
A new harpactorin hemipteran insect from the Miocene Dominican amber with fossula spongiosa on all three pairs of legs
PEIPEI ZHANG, YUNZHI YAO, ZHENGKUN HU, DONG REN, and YINGQI LIU
Zhang, P., Yao, Y., Hu, Z., Ren D., and Liu, Y. 2025. A new harpactorin hemipteran insect from the Miocene Dominican amber with fossula spongiosa on all three pairs of legs. Acta Palaeontologica Polonica 70 (2): 357–368.
A new genus and species of fossil harpactorin (Hemiptera: Reduviidae), Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov., is described from Miocene Dominican amber, representing the third fossil record of Harpactorinae. The new species is remarkable for exhibiting fossula spongiosa on all three pairs of legs, which is not only the first report in Harpactorinae but also rare within Reduviidae. This structure is considered to be related to the locomotor capabilities of assassin bugs, potentially enhancing their attachment and agility in navigating complex surfaces and vegetation. Furthermore, the present study suggests that fossula spongiosa exhibits remarkable plasticity within Reduviidae by integrating fossil and extant perspective.
Key words: Insecta, Reduviidae, Harpactorinae, assassin bug, fossula spongiosa, plasticity, Miocene, Dominican amber.
Peipei Zhang [2230801008@cnu.edu.cn; ORCID: https://orcid.org/0009-0006-7067-3464 ], Yunzhi Yao [yaoyz100@126.com; ORCID: https://orcid.org/0000-0003-4491-0260 ], Dong Ren [rendong@cnu.edu.cn; ORCID: https://orcid.org/ 0000-0001-8660-0901 ], and Yingqi Liu [yingqiliu0720@163.com; ORCID: https://orcid.org/0000-0003-0881-9670 ] (corresponding author), College of Life Sciences, Capital Normal University, Xisanhuanbeilu 105, Haidian District, Beijing 100048, China.
Zhengkun Hu [1428101617@qq.com; ORCID: https://orcid.org/0000-0002-7470-0830 ], Fanjingshan National Nature Reserve, Tongren 554400, China.
Received 27 November 2024, accepted 25 April 2025, published online 30 June 2025.
Copyright © 2025 P. Zhang et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Reduviidae are the most diverse predatory insects among Hemiptera, having evolved a wide range of morphological characters for capturing prey. The fossula spongiosa, a hairy pad-like attachment structure located on the ventral surface of the distal part of tibia, is the most typical (Haridass and Ananthakrishnan 1980; Maldonado-Capriles 1990; Cassis and Gross 1996; Weirauch et al. 2014). Weirauch (2007) investigated the distribution of fossula spongiosa on the three pairs of legs of 171 species across 22 reduviid subfamilies. The results showed that the distribution of fossula spongiosa in extant Reduviidae can be broadly divided into four types: (ⅰ) lack of fossula spongiosa; (ⅱ) fossula spongiosa only on the foreleg; (ⅲ) fossula spongiosa on the foreleg and midleg; and (ⅳ) fossula spongiosa on all three pairs of legs. Type (ⅳ) is exceedingly uncommon among Reduviidae, occurring solely in the genera of Microtriatoma Prosen & Martínez, 1952, and Parabelminus Lent, 1943, within Triatominae (Sherlock and Guitton 1982; Lent 1943; Lent and Jurberg 1984; Weirauch 2007; Bargues et al. 2017). It is noteworthy that all investigated species of Harpactorinae in the above study had no fossula spongiosa in all three pairs of legs, and only the distal portion of the tibiae of the foreleg was more or less densely covered with setae (Weirauch 2007). In particular, Masonick et al. (2024) showed that a key diagnostic character of Harpactorinae is the lack of fossula spongiosa on the tibiae in most taxa.
Harpactorinae with more than 2250 species in over 300 genera, represents the largest subfamily within Reduviidae (Masonick et al. 2024). However, the fossil record of this subfamily is very sparse, with only two genera and two species known from the Cenozoic. Aphelicophontes danjuddi Swanson, Heads, Taylor & Wang, 2021, from the Eocene Green River impression fossil in the United States, has tibiae which are incompletely preserved thus the distribution of fossula spongiosa is unknown; the other species is Apicrenus fossilis Capriles, Santiago-Blay & Poinar, 1993, from the Miocene Dominican amber, which does not have fossula spongiosa like most extant Harpactorinae species (Maldonado-Capriles et al. 1993a; Swanson et al. 2021).
This study presents a well-preserved specimen of Harpactorinae in Miocene Dominican amber, identified and described as a new genus and species. The new fossil preserves very distinct, pad-like fossula spongiosa on all three pairs of legs, being the first harpactorin species with fossula spongiosa on all three pairs of legs. This discovery not only enriches the fossil record of Harpactorinae to some extent but also broadens our understanding of the distribution pattern of this structure within Reduviidae. Furthermore, it provides valuable insights for further understanding the evolution of fossula spongiosa within both Harpactorinae and the entire Reduviidae.
Institutional abbreviations.—CNU, Capital Normal University, Beijing, China; FNNR, Fanjingshan National Nature Reserve, Tongren, China.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in Zoobank: urn:lsid:zoobank.org:pub:8750ADCD-8A50-42DA-B3CD-135721EB98D4.

Fig. 1. Harpactorin reduviid bug Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov., holotype, CNU-HET-DM2024011 from Burdigalian, Lower Miocene; Cordillera Septentrional, Dominican Republic. Dorsal (A1) and ventral (A2) views.
Material and methods
The holotype was purchased by Zhengkun Hu in the Dominican Republic in 2009 and subsequently donated to Professor Yunzhi Yao of CNU. The specimen originates from the Cordillera Septentrional, situated between Puerto Plata and Santiago in the northern Dominican Republic. Among the worldwide fossil resin occurrences, the Dominican amber deposits are considered to be one of the largest (Grimaldi 1995; Stach et al. 2021). They originated from deciduous trees of genus Hymenaea, which is similar to Acacia, but their age is still in a dispute, and is estimated as Miocene 20–15 Ma, 15.75–12.58 Ma, 25–20 Ma or Pliocene–Early Pleistocene (Stach et al. 2020, 2021; Hernández-Damián et al. 2024). These age discrepancies likely reflect limitations in dating methodologies or sample sources. Currently the more accepted view is that its geological age is Early Miocene (Schlee 1990; Grimaldi 1995; Iturralde-Vincent and Macphee 1996; Penney et al. 2010; Iturralde-Vincent and Macphee 2019; Stach et al. 2020, 2021).
This study conducted further morphological research on the materials using the Nikon SMZ25 microscope, which was connected in real-time to the Nikon DS-Ri2 digital photographic system. Sketches were drawn using the microscope’s drawing arm, scanned with a printer, and then imported into drawing software to create line reconstruction drawings. The morphological terminology used in this study primarily followed Lent and Wygodzinsky (1979), Weirauch (2008), and Zhang and Weirauch (2011).
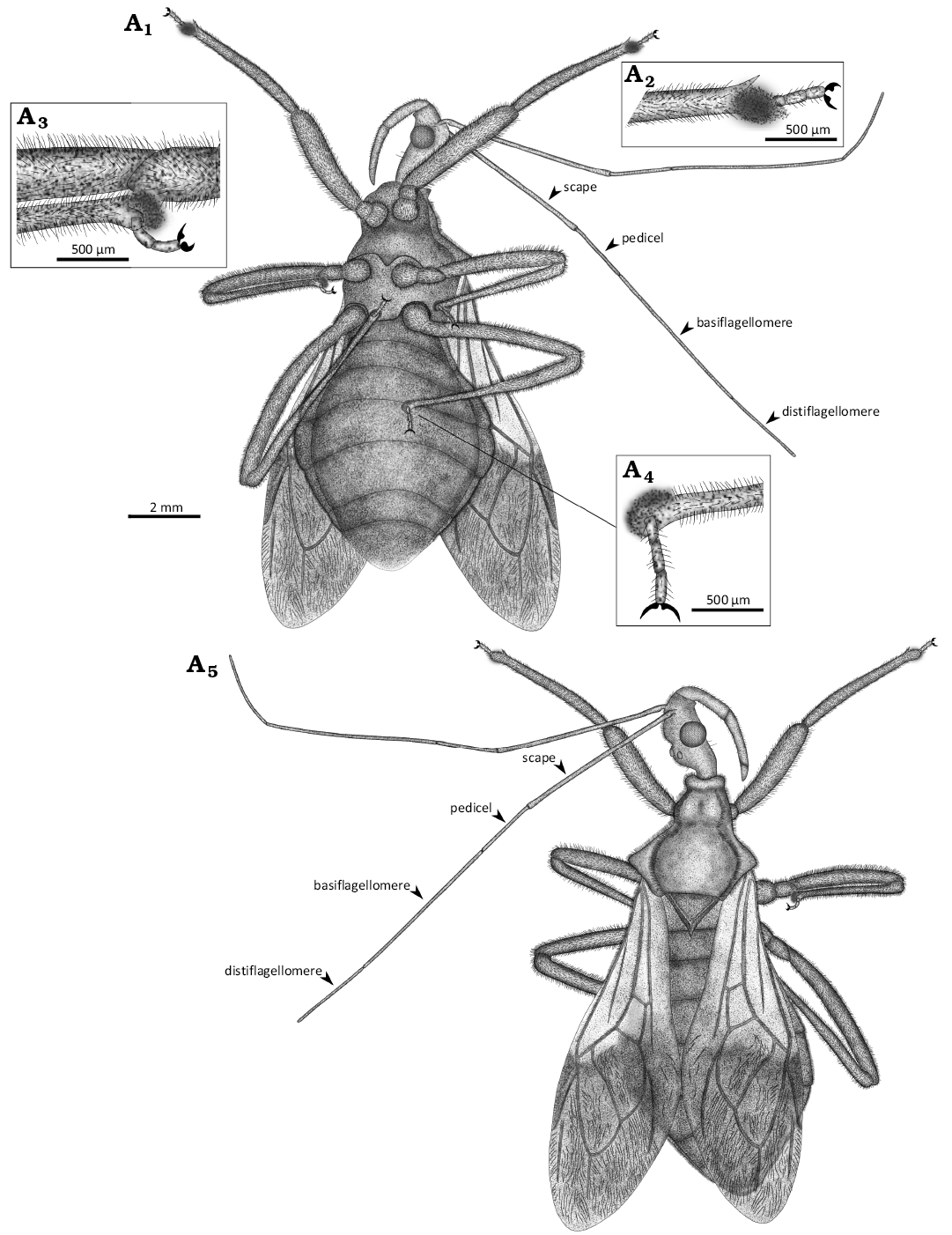
Fig. 2. The reconstruction of harpactorin reduviid bug Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov., holotype, CNU-HET-DM2024011, from Burdigalian Lower Miocene; Cordillera Septentrional, Dominican Republic. Line drawing of whole specimen in ventral (A1) and dorsal (A5) views; fossula spongiosa of foreleg (A2); fossula spongiosa of midleg (A3); fossula spongiosa of hindleg (A4).
Systematic palaeontology
Order Hemiptera Linnaeus, 1758
Suborder Heteroptera Latreille, 1810
Infraorder Cimicomorpha Leston, Pendergrast & Southwood, 1954
Family Reduviidae Latreille, 1807
Subfamily Harpactorinae Amyot & Serville, 1843
Tribe Harpactorini Amyot & Serville, 1843
Genus Trispongiosus Zhang, Yao, & Liu nov.
Zoobank LSID: urn:lsid:zoobank.org:act:21BB5754-B477-4848-98B4- F3E5CEF35C42.
Type species: Trispongiosus hui Zhang, Yao, & Liu, sp. nov., monotypic; see below.
Etymology: From Latin tri-, three, and spongiosus, spongy, in refrence to having fossula spongiosa on all three pairs of legs. Gender masculine.
Diagnosis.—As for the monotypic type species.
Remarks.—According to the studies of Weirauch et al. (2014), Schuh and Weirauch (2020), and Masonick et al. (2024), Trispongiosus Zhang, Yao, & Liu gen. nov. should be placed in Harpactorinae because of the following characters: (i) head with postocular part cylindrical; (ii) anteocular region noticeably shorter than postocular region; (iii) sclerotized antennal basiflagellomere and distiflagellomere lacking narrow membranous rings; (iv) foretibia with a well-developed apical spur and (v) hemelytron with quadrate cell distally on corium. Trispongiosus Zhang, Yao, & Liu gen. nov. is placed in Harpactorini on the basis of the following characters: (i) labium curved; (ii) apex of medial posterior process of scutellum blunt; (iii) forefemur without spined; (iv) claws with basal tooth and (v) posterior cell of membrane subequal in size to anterior cell, with apex bluntly pointed and subtriangular.
Trispongiosus Zhang, Yao, & Liu gen. nov. shares the following characters with Rocconota Stål, 1859 (Ramírez Silva et al. 2022), a genus within Harpactorini that is primarily distributed in Panama: (i) head elongate; (ii) postocular portion of head narrowing gradually to form collum in dorsal view; (iii) antennal scape longest; (iv) pedicel shortest; (v) basiflagellomere slightly shorter than scape; (vi) distiflagellomere slightly longer than pedicel; (vii) legs elongated and slender; (viii) female with a conspicuously enlarged abdomen, maximum width of abdomen significantly greater than width across humeral angles. Nevertheless, the two can be distinguished by the following characters: Rocconota with (i) short conical tubercle on anterior angles of pronotum and sharp spines on humeral angles; (ii) posterior lobe with two symmetrical spines on central posterior position; Trispongiosus Zhang, Yao, & Liu gen. nov. with (i) blunt, rounded, anterior and humeral angles; (ii) posterior lobe without two symmetrical spines. Hemelytron of Rocconota in addition is uniformly dark brown, without pterostigmata, and the coloration of corium and membrane of Trispongiosus Zhang, Yao, & Liu gen. nov. is uneven, with dark pterostigmata.

Fig. 3. The detailed partial view of harpactorin reduviid bug Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov., holotype, CNU-HET-DM2024011 from Burdigalian, Lower Miocene; Cordillera Septentrional, Dominican Republic. A. Pronotum and scutellum. B. Foreleg. C. Labium. D. Hemelytron. E. Midleg. F. Hindleg. G. Ocelli. Arrows in B2 and E2 indicates the basal tooth of claw, arrow in F points to the blunt tooth on the hind tarsal claw. Scale bars 1 mm. Abbreviations: Cu, cubitus; Cupcu1, Cupcu2, first and second vein formed by the fusion of the cubitus and postcubitus; M, media; Pcu, postcubitus; Pcu1A, fusion of the postcubitus and the first anal vein; R: radius; 1A: first anal vein.
Trispongiosus Zhang, Yao, & Liu gen. nov. shares the following characters with Parahiranetis Gil-Santana, 2015, which belongs to the wasp-mimicking Harpactorini in the Neotropical Region: (i) head elongate; (ii) postocular portion of head narrowing gradually to form collum in dorsal view, without postantennal spines; (iii) visible segment I of labium subequal to length of visible segment II; (iv) visible segment III shortest and (v) fore femur subequally longer than head and pronotum together. Nevertheless, the two can be distinguished by the following characters: Parahiranetis with (i) pubescence of long hairs on postocular portion; females of Parahiranetis with (ii) narrower abdomen and (iii) maximum width of abdomen subequal to width across humeral angles; (iv) hemelytron of Parahiranetis dark brown overall with bright yellow pterostigmata in specific positions. While in Trispongiosus Zhang, Yao, & Liu gen. nov., (i) the whole head is covered with hairs; (ii) the abdomen is conspicuously enlarged and (iii) maximum width of abdomen is significantly greater than width across humeral angles; (iv) hemelytron of retains pterostigmata in same positions but with color noticeably darker than surrounding wing surface. It is of particular significance that Trispongiosus Zhang, Yao, & Liu gen. nov. exhibits distinct fossula spongiosa on all three pairs of legs, which is quite unique within Harpactorinae. Therefore, a new genus was finally established based on the fossil specimen.
Stratigraphic and geographic range.—In the northern mountain range (Cordillera Septentrional) of the Dominican Republic between the cities of Puerto Plata and Santiago. Burdigalian, Lower Miocene.
Trispongiosus hui Zhang, Yao, & Liu sp. nov.
Figs. 1–3.
Zoobank LSID: urn:lsid:zoobank.org:act:6147E673-2167-4720-82EE-82FD9AD2B014.
Etymology: Dedicated to Zhengkun Hu of the Fanjingshan National Nature Reserve (Tongren, China) for his donation of this amber specimen.
Holotype: CNU-HET-DM2024011, an adult female.
Type locality: The northern mountain range (Cordillera Septentrional) of the Dominican Republic between the cities of Puerto Plata and Santiago. Details unknown.
Type horizon: Burdigalian, Lower Miocene.
Diagnosis.—Body medium-sized; head oval, length of head slightly shorter than length of pronotum; posterior lobe of pronotum with parentheses-shaped indentations on both sides, humeral angles blunt; pronotum and each leg densely covered with short, decumbent to suberect, curly setae; membrane with external cell longer and narrower than internal cell; basal teeth of tarsi of foreleg and midleg rounded.
Description.—Macropterous female. Body medium-sized.
Vestiture: Head with dense, short, suberect to erect, curly setae on dorsal and lateral surfaces; antennae densely covered with very short, decumbent to suberect setae; labium with short, suberect to erect setae; pronotum and legs densely covered with short, decumbent to suberect, curly setae.

Fig. 4. Fossula spongiosa on the forelegs and midlegs of 11 yet undescribed species of fossil reduviids from Upper Cretaceous Burmese amber. A. CNU-HET-MA2025003, sp. 1. B. CNU-HET-MA2025004, sp. 2. C. CNU-HET-MA2025005, sp. 3. D. CNU-HET-MA2025006, sp. 4. E. CNU-HET-MA2025007, sp. 5. F. CNU-HET-MA2025008, sp. 6. G. CNU-HET-MA2025009, sp. 7. H. CNU-HET-MA2025010, sp. 8. I. CNU-HET-MA2025011, sp. 9. J. CNU-HET-MA2025012, sp. 10. K. CNU-HET-MA2025013, sp. 11. H1, J1, K1, forelegs; H2, J2, K2, midlegs. Black arrows indicate the fossula spongiosa on the forelegs, while white arrows indicate those on the midlegs. Scale bars 250 µm.
Structure: Body oval. Head elongate, with dorsum weakly inflated and venter flattened; postocular part 1.2 times as long as anteocular part; eyes slightly protruding anterolaterally; ocelli small, slightly elevated. Antennal scape longest, 2.0 times as long as length of head, base of scape notched, pedicel shortest, 0.7 times as long as length of head, basiflagellomere slightly shorter than scape, 1.9 times as long as length of head, distiflagellomere as long as length of head; labium curved, slightly surpassing anterior margin of prosternum, visible segment I subequal to length of visible segment II, visible segment III shortest, conical, 0.3 times as long as visible segment I.
Collar processes obtuse horn-shaped, produced laterally; length of pronotum along midline just as long as width across humeral angles, 1.5 times as long as length of head; anterior margin slightly concave, anterior lobe round and bulged, 0.5 times as long as posterior lobe, with deep median longitudinal sulcus; posterior lobe separated from anterior lobe by faint transverse sulcus; posterior lobe with parentheses-shaped indentations on both sides, anterior region of posterior lobe with shallow median longitudinal groove, humeral angles blunt, posterior margin nearly straight, lateral margin finely concave. Scutellum partially exposed, Y-shaped ridges narrow, scutellar process rounded.
Fore coxa spherical, fore femur thickened, somewhat flattened laterally, fore tibia slender, slightly curved, 1.2 times as long as fore femur, fore tarsus three-segmented, with a pair of tiny claws at apex; mid and hind coxae widely separated from each other; mid and hind femora moderately and strongly thickened, mid tibia 1.1 times as long as mid femur, hind tibia 1.2 times as long as hind femur, mid and hind tarsi three-segmented, with a pair of small claws at apices, base of claw with a relatively rounded and blunt tooth. Each tibia with fossula spongiosa approximately 0.1 times as long as of total length of each tibia.
Hemelytron slightly extending beyond abdominal tip, corium with quadrate cubitalcell, membrane with external cell longer and narrower than internal cell.
Abdomen broad oval, length 1.6 times as long as its maximum width, connexivum clearly extended outwards on both sides.
Dimensions.—Table 1.
Stratigraphic and geographic range.—In the northern mountain range (Cordillera Septentrional) of the Dominican Republic between the cities of Puerto Plata and Santiago. Burdigalian, Lower Miocene.
Table 1. Measurements (in mm) of Trispongiosus hui (CNU-HET-DM2024011).
|
Body part |
Measurement |
|
|
body |
length |
14.27 |
|
head |
length |
2.43 |
|
anteocular part |
length |
0.84 |
|
postocular part |
length |
0.98 |
|
antennal segments I–IV |
length |
4.94; 1.57; 4.68; 2.51 |
|
visible labial segments I–III |
length |
1.61; 1.63; 0.55 |
|
anterior lobe of pronotum |
length |
1.23 |
|
width |
1.70 |
|
|
posterior lobe of pronotum |
length |
2.32 |
|
width |
3.50 |
|
|
scutellum |
length |
1.17 |
|
width |
1.74 |
|
|
hemelytron |
length |
10.05 |
|
abdomen |
length |
8.60 |
|
width |
5.24 |
|
|
fore: femur; tibia; tarsus |
length |
3.37; 4.16; 0.41 |
|
mid: femur; tibia; tarsus |
length |
3.59; 4.05; 0.78 |
|
hind: femur; tibia; tarsus |
length |
4.93; 6.12; 1.01 |
Discussion
Evolutionary plasticity of fossula spongiosa in Reduviidae.—The fossula spongiosa, a cushion-like expanded area on the tibia that is composed of thousands of minute hairs, and represents a key morphological character of Cimicomorpha (Weirauch 2007; Schuh et al. 2009). This structure is often conspicuous within Reduviidae, and is thought to be associated primarily with predatory behavior and sometimes with mating and locomotor functions (Haridass and Ananthakrishnan 1980; Weirauch 2007; Zhang et al. 2016). This study surveys the distribution of fossula spongiosa with hairy pad-like attachment structure on each leg of previously reported reduviid fossils, a newly described fossil within this paper, and 11 unpublished reduviid fossils from Burmese amber (Fig. 4, Table 2). The results show that the majority of currently known Cretaceous Reduviidae have fossula spongiosa on both the forelegs and midlegs (Fig. 5). A phylogenetic analysis of the evolution of raptorial leg in Reduviidae by Zhang et al. (2016) suggests that fossula spongiosa is a plesiomorphy of Reduviidae. Zhang et al. (2022) described the oldest fossils of Reduviidae, concluded that the presence of fossula spongiosa on both the foreleg and the midleg was the ancestral state of fossula spongiosa of Reduviidae. The statistical results of the present study further verify the above conclusions.
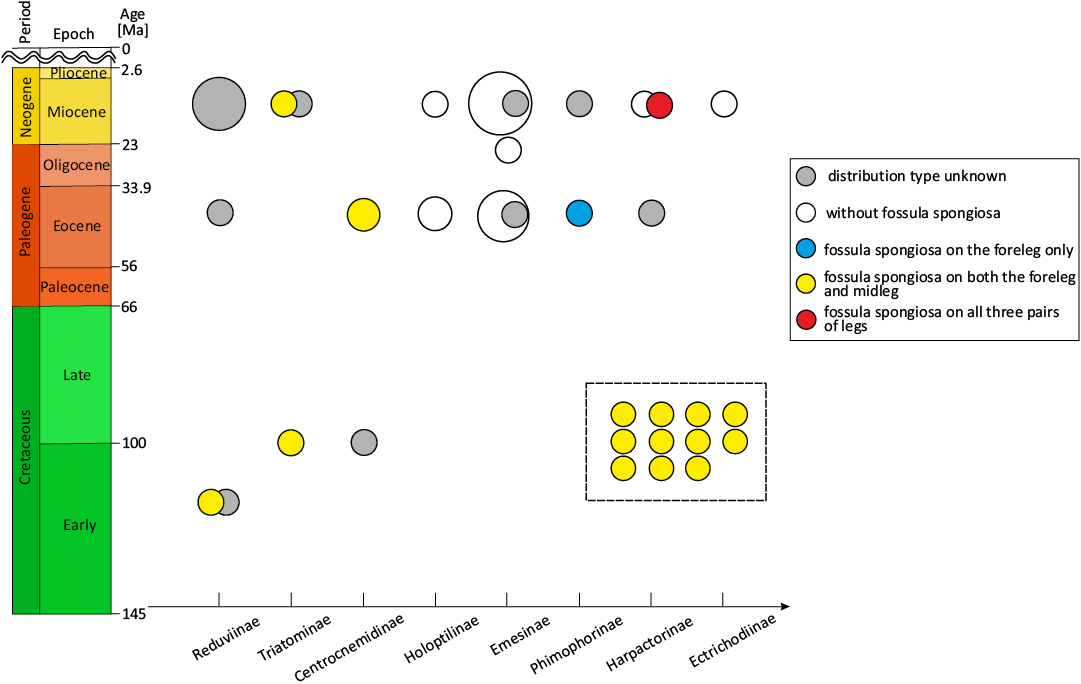
Fig. 5. Distribution pattern of fossula spongiosa in fossil reduviids. Circle size reflects the number of fossils discovered, with larger circles indicating more fossils. The dashed box indicate the distribution patterns of fossula spongiosa in 11 unpublished fossil reduviids. Fossils of which the subfamilies and fossula spongiosa type are both unknown were excluded from statistical analyses.
In addition, the results show that these fossils encompass all types of fossula spongiosa observed in extant Reduviidae: total lack of fossula spongiosa; only foreleg with fossula spongiosa; foreleg and midleg with fossula spongiosa; foreleg, midleg and hindleg with fossula spongiosa (Table 2). Excluding fossils with both undetermined subfamily and fossula spongiosa type, the types of fossula spongiosa in Reduviidae show a gradual trend of diversification over geological time (Fig. 5). During the Cretaceous, fossula spongiosa was relatively uniform in type, present only on the fore- and midlegs. However, since the Eocene, various types of fossula spongiosa began to appear in the fossil record, including the lack of fossula spongiosa, with fossula spongiosa only on the forelegs, and fossula spongiosa on both the foreleg and midleg. By the Miocene, the types of fossula spongiosa in Reduviidae became even more diverse, encompassing all types seen in extant species, ranging from no fossula spongiosa to fossula spongiosa on all three pairs of legs.
Table 2. Distribution of fossula spongiosa on each leg in Reduviidae fossils. + presence; – absence; ? unknown.
|
Epoch |
Subfamily |
Species |
Foreleg |
Midleg |
Hindleg |
Reference |
|
Miocene |
Reduviinae |
Reduvius diatomus |
? |
? |
? |
|
|
Reduvius shandongianus |
? |
? |
? |
|||
|
Triatominae |
Triatoma dominicana |
? |
? |
? |
||
|
Panstrongylus hispaniolae |
+ |
+ |
– |
|||
|
Holoptilinae |
Praecoris dominicana |
– |
– |
– |
||
|
Emesinae |
Emesites voigti |
? |
? |
? |
||
|
Empicoris copal |
– |
– |
– |
|||
|
Malacopus wygodzinskyi |
– |
– |
– |
|||
|
Alumeda antilliana |
– |
– |
– |
|||
|
Alumeda dominicana |
– |
– |
– |
|||
|
Alumeda nigricans |
– |
– |
– |
|||
|
Paleoploiariola venosa |
– |
– |
– |
|||
|
Empiploiariola inermis |
– |
– |
– |
|||
|
Phimophorinae |
Phimophorus chiodii |
? |
? |
? |
||
|
Harpactorinae |
Apicrenus fossilis |
– |
– |
– |
||
|
Trispongiosus hui |
+ |
+ |
+ |
this paper |
||
|
Harpactor chomeraciensis |
? |
? |
? |
|||
|
Arilus faujasi |
? |
? |
? |
|||
|
Ectrichodiinae |
Ectrichodiella electrina |
– |
– |
– |
||
|
unknown |
Oncocephalus astutus |
? |
? |
? |
||
|
Reduvius immitus |
? |
? |
? |
|||
|
Reduvius nicus |
? |
? |
? |
|||
|
Reduvius piceus |
? |
? |
? |
|||
|
Oligo-Miocene |
Emesinae |
Empicoris electricus |
– |
– |
– |
|
|
Oligocene |
unknown |
Rhinocoris michalki |
? |
? |
? |
|
|
Eocene |
Reduviinae |
Platymeris insignis |
? |
? |
? |
|
|
Centrocnemidinae |
Redubitus centrocnemarius |
+ |
+ |
– |
||
|
Redubinotus liedtke |
+ |
+ |
– |
|||
|
Holoptilinae |
Proptilocerus dolosus |
– |
– |
– |
||
|
Proptilocnemus longispinis |
– |
– |
– |
|||
|
Emesinae |
Collarhamphus mixtus |
? |
? |
? |
||
|
Amphibolus disponsi |
– |
– |
– |
|||
|
Danzigia christelae |
– |
– |
– |
|||
|
Emesopsis putshkovi |
– |
– |
– |
|||
|
Emesopsis similis |
– |
– |
– |
|||
|
Phimophorinae |
Koenigsbergia herczeki |
+ |
– |
– |
||
|
Harpactorinae |
Aphelicophontes danjuddi |
? |
? |
? |
||
|
unknown |
Tagalodes inermis |
? |
? |
? |
||
|
Poliosphageus psychrus |
? |
? |
? |
|||
|
Paleocene |
unknown |
Hymenopterites deperditus |
? |
? |
? |
|
|
Cretaceous |
Reduviinae |
Simplicivenius rectidorsius |
? |
? |
? |
|
|
Simplicivenius tuberculosus |
+ |
+ |
– |
|||
|
Triatominae |
Paleotriatoma metaxytaxa |
+ |
+ |
– |
||
|
Centrocnemidinae |
Acutiangulus densus |
? |
? |
? |
||
|
unknown |
sp. 1 |
+ |
+ |
– |
unpublished |
|
|
sp. 2 |
+ |
+ |
– |
unpublished |
||
|
sp. 3 |
+ |
+ |
– |
unpublished |
||
|
sp. 4 |
+ |
+ |
– |
unpublished |
||
|
sp. 5 |
+ |
+ |
– |
unpublished |
||
|
sp. 6 |
+ |
+ |
– |
unpublished |
||
|
sp. 7 |
+ |
+ |
– |
unpublished |
||
|
sp. 8 |
+ |
+ |
– |
unpublished |
||
|
sp. 9 |
+ |
+ |
– |
unpublished |
||
|
sp. 10 |
+ |
+ |
– |
unpublished |
||
|
sp. 11 |
+ |
+ |
– |
unpublished |
There are significant differences in the type of fossula spongiosa in extant Reduviidae, even within the same subfamily, e.g., the majority of species in Peiratinae have fossula spongiosa on the foreleg and midleg, whereas the genus Sirthenea Spinola, 1837, has fossula spongiosa on the foreleg only, with the exception of S. laevicollis Horváth, 1909, which occurs in Australia, possessing fossula spongiosa on both the foreleg and midleg (Weirauch 2007; Chłond et al. 2019). Additionally, the present study identifies a fossil within Harpactorinae that exhibits pad-like fossula spongiosa on all three pairs of legs, a feature that is traditionally absent in this subfamily. These observations suggest the significant diversity and plasticity of fossula spongiosa within Reduviidae. Consequently, the present study proposes that the evolution of fossula spongiosa within the Reduviidae was not following a linear direction by integrating fossil and extant perspectives. Zhang et al. (2016) suggested that the evolution of fossula spongiosa may be related to complex interrelated factors associated with prey type, predatory behavior, salivary toxicity, and morphological adaptation. Building on their work, this study further hypothesize that the plasticity of fossula spongiosa evolution in Reduviidae likely represents a flexible adaptation to varying ecological conditions and predatory needs.
Exploration of the adaptive functions of fossula spongiosa on all three pairs of legs.—It has been shown that fossula spongiosa enhances the gripping efficiency of the legs and is sexually dimorphic in some bedbugs (Haridass and Ananthakrishnan 1980; Reinhardt et al. 2019). Fossula spongiosa in some male bedbugs possess a higher density of setae compared to females, which results in greater adhesive strength (Reinhardt et al. 2019). This adaptation likely aids males in securely grasping females during traumatic insemination (Jung et al. 2023). Chłond et al. (2019) suggested that the distribution of fossula spongiosa in Peiratinae is closely related to their ecological environments, with the reduction of fossula spongiosa to only the forelegs in Sirthenea possibly being an adaptation to warm and humid ecological niches with abundant prey species and lower levels of competition.
The most remarkable character of Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov. is the presence of distinct and roughly uniformly-sized fossula spongiosa on all three pairs of legs, which is for the first time recorded in Harpactorinae. Currently, the presence of fossula spongiosa in all three pairs of legs within Reduviidae is also very rare, with similar phenomena observed only in species of two genera of Triatominae, i.e., Parabelminus and Microtriatoma (Lent 1943; Weirauch 2007; Bargues et al. 2017).
Parabelminus occurs in southeastern Brazil, while Microtriatoma species have a wider distribution, covering Central America (such as Costa Rica and Panama) and South America (such as Trinidad, Peru, Bolivia, as well as northern and southeastern Brazil) (Lent 1943; Sherlock and Guitton 1982; De la Riva et al. 2001; Abad-Franch and Monteiro 2007; Bargues et al. 2017; Dos Santos Souza et al. 2019; Hylton et al. 2020; Gil-Santana et al. 2021; Kieran et al. 2021; Galvão et al. 2024). It seems that Reduviidae with fossula spongiosa on all three pairs of legs are known to be distributed in tropical and subtropical regions. However, this is consistent with the ecological niche of Reduviidae with underdeveloped fossula spongiosa as indicated by Chłond et al. (2019). It should be noted that the study of Chłond et al. (2019) focused on Peiratinae, which is predominantly ground-dwelling (Weirauch et al. 2014). Given their habitat, Peiratinae may not rely heavily on fossula spongiosa to enhance grip on rough ground surfaces. The primary role of fossula spongiosa of Peiratinae is more likely to aid in predation. Nevertheless, most members of Harpactorinae are arboreal, living on plant surfaces (Weirauch et al. 2014). Many extant species within this subfamily lack fossula spongiosa, which may suggest a reduced reliance on fossula spongiosa for predation in Harpactorinae. Additionally, some lineages have evolved sticky predation strategies, which may have partially supplanted the predatory function of fossula spongiosa (Zhang and Weirauch 2013; Weirauch et al. 2014; Masonick et al. 2024). Triatominae members with fossula spongiosa on all three pairs of legs have been primarily found on smooth plant surfaces, such as Microtriatoma borbai Lent & Wygodzinsky, 1979, collected from a hybrid eucalyptus crop, and Microtriatoma trinidadensis Lent, 1951, found on palm leaves (Dos Santos Souza et al. 2019; Gil-Santana et al. 2021). As hematophagous insects, Triatominae rely less on their legs for capturing prey, but instead use their labium to feed on the blood of their prey (Weirauch et al. 2014; Masonick et al. 2024). These implies that fossula spongiosa may have a different major function in these two subfamilies compared to Peiratinae. Furthermore, the study by Haridass and Ananthakrishnan (1980) demonstrated that fossula spongiosa of some reduviids generate greater static tension on smooth surfaces. In conclusion, this study hypothesizes that Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov. with fossula spongiosa on all three pairs of legs is more likely to be arboreal. The presence of fossula spongiosa on all legs may be associated with locomotor functions, such as enhanced adhesion and movement. In tropical and subtropical regions which are characterized by warm and humid climates and a high diversity of plant species, assassin bugs may need to adapt to various plant surfaces, some of which can be relatively smooth, such as the leaves of certain palm species. The fossula spongiosa on all three pairs of legs can enhance the attachment ability of assassin bugs on these smooth surfaces, thereby increasing their flexibility in complex ecological niches and effectively aiding in prey pursuit and predator evasion. In addition, extant predatory Reduviidae usually immobilize their prey using the foreleg and midleg, and the fossula spongiosa of foreleg is generally slightly larger than the midleg (Weirauch 2007; Zhang et al. 2022). However, in Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov., and species of Parabelminus, and Microtriatoma, the hind legs, which are not involved in predation, also possess fossula spongiosa that are comparable in size to those on the forelegs and midlegs. This phenomenon further implies that fossula spongiosa present on all three pairs of legs may play a major role in locomotion. However, direct evidence for its locomotor function is still unavailable. Future research should incorporate extensive fossil records and experimental approaches to elucidate its precise role and evolutionary importance.
Conclusions
This study describes a new genus and species of Harpactorinae, Trispongiosus hui Zhang, Yao, & Liu gen. et sp. nov., discovered in Miocene Dominican amber. The new species is remarkable for possessing distinct fossula spongiosa on all three pairs of legs, a character recorded for the first time in Harpactorinae and also rare within Reduviidae. This study proposes that the structure could contribute to the locomotor abilities of assassin bugs, enhancing their grip and maneuverability on challenging surfaces and dense vegetation. Additionally, the research highlights the significant plasticity of the fossula spongiosa within Reduviidae, as demonstrated through the integration of fossil and extant perspectives. This discovery not only expands the fossil record of Harpactorinae but also provides new perspectives and evidence for understanding the evolution of fossula spongiosa within Reduviidae.
Acknowledgements
We are grateful to Qi Zuo (College of Life Sciences, Capital Normal University, Beijing, China) for her assistance in photographing the specimen. We also express our sincere gratitude to the editor and two anonymous reviewers for their constructive criticism and valuable suggestions. This research was funded by the National Natural Science Foundation of China (No. 32020103006).
Editor: Krzysztof Hryniewicz.
References
Abad-Franch, F. and Monteiro, F.A. 2007. Biogeography and evolution of Amazonian triatomines (Heteroptera: Reduviidae): implications for Chagas disease surveillance in humid forest ecoregions. Memórias do Instituto Oswaldo Cruz 102: 57–70. Crossref
Bargues, M.D., Schofield, C., and Dujardin, J.P. 2017. Classification and systematics of the Triatominae. In: J. Telleria and M. Tybairenc (eds.), American trypanosomiasis chagas disease (2nd Edition) One Hundred Years of Research, 113–143. Elsevier, London. Crossref
Boderau, M., Ngô-Muller, V.A., Peng, A.C., Engel, M.S., Garrouste, R., and Nel, A. 2024. Phimophorus chiodii sp. nov., first representative of a cryptic assassin bug subfamily from Dominican amber (Reduviidae: Phimophorinae) as revealed with traditional light microscopy and computed micro-tomographic reconstruction. Palaeoentomology 7: 299–305. Crossref
Bush, T., Berenger, J.M., Gil-Santana, H., Forthman, M., Hoey-Chamberlain, R., and Weirauch, C. 2024. Systematics of Ectrichodiella Fracker and Bruner, 1924, with description of the first fossil millipede assassin bug species (Insecta: Hemiptera: Reduviidae: Ectrichodiinae). American Museum Novitates 4025: 1–40. Crossref
Cassis, G. and Gross, G.F. 1996. Hemiptera: Heteroptera (Coleorrhyncha to Cimicomorpha). Journal of the New York Entomological Society 104: 113–115.
Chłond, D., Sawka-Gądek, N., and Żyła, D. 2019. Genetic data of museum specimens allow for inferring evolutionary history of the cosmopolitan genus Sirthenea (Heteroptera: Reduviidae). PeerJ 7: e6640. Crossref
De la Riva, J., Matias, A., Torrez, M., Martínez, E., and Dujardin, J.P. 2001. Adult and nymphs of Microtriatoma trinidadensis (Lent, 1951) (Hemiptera: Reduviidae) caught from peridomestic environment in Bolivia. Memórias do Instituto Oswaldo Cruz 96: 889–894. Crossref
Dos Santos Souza, É., Monte, G.L.S., Paiva, V.F., and Galvão, C. 2019. Microtriatoma trinidadensis (Lent, 1951) (Hemiptera, Reduviidae, Triatominae): first record in the state of Amazonas, Brazil. Check List 15: 905–909. Crossref
Galvão, C., Gil-Santana, H.R., and Oliveira, J.D. 2024. The taxonomic catalog of the Brazilian fauna: biodiversity and geographical distribution of Triatominae (Hemiptera: Reduviidae) in Brazil. Zoologia (Curitiba) 41 (2024): 1–28. Crossref
Germar, E.F. and Berendt, G.C. 1856. Die im bernstein befindlichen Hemipteren und Orthopteren der Vorwelt. 144 pp. Nicolaischen Buchhandlung, Berlin.
Grimaldi, D.A. 1995. The age of Dominican amber. In: K.B. Anderson and J.C. Crelling (eds.), Amber, Resinite, and Fossil Resins, 203–217. American Chemical Society, Washington, DC. Crossref
Gil-Santana, H.R., Martins, D.D.S., Silva, J.B.D., and Oliveira, J.D. 2021. First report of Microtriatoma borbai Lent & Wygodzinsky, 1979 (Hemiptera, Reduviidae, Triatominae) in the state of Espírito Santo, Brazil: would M. borbai be living in eucalyptus crops? Revista da Sociedade Brasileira de Medicina Tropical 54: e0147-2021. Crossref
Heer, O. 1870. Die Miocene flora und fauna Spitzbergens, Vol. 2. 140 pp. P. A. Norstedt & Söner, Stockholm.
Heiss, E. 2009. Über Holoptilinae in Bernstein (Heteroptera, Reduviidae). Denisia 26: 83–92.
Haridass, E.T. and Ananthakrishnan, T.N. 1980. Functional morphology of the fossula spongiosa in some reduviids (Insecta–Heteroptera–Reduviidae). Proceedings: Animal Sciences 89: 457–466. Crossref
Hernández-Damián, A.L., Rubalcava-Knoth, M.A., and Cevallos-Ferriz, S.R. 2024. A new extinct member of the resin producer group of the Mexican amber: Hymenaeaphyllum mirandae n. gen. n. sp. (Detarioideae–Leguminosae). Palaeoworld 33: 1710–1726. Crossref
Hylton, A., Fitzpatrick, D.M., Suepaul, R., Dobson, A.P., Charles, R.A., and Peterson, J.K. 2020. Preliminary characterization of triatomine bug blood meals on the island of Trinidad reveals opportunistic feeding behavior on both human and animal hosts. Tropical Medicine and Infectious Disease 5 (4): 166–180. Crossref
Iturralde-Vinent, M.A. and Macphee, R.D.E. 1996. Age and paleogeographic origin of Dominican amber. Science 273: 1850–1852. Crossref
Iturralde-Vinent, M.A. and Macphee, R.D.E. 2019. Remarks on the age of Dominican amber. Palaeoentomology 2 (3): 236–240. Crossref
Jung, S., Kim, J., Balvín, O., and Yamada, K. 2023. Molecular phylogeny of Cimicoidea (Heteroptera: Cimicomorpha) revisited: increased taxon sampling reveals evolution of traumatic insemination and paragenitalia. Insects 14 (3): 267–281. Crossref
Kinzelbach, R.K. 1970. Wanzen aus dem eozänen Ölschiefer von Messel (Insecta: Heteroptera). Notizblatt des Hessischen Landesamtes für Bodenforschung 98 (4): 9–18.
Kirkaldy, G.W. 1910. Three new Hemiptera–Heteroptera from the Miocene of Colorado. Entomological News 21 (3): 129–131.
Kieran, T.J., Gordon, E.R., Zaldívar‐Riverón, A., Ibarra‐Cerdeña, C.N., Glenn, T.C., and Weirauch, C. 2021. Ultraconserved elements reconstruct the evolution of Chagas disease‐vectoring kissing bugs (Reduviidae: Triatominae). Systematic Entomology 46: 725–740. Crossref
Lent, H. 1943. A new transmitter of Chagas’ disease in the city of Rio de Janeiro, Parabelminus carioca, gen. et sp. n. Memorias do Instituto Oswaldo Cruz 38: 497–516. Crossref
Lent, H. and Wygodzinsky, P. 1979. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas’ disease. Bulletin of the American Museum of Natural History 163 (3): 123–520.
Lent, H. and Jurberg, J. 1984. A genitália externa na tribo Bolboderini (Hemiptera, Reduviidae, Triatominae). Memórias do Instituto Oswaldo Cruz 79: 1–27. Crossref
Maldonado-Capriles, J. 1990. Systematic Catalogue of the Reduviidae of the World (Insecta: Heteroptera). 694 pp. Caribbean Journal of Science, special edition, University of Puerto Rico, Mayagüez.
Maldonado-Capriles, J., Santiago-Blay, J.A., and Poinar, G. Jr. 1993a. Apicrenus fossilis gen. & sp. n. (Heteroptera: Reduviidae: Apiomerinae) from Dominican amber (lower Oligocene–upper Eocene). Insect Systematics & Evolution 24 (2): 139–142. Crossref
Maldonado-Capriles, J., Santiago-Blay, J.A., and Poinar, G. Jr. 1993b. Paleoploiariola venosa, a new fossil Emesinae (Heteroptera: Reduviidae) genus and species from Dominican amber. The Journal of agriculture of the University of Puerto Rico 77 (1–2): 95–100. Crossref
Masonick, P.K., Knyshov, A., Gordon, E.R., Forero, D., Hwang, W.S., Hoey-Chamberlain, R., Bush, T., Castillo, S., Hernandez, M., Ramirez, J., Standring, S., Zhang, J.X., and Weirauch, C. 2024. A revised classification of the assassin bugs (Hemiptera: Heteroptera: Reduviidae) based on combined analysis of phylogenomic and morphological data. Systematic Entomology 50 (1): 102–138. Crossref
Penney, D., Green, D.I., and Mellish, C. 2010. Biodiversity of Fossils in Amber from the Major World Deposits. 304 pp. Siri Scientific Press, Manchester.
Poinar, G. Jr. 1991. Praecoris dominicana gen. n., sp. n. (Hemiptera: Reduviidae: Holoptilinae) from Dominican amber, with an interpretation of past behavior based on functional morphology. Insect Systematics & Evolution 22 (2): 193–199. Crossref
Poinar, G. Jr. 2005. Triatoma dominicana sp. n. (Hemiptera: Reduviidae: Triatominae), and Trypanosoma antiquus sp. n. (Stercoraria: Trypanosomatidae), the first fossil evidence of a triatomine-trypanosomatid vector association. Vector-Borne & Zoonotic Diseases 5 (1): 72–81. Crossref
Poinar, G. Jr. 2013. Panstrongylus hispaniolae sp. n. (Hemiptera: Reduviidae: Triatominae), a new fossil triatomine in Dominican amber, with evidence of gut flagellates. Palaeodiversity 6: 1–8.
Poinar, G. Jr. 2019. A primitive triatomine bug, Paleotriatoma metaxytaxa gen. et sp. nov. (Hemiptera: Reduviidae: Triatominae), in mid-Cretaceous amber from northern Myanmar. Cretaceous Research 93: 90–97. Crossref
Popov, Y.A. 1987a. A new species of the bug genus Empicoris Wolff from Dominican copal, with the redescription of E. nudus McAtee & Malloch (Heteroptera, Reduviidae, Emesinae). Stuttgarter Beiträge zur Naturkunde, Ser. B (Geologie und Paläontologie) 134: 1–9.
Popov, Y.A. 1987b. Synopsis of the Neotropical bug genus Malacopus Stål, with the description of a new fossil species from Dominican amber (Heteroptera: Reduviidae, Emesinae). Stuttgarter Beiträge zur Naturkunde, Ser. B (Geologie und Paläontologie) 130: 1–15.
Popov, Y.A. 1989. Alumeda n. g., a new bug genus elected for the fossil species from Dominican amber (Heteroptera: Reduviidae, Emesinae). Stuttgarter Beiträge zur Naturkunde, Ser. B (Geologie und Paläontologie) 150: 1–14.
Popov, Y.A. 1993. A new genus of Emesinae from Dominican amber (Heteroptera: Reduviidae). Acta Zoologica Cracoviensia 35: 435–443.
Popov, Y.A. 2003. Some fossil Reduviidae in Baltic amber (Heteroptera: Cimicomorpha: Reduviidae). Mitteilungen aus dem Museum für Geologie und Paläontologie der Universität Hamburg 87: 159–170.
Popov, Y.A. and Weitschat, W. 2005. Emesites voigti n. gen. n. sp.—the first Emesina (Insecta: Heteroptera, Reduviidae) from the Borneo amber. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 89: 173–178.
Popov, Y.A. and Chłond, D. 2015. The first fossil record of the Emesinae genus Emesopsis Uhler (Hemiptera: Heteroptera, Reduviidae) from Eocene Baltic amber. Zootaxa 4039: 566–574. Crossref
Putshkov, P.V. and Popov, Y.A. 1993. A remarkable nymph of a centrocneminous bug from the Baltic amber (Insecta: Heteroptera, Reduviidae). Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 75: 211–229.
Putshkov, P.V. and Popov, Y.A. 1995. Collarhamphus mixtus n. gen. n. sp. – Die erste Collartidina (Heteroptera: Reduviidae, Emesinae) aus dem Baltischen Bernstein. Mitteilungen aus dem Geologisch-Paläontologischen Inststitut der Universität Hamburg 78: 179–187.
Putshkov, P.V. and Popov, Y.A. 1998. Redubinotus liedtkei n. gen. n. sp.—the second Centrocnemina from the Baltic amber (Heteroptera: Reduviidae: Centrocnemidinae). Annals of the Upper Silesian Museum (Entomology) 8 (9): 205–210.
Ramirez, J., Bugaj-Nawrocka, A., Taszakowski, A., and Weirauch, C. 2024. New species and reclassification of the fossil assassin bug Koenigsbergia (Hemiptera: Reduviidae: Phimophorinae). Arthropod Systematics & Phylogeny 82: 369–384. Crossref
Reinhardt, K., Voigt, D., and Gorb, S.N. 2019. Evidence for a sexually selected function of the attachment system in bedbugs Cimex lectularius (Heteroptera, Cimicidae). Journal of Experimental Biology 222 (12): jeb206136. Crossref
Ramírez Silva, J.A., Forero, D., Santos Murgas, A., Osorio, P., and Zachrisson, B. 2022. First report of Ricollapallidinends and Rocconota tuberculigera (Hemiptera: Reduviidae: Harpactorinae) predatory Oebalus insularis (Hemiptera: Pentatomidae) in natural reserves of Darien, Panama. Idesia (Arica) 40 (1): 123–128. Crossref
Riou, B. 1999. Descriptions de quelques insectes fossiles du Miocène supérieur de la Montagne d’Andance (Ardèche, France). École Pratique des Hautes Études, Biologie et Evolution des Insectes 11 (12): 123–133.
Schlee, D. 1990. Das Bernstein–Kabinett. Begleitheft zur Bernsteinausstellung im Museum am Löwentor. Stuttgarter Beiträge zur Naturkunde Serie C 28: 1–100.
Sherlock, Í.A. and Guitton, N. 1982. Fauna Triatominae do Estado de Bahia, Brasil. V-Microtriatoma pratai sp. n. Hemiptera, Reduviidae, Triatominae, Bolboderini. Memórias do Instituto Oswaldo Cruz 77: 131–137. Crossref
Schuh, R.T., Weirauch, C., and Wheeler, W.C. 2009. Phylogenetic relationships within the Cimicomorpha (Hemiptera: Heteroptera): a total‐evidence analysis. Systematic Entomology 34 (1): 15–48. Crossref
Schuh, R.T. and Weirauch, C. 2020. True Bugs of the World (Hemiptera: Heteroptera). Classification and Natural History. Second Edition. 768 pp. Siri Scientific Press, Manchester.
Statz, G. and Wagner, E. 1950. Geocorisae (Landwanzen) aus den oberoligocänen Ablagerungen von Rott (Mitteilung aus dem Geologischen Institut der Universität Köln). Palaeontographica 98: 97–136.
Stach, P., Natkaniec-Nowak, L., Wagner, M., Dumańska-Słowik, M., Mroczkowska-Szerszeń, M., Wesełucha-Birczyńska, A., Drzewicz, P., George, C., and Garcia, E. 2020. A study on the formation environment of the La Cumbre Amber Deposit, from Santiago Province, the Northwestern Part of the Dominican Republic. Minerals 10 (9): 1–21. Crossref
Stach, P., Natkaniec-Nowak, L., Dumańska-Słowik, M., Kosakowski, P., Naglik, B., Drzewicz, P., Misiak, J., Pršek, J., George, C., and Ramírez Gómez, R.E. 2021. The study of Dominican amber-bearing sediments from Siete Cañadas and La Cumbre with a discussion on their origin. Scientific Reports 11 (1): 1–21. Crossref
Swanson, D.R., Heads, S.W., Taylor, S.J., and Wang, Y. 2021. A new remarkably preserved fossil assassin bug (Insecta, Heteroptera, Reduviidae) from the Eocene Green River Formation of Colorado. Papers in Palaeontology 7: 1459–1478. Crossref
Thomas, D.B. 1992. A fossil Empicoris Wolff (Reduviidae: Heteroptera) from Mexican amber with remarks on the phylogenetic status of the fossil genus Alumeda Popov. Journal of the New York Entomological Society 100: 535–539.
Wasmann, E. 1932. Eine ameisenmordende Gastwanze (Proptilocerus dolosus n. g. n. sp.) im baltischen Bernstein; beitrag zur Kenntnis der Myrmecophilen. Bernstein-Forschungen (Amber Studies) 3: 1–3.
Weirauch, C. 2007. Hairy attachment structures in Reduviidae (Cimicomorpha, Heteroptera), with observations on the fossula spongiosa in some other Cimicomorpha. Zoologischer Anzeiger 246 (3): 155–175. Crossref
Weirauch, C. 2008. Cladistic analysis of Reduviidae (Heteroptera: Cimicomorpha) based on morphological characters. Systematic Entomology 33: 229–274. Crossref
Weirauch, C., Bérenger, J.M., Berniker, L., Forero, D., Forthman, M., Frankenberg, S., Freedman, A., Gordon, E., Hoey-Chamberlain, R., Hwang, W.S., Marshall, S.A., Michael, A., Paiero, S.M., Udah, O., Watson, C., Yeo, M., Zhang, G., and Zhang, J. 2014. An illustrated identification key to assassin bug subfamilies and tribes (Hemiptera: Reduviidae). Canadian Journal of Arthropod Identification 26 (2): 1–115.
Zhang, G.Y. and Weirauch, C. 2011. Matching dimorphic sexes and immature stages with adults: resolving the systematics of the Bekilya group of Malagasy assassin bugs (Hemiptera: Reduviidae: Peiratinae). Systematic Entomology 36 (1): 115–138. Crossref
Zhang, G.Y. and Weirauch, C. 2013. Molecular phylogeny of Harpactorini (Insecta: Reduviidae): correlation of novel predation strategy with accelerated evolution of predatory leg morphology. Cladistics 30: 339–351. Crossref
Zhang, J.F. and Zhang, X.Y. 1990. Fossil insects of cicada (Homoptera) and true bugs (Heteroptera) from Shanwang, Shandong. Acta Palaeontologica Sinica 29 (3): 337–348.
Zhang J.X, Gordon, E.R., Forthman, M., Hwang, W.S., Walden, K., Swanson, D.R., Johnson, K.P., Meier, R., and Weirauch, C. 2016. Evolution of the assassin’s arms: insights from a phylogeny of combined transcriptomic and ribosomal DNA data (Heteroptera: Reduvioidea). Scientific Reports 6 (1): 1–8. Crossref
Zhang, J.F., Sun, B., and Zhang, X.Y. 1994. Miocene Insects and Spiders from Shanwang, Shandong [in Chinese]. 352 pp. Science Press, Beijing.
Zhang, P.P., Liu, Y.Q., Ren, D., and Yao, Y.Z. 2024. The first report of fossil of Centrocnemidinae (Hemiptera: Heteroptera: Reduviidae) in the Mesozoic. Palaeoentomology 7 (2): 314–323. Crossref
Zhang, P.P., Liu, Y.Q., Ren, D., and Yao, Y.Z. 2022. The oldest fossils from China provide the most direct evidence for the ancestral state of fossula spongiosa and stridulitrum of Reduviidae. Frontiers in Ecology and Evolution 10: 927537. Crossref
Acta Palaeontol. Pol. 70 (2): 357–368, 2025
https://doi.org/10.4202/app.01225.2024