
Post Cretaceous/Paleogene boundary recovery of the deep marine ecosystem recorded by benthic foraminiferal morphogroups
MICHAEL A. KAMINSKI, SYOUMA HIKMAHTIAR, and CLAUDIA G. CETEAN
Kaminski, M.A., Hikmahtiar, S., and Cetean, C.G. 2025. Post Cretaceous/Paleogene boundary recovery of the deep marine ecosystem recorded by benthic foraminiferal morphogroups. Acta Palaeontologica Polonica 70 (4): 641–648.
The uppermost 2 meters of the Maastrichtian Scaglia Rossa Fm. (Gubbio, Italy) record a notable change in the proportions of morphogroups of deep-water agglutinated foraminifera beginning about 1.4 m below the boundary, where we observe a gradual decrease in the abundance of the morphotype M4 (infauna) and a concurrent increase in the morphotype M1 (suspension feeders). This trend suggests a reduction in the flux of organic matter to the sea floor began approximately 114 k.y. before the boundary event. The abundance of M4 shows an abrupt decline from around 13% below the Cretaceous/Paleogene boundary, to a minimum of 7% in the beds directly above the boundary. This trend suggests a modest, but temporary reduction in organic matter flux to the sea floor following the Cretaceous/Paleogene boundary event, though not in the category of a “Strangelove Ocean”. Morphotype M1 shows a marked decrease across the boundary, with values decreasing from >70% to around 11%. The lower Paleocene displays strong fluctuations in the M4 morphogroup. In the same interval we observe a rise in Morphotype M2 (epifaunal detritivores) from values below 10% in the Maastrichtian to over 50% in the basal Paleocene, and a short-lived maximum in Morphotype M3, indicating a decline in particulate organic matter input from suspension and a concurrent increase in bacterial-derived organic matter in the early recovery phase of the Paleocene. The deep-marine ecosystem as witnessed by deep-water agglutinated foraminiferal morphogroups shows a prolonged recovery, with M1 abundance returning to Maastrichtian values approximately 2.8 meters above the Cretaceous/Paleogene boundary clay, corresponding to an age of 810 k.y. after the event. These findings support previous research from calcareous nannofossil studies, which suggested a prolonged recovery of the marine food web following the Cretaceous/Paleogene boundary event.
Key words: Foraminifera, ecology, Strangelove Ocean, Cretaceous, Paleogene, Apennines, Italy.
Michael A. Kaminski [kaminski@kfupm.edu.sa; ORCID: https://orcid.org/0000-0002-7344-5874 ], Geosciences Department, King Fahd University of Petroleum & Minerals, Dhahran 31261, Saudi Arabia.
Syouma Hikmahtiar [syouma.santoso@kfupm.edu.sa; ORCID: https://orcid.org/0000-0002-8481-1884 ], Geosciences Department, King Fahd University of Petroleum & Minerals, Dhahran 31261, Saudi Arabia.
Claudia G. Cetean [ceteanc@yahoo.com; ORCID: https://orcid.org/0009-0004-9882-0257 ], Micropress Europe, al. Mickiewicza 30, 30-059 Kraków, Poland.
Received 1 December 2024, accepted 28 August 2025, published online 30 October 2025.
Copyright © 2025 M.A. Kaminski et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Morphogroups of benthic foraminifera have been shown to be useful tools for paleoenvironmental analysis, as they vary in response to environmental gradients such as water depth (Corliss and Chen 1988), the dissolved oxygen content of bottom water (Kaiho 1991, 1994), and in particular to trophic conditions at the sea floor (Jorissen et al. 1995; Altenbach et al. 1999; Van der Zwaan et al. 1999). The agglutinated foraminifera can be categorized into four main morphogroups based on their test shape and feeding behavior (Jones and Charnock 1985). Morphotype M1 includes tubular suspension feeders that capture suspended food particles, while M2 and M3 represent coiled, multichambered forms that function as epifaunal detritivores. Morphotype M4 comprises elongated, rectilinear infaunal deposit feeders that feed as bacteriovores (Nagy et al. 1995; Van den Akker et al. 2000). The proportions of these morphogroups respond to changes in the flux of particulate organic matter (POM) to the seafloor, which in turn is influenced by water depth, currents, and surface-water phytoplankton production. In this study we use benthic foraminiferal morphogroup analysis to understand changes to the trophic structure across the Cretaceous/Paleogene boundary and in particular the dynamics and nature of organic matter flux to the seafloor.
We investigated the Gubbio area in the Italian Apennines, where a thick sequence of pelagic limestone of the Scaglia Rossa Formation exposes the Cretaceous/Paleogene (K/Pg) boundary (Luterbacher and Premoli Silva 1964; Alvarez et al. 1980). The Scaglia Rossa Formation in this region hosts diverse assemblages of deep-water agglutinated foraminifera (DWAF) (Kuhnt 1990; Kaminski and Gradstein 2005; Cetean 2009). Although some initial studies explored changes in morphogroup proportions across this boundary (e.g., Kuhnt and Kaminski 1996), these authors analyzed only a limited number of samples. Kaminski et al. (2011) later documented the Upper Cretaceous DWAF biostratigraphy from the Contessa Highway section near Gubbio but their data set terminates at the K/Pg boundary. By integrating the late Maastrichtian DWAF assemblages documented by Cetean (2009) with newer Paleocene findings by Hikmahtiar et al. (2022), we assembled a dataset that identifies 92 DWAF species in 73 samples collected across the K/Pg boundary interval (SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app70-Kaminski_etal_SOM.pdf), which includes some Paleocene taxa that have been recently described as new (Hikmahtiar and Kaminski 2022, 2023). This study aims to quantitatively track the shifts in community structure from the upper Maastrichtian through the lower Paleocene, using DWAF morphogroups to assess the impact of the K/Pg boundary event on the deep-sea ecosystem and to track recovery of the ecosystem following the K/Pg boundary event.
Other abbreviations.—DWAF, Deep-Water Agglutinated Foraminifera; K/Pg, Cretaceous/Paleogene; POM, Particulate Organic Matter.
Geological setting
The Umbria-Marche Basin in Italy hosts an extensive sequence of Cretaceous to Paleogene deep-water limestone formations, prominently exposed near Gubbio. This region is a crucial reference for the Cretaceous/Paleogene boundary, notably since Alvarez et al. (1980) first discovered the iridium anomaly in the Bottaccione Gorge section near Gubbio. The Contessa Highway Section (located at 43°22’47” N; 13°33’49” E) lies within this basin in central Italy a few km to the east of the Bottaccione Gorge section (Fig. 1). The Scaglia Rossa Formation, dating from the lower Turonian to the lower Eocene, is characterized by reddish-pink pelagic micritic limestones, with chert present at certain levels. This formation is part of a broader sequence of pelagic limestones, which range from the Cenomanian (Upper Cretaceous) to the upper Eocene and include interbedded marls within the lower Paleocene. Alvarez and Montanari (1988) provided a detailed description of the Scaglia Rossa Fm., noting its reddish pink micritic limestone composition and the presence of some cherty layers. They divided the formation into four members (R1–R4), ascending from the base to the top. An updated biostratigraphic and magnetostratigraphic framework of the Paleogene successions near Gubbio was presented by Coccioni et al. (2016), which included the outcrops sampled for this study.
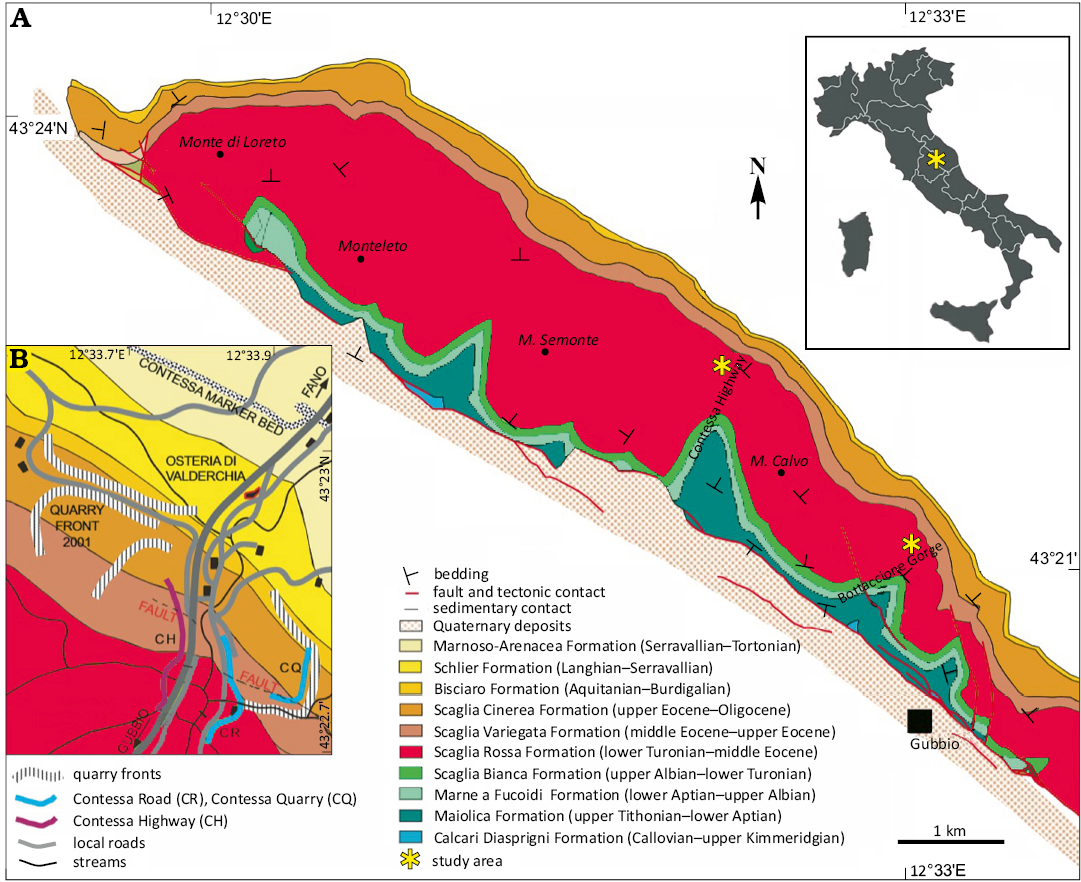
Fig. 1. Location of the Bottaccione Gorge section and the Contessa Highway section near Gubbio, Italy. Insert map shows the locations of exposed outcrops in the Contessa Quarry (CQ) and along the Contessa Highway (CH). Map redrawn after Jovane et al. (2007).
For the current study (43°21’56.2”N 12°34’57.0”E), we focused on sampling a 10 m composite section of the Maastrichtian R2 member and the lower portion of the Danian R3 member (extending from 4.4 m below the K/Pg boundary to 6.4 m above the boundary) of the Scaglia Rossa Fm. in the Contessa Highway section and Bottaccione Gorge section. The R2 member comprises Santonian to Maastrichtian pinkish limestone, transitioning to terminal whitish limestone. The overlying R3 member contains dark red marly limestones of the Paleocene. Both members are exposed along the east side of the Contessa Highway next to the small tunnel. This is the “Contessa Highway Lower Section” of Coccioni et al. (2016). A thicker interval of Maastrichtian limestone is exposed in the classic Bottaccione Gorge section, and this locality was newly sampled for this study. Our new data therefore constitute a composite record from the two sampled outcrops.
Material and methods
For this study, we analyzed 73 samples from the upper Maastrichtian and lower Paleocene of the Contessa Highway section and the Bottaccione Gorge section. We sampled the top 2 m of the Maastrichtian at 10 cm intervals at Bottaccione Gorge, with sample resolution of 20 cm below 2 m. In the Paleocene, we sampled at 10 cm intervals between 0 and 2 m and at 20 cm intervals from 2 to 6.4 m above the K/Pg boundary in the Contessa Highway section. The samples from the upper 2 m of the Maastrichtian were first cut into slabs horizontal to bedding with a diamond saw, and only the Cretaceous white limestone was sampled for micropaleontology, avoiding the pink burrow infillings that are present in the white limestone below the K/Pg boundary. Limestone samples of exactly 100 g weight were dissolved in dilute hydrochloric acid (HCl) and washed through a 63 µm sieve to extract the agglutinated foraminifera. We picked specimens from the >125 µm fraction and mounted them on cardboard microslides.
Our morphogroup analysis follows the classification models from Kaminski and Gradstein (2005) and Cetean et al. (2011), with minor modifications. The specimen slides are currently in the author’s collection but will be eventually archived in cabinet 14, drawers 22–24 at the European Micropalaeontological Reference Centre (Micropress Europe Foundation) at the AGH University of Kraków, Poland.
Results
Agglutinated foraminiferal morphogroups.—The population dynamics of benthic foraminifera are influenced by environmental factors, primarily mirroring the trophic gradient (Jorissen et al. 1995; Altenbach et al. 1999; Van der Zwaan et al. 1999). This study categorized the DWAF genera into morphogroups based on their overall morphology (Fig. 2). The classification of morphogroups used in this work follows the framework established by Cetean et al. (2011) for the Cretaceous DWAF genera and Kaminski and Gradstein (2005) and Setoyama et al. (2017) for the Paleocene forms, with some minor modifications. For example we now consider the pseudo-multichambered genera and Nothia to belong to Morphotype M2d (epifaunal detritivores).

Fig. 2. Morphotype model of Deep-Water Agglutinated Foraminifera from the Bottaccione and Contessa Highway sections.
To establish a robust chronostratigraphic framework for this study, we recalibrated the age vs. depth curve of Coccioni et al. (2016) to the new magnetostratigraphic age calibrations published in the 2020 Geological Time Scale (Gradstein et al. 2020). Our revised age model for the lower Paleocene of Contessa is shown in Fig. 3.
The newly obtained record of DWAF morphogroups across the K/Pg boundary is shown in Fig. 4. In the top 2 m of the Maastrichtian, there is a noticeable declining trend in the abundance of morphogroup M4, reaching minimum values 1.4 m below the K/Pg boundary. The low values persist in the beds directly above the boundary clay to a stratigraphic height of 0.5 m above the boundary clay, followed by more strongly fluctuating values in the lower Paleocene. Morphogroup M1 witnessed an increasing trend in abundance in the uppermost 1.4 m of the Maastrichtian and then an abrupt decline across the boundary, with minimum values in the first 0.5 m of the Paleocene. This coincides with a rapid increase in Morphogroups M2 and M3. At 0.3 m above the boundary the abundance of M2 exceeds any of the values observed in the uppermost 4.4 m of the Maastrichtian. The peak in M2 between 0.8 and 1.0 m is related to blooms of opportunistic species such as Spiroplectinella israelskyi (SOM 1). Morphogroup M1 returns to Maastrichtian levels around 3.0 to 3.4 meters above the boundary clay, suggesting a delayed recovery of the deep-sea ecosystem. According to our updated age model, recovery of the suspension-feeding morphogroup M1 to pre-K/Pg values occurred approximately 810 000 years after the K/Pg boundary event.
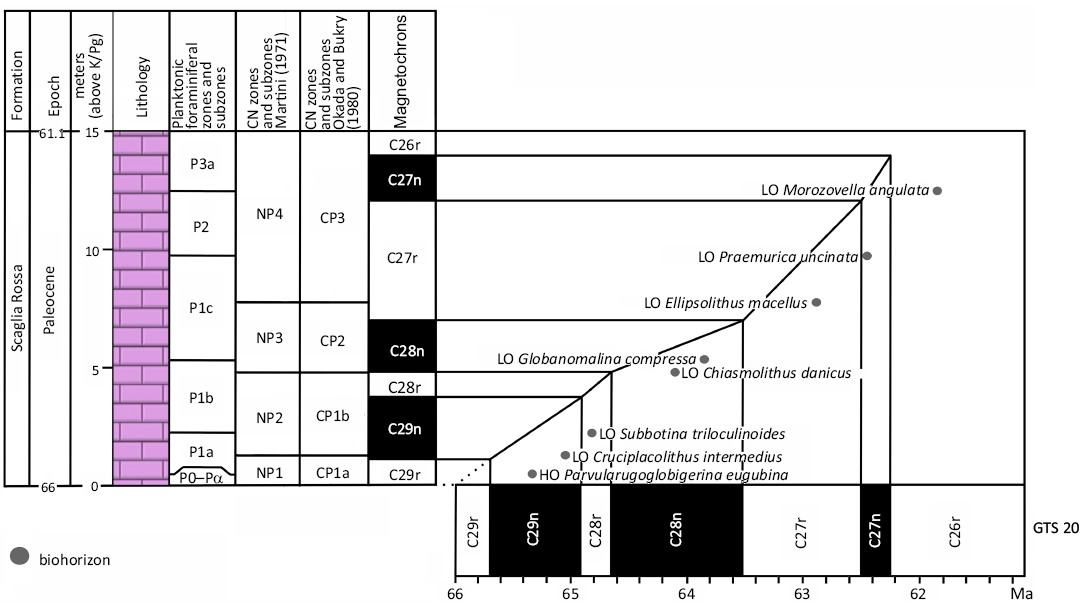
Fig. 3. Revised age depth plot for the Paleocene of the Contessa Highway section with correlation to the geomagnetic polarity time scale of Gradstein et al. 2020 (GTS ‘20), after Coccioni et al. (2016). Planktonic foraminiferal zones after Berggren et al. (1995).
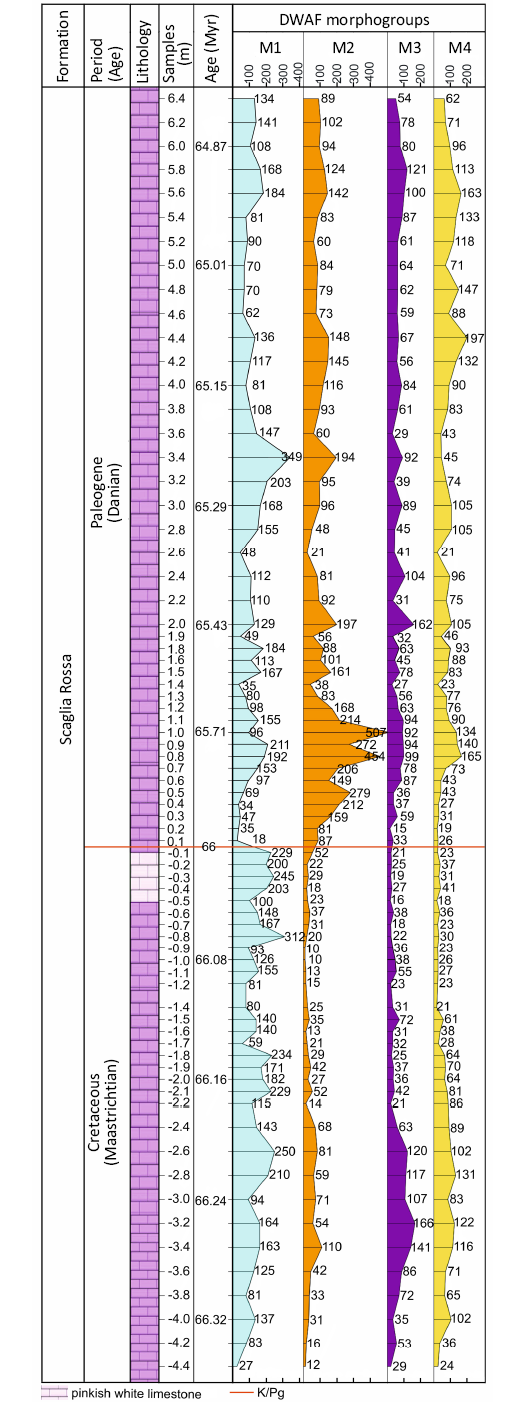
Fig. 4. Distribution of Deep-Water Agglutinated Foraminifera (DWAF) morphogroups across the K/Pg boundary at Gubbio.
Discussion
Kaminski and Gradstein (2005) presented a model in which agglutinated foraminifera display three levels of trophic activity in relation to the sea floor: tubular (the M1 morphotype) and globular (the M2a morphotype) agglutinated species first scavenge the organic matter reaching the sea floor, using their pseudopodia to capture food from the water column. Epifaunal and shallow infaunal taxa (M2b, M2c, M2d, M3a, M3b morphotypes) consume the organic matter that settles on the sediment surface, utilizing various active and passive feeding strategies. Bioturbators subsequently incorporate any remaining organic material into the sediment, making it accessible to deep infaunal species (M4a, M4b morphotypes).
At Bottaccione, we observed a decrease in the M4 morphogroup and a concurrent increase in M1 about 1.4 m below the K/Pg boundary, corresponding to about 114 kyr before the event. This trend is consistent with a reduction in the flux of organic matter to the sea floor that began approximately one Milankovitch cycle before the boundary event. The DWAF community before the boundary event was strongly dominated by suspension feeding tubular forms.
At Contessa, we observed a significant reduction in the M1 morphotype and a concurrent increase in M2 and M3 morphogroups across the K/Pg boundary. This shift signifies a transition from a Maastrichtian community dominated by epifaunal suspension feeders to one in the lower Paleocene dominated by epifaunal detritivores. The M4 morphogroup, primarily composed of elongated tapered forms, likely represents active deposit-feeders. The M4 trend shows a slight but steady decline in the upper two meters of the Maastrichtian, with continued low values in the lowermost 0.5 m of the Paleocene, followed by a gradual recovery to higher but less stable levels. The continued presence of the M4 morphogroup in the lowermost 0.5 m of the Paleocene and its rapid recovery in the overlying beds suggests that the overall flux of organic carbon to the sea floor did not completely collapse at the K/Pg boundary (i.e., “Strangelove Ocean”). Instead, the changes in the abundance of M1 and M2, M3 morphogroups imply a shift in the type of organic matter available to benthic organisms.
Previous studies of the carbon isotope record across the K/Pg boundary have suggested a collapse in productivity, leading to the “Strangelove Ocean” scenario (Hsü and MacKenzie 1985). Although the impact winter scenario may have caused a short-term decline in marine productivity, it does not fully account for the longer-term changes observed. Despite the extinction of calcareous nannoplankton at the boundary (MacLeod et al. 1997), organic-walled dinoflagellates survived and may have even increased in abundance in the earliest Paleocene (Hultberg 1986). Alegret et al. (2012) questioned the global nature of the “productivity collapse”, noting inconsistencies in paleoproductivity patterns across different regions. Based on our findings at Contessa, we estimate that the “Strangelove Ocean” was short-lived, lasting about one Milankovitch cycle. Birch et al. (2016) also found that while marine extinctions weakened the biological pump, the reduction in productivity was less severe and shorter in duration than previously suggested. Our results agree with these findings.
Although organic carbon flux may have fluctuated more intensely in the early Paleocene, as witnessed by the variable M4 morphogroup values, the reduction in productivity after the boundary event was minor compared to Late Cretaceous (M4 average values for the lowermost 0.5 m of the Paleocene = 11.5%, compared with the latest Maastrichtian average of 18.5%). The peak abundance of Repmanina charoides in samples CON0.9 and CON1.0 suggests slightly more oligotrophic conditions, as this species indicates oligotrophy in the modern Mediterranean Sea (De Rijk et al. 1999, 2000). Our findings are consistent with those of Gibbs et al. (2020), who observed reduced productivity from photosynthetic calcareous plankton and increased organic productivity driven by bacteria. The dominance of M2 and M3 morphogroups above the boundary supports their role as bacterial detritivores.
Conclusions
The DWAF assemblage structure based on morphogroups tracks the recovery of the deep-sea ecosystem across and following the K/Pg boundary event. The declining abundance of the M1 morphogroup (suspension feeders) reflects a reduction in the productivity of photosynthetic calcareous plankton at the boundary. At the same time, the increased abundance of the M2 and M3 morphogroups (epifaunal to shallow infaunal) suggest that productivity shifted towards bacterial sources. The small decline in the abundance of the M4 morphogroup (deep infaunal) in the basal Paleocene indicates a moderate but temporary reduction in the total sediment carbon flux across the K/Pg boundary. Our findings at Contessa do not suggest the existence of a “Strangelove Ocean” following the K/Pg boundary event. According to our revised age model, we observed reduced productivity and abundance of the deep-marine agglutinated foraminifera, with M1 returning to Maastrichtian levels approximately 2.8 meters above the K/Pg boundary, equivalent to about 810 kyr after the event. This estimate, although earlier than the changes in calcareous nannofossil assemblages reported by Gibbs et al. (2020), is consistent with the idea of a prolonged recovery of marine productivity after the K/Pg boundary event.
Acknowledgements
We thank the College of Petroleum and Geosciences at King Fahd University of Petroleum and Minerals (Dhahran, Saudi Arabia) for funding the fieldwork through a startup grant to MAK. We also thank Fabrizio Frontalini (Carlo Bo University, Urbino, Italy) for assisting with the fieldwork as well as Eiichi Setoyama (University of Utah, Salt Lake City, USA), Miroslav Bubík (Czech Geological Survey, Brno, Czech Republic) and Anna Waśkowska (AGH University of Kraków, Poland) for reviewing a draft of the paper.
Editor: Krzysztof Hryniewicz
References
Alegret, L., Thomas, E., and Lohmann, K.C. 2012. End-Cretaceous marine mass extinction not caused by productivity collapse. Proceedings of the National Academy of Sciences of the United States of America 109: 728–732. Crossref
Altenbach, A., Pflaumann, U., Schiebel, R., Thies, A., Timm, S., and Trauth, M. 1999. Scaling percentages and distributional patterns of benthic foraminifera with flux rates of organic carbon. Journal of Foraminiferal Research 29 (1): 173–185.
Alvarez, L.W., Alvarez, W., Asaro, F. and Michel, E.V. 1980. Extraterrestrial cause for the Cretaceous–Tertiary extinction. Science 208: 1095–1108. Crossref
Alvarez, W. and Montanari, A. 1988. Geologic framework of the Northern Apennines pelagic carbonate sequence. In: I. Premoli Silva, R. Coccioni, and A. Montanari (eds.), The Eocene–Oligocene Boundary in the Marche-Umbria Basin (Italy): Aniballi, Ancona, International Subcommission on Paleogene Stratigraphy Special Publication, 13–30. Anibelli Publishers, Ancona.
Berggren, W.A., Kent, D.V., Swisher III, C.C., and Aubry, M.P. 1995. A revised Cenozoic geochronology and chronostratigraphy. In: W.A. Berggren, D.V. Kent, and J. Hardenbol (eds.), Geochronology, Time Scales and Global Stratigraphic Correlations: A unified Temporal Framework for a Historical Geology. SEPM Special Volume 54: 129–212. Crossref
Birch, H.S., Coxall, H.K., Pearson, P.N., Kroon, D., and Schmidt D.N. 2016. Partial collapse of the marine carbon pump after the Cretaceous–Paleogene boundary. Geology 44: 287–290. Crossref
Cetean C.G. 2009. Cretaceous Foraminifera from the southern part of the Eastern Carpathians, between Stoeneşti and Cetăţeni, Paleocology and Biostratigraphy. 214 pp. Unpublished Ph.D. Thesis, Babeş-Bolyai University, Cluj-Napoca.
Cetean, C.G., Bӑlc, R., Kaminski, M.A., and Filipescu, S. 2011. Integrated biostratigraphy and palaeoenvironments of an upper Santonian–upper Campanian succession from the southern part of the Eastern Carpathians, Romania. Cretaceous Research 32: 575–590. Crossref
Coccioni, R., Catanzariti, R., Frontalini, F., Galbrun, B., Jovane, L., Montanari, A., Savian, J., and Sideri, M. 2016. Integrated magnetostratigraphy, biostratigraphy, and chronostratigraphy of the Paleogene pelagic succession at Gubbio (central Italy). In: M. Menichetti, R. Coccioni, and S. Montanari (eds.), The Stratigraphic Record of Gubbio: Integrated Stratigraphy of the Late Cretaceous Paleogene Umbria-Marche Pelagic Basin. Geological Society of America Special Paper 524: 139–160. Crossref
Corliss, B.H. and Chen, C. 1988. Morphotype patterns of Norwegian Sea deep-sea benthic foraminifera and ecological implications. Geology 16: 716–719. Crossref
De Rijk, S., Jorissen, F.J., Rohling, E.J., and Troelstra, S.R. 2000. Organic flux control on bathymetric zonation of Mediterranean benthic foraminifera. Marine Micropaleontology 40: 151–166. Crossref
De Rijk, S., Troelstra, S.R., and Rohling, E.J. 1999. Benthic foraminiferal distribution in the Mediterranean Sea. Journal of Foraminiferal Research 29 (2): 93–103. Crossref
Gibbs, S.J., Bown, P.R., Ward, B.A., Alvarez, S.A., Kim, H., Archontikis, O.A., Sauterey, B., Poulton, A.J., Wilson, J., and Ridgwell, A. 2020. Algal plankton turn to hunting to survive and recover from end-Cretaceous impact darkness. Science Advances 6: eabc9123. Crossref
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M. 2020. Geologic Time Scale 2020. Vols 1 and 2. 1357 pp. Elsevier, Amsterdam.
Hikmahtiar, S. and Kaminski, M.A. 2022. A new agglutinated foraminiferal species (Arenoturrispirillina waskowskae n. sp.) from the Danian of Contessa, Italy. Geology, Geophysics and Environment 48 (4): 405–411. Crossref
Hikmahtiar, S. and Kaminski, M.A. 2023. Ammopemphix contessaensis sp. nov., a pseudocolonial agglutinated foraminifera from lower Paleocene of the Western Tethys, Contessa, Italy. Bolletino della Società Paleontologica Italiana 62 (3): 281–287.
Hikmahtiar, S., Kaminski, M.A., and Cetean, C.G. 2022. Lower Paleocene Deep-Water Agglutinated Foraminifera from the Contessa Highway Section (Umbria-Marche Basin, Italy): Taxonomy, stratigraphic distribution and assemblage turnover across the Cretaceous/Paleogene Boundary. Rivista Italiana de Paleontologia e Stratigrafia 128: 719–744. Crossref
Hsü, K. and MacKenzie, J. 1985. A Strangelove Ocean in the Early Tertiary. In: E.T. Sundquist and W.S. Broecker (eds.), The Carbon Cycle and Atmospheric CO2: Archean to Present. Geophysical Monograph 32: 487–492. Crossref
Hultberg, S.U. 1986. Danian dinoflagellate zonation, the C-T boundary and the stratigraphical position of the fish clay in southern Scandinavia. Journal of Micropalaeontology 5: 37–47. Crossref
Jones, R.W. and Charnock, M.A. 1985. “Morphogroups” of agglutinating foraminifera. Their life position and feeding habits and potential applicability in (paleo) ecological studies. Revue de Paleobiologie 4: 311–320.
Jorissen, F.J., de Stigter, H.C., and Widmark, J.G.V. 1995. A conceptual model explaining benthic foraminiferal microhabitats. Marine Micropaleontology 26: 3–15. Crossref
Jovane, L., Florindo, F., Coccioni, R., Dinarès-Turell, J., Marsili, A., Monechi, S., Roberts, A.P., and Sprovieri, M. 2007. The Middle Eocene Climatic Optimum (MECO) event in the Contessa Highway Section, Umbrian Apennines, Italy. Geological Society of America Bulletin 119: 413–427. Crossref
Kaiho, K. 1991. Global changes of Paleogene aerobic/anaerobic benthic foraminifera and deep-sea circulation. Palaeogeography Palaeoclimatology Palaeoecology 83: 65–85. Crossref
Kaiho, K. 1994. Benthic foraminiferal dissolved oxygen index and dissolved oxygen levels in the modern ocean. Geology 22: 719–722. Crossref
Kaminski, M.A. and Gradstein, F.M. 2005. Atlas of Paleogene Cosmopolitan deep-water Agglutinated Foraminifera. Grzybowski Foundation Special Publication 10: 1–547.
Kaminski, M.A., Cetean, C.G., Bălc, R., and Coccioni, R. 2011. Upper Cretaceous Deep-Water Agglutinated Foraminifera from the Contessa Highway Section, Umbria-Marche Basin, Italy: Taxonomy and Biostratigraphy. In: M.A. Kaminski and S. Filipescu (eds.), Proceedings of the Eighth International Workshop on Agglutinated foraminifera. Grzybowski Foundation Special Publication 16: 71–106.
Kuhnt, W. 1990. Agglutinated foraminifera of western Mediterranean Upper Cretaceous pelagic limestones (Umbrian Apennines, Italy, and Betic Cordillera, Southern Spain). Micropaleontology 36: 297–330. Crossref
Kuhnt, W. and Kaminski, M.A. 1996. The response of benthic foraminifera to the K/T boundary Event: a review. In: S. Jardine, I. De Klasz, and J.P. Debenay (eds.), Geologie de l’Afrique et de l’Atlantique Sud – Compte-rendu des colloques de geologie d’Angiers. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, Mémoire 16: 433–442.
Luterbacher, H.P. and Premoli-Silva, I. 1964. Biostratigrafia del limite Cretaceo-Terziaro nell’Appennino centrale. Rivista Italiana di Paleontologia e Stratigrafia 70: 67–128.
MacLeod, N., Rawson, P.F., Forey, P.L., Banner, F.T., Boudagher-Fadel, M.K., Bown, P.R., Burnett, J.A., Chambers, P., Culver, S., Evans, S.E., Jeffrey, C., Kaminski, M.A., Lord, A.R., Milner, A.C., Milner, A.R., Morris, N., Owen, E., Rosen, B.R., Smith, A.B., Taylor, P.D., Urquhart, E., and Young, J.R. 1997. The Cretaceous–Tertiary biotic transition. Journal of the Geological Society, London 154: 265–292. Crossref
Martini, E. 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation. In: A. Farinacci (ed.), Proceedings 2nd International Conference Planktonic Microfossils Roma 2: 739–785. Tecnosci, Rome.
Nagy, J., Gradstein, F.M., Kaminski, M.A., and Holbourn, A.E. 1995. Foraminiferal morphogroups, paleoenvironments and new taxa from Jurassic to Cretaceous strata of Thakkhola, Nepal. In: M.A. Kaminski, S. Geroch, and M.A. Gasinski (eds.), Proceedings of the Fourth International Workshop on Agglutinated Foraminifera. Grzybowski Foundation Special Publication 3: 181–209.
Okada, H. and Bukry, D. 1980. Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation (Bukry, 1973; 1975). Marine Micropaleontology 5: 321–325. Crossref
Setoyama, E., Kaminski, M.A., and Tyszka, J. 2017. Late Cretaceous–Paleogene foraminiferal morphogroups as palaeoenvironmental tracers of the rifted Labrador margin, northern proto-Atlantic. In: M.A. Kaminski and L. Alegret (eds.), Proceedings of the Ninth International Workshop on Agglutinated Foraminifera. Grzybowski Foundation Special Publication 22: 171–213.
Van den Akker, T.J.H.A., Kaminski, M.A., Gradstein, F.M., and Wood, J. 2000. Campanian to Palaeocene biostratigraphy and palaeoenvironments in the Foula Sub-basin, west of the Shetland Islands, UK. Journal of Micropalaeontology 19: 23–43. Crossref
Van der Zwaan, G.J., Duijnstee, I.A.P., den Dulk, M., Ernst, S.R., Jannink, N.T., and Kouwenhoven, T.J. 1999. Benthic foraminifers: proxies or problems? a review of paleocological concepts. Earth Science Reviews 46: 213–236. Crossref
Acta Palaeontol. Pol. 70 (4): 641–648, 2025
https://doi.org/10.4202/app.01227.2024