
The first fossil representative of the dragonfly family Synthemistidae
ANDRE NEL, VALERIE NGO MULLER, ROMAIN GARROUSTE, and MATHIEU BODERAU
Nel, A., Muller, V.N., Garrouste, R., and Boderau, M. 2025. The first fossil representative of the dragonfly family Synthemistidae. Acta Palaeontologica Polonica 70 (2): 253–258.
Gallosynthemis bechlyi gen. et sp. nov., described from the Paleocene maar of Menat (Puy-de-Dôme, France) is the first-ever fossil record of the dragonfly family Synthemistidae. It shows the main synapomorphies of the family, viz. hind wing with 3–5 costal braces (complete antenodal crossveins) alternating with non-aligned antenodals; presence of crossveins in median area and between CuP and PsA; absence of postsubnodal crossveins below first postnodal crossveins; in hind wing, a very large and elongate anal loop without a well-defined mid-rib; and CuAa without posterior branches. It belongs to the stem group of this family and is attributed to its own subfamily Gallosynthemistinae subfam. nov., characterized by a very long stem of hind wing cubitus anterior vein (autapomorphy), plus some symplesiomorphies such as sectors of arculus separated at their bases. While extant Synthemistidae sensu Bechly (2016) are Australasian, the present new taxon demonstrates that the family may have been much more widespread during the Paleocene. These dragonflies probably became extinct in the Northern Hemisphere in relation with the Cenozoic dramatic episodes of cooling.
Key words: Insecta, Odonata, Anisoptera, Libelluloidea, new taxa, Paleocene, Menat, France.
André Nel [anel@mnhn.fr; ORCID: https://orcid.org/0000-0002-4241-7651 ], Valérie Ngô Muller [valerie.ngo-muller@ u-paris.fr; https://orcid.org/0000-0002-9456-6667 ], Romain Garrouste [romain.garrouste@mnhn.fr; ORCID: https://orcid.org/0000-0002-0880-5781 ], Mathieu Boderau [mathieuboderau@gmail.com; ORCID: https://orcid.org/0000-0001-7088-9560 ], Institut de Systématique, Évolution, Biodiversité (UMR 7205) Muséum national d’Histoire naturelle, CNRS, Sorbonne Université, EPHE-PSL, Université des Antilles, F-75005 Paris, France.
Valérie Ngô Muller UFR Sciences du Vivant, Faculté des Sciences, Université Paris-Cité, Paris, France.
Received 9 January 2025, accepted 9 April 2025, published online 9 May 2025.
Copyright © 2025 A. Nel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Synthemistidae Tillyard, 1917 (sensu Bechly 1996, 2016) is an extant dragonfly family nowadays restricted to Australia, Tasmania, New Caledonia, Fiji, Solomon, and Papua New Guinea (Carle 1995; Theischinger and Richards 2013). Carle et al. (2015) proposed an expanded concept of the “GSI/Synthemistidae” to include the Gomphomacromiidae Tillyard & Fraser, 1940, Pseudocorduliidae Lohmann, 1996b, and several other taxa considered in separate families by Bechly (2016). More precisely, Carle et al. (2015: 295) proposed a concept of a “further expanded Synthemistidae which includes all of Fraser’s Corduliidae except Macromiinae and Corduliinae ” (i.e., the GSI of Ware et al. 2007). GSI is an abbreviation proposed by Ware et al. (2007: 298) for the group “Gomphomacromiinae + Synthemistinae + Cordulephyinae + Idionychinae”. Davis et al. (2011) recovered the Synthemistidae as the sister lineage to all other Libelluloidea. Carle et al. (2015) considered the “GSI/Synthemistidae” as a much wider set of taxa, sister group to the (Macromiidae + (Corduliidae + Libellulidae)), while Bechly (2016) considered the Synthemistidae sensu stricto as sister group to the Neolamellida Bechly, 1996 (a group that would comprise the same last families plus those included into the “GSI/Synthemistidae” sensu Carle et al. (2015) that do not fall in the Synthemistidae sensu Bechly (2016). Fleck and El Adouzi (2013) separated the Synthemistidae into Synthemistinae (= Synthemistidae sensu Bechly 1996, 2016) and Gomphomacromiinae. As the crown Cavilabiata (Libelluloidea sensu lato) are dated at at least 168 Ma, and the stem Libellulidae are dated at 93.9–89.8 Ma (Kohli et al. 2016), the most recent common ancestor of Synthemistidae sensu stricto is certainly Mesozoic too. To date, no fossil Synthemistidae sensu Bechly (2016) have ever been recorded.
The fact that the extant Synthemistidae sensu Bechly (1996, 2016) are Australasian does not imply that the family could not be found as fossils in other regions. The termite Mastotermitidae is a spectacular example of a strictly Australasian family that is now known from fossils on all continents, except Antarctica (Jouault et al. 2022).
Here we describe the first-ever fossil representative of the Synthemistidae sensu Bechly (1996, 2016), which was collected from the Paleocene of Menat (France). This first occurrence may provide a time point for calibration in phylogenetic analyses.
The odonatan fauna from Menat is rather diverse (Garrouste et al. 2017; Nel and Jouault 2022; Doriath-Döhler et al. 2023), with at least representatives of the families Epigomphidae Fraser, 1934, Dysagrionidae Cockerell, 1908 and superfamilies Lestinoidea Calvert, 1901, and Amphipterygoidea Selys -Longshamps, 1853. Several undescribed species of Odonata are also recorded by very fragmentary wings.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank:urn:lsid:zoobank.org:pub: urn: 5A082E47-B4FF-47CB-9EFD-81281ED5F92F.
Institutional abbreviations.—MNT, Musée de Paléontologie de Menat¸ Menat, Puy-de-Dôme, France.
Other abbreviations.—AA/P, anal anterior/posterior; a.l., anal loop; Arc, arculus; Ax, primary antenodal crossvein; C, costa; CuA/P, cubitus anterior/posterior; CuAa/b anterior/posterior branch of CuA; GSI, Gomphomacromiinae + Synthemistinae + Cordulephyinae + Idionychinae; h, hypertriangle; IR, intercalary longitudinal veins between main branches of RP; m, median space; MA/P, median anterior/posterior; N, nodus; O, oblique crossvein; PsA, Pseudo-Anal vein; RA/P radius anterior/posterior; ScP, subcostal posterior; sm, submedian space; Sn, subnodus; t, discoidal triangle.
Material and methods
The lacustrine Lagerstätte of Menat (Puy-de-Dôme, France) is a rather small maar (about 1 km diameter), filled with spongodiatomites. It is among the oldest Paleocene outcrops providing insect fossils. According to palynology, mammalian biostratigraphy, and radiometric K/Ar analyses its age was estimated to ca. 56 Ma (Kedves and Russel 1982) vs macrofloral datings indicating the age of 60–61 Ma (Wappler et al. 2009), corresponding to the Selandian–Thanetian (Uhl et al. 2024). It is well known for its very rich and diverse floras (e.g., Laurent 1912; Piton 1940) and faunas (vertebrates and insects). Piton (1940) was the first to extensively study this Lagerstätte. Since then, several works have been done on the entomofauna, mainly during the last twenty years by the group from MNHN (see list in Nel 2022). The insect fauna is abundant and diverse, mainly comprising beetles, cockroaches, and bugs.
The composition of faunal and floral remains suggests that this lake was surrounded by a forest and that the palaeoenvironment was warm and humid (Wedmann et al. 2018).
The new discovery made during field works in 2019 was possible thanks to a careful examination of all the rock fragments during field and laboratory research. Insects are notoriously hard to detect on the dark brown rocks from this locality. The fossil studied herein is a dragonfly specimen that may have been regurgitated by a vertebrate predator as suggested by folded and crumpled wing remains and two abdominal segments. Nevertheless, a reconstruction of the basal half of the hind wing was possible.
The fossil was studied using stereomicroscopes (Olympus SCX9 and a Nikon SMZ1500), drawn with a camera lucida under a Nikon SMZ1500. It was photographed with a Canon MP-E 65 mm f/2.8 1–5× macro photo lens attached to a Canon EOS 6D camera, mounted on a semi-automatic Cognises Rail macro Stack Shot controlled by Helicon Remote software. The reconstruction was created through drawings of both part and counterpart. Photographs were focus-merged using Helicon Focus 6.7 software and were exported in TIF format. Final images, and drawings were realized using Pixelmator Pro 3.6.13 Archipelago software.
We follow the wing venation nomenclature of Riek and Kukalová-Peck (1984), modified by Nel et al. (1993), Bechly (1996), and Jacquelin et al. (2018). We follow Bechly (2016)’s classification of the Synthemistidae (= Synthemistinae sensu Fleck and El Adouzi 2013).
Systematic palaeontology
Class Insecta Linnaeus, 1758
Order Odonata Fabricius, 1793
Suborder Anisoptera Selys in Selys-Longschamps & Hagen, 1854
Family Synthemistidae Tillyard, 1917
(sensu Bechly 1996, 2016)
Subfamily Gallosynthemistinae nov.
Zoobank LSID: urn:lsid:zoobank.org:act:158E1DA8D0E3-4692-808B- 2B4BB0BC118D.
Type genus: Gallosynthemis gen. nov.; see below.
Diagnosis.—As for the type species of the type genus, by monotypy.
Genus Gallosynthemis nov.
Zoobank LSID: urn:lsid:zoobank.org:act:2261A631-35B1-4329-9AAE -DFBBBD4CD9AA.
Type species: Gallosynthemis bechlyi sp. nov., see below; monotypic.
Etymology: Named after Gallia, Latin name for France and the genus name Synthemis. Gender feminine.
Diagnosis.—As for the type species, by monotypy.
Gallosynthemis bechlyi sp. nov.
Fig. 1.
Zoobank LSID: urn:lsid:zoobank.org:act:BAC7293E-5A46-4C6C-954 E-084393E9749F.
Etymology: After our friend and colleague Günter Bechly (1963–2025), who passed away in a dramatic accident at the beginning of January 2025, in recognition to his impressive and crucial contribution to palaeoentomology, especially on dragonflies.
Holotype: MNT Nel 2013 (part and counterpart).
Type locality: Menat, Puy-de-Dôme, France.
Type horizon: Selandian–Thanetian, Paleocene.
Material.—Holotype only.
Diagnosis.—Hind wing characters only. A very long stem of CuA, covering five large cells between CuA and MP (autapomorphy); discoidal cell elongate, not equilateral; sectors of arculus not stalked; anal loop transversely elongate with a strongly zigzagged midrib.
Description.—Only basal half of hind wing can be described. Distance from base to nodus 10.4 mm, from base to arculus 3.2 mm, wing 6.4 mm wide; two primary antenodal crossveins visible, Ax1 situated 0.9 mm basal to arculus, Ax2 1.1 mm distal to it, a third secondary antenodal complete, other secondary antenodals of first row not aligned with those of second row; median area with two distal crossveins; submedian area with one or two crossveins between CuP and PsA (pseudo anal vein); sectors of arculus touching only at their bases; hypertriangle free; discoidal triangle free, elongate, not equilateral; two crossveins between MAa and RP basal of RP3/4; antesubnodal crossveins not discernable; base of RP2 aligned with subnodus; oblique vein O close to base of RP2; two postnodal crossveins discernable but no corresponding postsubnodals; postdiscoidal area with one row of cells just distal to discoidal triangle and two rows more distally; area between MP and CuA/CuAa with one row of cells basally, two rows distally and more distally only one row; stem of CuA very long; CuAb short fused with AA and closing posteriorly anal loop; CuAa smoothly curved and parallel to MP, without well-defined posterior branch; cubito-anal area rather narrow, with three rows of cells between CuAa and posterior margin of wing; anal loop obliquely elongate, with a poorly defined zigzagged midrib; at most two rows of cells between anal loop and posterior margin of wing.
Stratigraphic and geographic range.—Selandian–Thanetian, Paleocene, Menat, Puy-de-Dôme, France.
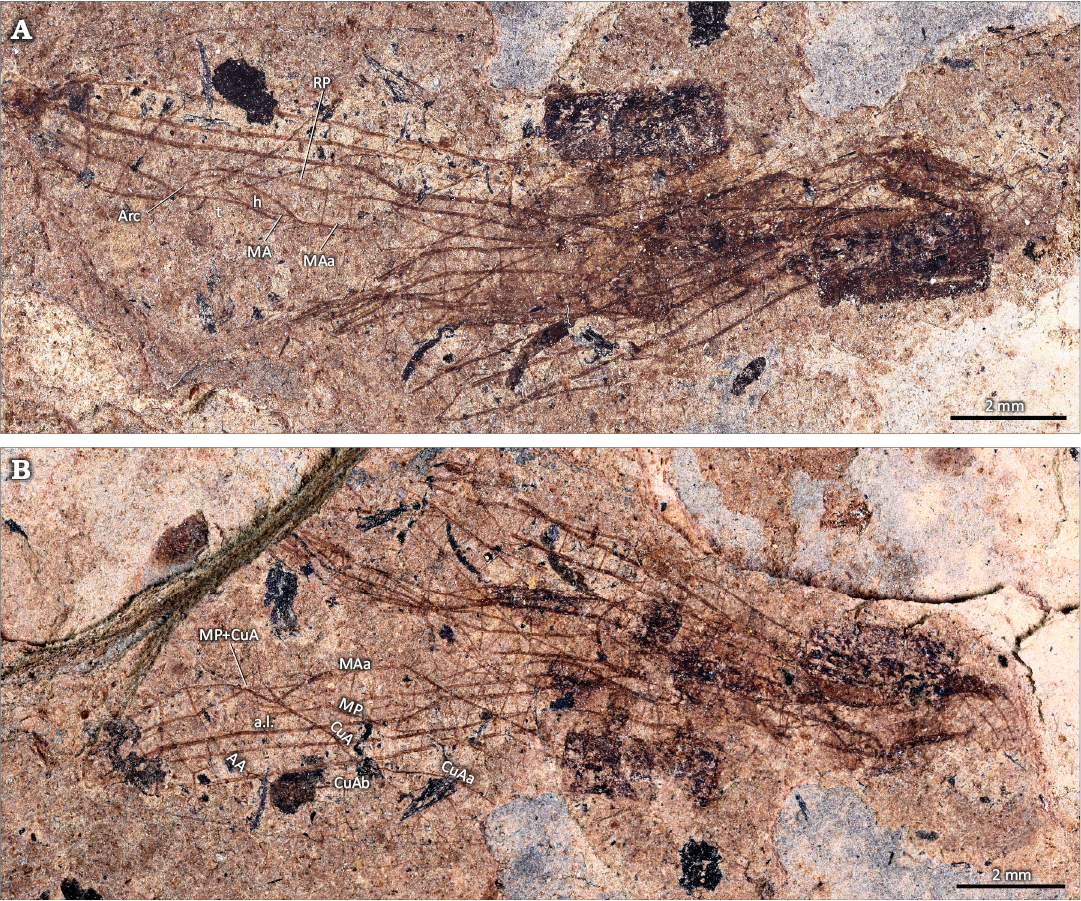
Fig. 1. Synthemistid dragonfly Gallosynthemis bechlyi gen. et sp. nov, holotype MNT Nel 2013 from Selandian–Thanetian, Paleocene, Menat, Puy-de-Dôme, France. Photograph of part (A) and counterpart (B). Abbreviations: AA, anal anterior; a.l., anal loop; Arc, arculus; CuA, cubitus anterior; CuAa/b, anterior/posterior branch of CuA; h, hypertriangle; MA/P, median anterior/posterior; RP radius posterior; t, discoidal triangle.
Discussion
In the hind wing, the very large and elongate anal loop without a well-defined mid-rib, the CuAa without posterior branches, the presence of crossveins in the median area and between CuP and PsA (putative synapomorphies), and the absence of postsubnodal crossveins below the first postnodal crossveins are characters of the family Synthemistidae sensu Bechly (2016). Chlorogomphidae also displays crossveins in the median and submedian spaces and a rather similar anal loop, but these strongly differ from the new fossil by the presence of a strong posterior branch of CuAa (Fraser 1957; Bechly and Ueda 2002). Extant Chlorogomphidae also have postsubnodal crossveins just distad of the subnodus, absent in the Cretaceous Araripechlorogomphidae Bechly & Ueda, 2002. The Araripechlorogomphidae have no median and submedian supplementary crossveins.
Carle (1995: 408) proposed the character state “hindwing with 3–5 costal braces [antenodal crossveins] alternating with non-aligned antenodals” as synapomorphy of the Synthemistidae, which is present in Gallosynthemis bechlyi gen. et sp. nov.. But he also proposed the character “sectors of arculus stalked” as apomorphy, which is not the case for Gallosynthemis bechlyi. Nevertheless, the character state “unstalked sectors of arculus” is likely plesiomorphic, and does not exclude a position of the new fossil G. bechlyi in the stem group Synthemistidae sensu Bechly (2016).
The valvulidan families Gomphomacromiidae and Pseudocorduliidae have an anal loop rather similar to that of the new fossil, but they differ in having a more posteriorly curved CuAa, with a rudimentary secondary branch (vs. more longitudinal and with no branch at all), and the antenodal crossveins all complete, which may be a putative apomorphy of all the Valvulida Bechly, 1996 (Ellenrieder and Garrison 2005; Bechly 2016). The Pseudocorduliidae also have no crossvein in the median area.
All extant genera of Synthemistidae sensu Bechly (2016) (Palaeosynthemis Förster, 1903, Eusynthemis Förster, 1903, Choristhemis Tillyard, 1910, Austrosynthemis Carle, 1995, Tonyosynthemis Theischinger, 1998, Synthemis Selys, 1870, Parasynthemis Carle, 1995, Calesynthemis Carle, 1995), except Synthemiopsis gomphomacromioides Tillyard, 1910, differ from the new fossil genus Gallosynthemis in having the hind wing stem of CuA much shorter, and the discoidal cell nearly equilateral (Förster 1903; Tillyard 1910; Fraser 1957; Moulds 1985; Carle 1995; Theischinger 1998; Theischinger and Richards 2013, 2016; Fleck 2024).
In Synthemiopsis Tillyard, 1910 (unique representative of the Synthemiopsinae sensu Carle 1995), the stem of CuA is distinctly longer than in the other Synthemistidae sensu Bechly (2016), even if shorter than in the new fossil genus Gallosynthemis, the anal loop is obliquely elongate as in Gallosynthemis, with a rudimentary strongly zigzagged mid-rib as in Gallosynthemis, and the hind wing discoidal cell is elongate, not equilateral as in Gallosynthemis. Nevertheless, it differs from the new fossil in the sectors of arculus with a long stalk, vs. separated at base.
Gallosynthemis bechlyi gen. et sp. nov. is thus attributed to the stem group of the Synthemistidae sensu Bechly (2016). It can be separated from the crown group in the very long stem of CuA (autapomorphy), discoidal cell elongate and sectors of arculus not stalked (putative symplesiomorphies).
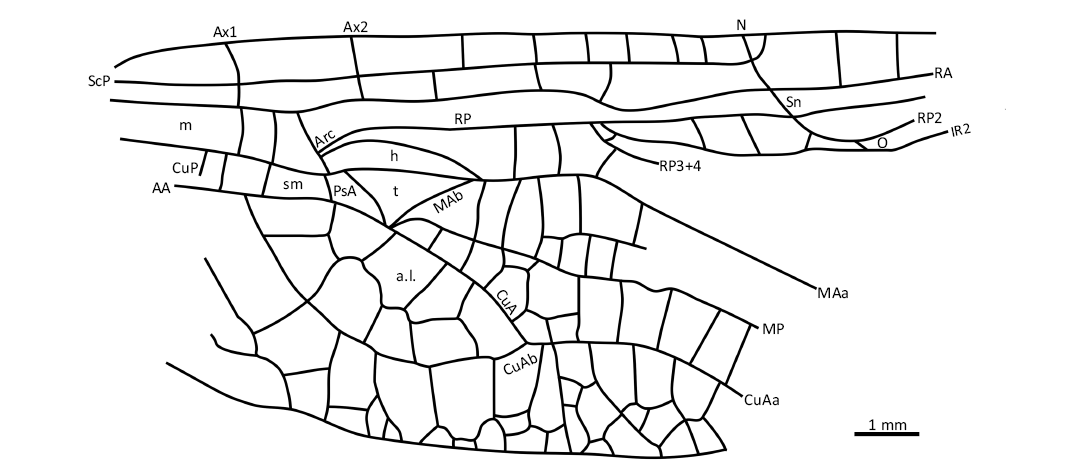
Fig. 2. Synthemistid dragonfly Gallosynthemis bechlyi gen. et sp. nov, holotype MNT Nel 2013 from Selandian–Thanetian, Paleocene, Menat, Puy-de-Dôme, France. Hind wing venation reconstruction. Arrow indicates an oblique vein. Abbreviations: AA, anal anterior; a.l., anal loop; Arc, arculus; Ax, primary antenodal crossvein; CuA/P, cubitus anterior/posterior; CuAa/b anterior/posterior branch of CuA; h, hypertriangle; IR, intercalary longitudinal veins between main branches of RP; m, median space; MA/P, median anterior/posterior; N, nodus; O, oblique crossvein; PsA, pseudo-anal vein; RA/P, radius anterior/posterior; ScP, subcostal posterior; sm, submedian space; Sn, subnodus; t, discoidal triangle.
Conclusions
Gallosynthemis bechlyi gen. et sp. nov., first fossil record of the dragonfly family Synthemistidae, is described from the Paleocene maar of Menat (Puy-de-Dôme, France). It belongs to the stem group Synthemistidae and is attributed to its own subfamily Gallosynthemistinae subfam. nov., characterized by a very long stem of hind wing cubitus anterior vein (autapomorphy). This finding is the first occurrence of a stem group Synthemistidae sensu Bechly (2016) (= Synthemistinae sensu Fleck & El Adouzi, 2013), and provides a key calibration point for the family Synthemistidae sensu Carle et al. (2015), because it belongs to the crown group of this clade.
While the extant Synthemistidae sensu stricto are Australasian, the description of the new species demonstrates that this family may have been much more widespread during the Paleocene. The fact that the Gomphomacromiidae (a part of the Synthemistidae sensu lato) are distributed in South America and Australia also supports a wider distribution of the Synthemistidae in the deep past compared to the present. This situation can be likened to that of the past distribution of the Mastotermitidae (Isoptera), nowadays strictly Australasian, and that were globally distributed during the Cretaceous–Paleogene. The extant Synthemistidae larvae are known to occur in lotic environments (Fleck 2005), permanent small streams (Fleck and El Adouzi 2013), river springs, or on seepages (Theischinger and Hawking 2000; Theischinger 2003). The presence of fishes in the paleolake of Menat indicates that it could have been linked to at least a river and likely springs.
Acknowledgments
We sincerely thank Mario Schädel (University of Tübingen, Germany) and an anonymous referee for their useful comments on the first version of the paper. We had the pleasure to receive generous assistance from Bernard Duverger (President of the Communauté de Communes of Menat region), Clotilde Berger-Pompili and Mathilde Leygnac (directors of the EHPAD du pays de Menat), for their kind help and authorizations to collect fossil insects in the small but rich outcrop near the village of Menat, Puy-de-Dôme, France. We are thankful towards Antoine Mantilleri and Christophe Rivier (MNHN, Paris) for their kind help to make the photographs and for providing access to the Entomology collection photography equipment.
Editor: Krzysztof Hryniewicz.
References
Bechly, G. 1996. Morphologische Untersuchungen am Flügelgeäder der rezenten Libellen und deren Stammgruppenvertreter (Insecta; Pterygota; Odonata), unter besonderer Berücksichtigung der Phylogenetischen Systematik und des Grundplanes der Odonata. Petalura Special Volume 2: 1–402.
Bechly, G. 2016. Phylogenetic Systematics of Odonata [available online, https://bechly.lima-city.de/phylosys.htm]
Bechly, G., and Ueda, K. 2002. The first fossil record and first New World record for the dragonfly clade Chlorogomphida (Insecta: Odonata: Anisoptera: Araripechlorogomphidae n. fam.) from the Crato Limestone (Lower Cretaceous, Brazil). Stuttgarter Beiträge zur Naturkunde Serie B 328: 1–11.
Calvert, P.P. 1901. Odonata. In: A.E. Eaton (ed.), Biologia Centrali-Americana: Zoology, Botany and Archaeology. Insecta. Neuroptera, 17–410. Horman-Fisher, Maud.
Carle, F.L. 1995. Evolution, taxonomy, and biogeography of ancient Gondwanian libelluloides, with comments on anisopteroid evolution and phylogenetic systematics (Anisoptera: Libelluloidea). Odonatologica 24: 383–424.
Carle, F.L., Karl M. Kjer, K.M., and May, M.L. 2015. A molecular phylogeny and classification of Anisoptera (Odonata). Arthropod Systematics & Phylogeny 73: 281–301. Crossref
Cockerell, T.D.A. 1908. Fossil insects from Florissant, Colorado. Bulletin of the American Museum of Natural History 24, 59–69.
Davis, R.B., Nicholson, D.B., Saunders, E.L.R., and Mayhew, P.J. 2011. Fossil gaps inferred from phylogenies alter the apparent nature of diversification in dragonflies and their relatives. BMC Evolutionary Biology 11: 252–261. Crossref
Doriath-Döhler, A., Hervet, S., and Béthoux, O. 2023. Palaeophylolestes distinctus n. gen., n. sp., a new malachite damselfly (Odonata: Zygoptera: Synlestidae) from the Paleocene Menat locality (France). Geodiversitas 45 (14): 401–407. Crossref
Ellenrieder, N. von, and Garrison, R.W. 2005. A synopsis of the South American genus Gomphomacromia (Odonata: Gomphomacromiinae). International Journal of Odonatology 8: 81–96. Crossref
Fabricius, J.C. 1793. Entomologia systematica emendata et aucta, secundum classes, ordines, genera, species, adjectis synonymis, locis, observationibus, descriptionibus. 519 pp. C.G. Proft, Hafniae [= Copenhagen]. Crossref
Fleck, G. 2005. Contribution à la connaissance des Odonates de Nouvelle-Calédonie: notes sur la larve supposée de Synthemis ariadnes Lieftinck, 1975 (Odonata, Anisoptera, Synthemistidae). Bulletin de la Société Entomologique de France 110: 165–166. Crossref
Fleck, G. 2024. A remarkable new synthemistid from New Caledonia (Odonata: Anisoptera: Synthemistidae s. str.). Taxonomic and phylogenetic note on New Caledonian Synthemistidae and erection of a new genus. Zootaxa 5403: 320–330. Crossref
Fleck, G. and El Adouzi, M. 2013. The larva of the genus Palaeosynthemis Förster, 1903 (Odonata: Anisoptera: Synthemistidae) and a generic key to the larvae of non-New Caledonian Synthemistidae. Zootaxa 3619: 589–894. Crossref
Förster, F. 1903. Odonaten aus Neu-Guinea. 3. Annales Musei Nationalis Hungarici 1: 509–554.
Fraser, F.C. 1934. Odonata. In: J. Stephenson (ed.), The Fauna of British India Including Ceylon and Burma, Vol. 2, 1–398. Taylor and Francis, London.
Fraser, F.C. 1957. A reclassification of the order Odonata, based on some new interpretations of the venation of the dragonfly wing. Handbook of the Royal Zoological Society of New South Wales 12: 1–133.
Garrouste, R., Wedmann, S., Pouillon, J.-M., and Nel, A. 2017. The oldest ‘amphipterygid’ damselfly of tropical affinities in the Paleocene of Menat (Zygoptera: Eucaloptera). Historical Biology 29 (6): 818–821. Crossref
Jacquelin, L., Desutter-Grandcolas, L., Chintauan-Marquier, I., Boistel, R., Zheng, D., Prokop, J., and Nel, A. 2018. New insights on basivenal sclerites using 3D tools and homology of wing veins in Odonatoptera (Insecta). Scientific Reports 8 (238): 1–7. Crossref
Jouault, C., Engel, M.S., Legendre, F., Huang, D., Grandcolas, P., and Nel, A. 2022. Incrementing and clarifying the diversity and early evolution of termites (Blattodea: Isoptera). Zoological Journal of the Linnean Society 196: 608–629. Crossref
Kedves, M. and Russell, D.E. 1982. Palynology of the Thanetian layers of Menat. The geology of the Menat Basin, France. Palaeontographica B 182: 87–150.
Kohli, M.K., Ware, J.L., and Bechly, G. 2016. How to date a dragonfly: Fossil calibrations for odonates. Palaeontologia Electronica 19.1.1FC: 1–14. Crossref
Laurent, L. 1912. Flore fossile des schistes de Menat (Puy-de- Dôme). Annales du Musée d’Histoire Naturelle de Marseille 14: 1–240.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Lohmann, H. 1996a. Das phylogenetische System der Anisoptera (Odonata) (first part). Entomologische Zeitschrift 106: 209–252.
Lohmann, H. 1996b. Das phylogenetische System der Anisoptera (Odonata) (second part). Entomologische Zeitschrift 106: 253–296.
Lohmann, H. 1996c. Das phylogenetische System der Anisoptera (Odonata) (third part). Entomologische Zeitschrift 106: 360–367.
Moulds, M.S. 1985. A new species of Choristhemis Tillyard (Odonata Synthemistidae). Journal of the Australian Entomological Society 24: 113–116. Crossref
Nel, A. 2022. The oldest fossil of the sawfly subfamily Nematinae (Hymenoptera: Tenthredinidae) from the Paleocene of Menat (France). Palaeoentomology 5: 289–293. Crossref
Nel, A. and Jouault, C. 2022. The odonatan insects from the Paleocene of Menat, central France. Acta Palaeontologica Polonica 67: 631–648. Crossref
Nel, A., Martínez-Delclòs, X., Paicheler, J.-C., and Henrotay, M. 1993. Les “Anisozygoptera” fossiles. Phylogénie et classification (Odonata). Martinia Numéro Hors Série 3: 1–311.
Piton, L. 1940. Paléontologie du gisement éocène de Menat (Puyde- Dôme), flore et faune. Mémoire de la Société d’Histoire Naturelle d’Auvergne 1: 1–303.
Riek, E.F. and Kukalová-Peck, J. 1984. A new interpretation of dragonfly wing venation based upon Early Carboniferous fossils from Argentina (Insecta: Odonatoidea) and basic characters states in pterygote wings. Canadian Journal of Zoology 62: 1150–1166. Crossref
Selys-Longchamps, E. de 1870. Annonce du Synopsis des Corduliines. Comptes-Rendus de la Société Entomologique de Belgique 14: iv–vii. Crossref
Selys-Longchamps, E. de 1853. Synopsis des Caloptérygines. Bruxelles. (preprinted before the “original” in Bulletin de l’Académie Belge 20, Annexe: 1–73.) [also as 1854 Bulletin de l’Académie Belge 20 Annexe]
Sélys-Longchamps, E. de and Hagen, H. 1854. Synopsis des Gomphines. Bulletin de l’Académie Royale des sciences et belles-lettres de Bruxelles 21 (2): 23–114.
Theischinger, G. 1998. Tonyosynthemis, a new dragonfly genus from Australia (Insecta: Odonata: Synthemistidae). Linzer Biologische Beiträge 30 (1): 139–142.
Theischinger, G. 2003. The larva of Choristhemis olivei (Tillyard) (Odonata: Synthemistidae). Linzer Biologische Beiträge 35 (1): 657–660.
Theischinger, G. and Hawking, J.H. 2000. The larva of Eusynthemis ursula Theischinger (Odonata: Synthemistidae). Linzer Biologische Beiträge 32 (1): 247–251.
Theischinger, G. and Richards, S.J. 2013. Palaeosynthemis elegans spec. nov., a new dragonfly from Papua New Guinea (Anisoptera: Synthemistidae). Odonatologica 42: 63–66.
Theischinger, G. and Richards, S.J. 2016. Palaeosynthemis opaca sp. nov., a new dragonfly from Papua New Guinea (Anisoptera: Synthemistidae). IDF-Report 99: 1–8.
Tillyard, R.J. 1910. Monograph of the genus Synthemis. Proceedings of the Linnean Society of New South Wales 35: 312–377. Crossref
Tillyard, R.J. 1917. The Biology of Dragonflies (Odonata or Paraneuroptera). xii + 396 pp. Cambridge University Press, London. Crossref
Tillyard, R.J. and Fraser, F.C. 1940. A reclassification of the order Odonata based on some new interpretations of the venation of the dragonfly wing. Part 3. The Australian Zoologist, Sydney 9: 359–396.
Uhl, D., Wuttke, M., Smith, K.T., Wedmann, S., and Lehmann, T. 2024. Pre-Quaternary maar lakes/volcanogenic lakes as Konservat Lagerstätten—Messel and beyond. Palaeobiodiversity and Palaeoenvironments 104: 753–761. Crossref
Wappler, T., Currano, E.D., Wilf, P., Rust, J., and Labandeira, C.C. 2009. No post-Cretaceous ecosystem depression in European forests? Rich insect-feeding damage on diverse middle Palaeocene plants, Menat, France. Proceedings of the Royal Society B 276: 4271–4277. Crossref
Ware, J.L., May, M., and Kjer, K. 2007. Phylogeny of the higher Libelluloidea (Anisoptera: Odonata): An exploration of the most speciose superfamily of dragonflies. Molecular Phylogenetic & Evolution 45: 289–310. Crossref
Wedmann, S., Uhl, D., Lehman, T., Garrouste, R., Nel, A., Gomez, B., Smith, K., and Schaal, S.F.K. 2018. The Konservat-Lagerstätte Menat (Paleocene; France)—an overview and new insights. Geologica Acta 16: 1–31.
Acta Palaeontol. Pol. 70 (2): 253–258, 2025
https://doi.org/10.4202/app.01236.2025