
Dercetid fishes from the Maastrichtian type area (Upper Cretaceous) of Belgium and the Netherlands
JELLE J.A. HEERE, JONATHAN J.W. WALLAARD, MAARTEN DE RIJKE, and JOHN W.M. JAGT
Heere, J.J.A., Wallaard, J.J.W., De Rijke, M., and Jagt, J.W.M. 2025. Dercetid fishes from the Maastrichtian type area (Upper Cretaceous) of Belgium and the Netherlands. Acta Palaeontologica Polonica 70 (4): 661–697.
The family Dercetidae comprises a diverse array of marine aulopiform actinopterygians. Until now, merely seven specimens, assigned to five species, were recorded from the Maastrichtian type area. Here, around two dozen dercetid fossils, originating from a number of quarries and outcrops in north-eastern Belgium and the south-eastern Netherlands are described and illustrated. This material comprises two new genera, Beukidercetis gen. nov. and Petrodercetis gen. nov., and four new species: Caudadercetis taverni sp. nov., Beukidercetis lissus gen. et sp. nov., Cyranichthys sideralis sp. nov., and Petrodercetis bidirectus gen. et sp. nov. Newly collected specimens of Cyranichthys jagti, Dercetis triqueter and Ophidercetis italiensis are also added, because these extend their respective stratigraphical ranges in the study area. Moreover, material here assigned to Dercetoides venator and a specimen resembling Cyranichthys ornatissimus markedly expand their geographical and stratigraphical ranges. In general, the present material reflects an underappreciated diversity of dercetids in the Maastrichtian type area, even though that they fossilize well compared to other late Maastrichtian actinopterygians from the study area. This high degree of preservation may be related to a benthic lifestyle, or to their slender, scute-covered bodies that may have been less vulnerable to disruption than those of other actinopterygians.
Key words: Actinopterygii, Aulopiformes, Enchodontoidei, Dercetidae, scutes, Maastrichtian, Cretaceous, Belgium, the Netherlands.
Jelle J.A. Heere [jelle.heere@ziggo.nl; ORCID: https://orcid.org/0000-0002-7615-1716], Leiden University, P.O. Box 9500, 2300 RA Leiden, the Netherlands, and Het Nationaal Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, the Netherlands.
Jonathan J.W. Wallaard [curator@oertijdmuseum.nl;ORCID: https://orcid.org/0000-0002-7752-1598] and Maarten de Rijke [maarten@oertijdmuseum.nl; ORCID: https://orcid.org/0000-0003-4652-3782], Het Nationaal Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, the Netherlands.
John W.M. Jagt [john.jagt@maastricht.nl; ORCID: https://orcid.org/0000-0001-6216-1991], Natuurhistorisch Museum Maastricht, De Bosquetplein 6-7, 6211 KJ Maastricht, the Netherlands.
Received 12 March 2025, accepted 30 July 2025, published online 1 December 2025.
Copyright © 2025 J.J.A. Heere et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Vertebrate assemblages that existed in the shallow, warm sea that covered the present-day Maastricht area in the Netherlands during the latest Cretaceous comprised a fair range of small- to medium-sized cartilaginous fishes (sharks, rays, skates, sawfish, and chimaeras), and actinopterygians (Herman 1977; Duffin and Reynders 1995; Friedman 2012). Bony fish material is predominantly known from the upper Gulpen Formation (Lanaye Member) and the entire Maastricht and Kunrade formations, all dated as late Maastrichtian (Vellekoop et al. 2022). The latest methods to assess fish diversity in the study area during the latest Cretaceous involved silicified otoliths (Schwarzhans and Jagt 2021).
Although bony fish fossils are not rare in the Maastrichtian type area (Fig. 1), their record is confined mostly to isolated teeth, skull bones (occasionally associated, yet displaced), vertebrae, scales, and scutes. Particularly in the lower portion of the Maastricht Formation, the ichnofossil (burrow) Lepidenteron lewesiensis (Mantell, 1822), which contains numerous bony fish remains, including, albeit rarely, those of dercetids. More or less complete bodies or associated remains of skull, fins and body outside burrows, as the examples described herein, are very rare (Friedman 2012). Dercetidae are a Late Cretaceous family of bony fish (Taverne and Goolaerts 2015), sometimes referred to as “needle fish”, although that vernacular name is also used for the extant, unrelated beloniform family Belonidae. Dercetidae are characterised by having a long, pointed rostrum (i.e., an extension of the upper and lower jaws), one or two pairs of transverse processes on each abdominal vertebral centrum, and an eel-shaped body covered with cordiform or tripartite scutes (Silva and Gallo 2011).
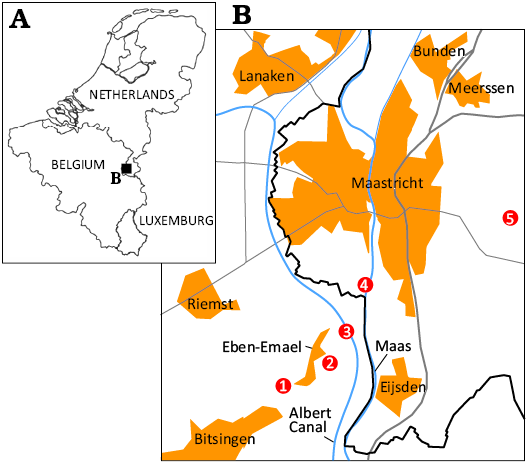
Fig. 1. A. Sketch map of the Netherlands and Belgium with study area indicated. B. Map of the Maastrichtian type area in north-east Belgium and the south-east Netherlands, with indications of quarries at which specimens were collected. 1, CBR-Romontbos quarry (Eben-Emael, Bassenge, province of Liège); 2, Marnebel quarry (Eben-Emael, Bassenge, province of Liège); 3, Albert Canal, between Kanne and Vroenhoven (province of Limburg); 4, former ENCI-HeidelbergCement Group quarry (Sint-Pietersberg, Maastricht); 5, former ‘t Rooth quarry (Bemelen).
Until the publications by Lambers (1998) and Friedman (2012), dercetids were unknown from upper Maastrichtian strata in southern Limburg and Liège (north-east Belgium), although there was a single record (under the name of Leptotrachelus sp., later placed in the genus Dercetis by Siegfried 1966) from the lower Campanian Vaals Formation at Aachen-Linzenhäuschen (Germany) by Albers and Weiler (1964). More recently, Taverne and Goolaerts (2015) recorded four genera and four species of dercetid, two of them new, based on incomplete, but associated remains. Finally, Wallaard et al. (2019) recorded a large-bodied new species, attributed therein to the genus Pelargorhynchus (Von der Marck, 1858), but potentially meriting the erection of a genus if more material were found. Here, we describe 27 specimens of Dercetidae from the Maastrichtian type area, re-attribute Pelargorhynchus grandis to a genus new to science, add two new genera in total, and add four new species to the known diversity of Dercetidae.
Institutional abbreviations.—MAB, Het Nationaal Oertijdmuseum, Boxtel, the Netherlands; NHMM, Natuurhistorisch Museum Maastricht, Maastricht, the Netherlands.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in Zoobank LSID: urn:lsid:zoobank.org:pub:D3502BEA-18DD-41EB-B785-5A0AED336E43.
Geological setting
On micro- and macrofossil evidence, the entire Maastricht Formation (Fig. 2) has been dated as late Maastrichtian, with the exception of the highest subunit (IVf-7) of the Meerssen Member, between the Berg en Terblijt and Vroenhoven horizons, which is of earliest Danian age (Schiøler et al. 1997; Vellekoop et al. 2020). Principal index macrofossils of correlative value include various heteromorph and non-heteromorph ammonites, belemnitellid coleoids and “tegulated” inoceramid bivalves (Keutgen et al. 2017; Jagt and Jagt-Yazykova 2018, 2019). These allow correlations with both Boreal and Tethyan sections in northern/north-eastern and southern Europe, respectively. Some crinoid and cirripede taxa hint at links with the Gulf Coast of North America.
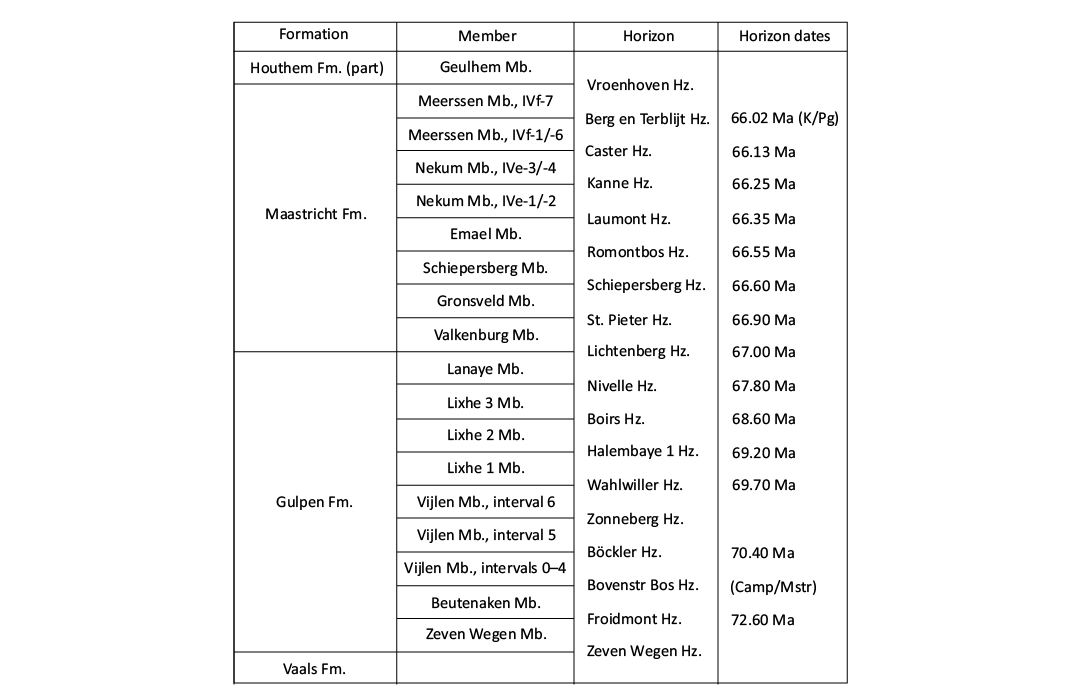
Fig. 2. Local lithostratigraphy of the Campanian–lowermost Paleocene strata in the extended Maastrichtian type area (see Fig. 1), with all formations (Fm.), members (Mb.), and horizons (Hz.) shown, as well as dating (in millions of years) as recently published by Vellekoop et al. (2022). Abbreviations: K/Pg, Cretaceous/Paleogene boundary; Camp/Mstr, Campanian/Maastrichtian boundary.
In the lower part of the Maastricht Formation, there are two prominent, fossil-rich levels, directly on top of the St. Pieter and ENCI horizons, the latter representing a storm-generated echinoderm Lagerstätte (Jagt et al. 1998). Keutgen (2018) dated the St. Pieter Horizon at 66.7 Ma, which is less than 1 myr from the terminal K/Pg extinction. Prior to 1975 (Felder 1975a, b), the Valkenburg, Gronsveld, Schiepersberg, and Emael members together were referred to as units Ma and Mb, which Villain (1977) considered to represent a gravelly intrabiomicrosparite, with regional currents constant enough to transport sediment particles over the entire platform, at shallow palaeodepths of 20–40 m, and free from oceanic influence. Sediment reworking resulted in homogenisation over depths of some decimetres, leaving a relatively firm sea floor and clear waters. Liebau (1978) and Sprechmann (1981) characterised this setting as fully marine (middle sublittoral, lagoon-like with near flat coastline and low hinterland), invariably above wave base in the euphotic zone, under subtropical temperatures and typified by the occurrence of seagrass communities and rich thallophyte assemblages, inclusive of washed-in terrestrial plants (Van der Ham et al. 2010).
In general, faunal diversity increases upwards throughout the Maastricht Formation, culminating in the shallow-water, bioherm-like structures of scleractinian corals, hippuritid and radiolitid bivalves, and bryozoans of the middle portion of the Meerssen Member. A combination of “Tethyan” migratory pulses reaching far into northern Europe, attributable to favourable ocean currents, intimately linked with inversion tectonics in some areas and suitable substrates/preferred temperature ranges in (extremely) shallow-water settings, may offer explanations for this trend (Bless et al. 1987; Bless 1989; Jagt 1999, 2000).
Distribution and taxonomy
of dercetid fishes
The family Dercetidae constitutes an extinct group of marine aulopiform actinopterygians that are characterised by a slender, elongated body and a rostrum, plus typical heart-shaped or triradiate scutes arranged in lines along the body (Taverne and Goolaerts 2015). Dercetids ranged from the early Late Cretaceous (Cenomanian) to the earliest Paleocene (Danian) (Davis 1890; Taverne 1987), while their palaeobiogeographical distribution is known to have covered most of the globe during the Cretaceous, with records from Europe, Asia, Africa, and North and South America (Table 1; see Agassiz 1834; Hay and Day 1903; Taverne 1987; Taverne and Goolaerts 2015; Vernygora et al. 2017; Argyriou and Davesne 2021; Argyriou et al. 2022; Díaz-Cruz et al. 2022). Most dercetid taxa have been recorded from the area of the former eastern Tethys Ocean (Europe, Africa, and Asia). Notable records include material recovered from Cenomanian sandstone of Kipali (Congo) (Casier 1965; Taverne 1987), lower Cenomanian carbonates at Ein-Yabrud (Israel) and Haqel (Lebanon) (Hay and Day 1903; Chalifa 1989), Turonian–Santonian chalks in southern England (Agassiz 1834), Campanian siliceous limestones of Komorów (Poland) (Bieńkowska-Wasiluk et al. 2015), Campanian–Maastrichtian carbonates near Nardò (Italy) (Taverne 2005a, b, 2006a, b), biocalcarenites in the Maastrichtian type area (Belgium, the Netherlands) (Friedman 2012; Taverne and Goolaerts 2015; Wallaard et al. 2019), and the Danian Saltholm Limestone in south-west Sweden (Davis 1890).
Table 1. List of all valid species currently included in the Dercetidae, with their known biogeographical distribution and stratigraphical ranges, sorted based on their known stratigraphical ranges. References list the original description of any given taxon and any relevant redefinitions (adapted from Taverne et al. 2005a; Silva and Gallo 2011; Wallaard et al. 2019; Chida et al. 2023).
|
Stratigraphy |
Species |
Distribution |
Reference |
|
Danian |
Scandiadercetis limhamnensis |
Denmark, Sweden |
|
|
Maastrichtian |
Beukidercetis grandis |
the Netherlands |
|
|
Maastrichtian |
Cyranichthys jagti |
the Netherlands |
|
|
Maastrichtian |
Apuliadercetis indeherbergei |
Belgium |
|
|
Upper Senonian? |
Pelargorhynchus dercetiformes |
Germany |
|
|
Campanian–Maastrichtian |
Ophidercetis italiensis |
Belgium and Italy |
|
|
Campanian–Maastrichtian |
Nardodercetis vandewallei |
Italy |
|
|
Campanian–Maastrichtian |
Caudadercetis bannikovi |
Italy |
|
|
Campanian–Maastrichtian |
Apuliadercetis tyleri |
Italy |
|
|
Campanian |
Dercetis magnificus |
Canada |
|
|
Campanian |
Apuliadercetis gonzalezae |
Mexico |
|
|
Santonian–Maastrichtian |
Dercetis triqueter |
Belgium, Lebanon, Poland |
|
|
Santonian–Campanian |
Benthesikyme armatus |
Germany |
|
|
Santonian |
Benthesikyme rostralis |
Italy and Lebanon |
|
|
?Turonian–Santonian |
Dercetis elongatus |
England |
|
|
Turonian |
Hastichthys regio |
Mexico |
Blanco and
Alvarado-Ortega 2006; |
|
Turonian |
Hastichthys totonacus |
Mexico |
|
|
Turonian |
Candelarhynchus padillai |
Colombia |
|
|
Turonian |
Brazilodercetis longirostris |
Brazil |
|
|
Cenomanian |
Hastichthys gracilis |
Israel |
|
|
Cenomanian |
Dercetoides venator |
Israel |
|
|
Cenomanian |
Rhynchodercetis serpentinus |
Lebanon |
|
|
Cenomanian |
Rhynchodercetis gortanii |
Italy |
|
|
Cenomanian |
Rhynchodercetis yovanovitchi |
Italy and Morocco |
|
|
Cenomanian |
Rhynchodercetis hakelensis |
Lebanon |
|
|
Cenomanian |
Cyranichthys ornatissimus |
Congo |
Documentation from the area of the western Tethys Ocean (North and South America) is currently much less detailed, which is presumably linked to collection failure and/or poor fossilisation potential (Vernygora et al. 2017). This lack of detailed data may lead to complications when trying to create a comprehensive biogeographical and/or phylogenetic review of dercetids. However, there are a handful of records from Turonian shale and siltstone levels in the Pelotas Basin of Brazil (de Figueiredo and Gallo 2006) and from coeval mudstones near Boyacá (Colombia) (Vernygora et al. 2017), Campanian marly limestones at Tzimol (Mexico) (Díaz-Cruz et al. 2022) and Campanian strata near Lethbridge in Canada (Chida et al. 2023). Finally, a recent report by Boles et al. (2024) discusses indeterminate dercetid scutes from the Maastrichtian–Danian greensands in Mantua Township, New Jersey (USA), where they persist across the K/Pg boundary.
Historically, dercetids have been included in the extinct suborder Enchodontoidei. However, this grouping is currently considered to be paraphyletic, because members of the outgroup had been retrieved within the ingroup in several analyses (Silva and Gallo 2011; Alvarado-Ortega and Díaz-Cruz 2021; Chida et al. 2023). Other assessments have found dercetids as a monophyletic group, and supported their position within the order Aulopiformes (Silva and Gallo 2011; Díaz-Cruz et al. 2016; Vernygora et al. 2017; Alvarado-Ortega and Díaz-Cruz 2021). However, in some recent studies doubts have been raised over their monophyly, arguing that the morphological traits used to define the group, such as an elongated rostrum and reduced neural spines on the vertebrae, are in fact homoplastic (Alvarado-Ortega and Díaz-Cruz 2021; Chida et al. 2023). In short, this would mean that there currently are no synapomorphies to define the Dercetidae.
At present, up to eighteen genera are placed in this family, although not all are widely accepted amongst ichthyologists (Silva and Gallo 2011; Chida et al. 2023). A review of the current consensus in dercetid taxonomy is provided in Table 1. Dercetid interrelationships are poorly resolved, although all of the commonly accepted genera are usually retrieved as monophyletic within phylogenetic analyses (Silva and Gallo 2011; Díaz-Cruz et al. 2016; Vernygora et al. 2017; Alvarado-Ortega and Díaz-Cruz 2021; Chida et al.2023).
Dercetidae in the Maastrichtian type area
The present paper provides descriptions and illustrations of 22 dercetid individuals from upper Maastrichtian strata in the Maastrichtian type area in north-east Belgium and the south-east Netherlands (Figs. 1, 2). These levels represent a shallow, locally seagrass-based, marine ecosystem, a few million years prior to the Cretaceous/Paleogene (K/Pg) extinction event (Van der Ham et al. 2007; Vellekoop et al. 2022). Prominent vertebrates in these strata comprise mosasaurs, crocodiles, plesiosaurs, sea turtles and fish (Mulder 2003; Friedman 2012; Heere et al. 2023). To date, five dercetid fish species have been documented from the Maastrichtian type area, namely Apuliadercetis indeherbergei, Cyranichthys jagti, Dercetis triqueter, Ophidercetis italiensis, and Pelargorhynchus grandis (reassigned herein to Beukidercetis gen. nov.) (Friedman 2012; Taverne and Goolaerts 2015; Wallaard et al. 2019). For now, A. indeherbergei, Cyranichthys jagti, and Pelargorhynchus grandis are known exclusively from the study area. This, and the erection of five species based on merely seven specimens, clearly illustrates the high diversity of dercetid fish in the region.
Dercetid fossils from the Maastrichtian type area are often misidentified as burrows filled with a range of skeletal remains of fish (such as vertebrae, scutes, teeth, scales, and ribs), in particular as the ichnofossil taxon Lepidenteron lewesiensis (see Suhr 1988). This trace fossil has a convoluted nomenclatural history, having been previously referred to as a petrified eel, a dercetid fish (Dercetis elongatus) or the polychaete worm Terebella (Agassiz 1843; Davies 1879; Jurkowska and Uchman 2013). Lepidenteron lewesiensis is defined as an unbranching burrow that lacks walls, usually filled with a variety of undigested bony fish remains, in particular scales (Jurkowska and Uchman 2013; Bieńkowska-Wasiluk et al. 2015). Possible producers of this type of burrow include worms, anguilliform fish or decapod crustaceans. The mislabelling of individual specimens of dercetids as Lepidenteron lewesiensis has led to underappreciation of the abundance and diversity of these fish in the Maastrichtian type area. There are three key differences between dercetid remains and the ichnofossil Lepidenteron lewesiensis, namely the colour of the fossil, its overall structure and composition. Dercetid remains are often black in colour, while the majority of burrows contain reddish-brown skeletal elements. In addition, examples of Lepidenteron are often preserved in characteristic tube shapes, while dercetid fish remains are more scattered. Lastly, associated dercetid remains may be ascribed to a single individual, whereas burrows contain multiple indeterminate bony fish taxa.
Most dercetid specimens in the Maastrichtian type area are disarticulated and broken up into pieces, due to the fragile nature of the material. In view of this, scute morphology is the most reliable criterion for identifying the material. Some dercetid scutes currently known from the Maastrichtian were previously illustrated elsewhere (Fig. 3). The material presented herein greatly increases dercetid diversity in the Maastrichtian type area, by comprising two new genera and four new species. It also provides more details concerning dercetid stratigraphical ranges in the region.
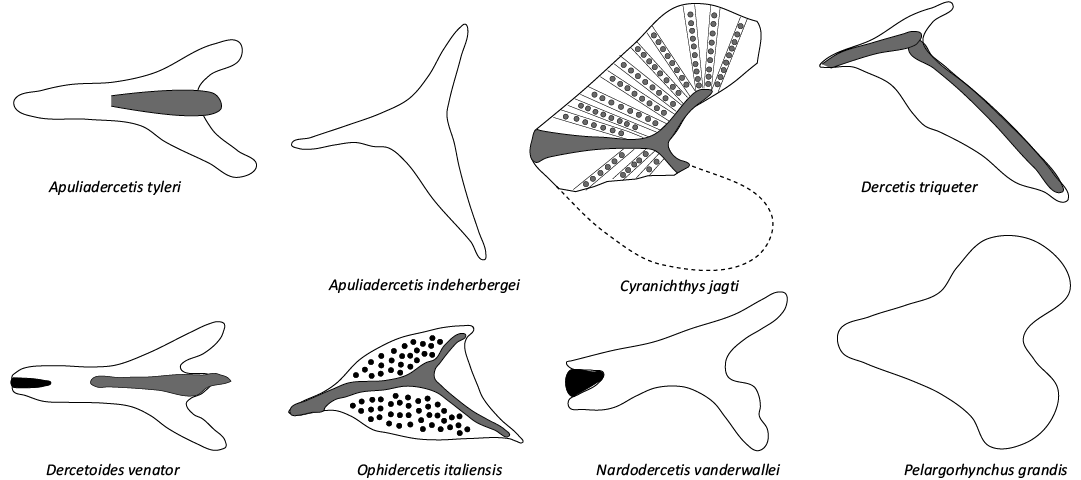
Fig. 3. Schematic drawings of scute morphology in various dercetids from the type Maastrichtian, plus others that are relevant here. Grey infills represent convex surfaces (e.g., ridges/tubercles), black infills stand for concavities (e.g., pits/canals). Modified from Chalifa (1989: fig. 11), Taverne (2005a: fig. 8), Taverne (2005b: fig. 9), Taverne (2006a: fig. 11), Taverne and Goolaerts (2015: figs. 7, 14, 22), and Wallaard et al. (2019: fig. 5).
Material and methods
Twenty-two individual finds of dercetid fish, recovered from five localities (Fig. 1) in the Maastrichtian type area, are described and illustrated. In the field, these specimens were collected from exposed blocks of biocalcarenite, occasionally at surfaces after quarrying activities or by splitting blocks along bedding planes. At the laboratory, further preparation was executed by two of us (JJAH and JJWW) and Osteofix was used as a consolidating agent.
Photographs were taken with a Canon 60D camera and Samsung A53 phone camera. Detailed pictures of specific anatomical features were taken using a camera-equipped binocular and subsequently compounded by the stacking software Helicon Focus. Reconstructions were produced by tracing techniques in Inkscape. Systematics and taxonomy were performed in accordance with the ICZN (Ride et al. 1999).
Systematic palaeontology
Class Actinopterygii Klein, 1885
Order Aulopiformes Rosen, 1973
Suborder Enchodontoidei Rosen, 1973
Family Dercetidae Woodward, 1901
Genus Beukidercetis nov.
Zoobank LSID: urn:lsid:zoobank.org:act:6AD52087-6BC6-433E-A868- 9552A9C5AAD7.
Etymology: In memoriam of Paul Beuk (1965–2025), curator of biology at the Natural History Museum Maastricht (NHMM) for 15+ years, who very suddenly passed away during this project.
Type species: Pelargorhynchus grandis Wallaard et al., 2019, designated herein; see below.
Species included: Beukidercetis grandis (Wallaard et al., 2019), Beukidercetis lissus sp. nov.
Diagnosis.—Skull roof bearing a crest posteriorly. Curved and toothless rostrum. Premaxillae unfused. Medio-posterior side of ribs ornamented with tubercles. Squamation consits of large, cordiform scutes, which are partially or completely unornamented, in addition to smaller scales ornamented with spikes and ridges.
Stratigraphic and geographic range.—Upper Maastrichtian (Upper Cretaceous) of Belgium and the Netherlands.
Beukidercetis grandis (Wallaard et al., 2019)
2019 Pelargorhynchus grandis sp. nov; Wallaard et al. 2019: 2, figs. 3–9.
Holotype: MAB3688, disarticulated individual, showing a partial cranium and rostrum, scute and smaller scales.
Type locality: Former ‘t Rooth quarry, Bemelen, province of Limburg, the Netherlands.
Type horizon: Maastricht Formation, probably Emael Member, upper Maastrichtian.
Original diagnosis.—Body squamation consisting of one or more lateral rows of smooth, cordiform scutes, combined with small, ornamented scales; scale ornamentation comprising several small spines and ridges originating from median crest; skull bones with high degree of curvature; premaxillae not fused, maxilla and premaxilla both lacking teeth.
Material.—The holotype, MAB3688, retains elements of the skull and rostrum, in addition to scutes and scales. Wallaard et al. (2019) assigned it tentatively to the genus Pelargorhynchus Von der Marck, 1858.
Description .—See Wallaard et al. 2019.
Remarks.—The curved upper rostrum, lack of teeth and scute morphology of the holotype MAB3688 were indicated by Wallaard et al. (2019), to potentially merit the erection of a new genus. At the time, however, the available material was deemed insufficient to draw any definitive conclusions. Here, the newly collected material of NHMM K 3805 (see below), in combination with the original data from MAB3688, did justify the erection of a new genus. Pelargorhynchus grandis is thus designated as the type species of Beukidercetis gen. nov.
Stratigraphic and geographic range.—Maastrichtian (Upper Cretaceous) of the Netherlands.
Beukidercetis lissus sp. nov.
Fig. 4.
Zoobank LSID: urn:lsid:zoobank.org:act:DB5D889B-923E-4C60-8A38- F4FCE5E781E8.
Etymology: From the Ancient Greek λίσσος, smooth; referring to the smooth rims of the scutes.
Holotype: NHMM K 3805 (M.M.M./J. Kuypers Collection), partially articulated individual, preserving cranial and rostral remains, as well as ribs and scutes.
Type locality: CBR-Romontbos quarry, Eben-Emael, province of Liège, Belgium.
Type horizon: Maastricht Formation, basal Emael Member, upper Maastrichtian.
Diagnosis.—Skull roof largely flat, bearing a crest posteriorly. Medio-posterior side of ribs sparsely ornamented with tubercles. Large cordiform scutes, bearing a rounded midline, originating from centre and opening up and bifurcating anteriorly. Scute surface ornamented with tubercles, unfused and arranged in lines, except for smooth rim.
Material.—The holotype, NHMM K 3805, constitutes the largest dercetid known from the Maastrichtian type area to date, judging from the size of scutes and rostrum. The material includes cranial and rostral remains, in addition to ribs and isolated scutes. Associated with the specimen are several bivalves, cheilostome bryozoan colonies, bourgeticrinid crinoid stalks, the lateral armplates of an ophiomusaid brittle star, an echinoid shell piece of Diplodetus sp., and finally the irregular echinoid Oolopygus pyriformis (Leske, 1778) (Fig. 4A1).
Description.—Cranium: Posterior right region of skull roof is preserved, containing parts of supraoccipital, right parietal (and right pterotic?) (Fig. 4A2). Fairly broad and flattened, with minute wavy structures radiating from centre. Along midline is a small crest, which appears to be partially hollow. Branchiostegal rays of various sizes also present (Fig. 4A2, A3, A6), showing various degrees of curvature and broadening along their length.
Rostrum: Portions of frontals (Fig. 4A3, A4) preserved, both fractured. Frontal with convex shape, being thicker dorso-medially. Dorsal surface of frontal ornamented with tubercles, slightly larger than those on scutes. The ornamented surface also has thick, rounded ridge that extends along antero-posterior axis. Lateral side of frontal consisting of a thin, curved sheet of bone. Most of rostrum preserved merely as fractured pieces. A large, rounded piece of the left premaxilla exposes its dorsal side (Fig. 4A1). The surface of this bone is fairly smooth. It is unfused to the other premaxilla and shows no signs of association with a counterpart either. The posterior end of the bone curves slightly distally, presumably creating a slot for the mesethmoid when associated with its right counterpart.

Fig. 4. Dercetid fish Beukidercetis lissus gen. et sp. nov., NHMM K 3805 (holotype) from the Maastrichtian, Upper Cretaceous, of Eben-Emael, Belgium. General view of the specimen (A1–A4), left mandible (A5), ribs and branchiostegal rays (A6), scute (A7).
Part of the posterior half of the left mandible is preserved (Fig. 4A5). Its surface is smooth, showing no signs of mandibular canals. The mandible has a rounded shape posteriorly, but broadens into a flatter structure anteriorly. At the posterior end, part of the articulation with the angular-articular complex is preserved, showing a conical structure. Both premaxilla and mandible are edentulous.
Axial skeleton: No vertebrae are preserved, but several ribs are present, identified by their curved nature and smaller size relative to pieces of the rostrum (Fig. 4A6). None of the ribs are fully preserved, but their morphology is well defined, being relatively straight and bearing a knob-like process at their upper end, with sharp edges and a fairly large attachment area for the vertebrae. The medio-posterior surface of the rib appears to be ornamented with sparse, minute striations.
Scutes: Several large (>50 mm in length) cordiform scutes are preserved (Fig. 4A1, A2, A7). Corners are sharp in some, but blunt in most. Scutes carry a median ridge, which originates around the centre, being roughly equal in height across the length of the scute and not bifurcating or extending past the scute margin. The surface of the scutes is ornamented with tubercles, arranged tightly in rows, but not fused. The anterior and lateral rims of the scute are smooth, lacking ornamentation. At the anterior end of the median ridge lies a vascular pit, opening up anteriorly. The walls of this canal extend antero-laterally, onto the smooth surface of the scutes, forming a crescentic cross section.
Remarks.—NHMM K 3805 is identified as a dercetid based on its cordiform, ornamented scutes and the elongate rostrum and frontals. However, the scutes of this specimen have a morphology that is atypical for any dercetid described to date, thus warranting the erection of a new genus. The closest possible analogy, as far as scute morphology is concerned, is Cyranichthys, which has generally cordiform shaped scutes with an ornamentation consisting of unfused tubercles (Figs. 6A5, 7A4, 8A4, A5, B2, 10A3; compare Taverne 1987; Taverne and Goolaerts 2015). It is different from Cyranichthys, however, in the notably unornamented rim of the scute and the ridge which opens up anteriorly.
Dercetid ribs are usually only partially preserved, if at all, which is linked to the fragile nature of most specimens. The notably large size of the present dercetid probably explains why ribs are preserved fairly well. Although no vertebrae are retained, it may be assumed that they would have been fairly large as well (>50 mm). Unfortunately, all diagnostic characters of the axial skeleton in dercetids involve the processes on vertebrae, which explains why a comparison of Beukidercetis lissus gen. et sp. nov. and other dercetids based on the axial skeleton is impossible.
Stratigraphic and geographic range.—Maastrichtian (Upper Cretaceous) of Belgium and the Netherlands.
Genus Caudadercetis Taverne, 2006b
Type species: Caudadercetis bannikovi Taverne, 2006b, by original designation; Campanian–Maastrichtian, Nardò, Italy.
Stratigraphic and geographic range.—Campanian–Maastrichtian of Italy; Maastrichtian of the Netherlands and Belgium. All Upper Cretaceous.
Caudadercetis taverni sp. nov.
Fig. 5.
Zoobank LSID: urn:lsid:zoobank.org:act:7ADC140F-5522-45A7-BE40- 0BAC1878D46A.
2012 Dercetidae indet.; Friedman 2012: 130, fig. 7C.
2015 Cyranichthys jagti sp. nov.; Taverne and Goolaerts 2015: 25, fig. 15A.
Etymology: In honour of Louis Taverne for his numerous contributions to palaeoichthyology, and in particular for his work on dercetids from Maastrichtian strata in Belgium, Italy, and the Netherlands.
Holotype: NHMM JJ 16220, a disarticulated individual with vertebrae and scutes.
Type locality: Former ENCI-HeidelbergCement Group quarry, Sint-Pietersberg, Maastricht, the Netherlands.
Type horizon: Maastricht Formation, Emael Member, upper Maastrichtian.
Diagnosis.—Vertebrae bear short neural arches, with forward-facing anterior process and large haemal arch with two tall haemophyses. Scutes bear steep ridges on midline, ending posteriorly in pointed extensions. Scute surface ornamented with fused tubercles, forming ridges that radiate from midpoint and do not reach scute edges.
Material.—The holotype, NHMM JJ 16220 (Fig. 5A), represents a disarticulated individual retaining vertebrae and scutes (found by JWMJ). The paratype, NHMM K 2372 (Fig. 5B), from the basal Emael Member in an unidentified quarry near Eben-Emael (Belgium), is an isolated large scute (10 mm in length), previously described by Friedman (2012) and Taverne and Goolaerts (2015). The latter authors designated this single scute as paratype of a new species, Cyranichthys jagti. It is here transferred to a new species of Caudadercetis (see below).
Description.—Axial skeleton: Several eroded vertebrae are preserved (Fig. 5A2), their length being about twice their height. They possess a short neural arch with short, forward-facing anterior processes. There is a fairly large haemal arch, covering the entire vertebral centre, containing two pairs of haemophyses, the anterior of which is directed ventrally, the posterior one laterally (Fig. 5A2, B).

Fig. 5. Dercetid fish Caudadercetis taverni sp. nov. from the Maastrichtian (Upper Cretaceous) of Maastricht, the Netherlands (A) and basal Emael Member in an unidentified quarry near Eben-Emael, Belgium (B). A. NHMM JJ 16220 (holotype), specimen in general view (A1) and details of isolated vertebrae and scutes (A2, A3). B. NHMM K 3272 (paratype), isolated scute. C. Schematic drawing of vertebral morphology (lateral view). D. Schematic drawing of a scute exposed in A1, showing tubercles and ridges (grey) that make up its ornamentation. C, D not to scale.
Scutes: Two large (about 5 mm in length), cordiform scutes are preserved in NHMM JJ 16220 (Fig. 5C); NHMM K 2372 is another (Fig. 5B). Scutes in NHMM JJ 16220 are slightly wider than long, with broad posterior limbs. Scutes have a steep median ridge that ends posteriorly in a pointed extension, which presumably overlaps the anterior end of the subsequent scute. Scutes are ornamented with radiating striations emanating from centre of the scute (Fig. 5A3, B). These lines are situated at roughly equal distances from each other and consist of fused tubercles. Lines may occasionally bifurcate near the posterior side (Fig. 5A3, D). Ornamentation does not reach the edge of scutes in either specimen, leaving a smooth area along the outer edge. There are no visible perforations of the sensory canal.
Remarks.—NHMMJJ 16220 is assigned to Caudadercetis based on its vertebral morphology, bearing two characteristic haemophyses (Fig. 5A2, C), and identified as a new species on account of its squamation (Fig. 5A3). Unfortunately, no detailed description or figure of the scutes in the type species, Caudadercetis bannikovi, from the Campanian–Maastrichtian of Italy (Taverne 2006b) has been published. However, squamation in NHMM JJ 16220 is compatible with the brief description given of cordiform scutes that bear an ergot-shaped posterior protuberance in Caudadercetis bannikovi. Caudadercetis taverni sp. nov. can be distinguished from the type species by the presence of tubercles on the scutes (Taverne 2006b).
NHMM K 3272 (Fig. 5B) was previously designated a paratype of Cyranichthys jagti (Taverne and Goolaerts 2015). However, upon closer inspection, the morphology of the scute appears dissimilar from those of Cyranichthys, yet remarkably similar to those of NHMMJJ 16220. It bears a midline, which does not bifurcate and carries a hook-shaped extension posteriorly (Fig. 5B). In addition, tubercles are fused tightly, forming radiating lines, as opposed to unfused and more widely spaced tubercles in both Cyranichthys jagti and Cyranichthys ornatissimus (Figs. 3, 6A5; Taverne and Goolaerts 2015). It should be noted that the isolated scute (NHMM K 2372) does differ from the holotype of Caudadercetis taverni sp. nov. in being longer than wide. This difference may be explained by the placing along the body.
Stratigraphic and geographic range.—Maastrichtian (Upper Cretaceous) of Belgium and the Netherlands.
Genus Cyranichthys Taverne, 1987
Type species: Dercetis ornatissimus Casier, 1965; Cenomanian of Kipali (Congo).
Stratigraphic and geographic range.—Cenomanian (Upper Cretaceous) of Congo and Maastrichtian (Upper Cretaceous) of the Netherlands and Belgium.
Cyranichthys jagti Taverne & Goolaerts, 2015
Figs. 6, 7.
2012 ?Ophidercetis Taverne, 2005; Friedman 2012: 132, fig. 7D–F.
2015 Cyranichthys jagti sp. nov.; Taverne and Goolaerts 2015: 25, figs. 11–15.
Material.—NHMM 2024 011 (Fig. 6) from Lanaye Member Gulpen Formation, Maastrichtian, Upper Cretaceous at Marnebel quarry, Eben-Emael (Fig. 1), is an articulated specimen, with several cranial and rostral bones, four anatomically aligned vertebrae, as well as several isolated ones and a few isolated, cordiform scutes (found by Dirk Eysermans). NHMM JJ 16594a–c (Fig. 7), basal Valkenburg Member, Maastricht Formation, Maastrichtian, Upper Cretaceous at the former ENCI-HeidelbergCement Group quarry, is an individual in partial anatomical preservation, with rows of vertebrae and scutes, in addition to portions of the rostrum.
Description.—Rostrum: A pair of elongate, unfused and edentulous bones, preserved in NHMM 2024 011 (Fig. 6A4), may be identified as the mandible, based on their triangular shape. It has flipped, leaving the anterior end pointing posteriorly, and its dorsal surface has a long concave trough, which lacks any teeth or alveoli. The ventral surface is ornamented with minute longitudinal striations. In addition, there is a small, striated piece of the right frontal/premaxilla (Fig. 6A2), identified as such on account of the curvature of the bone and its wider posterior end. Finally, there is an unidentified cranial element, which could represent either the frontal or parietal (Fig. 6A3); this has a curved L-shape and a concave surface on its dorsal side. In NHMM JJ 16594, only portions of the mandible appear to be preserved, judging from their size and the concave trough that is also seen in NHMM 2024 011 (Fig. 6A4).

Fig. 6. Dercetid fish Cyranichthys jagti (Taverne & Goolaerts, 2015) (NHMM 2024 011) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Complete specimen (A1), right frontal/premaxilla (A2), left parietal/frontal (A3), mandible and portion of vertebral column (A4), isolated scute (A5).
Axial skeleton: Four vertebrae are preserved in anatomical alignment in NHMM 2024 011 (Fig. 6A4), alongside several isolated, fractured ones. The first few vertebrae are about twice as long as successive ones. The first vertebrae are poorly preserved. The more posterior, isolated vertebrae have a large, round haemophysis and bear two pairs of thick, flat posterior, transverse processes. Their neural arches are rounded and have notably thick walls. The vertebrae in NHMM JJ 16594a–c are also preserved in anatomical alignment, with two separate rows of 5+ vertebrae (Fig. 7A3). Although their preservation is poorer, the thick neural arch and remains of the haemophyses are also visible.
Scutes: Several isolated cordiform scutes are preserved in NHMM 2024 011. These are convex and carry a median ridge that ends in a minute pointed extension (Fig. 6A5). The surface of the scutes is ornamented with ridges and tubercles that are arranged in lines radiating from the midline. Scutes in NHMM JJ 16594 are only partially prfeserved, but do show a similar morphology. A few of the fragmentary scutes are still preserved in their original alignment, with the anterior end of each scute overlapping the posterior end of the previous (Fig. 7A4).
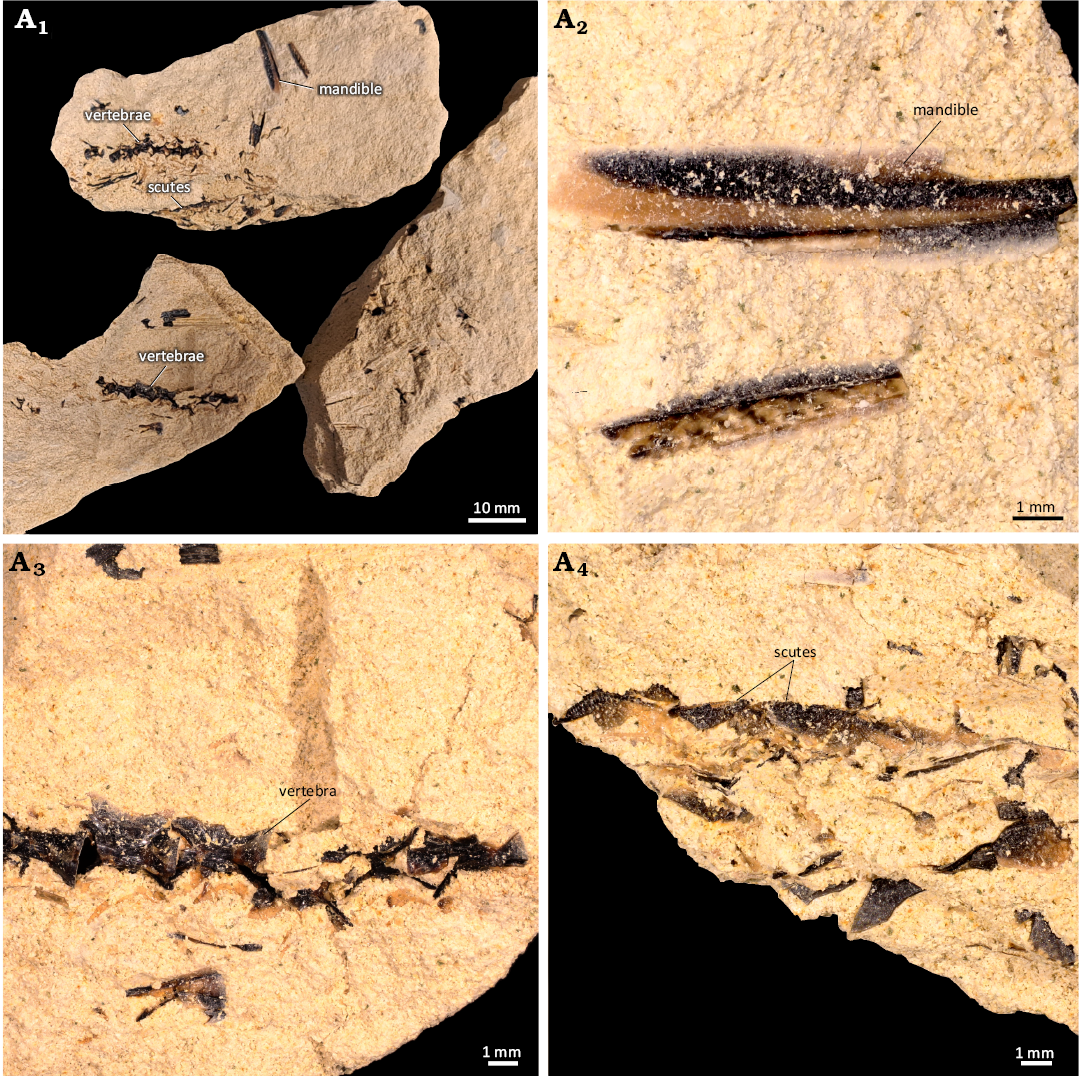
Fig. 7. Dercetid fish Cyranichthys jagti (Taverne & Goolaerts, 2015) (NHMM JJ 16594) from the Maastrichtian (Upper Cretaceous) of Maastricht, the Netherlands. Complete specimen (A1), rostrum (A2), vertebrae (A3), scutes (A4).
Remarks.—NHMM 2024 011 is identified as Cyranichthys jagti on the basis of its squamation, with convex, cordiform scutes that are ornamented with tubercles that radiate in lines from a central ridge (Fig. 6A5). This adds to our understanding of Cyranichthys jagti, because the holotype of that species does not retain the mandible or vertebral anatomy (Taverne and Goolaerts 2015). It is shown here that the mandible, like the upper jaw, is toothless and that the vertebrae have rounded haemophyses and two pairs of thick, flat transverse processes. NHMM JJ 16594 is referred to Cyranichthys jagti on account of its squamation and concave, toothless mandible fragments (Fig. 7A2). It does show an interesting pattern of preservation in scutes (Fig. 7A4), which lie in a straight line, partially overlapping one another. It is unclear whether this was the case in life or whether this is a preservational (taphonomic) factor.
Stratigraphic and geographic range.—Maastrichtian (Upper Cretaceous) of Belgium and the Netherlands.
Cyranichthys sideralis sp. nov.
Figs. 8–10.
Zoobank LSID: urn:lsid:zoobank.org:act:DDCD1264-7D04-4F31-999 F-866F6E9F5170.
Etymology: From Latin sideralis, related to the stars or outer space; referring to the characteristic scutes, which have sharp edges and resemble the contours of a jet plane.
Holotype: MAB 16755, partially articulated individual, preserving cranial and rostral remains, as well as ribs and scutes.
Type locality: CBR-Romontbos quarry, Eben-Emael, province of Liège, Belgium.
Type horizon: Maastricht Formation (unspecified level), upper Maastrichtian.
Diagnosis.—Branchiostegal rays ornamented on lateral surface and penetrated by vascular network ventrally. Vertebrae bearing broad neural arches, penetrated by a vascular network. Haemal arch strongly or completely reduced, a pair of rounded haemophyses on either side. Premaxilla ornamented with longitudinal striations over entire length. Mandible toothless, labial ridge increasing in height markedly near posterior end. Jet plane-shaped scutes with sharp edges, bearing a ridge on their midline that ends in a pointed extension and bifurcates in the largest scutes. Surface of scute is ornamented with tubercles that are arranged semi-randomly.
Material.—MAB16755, the holotype (Fig. 8A), originates from an unspecified level within the lower Maastricht Formation (found by JJWW). Based on the presence of a serpulid tube and the small bivalve Syncyclonema haeggi Dhondt, 1971, this could hail from either the Gronsveld, Schiepersberg or Emael members. Of note is that this individual is three-dimensionally preserved, featuring many jet plane-shaped scutes, around ten isolated vertebrae and portions of the rostrum. MAB16756, a paratype (Fig. 8B1, B2, B3), from the Marnebel quarry, Eben-Emael, Belgium (found by Maria Verschelling-Wallaard), preserves four vertebrae in anatomical alignment, alongside several isolated vertebrae and a single scute. MAB16757 (Fig. 10), from the Meerssen Member at the ENCI quarry, the Netherlands (found by René H.B. Fraaije), preserves a single jet plane-shaped scute, partial rostral remains and several well-preserved vertebrae.
Description.—Cranium: MAB 16755 retains a characteristic, curved, hemicylindrical cranial element, which appears to be the posterior part of a branchiostegal ray (Fig. 8B2). It has an ornamented medial ridge and a lateral ridge, the latter increasing in height posteriorly. It originates from the right side of the skull and has its medial side exposed. The ventral interior surface of this element is perforated irregularly by a vascular system.

Fig. 8. Dercetid fish Cyranichthys sideralis sp. nov., from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. A. MAB16755 (holotype), general view (A1), branchiostegal ray (A2) mandible (A3), partial mandible? and scutes (A4), premaxilla and scutes (A5). B. MAB16756 (paratype), general view (B1), isolated scute (B2), isolated vertebra (B3).
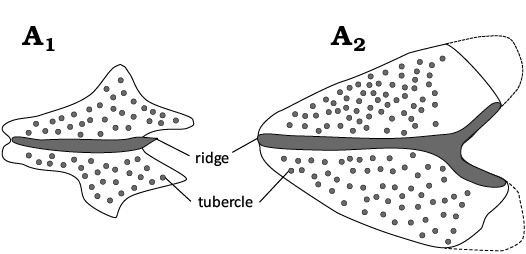
Fig. 9. Dercetid fish Cyranichthys sideralis sp. nov., from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Schematic drawings of scute morphology, with both small (A1) and large (A2) scutes. Grey infills represent tubercles and ridges; drawn on the basis of scutes visible in Fig. 8A3.
Rostrum: At least three pieces of the rostrum are preserved in MAB16755. The first is a straight bone that narrows over its length and is ornamented with longitudinal striations (Fig. 8A5). This element is identified as the right premaxilla, the left aspect of its dorsal surface exposed. The supposed border with the left premaxilla remains embedded in the sediment and is covered with several well-preserved scutes (Fig. 8A5). Thus, the specimen cannot be prepared further, and it is impossible to determine whether or not (and how) the premaxillae were fused. The second is another striated element that is much thinner and appears to be less round (Fig. 8A4). This is tentatively identified as part of the dentary, although it is only fragmentary. The third is an element that has its medial side exposed (Fig. 8A3). This element is identified as the right dentary, based on its articulations and morphology. It is only partially preserved, being fractured on its anterior side and eroded on its posterior. The mandible carries no teeth, nor dental alveoli. It has a steep labial ridge, which markedly increases in height and thickness on the posterior side. There is no lingual ridge, the medial side of the element is broad and flattened. Finally, there is a trough along the entire inner surface of the mandible. On the posterior side eroded remnants of the articulation point with the surangular on the dorso-medial side are seen.
Axial skeleton: As far as the axial skeleton is concerned, neural arches are fairly thick and appear to be perforated by a vascular system (Fig. 8B3). They carry neural spines of considerable length (Fig. 10A2). The haemal arch is strongly or completely reduced. There are two pairs of haemophyses on each vertebral centre, the anterior of which is sharp, and directed laterally and the posterior rounder and directed ventrally (Fig. 10A2). There are also several thin ribs present in MAB16755 and a single rib in paratype MAB16756 (Fig. 8B3).

Fig. 10. Dercetid fish Cyranichthys sideralis sp. nov. (MAB16757) from the Maastrichtian (Upper Cretaceous) of Maastricht, the Netherlands. General view (A1), isolated vertebra (A2), isolated scute (A3).
Scute: Shape of the scute in this form is unique for dercetids, being cordiform in shape, but with notably sharp edges and a much more acute anterior end (Figs. 8A4, A5, B2, 10A3). This renders a rough outline to the scutes, similar to fighter jets. Smaller scutes are more star-like in shape, resembling a McDonnell-Douglass F-15, and larger scutes are more triangular, closer to a Lockheed F-117 Nighthawk. A blunt ridge runs along the midline of the scutes, bifurcating posteriorly in the largest scutes or ending in a pointed extension in smaller ones (Fig. 8A4, A5). The surface of the scutes is ornamented with numerous, semi-randomly arranged tubercles. Most scutes are slightly convex, best seen in medial views of some of the exposed scutes in MAB16755.
Remarks.—All three specimens are identified as Cyranichthys sideralis sp. nov. on account of the unique, jet plane-shaped morphology of their scutes. This feature is highly consistent throughout the material (Figs. 8A1, A4, A5, B2, 10A3). The scutes are large, carry a ridge on their midline and are ornamented with semi-randomly arranged tubercles in the holotype (MAB16755) and paratype (MAB16756). This scute morphology resembles the description provided for the species of Cyranichthys (Taverne 1987), but differs from that of congeners: Cyranichthys jagti Taverne & Goolaerts, 2015, and Cyranichthys ornatissimus (Casier, 1965). Cyranichthys jagti has scutes that are more cordiform in morphology and are ornamented with tubercles arranged in lines radiating from the midline (Fig. 6A5; Taverne and Goolaerts 2015). Scute morphology in Cyranichthys ornatissimus is more variable, with both rounded, cordiform and sharper, star-shaped scutes occurring (Taverne 1987; Taverne and Goolaerts 2015), ornamented with ridges that radiate from the midline and occasionally bear tubercles.
The branchiostegal ray in MAB16755 (Fig. 8A2) has a comparable morphology to that of the branchiostegal rays in Cyranichthys ornatissimus (Taverne 1987). The premaxilla resembles that of Cyranichthys jagti, but is notably striated, a feature that is not as prevalent in NHMM 1993 119, the holotype of Cyranichthys jagti (Taverne and Goolaerts 2015). The axial skeleton may also be described here, with well-preserved vertebrae in both the paratype MAB16756 and MAB16757 (Figs. 8B3, 10A2). These vertebrae resemble those of Cyranichthys jagti, but differ in having two haemophyses rather than one (Figs. 6A4, 7A3).
Stratigraphic and geographic range.—Maastrichtian (Upper Cretaceous) of Belgium and the Netherlands.
Cyranichthys cf. jagti Taverne & Goolaerts, 2015
Fig. 11.
Material.—NHMM JJ 16069, from above the Lava Horizon, Emael Member, Maastrichtian, Upper Cretaceous at the former ENCI-HeidelbergCement Group quarry (Maastricht, the Netherlands) represents a disarticulated individual with partially preserved vertebrae and fragments of scutes exposed.
Description.—Axial skeleton: Three vertebrae are preserved in articulation, but in general most vertebrae are poorly preserved (Fig. 11A2–A4), still showing a rounded neural arch, reduced haemal arches and an anterior pair of laterally directed haemophyses.

Fig. 11. Dercetid fish Cyranichthys cf. jagti (Taverne & Goolaerts, 2015) (NHMM JJ 16069a–c) from the Maastrichtian (Upper Cretaceceous) of Maastricht, the Netherlands. General view (A1), three vertebrae in anatomical alignment (A2), fragmentary vertebrae and scutes (A3–A5).
Scute: A few large, convex, scute fragments are exposed (Fig. 11A4, A5). The scutes carry a median crest, which bifurcates posteriorly. Anterior to the bifurcation, the surface of the scutes is lined with tubercles that are arranged in a semi-regular structure. Posteriorly, the surface is smooth and thin.
Remarks.—It is difficult to determine the affinity of the present specimen below the generic level, because most vertebrae have lost their processes and scutes are preserved only fragmentarily. The ornamentation of the scutes most closely resembles that of Cyranichthys jagti, in having tubercles arranged in lines radiating from the midline (Taverne and Goolaerts 2015).
Genus Dercetis Agassiz, 1834
Type species: Dercetis elongatus Agassiz, 1834, by monotypy; Turonian? near Lewes, southern England.
Stratigraphic and geographic range.—Turonian–Santonian (England); Santonian (Lebanon); Campanian (Canada); Maastrichtian (Belgium, the Netherlands, and Poland).
Dercetis triqueter Pictet, 1850
Figs. 12–16.
1850 Dercetis triqueter sp. nov.; Pictet 1850.
1866 Dercetis triqueter Pictet, 1850; Pictet and Humbert 1866: 3–7.
1878 Leptotrachelus virgulatus sp. nov.; Cope 1878: 67–68.
2005 Dercetis triqueter Pictet 1850; Taverne 2005a: 64, figs. 10–13.
2012 ?Dercetis sp.; Friedman 2012: 131, fig. 7A.
2012 Dercetidae indet.; Friedman 2012: 131, fig. 7B.
2015 Dercetis triqueter Pictet 1850; Taverne and Goolaerts 2015: 22, figs. 4–7.
2015 Dercetis triqueter; Bieńkowska-Wasiluk et al. 2015: 801, fig. 5I
Material.—Two specimens from the Maastrichtian, Upper Cretaceceous, CBR-Romontbos quarry, Eben-Emael, Belgium: MAB16758 (Fig. 12), exposes the right lateral side of the body (found by Arno Savelkoul). Visible are a partially articulated vertebral column, separated into two pieces, an articulated line of triradiate scutes, two pieces of the skull and a few intermuscular bones and/or ribs. Part of the vertebral column is missing. MAB16120a–c (Fig. 13), consists of three parts (found by JJWW); MAB16120a is a small piece of sediment with disarticulated remains of the rostrum, a partial frontal and several vertebrae, plus triradiate scutes. MAB16120b is a piece containing a few vertebrae and a few cranial bones, while MAB16120c is a massive block containing a few triradiate scutes, disarticulated remains of the cranium and vertebrae, plus an unidentified piece of the cranium.
MAB16121 (Fig. 14), from the Maastrichtian, Upper Cretaceceous, Meerssen Member along the Albert Canal between Kanne and Vroenhoven, Belgium (found by Robert Frijns), preserves a few triradiate scutes, a partial frontal, several pieces of the rostrum and a few vertebrae. Two specimens from Maastrichtian, Upper Cretaceceous, Marnebel quarry, Eben-Emael, Belgium: MAB16759 (Fig. 15), represents a burrow infill containing the remains of at least two species of dercetid, including triradiate scutes, two different types of vertebrae and two pieces of the rostrum that show a great variety in size (found by Maria Verschelling-Wallaard). NHMM 2025 017 (Fig. 16), retains the rostrum, a few isolated scutes and approximately ten vertebrae in anatomical alignment (found by Lars P.J. Barten).
Description.—Cranium: The posterior part of the skull is preserved in MAB16758 (Fig. 12A1, A2), including the supra-occipital, epioccipitals, parietals and pterotics. Parts of the anterior and lateral margins of the skull are not preserved, but the structure is generally symmetrical and bat shaped. The lateral edges of the pterotics are ornamented with a minor ridge. A minute posterior extension originates from the posterior side of the supra-occipital. The articulation surface for the pterotic is partially disarticulated.
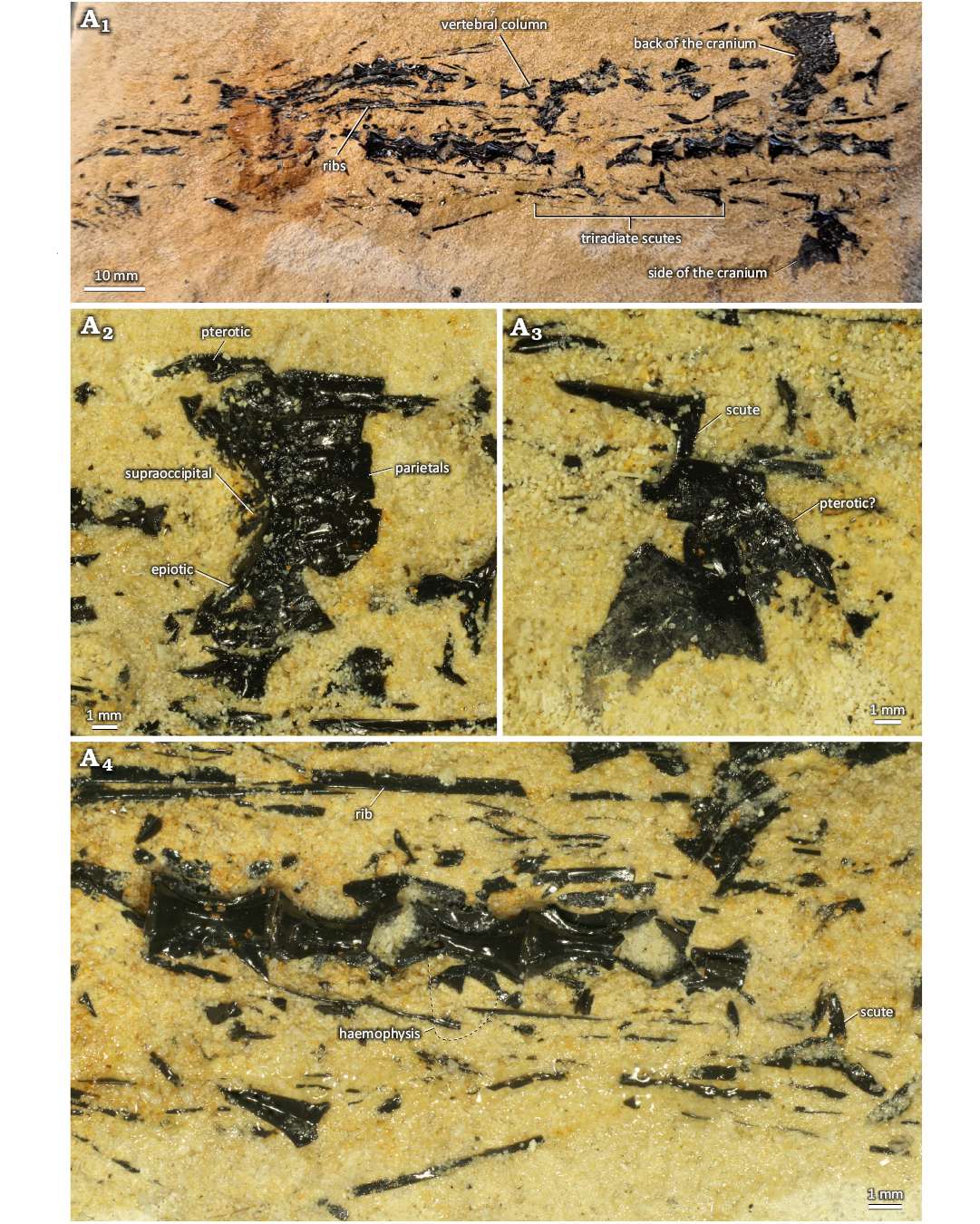
Fig. 12. Dercetid fish Dercetis triqueter (Pictet, 1850) (MAB16758) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Complete specimen (A1), posterior portion of cranium and first vertebra (A2), right lateral side of cranium (A3), part of vertebral column (A4).
Ventral to the collapsed vertebral column in MAB16758 (Fig. 12A3) lies a thick, partially eroded cranial bone, which is tentatively identified as part of the pterotic. The lateral side of this bone is exposed, showing several tall ridges, the most notable of which is a triradiate structure. Ventrally, on the lateral surface, lies a concave pit.
The frontal is preserved in MAB16120, in two parts: the left posterior half in MAB16120a and part of the anterior in MAB16120b (Fig. 13A3, A4). It is also partially preserved in MAB16121a (Fig. 14A2). Combined, these parts make an elongate frontal, with a broadened posterior platform, which is ornamented with longitudinal striations. On the left lateral surface lies an opening for the supra-occipital sensory canal. The frontal expands laterally around the level of the orbit.
The posterior end of the parasphenoid is preserved in MAB16120b (Fig. 13A6), with the anterior end presumably remaining hidden in matrix. Two parallel ridges run along the antero-posterior axis of this bone. In addition, the right sphenoid is visible in MAB16120b (Fig. 13A2). It is a smooth, thin, and curved bone, with a fairly tall and rounded ridge on its lateral surface, along the posterior margin of the orbit. Ventral to this bone lies another curved bone, which could potentially be one of the infraorbitals (Fig. 13A2).

Fig. 13. Dercetid fish Dercetis triqueter (Pictet, 1850) (MAB16120) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Complete specimen (A1), isolated pieces of cranium (A2–A6), disarticulated pieces of rostrum, alongside some scutes and vertebrae (A7).
There are two other cranial elements: a partial branchiostegal ray (MAB16120b; Fig. 13A3), which has a mostly flat shape, with rounded edges that curve up dorsally and abraded processes on the lateral surface, which probably served as muscle attachments; and an unidentified elongate bone (MAB16120; Fig. 13A5). The surface of this bone is ornamented with longitudinal striations, and it narrows near its tip; this might represent a branchiostegal ray.
The posterior part of the left dentary (lateral view exposed) and a few portions of the right dentary (dorsal view) are preserved in MAB16120a (Fig. 13A7). The elongate part of the left dentary and right articular is also preserved in MAB 16121a (Fig. 14A2). Together, these allow for the reconstruction of an elongate dentary, which ends anteriorly in a fairly sharp angle. The lateral surface of the dentary is fairly smooth but bifurcates posteriorly to form the articulation with the articular. At the start of this bifurcation, the dentary also contains an opening for the mandibular sensory canal. The articular bone itself is a tall bone, with a thick ridge running along the midline of the lateral surface. Dorsal to this ridge lies a concave area of roughly equal depth. The anterior end of the dentary is roughly flat, with a bulging ridge on the lateral side, rendering an L-shaped cross section to the dentary. The dentary is lined with two types of teeth (Figs. 13A7, 14A2): larger and sharper teeth on the lateral side, most of which have been lost and only recognisable by alveoli in the material, and smaller, more numerous, bulbous teeth, arranged in several semi-regular rows.
The anterior part of the right premaxilla is also preserved in MAB16120a (Fig. 13A7). This is a thin, smooth bone, which bulges briefly on the anterior side, to form the articulation surface with the bifurcated mesethmoid (not preserved here). The medial side remains embedded in the sediment, so it is not possible to determine the morphology of the premaxillary teeth in this particular specimen. There is a small, toothed element in MAB16120a (Fig. 13A7); a thin, partially flat bone, which carries two large, flattened teeth. A ridge runs along the exposed surface of this element. Too little of this bone is preserved to identify it with certainty, but this could be a part of the maxilla, which lies unattached behind the premaxilla in Dercetis triqueter (see Taverne 2005a: fig. 10). The anterior end of this structure is not preserved, precluding definite identification of this bone.
Part of the right articular and angular is preserved in MAB16121b (Fig. 14A3). They form an elongate structure that carries a ridge on its lateral surface near the ventral side, which is hollow and contains the mandibular sensory canal. Dorsal to this ridge is a trough and further dorsal to that the bone curve outwards laterally. Finally, there is a large convex element in MAB16121b (Fig. 14A3) with numerous small teeth, resembling the interior teeth of the mandible. This is most likely the ectopterygoid bone, which makes up the base of the orbit and is covered with small teeth in Dercetis triqueter.

Fig. 14. Dercetid fish Dercetis triqueter (Pictet, 1850) (MAB16121a, b) from the Maastrichtian (Upper Cretaceous) of Eben- Emael, Belgium. Complete specimen (A1), rostrum, alongside isolated vertebrae and scutes (A2,A3).
NHMM 2025 017 preserves the mesethmoid, which narrows markedly towards the anterior and carries a characteristic bifurcated articulation point anteriorly that connected to the sharp premaxillae (Fig. 16). Its surface is ornamented with very minute longitudinal striations. It also preserves the anterior end of the right? frontal, which has an elevated platform carrying striations. Also preserved are various portions of the dentary, judging from their thin nature and the alveolar scars indicating two different types of teeth (Fig. 16). Several teeth are visible, although none are attached to the rostrum. Teeth are generally sharp, thin and curve backwards.
Axial skeleton: Twenty to thirty vertebrae of the abdominal region are preserved in two articulated rows in MAB16758 (Fig. 12A1, A4). It appears that the vertebral column of this fish became folded over, because the two pieces of the column are not antero-posteriorly aligned. Little of the neural arches is preserved in MAB16758 (Fig. 12A4), but the vertebrae in MAB16120 do show a prominent neural arch that covers the length of the vertebral centre and carries a thin, diagonally pointed neural spine posteriorly (Fig. 13A7). Each vertebral haemal arch is as long as its corresponding centrum, the remnants of which show that the vertebrae bore just one pair of haemophyses near the posterior side (Figs. 12A4, 13A7). In addition, a few thin intermuscular bones and ribs are preserved in MAB16758 (Fig. 12A1, A4), although they do not show any distinguishable morphological traits. In specimen NHMM 2025 017 about 10–12 vertebrae are preserved in anatomical alignment (Fig. 16). Their right lateral view is exposed, which is evident from the location and direction of the anterior-pointing haemophysis. Most of the haemophysis have been lost, but their remnants are visible, as are the haemal and neural arches.
Scutes: Triradiate scutes are arranged in a single line on both sides of the body. In MAB16758 (Fig. 12A1, A4), seven of these scutes are preserved ventral to the vertebral column. Most of these scutes have their medial side exposed, evidenced by their surfaces being hollow and not bearing any ridges. This indicates that this row of scutes originated in the lateral right side of the body. The anterior limb varies in size along the line, ranging from roughly equal in length to the posterior limbs to being much shorter than them, seemingly in no particular pattern. One scute is flipped over, showing its lateral surface (Fig. 12A3). This scute, and several of the scutes in MAB16120 (Fig. 13A7), show a marked ridge running along the dorsal margin of the anterior and ventral posterior limbs of the scutes. The posterior limbs of the scutes are asymmetrical, with the ventral limb being larger and tapering downwards. Scutes in NHMM 2025 017 are notably thicker, considering their size, than in other specimens, but share the same triradiate morphology with a ridge (Fig. 16).

Fig. 15. Dercetid fish Dercetis triqueter (Pictet, 1850) and Ophidercetis? sp. (MAB16759) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Specimen contains a mix of scutes, rostral remains and vertebrae, belonging to at least two different individuals.

Fig. 16. Dercetid fish Dercetis triqueter (NHMM 2025 017), from the Maastrichtian (Upper Cretaceous) of Eben- Emael, Belgium. Specimen contains a largely intact rostrum and partially intact vertebral column.
Remarks.—MAB16758 is identified as Dercetis triqueter based on scute morphology (i.e., no ornament other than a ridge on the dorsal margin) and vertebral morphology (i.e., first vertebrae twice as long as deep). The back of the cranium in MAB16758 (Fig. 12A2) closely resembles that of Dercetis elongatus (Agassiz, 1834), as illustrated by Taverne (2005a). The dorsal side of the skull of Dercetis triqueter has not yet been described at length, but the present material shows that it closely resembles Dercetis elongatus.
MAB16120 is identified as Dercetis triqueter based on scute and rostral morphology (mandible bearing two distinct types of teeth in lines next to one another). The nature of the striated element in MAB16120c (Fig. 13A5) is unknown as there are no reports of striated elements in the cranium outside of the frontals, but these do not narrow so rapidly across their length. It is here preliminarily identified as a branchiostegal ray, which had not yet described for Dercetis triqueter (compare Taverne 2005a).
MAB16121 is referred to Dercetis triqueter on account of rostral and frontal morphology. It also preserves teeth (Fig. 13A2), the larger of which resemble those illustrated by Taverne and Goolaerts (2015: figs. 3, 5). In general, this specimen expresses well the complex morphology and dentition of the rostrum in species of Dercetis, as opposed to those of other, toothless genera from the Maastrichtian type area, such as Beukidercetis gen nov. and Cyranichthys (compare Figs. 6 and 7; Taverne 1987; Taverne and Goolaerts 2015; Wallaard et al. 2019).
MAB16759 consists of several individuals. Parts identified without a doubt as pertaining to Dercetis triqueter are several unornamented triradiate scutes, a flat, triangular left mandible, and a vertebra that bears a single pair of haemophyses which emanate from the center of the vertebrae. It is unclear how many other individuals are present in the material, but a much smaller right mandible with sharp, tightly packed dentition and a vertebra bearing two pairs of fused? haemophyses that do not belong to Dercetis triqueter and resemble those of Ophidercetis italiensis (see Taverne 2005a).
Specimen NHMM 2025 017 is identified as Dercetis triqueter based on the shape of the mandibles, forked mesethmoid, triradiate, unornamented scutes and vertebrae bearing a singular anterior-facing haemophysis (Fig. 15). Interestingly, the rostrum in NHMM 2025 017 shows a closely similar pattern of preservation to an individual of Dercetis triqueter discussed above, namely NHMM K 565 (see Taverne and Goolaerts 2015: figs. 3, 5).
Stratigraphic and geographic range.—Santonian of Lebanon, Campanian–Maastrichtian of Italy. Maastrichtian of Belgium and the Netherlands, all Upper Cretaceous.
Dercetis sp.
Fig. 17.
Material.—MAB16760, from the Maastrichtian, Upper Cretaceous, Marnebel quarry, Eben-Emael, Belgium (found by JJWW), representing an isolated vertebra. Due to its incredibly fragile nature, the specimen was only partially prepared and quickly solidified, leaving it only partially exposed.
Description.—Axial skeleton: Isolated vertebra eroded, only preserving the lateral face of a fairly broad neural and the start of a singular thin, flat haemophysis that originates from the vertebral centre. It is three times longer than high, and fairly narrow.

Fig. 17. Dercetid fish Dercetis sp. (MAB 16760) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Specimen represents an isolated vertebra with a partially preserved haemophysis.
Remarks.—MAB16760 undeniably is a dercetid based on its vertebral morphology and is here assigned to the genus Dercetis, on account of the singular flat haemophysis emanating from the centre. It is not possible to identify MAB16760 at the species level, as apomorphies that distinguish Dercetis triqueter and Dercetis elongatus lie in the rostrum and scutes, which are not preserved here (Taverne 2005).
Genus Dercetoides Chalifa, 1989
Type species: Dercetoides venator Chalifa, 1989; Cenomanian of Ein- Yabrud, Israel.
Stratigraphic and geographic range.—Cenomanian of Ein-Yabrud, Israel. Maastrichtian of Belgium. Both Upper Cretaceous.
Dercetoides venator Chalifa, 1989
Fig. 18.
1989 Dercetoides venator sp. nov.; Chalifa 1989: 320, figs. 6–11.
Material.—MAB16117 from Maastrichtian, Upper Cretaceous, CBR-Romontbos quarry, Eben-Emael, Belgium (found by MdR), consisting of two roughly equal-sized chunks of sediment, which contain remains of two cranial bones, some vertebrae and several isolated scutes of two different morphologies.
Description.—Cranium: Right hyomandibular is preserved (Fig. 18A2), identified by its general shape and the location of the ridge running along its antero-posterior axis. The hyomandibular is broader dorsally and narrower at its ventral end. A tall ridge runs roughly along the centre of this bone, which tapers ventrally on the posterior side and develops into a broader fold there. There is another, smooth and triangular bone that has a hollow interior and pointed tip (Fig. 18A3). It is relatively featureless, but does resemble the interoperculum as depicted by Chalifa (1989) in relative shape and size, when compared with the hyomandibular in the same specimen.

Fig. 18. Dercetid fish Dercetoides venator (Chalifa, 1989) (MAB16117) from the Maastricthian (Upper Cretaceous) of Eben-Emael, Belgium. Complete specimen (A1), hyomandibular and two isolated vertebrae (A2), interoperculum (A3), two morphotypes of scutes (A4, A5).
Axial skeleton: A few precaudal vertebrae are preserved in MAB16117 (Fig. 18A2), which show that the vertebral centres are twice as long as deep. These vertebrae have prominent neural arches that cover the length of the entire vertebral centre and carry a short, sharp neural spine posteriorly. The vertebrae possess a single transverse ridge on their dorsal half, that is up to twice the width of the vertebral centre. No haemophyses can be seen, possibly linked to the state of preservation of the specimen.
Scutes: Two morphotypes of scutes are preserved in MAB16117 (Fig. 18A4, A5). The first is thin, triradiate with asymmetrical posterior limbs and a ridge running along the dorsal margin of the anterior limb (Fig. 18A4). The second has a more lanceolate morphology, with symmetrical posterior limbs (Fig. 18A5). The anterior limb of these scutes carries a tall and sharp ridge along the midline, which surpasses the bifurcation of the posterior limbs and likely overlapped with the anterior limb of the successive scute.
Remarks.—The cranial remains in MAB16117 (Fig. 18A2, A4) closely resemble those of Dercetoides venator as described by Chalifa (1989). In addition, the vertebral morphology, although not completely preserved, is also comparable to that of Dercetoides venator in having short and sharp neural spines and a pair of flattened transverse processes on either side. There are two types of scutes in MAB16117: the first are lanceolate (Fig. 18A5) and resemble those of Dercetoides venator (compare Chalifa 1989), yet appear to lack a perforation for the lateral line canal; however, only one of these scutes has its anterior limb properly exposed in MAB16117. The second type is triradiate (Fig. 18A4) and resembles scutes of Dercetis, but are notably quite sharp and thin when compared to specimens of Dercetis triqueter from the Maastrichtian type area (see Figs. 11, 13; compare Taverne and Goolaerts 2015). It is likely that this second morphotype of scutes originated from an area of the body other than the lateral sensory line.
Stratigraphic and geographic range.—Cenomanian of Ein-Yabrud, Israel. Maastrichtian of Belgium. Both Upper Cretaceous.
Genus Ophidercetis Taverne, 2005
Type species: Ophidercetis italiensis Taverne, 2005, Campanian–Maastrichtian, Nardò, Italy.
Stratigraphic and geographic range.—Campanian–Maastrichtian of Belgium, Italy, and the Netherlands.
Ophidercetis italiensis Taverne, 2005
Figs. 19, 20.
2005 Ophidercetis italiensis sp nov.; Taverne 2005a: 57, figs. 1–8.
2015 Ophidercetis italiensis Taverne, 2005; Taverne and Goolaerts 2015: 23, figs. 8–10.
Material.—NHMMJJ 15235 (Fig. 19), from the Maastrichtian, Upper Cretaceous, Valkenburg Member, former ENCI-HeidelbergCement Group quarry, Maastricht (the Netherlands), represents a partially articulated individual with cranial and rostral remains and several vertebrae preserved in articulation. The cranial and rostral remains were partially fractured and displaced during fossilisation.
NHMMJJ 16217 (Fig. 20A), from the Maastricthian, Upper Cretaceous, Emael Member, former ENCI-HeidelbergCement Group quarry, Maastricht (the Netherlands), is a disarticulated individual with several vertebrae and scutes exposed. MAB16119 (Fig. 20B), from the Maastricthian, Upper Cretaceous, CBR-Romontbos quarry, Eben-Emael, Belgium (found by JJWW), is a single isolated scute.
Description.—Cranium: Only frontals and a parietal? are preserved in NHMMJJ 1235a (Fig. 19A2). The right frontal is intact, while the left one is fractured. The frontal has a broad, largely flat surface, with a raised surface dorso-medially. This surface elongates into a rod-like extension, ornamented with longitudinal striations that radiate posteriorly. The lateral side of the frontal shows a depression, which forms the articulation surface with the wedge-like sphenoid. Additionally, a small, convex bone is preserved, which could be a parietal, judging from its relative size, shape and position to the frontal, but the bone itself is relatively featureless (Fig. 19A2).

Fig. 19. Dercetid fish Ophidercetis italiensis (Taverne, 2005) (NHMMJJ 15235) from the Maastrichtian (Upper Cretaceous) of Maastricht, the Netherlands. Complete specimen (A1), partial cranium and rostrum (A2), partial vertebral column (A3).
Rostrum: Only the premaxilla is exposed in NHMMJJ 15235a (Fig. 19A2). The dorsal surface is exposed, showing a rounded surface, with very minute longitudinal striations across its length. The premaxilla remains separate across its entire length here, only remaining fused at the very anterior end. Anteriorly, it ends in a symphysial process, which has a knob-like structure, but is not fully preserved in NHMMJJ 15235a.
Axial skeleton: In the axial skeleton vertebrae carry a short neural arch that covers the entire length of the vertebral centre (Fig. 19A3). It is not possible to tell from the present material whether or not neural spines were present. The vertebrae carry two fused pairs of haemophyses (Figs. 19A3, 20A2), which are directed ventro-laterally and have a rounded appearance. The fusion between these two haemophyses is lined with two parallel ridges on the dorsal surface of the structure.
Scutes: The large (>5 mm in length) lanceolate scutes show a fairly tall median ridge that bifurcates posteriorly and bears a sharp, upright extension on the bifurcation point (Fig. 20A2 B). The anterior surface of the scutes is ornamented with numerous small pits. The mineralised tissues posterior to the bifurcation are thin and smooth.

Fig. 20. Dercetid fish Ophidercetis italiensis (Taverne, 2005) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium (A), and Maastricht, the Netherlands (B). A. NHMMJJ 16217, general view (A1), isolated scutes and vertebrae (A2). B. MAB16119, scute.
Remarks.—NHMMJJ 15235 (Fig. 19A3) preserves no scutes, and only partially retains the vertebrae. However, the clearly recognisable morphology of the frontal and partial remains of the vertebrae align with the material of Ophidercetis italiensis as presented by Taverne (2005a). Scutes and vertebrae in NHMMJJ 16217 are severely eroded (Fig. 20A2), but the large, round haemophyses and defined pits on the scutes allow for a confident identification. The isolated scute in MAB16119 also strongly resembles the scutes of O. italiensis (see Taverne 2005a: fig. 8).
Stratigraphic and geographic range.—Campanian–Maastrichtian of Italy. Maastricthian of Belgium and the Netherlands. All Upper Cretaceous.
Genus Petrodercetis nov.
Zoobank LCID: urn:lsid:zoobank.org:act:D1A744B7-D6E2-40D2-A 445-3078B0B1B46B.
Etymology: Dedicated to Saint Peter (Petrus), one of the twelve apostles of Jesus Christ, after whom the “Sint-Pietersberg”, which is home to several of the quarries in the Maastrichtian type area, was named.
Type species: Petrodercetis bidirectus sp. nov., monotypic; see below.
Diagnosis.—Same as for the monotypic type species.
Stratigraphic and geographic range.—Upper Maastrichtian, the Netherlands.
Petrodercetis bidirectus sp. nov.
Fig. 21.
Zoobank LCID: urn:lsid:zoobank.org:act:27A304B5-869F-434B-B1F6- ED053CC5F184.
Etymology: Referring to the bidirectional extension of the median ridge on the scutes.
Holotype: NHMM JJ 16219, a disarticulated individual with rostral remains, vertebrae and scutes.
Type locality: Former ENCI-HeidelbergCement Group quarry, Sint-Pietersberg, Maastricht, the Netherlands.
Type horizon: Emael Member (Maastricht Formation), upper Maastrichtian.
Material.—The holotype only.
Diagnosis.—Curved frontal which ends in an arched platform. Premaxilla forms a hollow interior surface, exterior surface is smooth and rounded. Unfused triangular mandible that bears a single row of small teeth on its lateral side. Vertebrae with two pairs of transverse processes: an anterior rod-shaped process and a short posterior flattened one. Cordiform scutes that are broader than long, midline that extends both anteriorly and posteriorly, surface ornamented with ridges and tubercles.
Description.—Cranium: A bone which appears to be the left frontal is preserved (Fig. 21A2), having an elongate, flattened shape, which curves posteriorly into an arched shelf. There are a few other partial cranial bones, although they are not complete enough to be identified.

Fig. 21. A. Dercetid fish Petrodercetis bidirectus gen. et sp. nov., NHMM JJ 16219 (holotype) from the Maastrichtian (Upper Cretaceous) of Maastricht, the Netherlands. Complete specimen (A1), rostrum (A2), vertebrae (A3), scutes (A4), schematic drawing of scute morphology (A5), based on scute in A4. B. Schematic drawing of P. bidirectus (sacral?) vertebral morphology (dorsal view), based on several different vertebrae (arrow indicates anterior side). Grey infills represent ridges and tubercles. C. Schematic figure of how the scutes of P. bidirectus might have been arranged in life: either with body space in between them (C1) or in diagonal lines along the body (C2).
Rostrum: Several pieces of the rostrum are preserved in NHMM JJ 16219 (Fig. 21A2). The right? premaxilla has its dorsal side exposed. It has a rounded shape dorso-medially, and a smooth surface, bearing no striations. The lateral side extends into a shelf, which has a convex internal surface. It is not possible to determine whether or not the premaxilla bore teeth. The other half of the premaxilla is not visible. The right mandible has its dorsal side exposed, and the left mandible has flipped over and has its ventral side exposed. The unfused mandible is flat and triangular in shape and ends in a pointed extension anteriorly. The lateral side of the mandible is lined with minute alveoli, showing it to have carried a single row of small teeth. The latero-medial surface is smooth, bearing no teeth. There is another elongate bone pertaining to the rostrum, although its nature in unclear. It is flattened and has a tall, twisted posterior end. It appears to have minute alveoli on its medial surface. This element could represent the maxilla, but its anterior end is not preserved.
Axial skeleton: Some smaller vertebrae are preserved in NHMM JJ 16219 (Fig. 21A3). These bear two pairs of transverse processes, the anterior being long and rod-like, and the posterior short, sharp and flattened. There is a second morphotype of vertebrae (Fig. 21A3), which is slightly larger than the first. These vertebrae are about as broad as long. Their surface is penetrated by a vascular system. The haemal arch is strongly reduced, but the vertebrae do bear two large, rounded haemophyses, which are not fully preserved in any of the vertebrae exposed in NHMM JJ 16219. This second type of vertebrae could have originated in the caudal region.
Scutes: Several large (around 5 mm long), broad (around 7 mm wide), cordiform scutes are preserved (Fig. 21A2). These scutes carry a median ridge that increases in height posteriorly and ends in a pointed extension posteriorly. In some scutes, this extension also features an anteriorly directed, pointed extension. The surface of the scutes is ornamented with numerous pits and striations, radiating semi-randomly from the midline (Fig. 21A4).
Remarks.—The axial skeleton in the present form shows different morphotypes (Fig. 21A3, B). This has also been recorded from other dercetids, such as Benthesikyme armatus, Caudadercetis bannikovi, Dercetoides venator, and Hastichthys gracilis (Chalifa 1989; Taverne 2005a, 2006b). However, the notable size difference between the two types of vertebrae is very unusual, especially for a dercetid. It is for this reason that these vertebrae were not considered when formulating the generic and specific diagnoses.
Scute morphology of NHMMJJ 16219 is unique and differs utterly from all species of Dercetidae described before, especially in having a bidirectional extension of the ridge on the midline (Fig. 21A4, A5). This, alongside a unique vertebral morphology, justifies the erection of a new genus. The rostrum in NHMM JJ 16219 bears a few similarities to that of Dercetis triqueter, but its mandibles have just one row of teeth, and the premaxilla is less triangular (Taverne 2005a).
The morphology of the scutes would seem to counter identification as a dercetid, considering that their scutes are usually interpreted to be arranged in straight lines along the body (Taverne and Goolaerts 2015). For such to be the case in Petrodercetis gen. nov., with pointed extensions of the scute midline both anteriorly and posteriorly, there would either have to be more body space between the scutes in life (Fig. 21C1), or these scutes would have had to be slightly angled, creating diagonal lines along the body (Fig. 21C2).
Stratigraphic and geographic range.—Maastrichtian (Upper Cretaceous) of the Netherlands.
Dercetidae gen. et sp. indet. 1
Fig. 22.
Material.—MAB17079, from the Maastrichtian, Upper Cretaceous, Marnebel quarry, Eben-Emael, Belgium (found by Maria Verschelling-Wallaard), comprises several pieces of the cranium, isolated and fragmentary vertebrae and isolated scute fragments.
Description.—Cranium: MAB17079 preserves a few indeterminate cranial bones. In addition, a partial parietal? is retained, with a shallow ridge running along the longitudinal axis (Fig. 22A2).
Aaxial skeleton: MAB17079 retains a few heavily fractured vertebrae (Fig. 22A3). Vertebrae are particularly narrow. Most of the vertebral processes have been lost, but visible are a rounded neural arch and the onset of thin and flat haemophyses on one side of the vertebra.

Fig. 22. Dercetidae gen. et sp. indet. 1 (MAB17079) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Complete specimen (A1), parietal? (A2), isolated vertebra (A3), fragmentary scutes (A4).
Scutes: MAB17079 retains several fragments of isolated scutes (Fig. 22A4). These are fairly large (>10 mm in length) and convex, bearing minute tubercles, but not enough scute surface is preserved to determine any patterning. There also appears to be a ridge running along the midline in one of these scutes, but this is not entirely clear.
Remarks.—MAB17079 is identified as a dercetid based on its cordiform scutes and narrow vertebrae. However, preservation is insufficient, both in quality and number of skeletal elements, to make identification at the generic level possible. Based on the fragmentary scutes (Fig. 22A4), it most closely resembles either species of Caudadercetis or of Cyranichthys, in view of the larger scutes and tubercular ornamentation.
Dercetidae gen. et sp. indet. 2
Fig. 23.
Material.—MAB16118, from the Maastrichtian, Upper Cretaceous, CBR-Romontbos quarry, Eben-Emael, Belgium (found by JJWW), comprises several chunks of biocalcarenitic rock containing several cranial/rostral bones.
Description.—Cranium: The left operculum is preserved in MAB16118 (Fig. 23A3). It is a thin, flat bone, with a roughly rounded shape. The anterior edge is straight and accentuated by a rounded ridge, from which minor striations radiate laterally. The most ventral striations appear to curve ventrally, although not all of the operculum is preserved in this position. There are also some disarticulated pieces of what appear to be branchiostegal rays (Fig. 23A4). Their length is roughly equal to that of the operculum, although their morphology is difficult to determine. Finally, there are some lateral cranial remains (Fig. 23A2). Both elements have a convex shape, but are difficult to identify for certain. The first has a rounded, smooth surface, while the other has a more elongate shape and is perforated by a sensory canal along its length. Potentially, this could be the pterotic, containing the otic sensory canal.
Rostrum: A partial, heavily disarticulated premaxilla? is preserved (Fig. 23A5). It has an elongate shape and is ornamented with minor longitudinal striations, that increase markedly in height posteriorly. A single, conical tooth (Fig. 23A6) is retained, showing a laterally flattened morphology. It is unknown from where in the rostrum this tooth originated.

Fig. 23. Dercetidae gen. et sp. indet. 2 (MAB16118) from the Maastrichtian (Upper Cretaceous) of Eben-Emael, Belgium. Complete specimen (A1), lateral side of cranium (A2), left operculum (A3), potential branchiostegal ray (A4), rostrum (A5), isolated tooth (A6).
Remarks.—MAB16118 undoubtedly represents a dercetid, based on the elongate, striated rostrum (Fig. 23A5). Due to its poor preservation, it is not possible to make any definitive conclusions about its generic and specific identity. However, the morphology of the operculum (Fig. 10A3), with its triangular shape and radiating striations, resembles that of Cyranichthys ornatissimus. Cyranichthys ornatissimus was first described from the Cenomanian of Congo by Casier (1965), as Dercetis ornatissimus, later redescribed by Taverne (1987). Both authors published based on very incomplete remains and thus, no detailed comparison can be made with MAB16118.
The laterally flattened tooth (Fig. 23A6) in MAB16118 represents an interesting curiosity. Its morphology distinguishes this particular find from most of the species previously described from the Maastrichtian type area. The rostrum of Ophidercetis italiensis (Taverne, 2005a) has more pointed, conical teeth (Taverne 2005a), while two distinct types of teeth have been described for Dercetis triqueter; both differ significantly from the tooth in MAB16118 (Figs. 13A7, 14A2, A3; compare Taverne 2005a; Taverne and Goolaerts 2015). Cyranichthys jagti, Cyranichthys sideralis sp. nov. and species of Beukidercetis gen. nov. have toothless rostra (Figs. 5–10; Taverne and Goolaerts 2015; Wallaard et al. 2019). Thus, it is highly likely that MAB 16118 represents a different species. However, it should be mentioned in this respect that, to date, there are no specimens of Apuliadercetis indeherbergei (see Taverne and Goolaerts 2015) known that preserve the rostrum and that tooth morphology in Petrodercetis bidirectus gen. et sp. nov. (Fig. 21) is not known either. For now, the present specimen will have to remain in open nomenclature.
Dercetidae from directly above
the Lichtenberg Horizon
In addition to specimens mentioned above, we here also record an assemblage of dercetid scutes, collected by Ken Cardon from the fossil hash directly overlying the Lichtenberg Horizon (basal Valkenburg Member, Maastricht Formation) at the former ENCI-HeidelbergCement Group quarry (Maastricht, the Netherlands). Scutes (fragments) are identified as dercetids on the basis of tubercles and ridges and their relative sturdiness. It is difficult to assign many of the fragmentary scutes to genera and/or species, but four forms are recognised in this assemblage (Fig. 24).
Of Caudadercetis taverni sp. nov. there is a scute fragment that bears a midline which ends in a sharp extension, and has a surface ornamented with lines, radiating from the midline, and composed of fused tubercles (Fig. 24A). Cyranichthys sideralis sp. nov. is recognised on the basis of a convex scute fragment that carries a rounded midline ridge, surrounded by semi-randomly arranged tubercles (Fig. 24B). To Dercetis triqueter a posterior fragment of a scute from the right flank is referred; this has a triradiate shape, although its anterior limb is lost (Fig. 24C). The surface is smooth, ornamented merely by a sharp ridge that bifurcates posteriorly. Finally, of Ophidercetis italiensis there is a more triangular scute fragment, with a sharp anterior edge, a midline that ends in a small extension and then bifurcates posteriorly, and a surface ornamented with numerous regularly spaced pits (Fig. 24D).

Fig. 24. Isolated dercetid fish scutes, collected from directly above the Maastrichtian (Upper Cretaceous) Lichtenberg Horizon (basal Valkenburg Member) of the former ENCI-HeidelbergCement Group quarry (Maastricht, the Netherlands). A. Caudadercetis taverni sp. nov. (MAB 18273). B. Cyranichthys sideralis sp. nov. (MAB 18272). C. Dercetis triqueter (Pictet, 1850) (MAB 18274). D. Ophidercetis italiensis (Taverne, 2005) (MAB 18271).
Discussion
Dercetids in the Maastrichtian type area are occasionally difficult to identify below the generic level, because the remains usually are preserved disarticulated and fractured. This explains why only few cranial and vertebral apomorphies used to describe species are often undetectable in specimens, making scute morphology the most reliable way to identify specimens. Despite these constraints, it is clear there was a great, previously underappreciated dercetid diversity in the study area. The material described here includes more specimens of three previously described taxa (Cyranichthys jagti, Dercetis triqueter, and Ophidercetis italiensis; see Figs. 6, 7, 12–16, 19, 20), one species which was not previously known from the Maastrichtian type area (Dercetoides venator; Fig. 18), while two genera and four new species are erected herein, namely Caudadercetis taverni sp. nov., Beukidercetis lissus gen. et sp. nov., Cyranichthys sideralis sp. nov., and Petrodercetis bidirectus gen. et sp. nov. (see Figs. 4, 5, 8, 9, 10, 21).
This material clearly underscores the diversity of the genus Cyranichthys in the Maastrichtian type area. This genus was first described from the Cenomanian of Congo based on poorly preserved material (Casier 1965; Taverne 1987). Prior to this study, only Cyranichthys jagti had been recognised from the Maastrichtian type area. This species differs from type species, Cyranichthys ornatissimus, mainly in scute ornamentation and the shape of its operculum (Taverne 1987; Taverne and Goolaerts 2015). Here, more material of Cyranichthys jagti is described, which affirms claims made by Taverne and Goolaerts (2015), and that provides additional diagnostic characters, such as the toothless triangular mandible and paired vertebral haemophyses (Figs. 6, 7). In addition, Cyranichthys sideralis sp. nov. is erected, based on a specimen with a markedly striated premaxilla and defined, jet plane-like scutes (Fig. 6). Finally, an indeterminate dercetid with a triangular operculum that closely resembles that of Cyranichthys ornatissimus is recorded herein (Fig. 23). This specimen also includes an isolated tooth, whereas all other records of Cyranichthys from the Maastrichtian type area have been consistently shown to be toothless (Figs. 6–9; compare Taverne and Goolaerts 2015). Taken together, these findings highlight the high diversity within the genus Cyranichthys in the Maastrichtian type area. However, more data are needed to identify clear plesio- and apomorphies for the three species now assigned to the genus.
There is a wide range in dercetid scute morphology in the material from the Maastrichtian type area (Figs. 4A1, A2, A7, 5A3, B, 6A5, 8A4, A5, B2, 11A4, A5, 12A3, A4, 13A7, 16, 18A4, A5, 20A2, B, 21A4, 24; compare Taverne and Goolaerts 2015; Wallaard et al. 2019), ranging from triradiate scutes (Apuliadercetis indeherbergei, Dercetis triqueter, and Dercetoides venator), via cordiform ones (Caudadercetis taverni sp. nov., Cyranichthys jagti, Petrodercetis bidirectus gen. et sp. nov., and Beukidercetis grandis), lanceolate ones (Ophidercetis italiensis) and even jet plane-shaped ones recorded here (Cyranichthys sideralis sp. nov.). Species are fairly readily distinguished on the basis of ornamentation, including pits, tubercles and ridges, which may or may not bifurcate and/or end in pointed extensions (Fig. 3). The new taxa introduced here, were erected mainly based on scute morphology and ornamentation. An updated view of dercetid scute diversity, including taxa introduced herein, is provided in Fig. 25.

Fig. 25. Schematic drawings of scutes for all the dercetid species known to date from the Maastrichtian type area. Grey infills represent convex surfaces (e.g., ridges/tubercles), black infills represent concavities (e.g., pits/canals). Modified from Chalifa (1989: fig. 11), Taverne (2005a: fig. 8), Taverne (2005b: fig. 9), Taverne (2006a: fig. 11), Taverne and Goolaerts (2015: figs. 7, 14, 22) and Wallaard et al. (2019: fig. 5). Not to scale.
Dercetid scutes from the Maastrichtian type area are often preserved in isolation, outside of anatomical alignment. Current reconstructions place these characteristic scutes in rows along the body, often associated with the lateral line (Davis 1890; Chalifa 1989; Taverne 2005b; Figueiredo and Gallo 2006; Taverne and Goolaerts 2015; Alvarado-Ortega and Díaz-Cruz 2021; Díaz-Cruz et al. 2022). However, the material of Cyranichthys sideralis sp. nov. (Figs. 8–10), Dercetoides venator (Fig. 18), Petrodercetis bidirectus gen. et. sp. nov. (Fig. 21) described herein, suggests there may be more complex patterns of dermal covering on the body of dercetids. Examples are scutes in arched lines along the body as in Petrodercetis bidirectus gen. et sp. nov., or variation in scute type along the body in Cyranichthys sideralis sp. nov. and Dercetoides venator.
It is difficult to draw any palaeoecological conclusions based on these scutes. However, genera with larger, cordiform scutes such as Caudadercetis, Cyranichthys, Ophidercetis, and Beukidercetis gen. nov. are likely to have had a much higher degree of dermal coverage and thus, a tougher armour, than genera with thin scutes such as Apuliadercetis, Dercetis, and Dercetoides (Fig. 3). This artificial grouping does become more apparent, when considering recent reports that suggest there are no synapomorphies to define the family Dercetidae (Alvarado-Ortega and Díaz-Cruz 2021; Chida et al. 2023). An extensive taxonomic reassessment of this family as a whole is needed before any further claims can be made.
There is also great variation in the dentition (or rather lack thereof) of dercetid species found in the Maastrichtian type area, although only a few specimens actually preserve teeth. All species of Cyranichthys and Beukidercetis gen. nov. with known rostral elements are edentulous (compare Taverne and Goolaerts 2015; Wallaard et al. 2019), while Dercetis triqueter bears two types of teeth: an exterior row of sharp, irregularly spaced, and curved teeth, and several rows of interior, short and stumpy teeth. Finally, Ophidercetis has tightly packed teeth arranged in several rows along the rostrum (Taverne 2005a). There are no specimens of Apuliadercetis indeherbergei with rostral preservation, but reports from a congener, A. gonzalezae, suggest that it might have had small conical teeth, tightly spaced in several rows (Díaz-Cruz et al. 2022). Isolated dercetid teeth, such as the ones seen in MAB16118 (Fig. 23A6), are unreliable indicators, however. Tooth morphology appears to differ even within species, with examples for Dercetis triqueter and Ophidercetis italiensis (Taverne 2005b; Taverne and Goolaerts 2015).
The material discussed herein contributes to a refinement of the stratigraphical ranges of dercetid fish from the Maastrichtian type area (Fig. 25). Firstly, the temporal distribution of Dercetis triqueter and Ophidercetis italiensis are expanded, with the former now documented from the entire Maastricht Formation (Valkenburg up to and including Meerssen members), and the latter from the Valkenburg to Emael members of this formation (Taverne and Goolaerts 2015). New material of Cyranichthys jagti extends its range downwards into the uppermost Gulpen Formation (Lanaye Member). However, the stratigraphical range of Cyranichthys jagti in the Maastricht Formation has become less clear with the transfer of NHMM K 2372 (Fig. 5B) to Caudadercetis taverni sp. nov. herein.
The holotype of Cyranichthys jagti, NHMM 1993 119, is from an unidentified level within the lower Maastricht Formation (Taverne and Goolaerts 2015). In addition, material of Cyranichthys sideralis sp. nov. (Figs. 8–10) suggests that two species of Cyranichthys co-existed in the Maastrichtian type area. Finally, a specimen of Dercetoides venator (Fig. 18) presents the second ever record of a species previously known only from a single individual from the Cenomanian of Israel (Chalifa 1989). This find greatly increases the spatial and temporal distribution of the species. It also creates a large gap in its distribution, which cannot yet be explained.
The present records from the fossil hash overlying the Lichtenberg Horizon (Fig. 24) document that dercetid assemblages persisted across the boundary level between the Gulpen and Maastricht formations, which constitutes a stratigraphical hiatus (Vellekoop et al. 2022). This dercetid assemblage, while difficult to determine at generic or specific level, extends the stratigraphical ranges of several of the taxa.
Dercetids constitute the sole actinopterygian family in the Maastrichtian type area of which semi-complete remains have so far been collected (Friedman 2012; Taverne and Goolaerts 2015; Wallaard et al. 2019). Most other forms, such as the aulopiform Enchodus, are almost exclusively known from isolated remains, generally isolated teeth, anterior fangs and partial jaws (Friedman 2012; Kovalchuk et al. 2022). Other areas that have yielded dercetid fossils in similar Maastrichtian chalk deposits, such as Nardò (Italy) or Eurytania and Gavdos Island (Greece), do not show a pattern of Dercetidae having a great potential of preservation in comparison to other actinopterygians (Sorbini and Bannikov 1991; Taverne 2005a, b, 2006a, b; Mayrinck et al. 2015; Argyriou and Davesne 2015; Argyriou et al. 2022). However, despite a higher percentage of preservation, articulated remains of dercetids from the Maastrichtian type area are rare, as is clear from the material presented herein. The disarticulated state of most of the material is one of the reasons why specimens of Dercetidae from the Maastrichtian type area can be erroneously identified as the burrow trace fossil Lepidenteron lewesiensis.
There are several possible explanations for the high abundance and preservation of dercetid fossils compared to other actinopterygians in the Maastrichtian type area. First, dercetids in the area could have exhibited a more purely benthic or burrowing lifestyle. This would have resulted in a better chance at fossilisation as the area was a shallow, high-energy environment in which the top layers of the sediment were likely highly perturbed (Jagt and Jagt-Yazykova 2012; Vellekoop et al. 2022). There are few descriptions of the palaeoecology of dercetid fish, except for the notion that they are predatory species (Chalifa 1989). The elongate body plan of dercetids is usually characteristic of species with an “ambush predator” lifestyle, such as modern eels, which hide in burrows or holes and lash out to capture their prey (Maxwell and Wilson 2013; Catania 2019). However, unlike modern eels, dercetids had a tough dermal body coverage, reducing their overall flexibility. This makes a burrowing life-style unlikely, when opposed to hiding in pre-existing burrows. Secondly, it might be possible that dercetids were less easy to digest for local predators, due to their slender bodies, a relatively high degree of scale coverage and characteristic, tough scutes (Taverne 2005a; Taverne and Goolaerts 2015; Wallaard et al. 2019). The Maastrichtian type area featured a wide range of nektic predators, including sharks, sea turtles, mosasaurs and plesiosaurs (Mulder 2003; Jagt et al. 2008; Van Baal et al. 2013; Mulder et al. 2016; Miedema et al. 2019; Donovan and Jagt 2020). If indeed dercetids had tougher bodies that were less digestible compared to other actinopterygian fishes in the area, they could have stood a better chance of being fossilised more completely, supported by the fact that dermal remains are some of the best-preserved parts of dercetids in the area.
Overall, the present study has highlighted an underappreciated diversity of Dercetidae in the Maastrichtian type area, with the erection of two new genera and four new species, and documentation of a higher degree of preservation potential in comparison to other actinopterygians found in the study area. Several species of Dercetidae co-existed in the Maastrichtian type area during the Late Cretaceous. To date, no Dercetidae have been recorded from the Danian of the Maastrichtian type area, but in similar strata in New Jersey, dercetids persisted across the K/Pg boundary into the Danian (Boles et al. 2024). Thus, it remains possible that one day they will be found in the Geulhem Member (Houthem Formation) of early and middle Danian age in the Maastrichtian type area. It is highly likely that more specimens of Dercetidae exist in private and institutional collections alike, potentially labelled as indeterminate fish remains or worm burrows. Furthermore, these findings, combined with extensive reports from Nardò in Italy (Taverne 2005a, b, 2006a, b), suggest that similar Maastrichtian deposits such as those in Greece and Poland could prove to have an equally high diversity of Dercetidae (Bieńkowska-Wasiluk et al. 2015; Argyriou and Davesne 2021; Argyriou et al. 2022). It is therefore of importance that this group receive more attention in order to discern more about their ecology and understand more about their preservation.
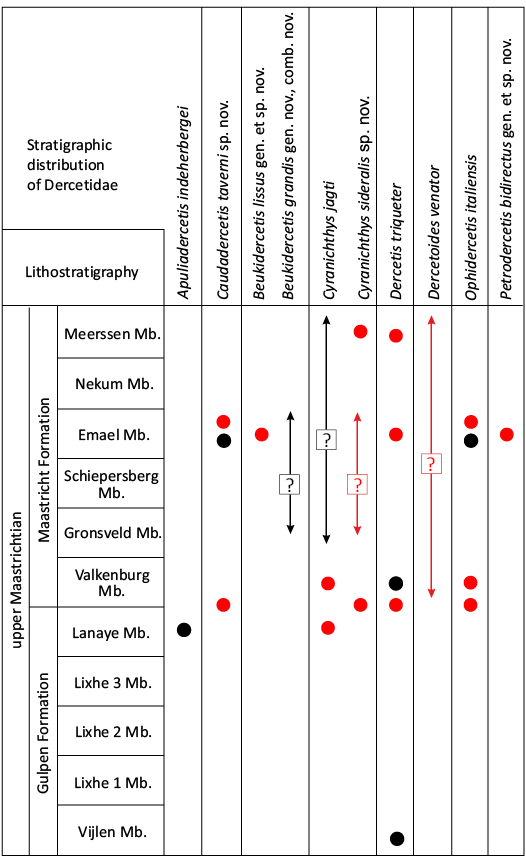
Fig. 26. Stratigraphical distribution of dercetid fish in the Maastrichtian type area. Black indicate finds described by Taverne and Goolaerts (2015) and Wallaard et al. (2019). Red indicate the specimens described herein. Note that NHMM K 2372 (Fig. 5B) was transferred from Cyranichthys jagti to Caudadercetis taverni sp. nov. and Pelargorhynchus grandis reassigned herein to Beukidercetis gen. nov. (modified from Taverne and Goolaerts 2015: fig. 2). Abbreviation: Mb., Member.
Conclusions
Twenty-two dercetid fish specimens from the Maastrichtian type area in Belgium and the Netherlands are recorded. The material includes several specimens of Dercetis triqueter and Ophidercetis italiensis, which improve current knowledge of the stratigraphical distribution of these taxa in the Maastrichtian type area. Of the genus Cyranichthys, two species (Cyranichthys jagti and Cyranichthys sideralis sp. nov.) possibly co-existed, in addition to a specimen that bears a resemblance to the type species, Cyranichthys ornatissimus. Additionally, the present project has documented an increased geographical and stratigraphical ranges for the genus Dercetoides, originally recorded from the Cenomanian of Israel. In addition, new genera and species are erected on the basis of the present material: Caudadercetis taverni sp. nov., Beukidercetis lissus gen. et sp. nov., Cyranichthys sideralis sp. nov., and Petrodercetis bidirectus gen et. sp. nov., relying mostly on unique scute morphologies. Dercetids from the Maastrichtian type area are preserved much better than other actinopterygian fish, suggesting that they either exhibited a benthic lifestyle, or were less digestible due to their slender bodies and high scute coverage. In general, dercetid diversity has been underappreciated, which makes it likely that more species will turn up in similar strata in the future.
Note added in proof
Recently, Gallo et al. (2025) described the first ever dercetid fish known from from the upper Campanian (Snow Hill Formation) of James Ross Island, Antarctica, as a new genus and species, Antarctichthys longipectoralis.
Acknowledgements
We wish to thank Arno Savelkoul (Nieuwstadt, the Netherlands), Dirk Eysermans (Vosselaar, Belgium), Ken Cardon (Het Nationaal Oertijdmuseum, Tongeren, Belgium), Lars P.J. Barten (University of Nijmegen, Rijkevoort, the Netherlands), Maria Verschelling-Wallaard (Boven-Hardinxveld, the Netherlands), and the late Robert Frijns for donating specimens for the present study. One of us (JJAH) also acknowledges Lars W. van den Hoek Ostende (Universiteit Leiden, the Netherlands) for his supervision during the MSc Research Project that laid the foundation for the present paper. Finally, we want to thank Oleksandr Kovalchuk (National Museum of Natural History, Kyiv, Ukraine and University of Wroclaw, Poland) and the anonymous reviewer for their comments.
Editor: Andrzej Kaim
References
Agassiz, L.1834. Abgerissene Bemerkungen über fossile Fische. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1834: 379–390.
Albers, H. and Weiler, W. 1964. Eine Fischfauna aus der oberen Kreide von Aachen und neuere Funde von Fischresten aus dem Maestricht [sic] des angrenzenden belgisch-holländischen Raumes. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 120: 1–33.
Alvarado-Ortega, J. and Díaz-Cruz, J.A. 2021. Hastichthys totonacus sp. nov., a North American Turonian dercetid fish (Teleostei, Aulopiformes) from the Huehuetla quarry, Puebla, Mexico. Journal of South American Earth Sciences 105: 102900. Crossref
Arambourg, C. 1943. Note préliminaire sur quelques poissons fossiles nouveaux. Bulletindela Société géologique de France 5: 281–285. Crossref
Argyriou, T., Alexopoulos, A., Carrillo-Briceño, J.D., and Cavin, L. 2022. A fossil assemblage from the mid-late Maastrichtian of Gavdos Island, Greece, provides insights into the pre-extinction pelagic ichthyofaunas of the Tethys. PLoS ONE 17 (4): e0265780. Crossref
Argyriou, T. and Davesne, D. 2021. Offshore marine actinopterygian assemblages from the Maastrichtian–Paleogene of the Pindos Unit in Eurytania, Greece. PeerJ 9: e10676. Crossref
Bieńkowska-Wasiluk, M., Uchman, A., Jurkowska, A., and Świerczewska-Gładysz, E. 2015. The trace fossil Lepidenteron lewesiensis: a taphonomic window on diversity of Late Cretaceous fishes. Paläontologische Zeitschrift 89: 795–806. Crossref
Blanco, A. and Alvarado-Ortega, J. 2006. Rhynchodercetis regio, sp. nov., a dercetid fish (Teleostei: Aulopiformes) from Vallecilo, Nuevo Léon State, northeastern Mexico. Journal of Vertebrate Paleontology 26: 552–558. Crossref
Bless, M.J.M. 1989. Event-induced changes in Late Cretaceous to Early Paleocene ostracode assemblages of the SE Netherlands and NE Belgium. Annales de la Société géologique de Belgique 112: 19–30.
Bless, M.J.M., Felder, P.J., and Meessen, J.P.M.T. 1987. Late Cretaceous sea level rise and inversion: their influence on the depositional environment between Aachen and Antwerp. In: M.J.M. Bless, M. Dusar, and M. Streel (eds.), Some Aspects of the Late Cretaceous in NW Europe. Annales de la Société géologique de Belgique 109 (for 1986): 333–355.
Boles, Z., Ullmann, P.V., Putnam, I., Ford, M., and Deckhut, J.T. 2024. New vertebrate microfossils expand the diversity of the chondrichthyan and actinopterygian fauna of the Maastrichtian–Danian Hornerstown Formation in New Jersey. Acta Palaeontologica Polonica 69: 173–198. Crossref
Casier, E.1965. Poissons fossiles de la Série du Kwango (Congo). Musée royal de l’Afrique centrale, Séries in 8. Sciences géologiques 50: 1–64.
Catania, K.C. 2019. The astonishing behavior of electric eels. Frontiers in Integrative Neuroscience 13: art. 23. Crossref
Chalifa, Y. 1989. Two new species of longirostrine fishes from the early Cenomanian (Late Cretaceous) of Ein-Yabrud, Israel, with comments on the phylogeny of the Dercetidae. Journal of Vertebrate Paleontology 9: 314–328. Crossref
Chida, M., Brinkman, D.B., and Murray, A.M. 2023. A large, new dercetid fish (Teleostei: Aulopiformes) from the Campanian Bearpaw Formation of Alberta, Canada. Cretaceous Research 150: 105579. Crossref
Cope, ED. 1878. Description of Fishes from the Cretaceous and Tertiary Deposits West of the Mississippi River, Vol. 4. 11 pp. Government Printing Office, Washington D.C.
d’Erasmo, G. 1946. L’ittiofauna cretaceae dei dintorni di Comeno del Carso Triestino. 136 pp. Stabilimento tipografico G. Genovese, Napoli.
Davies, W. 1879. On some fish exuviæ from the Chalk, generally referred to Dercetis elongatus, Ag.; and on a new species of fossil annelide, Terebella lewesiensis. Geological Magazine 6: 145–148. Crossref
Davis, J.W. 1890. On the fossil fish of Cretaceous formation of Scandinavia. The Scientific Transactions of the Royal Dublin Society 4: 363–434.
de Figueiredo, F.J. and Gallo, V.2006. A new dercetid fish (Neoteleostei: Aulopiformes) from the Turonian of the Pelotas Basin, southern Brazil. Palaeontology 49: 445–456. Crossref
Dhondt, A.V. 1971. Systematic revision of Entolium, Propeamussium (Amusiidae) and Syncyclonema (Pectinidae, Bivalvia, Mollusca) of the European boreal Cretaceous. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre 47 (32): 1–95.
Díaz-Cruz, J.A., Alvarado-Ortega, J., and Cantalice, K.M. 2022. Apuliadercetis gonzalezae sp. nov., a North American Campanian dercetid fish (Teleostei, Aulopiformes) from Tzimol, Chiapas, Mexico. Cretaceous Research 130: 105060. Crossref
Díaz-Cruz, J.A., Alvarado-Ortega, J., and Carbot-Chanona, G. 2016. The Cenomanian short snout enchodontid fishes (Aulopifomes, Enchodontidae) from Sierra Madre Formation, Chiapas, southeastern Mexico. Cretaceous Research 61: 136–150. Crossref
Donovan, S.K. and Jagt, J.W.M. 2020. Ichnology of Late Cretaceous echinoids from the Maastrichtian type area (the Netherlands, Belgium). 4. Shark versus echinoid: failed predation on the holasteroid Hemipneustes. Bulletin of the Mizunami Fossil Museum 47: 49–57.
Duffin, C.J. and Reynders, J.P.H. 1995. A fossil chimaeroid from the Gronsveld Member (Late Maastrichtian, Late Cretaceous) of northeast Belgium. Belgian Geological Survey, Professional Paper [Elasmobranches et stratigraphie] 278: 111–156.
Felder, W.M. 1975a. Lithostratigrafie van het Boven-Krijt en het Dano-Montien in Zuid-Limburg en het aangrenzende gebied. In: W.H. Zagwijn and C.J. van Staalduinen (eds.), Toelichting bij geologische overzichtskaarten van Nederland, 63–72. Rijks Geologische Dienst, Haarlem.
Felder, W.M. 1975b. Lithostratigraphische Gliederung der Oberen Kreide in Süd-Limburg (Niederlande) und den Nachbargebieten. Erster Teil: Der Raum westlich der Maas, Typusgebiet des ‘Maastricht’. Publicaties Van Het Natuurhistorisch Genootschap in Limburg 24: 1–43.
Friedman, M. 2012. Ray-finned fishes (Osteichthyes, Actinopterygii) from the type Maastrichtian, the Netherlands and Belgium. In: J.W.M. Jagt, S.K. Donovan, and E.A. Jagt-Yazykova (eds.), Fossils of the Type Maastrichtian. Part 1. Scripta Geologica, Special Issue 8: 113–142.
Gallo, V., de Figueiredos, F.J., Souza Brum, A., Bento Soares, M., Manso Sayao, J., Piacentini Pinheiro, A.E., Figueiredo, R.G., Lopes, R.T., Oliveira de Araujo, O.M., and Kellner, A.W.A. 2025. First dercetid fish from the Upper Cretaceous of Antarctica. Scientific Reports 15: 26650. Crossref
Hay, O.P. and Day, A. 1903. On a collection of Upper Cretaceous fishes from Mount Lebanon, Syria, with descriptions of four new genera and nineteen new species. Bulletin of the American Museum of Natural History 19: 395–452.
Heere, J.J.A., Wallaard, J.J.W., Mulder, E.W.A., Ponstein, J., and Schulp, A.S. 2023. The first report of Chelonioidea cf. Ctenochelys from the Late Cretaceous of the Maastrichtian type area. Netherlands Journal of Geosciences 102: e6. Crossref
Herman, J. 1977. Les sélaciens des terrains néocrétacées & paléocènes de Belgique & des contrées limitrophes. Eléments d’une biostratigraphie intercontinentale. Mémoires explicatives des Cartes géologiques et minières de la Belgique 15 (for 1975): 5–401.
Jagt, J.W.M. 1999. Late Cretaceous–Early Palaeogene echinoderms and the K/T boundary in the southeast Netherlands and northeast Belgium – Part 1: Introduction and stratigraphy; Part 2: Crinoids. Scripta Geologica 116: 1–57, 59–255.
Jagt, J.W.M. 2000. Late Cretaceous–Early Palaeogene echinoderms and the K/T boundary in the southeast Netherlands and northeast Belgium – Part 3: Ophiuroids. With a chapter on Early Maastrichtian ophiuroids from Rügen (eastern Germany) and Møn (Denmark) by Manfred Kutscher & John W.M. Jagt; Part 4: Echinoids; Part 5: Asteroids; Part 6: Discussion and conclusions. Scripta Geologica 121: 1–179, 181–375, 377–503, 505–577.
Jagt, J.W.M. 2019. Met visschubben bekleed – het sporenfossiel Lepidenteron lewesiensis uit het Krijt van Luik-Limburg. Gea 52: 15–18.
Jagt, J.W.M. and Jagt-Yazykova, E.A. 2012. Stratigraphy of the type Maastrichtian—a synthesis. In: J.W.M. Jagt, S.K. Donovan, and E.A. Jagt-Yazykova (eds.), Fossils of the Type Maastrichtian. Part 1. Scripta Geologica, Special Issue 8: 5–32.
Jagt, J.W.M. and Jagt-Yazykova, E.A. 2018. Stratigraphical ranges of tegulated inoceramid bivalves in the type area of the Maastrichtian Stage (Belgium, the Netherlands). In: E.A. Jagt-Yazykova, J.W.M. Jagt, and R.N. Mortimore (eds.), Advances in Cretaceous Palaeontology and Stratigraphy—Christopher John Wood Memorial Volume. Cretaceous Research 87: 385–394. Crossref
Jagt, J.W.M. and Jagt-Yazykova, E.A. 2019. Late Cretaceous and Cenozoic cephalopods from the southern North Sea Basin: stocktaking and future directions. Vita Malacologica 18: 1–33.
Jagt, J.W.M., Donovan, S.K., Deckers, M.J.M., Dortangs, R.W., Kuypers, M.M.M., and Veltkamp, C.J. 1998. The Late Maastrichtian bourgueticrinid crinoid Dunnicrinus aequalis(d’Orbigny, 1841) from the Netherlands and Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre 68: 129–154.
Jagt, J.W.M., Cornelissen, D., Mulder, E.W.A., Schulp, A.S., Severijns, J., and Verding, L. 2008. The youngest in situ record to date of Mosasaurus hoffmanni (Squamata, Mosasauridae) from the Maastrichtian type area, the Netherlands. In: M.J. Everhart (ed.), Proceedings of the Second Mosasaur Meeting, 73–80. Fort Hays State University, Hays.
Jurkowska, A. and Uchman, A. 2013. The trace fossil Lepidenteron lewesiensis (Mantell, 1822) from the Upper Cretaceous of southern Poland. Acta Geologica Polonica 63: 611–623. Crossref
Keutgen, N. 2018. A bioclast-based astronomical timescale for the Maastrichtian in the type area (southeast Netherlands, northeast Belgium) and stratigraphic implications: the legacy of P.J. Felder. Netherlands Journal of Geosciences 97: 229–260. Crossref
Keutgen, N., Remin, Z., and Jagt, J.W.M. 2017. The late Maastrichtian Belemnella kazimiroviensis group (Cephalopoda, Coleoidea) in the Middle Vistula valley (Poland) and the Maastricht area (the Netherlands, Belgium)—taxonomy and palaeobiological implications. Palaeontologia Electronica 20: 2.38A Crossref
Kovalchuk, O., Barkaszi, Z., and Anfimova, G. 2022. Records of Enchodus (Teleostei, Aulopiformes) from the Cenomanian of Ukraine in the light of European distribution of enchodontid fishes. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 303: 295–307. Crossref
Klein, E.F. 1885, Beiträge zur Bildung des Schädels der Knochenfische II. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 41: 107–261.
Lambers, P. 1998. Beenvissen. In: J.W.M. Jagt, J. Leloux, and A.V. Dhondt (eds.), Fossielen van de St. Pietersberg [Limburgnummer 9B]. Grondboor & Hamer 52: 142–143.
Leske, N.G. 1778. Jacobi Theodori Klein Naturalis dispositio Echinodermatum, edita et descriptionibus novisque inventis et synonymis auctorum aucta. 278 pp. G.E. Beer, Leipzig.
Liebau, A. 1978. Paläobathymetrische und paläoklimatische Veränderungen im Mikrofaunenbild der Maastrichter Tuffkreide. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 157: 233–237. Crossref
Maxwell, E.E. and Wilson, L.A. 2013. Regionalization of the axial skeleton in the ‘ambush predator’ guild—are there developmental rules underlying body shape evolution in ray-finned fishes? BMC Evolutionary Biology13: art. 265. Crossref
Mantell, G.A. 1822. The Fossils of the South Downs or Illustrations of the Geology of Sussex. 327 pp. Lupton Relfe, London. Crossref
Mayrinck, D., Brito, P.M., and Otero, O. 2015. Review of the osteology of the fossil fish formerly attributed to the genus †Chanoides and implications for the definition of otophysan bony characters. Journal of Systematic Palaeontology 13: 397–420 [published online in 2014]. Crossref
Miedema, F., Schulp, A.S., Jagt, J.W.M., and Mulder, E.W.A. 2019. New plesiosaurid material from the Maastrichtian type area, the Netherlands. Netherlands Journal of Geosciences 98: e3. Crossref
Mulder, E.W.A. 2003.On latest Cretaceous tetrapods from the Maastrichtian type area.Publicaties van het Natuurhistorisch Genootschap in Limburg 44 (1): 1–188.
Mulder, E.W.A., Jagt, J.W.M., and Stroucken, J.W. 2016. New records of latest Cretaceous neosuchian crocodyliforms from the Maastrichtian type area (southern Limburg, the Netherlands). In: R.M. Sullivan and S.G. Lucas (eds.), Fossil Record 5. New Mexico Museum of Natural History and Science Bulletin 74: 169–172.
Pictet, F.J. 1850. Description de quelques poissons fossiles du Mont Liban. 86 pp. Jules-Guillaume Fick, Genève. Crossref
Pictet, F.J. and Humbert, A. 1866. Nouvelles recherches sur les poissons fossiles du Mont Liban. 114 pp. Jean-Baptiste Baillière, Paris. Crossref
Ride, W.D.L., Copper, H.G., Dupuis, C., Kraus, O., Minelli, A., Thompson, F.C., and Tubbs, P. K. 1999. International Code of Zoological Nomenclature, 4th Edition. 140 pp. The International Trust for Zoological Nomenclature, London.
Rosen, D.E. 1973. Interrelationships of higher euteleostean fishes. In: P.H. Greenwood, R.S. Miles, and C. Patterson (eds.), Interrelationships of Fishes. 536 pp. Zoological Journal of the Linnean Society 53 (Supplement 1): 397–513.
Schiøler, P., Brinkhuis, H., Roncaglia, L., and Wilson, G.J. 1997. Dinoflagellate biostratigraphy and sequence stratigraphy of the Type Maastrichtian (Upper Cretaceous), ENCI Quarry, the Netherlands. Marine Micropaleontology 31: 65–95. Crossref
Schwarzhans, W.W. and Jagt, J.W.M. 2021. Silicified otoliths from the Maastrichtian type area (Netherlands, Belgium) document early gadiform and perciform fishes during the Late Cretaceous, prior to the K/Pg boundary extinction event. Cretaceous Research 127: 104921. Crossref
Siegfried, P. 1966. Zur Osteologie der Gattung Dercetis Agassiz (Teleostei, Pisces). Paläontologische Zeitschrift 40: 205–217. Crossref
Signeux, J.1954. Notes paléoichthyologiques. IV. Leptotrachelus nouveau du Sénonien de Sahel Alma (Liban). Bulletin du Muséum national d’Histoire naturelle, Paris (2) 26: 642–643.
Silva, H.M.A. and Gallo, V.2011. Taxonomic review and phylogenetic analysis of Enchodontoidei (Teleostei: Aulopiformes). Anais da Academia Brasileira de Ciências 83: 483–511. Crossref
Sorbini, L. and Bannikov, A.F.1991. The Cretaceous fishes of Nardò 2°. An enigmatic spiny-rayed fish. Bollettino della Società Paleontologica Italiana 30: 239–249.
Sprechmann, P. 1981. Paleocommunities and paleobathymetry of Maastrichtian sublittoral benthonic foraminifera from western Europe. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 162: 188–230. Crossref
Suhr, P. 1988. Taxonomie und Ichnologie fossiler Wohnröhren terebelloider Würmer. Freiberger Forschungshefte 419: 81–87.
Taverne, L.1987. Ostéologie de Cyranichthys ornatissimus nov. gen. du Cénomanien du Zaïre et de Rhynchodercetis yovanovitchi du Cénomanien de l’ Afrique du Nord. Les relations intergénériques et la position systématique de la famille néocrétacique marine des Dercetidae (Pisces, Teleostei). Annales du Musée royal de l’Afrique Centrale, Rapport Annuel 1985–86: 93–112.
Taverne, L. 1991. New considerations on the osteology and phylogeny of the Cretaceous marine teleost family Dercetidae. Biologisch Jaarboek Dodonea 58: 94–112.
Taverne, L. 2005a. Les poissons crétacés de Nardò. 21°. Ophidercetis italiensis gen. et sp. nov. (Teleostei, Aulopiformes, Dercetidae). Une solution ostéologique au problème des genres Dercetis et Benthesikyme (= Leptotrachelus). Bollettino del Museo civico di Storia naturale di Verona, Geologia Paleontologia Preistoria 29: 55–79.
Taverne, L. 2005b. Les poissons crétacés de Nardò. 22°. Nardodercetis vandewallei gen. et sp. nov. (Teleostei, Aulopiformes, Dercetidae). Bollettino del Museo civico di Storia naturale di Verona, Geologia Paleontologia Preistoria 29: 81–93.
Taverne, L. 2006a. Les poissons crétacés de Nardò. 23°. Apuliadercetis tyleri gen. et sp. nov. (Teleostei, Aulopiformes, Dercetidae). Bollettino del Museo civico di Storia naturale di Verona, Geologia Paleontologia Preistoria 30: 11–26.
Taverne, L. 2006b. Les poissons crétacés de Nardò. 24°. Caudadercetis bannikovi gen. et sp. nov. (Teleostei, Aulopiformes, Dercetidae). Considérations sur la phylogénie des Dercetidae. Bollettino del Museo civico di Storia naturale di Verona, Geologia Paleontologia Preistoria 30: 27–48.
Taverne, L. 2013. Les poissons du Santonien (Crétacé supérieur) d’Apricena (Italie du Sud). 5°. Nardodercetis garganoi sp. nov. (Teleostei, Aulopiformes, Dercetidae). Bollettino del Museo civico di Storia naturale di Verona, Geologia Paleontologia Preistoria 37: 67–72.
Taverne, L. and Goolaerts, S. 2015. The dercetid fishes (Teleostei, Aulopiformes) from the Maastrichtian (Late Cretaceous) of Belgium and the Netherlands. Geologica Belgica 18: 21–30.
van Baal, R.R., Janssen, R., Van der Lubbe, H.J.L., Schulp, A. S., Jagt, J.W.M., and Vonhof, H.B. 2013. Oxygen and carbon stable isotope records of marine vertebrates from the type Maastrichtian, the Netherlands and northeast Belgium (Late Cretaceous). Palaeogeography, Palaeoclimatology, Palaeoecology 392: 71–78. Crossref
Van der Ham, R.W.J.M., Jagt, J.W.M., Renkens, S., and van Konijnenburg-van Cittert, J.H.A. 2010. Seed-cone scales from the upper Maastrichtian document the last occurrence in Europe of the Southern Hemisphere conifer family Araucariaceae. Palaeogeography, Palaeoclimatology, Palaeoecology 291: 469–473. Crossref
Van der Ham, R.W.J.M., van Konijnenburg-van Cittert, J.H.A., and Indeherberge, L. 2007. Seagrass foliage from the Maastrichtian type area (Maastrichtian, Danian, NE Belgium, SE Netherlands). Review of Palaeobotany and Palynology144: 301–321. Crossref
Vellekoop, J., Kaskes, P., Sinnesael, M., Huygh, J., Déhais, T., Jagt, J.W.M., Speijer, R.P., and Claeys, P. 2022. A new age model and chemostratigraphic framework for the Maastrichtian type area (southeastern Netherlands, northeastern Belgium). Newsletters on Stratigraphy 55: 479–501. Crossref
Vellekoop, J., Van Tilborgh, K.H., van Knippenberg, P., Jagt, J.W.M., Stassen, P., Goolaerts, S., and Speijer, R.P. 2020. Type-Maastrichtian gastropod faunas show rapid ecosystem recovery following the Cretaceous–Palaeogene boundary catastrophe. Palaeontology 63: 349–367. Crossref
Vernygora, O., Murray, A.M., Luque, J., Ruge, M.L.P., and Fonseca, M.E.P. 2017. A new Cretaceous dercetid fish (Neoteleostei: Aulopiformes) from the Turonian of Colombia. Journal of Systematic Palaeontology 16: 1057–1071. Crossref
Villain, J.-M. 1977. Le Maastrichtien dans sa région type (Limbourg, Pays-Bas). Étude stratigraphique et micropaléontologique. Palaeontographica A 157: 1–87.
Von der Marck, W. 1858. Über einige Wirbeltiere, Kruster und Cephalopoden der westfälischen Kreide. Zeitschrift der Deutschen geologischen Gesellschaft 10: 231–271.
Von der Marck, W. 1863. Fossile Fische, Krebse und Pflanzen aus dem Plattenkalke der jüngsten Kreide in Westphalen. Palaeontographica 11: 1–83.
Wallaard, J.J.W., Fraaije, R.H.B., Diependaal, H.J., and Jagt, J.W.M. 2019. A new species of dercetid (Teleostei, Aulopiformes) from the type Maastrichtian of southern Limburg, the Netherlands. Netherlands Journal of Geosciences 98: e2. Crossref
Woodward, A.S. 1901. Actinopterygian Teleostomi of the Suborders Isospondyli (in Part), Ostariophysi, Apodes, Percesoces, Hemibranchii, Acanthopterygii, and Anacanthini. 636 pp. British Museum (Natural History), London. Crossref
Acta Palaeontol. Pol. 70 (4): 661–697, 2025
https://doi.org/10.4202/app.01253.2025