
Lower jaw morphology of the last surviving tritylodontid Fossiomanus sinensis from the Early Cretaceous Jehol Biota, Liaoning Province, China
HAIBING WANG, JUNFANG XIE, ZHIQIANG YU, LUN HAI, ZIFAN ZHU, WENJIE ZHENG, and YUANQING WANG
Tritylodontids are close relatives of mammals with specialized teeth adapted for herbivory. Despite their diversification during the Jurassic, the fossil record of this clade suggests they declined significantly in the Cretaceous when they are mainly represented by fragmentary dental remains. The exception is the Early Cretaceous taxon Fossiomanus sinensis Mao et al., 2021. Here we describe a new mandible of this species from the same locality as the holotype specimen. The new specimen provides more complete information on mandible shape and tooth morphology, filling a knowledge gap for this iconic Cretaceous tritylodontid, given that cranial morphology in the holotype of F. sinensis remains insufficiently studied. Additionally, the fossil record of F. sinensis represents the youngest known tritylodontid (~119 Ma, Aptian) and the latest non-mammaliaform cynodont, shedding light on the evolutionary history of early mammalian relatives.
Introduction
Tritylodontids are a group of mammaliamorphs that were predominantly specialized herbivores (Kalthoff et al. 2019), characterized by multi-cusped cheek teeth (postcanines). It is generally accepted that this clade is closely related to mammaliaforms due to dental morphological similarities (complexity in cusp pattern and occlusal configuration), although their exact phylogenetic position, whether as the sister clade to mammaliaforms or another related group, remains debated (Rowe 1988; Angielczyk and Kammerer 2018; Abdala 2021). Tritylodontids were among the most derived and diverse cynodonts, with a wide geographic distribution and an extensive temporal range, from the Late Triassic to the Early Cretaceous (Kühne 1956; Fedak et al. 2015; Matsuoka et al. 2016; Velazco et al. 2017; Angielczyk and Kammerer 2018; Mao et al. 2021). The unequivocal record of this clade in the Triassic is sparse, e.g., Oligokyphus discovered in the Upper Triassic (Rhaetian) Scots Bay Member of the McCoy Brook Formation at Wasson Bluff in the Fundy rift basin in Nova Scotia and NewBrunswick, Canada (Fedak et al. 2015). Tritylodonts flourished during the Early Jurassic, but experienced a sharp decline by the end of the Jurassic (Kühne 1956; Sues 1985b, 1986b; Matsuoka and Setoguchi 2000; Matsuoka et al. 2016). The Early Cretaceous tritylodontids have been reported from Siberia (Tatarinov and Matchenko 1999), central Japan (Matsuoka et al. 2016), and northeastern China (Mao et al. 2021). Most of these fossil remains are only dentition, with the exception of Fossiomanus sinensis Mao et al., 2021, which is represented by a complete skeleton (Mao et al. 2021). However, in the holotype of F. sinensis, the crania and mandible are poorly preserved, obscuring the details of the morphology of this iconic tritylodontid taxon. Here, we report a new mandible of this taxon discovered in Liaoning Province from the Jiufotang Formation, the late stage of the Early Cretaceous Jehol Biota (Zhou et al. 2021; Yu et al. 2023).
Institutional abbreviations.—CVEB, Centre for Vertebrate Evolutionary Biology, Yunnan University, Kunming, China; JZPM, Jinzhou Paleontological Museum; ZMNH, Zhejiang Museum of Natural History, Hangzhou, Zhejiang Province, China.
Material and methods
The specimen, ZMNH M31795, was discovered in the same locality as the holotype: in the Jiufotang Formation at the Lamadong site, Jianchang County, Chaoyang City, Liaoning Province, China. It is currently housed in the Zhejiang Museum of Natural History, Hangzhou, Zhejiang Province. High-resolution micro-computed tomography (micro-CT) was conducted at the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing using a GE V|Tome|xm dual tube. The mandible was scanned at a resolution of 15.734 μm (160 kV, 110 μA). The software package VGStudio Max 3.0 was used for virtual segmentation and visualization. The 3D files for the lower jaws are deposited in figshare (https://figshare.com/s/bff16d85764872847655). The cusp terminology of lower cheek teeth used in this study follows Watabe et al. (2007), in which cusps are labelled as the buccal (b) and lingual (l) rows and numbered mesiodistally as cusp 1 and cusp 2 in each cusp row. The measurements of the lower jaws and teeth were taken digitally in ImageJ.
Systematic palaeontology
Genus Fossiomanus Mao et al., 2021
Type locality: Lamadong, Jianchang County, Liaoning province, China; Aptian, Lower Cretaceous.
Fossiomanus sinensis Mao et al., 2021
Fig. 1.
Type material: ZGY0052, a nearly complete skeleton.
Type locality: Lamadong, Jianchang County, Liaoning province, China.
Type horizon: Aptian, Lower Cretaceous Jiufotang Formation.
Material.—Holotype and the new specimen (ZMNH M31795), a partial mandible discovered from the Jiufotang Formation at Lamadong site.
Description (mandible only).—The mandible, ZMNH M31795, is mainly represented by the dentaries, and the tooth row is badly damaged except for the lower incisors and one displaced left postcanine (Fig. 1A1, A2). The dentaries are slightly compressed dorsoventrally. The length of the mandible is 51.3 mm. The mandibular symphysis is transversely broad and not fused with a longitudinal suture in the anterior portion between the left and right dentaries. The suture exhibits an interlocking connection on the posteroventral portion of the symphysis, likely suggesting a partial fusion between the dentaries. On the ventral side of the symphysis, a zig-zag suture is present (Fig. 1A4). The posterior margin of the symphyseal facets is anteriorly concave and extends posteriorly to the level of the anterior-most part of the tooth row. The ventral margin of the dentaries is rounded and broad in the symphysis region. In lateral view, a mental foramen is visible on the lateral surface of the left dentary dorsal to the anterior part of the tooth row (Fig. 1A3).
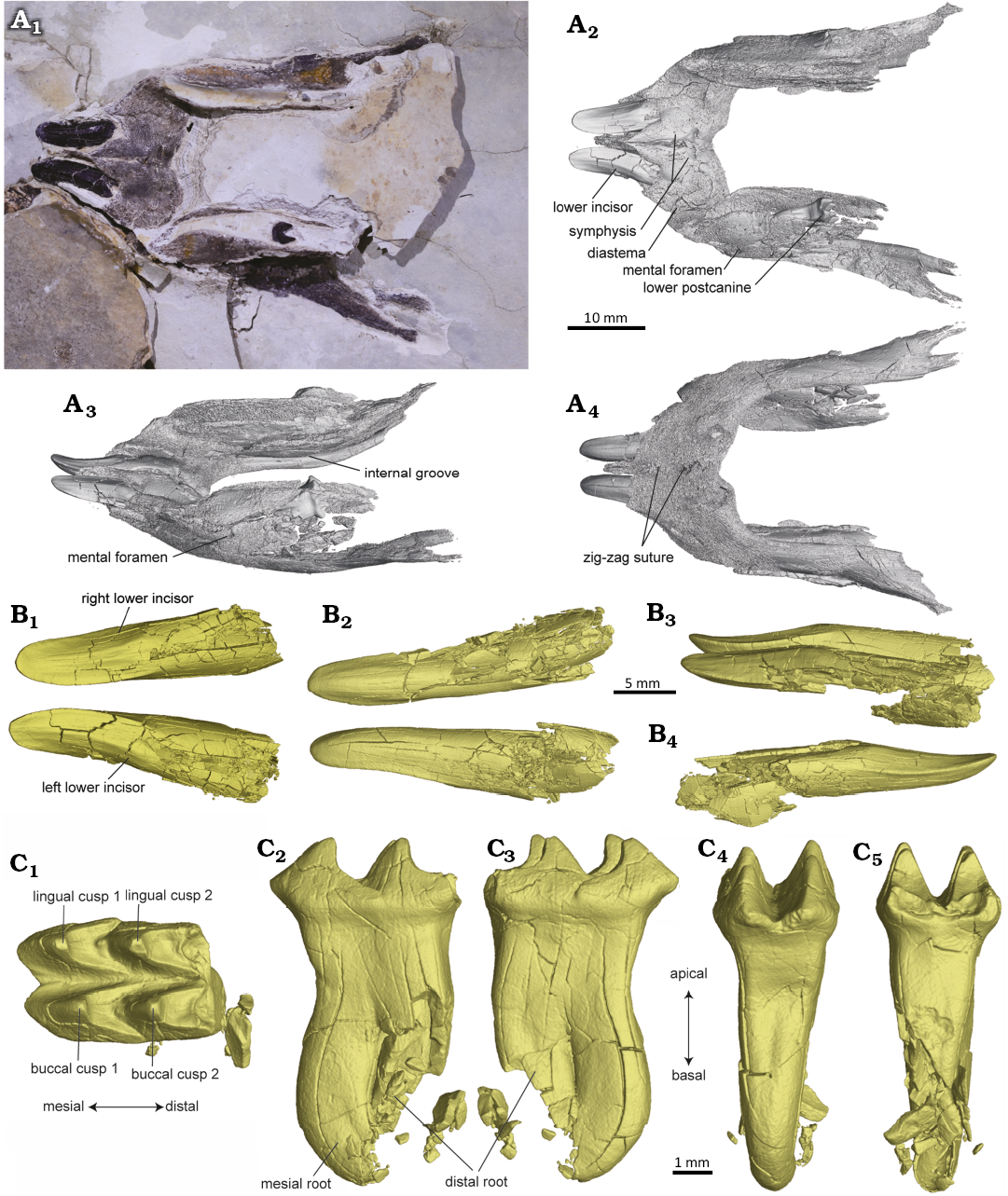
Fig. 1. The mandible of tritylodontid mammal Fossiomanus sinensis Mao et al., 2021, from Aptian, Lower Cretaceous Jiufotang Formation at the Lamadong site, with two lower incisors preserved in their alveoli and one lower postcanine displaced (ZMNH M31795) from the Jiufotang Formation at the Lamadong site. A. The new specimen preserved in the slab in dorsal view (A1), segmented mandible based on CT data in dorsal (A2), dorsolateral (A3), and ventral (A4) views. B. Lower incisors in occlusal (B1), ventral (B2), left lateral (B3), and right lateral (B4) views. C. Lower left postcanine in occlusal (C1), buccal (C2), lingual (C3), mesial (C4), and distal (C5) views.
Only one lower incisor is present in each dentary of the mandible, characterized by an enlarged and procumbent crown (Fig. 1A2, B). No additional small or developing lower incisors are present near this tooth locus on each side. The apical portion of the lower incisors is linguolabially compressed, featuring a longitudinal ridge along the midline of the crown, with sharp medial and lateral margins (Fig. 1B4). The basal portion of the lower incisors is more robust and rounded, and the roots appear to be open as indicated by the incomplete outline observed in the distal region. The lower canine is absent, as in all tritylodontids, and a distinct diastema separates the lower incisor from the postcanine row on each side. The diastema has a ridged dorsal margin. The postcanine rows run relatively straight along the length of each dentary with minimal twisting between the anterior and posterior teeth. A distinct internal groove is present on the medial surface of each dentary (Fig. 1A3).
One left lower postcanine is preserved, though slightly displaced (Fig. 1A2, C). The tooth measures 2.5 mm in length, 1.6 mm in width, and 4.4 mm in height (including both the crown and root). The postcanine tooth exhibits a cusp formula of 2:2, consisting of two longitudinal rows of equal-sized cusps (Fig. 1C1). The cusps are distally concave and separated by a deep, open groove. The postcanine tooth has two roots, mesial and distal, which are fused in their proximal half and only separate in their distal half (Fig. 1C2, C3). The tip of the mesial root curves mesially, while the tip of the root shows damage to the distal region.
Remarks.—We assign this specimen to the recently reported species Fossiomanus sinensis based on the similarities of dental morphology and shared locality of discovery. The curvature, fusion of roots, and the cusp formula of lower postcanines closely resembles that of the holotype of F. sinensis, JZMP 2107500093. The preserved lower postcanine of ZMNH M31795 (2.5 mm in length) is much smaller than that of the holotype (approximately 4.5 mm in length, as measured from figures) (Mao et al. 2021: extended fig. 3E). The replacement pattern of postcanines in tritylodontids is featured by teeth moving anteriorly, with oldest teeth lost at the anterior end of the postcanine row and newly erupted teeth being added at the posterior end of the postcanine row, so-called the “conveyor belt” system (Kühne 1956; Matsuoka and Setoguchi 2000; Panciroli et al. 2017). Given that intraspecific variation in postcanine size is distinct in tritylodontids due to ontogenetic differences (e.g., Panciroli et al. 2017), the size difference of lower postcanines between these two specimens suggests that ZMNH M31795 likely represents a younger individual of F. sinensis. The lower postcanine in ZMNH M31795 is likely an anteriorly implanted, soon-to-be lost tooth. Fossiomanus sinensis with the preserved dentary length of 51.3 mm in ZMNH M31795, represents a medium-sized tritylodontid. This size of the mandible within the clade ranges from small taxa like Lufengia (CVEB 12001, dentary length about 23 mm, cranium length estimated about 31 mm) (Liu et al. 2022) to large-sized taxa like Kayentatherium (skull length over 220 mm) (Sues 1986b). The mandibular symphysis in Fossiomanus is transversely wider than in most Jurassic taxa, such as Oligokyphus, Dinnebitodon, and Kayentatherium (Kühne 1956; Sues 1985a, 1986a, b). A zig-zag suture of the mandibular symphysis was noted in the holotype of F. sinensis by Mao et al. (2021), whereas the new specimen exhibits a longitudinally oriented suture in the anterior portion of the symphysis. In addition, the posteroventral portion of the symphysis tends to be partially fused with an interlocking suture in dorsal view. Recent studies in fossorial rodents suggest that compared to the cranium, mandible morphology is more adaptable to functional selective pressures (McIntosh and Cox 2019). The fossorial adaptation to scratch-digging of Fossiomanus, as reflected by its postcranial morphology (Mao et al. 2021), might also be enhanced by the shovel-like lower incisor and the broad, partially fused mandibular symphysis, which likely contributes to the stability of the mandible and the efficiency of penetrating and soil-grasping of the lower incisors, a similar mechanism as shown in fossorial rodents (Kley and Kearney 2007; Van Wassenbergh et al. 2017; Mclntosh and Cox 2019).
Only one pair of lower incisors is present in Fossiomanus (Fig. 1A2, B), similar to Bienotherium (Young 1947; Cui and Sun 1987), and Kayentatherium (Sues 1986b), but different from Bienotherodes (Sun 1984; Cui and Sun 1987; Watabe et al. 2007) and Lufengia (two pair of lower incisors; Liu et al. 2022), and Oligokyphus (three pair of lower incisors; Kühne 1956). In lower incisors, the tooth crown is more laterally compressed, and the root is open posteriorly in Fossiomanus, different from that of the Early Cretaceous Montirictus from central Japan (Matsuoka et al. 2016). Tritylodontids exhibit a pattern of horizontal tooth replacement, with the addition of newly erupting postcanines at the distal end of the tooth row (Kühne 1956; Matsuoka and Setoguchi 2000; Jasinoski and Chinsamy 2012). Differences in tooth morphology are an important aspect in evolutionary morphology of tritylodontids, and distinct variations have been observed in height, number, fusion, and curvature of postcanine roots (e.g., Cui and Sun 1987; Averianov et al. 2017; Panciroli et al. 2017; Liu et al. 2022). The curvature of the roots in the lower postcanines increases anteriorly, whether the roots of lower postcanines are fused or not. In Fossiomanus, the double-rooted and partially fused condition of the lower postcanine roots differs from the long, separate, and distinctly curved condition in Bienotherium (Cui and Sun 1987) and from the slighter shorter and less curved condition in Lufengia (Liu et al. 2022). The fusion of the proximal root in the lower postcanine of Fossiomanus is intermediate between that seen in the Middle Jurassic Stereognathus and Early Cretaceous Montirictus (Matsuoka et al. 2016; Averianov et al. 2017). The fusion of lower postcanine roots tends to be more developed in some specimens of the Middle Jurassic derived Stereognathus sp. (with nearly fully fused roots; Averianov et al. 2017) and Bienotheroides (single-rooted with complete fusion; Cui and Sun 1987).
Stratigraphic and geographic range.—Aptian, Lower Cretaceous Jiufotang Formation in Liaoning Province, China.
Concluding remarks
Fossiomanus represents the latest survival tritylodontid taxon and the youngest record of a non-mammaliaform cynodont. Among the few Early Cretaceous tritylodontid localities, the Ilek Formation in western and central Siberia is currently considered to date from Barremian to Aptian stages (Tatarinov and Matchenko 1999; O’Connor et al. 2014). The Kuwajima Formation in central Japan is estimated between 130.7 Ma and 121.2 Ma by zircon U-Pb ages, indicating that the Kuwajima Formation formed between late Hauterivian and early Aptian. The lower part of the Kuwajima Formation yields a zircon U-Pb age of 130.7 ± 0.8 Ma (Matsuoka and Setoguchi 2000; Matsuoka et al. 2016), while a LA-ICP-MS zircon U-Pb date of 121.2 ± 1.1 Ma was obtained from the tuff of the overlying Akaiwa Formation (Matsuoka et al. 2016; Sakai et al. 2019), providing a minimum age estimate for the Kuwajima Formation (Matsuoka et al. 2016; Sakai et al. 2019). Recently, a series of zircon U-Pb ages have been reported for the Jiufotang Formation, which represents the later stage of the Early Cretaceous Jehol Biota, from two main sections: the Sanmendian section and the Xiaotaizi section (Yu et al. 2021, 2023) using secondary ion mass spectroscopy (SIMS). One tuff from the Sanmendian section, nearly the lowest part of the Jiufotang Formation, constrains the lower limit of the depositional age to an (SIMS) age of 122.0 ± 0.9 Ma. Additional age constraints are also available, including U-Pb ages of 118.9 ± 0.8 Ma, 118.8 ± 0.6 Ma, and 118.6 ± 1.1 Ma for the Jiufotang Formation at the Xiaotaizi section (Yu et al. 2021). Additionally, an age constraint of 120.3 ± 0.7 is reported for the Shangheshou section of the Jiufotang Formation based on 40Ar/39Ar dating (He et al. 2004). These results suggest that the depositional age of Jiufotang Formation is approx. 123–119 Ma (He et al. 2004; Yu et al. 2021, 2023). Fossiomanus, discovered at the Lamadong site, can be estimated in age by the geological dating in the Xiaotaizi section, i.e., approx. 119 Ma (Aptian) (Gale et al. 2020; Yu et al. 2021). The young age of Fossiomanus makes it a significant taxon for understanding the evolutionary history of tritylodontids, and of cynodonts as a whole. Tritylodontids is one of the most successful clades in the evolutionary history of cynodonts, which can be attributed to their wide dietary range, efficient mastication, fossorial adaptation, and reproduction and growth pattern (Hoffmann and Rowe 2018; Kalthoff et al. 2019; Mao et al. 2021). Current evidence indicates a dramatic decline of this clade in the Early Cretaceous, coinciding with the rise of multituberculates, another group of specialized herbivores with more derived life history strategies (Hoffmann and Rowe 2018; Wang et al. 2019; Weaver et al. 2022). This suggests that the diversification of multituberculates, along with other small vertebrates occupying similar ecological niches, may have intensified competition for resources and ecological space, ultimately leading to the extinction of tritylodontids.
Acknowledgements.—We thank Yemao Hou and Pengfei Yin (both Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China) for help with computed tomography scans. This work was supported by the National Natural Science Foundation of China (42288201, 42272017) and the Youth Innovation Promotion Association, CAS (No. 2021068). We thank Elsa Panciroli (National Museums Scotland, Edynburg, UK) and Julia A Schultz (University of Bonn, Germany) for their constructive suggestions on the manuscript.
Editor: Eli Amson.
References
Abdala, F. 2021. Permo-Jurassic cynodonts: the early Road to mammalness. In: D. Alderton and E. Scott (eds.), Encyclopedia of Geology, 2nd Edition. Vol. 3, 206–226. Academic Press, London. Crossref
Angielczyk, K.D. and Kammerer, C.F. 2018. Non-mammalian synapsids: the deep roots of the mammalian family tree. In: F. Zachos, and R.J. Asher (eds.), Handbook of Zoology: Mammalia, 117–198. Walter de Gruyter, Berlin. Crossref
Averianov, A.O., Martin, T., Lopatin, A.V., Schultz, J.A., Skutschas, P.P., Schellhorn, R., and Krasnolutskii, S.A. 2017. A tritylodontid synapsid from the Middle Jurassic of Siberia and the taxonomy of derived tritylodontids. Journal of Vertebrate Paleontology 37 (5): e1363767. Crossref
Cui, G.H. and Sun, A.L. 1987. Postcanine root system in tritylodonts. Vertbreta PalAsiatica 25 (4): 245–259. Crossref
Fedak, T.J., Sues, H.D., and Olsen, P.E. 2015. First record of the tritylodontid cynodont and cynodont postcranial bones from the McCoy Brook Formation of Nova Scotia, Canada. Canadian Journal of Earth Sciences 52 (4): 244–249. Crossref
Gale, A.S., Mutterlose, J., Batenburg, S., Gradstein, F.M., Agterberg, F.P., Ogg, J.G., and Petrizzo, M.R. 2020. The Cretaceous period. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, G.M. Ogg (eds.), Geologic Time Scale 2020, 1023–1086. Elsevier, Amsterdam. Crossref
Hoffman, E.A. and Rowe, T.B. 2018. Jurassic stem-mammal perinates and the origin of mammalian reproduction and growth. Nature 561: 104–108. Crossref
He, H.Y., Wang, X.L., Zhou, Z.H., Wang, F., Boven, A., Shi, G.H., and Zhu, R.X. 2004. Timing of the Jiufotang Formation (Jehol Group) in Liaoning, northeastern China, and its implications. Geophysical Research Letters 31 (12): 1–4. Crossref
Jasinoski, S.C. and Chinsamy, A. 2012. Mandibular histology and growth of the nonmammaliaform cynodont Tritylodon. Journal of Anatomy 220: 564–579. Crossref
Kalthoff, D.C., Schulz-Kornas, E., Corfe, I., Martin, T., McLoughlin, S.Schultz, J.A. 2019. Complementary approaches to tooth wear analysis in Tritylodontidae (Synapsida, Mammaliamorpha) reveal a generalist diet. PLOS One 14 (7): e0220188. Crossref
Kley, N. Kearney, M. 2007. Adaptations for digging and burrowing. In: B.K. Hall (ed.), Fins into Limbs: Evolution, Development, and Transformation, 284–309. University of Chicago Press, Chicago.
Kühne, W.G. 1956. The Liassic Therapsid Oligokyphus. 149 pp. British Museum (Natural History), London.
Liu, J., Wang, L., Beatty, B.L., Zhang, G., Wang, T., and Bi, S. 2022. New material (Tritylodontidae, Mammaliamorpha) from the lower Jurassic of Lufeng and its implication on the taxonomy of Lufengia. Historical Biology 35: 1605–1615. Crossref
Mao, F., Zhang, C., Liu, C., and Meng, J. 2021. Fossoriality and evolutionary development in two Cretaceous mammaliamorphs. Nature 592: 577–582. Crossref
Matsuoka, H. and Setoguchi, T. 2000. Significance of Chinese tritylodonts (Synapsida, Cynodontia) for the systematic study of Japanese materials from the Lower Cretaceous Kuwajima Formation, Tetori Group of Shiramine, Ishikawa, Japan. Asian Paleoprimatology 1: 161–176.
Matsuoka, H., Kusuhashi, N., and Corfe, I.J. 2016. A new Early Cretaceous tritylodontid (Synapsida, Cynodontia, Mammaliamorpha) from the Kuwajima Formation (Tetori Group) of Central Japan. Journal of Vertebrate Paleontology 36 (4): e1112289. Crossref
McIntosh, A.F., Cox, P.G. 2019. The impact of digging on the evolution of the rodent mandible. Journal of Morphology 280: 176–183. Crossref
O’Connor, J.K., Averianov, A.O., and Zelenkov, N.V. 2014. A confuciusornithiform (Aves, Pygostylia)-like tarsometatarsus from the Early Cretaceous of Siberia and a discussion of the evolution of avian hind limb musculature. Journal of Vertebrate Paleontology 34: 647–656. Crossref
Panciroli, E., Walsh, S., Fraser, N.C., Brusatte, S.L., and Corfe, I. 2017. A reassessment of the postcanine dentition and systematics of the tritylodontid Stereognathus (Cynodontia, Tritylodontidae, Mammaliamorpha), from the Middle Jurassic of the United Kingdom. Journal of Vertebrate Paleontology 37: e1351448. Crossref
Rowe, T. 1988. Definition, diagnosis and origin of Mammalia. Journal of Vertebrate Paleontology 8: 241–264. Crossref
Sun, A. 1984. Skull morphology of the tritylodont genus Bienotheroides of Sichuan. Scientia Sinica 27: 970–984.
Sakai, Y., Tsutsumi, Y., Kusuhashi, N., Sonoda, T., Horie, K., and Matsuoka, A. 2019. Zircon LA-ICP-MS U-Pb age of a tuff from the Akaiwa Formation of the Tetori Group in the Shiramine area, Ishikawa Prefecture, central Japan. The Journal of the Geological Society of Japan 125: 255–260. Crossref
Sues, H.-D. 1985a. First record of the tritylodontid Oligokyphus (Synapsida) from the Lower Jurassic of Western North America. Journal of Vertebrate Paleontology 5 (4): 328–335. Crossref
Sues, H.-D. 1985b. The relationships of the Tritylodontidae (Synapsida). Zoological Journal of the Linnean Society 85: 205–217. Crossref
Sues, H.-D. 1986a. Dinnebitodon amarali, a new tritylodontid (Synapsida) from the Lower Jurassic of western North America. Journal of Paleontology 60: 758–762. Crossref
Sues, H.-D. 1986b. The skull and dentition of two tritylodontid synapsids from the Lower Jurassic of western North America. Bulletin of the Museum of Comparative Zoology 151: 217–268.
Tatarinov, L.P. and Matchenko, E.N. 1999. A find of an aberrant tritylodont (Reptilia, Cynodontia) in the lower Cretaceous of the Kemerovo region. Paleontological Journal 4 (4): 85–92.
Van Wassenbergh, S., Heindryckx, S., and Adriaens, D. 2017. Kinematics of chisel-tooth digging by African mole-rats. Journal of Experimental Biology 220: 4479–4485. Crossref
Velazco, P.M., Buczek, A.J., and Novacek, M.J. 2017. Two new tritylodontids (Synapsida, Cynodontia, Mammaliamorpha) from the Upper Jurassic, southwestern Mongolia. American Museum Novitates 3874: 1–36. Crossref
Wang, H., Meng, J., and Wang, Y.Q. 2019. Cretaceous fossil reveals a new pattern in mammalian middle ear evolution. Nature 576: 102–105. Crossref
Watabe, M., Tsubamoto, T., and Tsogtbaatar, K. 2007. A new tritylodontid synapsid from Mongolia. Acta Palaeontologica Polonica 52: 263–274.
Weaver, L.N., Fulghum, H.Z., Grossnickle, D.M., Brightly, W.H., Kulik, Z.T., Wilson Mantilla, G.P., and Whitney, M.R. 2022. Multituberculate mammals show evidence of a life history strategy similar to that of placentals, not marsupials. American Naturalist 200: 383–400. Crossref
Young, C.C. 1947. Mammal-like reptiles from Lufeng, Yunnan, China. Proceedings of the Zoological Society of London 117: 537–597. Crossref
Yu, Z.Q., Wang, H.B., Zhang, C., Dong, L.P., Huyskens, M.H., Cui, Z., Cary, P., Di, Y., Amelin, Y., Li, G., Li, Q.L., Xia, X.P., Deng, C.L., Wang, Y.Q., He, H.Y., and Yin, Q.-Z. 2023. Temporal framework for the Yanliao Biota and timing of the origin of crown mammals. Earth and Planetary Science Letters 617 (1): 118246. Crossref
Yu, Z.Q., Wang, M., Li, Y.J., Deng, C.L., and He, H.Y. 2021. New geochronological constraints for the Lower Cretaceous Jiufotang Formation in Jianchang Basin, NE China, and their implications for the late Jehol Biota. Palaeogeography Palaeoclimatology Palaeoecology 583: 110657. Crossref
Zhou, Z., Meng, Q., Zhu, R., and Wang, M. 2021. Spatiotemporal evolution of the Jehol Biota: Responses to the North China craton destruction in the Early Cretaceous. Proceedings of the National Academy of Sciences of the United States of America 118 (34): e2107859118. Crossref
Haibing Wang [wanghaibing@ivpp.a.c.cn, ORCID: https://orcid.org/0000-0001-6811-7262 ], Zifan Zhu [zhuzifan24@mails.ucas.ac.cn; ORCID: https://orcid.org/0009-0008-6938-4774 ], and Yuanqing Wang [wangyuanqing@ivpp.ac.cn, ORCID: https://orcid.org/0000-0002-7467-6586 ], Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, 100044, China. College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing, 100049, China.
Wenjie Zheng [zhengwj@zmnh.com, ORCID: https://orcid.org/0000-0002-3610-806X ] and Junfang Xie [xiejf@zmnh.com; ORCID: https://orcid.org/0009-0001-7629-9972 ], Zhejiang Museum of Natural History, Hangzhou, Zhejiang Province, 310014, China.
Zhiqiang Yu [zhiqiangyu@mail.iggcas.ac.cn; ORCID: https://orcid.org/0000-0001-5486-8372 ], Key Laboratory of Planetary Science and Frontier Technology, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing 100029, China.
Lun Hai [hailun@ivpp.ac.cn; ORCID: https://orcid.org/0009-0007-7610-4323 ], Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, 100044, China. School of Earth Sciences and Resources, China University of Geosciences, Beijing, 100083, China.
Received 12 December 2024, accepted 6 March 2025, published online 28 May 2025.
Copyright © 2025 H. Wang et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 70 (2): 285–289, 2025
https://doi.org/10.4202/app.01232.2024