
A new meiolaniform turtle from the Maastrichtian of Northern Patagonia, Argentina
FEDERICO L. AGNOLIN, MAURO ARANCIAGA ROLANDO, JULIANA STERLI, NICOLÁS R. CHIMENTO, FERNANDO E. NOVAS, and GONZALO L. MUÑOZ
Agnolín, F.L., Aranciaga Rolando, M., Sterli, J., Chimento, N.R., Novas, F.E., and Muñoz, G.L. 2026. A new meiolaniform turtle from the Maastrichtian of Northern Patagonia, Argentina. Acta Palaeontologica Polonica 71 (1): 173–184.
Meiolaniformes are a group of chelonians including the famous horned-turtles Niolamia argentina (Patagonia) and Meiolania platyceps (Australia). In South America, the Late Cretaceous meiolaniforms are represented by two named taxa: Patagoniaemys gasparinae coming from Campanian–Maastrichtian beds of the La Colonia Formation, Chubut province, and Trapalcochelys sulcata from the Campanian–Maastrichtian beds of the Allen Formation, Río Negro Province. The aim of the present contribution is to describe a new meiolaniform chelonian coming from the Upper Cretaceous (Maastrichtian) Los Alamitos Formation, at Río Negro Province, Argentina. The material is represented by a partial basicranium, incomplete carapace and fragmentary postcranial bones (MPMIK 1839/P/33) belonging to a new species Patagoniaemys aeschyli. To this new species we refer all the material previously referred as indeterminate meiolaniid, meiolaniform and cf. Niolamia sp., coming from the same site and locality, and described by different authors since the 1980s. This new species differs from the type species P. gasparinae, particularly by the presence of longitudinal ridges in the nuchal bone. We also include brief comments about meiolaniform palaeobiogeography and the impact of K/Pg extinction event in Patagonian chelonians.
Key words: Testudinata, Meiolaniformes, Los Alamitos Formation, Maastrichtian, Cretaceous, Patagonia, Argentina.
Federico L. Agnolín [fedeagnolin@yahoo.com.ar, ORCID: https://orcid.org/0000-0001-5073-561X], Laboratorio de Anatomía Comparada y Evolución de los Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. Fundación de Historia Natural “Félix de Azara”, Universidad Maimónides, Hidalgo 775, C1405BDB Buenos Aires, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
Mauro Aranciaga Rolando [mauro.a_guido@hotmail.com, ORCID: https://orcid.org/0000-0002-0833-6866] and Gonzalo L. Muñoz [gonza_25_rnr@hotmail.com, ORCID: https://orcid.org/0009-0000-5041-1009], Laboratorio de Anatomía Comparada y Evolución de los Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
Juliana Sterli [jsterli@mef.org.ar, ORCID: https://orcid.org/0000-0003-2942-5558], Museo Paleontológico Egidio Feruglio, Av. Fontana 140, 9100 Trelew, Chubut, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
Nicolás R. Chimento [nicochimento@hotmail.com, ORCID: https://orcid.org/0000-0002-6340-4358] (corresponding author), Laboratorio de Anatomía Comparada y Evolución de los Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. Fundación de Historia Natural “Félix de Azara”. Museo de Ciencias Naturales y Arqueología “Prof. Manuel Almeida”. 25 de Mayo 533 (2820), Gualeguaychú, Entre Ríos, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
Fernando E. Novas [fernovas@yahoo.com.ar, ORCID: https://orcid.org/0000-0002-6901-8677], Fundación de Historia Natural “Félix de Azara”, Universidad Maimónides, Hidalgo 775, C1405BDB Buenos Aires, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
Received 4 June 2025, accepted 22 January 2026, published online 26 March 2026.
Copyright © 2026 F.L. Agnolin et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Meiolaniformes represent a group of turtles that includes the well-known horned species Niolamia argentina (Patagonia) and Meiolania platyceps (Australia). Uncontroversial meiolaniforms are known from Early Cretaceous to Pleistocene times in southern landmasses, including South America, Australia, and New Caledonia (Sterli 2015). In addition, several authors support that the Late Cretaceous Kallokibotion bajazidi from Romania and Mongolochelys efremovi from Mongolia (Nopcsa 1923; Khozatskii 1997), among others, should be included among Meiolaniformes (Ckhikvadzé 1987; Hirayama et al. 2000; Joyce 2007; Sterli 2008; Anquetin 2011; Sterli and de la Fuente 2011a, b, 2019; Sterli et al. 2013; but see Joyce et al. 2016 and Pérez-García and Codrea 2018). The increasing number of extinct turtles potentially related to Meiolaniidae highlights the fact that the group was very diverse and that it had a long and complex evolutionary history.
Until recently, the better-known meiolaniform was the derived clade, the Meiolaniidae, from the Cenozoic, with its most iconic species, Meiolania platyceps described in great detail by Gaffney (1983, 1985, 1996). In South America, Late Cretaceous meiolaniforms are known from two described species: Patagoniaemys gasparinae, which originates from the Campanian–Maastrichtian deposits of the La Colonia Formation in Chubut Province (Sterli and De la Fuente 2011a), and Trapalcochelys sulcata, found in the Campanian–Maastrichtian strata of the Allen Formation in Río Negro Province (Sterli et al. 2013). Recently discovered specimens of Patagoniaemys gasparinae makes this species the most complete non-meiolaniid meiolaniform from Gondwana (Sterli et al. 2021; Oriozabala et al. 2025).
In addition, some scattered remains of several indeterminate meiolaniforms were described from several localities in Patagonia, indicating that the clade is far more abundant and geographically widespread than previously thought (Broin 1987; Broin and de la Fuente 1993; Gasparini and de la Fuente 2000; Sterli et al. 2013, 2021; Sterli 2015; Moyano-Paz et al. 2022). Particularly, Broin (1987) recognized the occurrence of meiolaniids (nowadays recognized as meiolaniforms, see Sterli et al. 2013) in the Maastrichtian beds of the Los Alamitos Formation, at Nueva Poupée farm (Los Alamitos farm at that time), which constitute the first Cretaceous meiolaniforms recognized in South America. She supported the idea that the specimens (consisting of a humerus and isolated carapace fragments and bones) were closely related to the Cenozoic Patagonian meiolaniid, Niolamia argentina. Later, Broin and de la Fuente (1993), Gaffney et al. (2007), and Sterli et al. (2013) recognized new fragmentary meiolaniform specimens from the same locality and age. Because of their isolated and fragmentary nature, all the specimens remained as indeterminate Meiolaniformes.
The aim of present contribution is to describe a partial carapace, cranial, and postcranial bones belonging to an associated meiolaniform specimen coming from the Maastrichtian beds of Los Alamitos Formation, at Nueva Poupeé farm, Río Negro Province, Argentina. The new specimen belongs to a new species of the genus Patagoniaemys, which sheds light on the still poorly known Late Cretaceous record of meiolaniforms.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:8C16458D-1D80-4B4C-8957-21010FB6769E
Institutional abbreviations.—MACN-Pv RN, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Colección Nacional Paleovertebrados-Río Negro, Buenos Aires, Argentina; MPMIK, Museo Provincial “María Inés Kopp”, Valcheta, Río Negro, Argentina.
Other abbreviations.—SQS, shareholder quorum subsampling.
Geological setting
The specimens here described come from the base of Cerro Cuadrado (41° 57’13.5” S 66° 19’ 20.3” W; 582 m a.s.l.), at Nueva Poupée Farm (ex Los Alamitos Farm), approximately at 60 km from Arroyo Ventana locality, Valcheta Department, Río Negro Province, Patagonia, Argentina (Fig. 1A, B). The fossil bearing bed is a non-calcareous pelitic facies from the lower section of the Los Alamitos Formation (sensu Andreis 1987; see Fig. 1C, D). This bed contains almost all the microvertebrates found in the Formation (Bonaparte et al. 1984; Bonaparte 1987). The specimens described here were found together with abundant microvertebrates, including diverse fishes, anurans, snakes, sphenodonts, chelid turtles, birds, sauropods, theropods and hadrosaurid teeth, and abundant sauropod eggshells, as well as an unprecedented mammalian diversity (see Bonaparte et al. 1984; Albino 1987, 2000; Báez 1987; Bonaparte 1987, 2002; Broin 1987; Cione 1987; Powell 1987; Apesteguía 2005; Agnolín and Martinelli 2009; Gómez 2016; Rougier et al. 2021; Rozadilla et al. 2021; Agnolín et al. 2024; Panzeri et al. 2024).
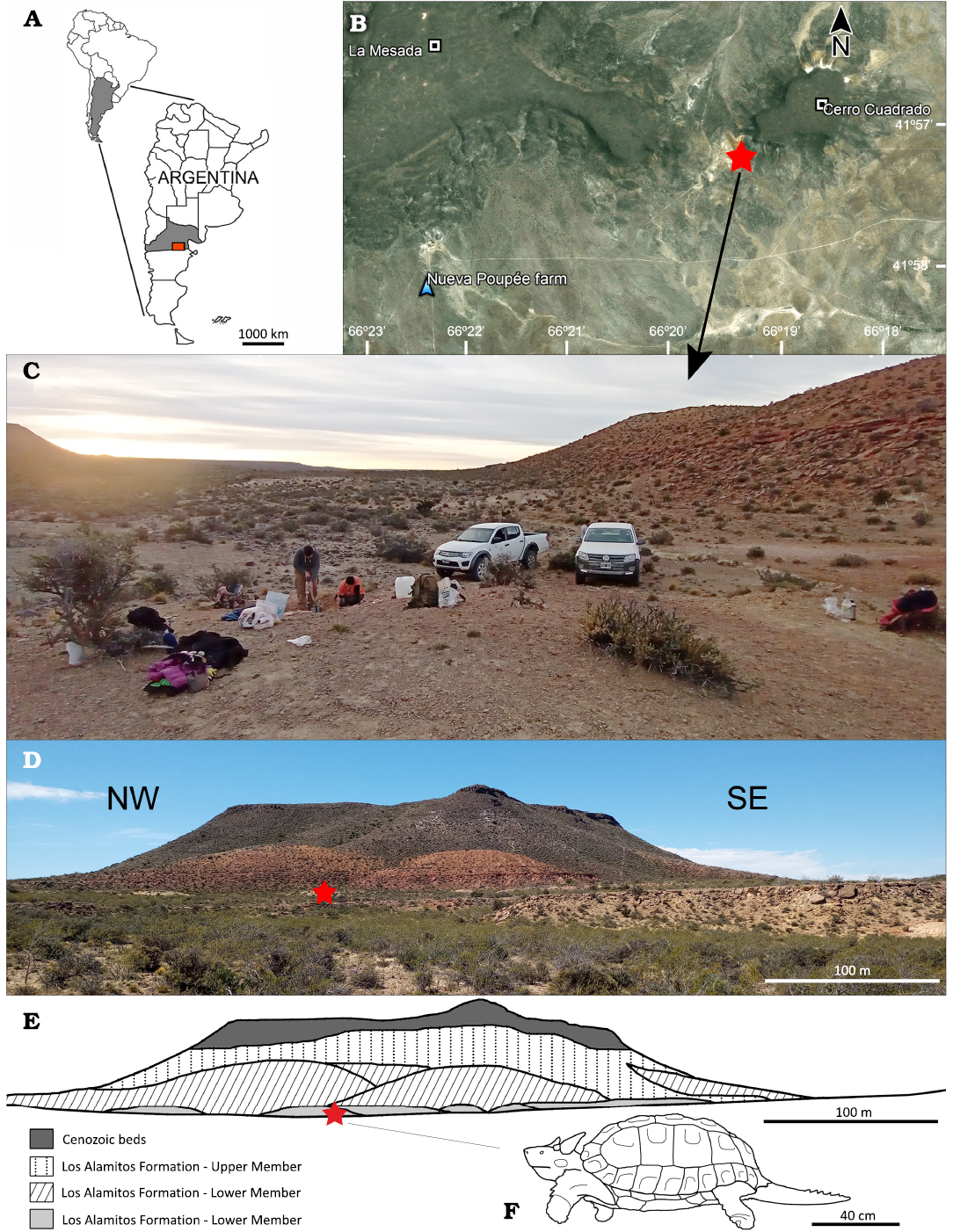
Fig. 1. A. Map showing geographic location of the study area within Argentina. B. Satellite map showing the location of Cerro Cuadrado. C. Photograph of the site showing the excavation work. D. Panoramic photograph of the discovery site. E. Line diagram showing outcrops in the type section of the Los Alamitos Formation southwest of Cerro Cuadrado. The red star marks the find site. F. Reconstruction of Patagoniaemys aeschyli sp. nov. (artwork by NRC)
Material and methods
Mechanical preparation.—The specimen was prepared using mechanical methods. The carapace was excavated and placed into two plaster jackets, which were prepared in two main stages. On the first stage a pin vise with a tungsten needle was used, removing small fragments of sediment containing some scattered materials of small vertebrates, taking advantage of the small fragments of the sediment, with the aim of getting close to the carapace elements while removing and protecting the delicate and small vertebrate remains. A second stage consisted of preparing the carapace materials using air scribes, particularly Micro Jack 3 and 4 and also the pin vise with a sharp tungsten needle and a 45° chisel shaped needle to give some detail. Because of the fragmentary state of the carapace, cyanoacrylate adhesives were applied when needed. Long bones and vertebrae were prepared mechanically with air scribes Micro Jack 3 and 4.
Comparisons.—The specimens were compared to other meiolaniforms from Patagonia, Australia, and Asia. Those comparisons were based on firsthand observations by JS (Chubutemys copelloi, Patagoniaemys gasparinae, Trapalcochelys sulcata, specimens from Los Alamitos (Peligrochelys walshae, Niolamia argentina, Gaffneylania auricularis, Otwayemys cunicularius, and Meiolania platyceps), photographs (Mongolochelys efremovi), and the literature (Gaffney 1983, 1985, 1996; Gaffney et al. 1998, 2007; Sterli and de la Fuente 2011a, 2013, 2019; Sterli 2015; Sterli et al. 2013, 2015).
For the diversity analysis, we downloaded the data from the Paleobiology Database on August 25, 2025 (see SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app71-Agnolin_etal_SOM.pdf). We downloaded the species diversity count (SOM 1) from Patagonia listing the Argentinean provinces of Mendoza, Neuquén, Río Negro, Chubut, Santa Cruz, and Tierra del Fuego. The Chilean Patagonia was not included because there are no species recognized in the Cretaceous–Paleogene. The time rule used was “major”. We also downloaded the list of occurrences at species level to account for the list of species recognized in each time bin (SOM 2). We calculated three diversity estimates following Vlachos (2024; and references therein): raw species diversity, range-through species diversity, and shareholder quorum subsampling (SQS). SQS was applied to account for uneven sampling intensity among time bins, as it standardizes samples based on coverage (the proportion of true diversity represented in a sample) rather than on the absolute number of occurrences or individuals. Coverage was calculated for each time bin. The SQS correction was calculated using the divDyn package (Kocsis et al. 2019, 2022) in R (R Core Team 2024) that calculates the SQS as in Alroy (2014). We used the sampled-in-bin richness (divSIB in the divDyn package; Koscis et al. 2022) and the SQS-corrected for 0.2 quorum levels (SOM 3). The diversity curves were calculated in Excel and subsequently edited in Adobe Illustrator for graphical purposes only to improve clarity and readability.
Systematic palaeontology
Testudinata Klein, 1760
Meiolaniformes Sterli & de la Fuente, 2013
Genus Patagoniaemys Sterli & de la Fuente, 2011a
Type species: Patagoniaemys gasparinae Sterli & de la Fuente, 2011a; Chubut Province, Argentina, Campanian–Maastrichtian.
Patagoniaemys aeschyli sp. nov.
Fig. 2–4.
ZooBank LSID: urn:lsid:zoobank.org:pub:8C16458D-1D80-4B4C-8957-21010FB6769E
Etymology: In reference to Aeschylus (525–455 BC), an ancient Greek writer often described as the father of tragedy. During a visit to the city of Gela (Italy), he was killed outside the city by a tortoise dropped by an eagle, which had mistaken his bald head for a rock suitable for shattering the shell.
Holotype: MPMIK 1839/P/33 (Figs. 2, 3, 4A1–A10, C1–C10), associated specimen including the posterior-most part of the basicranium formed by the basioccipital and parts of both exoccipitals, anterior margin of the dorsal carapace with part of the nuchal, right peripherals 1–4 and left peripherals 1–3 and part of right costal 1, three posterior peripherals, plastral bridge from the right side, including partial left hyoplastron, two highly incomplete neural arches of anterior cervical vertebrae, incomplete cervical 4, two incomplete centra of posterior cervical vertebrae, two fused thoracic vertebral centra, ?ilium, right pubis, and distal end of left humerus. The specimen was recovered partially articulated at a very restricted area (approximately 1 m2) and was collected by members of our team during a fieldtrip in 2022.
Type locality: Cerro
Cuadrado (41°57’13.5” S, 66°19’20.3” W;
582 m a.s.l.), Valcheta department, Río Negro Province, Argentina.
Type horizon: Los Alamitos Formation, Maastrichtian, Upper Cretaceous.
Material.—MPMIK 1839/P/34, isolated caudal vertebra (Fig. 4B); MACN-RN 36, left humerus (Broin 1987: pl. 1: 1); MACN-RN 37, bridge peripheral (Broin 1987: pl. 1: 2); MACN-RN 38, left anterior peripheral (Broin 1987: pl. 1: 3); MACN-RN 39, left bridge peripheral (Broin 1987: pl. 1: 4); MACN-RN 40, posterior peripheral (Broin 1987: pl. 1: 5); MACN-RN 41, left posterior peripheral (Broin 1987: pl. 1: 6); MACN-RN 63, peripheral 7 or 8; MACN-RN 64, right anterior peripheral 2 or 3; MACN-RN 66, fragment indet.; MACN-RN 67, fragment indet. or ?right bridge peripheral; MACN-RN 68, fragment indet.; MACN-RN 69, left peripheral 1 or 2; MACN-RN 70, peripheral indet.; MACN-RN 101, fragment of proximal end of left humerus; MACN-RN 709, fragment indet; MACN-RN 907, incomplete cervical 4 (Broin and de la Fuente 1993: fig. 5a–e); MACN-RN 908, incomplete cervical 8 (Broin and de la Fuente 1993: fig. 5f–j); MACN-RN 909, thoracic vertebra 10; MACN-RN 910, neural plate and waved costal; MACN-RN 918 fragment of right peripheral 1 or 2; MACN RN unnumbered, several caudal vertebrae. All from the type locality and horizon.
Diagnosis.—Large chelonian (maximum anterior carapace width about 60 cm, maximum inferred carapace length 80 cm). Referable to meiolaniforms by several features, namely the presence of small pits and grooves ornamenting carapace bones, anteriorly curved grooves among marginal scutes, opisthocoelous caudal vertebrae, and carapace-plastron connection through ligaments (Gaffney 1996; Hirayama et al. 2000; Sterli and de la Fuente 2013; Sterli et al. 2013). It is referred to Patagoniaemys by having a basioccipital with well-developed basal tubera with a concave surface between them, posteriorly flat occipital condyle, a shallowly notched anterior carapace margin, presence of a thickened bump on the posterior part of anterior marginal scutes, biconvex and narrow cervical 4 having a notably ventrally keeled centrum and disparate-sized articular surfaces with the anterior one bigger than the posterior one (Sterli and De la Fuente 2011; Oriozabala et al. 2025). P. aeschyli sp. nov. differs from the only previously known species of the genus, P. gasparinae in the presence of at least 5 longitudinal ridges on the nuchal bone (a feature unknown in other meiolaniforms) and the presence of a trapezoidal and narrow cervical scute with a posterior groove (lyra-shaped in contour in P. gasparinae; Oriozabala et al. 2025).
Description.—The description is based on the MPMIK 1839/P/33 (holotype) and MPMIK 1839/P/34 (caudal vertebra). Remaining specimens have been described elsewhere by previous authors (see de Broin 1987 and Sterli 2015).
Basicranium (Fig. 2): Only the posteriormost part of the basicranium is preserved. It is formed by the basioccipital and parts of the exoccipitals. In this part of the skull, the occipital condyle, the ventral rim of the foramen magnum, the basal tubera, and the foramina for the cranial nerve XII can be identified. The occipital condyle is kidney-shaped and it is formed by the basioccipital with contributions of both exoccipitals. The articular surface is flat. In ventral view, one of the basal tubera is preserved (the other is broken). The preserved basal tuber is well-developed latero-ventrally. In anterior view, the articular/sutural surface with the basisphenoid is partially preserved. In dorsal view, we can recognize at least two pairs of foramina nervi hypoglossi (cranial nerve XII), the foramen jugulare anterius (through which the cranial nerves X and XI exit the skull as well as the vena capitis cerebralis; Gaffney 1979), and the basis tuberculi basalis (where the bifid ligament of the medulla attaches; Kesteven 1910).
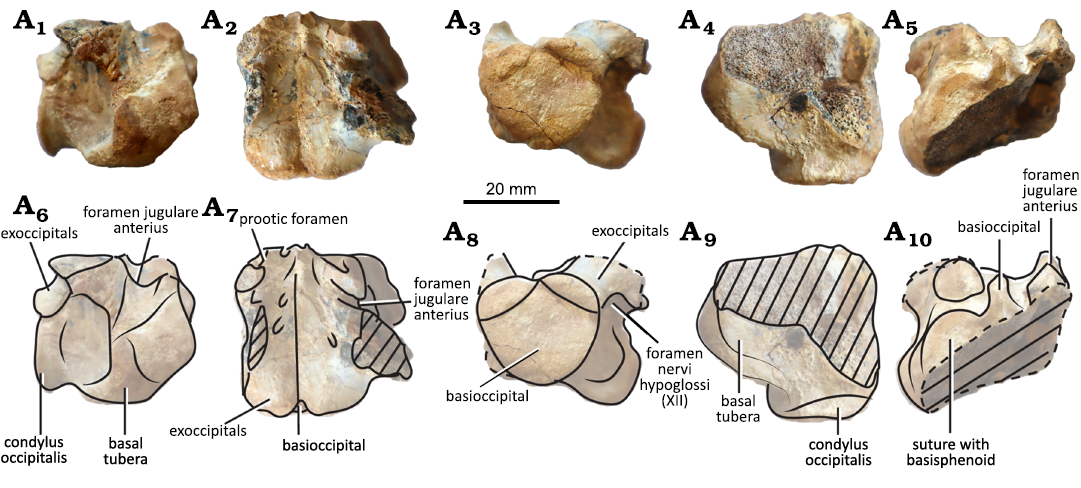
Fig. 2. Meiolaniform turtle Patagoniaemys aeschyli sp. nov. holotype (MPMIK 1839/P/33); Cerro Cuadrado locality, Argentina, Los Alamitos Formation Maastrichtian (Upper Cretaceous). Basioccipital in lateral (A1, A6), dorsal (A2, A7), posterior (A3, A8), ventral (A4, A9), and anterior (A5, A10) views. Photographs (A1–A5), explanatory drawings (A6–A10). The dashed areas correspond to missing parts.
Carapace (Fig. 3): The anterior margin of the carapace is smoothly notched. The margins of the peripheral bones are gently dorsally curved (Fig. 3A1–A4). Preserved maximum anterior carapace width is about 60 cm, with an estimated total length of about 80 cm. The carapace is relatively thick at the base of the peripheral plates, where it shows a series of robust thick bumps, especially in the posterior parts of marginal (Fig. 3A5, A6). The external surface of the carapace ornamentation is composed of small pits and grooves.
Plates: The nuchal bone is incompletely preserved, but it was relatively wide and trapezoidal in contour (Fig. 3A7, A8). The preserved contacts are with peripheral 1 and costal 1 laterally. It has at least 5 longitudinal ridges, with the middle one the most prominent. Peripheral 1 is triangular or sub-trapezoidal in shape. It contacts peripheral 2 laterally, the nuchal medially, and costal 1 posteriorly. Peripheral 2 is rectangular, being longer than wide. It contacts peripheral 1 medially, peripheral 3 laterally and costal 1 posteriorly. Peripheral 3 is squarish and contacts peripheral 2 anteriorly, costal 1 posteromedially, and peripheral 4 posteriorly. The carapace peripheral bridge starts in this peripheral, where the axillary buttress abuts. Peripheral 4 is a bridge peripheral, rectangular in contour and C-shaped in cross-section. The only preserved contact is the anterior one with peripheral 3. Only right costal 1 is partially preserved. The preserved contacts are with the nuchal anteriorly and with peripherals 1 to 3 laterally. In ventral view, remains of the thoracic rib 2 are preserved and the sutural area for thoracic rib 1 is seen. The axillary buttress seems to extend over the costal 1.
Scutes: The cervical scute was present. Its posterior sulcus is not preserved, consequently its complete shape cannot be assessed. The lateral sulci converge anteriorly. The cervical scute articulated on an inverted V-shaped protuberance made by the nuchal. Cervical contacted marginal 1 laterally, and probably vertebral 1 posteriorly. Only the lateral sulci of marginal 1 are preserved. Marginal 1 covered part of the nuchal and peripheral 1. The intermarginal sulci have a strong, anterior inflection in the rim of the carapace. Marginal 2 was squarish and contacted marginal 1 medially, marginal 3 laterally, and, at least, pleural 1 posteriorly. It covered the posterior part of peripheral 1, the tip of the nuchal, and the anterior part of peripheral 2. Marginal 3 covered the posterior part of peripheral 2 and the anterior part of peripheral 3. It contacted marginal 2 anteromedially, pleural 1 posteromedially, and marginal 4 laterally. Marginal 4 was rectangular and covered the posterior part of peripheral 3 and the anterior part of peripheral 4. It contacted marginal 3 anteriorly and marginal 5 posteriorly. Marginal 5 was covering the posterior part of peripheral 4. The only preserved contact was with marginal 4. The posterior part of marginals 1–3 covered bony protuberances of the underlaying plates.

Fig. 3. Meiolaniform turtle Patagoniaemys aeschyli sp. nov. holotype (MPMIK 1839/P/33); Cerro Cuadrado locality, Argentina, Los Alamitos Formation Maastrichtian (Upper Cretaceous). A. Carapace; anterior border in dorsal (A1, A2) and ventral (A3, A4) views (grey lines, limits of the scutes; black lines, limits of the plates; dashed lines, possible limits of the plates), anterior view of the left side of the anterior margin (A5), enlargement showing the protuberances that are typical of the genus (A6), detail of the anterior margin in ventral view (A7, A8), showing the presence of longitudinal ridges that characterize the new species (light grey, nuchal bone; dark grey, broken surfaces). B. Left hyoplastron fragment in ventral (B1) and visceral (B2) views. Photographs (A1, A3, A5–A7, B), explanatory drawings (A2, A4, A8). Scale bars 100 mm.
Plastron: The hyoplastron forms part of the axillary buttress and is represented by a sheet of bone curving distally around the forelimb opening. Its incomplete nature does not clarify whether a plastral fontanelle was present (Fig. 3B).
Vertebral column: Two incomplete neural arches of cervical
vertebrae have been preserved. There is no well-
differentiated neural spine. A nearly complete centrum of
cervical 4 is preserved (Fig. 4A). It is biconvex and is notably
transversely compressed and shows a very prominent and
narrow ventral longitudinal keel. The posterior condyle is ovoidal,
higher than wide. The anterior condyle is wider than the posterior one,
but as it is broken, its complete contour is unknown.
There are two incompletely preserved but strongly sutured (through synchondrosis) thoracic vertebral centra. They are longer than wide and hourglass-shaped in ventral view.
The caudal MPMIK 1839/P/34 is opisthocoelous (Fig. 4B). The centrum in ventral view is anteroposteriorly longer than transversely wide. The condyle is oval with its main axis horizontally oriented and the cotyle is almost round, slightly taller than wide. On its posteroventral edge, the wide and well-defined double articulation for the chevron is seen. There is a wide, longitudinal, ventral crest on the centrum. The transverse process is very thick and subquadrangular in cross-section at its base. It is subtriangular in contour and strongly anteriorly oriented. The neural crest is present but low.
Humerus: Only the distal end of a left humerus was found (Fig. 4C). The canal of the enclosed ectepicondylar foramen is seen in the capitular view. In the intertubercular view, it shows two distal condyles, one for the radius (capitellum) and one for the ulna (trochlea). The condyles are bulbous, subequal in size and extension. Both are separated by a narrow and shallow, but well-defined, intercondylar groove. The ectepicondylar foramen exits in the intertubercular view, next to the capitellum.
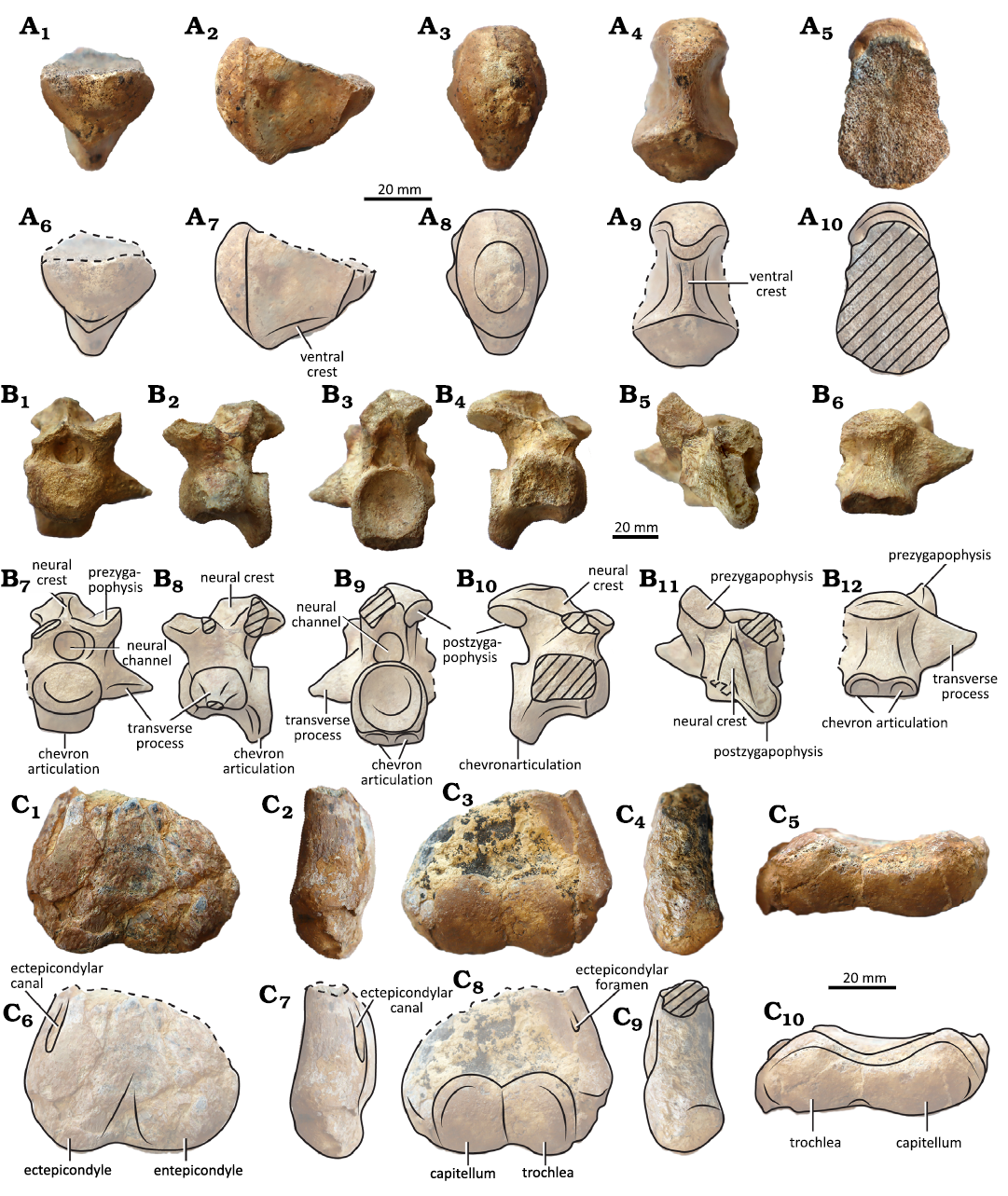
Fig. 4. Meiolaniform turtle Patagoniaemys aeschyli sp. nov., Cerro Cuadrado locality, Argentina, Los Alamitos Formation Maastrichtian (Upper Cretaceous). A. MPMIK 1839/P/33 (holotype), cervical vertebra 4 in anterior (A1, A6), lateral (A2, A7), posterior (A3, A8), ventral (A4, A9), and dorsal (A5, A10) views. B. MPMIK 1839/P/34, anterior caudal vertebra in anterior (B1, B7), right lateral (B2, B8), posterior (B3, B9), left lateral (B4, B10), dorsal (B5, B11), and ventral (B6, B12) views. C. MPMIK 1839/P/33 (holotype), distal end of left humerus in capitular (C1, C6), ulnar (C2, C7), intertubercular (C3, C8), radial (C4, C9), and distal (C5, C10) views. Photographs (A1–A5, B1–B6, C1–C5), explanatory drawings (A6–A10, B7–B12, C6–C10). The dashed areas correspond to missing parts.
Remarks.—As outlined in the diagnosis and description, we consider this new species to present a combination of unique features sufficient to distinguish it from all other known taxa. The holotype preserves much of the anterior end of the carapace, a region that bears numerous diagnostic characters in meiolaniforms. Fortunately, Campanian–Maastrichtian meiolaniforms from Patagonia, namely Trapalcochelys sulcata and Patagoniaemys gasparinae include materials that could be directly compared with the material of the new species (Sterli et al. 2013; Oriozabala et al. 2025). Among meiolaniforms, Patagoniaemys aeschyli sp. nov. shares with P. gasparinae (Oriozabala et al. 2025), and probably Trapalcochelys sulcata and Peligrochelys walshae, a shallow notch in the anterior margin of the carapace. An anteriorly notched carapace is absent in the basal forms Chubutemys copelloi and probably Otwayemys cunicularius (Gaffney et al. 1998; Sterli et al. 2015). Patagoniaemys aeschyli sp. nov. shares with P. gasparinae the presence of thickened bumps on the posterior part of anterior marginal scutes, features absent in Mongolochelys efremovi (see Sukhanov 2000; Oriozabala et al. 2025). Otherwise, Patagoniaemys aeschyli sp. nov. and P. gasparinae share with Mongolochelys efremovi the presence of a wide, trapezoidal nuchal that almost prevents the contact of peripheral 1 and costal 1. The nuchal bone of Patagoniaemys aeschyli sp. nov. differs from P. gasparinae in the presence of at least 5 longitudinal ridges and the presence of a narrow cervical scute with a posterior groove, that strongly differs from the lyra-shaped contour present in P. gasparinae (Oriozabala et al. 2025).
Patagoniaemys aeschyli sp. nov. shares with Otwayemys cunicularius and Mongolochelys efremovi, a long thoracic rib 1 that reached or almost reached the axillary buttress. In P. aeschyli sp. nov. the bridge starts in peripheral 3 close to the anterior suture with peripheral 2, as in M. efremovi. In Patagoniaemys gasparinae, the axillary buttress reached peripheral 2 in one of the sides (JS personal observations). In spite of being not completely preserved, the low and wide carapace of P. aeschyli sp. nov. differs from that of derived meiolaniids (e.g., Meiolania) which show a strongly domed carapace (Gaffney 1996; Gaffney et al. 1992; Sterli and de la Fuente 2011a).
In Patagoniaemys aeschyli sp. nov., only one basal tuber of the basioccipital is completely preserved and well developed, whereas the other is broken. Based on the morphology and orientation of the preserved tuber, we infer that the original condition involved well-separated basal tubera with a concave surface between them, as observed in some specimens of Peligrochelys walshae (Sterli and de la Fuente 2019) and Patagoniaemys gasparinae (JS personal observations based on unpublished material). In contrast, some specimens of Peligrochelys walshae (e.g., MACN-PV CH 2017) exhibit a longitudinal ridge between the basal tubera. The occipital condyle of Patagoniaemys aeschyli sp. nov. is flat posteriorly as in P. gasparinae (JS personal observations based on unpublished material) and Chubutemys copelloi (Gaffney et al. 2007; Sterli et al. 2015). The occipital condyle in P. aeschyli sp. nov. is formed by the basioccipital and both exoccipitals. Due to the fusion of basioccipital and exoccipital in the condyle area, the contribution of the exoccipitals to the occipital condyle cannot be assessed for Chubutemys copelloi, Peligrochelys walshae, Patagoniaemys gasparinae, and Mongolochelys efremovi.
A common trait shared by Patagoniaemys aeschyli sp. nov., P. gasparinae, and Peligrochelys walshae is the biconvex and narrow cervical 4 having a notably ventrally keeled centrum and disparately-sized articular surfaces (Sterli and de la Fuente 2011a). In both P. aeschyli sp. nov. and P. gasparinae the anterior condyle of cervical vertebra 4 is dorsoventrally taller and transverse wider than the posterior one. In Peligrochelys walshae both condyles of cervical vertebra 4 are similar in shape and size (Sterli and de la Fuente 2019). This morphology differs from the box-shaped condition exhibited by species of Meiolania (Gaffney 1985).
Anterior caudal vertebrae in meiolaniforms are opisthocoelous and the posteroventral end of the centra houses the articular area for the chevrons. This morphology is observed in T. sulcata, P. aeschyli sp. nov., P. gasparinae, Peligrochelys walshae, Mongolochelys efremovi and in the meiolaniids Gaffneylania auricularis and Meiolania platyceps. The lack of complete and/or undeformed specimens makes comparisons among taxa difficult. Meiolaniforms also have an enclosed ectepicondylar foramen in the humerus. This condition is seen in P. aeschyli sp. nov., P. gasparinae, Peligrochelys walshae, Mongolochelys efremovi, Gaffneylania auricularis, and Meiolania platyceps.
The distal end of the humerus in P. aeschyli sp. nov. shows a narrow distal end with poorly developed epicondyles and separated distal condyles not forming a pulley-like continuous surface, features different from terrestrial chelonians, including meiolaniids in which the distal end of the bone forms a pulley-like articulation (Walker 1973; Nakajima et al. 2014; Krahl et al. 2019; see Gaffney 1996; Sterli et al. 2015). The ectepicondyle and entepicondyle are subequal in size and are much more transversely narrow than in Peligrochelys walshae (Sterli and de la Fuente 2019). Due to the fragmentary nature of the humerus in P. aeschyli sp. nov., no further comparisons are possible.
Stratigraphic and geographic range.—Cerro Cuadrado locality, Argentina, Los Alamitos Formation Maastrichtian (Upper Cretaceous).
Discussion
The discovery of meiolaniids in South America and Australia in the 19th century sparked a great debate. The discovery of such large and terrestrial forms on now separated landmasses was taken as a proof of ancient connection between the continents by Moreno (1899). This view was strongly criticized by Simpson (1938, 1939) who indicated that the common presence of meiolaniids was the result of parallel invasions from North America and Asia, respectively. Under this scheme, there occurred a single, or just a few, old biotic dispersal events between Australia and South America (e.g., Simpson 1980; Pascual et al. 1992; Szalay and Sargis 2001; Meredith et al. 2009; Wilf et al. 2013).
On the other hand, the common occurrence of basal meiolaniforms and derived meiolaniids in Patagonia and Australia is congruent with other faunistic evidence supporting the strong biotic ties between both landmasses during the Cenozoic (see Gaffney 1996; Beck 2012; Sterli et al. 2013; Ezcurra and Agnolín 2012; Agnolín 2024), and puts into question the occurrence of a single or a few dispersal events between both landmasses.
Vlachos et al. (2018) suggested that turtles in South America were seriously affected by the K/Pg mass extinction, in contrast with what is observed for North American turtles (e.g., Holroyd and Hutchison 2002; Hutchison and Holroyd 2003; Lyson et al. 2011; Holroyd et al. 2014). According to Archibald (1996), this turtle fauna includes a mix of extinct (Adocidae, Baenidae, Macrobaenidae, Pleurosternidae, Nanhsuingchelydidae) and extant (e.g., Chelydridae, Kinosternidae, Trionychidae) lineages, with most extinctions occurring in strictly terrestrial lineages, whereas aquatic ones were poorly affected by the K/Pg event. Explanations for such differences range from authors who suggested that the turtles were able to survive because aquatic environments are more heterogeneous than terrestrial ones, allowing some taxa to survive in natural refuges (Holroyd and Hutchison 2002), or contrarily, because they are much more stable and are not strongly affected by large-scale extinction events (Bakker 1977). In any case, aquatic and semiaquatic turtles seem to be more likely to survive mass extinctions. In the analyzed raw diversity of Cretaceous–Paleogene chelonians from Patagonia (Fig. 5), there is a peak of diversity in the Cenomanian with 4 species and then, the diversity drops during the “mid-Cretaceous”. In the Campanian, the diversity increases again to 7 species, then descends to 4 in the Maastrichtian, increasing slightly again in the Danian with 5 species. In the raw diversity diagram, the K/Pg extinction event is not remarkable, because the major drop is seen between the Campanian and Maastrichtian. This drop, and later stabilization between the Maastrichtian and Danian, is also recovered in the range-through diversity and in the SQS 0.2 analysis (Fig. 5; SOM 3). Although raw and corrected diversities were used herein, it is interesting to note that Vlachos (2024) noticed that the coverage for the Campanian, Maastrichtian, and Danian is similar (0.88, 0.87, and 0.82, respectively).
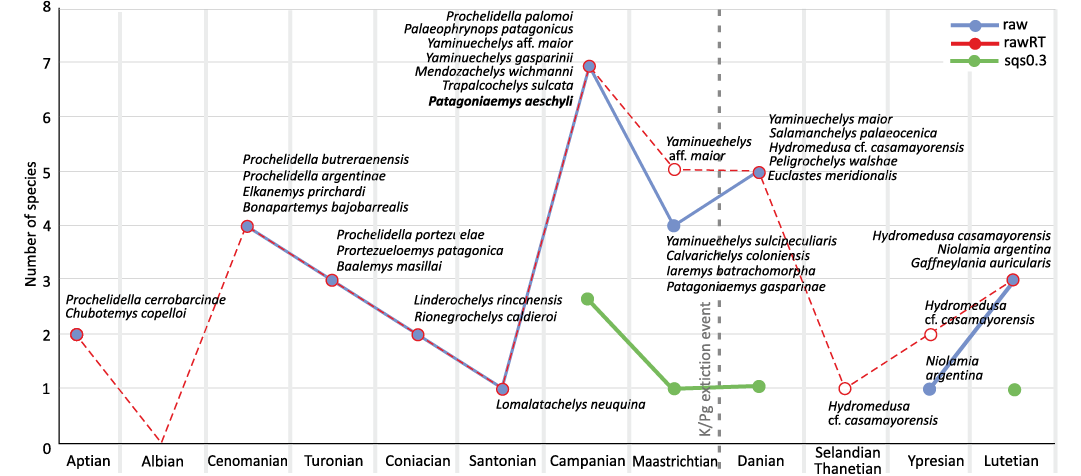
Fig. 5. Raw, range-through, and SQS 0.2 corrected species’ diversity of turtles from the Cretaceous to the Paleogene in Patagonia. Data extracted from the Paleobiology Database (August 25, 2025).
In this regard, it is worth mentioning that similarities between chelonian assemblages of the uppermost Cretaceous (Maastrichtian Allen, La Colonia, and Los Alamitos Formations) and the Lower Paleocene (Danian Banco Negro Inferior of the Salamanca Formation and Roca Formation) strata are impressive (Fig. 5). In the Banco Negro Inferior, chelonians are represented by the chelids Yaminuechelys maior (Staesche, 1929), cf. Hydromedusa sp. and Salamanchelys palaeocenica Bona, 2006 (Bona and de la Fuente 2005; Bona 2006; Maniel and de la Fuente 2016), and meiolaniforms by Peligrochelys walshae (Sterli et al. 2015). All these forms have representatives in Maastrichtian deposits, including Yaminuechelys species (present in Allen, La Colonia, and Los Alamitos formations; Broin 1987; Broin and de la Fuente 1993; de la Fuente et al. 2014; Gasparini et al. 2015; Sterli et al. 2021). Salamanchelys is very similar to indeterminate chelids described for these Maastrichtian units (Bona 2006), cf. Hydromedusa sp. was also reported previously in latest Cretaceous assemblages (e.g., Broin 1987), and Peligrochelys is very closely related to Patagoniaemys (Sterli et al. 2015). This is also supported by the fact that at least two lineages of meiolaniform turtles survived the K/Pg boundary, one including derived meiolaniids and the other includes large basal meiolaniforms (e.g., Patagoniaemys and Peligrochelys; Sterli and de la Fuente 2013; Sterli et al. 2015).
The substantial number of taxa shared between diverse Maastrichtian localities and the Paleogene Punta Peligro site have been previously noted for plants, mammals and reptiles (Stiles et al. 2020; Agnolín 2024; Bravo et al. 2025). This pattern suggests that the K/Pg extinction event may not have severely impacted certain lineages, particularly chelonians, and further supports the interpretation that the Punta Peligro site represents one of the earliest Paleogene localities in South America (see del Campo et al. 2025).
Conclusions
The material from the Maastrichtian Los Alamitos Formation supports the recognition of Patagoniaemys aeschyli sp. nov. as a new basal meiolaniform that expands both the diversity and morphological disparity of the clade in the Late Cretaceous of Patagonia. The combination of cranial, vertebral, and humeral features (particularly the distinctive longitudinal ridges on the nuchal) substantiates the taxonomic validity of the new species and confirms the presence of at least two meiolaniform lineages in the latest Cretaceous of southern South America. the marked taxonomic continuity between Maastrichtian and Danian assemblages suggests that the K/Pg extinction event did not severely impact Patagonian turtles, supporting a scenario of lineage persistence and limited faunal turnover among southern chelonians.
Acknowledgements
We deeply thank the crew that formed part of the expedition to the Los Alamitos Formation, including Jordi García-Marsà, Ana Moreno Rodríguez, Santiago Miner, David Piazza, Marcelo Isasi, and Gerardo Álvarez-Herrera (all MACN). Special thanks to Sergio Collazo (MACN), who skillfully prepared the specimen. The holotype of the new species was found by A. Moreno Rodríguez and S. Miner, and specimen MPMIK 1839/P/34 was found by A. Moreno Rodríguez. We thank Romina Rial (Museo Provincial “María Inés Kopp”, Valcheta town, Río Negro Province, Argentina) for their valuable help in the museum. The Secretariat of State for Culture of the Province of Río Negro (especially Mariano Zacharonok, Romina Brunoni, and Pamela Romero) provided valuable support for the paleontological explorations carried out here. Special thanks to José Chaquib Hamzé, Wadid Rechene, and José Manuel Hamzé Capaccioni, the owners of Nueva Poupée (Valcheta department, Rio Negro province, Argentina), who kindly allowed camping on their farm. We thank all the enterers and authorizers that contributed to the Paleobiology Database for uploading the data used herein. We thank Evangelos Vlachos (Museo Paleontológico Egidio Feruglio, Trelew, Chubut, Argentina) who helped in the calculation of the SQS corrected diversity. The field trip was financed by the Mamont Scholar Grant 2020 program of The Explorers Club to NC, and NGS-92822R-22 “The end of the Dinosaur Era in Patagonia” to Diego Pol. We would like to thank the reviewers of this manuscript, Ignacio Maniel (Instituto de Evolución, Ecología Histórica y Ambiente, San Rafael, Mendoza, Argentina) and Walter Joyce (University of Fribourg, Switzerland), as well as the editor, Daniel Barta (Oklahoma State University College of Osteopathic Medicine at the Cherokee Nation, Tahlequah, USA), whose comments and suggestions greatly improved the manuscript.
Editor: Daniel Barta
References
Agnolín, F. 2024. History of Cenozoic Mammals from South America: A New Model. 318 pp. Springer Nature Switzerland AG, Cham. Crossref
Agnolín, F.L. and Martinelli, A.G. 2009. Fossil birds from the Late Cretaceous Los Alamitos Formation, Río Negro province, Patagonia. Journal of South American Earth Sciences 27: 42–49. Crossref
Agnolín, F.L., Rolando, A.M.A., Chimento, N.R., and Novas, F.E. 2024. New small reptile remains from the Late Cretaceous of Patagonia increase morphological diversity of sphenodontids (Lepidosauria). Proceedings of the Geologists’ Association 135: 36–44. Crossref
Albino, A. 1987. The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Part V. Ophidians. In: J.F. Bonaparte (coord.), The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 3 (3): 141–146.
Albino, A. 2000. New record of snakes from the Cretaceous of Patagonia (Argentina). Geodiversitas 22: 247–253.
Alroy, J. (2014). Accurate and precise estimates of origination and extinction rates. Paleobiology 40: 374–397. Crossref
Andreis, R.R., 1987. Stratigraphy and paleoenvironment. The late Cretaceous fauna of Los Alamitos, Patagonia, Argentina. In: J.F. Bonaparte (coord.), The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 3 (3): 103– 110.
Anquetin, J. 2011. Evolution and palaeoecology of early turtles: a review based on recent discoveries in the Middle Jurassic. Bulletin de la Société géologique de France 182: 231–240. Crossref
Apesteguía, S. 2005. A late Campanian sphenodontid (Reptilia, Diapsida) from northern Patagonia. Comptes Rendus Palevol 4: 663–669. Crossref
Archibald, J.D. 1996. Dinosaurs—Extinction and the End of an Era. What the Fossils Say. 237 pp. Columbia University Press, New York.
Báez, A.M. 1987. Anurans. In: J.F. Bonaparte (coord.), The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 3 (3): 121–130.
Beck, R.M. 2012. An ‘ameridelphian’ marsupial from the early Eocene of Australia supports a complex model of Southern Hemisphere marsupial biogeography. Naturwissenschaften 99: 715–729. Crossref
Bakker, R.T. 1977. Tetrapod mass extinctions- model of the regulation of speciation rates and immigration by cycles of topographic diversity. Developments in Palaeontology and Stratigraphy 5: 439–468. Crossref
Bona, P. 2006. Paleocene (Danian) chelid turtles from Patagonia, Argentina: taxonomic and biogeographic implications. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 241: 303–323. Crossref
Bona, P. and de la Fuente, M.S. 2005. Phylogenetic and paleobiogeographic implications of Yaminuechelys maior (Staesche, 1929) new comb., a large long-necked chelid turtle from the early Paleocene of Patagonia, Argentina. Journal of Vertebrate Paleontology 25: 569–582. Crossref
Bonaparte, J.F. (coord.). 1987. The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 3 (3).76 pp. Museo Argentino de Ciencias Naturales, Buenos Aires.
Bonaparte, J.F. 2002. New Dryolestida (Theria) from the Late Cretaceous of Los Alamitos Formation, Argentina, and paleogeographical comments. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 224: 339–371. Crossref
Bonaparte, J.F., Franchi, M.R., Powell, J.E., and Sepulveda, E.G. 1984. La Formación Los Alamitos (Campaniano–Maastrichtiano) del sudeste de Río Negro, con descripción de Kritosaurus australis n. sp. (Hadrosauridae). Significado paleogeográfico de los vertebrados. Revista de la Asociación Geológica Argentina 39 (3–4): 284–299.
Bravo, G.G., Pol, D., Leardi, J.M., Krause, J.M., Nicholl, C.S., Rougier, G., and Mannion, P.D. 2025. A new notosuchian crocodyliform from the Early Palaeocene of Patagonia and the survival of a large-bodied terrestrial lineage across the K/Pg mass extinction. Proceedings of the Royal Society B 292 (2043): 20241980. Crossref
Broin, F. de 1987. The Late Cretaceous fauna of Los Alamitos, Patagonia, Argentina. Chelonia. In: J.F. Bonaparte (coord.), The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 3 (3): 131–178.
Broin, F. de and de la Fuente, M. S. 1993. Les tortues fossiles d’Argentine: synthèse. Annales de Paléontologie 79: 169–232.
Cione, A.L. 1987. The Late Cretaceous fauna of los Alamitos, Patagonia, Argentina. The fishes. In: J.F. Bonaparte (coord.), The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 3 (3): 111–120.
Ckhikvadzé, V.M. 1987. Sur la classification et les caractères de certaines tortues fossiles d’Asie, rares et peu etudiées. Studia Palaeocheloniologica 2: 55–86
de la Fuente, M.S., Sterli, J., and Maniel, J.I. 2014. Origin, Evolution and Biogeographic History of South American Turtles. 168 pp. Springer Nature Switzerland AG, Cham. Crossref
del Campo, E.N., Chimento, N.R., and
Agnolín, F.L. 2025. Postcranial remains of Eutherian mammals from Punta
Peligro (Paleocene), Patagonia, Argentina. Historical
Biology. [published online https://doi.org/ Crossref
10.1080/08912963.2025.2537151]
Ezcurra, M.D. and Agnolín, F.L. 2012. A new global palaeobiogeographical model for the late Mesozoic and early Tertiary. Systematic Biology 61: 553–566. Crossref
Gaffney, E.S. 1979. Comparative cranial morphology of the recent and fossil turtles. Bulletin of the American Museum of Natural History 164: 65–375.
Gaffney, E.S. 1983. The cranial morphology of the extinct horned turtle Meiolania platyceps, from the Pleistocene of Lord Howe Island, Australia. Bulletin of the American Museum of Natural History 175: 361–480.
Gaffney, E.S. 1985. The cervical and caudal vertebrae of the cryptodiran turtle, Meiolania platyceps, from the Pleistocene of Lord Howe Island, Australia. American Museum Novitates 2805: 1–22.
Gaffney, E.S. 1996. The postcranial morphology of Meiolania platyceps and a review of the Meiolaniidae. Bulletin of the American Museum of Natural History 229: 1–165.
Gaffney, E.S., Archer, M., and White, A. 1992. Warkalania, a new meiolaniid turtle from the Tertiary Riversleigh deposit of Queensland, Australia. The Beagle, Records of the Northern Territory Museum of Arts and Sciences 9: 35–48. Crossref
Gaffney, E.S., Kool, L., Brinkman, D.B., Rich, T.H., and Vickers-Rich, P. 1998. Otwayemys, a new cryptodiran turtle from the early Cretaceous of Australia. American Museum Novitates 3233: 1–28.
Gaffney, E.S., Rich, T.H., Vickers-Rich, P., Constantine, A., Vacca, R., and Kool, L. 2007. Chubutemys, a new eucryptodiran turtle from the Early Cretaceous of Argentina, and the relationships of the Meiolaniidae. American Museum Novitates 3599: 1–35. Crossref
Gasparini, Z. and de la Fuente, M.S. 2000. Tortugas y plesiosaurios de la Formación La Colonia (Cretácico Superior) de Patagonia. Revista Española de Paleontología 15: 23–35. Crossref
Gasparini, Z., Sterli, J., Parras, A., O’Gorman, J.P., Salgado, L., Varela, J., and Pol, D. 2015. Late Cretaceous reptilian biota of the La Colonia Formation, central Patagonia, Argentina: occurrences, preservation and paleoenvironments. Cretaceous Research 54: 154–168. Crossref
Gómez, R.O. 2016. A new pipid frog from the Upper Cretaceous of Patagonia and early evolution of crown-group Pipidae. Cretaceous Research 62: 52–64. Crossref
Hirayama, R., Brinkman, D.B., and Danilov, I.G. 2000. Distribution and biogeography of non-marine Cretaceous turtles. Russian Journal of Herpetology 7: 181–198.
Holroyd, P.A. and Hutchison, J.H. 2002. Patterns of geographic variation in latest Cretaceous vertebrates: Evidence from the turtle component. Geological Society of America, Special Paper 361: 177–190. Crossref
Holroyd, P.A., Wilson, G.P., and Hutchison, J.H. 2014. Temporal changes within the latest Cretaceous and early Paleogene turtle faunas of northeastern Montana. Geological Society of America, Special Paper 503: 299–312. Crossref
Hutchison, J.H. and Holroyd, P.A. 2003. Late Cretaceous and early Paleocene turtles of the Denver Basin, Colorado. Rocky Mountain Geology 38: 121–142. Crossref
Joyce, W.G. 2007. Phylogenetic relationships of Mesozoic turtles. Bulletin of the Peabody Museum of Natural History 48: 3–102. Crossref
Joyce, W.G., Rabi, M., Clark, J.M., and Xu, X. 2016. A toothed turtle from the Late Jurassic of China and the global biogeographic history of turtles. BMC Evolutionary Biology 16 (1): 1–29. Crossref
Kesteven, H.L. 1910. The anatomy of the head of Chelone midas. Proceedings of the Royal Society of Yew South Wales 44: 368–400. Crossref
Khozatskii, L.I. 1997. Large turtles from the Late Cretaceous of Mongolia. Russian Journal of Herpetology 4: 148–154. Crossref
Klein, J.T. 1760. Klassification und kurze Geschichte der vierfüβigen Thiere. 381 pp. Jonas Schmidt, Lübeck.
Kocsis, Á.T., Reddin, C.J., Alroy, J., and Kiessling, W. 2019. The R package divDyn for quantifying diversity dynamics using fossil sampling data. Methods in Ecology and Evolution 10: 735–743. Crossref
Kocsis, Á.T., Reddin, C.J., and Kiessling, W. 2022. Handout to the R package divDyn v0. 8.2 for diversity dynamics using fossil sampling data. Available at https://cran.r-project.org
Krahl, A., Lipphaus, A., Witzel, U., Sander, P.M., Maffucci, F., and Hochscheid, S. 2019. Humerus osteology, myology, and finite element structure analysis of Cheloniidae. The Anatomical Record 303: 2177–2191. Crossref
Lyson, T.R., Joyce, W.G., Knauss, G.E., and Pearson, D.A. 2011. Boremys (Testudines, Baenidae) from the latest Cretaceous and early Paleocene of North Dakota: an 11-million-year range extension and an additional K/T survivor. Journal of Vertebrate Paleontology 31: 729–737. Crossref
Maniel, I.J. and de la Fuente, M.S. 2016. A Review of the Fossil Record of Turtles of the Clade Pan-Chelidae. Bulletin of the Peabody Museum of Natural History Yale University 57: 191–227. Crossref
Meredith, R.W., Krajewski, C., Westerman, M., and Springer, M.S. 2009. Relationships and divergence times among the orders and families of Marsupialia. Museum of Northern Arizona Bulletin 65: 383–406.
Moreno, F.P. 1889. Breve reseña de los progresos del Museo de la Plata, durante el segundo semestre de 1888. Boletín del Museo de La Plata 2: 1–44.
Moyano-Paz, D., Rozadilla, S., Agnolín, F., Vera, E., Coronel, M.D., Varela, A.N., Gómez-Dacal, A.R., Aranciaga Rolando, A.M., D’Angelo, J.S., Pérez-Loinaze, V., Richiano, S., Chimento, N.R., Motta, M.J., Sterli, J., Manabe, M., Takanobu, T., Isasi, M.P., Poiré, D.G., and Novas, F.E. 2022. The uppermost Cretaceous continental deposits at the southern end of Patagonia, the Chorrillo Formation case study (Austral–Magallanes Basin): Sedimentology, fossil content and regional implications. Cretaceous Research 130: 105059. Crossref
Nakajima, Y., Hirayama, R., and Endo, H. 2014. Turtle humeral microanatomy and its relationship to lifestyle. Biological Journal of the Linnean Society 112: 719–734. Crossref
Nopcsa, F. 1923. On the geological importance of the primitive reptilian fauna of the uppermost Cretaceous of Hungary; with a description of a new tortoise (Kallokibotion). Quarterly Journal of the Geological Society 79: 100–116. Crossref
Oriozabala, C., de la Fuente, M.S., and Sterli, J. 2025. The meiolaniform Patagoniaemys gasparinae from the Upper Cretaceous La Colonia Formation (Chubut, Argentina), Postcranial anatomy and paleoecology. Journal of Vertebrate Paleontology 45: e2554253.
Panzeri, K.M., Bogan, S., Raigemborn, M.S., Pagani, M.A., Chimento, N., Agnolín, F.L., and Martinelli, A.G. 2024. Purported crushing teeth of actinopterygian fishes from the Late Cretaceous of Patagonia reinterpreted as crustacean bio-gastroliths. Journal of South American Earth Sciences 149: 105179. Crossref
Pascual, R., Archer, M., Jaureguizar, E.O., Prado, J.L., Godthelp, H., and Hand, S.J. 1992. First discovery of monotremes in South America. Nature 356: 704–706. Crossref
Pérez-García, A. and Codrea, V. 2018. New insights on the anatomy and systematics of Kallokibotion Nopcsa, 1923, the enigmatic uppermost Cretaceous basal turtle (stem Testudines) from Transylvania. Zoological Journal of the Linnean Society 182: 419–443. Crossref
Powell, J.E. 1987. The Late Cretaceous fauna of Los Alamitos, Patagonia, Argentina. VI: The titanosaurids. In: J.F. Bonaparte (coord.), The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 3 (3): 147–153.
R Core Team. 2024. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
Rougier, G.W., Martinelli, A.G., and Forasiepi, A.M. 2021. Mesozoic Mammals from South America and Their Forerunners. 388 pp. Springer Nature Switzerland AG, Cham. Crossref
Rozadilla, S., Brissón-Egli,
F., Agnolín, F.L., Aranciaga Rolando, A.M., and Novas, F.E. 2021. A
new hadrosaurid (Dinosauria: Ornithischia) from the Late Cretaceous of
northern Patagonia and the radiation of South American hadrosaurids. Journal of Systematic Palaeontology 19:
1207–1235. Crossref
Simpson, G.G. 1938. Crossochelys, Eocene horned turtle from Patagonia. Bulletin of the American Museum of Natural History 74: 221–254.
Simpson, G.G. 1939. Antarctic as a faunal migration route. Proceedings, Sixth Pacific Congress 2: 755–768.
Simpson, G.G. 1980. Splendid Isolation. The Curious History of South American Mammals. 266 pp. Yale University Press, New Haven.
Staesche, K. 1929. Schildkrötenreste aus der oberen Kreide Patagoniens. Paläontographica 72: 103–112.
Sterli, J. 2008. A new,
nearly complete stem turtle from the Jurassic of South America with
implications for turtle evolution. Biology
Letters 4: 286–289. Crossref
Sterli, J. 2015. A review of the fossil record of Gondwanan turtles of the clade Meiolaniformes. Bulletin of the Peabody Museum of Natural History 56: 21–45. Crossref
Sterli, J. and de la Fuente, M.S. 2011a. A new turtle from the La Colonia Formation (Campanian–Maastrichtian), Patagonia, Argentina, with remarks on the evolution of the vertebral column in turtles. Palaeontology 54: 63–78. Crossref
Sterli, J. and de la Fuente, M.S. 2011b. Re-description and evolutionary remarks on the Patagonian horned turtle Niolamia argentina Ameghino, 1899 (Testudinata; Meiolaniidae). Journal of Vertebrate Paleontology 31: 1210–1229. Crossref
Sterli, J. and de la Fuente, M.S. 2013. New evidence from the Palaeocene of Patagonia (Argentina) on the evolution and palaeo-biogeography of Meiolaniformes (Testudinata, new taxon name). Journal of Systematic Palaeontology 11: 835–852. Crossref
Sterli, J. and de la Fuente, M. S. 2019. Cranial and post-cranial remains and phylogenetic relationships of the Gondwanan meiolaniform turtle Peligrochelys walshae from the Paleocene of Chubut, Argentina. Journal of Paleontology 93: 798–821. Crossref
Sterli, J., de la Fuente, M.S., and Cerda, I.A. 2013. A new species of meiolaniform turtle and a revision of the Late Cretaceous Meiolaniformes of South America. Ameghiniana 50: 240–256. Crossref
Sterli, J., de la Fuente, M.S. and Umazano, A.M. 2015. New remains and new insights on the Gondwanan meiolaniform turtle Chubutemys copelloi from the Lower Cretaceous of Patagonia, Argentina. Gondwana Research 27: 978–994. Crossref
Sterli, J., Vlachos, E., Krause, M., Puerta, P., and Oriozabala, C. 2021. Contribution to the diversity of the fossil record of turtles (Testudinata) from Chubut province (Argentina) and its significance in understanding the evolution of turtles in southern South America. Publicación Electrónica de la Asociación Paleontológica Argentina 21 (1): 118–160.
Stiles, E., Wilf, P., Iglesias, A., Gandolfo, M. A., and Cúneo, N. R. 2020. Cretaceous–Paleogene plant extinction and recovery in Patagonia. Paleobiology 46: 445–469. Crossref
Sukhanov, V.B. 2000. Mesozoic turtles of middle and central Asia. In: M.J. Benton, M.A. Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 309–367. Cambridge University Press, Cambridge.
Szalay, F.S. and Sargis, E.J. 2001. Model-based analysis of postcranial osteology of marsupials from the Palaeocene of Itaboraí (Brazil) and the phylogenetics and biogeography of Metatheria. Geodiversitas 23: 139–302.
Vlachos, E. 2024. Turtle species extinction across the Cretaceous/Paleogene boundary. Paleobiology 50: 641–647. Crossref
Vlachos, E., Randolfe, E., Sterli, J., and Leardi, J.M. 2018. Changes in the diversity of turtles (Testudinata) in South America from the late Triassic to the present. Ameghiniana 55: 619–643. Crossref
Walker, W.F. 1973. The locomotor apparatus of testudines. In: C. Gans and T.S. Parsons (eds.), Biology of Reptilia Vol. 4, 1–100. Academic Press, New York.
Wilf, P., Cúneo, N.R., Escapa, I.H., Pol, D., and Woodburne, M.O. 2013. Splendid and seldom isolated: the paleobiogeography of Patagonia. Annual Review of Earth and Planetary Sciences 41: 561–603. Crossref
Acta Palaeontol. Pol. 71 (1): 173–184, 2026
http://doi.org/10.4202/app.01268.2025