imelakei ng, n. sp., a brachiosaurid-like sauropod from the Middle Jurassic of Morocco alen. Comptes Rendus de l’Académie des Sciences, Paris, Series IIA, Sciences de la Terre et des Planètes 329 (7): 519–526. Crossref
Nicholl, C.S., Mannion, P.D., and Barrett, P.M. 2018. Sauropod dinosaur remains from a new Early Jurassic locality in the Central High Atlas of Morocco. Acta Palaeontologica Polonica 63: 147–157. Crossref
Oukassou, M., Klein, H., Lagnaoui, A., Charrière, A., Saber, H., Gierliński, G.D., Lallensack, J.N., Hminna, A., Boumaalif, A., Oussou, A., and Ouarhache, D. 2019. Polyonyx-like tracks from Middle–?Upper Jurassic red beds of Morocco: Implications for sauropod communities on southern margins of Tethys. Palaeogeography, Palaeoclimatology, Palaeoecology 536: 109394. Crossref
Oussou, A., Falkingham P.L., Butler R.J., Boumir K., Ouarhache, D., Ech-charay, K., Charrière, A., and Maidment, S.C.R. 2023. New Middle to ?Late Jurassic dinosaur tracksites in the Central High Atlas Mountains, Morocco. Royal Society Open Science 10 (9): 231091. Crossref
Pereda Suberbiola, X., Bardet, N., Iarochène, M., Bouya, B., and Amaghzaz, M. 2004. The first record of a sauropod dinosaur from the Late Cretaceous phosphates of Morocco. Journal of African Earth Sciences 40: 81–88. Crossref
Pol, D., Ramezani, J., Gomez, K., Carballido, J.L., Carabajal, A.P., Rauhut, O.W.M., Escapa, I.H., and Cúneo, N.R. 2020. Extinction of herbivorous dinosaurs linked to Early Jurassic global warming event. Proceedings of the Royal Society B 287: 20202310. Crossref
Poropat, S.F., Frauenfelder, T.G., Mannion, P.D., Rigby, S.L., Pentland, A.H., Sloan, T., and Elliott, D.A. 2022. Sauropod dinosaur teeth from the lower Upper Cretaceous Winton Formation of Queensland, Australia and the global record of early titanosauriforms. Royal Society Open Science 9 (7): 220381. Crossref
Remes, K., Ortega, F., Fierro, I., Joger, U., Kosma, R., Marín Ferrer, J.M., Project PALDES, Niger Project SNHM, Ide, O.A., and Maga, A. 2009. A new basal sauropod dinosaur from the Middle Jurassic of Niger and the early evolution of Sauropoda. PLOS ONE 4 (9): e6924. Crossref
Régent, V., Wiersma-Weyand, K., Wings, O., Knötschke, N., and Sander, P.M. 2024. The dentition of the Late Jurassic dwarf sauropod Europasaurus holgeri from northern Germany: ontogeny, function, and implications for a rhamphotheca-like structure in Sauropoda. PeerJ 12: e17764. Crossref
Reolid, M., Ruebsam, W., and Benton, M.J. 2022. Impact of the Jenkyns Event (early Toarcian) on dinosaurs: Comparison with the Triassic/Jurassic transition. Earth-Science Reviews 234: 104196. Crossref
Royo-Torres, R. and Upchurch, P. 2012. The cranial anatomy of the sauropod Turiasaurus riodevensis and implications for its phylogenetic relationships. Journal of Systematic Palaeontology 10: 553–583. Crossref
Royo-Torres, R., Cobos, A., and Alcalá, L. 2006. A giant European dinosaur and a new sauropod clade. Science 314: 1925–1927. Crossref
Royo-Torres, R., Cobos, A., Mocho, P., and Alcalá, L. 2021. Origin and evolution of turiasaur dinosaurs set by means of a new ‘rosetta’ specimen from Spain. Zoological Journal of the Linnean Society 191: 201–227. Crossref
Royo-Torres, R., Upchurch, P., Kirkland, J.I., DeBlieux, D.D., Foster, J.R., Cobos, A., and Alcalá, L. 2017. Descendants of the Jurassic turiasaurs from Iberia found refuge in the Early Cretaceous of western USA. Scientific Reports 7 (1): 14311. Crossref
Sereno, P.C., Beck, A.L., Dutheil, D.B., Larsson, H.C., Lyon, G.H., Moussa, B., Sadleir, R.W., Sidor, C.A., Varricchio, D.J., Wilson, G.P., and Wilson, J.A. 1999. Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs. Science 286: 1342–1347. Crossref
Sereno, P.C., Wilson, J.A., Larsson, H.C., Dutheil, D.B., and Sues, H.-D. 1994. Early Cretaceous dinosaurs from the Sahara. Science 266: 267–271. Crossref
Sharma, A., Singh, S., and Satheesh, S.R. 2022. The first turiasaurian sauropod of India reported from the Middle Jurassic (Bathonian) sediments of Jaisalmer Basin, Rajasthan, India. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 304: 187–203. Crossref
Smith, J.B. and Dodson, P. 2003. A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. Journal of Vertebrate Paleontology 23: 1–12. Crossref
Termier, H. 1942. New data on the Red Jurassic and dinosaurs of the Great and Middle Atlas (Morocco). Bulletin of the Geological Society of France 5 (4–6): 199–207. Crossref
Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43–103. Crossref
Upchurch, P. and Barrett, P.M. 2000. The evolution of sauropod feeding mechanisms. In: H.-D. Sues (ed.), Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record, 79–122. Cambridge University Press, Cambridge. Crossref
Upchurch, P. and Martin, J. 2003) The anatomy and taxonomy of Cetiosaurus (Saurischia, Sauropoda) from the Middle Jurassic of England. Journal of Vertebrate Paleontology 23: 208–231. Crossref
Upchurch, P., Barrett, P.M., and Dodson, P. 2004. Sauropoda. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria. 2nd Edition, 259–322. University of California Press, Berkeley. Crossref
Wilson, J. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 215–275. Crossref
Wilson, J.A. and Allain, R. 2015. Osteology of Rebbachisaurus garasbae Lavocat, 1954, a diplodocoid (Dinosauria, Sauropoda) from the early Late Cretaceous–aged Kem Kem beds of southeastern Morocco. Journal of Vertebrate Paleontology 35 (4): e1000701. Crossref
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Memoir of the Society of Vertebrate Paleontology 5: 1–79. Crossref
Wiersma, K. and Sander, P.M. 2017. The dentition of a well-preserved specimen of Camarasaurus sp.: implications for function, tooth replacement, soft part reconstruction, and food intake. Paläontologische Zeitschrift 91: 145–161. Crossref
Xing, L., Miyashita, T., Currie, P.J., You, H., Zhang, J., and Dong, Z. 2015. A new basal eusauropod from the Middle Jurassic of Yunnan, China, and faunal compositions and transitions of Asian sauropodomorph dinosaurs. Acta Palaeontologica Polonica 60: 145–154.
Zafaty, O., Oukassou, M., Riguetti, F., Company, J., Bendrioua, S., Tabuce, R., Charrière, A., and Pereda-Suberbiola, X. 2024. A new stegosaurian dinosaur (Ornithischia: Thyreophora) with a remarkable dermal armour from the Middle Jurassic of North Africa. Gondwana Research 131: 344–362. Crossref
Acta Palaeontol. Pol. 70 (3): 411–420, 2025
https://doi.org/10.4202/app.01214.2024