
Seagrass-associated Middle Miocene brachiopods from the Central Paratethys, with description of a new species of Bronnothyris
ALFRÉD DULAI
Dulai, A. 2025. Seagrass-associated Middle Miocene brachiopods from the Central Paratethys, with description of a new species of Bronnothyris. Acta Palaeontologica Polonica 70 (4): 649–660.
The Middle Miocene brachiopod fauna of the Mecsek Hills (Southern Hungary) is poorly known. In this paper the brachiopods of a new fossil site discovered by private collectors on the outskirts of Mecsekpölöske village is described. The studied assemblage from the lower Badenian clayey sand (Lajta Limestone Formation) is very abundant (5248 specimens), and represented partly by some species well-known from other shallow marine sites of the Central Paratethys (Joania cordata, Megathiris detruncata, Gryphus miocenicus, Megerlia truncata, and Discradisca sp.). However, the most abundant taxon is a new species of Bronnothyris (i.e., B. attilavorosi sp. nov.), characterised by subcircular outline; four to seven single, weak, rounded ribs; distinct cardinal process; strongly prominent triangular dorsal median septum, with septal flanges and four strong serrations on its anterior slope; short, slightly divergent inner socket ridges; narrow fused hinge plates. This is the first record of Bronnothyris from the Central Paratethys and from the Miocene. The Hungarian species extends the stratigraphic range of Bronnothyris: it is known from the Upper Cretaceous to the Middle Miocene. The taxonomic composition of the Mecsekpölöske brachiopod assemblage (with strong dominance of Megathyrididae), together with several elements of the associated fauna (e.g., gastropods, foraminifers, ostracods, otoliths, and fish teeth), suggests that the palaeoenvironment may have been a shallow marine seafloor covered with seagrass meadows. Both the recently published upper Pliocene brachiopod assemblage from Italy (Villalvernia, Piedmont) and this Middle Miocene assemblage presented here from Hungary confirm that micromorphic Megathyrididae brachiopods can be used as indirect palaeo-seagrass indicators (IPSIs). However, since they do not occur exclusively in this environment, other direct or indirect evidence of the former presence of seagrass meadows is also necessary.
Key words: Brachiopoda, Megathyrididae, seagrass meadows, indirect palaeo-seagrass indicator (IPSI), Badenian, Miocene, Mecsek Hills, Hungary.
Alfréd Dulai [dulai.alfred@nhmus.hu; ORCID: https://orcid.org/0000-0002-9366-5217 ], Department of Palaeontology and Geology, Hungarian National Museum Public Collection Centre – Hungarian Natural History Museum, Ludovika Square 2–6, H-1083 Budapest, Hungary.
Received 1 July 2025, accepted 26 August 2025, published online 30 October 2025.
Copyright © 2025 A. Dulai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Shallow marine fauna of the Middle Miocene Central Paratethys was extremely diverse, and the various sedimentary formations contain abundant fossil remains of many invertebrate and vertebrate groups (Moissette et al. 2006; Hyžný and Dulai 2021; Harzhauser et al. 2024). Although brachiopods generally play a subordinate role in the Cenozoic, they are regularly found in smaller to larger quantities in Neogene formations. This is particularly true for the small-sized, so-called micromorphic brachiopod species, which can sometimes be significantly enriched in screen-washed residues.
When discussing a large palaeogeographical unit, the knowledge and exploration of individual sub-areas can often vary greatly. This is the case for the Central Paratethys and the Mecsek Hills in southern Hungary. Compared to other Central Paratethyan areas, there are very few publications and limited data available on Miocene brachiopods from this region. Even the only major publication (Matyasovszky 1880) would need revision (Dulai 2025a).
The role of private collectors, who often discover previously unknown sites and valuable new fossils, is increasing. This was the case here, where the new fossil site at Mecsekpölöske was discovered by private collector Tamás Németh (Balatonkenese, Hungary), who drew attention to the extremely diverse fossil material found there.
The aim of this paper is to discuss the Middle Miocene brachiopod assemblage from the Mecsekpölöske site, to describe a new species of Bronnothyris from there and to interpret the palaeoenvironment on the basis of the taxonomic composition of the identified brachiopods and some associated fossil groups. In addition to reconstructing the local palaeoenvironment, the possible use of micromorphic Megathyrididae brachiopods as indirect palaeo-seagrass indicators in a broader sense is also examined.
Institutional abbreviations.—HNHM, Hungarian National Museum Public Collection Centre – Hungarian Natural History Museum, Budapest, Hungary.
Other abbreviations.—A, articulated specimen; D, dorsal valve; fr, fragment; IPSI, indirect palaeo-seagrass indicator; MNI, minimal number of individuals; V, ventral valve.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in Zoobank: urn:lsid:zoobank.org:pub:6766FAF0-DC54-4E4A-AF63-07E18DF428C8.
Geological setting
The Paratethys was a large inland sea intermittently connected to the Mediterranean (Rögl 1998; Popov et al. 2004). Its diverse marine fauna refers to a stable connection with the Mediterranean in the present-day Slovenia (Kókay 1985) (Fig. 1). However, the biogeographic differentiation necessitated the establishment of a regional stratigraphic scheme (Harzhauser and Piller 2007). The Badenian (16.303–12.829 Ma) is a regional stage used in the Central Paratethys for part of the Middle Miocene (Langhian to middle Serravallian) (Papp et al. 1978; Hohenegger et al. 2014).

Fig. 1. Palaeogeographic sketch map of the the Middle Miocene Central Paratethys (modified from Rögl 1998), indicating the studied locality in the Mecsek Hills (Mecsekpölöske, Southern Hungary).
During the Miocene, islands of Permian and Mesozoic rocks of the Mecsek Hills were surrounded by the Central Paratethys, leaving a variety of lagoonal, coastal and near-shore sediments (Hámor 1970; Chikán 1991; Sebe et al. 2015). Three Lower and Middle Miocene sedimentary cycles are observed in the Mecsek Hills (Sebe et al. 2015). The upper part of the second cycle is represented by the lower Badenian Pécsszabolcs Limestone Member of the Lajta Limestone Formation (Selmeczi et al. 2024), it consists of varied shallow marine sediments (conglomerate, glauconitic sandstone, bioclastic limestone, calcareous sandstone and sandy clays). The rich macrofaunal content of the red algal “lower Leitha limestone” consists mainly of mollusc, echinoid and bryozoan remains, which are indicative of the Chlamys elegans/Pecten revolutus Subzone of the Flabellipecten besseri Assemblage Zone (Bohn-Havas et al. 1987). Amphistegina, Miliolina, and Heterostegina species are the most common in the benthic Foraminifera assemblages (Korecz-Laky 1968).
The geology of the area around Mecsekpölöske is described in Strausz (1928) and Chikán (1991). The studied new site is located 1 km east of the village, in a gully on the western side of the Nagy Hill (46.22311° N, 18.22904° E; Kovács and Vicián 2021) (Fig. 2). The outcropping layers, which are about 3 m thick, are composed of mollusc-bearing limestone layers alternating with yellowish sandy clays (Fig. 3). The latter contain a very well-preserved and diverse fossil fauna, including molluscs and brachiopods. Additional faunal elements in the screen-washed residues are foraminifers, ostracods, bryozoans, decapods, worms, echinoids, otholiths, and fish teeth. Among the molluscs, representatives of the family Muricidae (Kovács 2020: 12 species, e.g., Purpurellus cyclopterus [Millet, 1865], Aspella subanceps [d’Orbigny, 1852], Murexsul sandbergeri [Hörnes, 1856], and Favartia suboblonga [d’Orbigny, 1852]) and the family Cancellariidae (Kovács and Vicián 2021: Bivetiella dertonensis [Bellardi, 1841], Scalptia hidasensis [Hoernes & Auinger, 1890]) have been published so far. The fragmentary colonial corals from Mecsekpölöske are mentioned briefly by Strausz (1923, 1928), Soós-Kablár (2013), and Dulai et al. (2021), without more detailed investigations. The processing of the fish teeth (Márton Szabó, personal communication 2025), as well as polyplacophoran (AD and Márton Szabó, unpublished data) assemblages recovered from the site are ongoing.
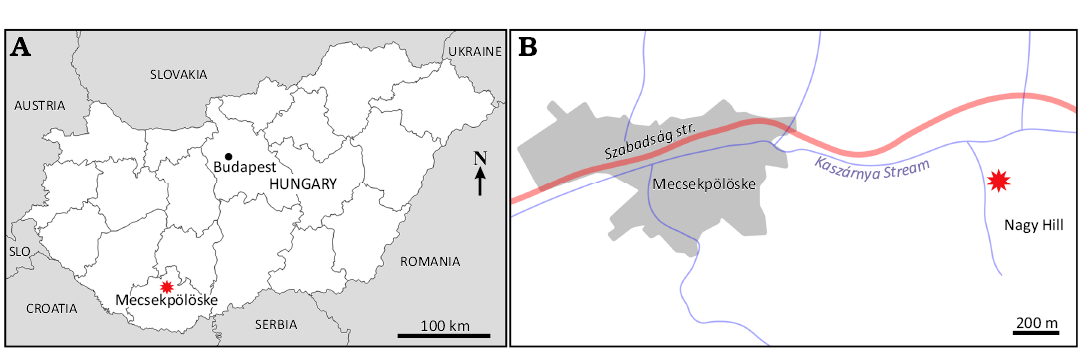
Fig. 2. Geographic location of Mecsekpölöske fossil site. A. Location of Mecsekpölöske in Southern Hungary. Abbreviation: Slo, Slovenia. B. Location of the fossil site on the western side of Nagy Hill.

Fig. 3. Mecsekpölöske fossil site. A. Outcropping Middle Miocene layers (lower Badenian Pécsszabolcs Limestone Member of the Lajta Limestone Formation) along the western side of the Nagy Hill, Mecsekpölöske. B. Soft yellowish sandy clays, containing abundant micromorphic brachiopod assemblage. Length of hammer 315 mm (photos by Márton Szabó).
Historical background
In the Neogene shallow marine environments, the screen-washed residues commonly yield an abundant but not very diverse micromorphic brachiopod fauna. These are dominated mainly by members of the families Megathyrididae and Thecideidae. The family Thecideidae is known from only two sites (Bivolare and Yasen) of the Central Paratethys in Bulgaria (Bitner 1993; Bitner and Motchurova-Dekova 2016). However, Megathyrididae taxa (Megathiris, Argyrotheca, Joania) are very common in the Central Paratethyan Miocene materials (e.g., Bitner 1990, 1993; Bitner and Pisera 2000; Bitner and Kaim 2004; Dulai 2007, 2015; and further references therein).
Although members of the family Megathyrididae belong to the most common shallow marine brachiopods, it contains few genera with rather uneven distribution. Phragmothyris Cooper, 1955, is limited to the Paleogene of Cuba (Cooper 1955), while Borellithyris was only recently described from the Upper Miocene of the Mediterranean by Dulai (2019). Bronnothyris Popiel-Barczyk & Smirnova, 1978, was restricted to the Upper Cretaceous and lower Paleocene of N-NW Europe (Denmark, England, Germany, and Poland) according to the revised Treatise (Lee et al. 2006), however, later it was also recorded from the upper Eocene of Ukraine (Bitner and Müller 2017) and from the Oligocene of Germany (Bitner and Kroh 2011; Dulai and von der Hocht 2020; Bitner and Müller 2022). Until now, it was not identified from the Miocene and from the Central Paratethys. The other three genera (Megathiris d’Orbigny, 1847; Argyrotheca Dall, 1900; and Joania Álvarez et al., 2008) are much more common and widely distributed in the Neogene brachiopod assemblages. Megathiris detruncata (Gmelin, 1791) is common almost everywhere in the European Neogene (Kocsis et al. 2020), and can therefore be found at numerous sites in the Central Paratethys (e.g., Bitner 1990; Dulai 2007; Bitner and Motchurova-Dekova 2016; and further references therein). Even more common were Argyrotheca (= Cistella, Argiope) records, for which numerous species names were used in the earlier literature. Without the need for completeness or details, only at the level of enumeration: Argyrotheca cistellula (Wood, 1841), A. cordata (Risso, 1826), A. cuneata (Risso, 1826), A. dertomutinensis (Sacco, 1902), A. interponens (Dreger, 1889), A. neapolitana (Scacchi, 1833), A. squamata (Eichwald, 1830), A. subcordata (Boettger, 1901), A. subcuneata (Boettger, 1901), and A. zboroviensis (Friedberg, 1921) were recorded from several Neogene sites. However, based on revisions of the last decades (e.g., Bitner 1990), only two valid Argyrotheca species remained in the Middle Miocene of the Central Paratethys: A. cuneata and A. cordata. The latter was reclassified in 2008 as the type species of the new genus Joania (Álvarez et al. 2008). These three Miocene Central Paratethyan megathyridid species (M. detruncata, A. cuneata, J. cordata) were later complemented by A. bitnerae Dulai in Dulai & Stachacz, 2011, described from some Middle Miocene sites in Poland (Dulai and Stachacz 2011), and this species was later found within the Central Paratethys in the Czech Republic (Hladilová et al. 2014; Kopecká et al. 2018), in Hungary (Zebegény; Dulai 2021) and in Romania (Buituri; Dulai 2021).
Material and methods
The Mecsekpölöske fossil site was brought to my attention by private collector Tamás Németh (Balatonkenese, Hungary), and a smaller part of the studied material comes from his collection. In addition to my own collecting efforts, Márton Szabó (HNHM Department of Palaeontology and Geology) has also screen-washed and selected a considerable amount of material from Mecsekpölöske.
The studied micromorphic brachiopods are deposited in the Palaeontological Collection of the Hungarian Natural History Museum (type material: HNHM PAL 2025.11.1–23.1; additional material: HNHM INV 2025.261–277).
The minimal number of individuals (MNI) was calculated in every species: A + V or D, depending on the higher value.
Middle Miocene brachiopod assemblage of Mecsekpölöske.—More than five thousand micromorphic brachiopods collected from the screen-washed residues are well preserved and comprise several articulated specimens. The majority of isolated valves were separated during the screen-washing treatment and ultrasonic cleaning. The brachiopod assemblage is not diverse (6 species), and the two dominant species, as well as a third species belong to the typical shallow-water family Megathyrididae. Among the well-known megathyridid species, Joania cordata is common, while Megathiris detruncata is rare (Fig. 4). The most common species in this assemblage is another member of Megathyrididae, a new species of Bronnothyris (see Systematic palaeontology section). In addition to the dominant Megathyrididae specimens, several juvenile Gryphus miocenicus, a few Megerlia truncata and a single Discradisca sp. fragment were also present in the studied samples. Detailed data of the brachiopods identified from the washed residues (articulated specimens, ventral valves, dorsal valves, total number of specimens, minimum number of individuals) are presented in Table 1.

Fig. 4. Middle Miocene micromorphic megathyridid brachiopods from Mecsekpölöske, Mecsek Hills, Hungary, Middle Miocene (Badenian). A–F. Joania cordata (Risso, 1826). A. Articulated specimen, HNHM INV 2025.261., ventral view. B. Articulated specimen, HNHM INV 2025.262., dorsal view. C. Articulated specimen, HNHM INV 2025.263., dorsal view. D. Dorsal valve, HNHM INV 2025.264., oblique lateral (D1) and internal (D2) views. E. Ventral valve, HNHM INV 2025.265., internal view. F. Dorsal valve, HNHM INV 2025.266., internal (F1) and oblique lateral (F2) views. G–J. Megathiris detruncata (Gmelin, 1791). G. Articulated specimen, HNHM INV 2025.267., dorsal view. H. Dorsal valve, HNHM INV 2025.268., external view. I. Ventral valve, HNHM INV 2025.269., internal view. J. Ventral valve, HNHM INV 2025.270, internal view. Scale bars 500 µm.
Table 1. Detailed data of Middle Miocene (Badenian) brachiopods identified from the screen-washed residues of Mecsekpölöske site (Mecsek Hills, S Hungary). Abbreviations: A, articulated specimen; D, dorsal valve; V, ventral valve; fr, fragment; MNI, minimal number of individuals (A+D or V).
|
Species |
Specimens (A, V, D, fr) |
Total |
MNI |
|
Discradisca sp. |
1 fr |
1 (0.02%) |
1 (0.03%) |
|
Gryphus miocenicus |
56 A, 31 V, 17 D |
107 (2.04%) |
90 (2.31%) |
|
Bronnothyris attilavorosi sp. nov. |
1046 A, 1271 V, 858 D |
3204 (61.05%) |
2325 (59.73%) |
|
Joania cordata |
728 A, 707 V, 447 D |
1892 (36.05%) |
1443 (37.08%) |
|
Megathiris detruncata |
22 A, 6 V, 4 D, 6 fr |
42 (0.8%) |
31 (0.8%) |
|
Megerlia truncata |
1A, 1 fr |
2 (0.04%) |
2 (0.05%) |
Independently of the screen-washed materials from the sandy clays, some larger brachiopod specimens were also found in the debris of the hillside, possibly derived from the more massive limestone layers and represent Gryphus miocenicus and Megerlia truncata.
Systematic palaeontology
Phylum Brachiopoda Duméril, 1805
Subphylum Rhynchonelliformea Williams, Carlson, Brunton, Holmer, & Popov, 1996
Class Rhynchonellata Williams, Carlson, Brunton, Holmer, & Popov, 1996
Order Terebratulida Waagen, 1883
Suborder Terebratellidina Muir-Wood, 1955
Superfamily Megathyridoidea Dall, 1870
Family Megathyrididae Dall, 1870
Genus Bronnothyris Popiel-Barczyk & Smirnova, 1978
Type species: Terebratula bronni Roemer, 1841, by original designation of Popiel-Barczyk and Smirnova (1978: 41); Rügen, Germany, Maastrichtian.
Bronnothyris attilavorosi sp. nov.
Figs. 5, 6.
ZooBank LSID: urn:lsid:zoobank.org:act:F633147E-1D3E-472B-90C9-CA578CE19F00.
Etymology: In honour of Attila Vörös (Hungarian Natural History Museum, Budapest; member of the Hungarian Academy of Sciences), eminent researcher of Mesozoic (mainly Jurassic) brachiopods, on the occasion of his 80th birthday.
Type material: Holotype, HNHM PAL 2025.11.1 (Fig. 5A), articulated specimen. Paratypes: HNHM PAL 2025.12.1 (Fig. 5B), articulated specimen; HNHM PAL 2025.13.1 (Fig. 5C), articulated specimen; HNHM PAL 2025.14.1 (Fig. 5D), articulated specimen; HNHM PAL 2025.15.1 (Fig. 6A), ventral valve; HNHM PAL 2025.16.1 (Fig. 6B), ventral valve; HNHM PAL 2025.17.1 (Fig. 6C), ventral valve; HNHM PAL 2025.18.1 (Fig. 6D), dorsal valve; HNHM PAL 2025.19.1 (Fig. 6E), dorsal valve; HNHM PAL 2025.20.1 (Fig. 6F), dorsal valve; HNHM PAL 2025.21.1 (Fig. 6G), dorsal valve; HNHM PAL 2025.22.1 (Fig. 6H), dorsal valve; HNHM PAL 2025.23.1 (Fig. 6I), dorsal valve. All from the type locality and horizon.
Type locality: Nagy Hill, Mecsekpölöske, Mecsek Hills, Hungary (46.22311° N, 18.22904° E).
Type horizon: Pécsszabolcs Limestone Member, Lajta Limestone Formation, Badenian, Middle Miocene.
Material.—Holotype, 12 paratypes, and additional rich material (1053 articulated specimens, 1272 ventral valves, 879 dorsal valves).
Diagnosis.—Small-sized Bronnothyris species with subcircular outline; shell surface with four to seven single, weak, rounded ribs; distinct cardinal process; strongly prominent triangular dorsal median septum, with short septal flanges and four strong serrations on its anterior slope; short, slightly divergent inner socket ridges; narrow, fused hinge plates.
Measurements.—See Table 2.
Table 2. Measurements (in mm) of Bronnothyris attilavorosi sp. nov. type specimens. Abbreviations: A, articulated specimen; D, dorsal valve; V, ventral valve.
|
Specimen |
Length |
Width |
Thickness |
|
Holotype (A) |
2.4 |
2.4 |
1.3 |
|
HNHM PAL 2025.12.1 (A) |
2.8 |
2.7 |
1.4 |
|
HNHM PAL 2025.13.1 (A) |
2.7 |
2.7 |
1.2 |
|
HNHM PAL 2025.14.1 (A) |
2.8 |
2.7 |
1.4 |
|
HNHM PAL 2025.15.1 (V) |
2.2 |
2.3 |
– |
|
HNHM PAL 2025.16.1 (V) |
2.5 |
2.5 |
– |
|
HNHM PAL 2025.17.1 (V) |
3.1 |
3.1 |
– |
|
HNHM PAL 2025.18.1 (D) |
1.8 |
2.4 |
– |
|
HNHM PAL 2025.19.1 (D) |
1.8 |
2.5 |
– |
|
HNHM PAL 2025.20.1 (D) |
1.7 |
2.3 |
– |
|
HNHM PAL 2025.21.1 (D) |
2.6 |
3.1 |
– |
|
HNHM PAL 2025.22.1 (D) |
1.4 |
1.8 |
– |
|
HNHM PAL 2025.23.1 (D) |
1.9 |
2.4 |
– |
Description.—External characters: Shell small (maximum observed length 3.1 mm), thin, equibiconvex in lateral profile (Fig. 5A2, C2). Slightly variable in outline, mostly subcircular (Fig. 5A1) or rounded subpentagonal (Fig. 5C1). Length and width more or less the same. Maximum width situated at mid-length or slightly moved forward the posterior third (Fig. 5A1, C1). Maximum thickness at mid-length to posterior third (Fig. 5C2). The apical angle varies between 95° and 110°. Hinge line long and straight, its length near to maximum width (Fig. 5A1, C1). Beak high, erect with sharp beak ridges (Fig. 5C2) and transversely striated, wide interarea (Figs. 5A1, C1, 7A, B). Beak truncated by large and wide subtriangular hypothyrid foramen, bordered by narrow, disjunct deltidial plates (Fig. 5A1, C1). Lateral commissures straight (Fig. 5C2), anterior commissure rectimarginate with shallow median sulcus on ventral valve (Fig. 5A2). Shell surface covered with four to seven single, weak, rounded ribs running through from umbo to anterior margin (Fig. 5A1, B, C1, D), space between ribs wider than ribs. No bifurcation or intercalation. The seventh shorter median rib only sometimes visible (Fig. 5C1, D). Distinct growth lines on some specimens, denser near to the anterior and lateral margins (Fig. 5A1, B, C1, D).

Fig. 5. Megathyridid brachiopod Bronnothyris attilavorosi sp. nov., Mecsekpölöske, Mecsek Hills, Hungary, Middle Miocene (Badenian); holotype (A) and paratypes (B–D); external morphological characters. A. Articulated specimen, HNHM PAL 2025.11.1, dorsal (A1) and anterior (A2) views. B. Articulated specimen, HNHM PAL 2025.12.1, ventral view. C. Articulated specimen, HNHM PAL 2025.13.1, dorsal (C1) and lateral (C2) views. D. Articulated specimen, HNHM PAL 2025.14.1, ventral view. Scale bars 500 µm.
Internal characters: Both valves coarsely endopunctate (Fig. 6A, D). Ventral valve interior with poorly developed short teeth lying parallel to hinge line (Fig 6A). Well-developed large, highly triangular pedicle collar, with dense transverse striation (Fig. 6A–C), supported by a low ventral median septum extending to mid-valve (Fig. 6A, B), sometimes longer (Fig. 6C). Anteriorly to ventral septum three ovoid depressions accommodate dorsal septum serrations (Fig. 6A, B). Dorsal valve interior with moderately deep subhorizontal sockets (Fig. 6D1, F, H1, I1); short, slender socket ridges situated near to each other and extending beyond posterior margin (Fig. 6D1, H1, I). Distinct cardinal process breaks down into parallel lamellae (Fig. 6D1, E, G1) and continued anteriorly in a triangular (Figs. 6E, G1, H1, I) or in a longer ridge (Fig. 6D1). Hinge plates fused mid-dorsally with valve floor to form a narrow platform (Fig. 6D1, E, I). Dorsal median septum beginning from anterior margin of hinge plates (Fig. 6D1, E), low posteriorly, becoming high at anterior third (Fig. 6G2); triangular in profile (Fig. 6F, G2, H2), the posterior edge rise at 45°, the anteroventral edge sloping towards anterior margin with four strong, sharp or rounded serrations (Fig. 6F, G2, H2). Short septal flanges extend posteroventrally from top of dorsal median septum (Fig. 6D, I1). Crura short, medially directed crural processes wide and relatively long (Fig. 6D1, E, F). Loop poorly calcified and poorly developed. Laterally curved descending branches united with valve floor (Fig. 6D1, E, F) and their anterior ends join to median septum (Fig. 6F, H2, I1).

Fig. 6. Megathyridid brachiopod Bronnothyris attilavorosi sp. nov., Mecsekpölöske, Mecsek Hills, Hungary, Middle Miocene (Badenian); paratypes; internal morphological characters. A. Ventral valve, HNHM PAL 2025.15.1, internal view. B. Ventral valve, HNHM PAL 2025.16.1, internal view. C. Slightly eroded ventral valve, HNHM PAL 2025.17.1, internal view. D. Dorsal valve, HNHM PAL 2025.18.1, internal view (D1) and enlarged part of dorsal median septum with septal flanges (D2). E. Dorsal valve, HNHM PAL 2025.19.1, oblique posterior view. F. Dorsal valve, HNHM PAL 2025.20.1, oblique lateral view. G. Dorsal valve, HNHM PAL 2025.21.1, internal (G1) and oblique lateral (G2) views. H. Dorsal valve, HNHM PAL 2025.22.1, internal (H1) and oblique lateral (H2) views. I. Dorsal valve, HNHM PAL 2025.23.1, internal view (I1) and enlarged part of the posterior part with sockets, oblique socket ridges, and cardinal process (I2). Scale bars 500 µm, except D2, and I2, 250 µm.
Remarks.—Within the megathyridid brachiopods found in Mecsekpölöske, the Megathiris and Joania specimens can be easily distinguished based on their internal morphological characters (Megathiris: three septa in the dorsal valve; Joania: series of tubercles on the inner margins of both valves). Two species of the genus Argyrotheca are known from the Central Paratethys. Argyrotheca bitnerae Dulai in Dulai & Stachacz, 2011, clearly differs from the Mecsekpölöske specimens by its subtrigonal outline and unornamented, smooth valves. A. cuneata, which is known from many sites since the end of the Paleogene to the present day, slightly resembles the new species described here, but can be clearly distinguished on the basis of short septal flanges on the high and triangular dorsal median septum and the fused hinge plates. Both characters refer to the genus Bronnothyris, previously unknown from both the Central Paratethys and the Miocene. In the case of strongly eroded Lower Miocene Argyrotheca cf. subradiata (Sandberger, 1862) specimens published by Bitner and Schneider (2009) from the Bavarian part of the Molasse Basin, their affiliation to the genus Bronnothyris cannot be confirmed due to the absence of septal flanges.
The Mecsekpölöske specimens can be clearly distinguished from all known Bronnothyris species, thus justifying the introduction of a new species for this material. The previously described taxa differ from the new species in the following characters:
–Bronnothyris bronni (Roemer, 1841) (Upper Cretaceous) in transversely slightly elongated bilobate outline; flat dorsal valve in adults, more numerous and stronger ribs extend slightly beyond the shell margin; not so high ventral beak, hinge plates form two separated and concave circular discs, indistinct, flat cardinal process (Steinich 1965; Surlyk 2025);
– Bronnothyris coniuncta (Steinich, 1965) (Upper Cretaceous) in transversely strongly elongated, “Spirifer-like” outline, more numerous and stronger ribs slightly extend beyond the shell margin, low ventral beak, fused hinge plates form a broad platform, wide and flat cardinal process, short dorsal median septum without anterior serrations (Steinich 1965; Surlyk 2025);
– Bronnothyris lacunosa (Steinich, 1965) (Upper Cretaceous) in rectangular outline, few rounded ribs, hinge plates form two concave discs, indistinct, flat cardinal process; anterior slope of dorsal median septum without strong serrations(Steinich 1965);
– Bronnothyris obstinata (Steinich, 1965) (Upper Cretaceous) in rectangular outline, few rounded ribs, indistinct, flat cardinal process, anterior slope of lower dorsal median septum without serrations (Steinich 1965);
– Bronnothyris stevensis(Nielsen, 1928) (Upper Cretaceous) in strongly inflated valves, much stronger ribs in two bundles, extend beyond the shell margin; hinge plates form two concave discs, higher septal flanges (Nielsen 1928; Simon 1998; Surlyk 2025);
– Bronnothyris rugicosta(Zelinskaya, 1975) (lower Paleocene) in more numerous, very strong and wide ribs, extend slightly beyond the shell margin, fused hinge plates form a broad platform, anterior slope of dorsal median septum without strong serrations (Zelinskaya 1975);
– Bronnothyris wansinensis (Vincent, 1923) (upper Paleocene) in transversely strongly elongated outline, fused hinge plates forming a broad platform; stronger ribs do not extend to the anterior margin (Vincent 1923; Dulai 2025b);
– Bronnothyris danaperensis Bitner & Müller, 2017 (upper Eocene) in transversely elongated outline, smooth surface, fused hinge plates forming a broad platform, eroded ventral beak area (Bitner and Müller 2017);
– Bronnothyris subradiata (Sandberger, 1862) (lower Oligocene) in transversely elongated, more irregular outline, more numerous and partly stronger ribs; strongly eroded ventral beak area, fused hinge plates forming a broad platform (Sandberger 1862; Bitner and Kroh 2011);
– Bronnothyris rugosa (Schreiber, 1871) (lower Oligocene) in much wider than long outline, lower and wider beak area, longer hinge line, more numerous, stronger and tuberculate ribs, broad and fused hinge plates forming a coherent platform (Bitner and Müller 2022).
The presence of septal flanges on the dorsal median septum and the fused hinge plates indicate a relationship between the Mecsekpölöske material and Bronnothyris. However, the narrowness of the fused hinge plates is striking when compared to previously known Cenozoic Bronnothyris species (B. wansinensis, Vincent 1923: fig. 3; B. danaperensis, Bitner and Müller 2017: fig. 7E–G, I, J; B. subradiata, Bitner and Kroh 2011: fig. 3C–I; B. rugosa, Bitner and Müller 2022: fig. 8E–K). After thorough examination of all the species classified in the genus Bronnothyris it might be concluded that the aspect of hinge plates is rather diverse. Three different morphotypes can be distinguished, what may result oversplitting into as many as three genera: (i) fused hinge plates forming a broad platform: Bronnothyris coniuncta, B. rugicosta, B. wansinensis, B. danaperensis, B. subradiata, B. rugosa; (ii) hinge plates form a narrow, single plate: B. obstinata, B. attilavorosi sp. nov.; (iii) hinge plates form two concave, separated circular discs: B. bronni, B. lacunosa, B. stevensis.
For the time being, I consider the new species B. attilavorosi, known from a single locality, to be a member of the genus Bronnothyris, reserving the possibility that the discovery of further similar materials or more detailed investigation of the available materials may justify the description of some new genera in the future within the family Megathyrididae.
Bitner and Kroh (2011) emphasised in the morphological description of B. subradiata that the ventral beak of most specimens is heavily eroded, which may indicate a very short pedicle and very tight attachment to the hard substrate. The situation was similar with B. danaperensis specimens from the Ukrainian upper Eocene (Bitner and Müller 2017). In the case of B. attilavorosi sp. nov., the vast majority of the specimens is in very good preservation, with no signs of erosion around the ventral beak. This may indicate that this Miocene species had a longer pedicle than older Cenozoic relatives, but it is more likely that the different palaeoenvironment and nature of preferred substrate could have played a more significant role (see the assessment of palaeoenvironment of Mecsekpölöske site in the discussion).
Stratigraphic and geographic range.—Middle Miocene (Badenian), Mecsek Hills (Southern Hungary).
Discussion
Palaeoenvironmental consideration on the Mecsekpölöske site.—The brachiopod assemblage collected from the screen-washed residues is strongly dominated by species of Megathyrididae generally typical of shallow-water environments (Bronnothyris + Joania + Megathiris = 97.91%), with negligible amounts of species typical of deeper waters (Gryphus + Megerlia = 2.07%) and the representatives of the latter group are mostly small-sized, juvenile specimens (Fig. 7). The fragmentary Discradisca sp. also tends to be more typical of shallow-water environments. The preservation of brachiopods is excellent, most of the specimens were originally articulated, but in several individuals the valves became separated during screen-washing and ultrasonic cleaning (because of the small ventral teeth and loose connection of valves). The percentage ratio of the number of specimens of each species and the calculated minimum number of individuals is almost identical (Table 1), also confirming that this is an autochthonous assemblage unaffected by significant transport.
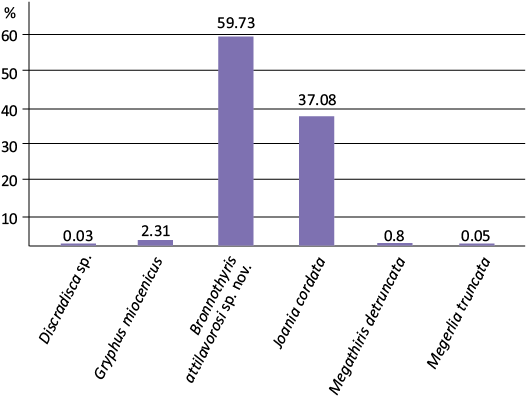
Fig. 7. Taxonomic composition of the Middle Miocene (Badenian) brachiopod assemblage from Mecsekpölöske, Mecsek Hills, Hungary.
Shallow-water micromorphic brachiopods are most commonly found in sheltered environments (caves, crevices, rocky bedrock) and on coralligenous substrates (Logan 1979). Following some rare previous records (e.g., Taddei Ruggiero 1985; Evangelisti et al. 2011), it has recently been confirmed that the rhizome level of Posidonia seagrass meadows provide excellent sheltered habitat for the attachment of micromorphic brachiopod assemblages with distinctive taxonomic composition. Albano and Stockinger (2019) described an abundant population of Joania cordata–Argyrotheca cuneata from the Posidonia oceanica meadows area in Plakias (SW Crete). Shortly afterwards, the first known fossil brachiopod assemblage associated with seagrass meadows from the Neogene of the Mediterranean was published from the Pliocene of Rio Vaccaruza, Italy by Pavia et al. (2022). In this latter assemblage, Joania cordata was dominant, while the few additional elements were Argyrotheca cuneata and Terebratula juv. sp. In that publication, Pavia et al. (2022) argued that some possible seagrass palaeoenvironments of the Central Paratethys could be interpreted based on their brachiopod assemblages dominated by Megathyrididae (e.g., Bitner 1990, 1993; Bitner and Pisera 2000; Bitner and Kaim 2004; Dulai 2007), but beside the taxonomic composition of the brachiopod fauna, these environmental assessments were supported by few additional evidences. For example, the presence of submarine flora was suggested by Pisera (1985) at Radwanówka (Roztocze Hills, Poland), some foraminifers indicated the presence of non-calcified submarine flora at Niechobrz, SE Poland (Gonera 1994) and several gastropod taxa referred to possible seagrass environment at Bánd, Hungary (Kókay 1966; Kovács and Vicián 2021).
The taxonomic composition of the brachiopod assemblage at Mecsekpölöske (dominanated by species of Bronnothyris and Joania, with some additional of Gryphus and Megathiris) and their excellent preservation may also be explained by a shallow marine bottom covered with seagrass meadows. This assessment of the palaeoenvironment is consistent with morphological observations showing that while in the Eocene B. danaperensis (Bitner and Müller 2017) and the Oligocene B. subradiata (Bitner and Kroh 2011) almost all specimens have heavily eroded ventral beak, whereas B. attilavorosi sp. nov. is almost without exception excellently preserved, very few specimens showing any signs of physical erosion. According to Bitner and Kroh (2011), the B. subradiata species had a very short pedicle, which allowed it to attach itself firmly to the solid surfaces, so that the beak in contact with the hard attachment surface was heavily eroded by the strong water movement prevailing in the shallow sea. On the other hand, B. attilavorosi sp. nov. was attached to the rhizomes of living seagrass or to smaller solid surfaces (e.g., fossil fragments) in the fine sandy sediment trapped in the rhizome level of seagrass meadows, so it did not erode significantly even if it did not have a longer pedicle than its older Cenozoic relatives.
Regardless of brachiopods, there are some other fossil groups in the Mecsekpölöske material that can also be considered indirect palaeo-seagrass indicators (Reich et al. 2015). The abundance of herbivorous gastropods recovered from the studied strata supports this interpretation. The seagrass environment is suggested by the presence of Bittium tani Harzhauser, Guzhov, & Landau, 2025 (very common), Bittium praescabrum Harzhauser, Guzhov, & Landau, 2025 (common), Thecidium zeuschneri (Pusch, 1836) (common), Oligodia bicarinata (Eichwald, 1830) (common), Mitrella viennensis Harzhauser & Landau, 2021 (very common), Conolithes brezinae (Hoernes & Auinger, 1879) (common), among many others. Some additional very common gastropod genera, such as Alvania, Manzonia, Rissoina, and Zebinella also occur in seagrass environments of the Recent seas (Zoltán Kovács, personal communication 2025). The almost monospecific Amphistegina mammilla (Fichtel & Moll, 1798) foraminifer assemblage, some epiphytic ostracod species (Emőke Mohr, personal communication 2025), otoliths of Gobiidae, Sparidae, and Gadidae fishes, which use seagrass meadows as a nursery habitat (Mariann Bosnakoff, personal communication 2025), as well as some sharks and other fishes: angel sharks (Squatinidae), reef sharks (Carcharhinidae), whiptail stingrays (Dasyatidae), wrasses (Labridae), porgies and seabreams (Sparidae) (Márton Szabó, personal communication 2025) also may indicate the presence of a palaeo-seagrass environment. A comprehensive palaeoenvironmental assessment of the Mecsekpölöske site is in preparation by AD and other colleagues.
Both the Italian Pliocene (Pavia et al. 2022) and the Hungarian Middle Miocene Megathyrididae assemblages confirm that these micromorphic brachiopods can be useful as indirect palaeo-seagrass indicators. Considering the criteria for the usefulness of IPSIs defined by Reich et al. (2015), Megathyrididae brachiopods: (i) have appropriate stratigraphic (Upper Cretaceous–Recent; Lee et al. 2006) and geographical (worldwide except for the polar regions; Logan 2007: fig. 1954) distribution; (ii) have excellent fossilization potential (Kocsis et al. 2020); (iii) based on known examples, they occur in sufficiently large numbers in palaeo-seagrass meadows (Pavia et al. 2022; this paper); however, (iv) they are not strictly limited to seagrass meadows and can also occur in many other shallow marine habitats (Logan 1979), therefore, additionally to well-preserved Megathyrididae brachiopods, further evidence from other groups is needed at the studied fossil sites.
Conclusions
An abundant but low-diversity micromorphic brachiopod fauna was found in the Badenian (Middle Miocene) sandy clays (Lajta Limestone Formation) of the Mecsekpölöske fossil site in southern Hungary. More than five thousand specimens were identified from the screen-washed residues, representing six brachiopod species. The taxonomic composition of the brachiopod assemblage was found to be fairly uniform, with almost 98% of the specimens representing the species of Bronnothyris, Joania, and Megathiris (Megathyrididae). The most common species was identified as a new species of Bronnothyris (B. attilavorosi sp. nov.). Bronnothyris was long thought to have disappeared after the end-Cretaceous mass extinction, but more and more Bronnothyris records are being found in the Cenozoic strata. This is the first known occurrence of Bronnothyris from the Central Paratethys and from the Miocene. The youngest known stratigraphic occurrence of the genus is B. attilavorosi sp. nov. from the Middle Miocene of Mecsekpölöske.
The dominance of the family Megathyrididae and the low abundance (2%) of deeper water forms (Gryphus, Megerlia; mainly juvenile specimens) clearly indicate a shallow water environment. The taxonomic composition of the brachiopod assemblage and the co-occurrence of some specific other fossils groups (e.g., gastropods, foraminifers, ostracods, otoliths, fish teeth) suggest that the shallow-marine substrate was covered by seagrass meadows. This is the first evidence that Bronnothyris, like other megathyridid genera (Argyrotheca and Joania), may have been associated with seagrass meadows. Reich et al. (2015) published an excellent summary of IPSI groups, but only mentioned brachiopods in passing, stating that although they occur in seagrass beds, no detailed studies have been published on them. Following their confirmed occurrence in Recent Posidonia meadows (Albano and Stockinger 2019), both the recently published similar Italian Pliocene Joania–Argyrotheca material (Pavia et al. 2022) and this Hungarian Miocene Bronnothyris–Joania–Megathiris assemblage confirmed that micromorphic Megathyrididae brachiopods can be used as indirect palaeo-seagrass indicator. However, since their distribution is not limited to this habitat, they can only be used for this purpose in conjunction with some further positive evidences from the studied site.
Acknowledgements
Part of the studied Mecsekpölöske brachiopod material was collected by Tamás Németh (Balatonkenese, Hungary) and Márton Szabó (Budapest, Hungary). I thank László Sővér (Bonyhád, Hungary) for allowing me to examine some of his larger brachiopod specimens found at Mecsekpölöske site. The SEM photos were taken at the SEM Laboratory of the HNHM in Budapest (by Márton Szabó) and at the Plant Protection Institute of the HUN-REN Centre for Agricultural Research in Martonvásár, Hungary (by Barna Páll-Gergely). Thanks to Márton Szabó for the locality map and the field photos. Many thanks to my friends and colleagues for the useful discussions on associated fossil groups: Zoltán Kovács, Márton Szabó, and Mariann Bosnakoff (HNHM) and Emőke Tóth (Eötvös University, Budapest, Hungary). The study was supported by Hungarian Scientific Research Fund (NKFI/OTKA K146962). I would like to thank the reviewers Maria Aleksandra Bitner (Insitute of Paleobiology PAS, Warsaw, Poland) and Finn Surlyk (University of Copenhagen, Denmark) for their constructive suggestions, which have significantly improved the quality of this publication. Last but not least, I would like to thank Professor Attila Vörös, who is congratulated on the occasion of his 80th birthday. A very significant part of my knowledge of brachiopods comes from Attila Vörös who was my external supervisor to both my M.Sc. and Ph.D. thesis and later my boss and colleague at Hungarian Natural History Museum, Budapest.
Editor: Andrzej Kaim
References
Albano, P.G. and Stockinger, M. 2019. The rhizome layer of Posidonia oceanica: an important habitat for Mediterranean brachiopods. Marine Biodiversity 49: 2467–2472. Crossref
Álvarez, F., Brunton, C.H.C., and Long, S.L. 2008. Loop ultrastructure and development in Recent Megathiridoidea, with description of a new genus, Joania (type species Terebratula cordata Risso, 1826). Earth and Environmental Science Transactions of the Royal Society of Edinburgh 98: 391–403. Crossref
Bitner, M.A. 1990. Middle Miocene (Badenian) brachiopods from the Roztocze Hills, south-eastern Poland. Acta Geologica Polonica 40: 129–157.
Bitner, M.A. 1993. Middle Miocene (Badenian) brachiopods from coral reefs of north-western Bulgaria. Acta Geologica Polonica 4: 147–155.
Bitner, M.A. and Kaim, A. 2004. The Miocene brachiopods from the silty facies of the intra-Carpathian Nowy Sącz Basin (Poland). Geological Quarterly 48: 193–198.
Bitner, M.A. and Kroh, A. 2011. First record of the genus Bronnothyris (Brachiopoda: Megathyrididae) from the Oligocene of the Mainz Basin (Germany). Geologica Carpathica 62: 203–209. Crossref
Bitner, M.A. and Motchurova-Dekova, N. 2016. Middle Miocene (Badenian) brachiopods from Yasen, northwestern Bulgaria: taxonomic composition and biogeographical significance. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 29: 7–22. Crossref
Bitner, M.A. and Müller, A. 2017. Late Eocene (Priabonian) brachiopod fauna from Dnipropetrovsk, eastern Ukraine. Bulletin of Geosciences 92: 211–231. Crossref
Bitner, M.A. and Müller, A. 2022. Early Oligocene brachiopods from the rocky-shore deposits at Mammendorf, Central Germany. Annales Societatis Geologorum Poloniae 92: 87–107. Crossref
Bitner, M.A. and Pisera, A. 2000. Brachiopod fauna from the Middle Miocene deposits of Niechobrz, south-eastern Poland. Tertiary Research 20: 7–15.
Bitner, M.A. and Schneider, S. 2009. The Upper Burdigalian (Ottnangian) brachiopod fauna from the northern coast of the Upper Marine Molasse Sea in Bavaria, Southern Germany. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 254: 117–133. Crossref
Bohn-Havas, M., Báldi, T., Kókay, J., and Halmai, J. 1987. Pectinid assemblage zones of the Miocene in Hungary. Jahrbuch der Ungarischen Geologischen Anstalt 70: 441–446.
Chikán, G. 1991. Die Käinozoischen Ablagerungen des westlichen Mecsekgebirges. Jahrbuch der Ungarischen Geologischen Anstalt 72: 1–281.
Cooper, G.A. 1955. New brachiopods from Cuba. Journal of Paleontology 29: 64–70.
Dulai, A. 2007. Badenian (Middle Miocene) micromorphic brachiopods from Bánd and Devecser (Bakony Mountains, Hungary). Fragmenta Palaeontologica Hungarica 24–25: 1–13.
Dulai, A. 2015. Central Paratethyan Middle Miocene brachiopods in Naturalis Biodiversity Center (Leiden, the Netherlands). Scripta Geologica 149: 185–211.
Dulai, A. 2019. New data on the Late Miocene brachiopod fauna of Tetti Borelli (Piedmont, Italy). Rivista Italiana di Paleontologia e Stratigrafia 125: 125–145.
Dulai, A. 2021. Szemelvények az európai kainozoós Brachiopoda fauna fejlődéstörténetéből. 409 pp, Unpublished D.Sc. Thesis, Hungarian Natural History Museum, Budapest.
Dulai, A. 2025a. Taxonomic revision of some Central Paratethyan Middle Miocene brachiopod species described by Matyasovszky in 1880 (Terebratulida; Megathyrididae; Terebratulidae). Annales Musei historico-naturalis hungarici 117: 121–130. Crossref
Dulai, A. 2025b. Emended description and generic attribution of Argyrotheca wansinensis Vincent, 1923 (Brachiopoda, Megathyrididae; upper Palaeocene, Belgium). Cainozoic Research 25 (2): 167–172.
Dulai, A., Henn, T., and Sebe, K. 2021. Middle Miocene (Badenian) macroinvertebrates from Pécs-Danitzpuszta (Mecsek Mts, SW Hungary). Földtani Közlöny 151: 329–334. Crossref
Dulai, A. and Stachacz, M. 2011. New Middle Miocene Argyrotheca (Brachiopoda; Megathyrididae) species from the Central Paratethys. Földtani Közlöny 141: 283–291.
Dulai, A. and von der Hocht, F. 2020. Upper Oligocene brachiopods from NW Germany, with description of a new Platidiinae genus, Germanoplatidia n. gen. Rivista Italiana di Paleontologia e Stratigrafia 126: 223–248.
Evangelisti, F., Albano, P.G., and Sabelli, B. 2011. Recent Brachiopoda of the Marine Protected Area “Secche di Tor Paterno”, Central Tyrrhenian Sea. Cahers de Biologie Marine 52: 193–202.
Gonera, M. 1994. Palaeoecology of Marine Middle Miocene (Badenian) in the Polish Carpathians (Central Paratethys). Foraminiferal record. Bulletin of the Polish Academy of Sciences, Earth Sciences 42: 107–125.
Hámor, G. 1970. Das Miozän des östlichen Mecsek-Gebirges. Jahrbuch der Ungarischen Geologischen Anstalt 53: 373–483.
Harzhauser, M. and Piller, W.E. 2007. Benchmark data of a changing sea —palaeogeography, palaeobiogeography and events in the Central Paratethys during the Miocene. Palaeogeography, Palaeoclimatology, Palaeoecology 253: 8–31. Crossref
Harzhauser, M., Landau, B., Mandic, O., and Neubauer, T.A. 2024. The Central Paratethys Sea—rise and demise of a Miocene European marine biodiversity hotspot. Scientific Reports 14: 16288. Crossref
Hladilová, Š., Nehyba, S., Zágoršek, K., Tomanová-Petrová, P., Bitner, M.A., and Demény, A. 2014. Early Badenian transgression on the outer flank of Western Carpathian Foredeep, Hluchov area, Czech Republic. Annales Societatis Geologorum Poloniae 84: 259–278.
Hohenegger, J., Ćorić, S., and Wagreich, M. 2014. Timing of the Middle Miocene Badenian Stage of the Central Paratethys. Geologica Carpathica 65: 55–66. Crossref
Hyžný, M. and Dulai, A. 2021. Badenian decapods of Hungary. 296 pp, GeoLitera, Szeged.
Kocsis, L., Dulai, A., Cipriani, A., Vennemann, T., and Yunsi, M. 2020. Geochemistry of Recent and fossil brachiopod calcite of Megathiris detruncata (Terebratulida, Megathyrididae): A modern baseline study to trace past environmental conditions. Chemical Geology 533: 119335. Crossref
Kókay, J. 1966. Geologische und paläontologische Untersuchung des Braunkohlengebietes von Herend – Márkó (Bakony-Gebirge, Ungarn). Geologica Hungarica, Series Palaeontologica 36: 1–147.
Kókay, J. 1985. Central and Eastern Paratethyan interrelations in the light of Late Badenian salinity conditions. Geologica Hungarica, Series Palaeontologica 48: 9–95.
Kopecká, J., Holcová, K., Nehyba, S., Hladilová, Š., Brzobohaty, R., and Bitner, M.A. 2018. The earliest Badenian Planostegina bloom deposit: reflection of an unusual environment in the westernmost Carpathian Foredeep (Czech Republic). Geological Quarterly 62: 18–37. Crossref
Korecz-Laky, I. 1968. Miozäne Foraminiferen des östlichen Mecsek-Gebirges. Jahrbuch der Ungarischen Geologischen Anstalt 52: 1–229.
Kovács, Z. 2020. New Muricidae (Neogastropoda) faunas from the Middle Miocene of Hungary. Földtani Közlöny 150: 449–468. Crossref
Kovács, Z. and Vicián, Z. 2021. New Cancellariidae (Neogastropoda) assemblage from the Middle Miocene of Hungary with description of a new species. Central European Geology 64: 59–73. Crossref
Lee, D.E., MacKinnon, D.I., and Smirnova, T.N. 2006. Megathyridoidea. In: R.L. Kaesler (ed.), Treatise on Invertebrate Palaeontology; Part H, Brachiopoda Revised, Volume 5: Rhynchonelliformea (part), 2217–2222. Geological Society of America and The University of Kansas, Boulder and Lawrence.
Logan, A. 1979. The Recent Brachiopoda of the Mediterranean Sea. Bulletin de l’Institut Océanographique de Monaco 72: 1–112.
Logan, A. 2007. Geographic distribution of extant articulated brachiopods. In: P.A. Selden (ed.), Treatise on Invertebrate Palaeontology, Part H, Brachiopoda Revised, Volume 6 (Supplement), 3082–3115. Geological Society of America and The University of Kansas, Boulder and Lawrence.
Matyasovszky, J. 1880. Paläontologische beiträge zur Kenntnis der jüngeren Mediterranen Schichten des Baranyaer Comitates. Természetrajzi Füzetek 4: 243–248.
Moissette, P., Dulai, A., and Müller, P. 2006. Bryozoan faunas in the Middle Miocene of Hungary: biodiversity and biogeography. Palaeogeography, Palaeoclimatology, Palaeoecology 233: 300–314. Crossref
Nielsen, K.B. 1928. Argiope-arterne i Danmarks senone, daniske og paleocaene Aflej-ringer. Meddelelser fra Dansk Geologisk Forening 7: 215–226.
Papp, A., Cicha, I., Seneš, J., and Steininger, F.F. 1978. M4 Badenien (Moravien, Wielicien, Kosovien). Chronostratigraphie und Neostratotypen, Miozän der Zentralen Paratethys. 594 pp. Verlag der Slowakischen Akademie der Wissenschaften, Bratislava.
Pavia, G., Dulai, A., Festa, A., Gennari, R., Pavia, M., and Carnevale, G. 2022. Palaeontology of the Upper Pliocene marine deposits of Rio Vaccaruzza, Villarvernia (Piedmont, NW Italy). Rivista Italiana di Paleontologia e Stratigrafia 128: 129–210. Crossref
Pisera, A. 1985. Paleoecology and lithogenesis of the Middle Miocene (Badenian) algal-vermetid reefs from the Roztocze Hills, south-eastern Poland. Acta Geologica Polonica 35: 89–155.
Popiel-Barczyk, E. and Smirnova, T.N. 1978. New genus megathyrid (Brachiopoda) from the Upper Cretaceous of Poland [in Russian]. Paleontologičeskij žurnal 1978 (3): 134–137.
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Scherba, I.G., and Kovač, M. (eds.) 2004. Lithological-paleogeographic maps of Paratethys. Curier Forschungsinstitut Senckenberg 250: 1–46.
Reich, S., Di Martino, E., Todd, J.A., Wesselingh, F.P., and Renema, W. 2015. Indirect paleo-seagrass indicators (IPSIs): A review. Earth-Science Reviews 143: 161–186. Crossref
Rögl, F. 1998. Palaeogeographic considerations for Mediterranean and Paratethys Seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien 99: 279–310.
Sandberger, C.L.F. 1858–1863. Die Conchylien des Mainzer Tertiärbeckens. 459 pp. C.W. Kreidel’s Verlag, Wiesbaden. Crossref
Sebe, K., Csillag, G., Dulai, A., Gasparik, M., Magyar, I., Selmeczi, I., Szabó, M., Sztanó, O., and Szuromi-Korecz, A. 2015. Neogene stratigraphy in the Mecsek region. Field trip guidebook. In: I.R. Bartha, Á. Kriván, I. Magyar, and K. Sebe (eds.), Programme, Abstracts, Field Guidebook, 6th Workshop on the Neogene of Central and South-Eastern Europe, 31 May – 3 June 2015, Orfű, Hungary, 102–124. Hungarian Geological Society, Budapest.
Selmeczi, I., Müller, P.M., Lelkes, Gy., Kókay, J., and Hámor, G. 2024. Lajta Limestone Formation. In: E. Babinszki, O. Piros, G. Csillag, L. Fodor, L. Gyalog, Z. Kercsmár, G. Less, R. Lukács, K. Sebe, I. Selmeczi, J. Szepesi, and O. Sztanó (eds.), Lithostratigraphic Units of Hungary II. Cenozoic Formations, 84, SARA, Budapest.
Simon, E. 1998. Maastrichtian brachiopods from Ciply: palaeoecological and stratigraphical significance. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 68: 181–232.
Soós-Kablár, J. 2013. Egy 19. századi mészégető kemence maradványa és ennek földtörténeti vonatkozásai Mecsekpölöskén. e-Acta Naturalia Pannonica 5: 1–7.
Steinich, G. 1965. Die artikulaten Brachiopoden der Rügener Schreibkreide (Unter-Maastricht). Paläontologische Abhandlungen, Abteilung A, Paläozoologie 2: 1–220.
Strausz, L. 1923. Über die geologischen Verhaltnisse der Gegend von Mecsekjánosi, Szopók und Mecsekpölöske (im Komitate Baranya). Földtani Közlöny 53: 136–138.
Strausz, L. 1928. Das Mediterran des Mecsekgebirges in Südungarn. Geologische und Paläontologische Abhandlungen, N.F. 15: 361–418.
Surlyk, F. 2025. Maastrichtian brachiopods from the chalk of Denmark. Bulletin of the Geological Society of Denmark 74: 49–118. Crossref
Taddei Ruggiero, E. 1985. Megathyrididae (Brachiopoda) attuali raccolti lungo le coste della Campania e della Puglia. Bollettino della Società dei naturalisti in Napoli 92: 347–413.
Vincent, E. 1923. Quelques remarques sur des Brachiopodes tertiaires de Belgique et description d’une espece nouvelle d’Argyrotheca. Annales de la Société Royale Malacologique de Belgique 53: 49–53.
Zelinskaya, V.A. 1975. Brakhiopody Paleogena Ukrainy. 148 pp, Naukova Dumka, Kiev.
Acta Palaeontol. Pol. 70 (4): 649–660, 2025
https://doi.org/10.4202/app.01272.2025