
Revision of haploceratid ammonoids from the Štramberk Limestone, Jurassic–Cretaceous boundary beds (Outer Western Carpathians)
ZDENĚK VAŠÍČEK and PETR SKUPIEN
Vašíček, Z. and Skupien, P. 2025. Revision of haploceratid ammonoids from the Štramberk Limestone, Jurassic–Cretaceous boundary beds (Outer Western Carpathians). Acta Palaeontologica Polonica 70 (3): 421–441.
Haploceratids from the Štramberk Limestones represent three genera Haploceras, Hypolissoceras, and Volanites. The most species reach genus is Haploceras. The semi-involute shells of the local haploceratids are almost smooth or only with a specific sculpture bound to the ventral region near the peristome. The whorls tend to be weakly arched or flat. Statistical elaboration of H/D, U/D and B/D values during shell growth shows no significant differences between these values, except perhaps for U/D. The external morphology plays a decisive role in the generic and species identification of haploceratids. It is known that haploceratids form dimorphic pairs, as evidenced by the differently shaped peristomes in addition to the different shell sizes. Dimorphic pairs have been demonstrated as new in the Štramberk material for the pairs Haploceras staszycii (microconch, m) and Haploceras elimatum (macroconch, M), as well as Hypolissoceras carachtheis (m) and Hypolissoceras woehleri (M). Haploceras tithonium and Volanites verrucosus possessed dimorphic pairs but their counterparts have not been found in the Štramberk Limestone. With the exception of Volanites verruciferus, the species described here are of no stratigraphical importance. Their stratigraphic range is from the lower Tithonian to the lower Berriasian.
Key words: Ammonoidea, Haploceratidae, taxonomy, Štramberk Limestone, Silesian Unit, Štramberk area.
Zdeněk Vašíček [zdenek.vasicek@ugn.cas.cz; ORCID: https://orcid.org/0009-0001-5472-9468 ], Institute of Geonics of Academy of Sciences of the Czech Republic, Studentská 1768, CZ -708 00 Ostrava-Poruba, Czech Republic.
Petr Skupien [petr.skupien@vsb.cz; ORCID: https://orcid.org/0000-0001-9158-466X ] (corresponding author), Department of Geological Engineering, VŠB – Technical University of Ostrava, 17. listopadu 15, CZ-708 00 Ostrava-Poruba, Czech Republic.
Received 6 December 2024, accepted 20 April 2025, published online 22 August 2025.
Copyright © 2025 Z. Vašíček and P. Skupien. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Palaeontological collections of Moravian-Silesian museums house a considerable number of macrofossils from the Štramberk Limestone collected by generations of collectors and available for studies. Significant part of the macrofossils from Štramberk are ammonites. Representatives of the family Haploceratidae constitute around 20% of all ammonites in these collections. The contributions by Oppel (1865) and Zittel (1868) concern primarily ammonites, including haploceratids from the Štramberk Limestone. These ammonites are taxonomically described in a monograph by Zittel (1868) and his ammonite collection, with only minor exceptions, was deposited in Munich, Germany (SNSB-BSPG).
In contrast to other groups of ammonites (see, e.g., Vašíček et al. 2018), haploceratids from Štramberk area were not investigated by any researchers, only with a minor exception of Václav Houša (in Špinar 1965). In recent years, especially during our so-called palaeontological salvage collecting in the Kotouč Quarry in Štramberk since 2002, haploceratids have been found again. Thanks to that abundant new material often found in well established stratigraphic levels we could investigate haploceratids in detail and check if they have any stratigraphical significance for distinguishing between Tithonian and Berriasian. The examined new material is represented by approximately one hundred favourably preserved, non-deformed specimens from our own collection and, from repositories of four Moravian-Silesian museums (Museum of Ostrava; Nový Jičín Regional Museum; Silesian Museum in Opava, and Moravian Museum in Brno). Additionally, we re-examined the specimens studied by Zittel (1868, 1870) and deposited in SNSB-BSPG.
From a large number of the ammonites deposited in Moravian-Silesian museums, we selected and loaned favourably preserved specimens of haploceratids with at least one measurable shell diameter and other standard size parameters. We processed all of the new material under study in the Silesian and Moravian repositories (B, MM, PL, and Z) using an electric vibration needle (mainly removing rock matrix from the umbilici). A considerable obstacle in studying the museum collections of the Štramberk Limestone fauna is, however, are poorly documented accurate points of collecting of the specimens. The original labels contain usually only general locality name Stramberg (German spelling) or Štramberk (Czech spelling). In total, the material under this study displays 7 species of haploceratids. We give special attention to the possible sexual dimorphism in these ammonoids. We also discuss posssible matches of shells and aptychi in haploceratids.
Institutional abbreviations.—B, Museum of Ostrava, Czech Republic; MM and Ge, Moravian Museum in Brno, Czech Republic; PL, Nový Jičín Regional Museum, Czech Republic; Z, Silesian Museum in Opava, Czech Republic; SNSB-BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany.
Other abbreviations.—A, adventive lobe; B, whorl breadth; B/D, whorl breadth to the diameter ratio; D, diameter; Dmax, maximal diameter; E, external lobe; H, whorl height; H/D, whorl height to the diameter ratio; m, microconchs; M, macroconchs; U, umbilicus width; Ul, U2, umbilical lobes; U/D umbilicus width to the diameter ratio.
Geological setting
The Štramberk Limestone is exposed in the form of carbonate megablocks (1 to 100 meter in size), breccias and conglomerates in several quarries (i.e., Kotouč, Municipal, Horní Skalka, and Castle Hill) in the immediate vicinity of the town of Štramberk (Figs. 1, 2). This unit represents deposits that formed during the latest Jurassic–earliest Cretaceous. Today, it is located in the Outer Western Carpathians. Block accumulations of the Štramberk Limestone form part of the continental-rise sediments of the Baška Facies in the Silesian Unit, which were deposited in the flysch trough of the Baška Subunit (for more details, see Picha et al. 2006).
The Štramberk Limestone represents sediments, formed on a carbonate platform during the Late Jurassic and earliest Cretaceous, along the northern Tethyan margin in the area of the Outer Western Carpathians. The Štramberk Limestone is whitish-grey in colour and was deposited in various settings across the carbonate platform and reef complex. The most common type of rock is biodetrital limestone. In some intervals sedimentary breccias occur with clasts up to some dozen of centimeters.
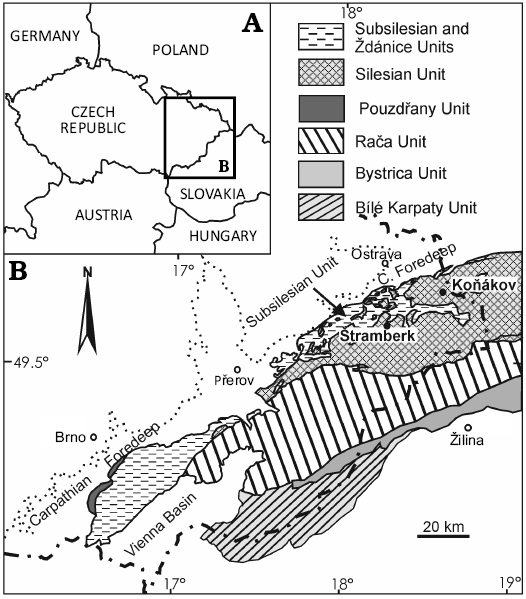
Fig. 1. A. Geographic location of the studied area. B. Tectonic map of the Outer Western Carpathian area of the Czech Republic (according to Skupien and Smaržová 2011).
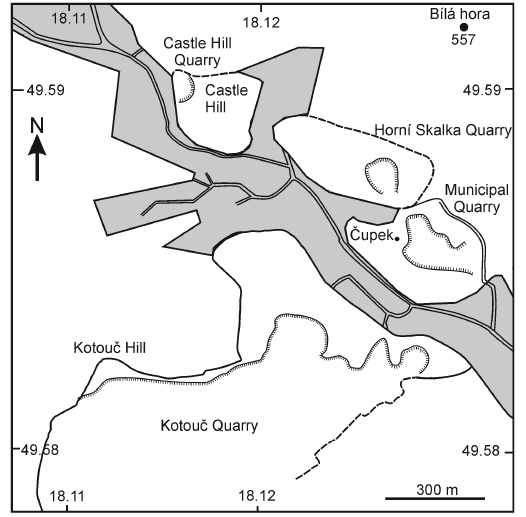
Fig. 2. Geographical setting of Štramberk Limestone quarries in the vicinity of Štramberk.
From about 1910 onwards, the quarry at Kotouč Hill has been the main source of macro- and microfossils that have been described in numerous palaeontological studies (see Vašíček and Skupien 2004, 2005 and references therein). The reefal limestone facies at the quarry varies widely, ranging from very coarse-grained to gravelly layers or lenses, formed by e.g., molluscan shells and corals, to very fine-grained micritic limestones and (most commonly) fine-grained biodetrital limestones (e.g., Houša and Vašíček 2004).
The age of the Štramberk Limestone was previously assumed to be the latest Kimmeridgian and Tithonian (e.g., Houša 1990; Houša in Houša and Vašíček 2004) but ammonites (Vašíček et al. 2013, 2017; Vašíček and Skupien 2013, 2014, 2016) from the limestone bodies indicate instead entire Tithonian and the earliest Berriasian.
Material
The advantage of ammonites from the Štramberk Limestone is their typically well-preserved, three-dimensional morphology without any deformation. In rare cases, their shells are only slightly deformed. These were not included in the statistical analysis. The core of our material comes from juveniles and ranges to adult specimens, with the diameter of the latter reaching almost 200 mm. There are three forms of proper preservation: recrystallized original tests, external and internal moulds (steinkerns). In the case of the last category, incomplete outer suture lines are usually preserved to various extents. Specimens are usually encased in detritic to coarse detritic, often organogenic limestone. In one specimen (Z1824) one side of the shell is covered by a sizeable number of small sessile remains of serpulid tubes, demonstrating that the ammonite shell had been lying on the bottom for quite a long time before it was covered with sediment.
The material processed by us is kept in museum collections and is designated in the systematic section below by means of an acronym of the relevant museum together with a deposit numbers marked on museum labels, as well as on individual specimens.
Historical background
The collection of ammnoids deposited now in SNSB-BSPG, was collected by a geologist Ludwig Hohenegger (1807–1854) mostly from a quarry below Castle Hill in Štramberk and from additional occurrences of Štramberk Limestone situated in the Silesian Unit of the Outer Western Carpathians, i.e., Koniakau (now Koňákov), Kotzobenz (now Chotěbuz), and Wilamowitz (now Wilamowice). Based on the Štramberk fauna, Oppel (1865) defined the Tithonian Stage. He identified and named some cephalopods from Štramberk Limestone, provided also brief descriptions but no illustrations. Subsequntly Zittel (1868) examined the collection of ammonites (and thus also haploceratids) from the Štramberk Limestone, described them in detail and illustrated—including the species established by Oppel in Zittel (1868) with no illustrations. Later on, Zittel (1870) created the genus Haploceras, which included Ammonites carachtheis and Ammonites staszycii established by Zejszner (1846). Zejszner (1846) illustrated these two species in his contribution on Tithonian brachiopods from Pieniny Klippen Belt, therefore he did not describe the ammonoids in detail. These species have been formally described in the monograph by Zittel (1870) and identified from the Štramberk Limestone. The name of the author of both species, however, have been provided in German spelling Zeuschner, while the original paper displays Polish spelling Zejszner.
In the 19th century and in the first half of the 20th century, the occurrence of Tithonian haploceratids under the generic name Haploceras was recorded in several localities throughout Europe. Breistroffer (1947) and subsequently many other authors proceeded to more specific, frequently different, varied and contradictory taxonomic concepts of haploceratids, above all in connection with Ammonites carachtheis at the subgeneric or generic level. Breistroffer (1947) defined the new subgenus Hypolissoceras with the type species A. carachtheis. Barthel (1962) assigned this latter species to Glochiceras Hyatt, 1900, a genus occurring in the Oxfordian and Kimmeridgian. Patrulius and Avram (1976) established the subgenus Neoglochiceras (synonym of Hypolissoceras Breistroffer, 1947); Olóriz Sáez (1978) used the subgenus Lingulaticeras Ziegler, 1958, for A. carachtheis; Sapunov (1979) used Glochiceras; Enay and Cecca (1986) added the new subgenus Volanites to the genus Haploceras; Főzy (1988) and lately Zeiss (2001) used subgenus Hypolissoceras for A. carachtheis. Kutek and Wierzbowski (1979, 1986) and Wierzbowski (1990) provided new data on the occurrence of several species of haploceratids, especially those from the lower and middle Tithonian strata of Rogoźnik (Pieniny Klippen Belt, Poland). Zeiss et al. (1994) referred to A. carachtheis as Hypoglochiceras carachtheis; however, the genus Hypoglochiceras has never been formally defined. Wright et al. (1996) included only two Cretaceous genera to the family Haploceratidae: Haploceras (and its synonyms) and Neolissoceras. Főzy and Scherzinger (2013) and Sarti (2020, 2024) reported also on some haploceratids from Trento Plateau (Southern Alp). All the species described in the papers by Sarti (2020, 2024) were classified in the genus Haploceras.
In remarks to the species Haploceras elimatum (Oppel Zittel, 1870), Barthel (1962) proposed that this species forms a dimorphic pair with Ha. staszycii. Olóriz Sáez (1978) described in detail Tithonian haploceratids from Spain. Enay and Cecca (1986) discussed in detail and in a somewhat non-traditional manner the evolution of Tithonian haploceratids. The basis of their analysis was Haploceras (Haploceras) carachtheis, which they considered as the evolutionary earliest species. First, they divided the collection of this species into two groups based on shell size and ornamentation. Furthermore, they subdivided Ha. carachtheis into smaller subgroups and named them morphs (e.g., Ha. carachtheis morph elimatum). In the framework of these morphs, they recognized microconchs (m) and macroconchs (M). For the species Haploceras verruciferum, Enay and Cecca (1986) established the new subgenus Volanites. Főzy (1988) subsequently studied the family Haploceratidae in detail, taking also into account possible dimorphism. He also defined a new species of Haploceras cassiferum. Interestingly, while establishing this new species, as well as discussing Ha. verruciferum, he did not mention the subgenus Volanites. At the beginning of this millennium, Zeiss (2001) again studied haploceratids. Among other matters, he reported that Ha. elimatum, which should represent microconchs, has allegedly developed peristome with rostra and lappets in all growth stages. Főzy and Scherzinger (2013) reported some similar observations.
Recently, Hoedemaeker (in Hoedemaeker et al. 2016) revised the systematics of Haploceras. His conception reflected the opinions of Enay and Cecca (1986); he also used morph as a taxonomic category, and regarded Haploceras carachtheis morph carachtheis as a microconch and Haploceras carachtheis morph elimatum as a macroconch.
Systematic palaeontology
In the superfamily- to family-level conception of haploceratids, we follow the historical classification by von Zittel (1884), with modifications proposed by Wright et al. (1996) with the type genus Haploceras. At the suborder level, we did not follow the older taxonomy, by e.g., Hyatt (1900) or Callomon (1981). According to the character of the suture lines of haploceratids by Beznosov and Mikhailova (1983, 1991) we have classified the haploceratids in the suborder Haploceratina.
In the 20th century, several other genera were established for the Late Jurassic period. Among them, in the presented paper, besides Haploceras, the Hypolissoceras and Volanites are considered as separate genera. Volanites was originally not very clearly established as a subgenus of the genus Haploceras in Enay and Cecca (1986).
For favourably preserved specimens of haploceratids, the usual size parameters were measured in mm. For a description and illustration of outer suture lines, we use newer terminology from Korn et al. (2003).
Stratigraphic data and ammonite zonation are based primarily on the data from Enay and Cecca (1986), Zeiss (2001, 2003), and Főzy and Scherzinger (2013).
Order Ammonitida von Zittel, 1884
Suborder Haploceratina Beznosov & Mikhailova, 1983
Superfamily Haploceratoidea von Zittel, 1884
Family Haploceratidae von Zittel, 1884
Genus Haploceras Zittel, 1870
Type species: Ammonites elimatus Oppel, 1865; Štramberk, Czech Republic, Tithonian, by subsequent designation of Spath 1923: 14.
Remarks.—Adult members of the genus possess smooth or indistinctly ribbed shells with ribs best developed on the venter and on the upper half of whorl flanks. Indistinct fine lines are sometimes apparent even near the umbilicus. Wright et al. (1996: 10) consider, in accordance with previous conventions, a specimen of Ammonites elimatus as the type species of the genus Haploceras. In the generic description, Wright et al. (1996) stated that Hypolissoceras Breistroffer, 1947, with type species Ammonites carachtheis Zejszner, 1846, is a synonym of the genus Haploceras. Wright et al. (1996) again illustrated the specimen of Zittel (1868: pl. 15: 1a–c) in fig. 6/1c–e and stated that it is Haploceras carachtheis, which should be considered a microconch to Haploceras elimatum. We do not agree with this concept, as discussed in detail below.
Recently, Hoedemaeker has discussed the systematics of the genus Haploceras (in Hoedemaeker et al. 2016) and returned back to the concept of morphs by Enay and Cecca (1986). He regards Haploceras carachtheis morph carachtheis as the microconch corresponding to the macroconch Ha. carachtheis morph elimatum. He does not discuss the entirely different type of ribbings in Ha. carachtheis and Ha. elimatum, respectively.
Haploceras elimatum (Oppel, 1865)
Figs. 3A, 4A, 5, 6F, G.
1865 Ammonites elimatus Opp.; Oppel 1865: 549.
1868 Ammonites elimatus Opp.; Zittel 1868: 79, pl. 13: 1a–c (lectotype), 2, 3a, b, 4, 5a–c, 6, 7a, b.
1870 Haploceras elimatum Opp. sp.; Zittel 1870: 51, pl. 27: 7a, b.
1890 Haploceras elimatum Oppel; Toucas 1890: 576, pl. 13: 4.
1923 Haploceras elimatum (Oppel); Spath 1923: 14.
1925 Haploceras elimatum (Oppel); Spath 1925: 153, pl. 1: 1a–c.
1953 Haploceras elimatum Oppel; Arnould-Saget 1953: 8, pl. 1: 9a–c.
?1953 Haploceras elimatum Opp.; Slavin 1953: 54, pl. 1: 9.
1956 Haploceras elimatum (Opp.); Książkiewicz 1956: 211, pl. 25: 6, text-fig. 25.
1960 Haploceras elimatum Opp.; Collignon 1960: pl. 142: 536, 537.
1960 Haploceras subelimatum Font.; Collignon 1960: pl. 142: 538.
1960 Haploceras elimatum Oppel; Drushchits and Kudryavtsev 1960: 267, pl. 13: 1, text-fig. 71.
1962 Haploceras elimatum (Oppel); Barthel 1962: 11, pl. 1: 12–15 (= Zittel 1870: pl. 27: 7a, b), t ext-figs. 15–17.
1965 Haploceras elimatum (Oppel); Houša in Špinar 1965: fig. VIII/254.
1966a Haploceras (Haploceras) elimatum; Wiedmann 1966a: 76, pl. 1: 1a, b.
1966b Haploceras (Haploceras) elimatum (Oppel); Wiedmann 1966 b: 61, text-figs. 42b, 43a.
1968 Haploceras (Haploceras) elimatum (Oppel); Wiedmann 1968: pl. 1: 3a, b.
1970 Haploceras elimatum (Oppel); Patrulius et al. 1970: 138, pl. 2: 11a, b, ?fig. 10.
1976 Haploceras (Haploceras) elimatum (Oppel); Avram 1976: 34, pl. 8: 1a, b.
1978 Haploceras elimatum (Oppel); Olóriz Sáez 1978: 12, pl. 1: 2, 3.
1979 Haploceras elimatum (Oppel); Sapunov 1979: 42, pl. 7: 1, 2a, b, text-fig. 4/5 (with synonymy).
1983 Haploceras (Haploceras) elimatum (Oppel); Vašíček 1983: 97, pl. 3: 1.
1984 Haploceras elimatum (Oppel); Rossi 1984: 88, pl. 31: 11, 12.
1986 (Haploceras (Haploceras) carachtheis (M) (Zeuschner) morph elimatum; Enay and Cecca 1986: pl. 4: 3, 4a, b, 5a, b; non pl. 4: 1a, b, 2a, b (= Haploceras staszyci Zejszner).
1988 Haploceras (Haploceras) elimatum (Oppel); Főzy 1988: 51, pl. 2: 1, non pl. 2: 2, 3 (= Haploceras tithonium Oppel).
?1994 Haploceras elimatum (Oppel); Főzy et al. 1994: 158, pl. 1: 10.
?1994 Haploceras elimatum (Oppel); Zeiss et al. 1994: 1: 4.
1995 Haploceras elimatum (Oppel); Főzy 1995: 136, pl. 20: 9.
1996 Haploceras elimatum (Oppel); Wright et al. 1996: 10: 6/1a, b (= Zittel 1868: pl. 13: 1a, b).
1997 Haploceras (Haploceras) carachtheis Zeuschner morph elimatum (Oppel); Benzaggagh and Atrops 1997: pl. 4: 3.
1999 Haploceras cf. elimatum (Oppel); Fatmi and Zeiss 1999: 39, pl. 43: 1.
non 1999 Haploceras elimatum (Oppel); Kvantaliani 1999: 74, pl. 5: 4 (= Haploceras staszyci Zejszner).
2000 Haploceras ex gr. elimatum (Oppel); Arkadiev et al. 2000: 99, pl. 2: 6a–c.
2001 Haploceras (Haploceras) elimatum (Oppel); Zeiss 2001: 37, pl. 5: 6, 7.
2005 Haploceras elimatum (Oppel); Boughdiri et al. 2005: 309, pl. 2: 1.
2009 Haploceras elimatum (Oppel); Klein et al. 2009: 257 (with synonymy).
2013 Haploceras elimatum (Oppel); Főzy and Scherzinger 2013: 214, pl. 4: 5a, b, ?pl. 5: 11, ? pl. 6: 7.
2013 Haploceras elimatum (Oppel); Szives and Fözy 2013: 294, pl. 3: 3.
2016 Haploceras carachtheis morph elimatum (Oppel); Hoedemaeker et al. 2016: 169, pl. 17: 7, 8.
2017 Haploceras elimatum (Oppel); Főzy 2017: pl. 27: 5, pl. 35: 7.
2020 Haploceras elimatum (Oppel); Sarti 2020: 86.
2024 Haploceras elimatum (Oppel); Sarti 2024: 86.
Type material: Lectotype SNSB-BSPG AS III 800 was designated by Spath (1925), originally figured by Zittel (1868: pl. 13: 1 a–c).
Type locality: Štramberk Limestone, Štramberk, Czech Republic.
Type horizon: Tithonian or lower Berriasian.
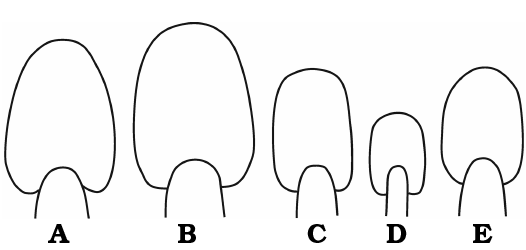
Fig. 3. Whorl sections of haploceratid ammonoids from Štramberk, Czech Republic, Tithonian–lower Berriasian. A. Haploceras elimatum (Oppel, 1865), SNSB-BSPG AS III 800 (Zittel 1868: pl. 13: 1a) at H = 33 mm. B. Haploceras staszycii (Zejszner, 1846), SNSB-BSPG AS III 805 (Zittel 1870: pl. 27: 3a) at H = 37 mm. C. Haploceras tithonium (Oppel, 1865), SNSB-BSPG AS III 798 (Zittel 1868: pl. 14: 2a) at H = 25 mm. D. Hypolissoceras carachtheis (Zejszner, 1846), SNSB-BSPG AS III 47 (Zittel 1868: pl. 15: 1c) at H = 17 mm. E. Hypolissoceras woehleri (Oppel, 1865), MM 06 at H = 27 mm.
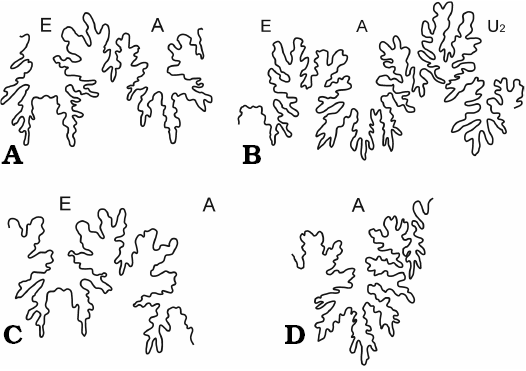
Fig. 4. Suture lines of haploceratid ammonoids from Štramberk, Czech Republic, Tithonian–lower Berriasian. A. Haploceras elimatum (Oppel, 1865). Almost complete outer suture line of Z4224 at H = 11.5 mm. B. Haploceras staszycii (Zejszner, 1846). Substantial part of outer suture line of PL5061 (6th level, middle) at H = 14 mm. C. Haploceras tithonium (Oppel, 1865). Outer and adventive lobes with accompanying saddles of PL5113 at H = 15 mm. D. Hypolissoceras woehleri (Oppel, 1865). Adventive lobe with parts of accompanying saddles of MM08 at H = 15 mm. A, adventive lobe; E, external lobe; U2, umbilical lobe.
Material.—In addition to the favourably preserved and published specimens deposited in Munich (SNSB-BSPG AS III 800–802), more than 30 other non-deformed specimens with sizes ranging from approximately 20 to 200 mm with various degrees of preservation quality were studied. Some specimens deposited in the Moravian-Silesian museums are phragmocones (some with preserved incomplete suture lines), and others are phragmocones with incomplete body chambers. The best preserved specimens are PL2520, PL5056, PL5058, PL5107–5112, B13569, B13831, Z1824, Z4224,. However, no completely preserved specimens have been found.
Description.—All specimens are semi-involute, with high, medium-broad whorls and a comparatively narrow umbilicus. The flanks of the whorl are arched. The whorl reaches its greatest breadth relatively close to the umbilical seam. The whorl declines from the area of the greatest arch to the umbilicus more steeply but continuously. No edge is developed on the line of coiling, but there is a narrow, strongly arched zone. Judging from the steinkerns, it is evident that the whorl still plunges inside below the line of coiling so that between the ultimate and penultimate whorls, an empty gap remains in the steinkerns. The flanks of the whorl decline gradually towards the venter. The venter is medium-broad and strongly arched.
The exceptionally well-preserved is medium-sized specimen PL5108 has a recrystallized original shell (Fig. 5D) and in comparison to others, is more conspicuously ribbed. Relatively broad, blunt, and rather widely placed ribs are present mainly on the upper half of the whorl in Haploceras elimatum. The ribs are S-shaped. Near the line of coiling, ribs are faint and concavely bent to the aperture in a short zone. At approximately half the whorl height, the blunt ribs are clearly convexly bent and on the outer side, again concave in a wide shallow arch. Over the venter, the ribs pass without interruption and are slightly inclined towards the peristome. On the original shells, small, fine, dense lines (Fig. 5C2) are usually preserved near the umbilicus. On the flanks and on the venter, as with the steinkerns, the ribs are usually only slightly pronounced. Very often, ribbing is not preserved at all, and thus, the specimens seem to be or are smooth.
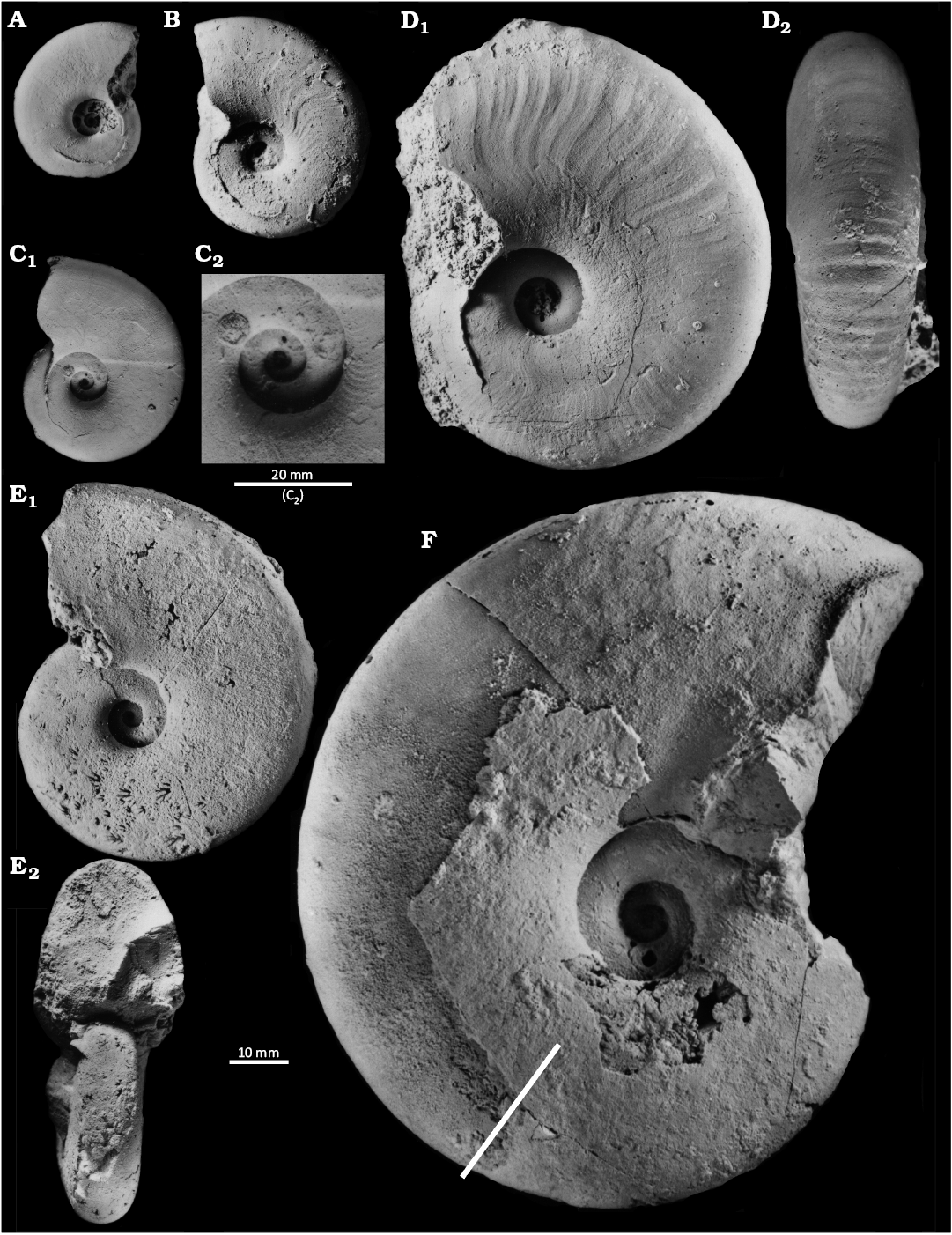
Fig. 5. Haploceratid ammonoid Haploceras elimatum (Oppel, 1865) from Kotouč Quarry, Tithonian–lower Berriasian, Upper Jurassic–Lower Cretaceous, from author’s own findings in the quarry or from the collection of Lubomir Martinásek (Kopřivnice, Czech Republic). A. PL5058, phragmocone with original test. B. PL5112, specimen with original test and with beginning of body chamber, lateral view. C. PL5110, phragmocone with original test, lateral view (C1), detail of sculpture around umbilicus (C2). D. PL5108, specimen with the original test and with a part of body chamber, lateral (D1) and ventral (D2) views. E. PL5107, external mould with beginning of body chamber in lateral view (E1), view on the area near aperture (E2). F. PL5109, internal mould in lateral view, with the body chamber inclusive of peristome (with rest of original test around umbilicus). The straight line perpendicular to periphery of specimen shows the margin between body chamber and phragmocone.
Zittel (1868: pl. 13: 2, 7) illustrated a peristome for two specimens. The smaller specimen, reaching a diameter of approximately 70 mm, has a simpler peristome than the larger specimen. The larger one, with a diameter of approximately 125 mm, displays a peristome with a well expressed lappet.
Almost entire outer suture line is preserved on the juvenile specimen Z4224 (D = 28.5 mm). The external lobe E bears a non-high secondary saddle of rectangular shape. The E/A saddle is two-branched, with its external branch higher than its internal branch. The mentioned saddle is lower than the A/U saddle. Adventive lobe A is broad and four finger-shaped at the base. On specimen B13831, an incomplete suture line (E and A) is preserved as well.
Measurements.—The parameters we measured on both juvenile and adult specimens (we measured approximately 30 specimens, see SOM 1, 2, in Supplementary Onine Material available at http://app.pan.pl/SOM/app70-Vasicek_Skupien_SOM.pdf). The lectotype at D = 61.3 mm (Dmax ca. 70 mm) has H/D = 0.49, U/D = 0.22, B/D = 0. 34. Across the entire size range of the shells, the ratios of the measured parameters remain in similar range: H/D ranges 0.46–0.50, U/D ca. 0.20–0.23, B/D ca. 0.33–0.38. It statistical investigations we initially set an arbitrary boundary between juvenile and adult specimens at a value around the diameter D of ca. 60 mm for practical reasons. In Fig. 7A–C, we have labelled the smaller specimens as “microconchs”, and the larger ones as “macroconchs”. The regression lines are similar in all cases, and the calculated correlation coefficients R are close to 1. Therefore it is implausible to define any statististically meaningful boundary between small and large specimens (see Fig. 7). We conclude therefore that all measured specimens belong to the macroconch category. The terminal size of the adult specimens is around the diameter of 105 mm. These results agree with data of Zeiss (2001).
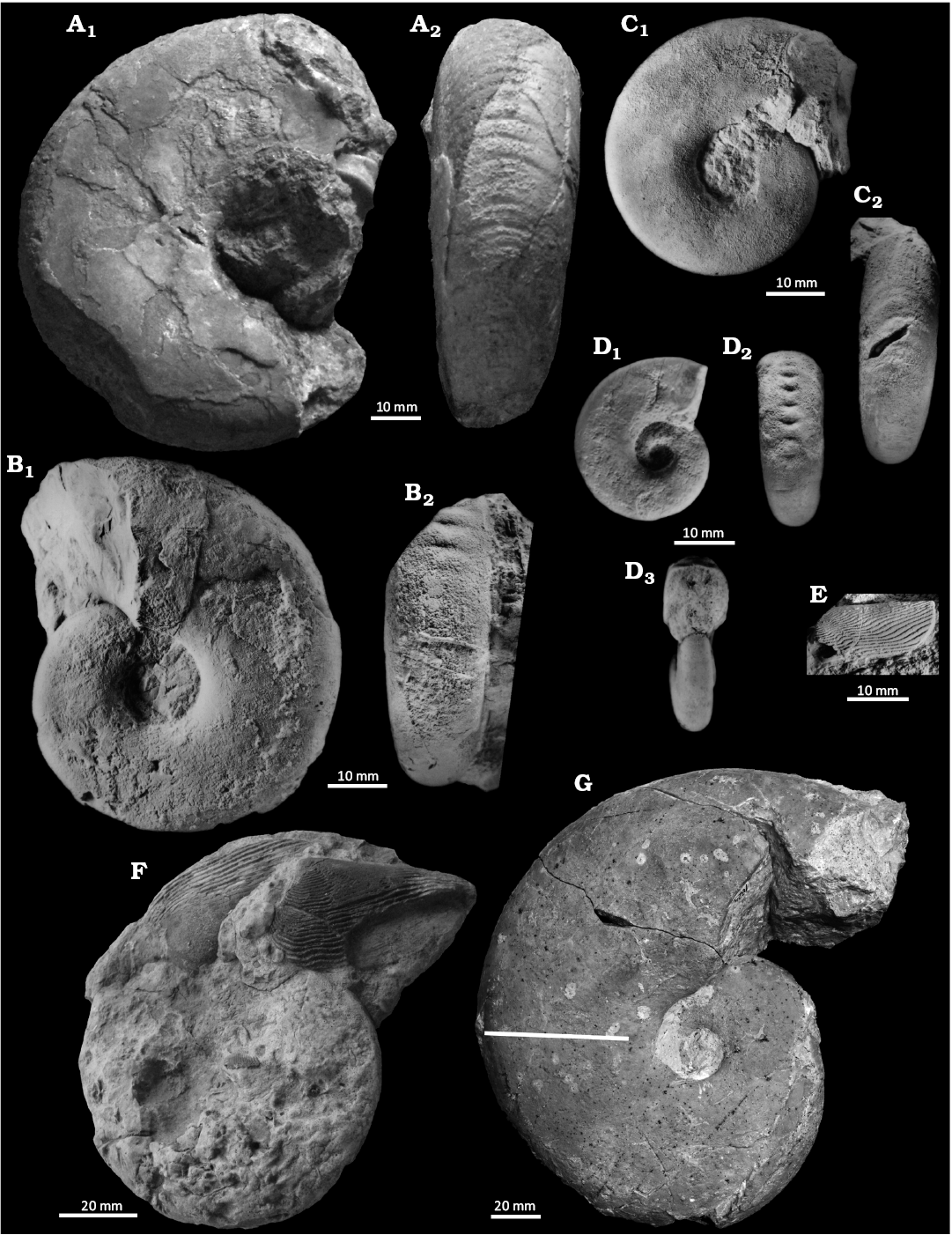
Fig. 6. Haploceratid ammonoids from Tithonian–lower Berriasian, Upper Jurassic–Lower Cretaceous. A, B. Hypolissoceras woehleri (Oppel, 1865). A. Holotype AS III 811 (real state of preservation), external mould, Štramberk, in lateral (A1) and ventral (A2) views. B. Z4171, external mould, Štramberk, in lateral (B1) and ventral (B2) views. C. Volanites verruciferus (Zittel, 1869), Z2981b, external mould, Štramberk, in lateral (C1) and ventral (C2) views. D. Hypolissoceras subtilius (Zittel, 1870), Z2767a, external mould, Štramberk, in lateral (D1) and ventral (D2) views, view of the area of peristome (D3). E. Beyrichilamellaptychus beyrichi beyrichi (Oppel, 1865), Ge0001, lateral view, from Měchová et al. (2010: fig. 9B), Kurovice. F, G. Haploceras elimatum (Oppel, 1865). F. Specimen with Punctaptychus punctatus (Zittel, 1868) in the body chamber, AS III 339, in lateral view, from Barthel (1962), according to Barthel’s hand-written label: “H. elimatum (Oppel) mit erhaltenen Aptychen, Tithon, Le Pouzin, Typloc. zu Toucas 1890”. G. Z1824, specimens carrying the calcitic shells of serpulid worms (white dots), Štramberk. The straight line perpendicular to periphery of specimen shows the margin between body chamber and phragmocone. Photos Kristýna Mezihoráková (Ostrava, Czechia), exept A, ZV; and G. Ondřej Malek, (Green Gas DPB, Paskov, Czechia).

Fig. 7. Relationship between size parameters of Haploceras elimatum (Oppel, 1865) macroconchs and microconchs. A. Height (H) of whorl on diameter (D). B. Umbilicus breadth (U) on diameter, C. Whorl breadth (B) on diameter.
Remarks.—We could not found any differences neither in the measured parameters nor their morphology and ornamentation between small and large specimens. The peristome in the small specimen illustrated as a drawing by Zittel (1868: pl. 13: 2) does not accurately reflect reality. Its shape, especially the size of the lappet is somewhat reduced. In contrast, in the peristome of the specimen illustrated by Zittel (1868: pl. 13: 7a), a seemingly unrealistic robust lappet is shown. Less conspicuous lappets in large specimens of Ha. elimatum are indicated by growth lines on the flank of the whorls of most specimens, as illustrated in Fig. 5F. This is also valid for the imperfectly preserved peristome of our adult specimen shown in Fig. 5F, which reaches a size similar to that of the specimen in Zittel (1868: pl. 13: 7). Growth lines, which should highlight the conspicuous lappet, are missing also in the lectotype.
Fontannes (1879) established Haploceras subelimatum, which according to its illustration (Fontannes 1879: pl. 2: 6) differs from Ha. elimatum merely in having narrower whorls. This is also confirmed by the data of B/D values (around 0.27–0.37) in the specimens reported by Sarti (2020). In contrast, Ha. subelimatum as illustrated by Collignon (1960: pl. 142: 538) did not have narrow whorls, and thus, it can be regarded as Ha. elimatum. According to a table showing the measurements of the specimens described by Kvantaliani (1999) as Ha. elimatum, the specimens have broader whorls and narrower umbilicus than both aforementioned species and apparently belong to Ha. staszycii.
Stratigraphic and geographic range.—Haploceras elimatum is known from the whole Mediterranean area from North Africa to the Himalayas and from Madagascar (for more information, see Főzy and Scherzinger 2013). According to figure in Enay and Cecca (1986: fig. 4), which marks the ammonite zonation of the Tithonian, Ha. elimatum has a considerably wide stratigraphic range from the basal Tithonian to the lower Berriasian inclusive (see also Sarti 2020). The lectotype comes from an unspecified part of the Štramberk Limestone in Štramberk. The lower Berriasian occurence is mentioned, e.g., by Arnould-Saget (1953), Olóriz Sáez (1978), Vašíček and Skupien (2013), and Szives and Főzy (2013). Spath (1925) stated that in Madagascar, this species occurs in the Kimmeridgian, but Collignon (1960) corrected it to the lower Tithonian (though also with an imprecise stratigraphy).
Our specimens from the Kotouč Quarry occur, according to the other accompanying ammonites, in both the lower and upper Tithonian (especially locality 4 in Vašíček and Skupien 2016), as well as sporadically in the lower Berriasian (Vašíček and Skupien 2013).
Another locality with Ha. elimatum in the Czech Republic is an abandoned quarry near the municipality of Roštín (Rača Nappe of the Magura Unit), namely, in a block conglomerate with grey marly limestones that, according to the accompanying ammonites, belong to the lower Tithonian (Vašíček 1983). Other historical occurrences in the Silesian Unit of the Outer Western Carpathians are known from so-called exotic conglomerates, in which the main component is Štramberk Limestone, in the localities of Koňákov, Chotěbuz, and other (Hohenegger 1861).
Haploceras staszycii (Zejszner, 1846)
Figs. 3B, 4B, 8A–D.
1846 Ammonites Staszycii Zejszner 1846: pl. 4: 3a–c.
1870 Haploceras Staszycii Zeuschner; Zittel 1870: 50, pl. 27: 2a, b, 3a, b (neotype), non figs. 4, 5 (?).
1876 Haploceras Staszycii Zeuschn.; Gemmellaro 1876: 34, pl. 7: 1–3.
1879 Haploceras staszycii Zeuschner; Fontannes 1879: 11, pl. 2: 4a, b.
1960 Haploceras staszycii Zeuschn.; Collignon 1960: pl. 142: 539.
1970 Haploceras stazsycii (Zeuschner); Patrulius et al. 1970: 138, pl. 2: 9a, b.
1976 Haploceras (Haploceras) staszycii (Zeuschner); Avram 1976: 35, pl. 8: 4a, b.
1978 Haploceras staszycii (Zeuschner); Olóriz Sáez 1978: 15, pl. 1: 1a, b.
1979 Haploceras staszycii (Zejszner); Sapunov 1979: 43, pl. 7: 3a, b, text-fig. 4/6.
1980 Haploceras staszici (Zejszner); Malinowska et al. 1980: 444, pl. 136: 5.
?1984 Haploceras staszycii (Zeuschner); Rossi 1984: 89, pl. 31: 8.
1986 Haploceras (Haploceras) carachtheis (M) Zeuschner) morph elimatum; Enay and Cecca 1986: pl. 4: 1a, b.
1986 Haploceras (Haploceras) carachtheis (M) (Zeuschner) morph staszycii; Enay and Cecca 1986: pl. 4: 2a, b.
1988 Haploceras (Haploceras) staszycii (Zeuschner); Főzy 1988: 53, text-fig. 7 (with additional synonymy).
1999 Haploceras elimatum (Oppel); Kvantaliani 1999: 74, pl. 5: 4.
1994 Haploceras staszycii (Zeuschner); Zeiss et al. 1994: 370, pl. 1: 1.
2001 Haploceras (Haploceras) staszycii (Zeuschner); Zeiss 2001: 38, pl. 5: 1-3.
2002 Haploceras (Haploceras) carachtheis (Zeuschner) staszycii (Zeuschner); Pavia et al. 2002: 233: 159.
2020 Haploceras staszycii (Zeuschner); Sarti 2020: 87, pl. 3: 5.
2024 Haploceras staszycii (Zeuschner); Sarti 2024: 87, pl. 3: 5.
Type material: The holotype of Zejszner (1846) according to Malinowska et al. (1980: 444) was lost. They designated as a lectotype the specimen from Zittel (1870: pl. 27: 3, i.e., SNSB-BSPG AS III 805) from Rogoźnik. But according to the ICZN she designated neotype rather than lectotype.
Type locality: Rogoźnik, Poland.
Type horizon: Pieniny Klippen Zone, Rogoża Coquina Member, Fallauxi ammonite Zone, lower Tithonian.
Material.—The material under this study consists of 8 specimens from Moravian-Silesian museums (e.g., Z1585, Z1484, MM 010, MM Ge30259, B13570, PL2447, PL5059, and PL5061). The set was supplemented by direct study of originals in the Zittel collection housed in Munich (SNSB-BSPG AS III 803, 805, 806). The initial half of the ultimate whorl of the specimen PL5059 belongs, according to the visible suture line, to the phragmocone, and the final half to the body chamber.
Description.—Semi-involute specimens, with a high, relatively broad whorl and a narrow umbilicus. The low umbilical wall declines obliquely to the line of coiling. It continuously passes through a rounded zone to the slightly arched flanks of the whorl. The flanks also pass through a rounded zone to the medium-broad venter. On the juvenile specimen PL2447 and further on the adult specimen Z1484, a siphonal furrow is preserved on the phragmocone. The shell is smooth at the beginning. On the body chambers in the specimens under study or where the original shell is peeled off to expose the steinkern, faint, blunt to indistinct, and relatively closely spaced ribs are visible.
Measurements.—The 16 measurable specimens range from 29 to 83 mm in diameter (SOM 3). The Dmax of PL5059 is 44.0 mm; the end of the phragmocone is present at a D of approximately 32.0 mm. According to our measurement from a copy of the illustration of the lost Zejszner’s holotype (1846: pl. 4: 3a, b), at Dmax = 56.5 mm, H = 27.2 (0.48), U = 10.0 (0.18), B = 21.7 (0.38). Neotype SNSB-BSPG AS III 805 at D = 73.6 mm shows H/D = 0.51, U/D = 0.19, B/D = 0.36. Values of H/D in the material under study range from 0.47–0.52, U/D from 0.17–0.22, B/D from 0.36–0.41. The regression lines H/D, U/D, B/D of Ha. staszycii are accompanied by the microconch lines of Ha. elimatum in the graphs (Fig. 9A–C). The only clear difference in the course of the two regression lines is evident for the U/D values.
Remarks.—Malinowska et al. (1980: 444) stated that the holotype of Ha. staszycii disappeared and established the lectotype (but actually neotype). Haploceras staszycii reaches a medium size. Characteristic features include a relatively narrow umbilicus. Ribbing, if preserved, is usually weakly developed with regard to the smaller sizes of specimens of Ha. staszycii. Haploceras tithonium (Oppel, 1865) is a species that reaches a minor size of specimens and has higher whorls.
Stratigraphic and geographic range.—Enay and Cecca (1986) state that the stratigraphic range is Hybonotum to Ponti zones. Zeiss (2001) mentions the Tithonian of the Tethyan area with emphasis on the upper Tithonian. With regard to the assumption that Ha. staszycii forms a dimorphic pair with Ha. elimatum, the stratigraphic range should be identical to that of Ha. elimatum, i.e., the Tithonian to lower Berriasian. This argument is supported by specimens in Kvantaliani (1999) from Crimea, determined, however, as Ha. elimatum. Haploceras staszycii occurs in many localities of the Mediterranean and sub-Mediterranean areas. The neotype comes from Rogoźnik in Poland (lower Tithonian of the Pieniny Klippen Zone); it is also known from France, Italy, Romania, Hungary, Crimea, and Madagascar.
Our own collections from Kotouč Quarry come from Level 8 from locality 4 (Vašíček and Skupien 2016) with accompanying ammonites from the Fallauxi and Microcanthum ammonite zones and from the debris on Level 9.
Haploceras tithonium (Oppel, 1865)
Figs. 3C, 4C, 8E, F.
1865 Ammonites tithonius Opp.; Oppel 1865: 549.
1868 Ammonites tithonius Opp.; Zittel 1868: 82, pl. 14: 1a, b (lectotype), 2a–c, 3a, b.
1953 Haploceras tithonium Opp.; Slavin 1953: 53, pl. 3: 11, 12, text-fig. 4
1978 Haploceras tithonium (Opp.); Olóriz Sáez 1978: 20, pl. 1: 4a, b, 5.
1988 Haploceras (Haploceras) tithonium (Oppel); Főzy 1988: 53, text-fig. 8.
2020 Haploceras tithonium (Zittel); Sarti 2020: 90, pl. 4: 2A, B.
2024 Haploceras tithonium (Zittel); Sarti 2024: 90, pl. 4: 2A, B.
Type material: Lectotype (established herein) SNSB-BSPG AS III 797 from Hohenegger collection in Munich, originally figured by Zittel (1868: pl. 14: 1) as Ammonites tithonius Oppel.
Type locality: Štramberk Limestone, Castle Hill in Štramberk, Czech Republic.
Type horizon: Tithonian or lower Berriasian.
Material.—Three specimens present in the Hohenegger collection in Munich (SNSB-BSPG AS III 797–799) and four favourably preserved specimens, partly preserved as steinkerns, partly as external moulds, or with a recrystallized original shell (PL5061, PL5113, PL5115, Z2761).
Description.—Specimens of small- to medium-sized, semi-involute, with high, medium-broad whorls and with a narrow umbilicus. The flanks of the whorl are slightly arched. They reach the greatest breadth at approximately one-third of the whorl height. From this height, the flanks slope gradually to the umbilicus. Near the umbilicus, a rather sharp transition to the steeper part of the whorl declining to the line of coiling is observed. Near the venter, the flanks of the whorl are separated unclearly from the venter. The venter is slightly arched and medium-broad.
Our specimens appear to be smooth or, rarely, very thin indistinct ribbing is observed.
Remarks.—The remains of weak, dense ribbing are clear in the area around the line of coiling in Zittel’s (1868: pl. 14: 1a) specimen SNSB-BSPG AS III 797, desinated here as lectotype.
In the juvenile specimen PL5061, three lobes (E, A, U2) are favourably preserved. In specimen PL5113, part of the outer suture line is preserved with the external lobe, adjacent saddle E/A and part of the adventive lobe. The preserved part is essentially identical to the suture line of Ha. elimatum. A more completle external suture of Ha. tithonium is also illustrated by Slavin (1953: text-fig. 4).
Measurements.—The lectotype: D = 61.0 mm has H/D = 0.565, U/D = 0.13, B/D = 0.38. Eight specimens under study have sizes from 25 to 55 mm in diameter. The other six specimens under study range 21.6 to 54.2 mm in diameter. Their parameters are as follows: H/D = 0.51–0.56, U/D = 0.13–0.19, B/D = 0.35–0.38 (SOM 4). In Z2761, the phragmocone ends at a diameter D = 42 mm.
Remarks.—The flanks of the whorl of the specimen illustrated by Zittel (1868: pl. 14: 2a, b) are not as flat in cross-section as illustrated in Zittel’s fig. 2a. Haploceras tithonium is characterised—in contrast to other species in this genus—by a narrower umbilicus and a high and relatively slender whorl. Vígh (1984: 176, pl. 1: 1a, b) described Haploceras tithonium perumbilicatum as a new morphologically similar subspecies that differs from Ha. tithonium in having a smaller whorl height (H/D = 0.44), broader umbilicus (U/D = 0.23) and narrower whorl (B/D = 0.32). It could, under the name Haploceras perumbilicatum, represent a dimorphic counterpart of Ha. tithonium. With regard to the sizes of both taxa, however, it is not possible to determine which of them is a macroconch and which is a macroconch.
Stratigraphic and geographic range.—Data on the stratigraphic position of Ha. tithonium are relatively scarce. Enay and Cecca (1986) state the uppermost lower Tithonian from Spain, and Főzy (1988), the upper Tithonian from Hungary. According to Slavin (1953), Ha. tithonium occurs abundantly in the Upper Jurassic of the Ukrainian Carpathians.
Our dated collection from the Štramberk Limestone in the Kotouč Quarry on Level 6 corresponds to the basal upper part of the lower Tithonian. The specimens described by Zittel (1868) come from Štramberk Limestone of the currently abandoned quarry below Castle Hill in Štramberk.
Genus Hypolissoceras Breistroffer, 1947
Type species: Ammonites carachtheis Zejszner, 1846 (pl. 4: 1) by monotypy. Tatra Mountains in Poland, probably lower Tithonian.
Remarks.—The genus is characterised in particular by a series of short, small ribs (crenulations) limited to the venter of adult specimens.
Hypolissoceras carachtheis (Zejszner, 1846)
Figs. 3D, 8G–J.
1846 Ammonites carachtheis Zejszner 1846: pl. 4: 1a–d.
1868 Ammonites carachtheis Zeuschner; Zittel 1868: 84, pl. 15: 1a-e, ?2a, b, ?3a, b.
1879 Haploceras carachtheis Zeuschner; Fontannes 1879: 10, pl. 2: 3a, b.
1880 Ammonites (Haploceras) carachtheis, Zeuschner; Favre 1880: 29, pl. 2: 10.
1890 Haploceras carachtheis Zeuschner sp.; Toucas 1890: 577, pl. 13: 5a, b, pl. 15, ?fig. 7a, b; non fig. 8 (= ?Ha. leiosoma Oppel).
1893 Haploceras carachtheis Zeuschn.; Retowski 1893: 242, pl. 9: 10, ?11.
1953 Haploceras carachtheis Zeuschner; Arnould-Saget 1953: 7, pl. 1: 8a, b, 11a, b.
non 1960 Haploceras carachtheis Zeuschner; Drushchits and Kudryavcev 1960: 268, pl. 13: 2a, b (= ?Ha. staszycii), ?3a, b, ?text-fig. 72.
1962 Glochiceras carachtheis (Zejszner); Barthel 1962: 17, pl. 2: 1–4, pl. 3: 1–3 (figs. 1, 2 = Zittel 1868, pl. 15: 1d, e): 6, 7; text-fig. 3c; non pl. 3: 4, 5.
1965 Glochiceras carachtheis (Zeuschner); Houša in Špinar 1965: fig. VIII-255.
1976 Haploceras (Neoglochiceras) carachtheis (Zeuschner); Avram 1976: 36, pl. 8: 9a, b.
?1976 Haploceras (Haploceras) aff. elimatum (Oppel); Avram 1976: pl. 8: 10a, b.
1976 Haploceras (Neoglochiceras) carachtheis (Zeuschner); Patrulius and Avram 1976: 168, pl. 3: 8.
1978 Haploceras (Lingulaticeras) carachtheis Zeuschner; Olóriz Sáez 1978: 124, pl. 10: 6a, b, 7, 8.
?1979 Glochiceras (Glochiceras) carachtheis (Zejszner); Sapunov 1979: 64, pl. 14: 2a, b.
1980 Glochiceras (Glochiceras) carachtheis (Zejszner); Malinowska et al. 1980: 445, pl. 136: 7a–c.
1983 “Haploceras” carachteis (Zeuschner); Cecca et al. 1983: 114, pl. 1: 3.
1983 Haploceras (Neoglochiceras) carachtheis (Zejszner); Vašíček 1983: 98, pl. 3: 2, 3.
1984 Haploceras carachtheis (Zeuschner); Cecca et al. 1984: pl. 6: 1.
1986 Haploceras (Haploceras) carachtheis (m) (Zeuschner), resp. Haploceras (Haploceras) carachtheis (m) (Zeuschner) morph carachtheis; Enay and Cecca 1986: 49, pl. 2: 1a–c, 3a, b, 4a, b, ?5a, b, 7, ?8a, b, 9a, b, 10a, b; non pl. 2: 2; pl. 3: ?6a, b, 7a, b, 8a, b, ?9a, b, 10a, b, 11a, b, 12a, b, ?13a,b, 14a, b, 15a, b, 18a, b; non pl. 3: 1, 2a, b (= Haploceras woehleri), non pl. 3: 16a, b, 17a, b, 19a, b.
1986 Glochiceras carachtheis (Zeusch.); Kutek and Wierzbowski 1986: tab. 2.
1988 Haploceras carachtheis (Zeuschner); Főzy 1988: 59, pl. 3: 3, 4.
1989 Haploceras carachtheis (Zeuschner); Khimshiashvili 1989: 31, pl. 9: 3, 5, 7.
1990 Haploceras carachtheis (Zeusch.); Wierzbowski 1990: fig. 3.
1991 Haploceras (Haploceras) carachtheis (m) (Zeuschner) morph carachtheis; Cecca and Enay 1991: pl. 1: 8a, b.
1994 Haploceras carachtheis (Zeuschner); Főzy et al. 1994: 158, pl. 1: 16, 22.
?1994 Haploceras (Hypoglochiceras) carachtheis (Zeuschner); Zeiss et al. 1994: 370, pl. 2: 3.
1995 Haploceras carachtheis (Zeuschner); Főzy 1995: 136, pl. 20: 12.
1996 Haploceras carachtheis (Zeuschner); Wright et al. 1996: 10: 6/1c–e.
?2001 Haploceras (Hypolissoceras) carachtheis (Zeuschner); Zeiss 2001: 38, pl. 5: 4, 5.
?2004 Haploceras carachtheis (Zeuschner); Ettachfini 2004: 155, pl. 5: 1.
2013 Haploceras carachtheis (Zeuschner); Főzy and Scherzinger 2013: 215, pl. 4: 4a, b, ?fig. 3a, b, ?pl. 5: 1a, b, 2, 10.
2013 Haploceras carachtheis (Zeuschner); Szives and Főzy 2013: 295, pl. 4: 3.
non 2016 Haploceras carachtheis morph carachtheis (Zeuschner); Hoedemaeker et al. 2016: 168, pl. 17: 4–6 (= Hypolissoceras leiosoma Oppel).
2017 Haploceras carachtheis (Zeuschner); Főzy 2017: pl. 1: 3a, b, ?pl. 32, fig 1a, b.
2020 Haploceras carachtheis carachteis (Zeuschner); Sarti 2020: 91, pl. 4: 7.
?2020 Haploceras carachtheis leiosoma (Zittel); Sarti 2020: 91, pl. 4: 6A, B.
2024 Haploceras carachtheis leiosoma (Zittel); Sarti 2024: 91, pl. 4: 6A, B.
Type material: Holotype of Zejszner (1846) is apparently lost (Malinovska et al. 1980). Neotype (established herein) SNSB-BSPG AS III 47, originally figured by Zittel (1868: pl. 15: 1a–e) as Ammonites carachtheis Zejszner, 1846, and figured herein in Fig. 8J. The Zittel’s (1868) depiction of this specimen is partly idealized (compare with Fig. 8J).
Type locality: Štramberk Limestone, locality of Koňákov (formerly Koniakau), Czech Republic (the locality no longer available).
Type horizon: Tithonian or lower Berriasian.
Material.—Four specimens from Zittel (1868) and Barthel (1962) collections (SNSB-BSPG AS III 45, 48, 49; SNSB-BSPG AS III 47 is the neotype) and eight relatively complete specimens preserved as external moulds (B12689, Z1503, Z2981a, PL4370, Ge30342, Ge30344b) and a steinkern with the remnant of peristome (Z1385).
Description.—Semi-involute specimens, rather small in size, with high, medium-broad whorls and a narrow umbilici. The flanks of the whorl are flat to slightly arched. In the area of the umbilicus, a funnel-shaped zone is observed. On the best-preserved steinkern, it is obvious that the base of the whorl ends in a narrowly rounded shape, and the whorl still continues below to the dorsum in the shape of a narrow, slot-like form. The flat venter is clearly separated from the flanks by a rounded zone.
The inner whorls are smooth. On favourably preserved juvenile specimens, growth lines can be observed on the upper parts of the whorl. In adult specimens, short ventral ribs are present in the siphonal area in the final fifth of the ultimate whorl.
Measurements.—Specimens under our study have maximum diameter of approximately 42 to 44 mm. The phragmocones usually terminate at diameters ranging from approximately 23 to 26 mm, and on the specimen in Cecca and Enay (1991: pl. 1: 8), even at 35 mm. In the specimens from Štramberk, the first ventral ribs appear at D = 30–36 mm. Our own measurement of 13 specimens was supplemented with several values from Sarti (2020) data (SOM 5). Parameters: H/D = 0.46–0.51, U/D = 0.17–0.22, B/D = 0.32–0.34, and B/H = 0.65–0.71. Neotype with diameter D = 44.0 mm has H/D = 0.46, U/D = 0.23, B/D = 0.315.
Remarks.—Based on information obtained from Lidia Malinowska in 1980 (see Vašíček 1983: 99), the type material, i.e., the holotype of Hy. carachtheis presented by Zejszner (1846), which was merely illustrated without diagnosis, is lost. Historically, the closest material to the non-existent type specimen is the specimen depicted in Zittel (1868), pl. 15: 1a–e. For this reason, we designate it as a neotype herein.
The basic features of Hy. carachtheis include a narrow umbilicus and slender whorls with flat flanks. The main morphological feature is the onset of ribbing confined to the venter that appears on the final half to third of the body chamber at a shell diameter of more than 30 mm. The favourably preserved specimens bear a complexly formed peristome. On the venter, a short rostrum is present, and on the flanks, conspicuous, tongue-shaped lappets occur. The whole peristome is usually bounded by a clear constriction that is accompanied by a swollen rib on the front side. The preserved portion of the peristome indicates that the specimens Z1385 (see Fig. 8G1 here) and Z2981a (Fig. 8I1) represents adult individuals.
The umbilicus is mostly narrow (U/D less than 0.23), whereas broader umbilici are commonly reported in the literature (e.g., Zittel 1868, for SNSB-BSPG AS III 47 in pl. 15/1d, e; Zeiss 2001; Fözy and Scherzinger 2013). Due to the lack of comparative material, it is not clear whether this is a case of species variability or a case of different species. Haploceras carachtheis, considered by us as a microconch of Hypolissoceras woehleri (Oppel, 1865), is compared with Hy. woehleri in the description of the latter species.
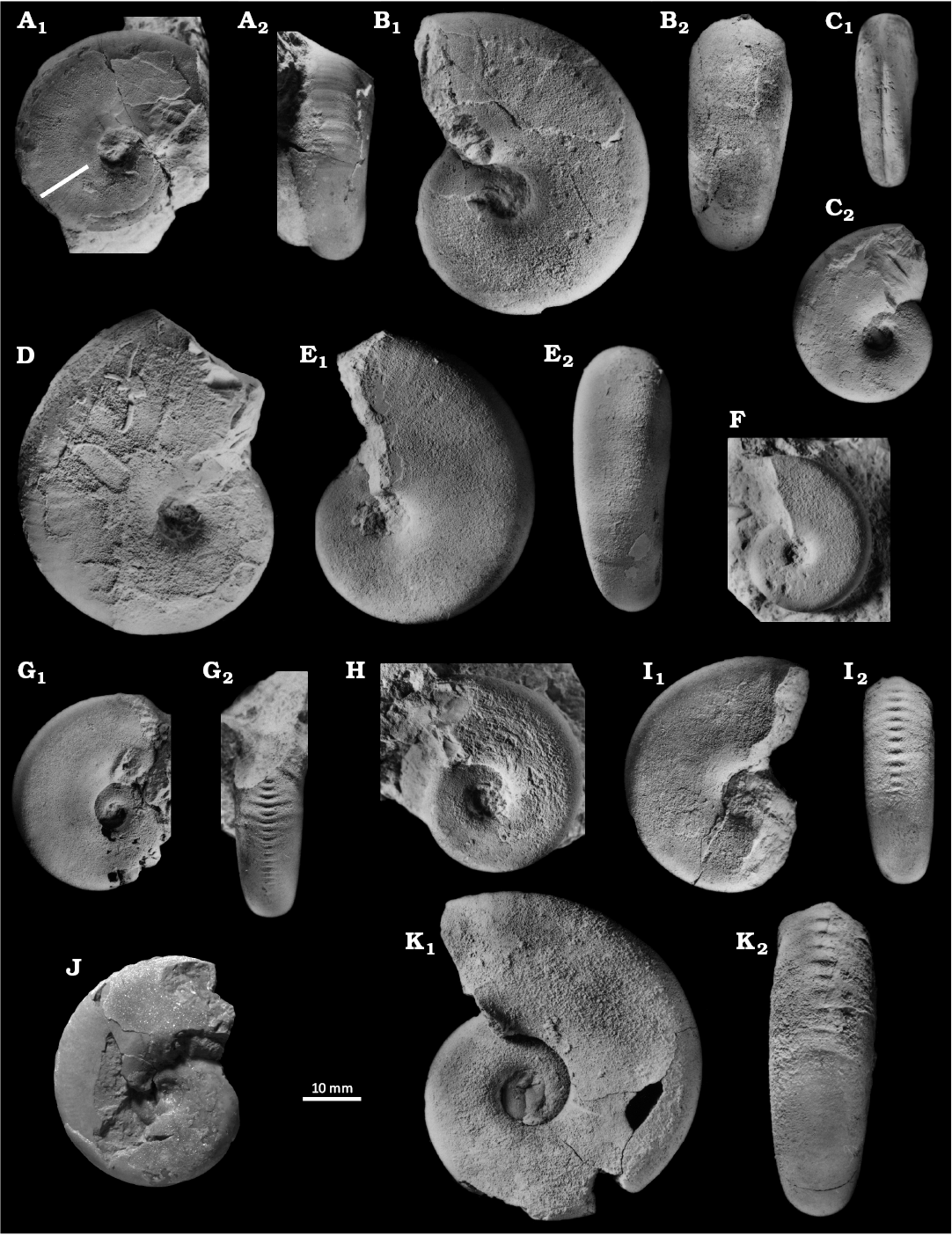
Fig. 8. Haploceratid ammonoid Haploceras staszycii (Zejszner, 1846) from Štramberk, Tithonian–lower Berriasian, Upper Jurassic–Lower Cretaceous. A. PL5059, lateral view (A1), ventral view demonstrating the ribbing on external mould (A2), Kotouč Quarry, debris on the 9th level. The straight line perpendicular to periphery of specimen show the margin between body chamber and phragmocone. B. Z1585, lateral (B1) and ventral (B2) views. C. PL2447, specimen with the siphonal groove, ventral (C1) and lateral (C2) views. D. MM010, lateral view. E, F. Haploceras tithonium (Oppel, 1865). E. PL5113, internal mould with incomplete outer suture line, lateral (E1) and ventral (E2) views, Kotouč Quarry, site No. 4 on the 8th level (Vašíček and Skupien 2016). F. PL5115, juvenile specimen, lateral view, Kotouč Quarry. G–J. Hypolissoceras carachtheis (Zejszner, 1846). G. Z1385 internal mould, lateral view (G1) with rest of peristome by umbilicus, ventral view (G2). H. B12689, external mould in lateral view. I. Z2981a external mould in lateral (H1) and ventral (H2) views. J. SNSB-BSPG AS III 47, lateral view of the lectotype (real state of preservation), Koňákov. K. Hypolissoceras woehleri (Oppel, 1865), MM06, external mould, lateral (K1) and ventral (K2) views. The straight lines perpendicular to periphery of specimen show the margin between body chamber and phragmocone. Photos Kristýna Mezihoráková (Ostrava, Czechia); except J, Alexander Nützel (SNSB-BSPG).
Stratigraphic and geographic range.—Haploceras carachtheis occurs in almost the whole Mediterranean and the sub-Mediterranean area, from the Himalayas to northwestern and eastern Africa. According to Enay and Cecca (1986), it occurs from the lower Tithonian (Hybonotum Zone) to the lower Berriasian inclusive. The lower Berriasian occurences are also mentioned by, e.g., Arnould-Saget (1953), Patrulius and Avram (1976), and Szives and Főzy (2013) with a detailed analysis of Ha. carachtheis occurrences.
Specimens described by Zittel (1868) come from the Štramberk Limestone in Štramberk and from Koňákov. We could not find this species in Štramberk area ourselves. In the Czech Republic, it is also known from the lower Tithonian of the Magura Group of nappes in Roštín (Vašíček 1983).
Hypolissoceras woehleri (Oppel, 1865)
Figs. 3E, 6A, B, 4D, 8K.
1865 Ammonites Wöhleri Opp.; Oppel 1865: 549.
1868 Ammonites Wöhleri Opp.; Zittel 1868: 84, pl. 14: 4a, c.
1986 Haploceras (Haploceras) carachtheis (m) (Zeuschner) morph carachtheis; Enay and Cecca 1986: 49, pl. 3: 1, 2a, b.
?1988 Haploceras (Hypolissoceras) carachtheis (Zeuschner); Főzy 1988: 59, pl. 3: 3, 4.
1988 Haploceras (Haploceras) wohleri (Oppel); Főzy 1988: 55, pl. 3: 1, 2, text-fig. 9.
Type material: The holotype by monotypy (SNSB-BSPG AS III 811) is a poorly preserved specimen (Zittel 1868: pl. 14: 4). The original illustration of which is quite idealized. The body chamber is almost completely preserved but the phragmocone is missing. The peristome, i.e., both the rostrum and especially the flank, is distorted. The greater part of the holotype (with exception of the area around peristome, where on both sides, remains of the original shell occur) is a steinkern. There is a prominent granulation on the venter, however, this is very likely a product of diagenesis.
Type locality: Štramberk Limestone, Štramberk, Czech Republic.
Type horizon: Tithonian or lower Berriasian.
Material.—In addition to the holotype, more than 10 adult specimens preserved as external moulds and one specimen with remains of the original test, partly split off from the body chamber (Z1817), are available. The best-preserved museum specimens are Z2305, Z4171, Z4206, Z4210, MM 04, MM 06–09, and Ge30355.
Description.—Specimens semi-involute, rather large in size, with relatively high whorls with slightly arched flanks, and a visibly, although unclearly, separated venter. The umbilicus is relatively narrow. The flanks of the whorl, which reach the greatest breadth at approximately the lower fifth of its height, decline steeply without any edge to the line of coiling. Around the umbilicus, a funnel-like shape is observed. The venter is only slightly arched.
The inner whorls are smooth. At the beginning of the final part of the ultimate whorl, unclear ribs may be seen on the venter. Near the peristome in all larger specimens, at least several (approximately 4–7) short, widely spaced, relatively strong blunt ventral ribs appear.
Suture line. On specimen MM08, some incomplete suture lines bound by merely an adventive lobe A and adjacent parts of saddles are preserved. The lobe is strongly articulated, formed similarly to the suture lines of the genus Haploceras.
Measurements.—The holotype reaches diameter of approximately 117 mm. Because of imperfect preservation, the measurements are rather ambiguous; at D = 116.0 mm, H is ca. 58.5 mm (0.50), U ca. 23.0 mm (0.20), B ca. 42.5 mm (0.37). The largest specimen from the Moravian-Silesian collection (Z4171) reaches a diameter of only approximately 61 mm. By D = 60.8 mm H/D is 0.435, U/D = 0.25, B/D = 0.33. Its phragmocone ends at D = 46 mm. The first ventral ribs appear at D = 53–60 mm. Usual parameters in the collection under study are as follows: H/D = 0.43–0.47, U/D = 0.24–0.26, B/D = 0.32–0.36. Főzy (1988) reports specimens larger in size, i.e., with a diameter of up to 146 mm. The measurements for constructing the charts on Fig. 10 are listed in SOM 6.
Remarks.—With reference to the morphology of the holotype, we assume that Hy. woehleri represents a macroconch to Hy. carachtheis. Hypolissoceras woehleri differs from Hy. carachtheis above all in different shell size. Figure 10A–C shows that the H/D and B/D values for both species are almost identical. A smaller difference is evidenced only by U/D values (Fig. 10B). The ventral ribs of Hy. woehleri, as shown by specimens from the Moravian Museum, begin only at shell diameters of approximately 53 to 60 mm, whereas in Hy. carachtheis, they begin at diameters of approximately 30–36 mm. The holotype of Hy. woehleri (Dmax of 117 mm) with ambiguously measurable parameters differs somewhat from the just-described macroconchs in the ratios H/D, U/D, and B/D. This is probably associated with the smaller sizes of specimens in the collection under study and with unfavourable preserved parameters of the holotype.
Stratigraphic and geographic range.—In contrast to Hy. carachtheis, data on the stratigraphic range and geographical distribution of Hy. woehleri are limited. The holotype comes from not well constrained stratigraphic position of Štramberk Limestone in Štramberk (probably from the currently non-existent Castle Quarry). According to Főzy (1988), this species occurs in the “middle” and upper Tithonian in Hungary. Given that we assume these species form a dimorphic pair, the stratigraphic range of Hy. woehleri should be the same as that for microconchs.
The holotype of Hy. woehleri comes from Štramberk, and the specimen in Enay and Cecca (1986: pl. 3: 1) from Le Pouzine, France (Fallauxi Zone). The specimens from the Moravian-Silesian museums lack detailed geographic and stratigraphic information.
Hypolissoceras subtilius (Zittel, 1870)
Fig. 6D.
1870 Haploceras carachtheis var. subtilior; Zittel 1870: 54, pl. 27: 11a–c.
1986 Haploceras carachtheis var. subtilior; Enay and Cecca 1986: 39, text-fig. 4.
?1986 Haploceras (Haploceras) carachtheis (m) (Zeuschner); Enay and Cecca 1986: pl. 2: 8a, b, pl. 3: 8a, b.
Type material: Holotype (by monotypy) SNSB-BSPG AS III 42 (Zittel 1870: pl. 27: 11)
Type locality: Rogoźnik, Poland.
Type horizon: Pieniny Klippen Zone, Rogoża Coquina Member, Fallauxi ammonite Zone, lower Tithonian.
Material.—Holotype and a single weathered, slightly corroded external mould (Z2767a), both small in size.
Description.—The semi-involute specimen has a medium-high whorl and a medium-broad umbilicus. The flanks of the whorl are slightly arched. The greatest breadth of the whorl is close to its base. From the mentioned area, the whorl declines, through a rounded zone, continuously to the line of coiling. The umbilical wall is very low. The flanks slope gradually to the venter. In the more juvenile part, the flanks pass continuously to a rounded venter; in the final part of the ultimate whorl, the venter is slightly arched with an indication of its separation from the flanks.
The juvenile half of the ultimate whorl is smooth. At the beginning of the second half, indistinct ribs are seen on the venter. Only in the vicinity of the aperture, 7 straight and very short but distinctive ventral ribs are observed. On the unphotographed, more poorly preserved opposite side, remains of the peristome are visible; they indicate that it is probably an adult specimen.
Measurements.—The holotype preserved as a steinkern has a maximum diameter of 23 mm. At D = 22.9 mm, H = 9.9 mm (0.43), U = 6.1 mm (0.27), and B mm = 7.0 (0.305). Specimen Z2767a reaches a maximum diameter of 30 mm. At D = 30.0 mm, H =12.6 mm (H/D = 0.42), U = 9.3 mm (U/D = 0.31), B = 9.4 mm (B/D = 0.31), and B/H = 0.75. The first ventral ribs appear at a D of approximately 27 mm.
Remarks.—Hypolissoceras subtilius differs from the similar small-sized Hypolissoceras leiosoma (Oppel in Zittel, 1868) in having a much broader umbilicus and smaller height and breadth of whorls. Adult specimens with the preserved peristome, designated by Enay and Cecca (1986: pl. 2: 8, pl. 3: 8) as Ha. carachtheis (m), probably also belong to Hy. subtilius.
Stratigraphic and geographic range.—According to Enay and Cecca (1986: text-fig. 4) Hy. subtilius occurs sporadically in the uppermost lower Tithonian (Ponti Ammonite Zone) in Spain, The holotype comes from Rogoźnik in Poland, most likely from the lower Tithonian. Z2767a from the museum collection in Opava is labelled with the locality name Štramberk only.
Genus Volanites Enay and Cecca, 1986
Type species: Due to the fact that Enay and Cecca (1986) did not fulfill all the prescribed requirements when determining the type species, we designate Ammonites verruciferus Zittel in Benecke 1869 herein, represented by the lectotype specimen SNSB-BSPG AS III 809, originally figured by Zittel (1870: pl. 27: 8). Tithonian, Volano, Italy.
Volanites verruciferus (Zittel, 1869)
Fig. 6C.
1869 Ammonites verruciferus Zittel in Benecke 1869: 145.
1870 Haploceras verruciferum Meneghini sp.; Zittel 1870: 52, pl. 27: 8 (lectotype), 9, 10.
1890 Haploceras verruciferum Meneghini sp.; Toucas 1890: 577, pl. 13: 7.
1978 Haploceras verruciferum Meneghini; Olóriz Sáez 1978: 23, pl. 1: 6–8.
1983 Haploceras verruciferum (Meneghini) in Zittel; Cecca et al. 1983: 116, pl. 1: 4.
1984 Haploceras verruciferum (Meneghini); Rossi 1984: 90, pl. 31: 7, pl. 32: 7.
1986 Haploceras (Volanites) verruciferum (Zittel); Enay and Cecca 1986: 48, pl. 1: 2-8, pl. 2: 11–15; non pl. 1: 1 and 9 ( = Haploceras cassiferum Főzy, 1988).
?1986 Haploceras verruciferum (Zittel); Sarti 1986: 490, pl. 1: 6.
?1986 Haploceras cf. verruciferum (Men.); Kutek and Wierzbowski 1986: tab. 2.
1988 Haploceras (Hypolissoceras) verruciferum (Zittel); Főzy 1988: 63, pl. 4: 3, 4, pl. 5: 1, pl. 6: 3–6, text-figs 13, 14 (cum syn.).
1989 Haploceras (Volanites) verruciferum Zittel; Khimshiashvili 1989: 31, pl. 9: 2.
1990 Haploceras verruciferum (Zittel); Főzy 1990: 327, pl. 1: 10, 11.
non 1991 Haploceras (Volanites) verruciferum (Zittel); Cecca and Enay 1991: 45, pl. 2: 1–4 (= ?V. cassiferum Főzy).
1994 Haploceras verruciferum (Zittel); Főzy et al. 1994: 158, pl. 1: 23.
1994 Haploceras (Volanites) verruciferum (Zittel); Zeiss et al. 1994: 370, pl. 2: 1.
1995 Haploceras verruciferum (Zittel); Főzy 1995: 136, pl. 20: 4, 7.
2002 Haploceras (Volanites) verruciferum (Zittel); Pavia et al. 2002: 234, figs. 160/1–3, ?4.
2013 Haploceras verruciferum (Zittel); Főzy and Scherzinger 2013: 216, pl. 5: 6–8, 12, ?pl. 4: 1, 2a,b, ?pl. 5: 9.
2017 Haploceras verruciferum (Zittel); Főzy 2017: pl. 1: 1, pl. 12: 3, pl. 23: 1, pl. 32: 17.
2020 Haploceras (Volanites) verruciferum (Zittel); Sarti 2020: 82, pl. 3: 3A, B, 4A, B.
2024 Haploceras (Volanites) verruciferum (Zittel); Sarti 2020: 82, pl. 3: 3A, B, 4A, B.
Type material: The lectotype (SNSB-BSPG AS III 809) originally designated by Enay and Cecca (1986: 49).
Type locality: Volano, Italy.
Type horizon: Tithonian or lower Berriasian.
Material.—Two specimens from Zittel collection (SNSB-BSPG AS III 807 and SNSB-BSPG AS III 809—lectotype), one relatively well-preserved juvenile external mould (Z2981b) without peristome. The specimen Z2981b is labelled only with the locality of Štramberk.
Description.—Semi-involute specimens, with medium-high whorls and a medium-broad umbilicus. The flanks of the ultimate whorl are moderately arched. They are broadest near the whorl base. From that point, the whorl declines, through a low rounded zone, to the line of coiling. The flanks of the whorl decline gradually towards the venter. The venter is rounded and separated from the flanks of the whorl.
The ultimate whorl is smooth. In one specimen (Z2981b) only blunt ribs convexly bent towards the peristome are observed.
Measurements.—The specimens of V. verruciferus illustrated in Zittel (1870: pl. 27: 8–10) reach, according to our measurements, sizes of 42–62.5 mm in diameter. In the studied small collection there are two size extreme specimens: the smallest with diameter about 38 mm, the largest with diameter 105 mm. The parameters of specimens in SOM 7 are as follows: H/D = 0.38–0.44, U/D = 0.30–0.33, and B/D = 0.29–0.32. The lectotype by D = 62.5 mm has H/D = 0.44, U/D = 0.30, B/D = 0.27. The largest specimen or The only specimen in our own collection (or Moravian-Silesian collections) has maximum size 51 mm.
Remarks.—We found only one specimen that should belong to V. verruciferus in the Moravian-Silesian collections studied by us. Judging from its size, it is a juvenile specimen without the characteristic final swollen rib on the venter near the peristome. However, the size parameters and morphology are similar to those of V. verruciferus. In other regions, a similar species but larger species, Volanites cassiferus (Főzy, 1988) occurs together with V. verruciferus. According to Főzy (1988), V. cassiferus reaches a diameter of approximately 100 mm or more. Főzy’s (1988) specimen J-8048 from Lókút section (Hungary) has measurements of H/D = 0.43, U/D = 0.27, and B/D = 0.30 at D = 105 mm. In accordance with Főzy (1988) and Főzy and Scherzinger (2013), we assume that V. cassiferus represents a macroconch of the microconch represented by V. verruciferus.
Stratigraphic and geographic range.—According to data in the literature, V. verruciferus occurs abundantly in the Mediterranean area (Verruciferus Zone) of southern Spain, southeastern France, Sicily, northern Italy, Hungary (Bakony, Gerecse), and Abkhazia. Főzy’s (1988) type material of V. cassiferus comes from the lower Tithonian of Lókút section, Hungary. According to Zeiss et al. (1994) and Zeiss (2003), V. verruciferus was previously considered an index species in the higher part of the lower Tithonian or near base of middle Tithonian. Detailed stratigraphic position of the specimen found in the area around Štramberk is not known.
Discussion
Considerable inconsistency in the classification of haploceratids is evident at the species and generic levels. In the present paper we propose an integrated concept based on dimorphic pairs of haploceratids. We recognize herein four dimorphic pairs, namely micro- and macroconchs, designated by us with individual species names. We abolished previous classification of the genus Haploceras based on subdivision into subgenera (Enay and Cecca 1986).
Our classification is based on the direct study of approximately one hundred favourably preserved specimens from the Štramberk Limestone and deposited in Moravian-Silesian museums, specimens collected by us, and study of published materials in which the morphology is sufficiently documented (including type series of Zittel 1868, 1870).
We furthermore performed the statistical analysis of the parameters H/D, U/D, and B/D based on the well-preserved specimens. We evaluated also other characters, e.g., whorl cross-sections, peristome shape, and sculpture. Several well-preserved steinkerns allowed the drawing of septal suture lines.
Based on the overall analysis of the available material and literature sources, we recognize four dimorphic pairs in the framework of three genera in Tithonian/early Berriasian haploceratids. The first pair is formed by Ha. elimatum (macroconchs) and Ha. staszycii (microconchs). The two species when preserved as external moulds, bear similar indistinct ribbing patterns on the venter and the flanks of the whorls. While the macroconchs reach diameters of up to 200 mm, microconchs reach diameters of only 80 mm. The microconchs have slightly broader whorls and somewhat narrower umbilicus (see Fig. 9). This dimorphic pair was already suggested by Barthel (1962: 12). The position of Ha. subelimatum (Fontannes, 1879) remains uncertain from the point of view of dimorphism, with specimens of a similar size. It has clearly narrower whorls than the two previous species.

Fig. 9. Relationship between size parameters of Haploceras elimatum (Oppel, 1865) microconchs and Haploceras staszycii (Zejszner, 1846). A. Height (H) of whorl on diameter (D). B. Umbilicus breadth (U) on diameter. C. Whorl breadth (B) on diameter.
The second possible dimorphic pair in Haploceras are Ha. tithonium and Ha. perumbilicatum Vígh, 1984. In this case, we have only specimens of Ha. tithonium in our own collection. Nevertheless, we consider Ha. tithonium perumbilicatum established by Vígh (1984), to be a dimorphic pair. The two morphologically similar species differ in the height and breadth of the whorls as well as in the breadth of the umbilicus. With reference to the insufficiently known maximum sizes of both species (according to the published data, approximately 50 mm in both cases), it is not yet possible to decide which species represents a macroconch and which a microconch.
The third pair is formed by Hypolissoceras carachtheis and Hy. woehleri. Zejszner’s original holotype (Hy. carachtheis) is lost, and thus we designated its neotype herein. Both species are characterised by specific ribbing that is developed only on the venter. Hyppolisoceras carachtheis (microconch) attains a smaller size, less than ca. 55 mm, while Hy. woehleri (macroconch) reaches a size of 120 to 140 mm. The cross-sections of whorls are also somewhat different; in Hy. carachtheis, the whorls are slenderer, whereas in Hy. woehleri, the whorls are lower, and the umbilicus is broader.
Another less perfectly clear dimorphic pair, due to the shortage of our own material, is based on accounts from published data. In accordance to Főzy (1988) and Főzy and Scherzinger (2013), we treat Volanites verruciferus and V. cassiferus (Főzy 1988: 59) as a possible dimorphic pair. Volanites verruciferus (microconch) reaches a maximum size of almost 70 mm, whereas V. cassiferus (macroconch) reaches a maximum size of approximately 100 mm.
In the collections of haploceratids from Štramberk deposited in the Moravian-Silesian museums, specimens very small in size (less than 30 mm) are missing, in contrast to the haploceratids from the Štramberk Limestone published by Zittel (1868, 1870) and from some other European localities. This may be explained by the fact that small specimens of ammonites were not attractive to the collectors of fossils in the Štramberk area. The exception in museum material is a single, small specimen of Hypolissoceras subtilius (originally Haploceras carachtheis var. subtilior Zittel, 1870). Its dimorphic pair could be Hypolissoceras leiosoma (Oppel in Zittel, 1868), with a distinctly narrower umbilicus. In both species, ventral ribbing or crenulation are less noticeable than that of Hy. carachtheis.
Peristomes are rarely preserved in Tithonian haploceratids; however, still and ample number of specimens of various sizes displays them, e.g., Zittel (1868: pl. 13: 2, 4, 7; pl. 14: 4b, ?5; pl. 15: 1b, 1e). Peristomes in his lithographic tables are usually complicated in nature and were usually drawn more or less embellished and even idealized. They are characterised by a rostrum on the venter and robust lateral tongue-shaped lappets. This holds true especially for the large specimens of Ha. elimatum (Zittel 1868: pl. 13: 7a) and Hy. woehleri (Zittel 1868: pl. 14: 4b) that are considered macroconchs. But it should be underlined that such terminal modifications of the aperture are normally typical of microconchs. It is possible that specimens like these started as females with a macroconchiate shape, but then changed their sex to males and finally formed the characteristic lappets. Examples of such phenomena were first described in Parent et al. (2008) and more extensively discussed by Frau and Boursicot (2021). Complicated peristomes were also illustrated by other authors as well (e.g., Barthel 1962: pl. 3: 1, 2; Olóriz Sáez 1978: pl. 10: 6a; Enay and Cecca 1986: pl. 2: 1, 8, 9; pl. 3: 1, 4; Zeiss 2001: pl. 5: 4), which should be considered as typical of microconchs.
In haploceratids, the possibility for sexual dimorphism is discussed in several papers (e.g., Zeiss 2001; Főzy and Scherzinger 2013). The theory of dimorphism and possible polymorphism was analysed thoroughly by Matyja (1986). Based on the study of Oxfordian ammonites, Matyja reported the possibility of trimorphism, where in addition to microconchs and macroconchs, specimens smaller than microconchs with lappets may occur. Such form named miniconchs, i.e., miniature specimens are considered by Matyja (1986) as an evidence of polymorphism. The haploceratids studied by us do not provide sufficient material to reach similar conclusions, although our material includes several small shells of Ha. elimatum. Our knowledge of dimorphic pairs, in the case where pairs cannot be established according to their peristomes, is based on different sizes of shells, similarity of sculpture, smaller differences in the parameters H/D, U/D, and B/D. The crowding of septa, demonstrating the adulthood of individuals, is proven in our material merely in several large steinkerns of Ha. elimatum and never in small specimens.
In the majority of Tithonian haploceratids, suture lines were already known in the period of publication of the monographs by Zittel (1868, 1870). The suture line of Ha. elimatum, illustrated by Zittel (1868: pl. 13: 6) has saddles accompanying lobe E drawn somewhat differently. Zittel’s right saddle by E has the all partial branches practically similar, typical specimens have the branches of different size. We regard the suture line of a juvenile specimen in Wiedmann (1966b: text-tab. 43a) with a trifid lobe A as an authoritative suture line for Ha. elimatum. Specimen Z4224, illustrated here in Fig. 4A, bears the same suture line. The suture line of Ha. staszycii illustrated by Zittel (1870) in pl. 27: 6, and in our Fig. 4B appears to be the same as that of Ha. elimatum.
The suture line of Ha. tithonium in Zittel (1868: pl. 14: 2b, c) is not drawn reliably. A suture line of the same type is well-illustrated by Slavin (1953: text-fig. 4). This suture line and the incomplete suture line of our specimen from Level 6 (Fig. 4C) are similar to the suture line of Ha. elimatum.
The suture line of Hypolissoceras carachtheis in the lost holotype was drawn by Zejszner (1846: pl. 4: 1d) and Zittel (1868) states that Zejszner’s illustration is not reliable. Zittel (1868: pl. 15: 3a, b) illustrated the suture line of Hy. carachtheis from a whorl fragment of his specimen. Neither Barthel (1962) nor Vašíček et al. (2018) succeeded in finding the latter specimen, which should be deposited in SNSB-BSPG. It cannot be determined if the suture lines of Hy. carachtheis illustrated by Zittel (1868) differ from the suture lines of the genus Haploceras. The same is true for the suture line of Hy. woehleri, in which only an adventive lateral lobe is preserved (Fig. 4D).
The incomplete suture line of Volanites verruciferus, which is illustrated by Zittel (1870: pl. 27: 10), if reliable, differs slightly from the suture lines of the two previously mentioned genera in terms of the shapes of most saddles and lobes.
Olóriz Sáez (1976: text-fig. on p. 33) summarily illustrates the suture lines of quite a number of species of haploceratids. However, due to the small scale of the illustrated suture lines and their schematic nature, they are not very helpful.
Statistics.—The measured values of the parameters (H, U, and B) in representatives of the genera Haploceras (Ha. elimatum, Ha. staszycii) and Hypolissoceras (Hy. carachtheis, Hy. woehleri) were compared in relation to the diameter (D) using multivariate regression analysis (see Shea and Vecchione 2002 for details on the statistical procedure). Statistical analyses were performed using the Excel software package. The dependence is expressed by the regression lines and the coefficient R2 determination (Hammer and Harper 2006). All measured values, from which the graphs were constructed, are presented in SOM 1–6. We have created statistical charts to compare dimension parameters between macroconchs and microconchs in Ha. elimatum (Fig. 7); between microconchs Ha. elimatum and Ha. staszycii (Fig. 9); and between Hy. carachtheis and Hy. woehleri (Fig. 10).

Fig. 10. Relationship between size parameters of Hypolissoceras carachtheis (Zejszner, 1846) and Hypolissoceras woehleri (Oppel, 1865). A. Height (H) of whorl on diameter (D). B. Umbilicus breadth (U) on diameter. C. Whorl breadth (B) on diameter.
A statistical proof of the dependence of whorl height and whorl breadth to diameter shows a high significant linear correlation (R2 more then 0.9). In the case of Ha. elimatum, the difference between microconchs and macroconchs cannot be observed. All measured specimens therefore belong to the macroconch category. In the adult specimens, the phragmocone ends at a diameter of approximately 105 mm. In the case of the macroconch, it is only possible to observe greater variability between the umbilicus breadth from a diameter of 80 mm.
The comparison of the microconch Ha. elimatum and Ha. staszycii (Fig. 9A–C), displays noticeable difference in umbilicus breadth. In the case of Ha. staszyci, the umbilicus breadth is smaller than in Ha. elimatum. It is also possible to observe more significant variability (linear correlation is 0.87).
Based on its morphology we assume that the holotype of Hy. woehleri represents a macroconch of Hy. carachtheis. Hypolissoceras woehleri differs from Hy. carachtheis above all in different shell size (Fig. 10A–C). Similarly, the difference in umbilicus breadth of Hy. carachtheis and Hy. woehleri is also noticeable. In the case of Hy. carachtheis, the umbilicus breadth is smaller than in Hy. woehleri.
Stratigraphy.—The majority of ammonoid specimens from the Štramberk Limestone from the Silesian Unit do not have accurate localizations and thus their stratigraphic positions are not known. It can be inferred from the data in the literature that the most stratigraphically important species described here is Volanites verruciferrus, which is usually given as an equivalent of the Semiforme Ammonite Zone from the higher part of the lower Tithonian (see, e.g., Zeiss et al. 1994; Zeiss 2003). Most data on the stratigraphic range are for the species Haploceras elimatum and Hypolissoceras carachtheis. Both species are, according to data based on our collection and in the literature, known from the whole Tithonian to the lower Berriasian. The occurrence of Ha. elimatum in the lower Berriasian is also confirmed by our newest findings in the Kotouč Quarry (Vaňková et al. 2019).
Aptychi.—Another peculiarity of the haploceratids is a strongly calcified lower element of the jaw apparatus in their body chambers. It consists of two symmetrical valves known as aptychi. In recent years, Engeser and Keupp (2002) and Schweigert (2009), among others documented their occurences. Thick-walled, ribbed calcite valves of aptychi, preserved in pairs, are most likely found in situ only in ammonites of the family Haploceratidae in Štramberk Limestone. Such examples with aptychi are usually only seldom illustrated in the literature. Finds of isolated (dispersed) specimens of aptychi are, in contrast, quite common.
In the Tithonian, specimens that belong, in the parataxonomic classification of aptychi (e.g., Měchová et al. 2010) to the genera Punctaptychus Trauth, 1927, and Beyrichilamellaptychus Turculet, 1994, occur more frequently. Some other genera from this artificial classification, similar to the latter genus, are included under the older common name lamellaptychi or in accordance with the proposal of Engesser and Keupp (2002), in aptychophoran aptychi (from Aptychophora), also occur.
In the collection of haploceratids processed by Barthel (1962), deposited in SNSB-BSPG, a specimen not illustrated by Barthel (1962) but labelled Haploceras elimatum (Tithonian from Le Pouzin) possesses two valves of Punctaptychus punctatus (Zittel, 1868) in the body chamber. Although the valves are not in paired positions (see Fig. 6F), with reference to their sizes (length L of ca. 48 mm) and the size of the body chamber of the ammonite, this finding can be regarded as evidence that the genus Haploceras is a bearer of aptychi of the genus Punctaptychus. Similarly, aptychi Beyrichilamellaptychus beyrichi (Oppel, 1865) may belong to Hypolissoceras woehleri based on equivalent dimensions of the body chamber and the maximum sizes of the valves (see an example in Fig. 6E).
The stratigraphic range of dimorphic pairs of the species of genera given here, i.e., Haploceras and Hypolissoceras, is from the Tithonian to the lower Berriasian. This corresponds essentially to the same time span of occurrence of aptychi of P. punctatus and B. beyrichi, for which Engeser and Keupp (2002) proposed the informal name Aptychophora.
Conclusions
According to our understanding, the haploceratids from the Štramberk Limestone belong to three genera: Haploceras, Hypolissoceras, and Volanites. A total of seven species have been identified and described. In the case of the lost holotypes, we designated lectotypes or neotypes.
Smooth juvenile and adult specimens of Haploceras elimatum analysed statistically as possible “microconchs” and “macroconchs” actually belong all to the to the macroconch of Ha. elimatum.
Haploceratids form dimorphic pairs. In our material, the dimorphism is evidenced by Haploceras staszycii (m) with Ha. elimatum (M), another pair is probably Haploceras tithonium with Haploceras subelimatum. The latter species is absent from our material. The determination of dimorphism is based on published data but we could not determine which species is macroconch and which is microconch.
A statistically documented dimorphic pair is also formed by Hypolissoceras carachtheis (m) and Hypolissoceras woehleri (M).
We assume that another dimorphic pair consists of Volanites verruciferus (m) and Volanites cassiferus (M). We were unable to find both species in Štramberk during our own sampling. The Silesian Museum in Opava has a single juvenile specimen deposited, possibly belonging to V. verruciferus.
From a stratigraphic point of view, the most important species should be V. verruciferus, sometimes considered as an index species. According to literature data, it occurs at the level of the Semiforme Ammonite Zone (base of the middle Tithonian).
The other haploceratid species described here from the Štramberk Limestone have a broad stratigraphic range from the lower Tithonian to the lower Berriasian inclusive. They are insignificant for the determination of the J/K boundary and detailed stratigraphy.
Acknowledgements
We are very grateful to Horacio Parent (Rosario University, Argentina) for his useful comments and remarks that helped to improve first version the manuscript. Authors would like to thank Günter Schweigert (State Museum of Natural History, Stuttgart, Germany) and Andrzej Wierzbowski (Warsaw University, Poland) for their helpful reviews and improvements. We would like to thank Andrzej Kaim and Krzysztof Hryniewicz (Institute of Paleobiology Polish Academy of Sciences, Warszawa, Poland) for their helpful comments and help with final editing of the manuscript. The authors are grateful to Kristýna Mezihoráková (Ostrava, Czech Republic) for taking photographs, Alexander Nützel (SNSB-BSPG) for photo on Fig. 8J, and Radmila Jelínková (Ostrava, Czech Republic) for the translation of the manuscript into English. The research is supported by Czech Science Foundation project no. 20-10035S and by grant m SP2025/086 from the Ministry of Education, Youth and Sports of the Czech Republic.
Editor: Andrzej Kaim
References
Arkadiev, V,V., Atabekian, A.A., Baraboshkin E.J., and Bogdanova, T.N. 2000. Stratigraphy and ammonites of Cretaceous deposits of South-West Crimea. Palaeontographica A255 (4–6): 85–128. Crossref
Arnould-Saget, S. 1953. Les ammonites pyriteuses du Tithonique supérieur et du Berriasien de Tunisie centrale. Annales des Mines et de la Géologie 10 (for 1951): 1–133.
Avram, E. 1976. Les fossiles du flysch éocrétacé et des calcaires tithoniques des hautes vallées de Doftana et du Tirlung (Carpathes Orientales). Mémoires de l’Institut de Géologie et de Géophysique Bucarest 24: 5–74.
Barthel, K.W. 1962. Zur Ammonitefauna und Stratigraphie der Neuburger Bankkalke. Bayerische Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse, Abhandlungen (Neue Folge) 105: 1–28.
Benecke, E.W. 1869. Geologische Beobachtungen aus dem Central Appenninen. Geognostisch-paläontologische Beiträge 2: 88–178.
Benzaggagh, M. and Atrops, F. 1997. Stratigraphie et association de faune d’ammonites des zones du Kimméridgien, Tithonien et Berriasien basal dans le Prérif interne (Rif, Maroc). Newsletter on Stratigraphy 35 (3): 127–163. Crossref
Besnosov, N.V. and Mikhailova, I.A. 1991. Higher taxa of Jurassic and Cretaceous Ammonitida. Paleontological Journal 25 (4): 1–19.
Beznosov, N.V. and Mikhailova, I.A. 1983. Evolution of the Jurassic-Cretaceous ammonoids [in Russian]. Doklady Akademii nauk SSSR, Paleontologiâ 269 (3): 733–737.
Boughdiri, M., Olóriz, F., Marques, B.L., Layeb, M., de Matos, J., and Sallouhi, H. 2005. Upper Kimmeridgian and Tithonian ammonites from the Tunesian «dorsale» (NE Tunesia): updated biostratigraphy from the Jebel Oust. Revista Italiana di Paleontologia e Stratigrafia: 111 (2): 305–316.
Breistroffer, M. 1947. Notes de nomenclature paléozoologique, I. – Ammonites jurassiques et cretacées. Proces-verbaux mensuels de la Societé Scientifique du Dauphiné 195: 1–5.
Callomon, J.H.C. 1981. Superfamily Haplocerataceae, superfamily Stephanocerataceae, superfamily Persphinctaceae, superfamily Spirocerataceae. In: M.R. House and J.R. Senior (eds.), The Ammonoidea, In: D.T. Donovan, J.H. Callomon and M.K. Howarth. Classification of Jurassic Ammonitina. The Systematic Association, Special Volume 18. 118–131 and 143–155. Academic Press, London.
Cecca, F. and Enay, R. 1991. Les ammonites des zones à semiforme et à fallauxi du Tithonique de l’Ardèche (sud-est de la France). Palaeontographica A 219: 1–87.
Cecca, F., Cresta, S., and Santantonio, M. 1983. Ammoniti del Malm dell’ Appennino marchigiano conservante nel Museo del Servizio Geologico d’Italia. Bolletino del Servizio geologico d’Italia 102 (for 1981): 109–132.
Cecca, F., Cresta, S., Pallini, G., and Santantonio, M. 1984. Biostratigrafia ed ammoniti del Dogger – Malm di Colle Tordina (Monti della Rossa, Appeninno Marchigiano). Bolletino del Servizio geologico d’Italia 104 (for 1983–1984): 177–204.
Collignon, M. 1960. Atlas des fossiles caractéristiques de Madagascar (Ammonites). Fascicule VI (Tithonique). 175 pp. Service Géologique, Ministère des Mines et de l’Energie de Republique Malgache, Tananarive.
Drushchits, V.V. and Kudryavtsev, M.P. (eds.) 1960. Atlas nižnemelovoj fauny Severnogo Kavkaza i Kryma. Ammonity. 396 pp. Vsesoûznyj Naučno-issledovatel’skij Institut Prirodnych Gazov, Moskva.
Enay, R. and Cecca, F. 1986. Structure et évolution des populations tithoniques du genre d’ammonites téthysien Haploceras Zittel, 1868. In: G. Pallini (ed.), Fossili Evoluzione Ambiente. Commemoracione di Rafaele Piccinini. Atti i Convegno Pergola 25–28 ottobre 1984: 37–53. Comitato Centenario Raffaele, Pesaro.
Engeser, T. and Keupp, H. 2002. Phylogeny of the aptychi-possesing Neoammonoidea (Aptychophora nov., Cephalopoda). Lethaia 34: 79–96. Crossref
Ettachfini, M. 2004. Les ammonites néocomiennes dans l’Atlas atlantique (Maroc). Biostratigraphie, paléontologie, paléobiogéographie et paléoécologie. Strata, Actes du Laboratoire de Géologie Sédimentaire et Paléontologie de l’Université Paul-Sabatier Toulouse Série 2, Mémoires 43: 1–225.
Fatmi, A.N. and Zeiss, A. 1999. First Upper Jurassic and Lower Cretaceous (Berriasian) ammonites from the Sembar Formation (Belemnite Shales), Windar Nai, Lasbela – Balochistan, Pakistan. Mémoires of the Geological Survey of Pakistan 19: 1–114.
Favre, E. 1880. Description des couches tithoniques des Alpes Fribourgeoises. Mémoires Suisses de Paléontologie 3: 1–74.
Fontannes, Ch.F. 1879. Description des ammonites des calcaires du Château de Crussol, Ardèche (zones à Oppelia tenuilobata et Waagenia beckeri). 122 pp. Lyon H. Georg, Libraire et Paris F. Savy, Libraire, Paris. Crossref
Főzy, I. 1988. Tithonian ammonites (Oppeliidae, Haploceratidae and Simoceratidae) from the Transdanubian Central Range, Hungary. Annales Universitatis Scientiarum Budapestensis de Rolando Eötvös nominate, Sectio Geologica 28: 43–119.
Főzy, I. 1990. Ammonite succession from three Upper Jurassic sections in the Bakony Mts. (Hungary). Atti del secondo convegno internazionale Fossili, Evoluzione, Ambiente, Pergola 1987: 323–339.
Főzy, I. 1995. Upper Jurassic ammonites from Seno di Guadaloca (Western Sicily). Hantkeniana, Géczy Jubilee Volume 1: 131–143.
Főzy, I. 2017. Oxfordian–Barremian (Late Jurassic–Early Cretaceous) Fauna, Biostratigraphy, Palaeoenvironment and Basin Evolution of the Transdanubian Range, Hungary. 205 pp. GeoLitera, Szeged.
Főzy, I. and Scherzinger, A. 2013. Systematic descriptions of Tithonian ammonites of the Gerecse Mountains. In: I. Főzy (ed.), Late Jurassic–Early Cretaceous Fauna, Biostratigraphy, Facies and Deformation History of the Carbonate Formations in the Gerecse and Pilis Mountains (Transdanubian Range, Hungary), 207–292. Institute of Geosciences, University of Szeged, GeoLitera Publishing House, Szeged.
Főzy, I., Kazmér, M., and Szente, I. 1994. A unique lower Tithonian fauna in the Gerecse Mts, Hungary. Palaeopelagos, Special Publication 1. Proceedings of the 3rd Pergola International Symposium, 155–165.
Frau, C. and Boursicot, P.-Y. 2021. Another lesson from beautiful monsters: the case of “sex reveals” in the Ammonoidea and their significance. BMC Ecology and Evolution 21: 1–13. Crossref
Gemmellaro, G.G. 1876. Studi paleontologici sulla fauna del calcare a Terebratula janitor del Nord di Sicilia. Giornale di Scienze naturali ed economiche 7: 179–185.
Hammer, Ø. and Harper, D.A.T. 2006. Paleontological Data Analysis. 368 pp. Blackwell, Oxford. Crossref
Hoedemaeker, Ph.J., Janssen, N.M.M., Casellato, C.E., Gardin, S., Reháková, D., and Jamrichová, M. 2016. Jurassic/Cretaceous boundary in the Río Argos succession (Caravaca, SE Spain). Revue de Paléobiologie 35 (1): 111–247.
Hohenegger, L. 1861. Die geognostische Verhältnisse der Nordkarpathen in Schlesien und den angrenzenden Theilen von Mähren und Galizien als Erläuterung der geognostischen Karte der Nordkarpathen. 50 pp. Justus Perthes Verlag, Gotha.
Houša, V. 1990. Stratigraphy and calpionellid zonation of the Štramberk Limestone and associated Lower Cretaceous beds. In: G. Pallini (ed.), Atti del secondo convegno internazionale Fossili, Evoluzione, Ambiente, Pergola 25–30 ottobre 1987, 365–370. Comitato centenario Raffaele Piccinini, Pergola.
Houša, V. and Vašíček, Z. 2004. Ammonoidea of the Lower Cretaceous deposits (late Berriasian, Valanginian, early Hauterivian) from Štramberk, Czech Republic. Geolines 18: 7–57.
Hyatt, A. 1900. Cephalopoda. In: K.A. Zittel (ed.), Textbook of Palaeontology, 502–592. Wiley, London.
Khimshiashvili, N.G. 1989. Berriasellidy Kavkaza. Titonskaâ fauna gory Lakorozi-Tau (bassejn r. Bzybi). 86 pp. Akademiâ nauk Gruzinskoj SSR, Institut Paleobiologii im. L.Š. Davitashvili, Tbilisi.
Klein, J., Hoffmann, R., Joly, B., Shigeta Z., and Vašíček, Z. 2009. Lower Cretaceous Ammonites IV – Boreophylloceratoidea, Phylloceratoidea, Lytoceratoidea, Tetragonitoidea, Haploceratoidea including the Upper Cretaceous representatives, Fossilum Catalogus I: Animalia. pars 146, 1–416. Backhuys Publishers, Leiden, Margraf Publishers, Weikersheim.
Korn, D., Ebbighausen, V., Bockwinkel, J., and Klug, C. 2003. The A-mode sutural ontogeny in prolecanitid ammonoids. Palaeontology 46: 1123–1132. Crossref
Książkiewicz, M. 1956. Jura i kreda Bachowic. Rocznik Polskiego Towarzystwa Geologicznego 24 (1954): 119–405.
Kutek, J. and Wierzbowski, A. 1979. Lower to middle Tithonian ammonite succession at Rogoźnik in the Pieniny Klippen Belt. Acta Geologica Polonica 29: 195–205.
Kutek, J. and Wierzbowski, A. 1986. A new account on the Upper Jurassic stratigraphy and ammonites of the Czorsztyn succession, Pieniny Klippen Belt, Poland. Acta Geologica Polonica 36: 289–316.
Kvantaliani, I.V. 1999. Berriasian cephalopod mollusks of Crimea and Caucasus [in Russian]. Trudy, novaă seriă, Akademiă nauk Gruzii, Geologičeskij Institut im. A.I. Dzhanelidze 112: 1–188.
Trudy, novaă seriă, Akademiă nauk Gruzii, Geologicheskii Institut im. A. I. Dzhanelidze
Malinowska, L. (ed.), Barczyk, W., Bielecka, W., Błaszyk, J., Brochwicz–Lewiński, W., Dadlez, R., Dajczak–Calikowska, K., Dembowska, J., Golonka, J. et al. 1980. Budowa geologiczna Polski, tom 3. Atlas skamieniaƚości przewodnich i charakterystycznych. część 2b, Mezozoik: jura, 436–493. Wydawnictwa Geologiczne, Warszawa.
Matyja, B. A. 1986. Developmental polymorphism in Oxfordian ammonites. Acta Geologica Polonica 36: 37–69.
Měchová, L., Vašíček, Z., and Houša, V. 2010. Early Cretaceous ribbed aptychi—a proposal for a new systematic classification. Bulletin of Geosciences 85: 219–274. Crossref
Olóriz Sáez, F. 1978. Kimmeridgense-Tithonico inferior en el Sector Central de las Cordilleras Beticas, Zona Subbetica. Paleontologia. Bioestratigrafia. 758 pp. Tesis doctorales de la Universidad de Granada, Granada.
Oppel, A. 1865. Die tithonische Etage. Zeitschrift der Deutschen geologischen Gesellschaft 1: 535–558.
Parent, H., Scherzinger, H., and Schweigert, G. 2008. Sexual phenomena in Late Jurassic Aspidoceratidae (Ammonoidea). Dimorphic correspondence between Physodoceras hermanni (Berckhemer) and Sutneria subeumela Schairer, and first record of possible hermaphroditism. Palaeodiversity 1: 181–187.
Patrulius, D. and Avram, E. 1976. Les Céphalopodes des couches de Carhaga (Tithonique supérieur – Barrémien inférieur). Mémoires de l’Institut de Géologie et de Géophysique Bucarest 24: 153–201.
Patrulius, D., Avram, E., and Matei, V. 1970. Notă asupra unei faune a Tithonicului inferieur (zona Contiguus) din Carpatii orientali (Valea Doftanei). Studii şi cercetąri de geologie, geofizica, geografie de Bucuresti, seria Geologie 15: 135–148.
Pavia, G, Cresta, S. (coords.), Cecca, F., d´Arpa, C., Galaácz, A., Goy, A., Henriquez, H., Macchioni, F., Martinez, G., Melendez, G., and Oloriz, F. 2002. Revision of Jurassic ammonites of the Gemmellaro collections. Quaderni del Museo Geologico “G.G. Gemmellaro”, Vol. 6, 406 pp. Dipartimento di geologia e geodesia, Università di Palermo, Palermo.
Picha, F.J., Stráník, Z., and Krejčí, O. 2006. Geology and hydrocarbon resources of the Outer Western Carpathians and their foreland: geology and hydrocarbon resources. American Association of Petroleum Geologists, Memoir 84: 49–175. Crossref
Retowski, O. 1893. Die tithonischen Ablagerungen von Theodosia. Ein Beitrag zur Paläontologie der Krim. Bulletin de la Société Imperiale des Naturalistes de Moscou (new series) 7: 206–301.
Rossi, F. 1984. Ammoniti del Kimmeridgiano superiore-Berriasiano inferiore del Passo del Furlo (Appennino Umbro-Marchigiano). Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 23 (3): 75–136.
Sapunov, I.G. 1979. Fossilite na B’lgarija, III. 3. Gorna yurska serija. Ammonoidea. 263 pp. Izdatelstvo na B’lgarskata akademija na naukite, Sofia.
Sarti, C. 1986. Fauna e biostratigrafia del Rosso Ammonitico del Trentino centrale (Kimmeridgiano–Titoniano). Bolletino della Società paleontologica Italiana 23 (3): 473–514.
Sarti, C. 2020. Il Titoniano del Trento Plateau (Alpi Meridionali): faune ad Ammoniti, Stratigrafia e variazioni paleo-ambientali. Studi Trentini di Scienze Naturali 99: 37–314.
Sarti, C. 2024. Il Titoniano del Trento Plateau (Alpi Meridionali). Faune ad ammoniti, stratigrafia e variazioni paleo-ambientali. 291 pp. Associazione Neroniade e del Museo Geo-paleontologico di Piobbico, Trento.
Schweigert, G. 2009. First three dimensionally preserved in situ record of an aptychophoran ammonite jaw apparatus in the Jurassic and discussion of the function of aptychi. Berliner paläobiologische Abhandlungen 10: 321–330.
Shea, E.K. and Vecchione, M. 2002. Quantification of ontogenetic discontinuities in three species of oegopsid squids using model II piecewise linear regression. Marine Biology 140: 971–979. Crossref
Skupien, P. and Smaržová, A. 2011. Palynological and geochemical response to environmental changes in the Lower Cretaceous in the Outer Western Carpathians; a record from the Silesian Unit, Czech Republic. Cretaceous Research 32: 538–551. Crossref
Slavin, V.I. 1953. Tithonian-Valanginian ammonoids of Carpathians [in Russian]. Trudy Instituta geologičeskih nauk, geologičeskaâ seriâ 149: 39–63.
Spath, L.F. 1923. A monograph of the Ammonoidea of the Gault, Part 1. Monographs of the Palaeontographical Society, London 75: 1–72. Crossref
Spath, L.F. 1925. Jurassic Cephalopoda from Madagascar. Bulletins of American Paleontology 11 (44): 1–29.
Špinar Z. (ed.) 1965. Systematická paleontologie bezobratlých. 1045 pp. Academia-Nakladatelství Československé akademie věd, Praha.
Szives, O. and Főzy, I. 2013. Systematic description of Early Cretaceous Ammonoidea of the carbonate formations of the Gerecse Mountains. In: I. Főzy (ed.), Late Jurassic–Early Cretaceous Fauna, Biostratigraphy, Facies and Deformation History in the Gerecse and Pilis Mountains (Transdanubian Range, Hungary), 293–342. Institute of Geosciences, University of Szeged, GeoLitera Publising House, Szeged.
Toucas, A. 1890. Étude sur la fauna des couches tithoniques de l’Ardèche. Bulletin de la Société géologique de France (series 3) 1: 560–629.
Trauth, F. 1927. Aptychenstudien I. Über die Aptychen im Allgemeinen. Annalen des Naturhistorischen Museums in Wien 41: 171–259.
Turculet, I. 1994. Asupra oportnitătii separării de parasubgenuri in cadrul paragenului Lamellaptychus (Cephalopoda, Ammonoidea. Studii şi Cercetări de Geologie, Geofizică, Geografie, Serie Geologica 39: 119–126.
Vaňková, L., Elbra, T., Pruner, P., Vašíček, Z., Skupien, P., Reháková, D., Schnabl, P., Košťák, M., Švábenická, L., Svobodová, A., Bubík, M., Mazuch, M., Čížková, K., and Kdýr, Š. 2019. Integrated stratigraphy and palaeoenvironment of the Berriasian peri-reefal limestones at Štramberk (Outer Western Carpathians, Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology 532: 1–21. Crossref
Vašíček, Z. 1983. Cephalopoden-Fauna aus dem Fundort Roštín bei Kroměříž (Unter-Tithon). Sborník vědeckých prací Vysoké školy báňské v Ostravě, Řada hornicko-geologická 28 (for 1982): 91–114.
Vašíček, Z. and Skupien, P. 2004. Historie geologických a paleontologických výzkumů svrchnojurských a spodnokřídových sedimentů na Štramberku. Sborník vědeckých prací Vysoké školy báňské v Ostravě, Řada hornicko-geologická 50: 83–102.
Vašíček, Z. and Skupien, P. 2005. Doplňky k historii geologických a paleontologických výzkumů na Štramberku. Sborník vědeckých prací Vysoké školy báňské – Technické univerzity Ostrava, Řada hornicko-geologická 51: 1–6.
Vašíček, Z. and Skupien, P. 2013. Early Berriasian ammonites from the Štramberk Limestone in the Kotouč Quarry (Outer Western Carpathians, Czech Republic). Annales Societatis Geologorum Poloniae 83: 329–342.
Vašíček, Z. and Skupien, P. 2014. Recent discoveries of Tithonian ammonites in the Štramberk Limestone (Kotouč Quarry, Outer Western Carpathians). Annales Societatis Geologorum Poloniae 84: 131–141.
Vašíček, Z. and Skupien, P. 2016. Tithonian–early Berriasian perisphinctoid ammonites from the Štramberk Limestone at Kotouč Quarry near Štramberk, Outer Western Carpathians (Czech Republic). Cretaceous Research 64: 12–29. Crossref
Vašíček, Z., Skupien, P., and Jagt, J.W.M. 2018. Current knowledge of ammonite assemblages from the Štramberk Limestone (Tithonian–lower Berriasian) at Kotouč Quarry, Outer Western Carpathians (Czech Republic). Cretaceous Research 90: 185–203. Crossref
Vašíček, Z., Reháková, D., and Skupien, P. 2017. Some perishinctoid ammonites of the Štramberk Limestone and their dating with associated microfossils (Tithonian to lower Berriasian, Outer Western Carpathians, Czech Republic). Geologica Carpathica 68: 583–605. Crossref
Vašíček, Z., Skupien, P., and Jirásek, J. 2013. The northernmost occurrence of the lower Berriasian ammonite Pseudosubplanites grandis (Štramberk Limestone, Czech Republic). Geologica Carpathica 64: 461–466. Crossref
Vígh, G. 1984. Die biostratigraphische Auswertung einiger Ammoniten-Faunen aus dem Tithon des Bakonygebirges sowie aus dem Tithon–Berrias des Gerecsegebirges. Annales Instituti Geologici Publici Hungarici 67: 1–210.
Wiedmann, J. 1966a. Stammesgeschichte und System der posttriadischen Ammonoideen, ein Überblick (1. Teil). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 125: 49–79.
Wiedmann, J. 1966b. Stammesgeschichte und System der posttriadischen Ammonoideen, ein Überblick (2. Teil). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 127: 13–81.
Wiedmann, J. 1968. Evolucion y clasification de los amonites del Cretácico. Boletin de Geologia 24: 23–49.
Wierzbowski, A. 1990. The taxonomy and phylogenetic significance of Early Tithonian ammonites of the genus Parancyloceras Spath from the Pieniny Klippen Belt (Carpathians, Poland. Atti del secondo convegno internazionale Fossili, Evoluzione, Ambienta, Pergola 1987: 479–489.
Wright, C.W., Callomon, J.H., and Howarth, M.K. 1996. Cretaceous Ammonoidea. Treatise on Invertebrate Paleontology. Part L, Mollusca 4 Revised, 362 pp. The Geological Society of America and The University of Kansas Boulder, Colorado and Lawrence, Kansas.
Zeiss, A. 2001. Die Ammonitenfauna der Tithonklippen von Ernstbrunn, Niederösterreich. Neue Denkschriften des Naturhistorischen Museums in Wien 6: 1–115.
Zeiss, A. 2003. The Upper Jurassic of Europe. Its subdivision and correlation. Bulletin of Geological Society of Denmark and Greenland 1: 75–114. Crossref
Zeiss A., Benetti, A., and Pezzoni, N. 1994. A new fauna from the Tithonian (Semiformiceras/Verruciferum Zone) of the Lessinian Alps, Verona Province, Northern Italy. Palaeopelagos, Special Publication 1: 367–381.
Zejszner, L. 1846. Nowe lub niedokładnie opisane gatunki skamieniałości Tatrowych. 32 pp. Poszyt I i II. The Author, Warszawa.
Ziegler, B. 1958. Monographie der Ammonitengattung Glochiceras im epikontinentalen Weißjura Mitteleuropas. Palaeontographica Abteilung A 110 (4–6): 93–164.
Zittel, K.A. 1868. Die Cephalopoden der Stramberger Schichten. Paläontologische Mittheilungen aus dem Museum des Königlich-Bayerischen Staates 2: 33–118.
Zittel, K.A. 1869. Geologische Beobachtungen aus dem Central Appenninen. Geognostisch-paläontologische Beiträge 2: 88–178.
Zittel, K.A. 1870. Die Fauna der aeltern cephalopodenfuehrenden Tithonbildungen. Palaeontographica (Supplement 1): 1–192.
Zittel, K.A. von 1884. Cephalopoda. In: K.A. v. Zittel (ed.), Handbuch der Paläontologie. I. Abtheilung, Paläozoologie. II. Band. Mollusca und Arthropoda, 329–522. Verlag R. Oldenbourg, München.
Acta Palaeontol. Pol. 70 (3): 421–441, 2025
https://doi.org/10.4202/app.01230.2024