
Middle Devonian and Frasnian bryozoan fauna from the Holy Cross Mountains, Poland
ANDREJ ERNST, GRZEGORZ RACKI, and PATRICK N. WYSE JACKSON
Ernst, A., Racki, G., and Wyse Jackson, P.N. 2026. Middle Devonian and Frasnian bryozoan fauna from the Holy Cross Mountains, Poland. Acta Palaeontologica Polonica 71 (1): 1–17.
The present paper describes a bryozoan fauna from the Devonian of Holy Cross Mountains (Poland) which includes nine species. Six species were described from the Dobruchna Member of the Skały Formation (Eifelian) of Skały section: Cyclotrypa sp., Fistuliphragma gracilis, Leioclema passitabulatum, Intrapora leunisseni, Laxifenestella sp., Hemitrypella nodulosa. The cryptostome species Bigeyella indigena was identified from the Laskowa Góra Beds (upper Givetian) of Józefka Hill at Górno. The fenestrate Rectifenestella sp. is described from the lower Frasnian Kowala Formation (Kadzielnia Member) of Bolechowice and Kielce. The new monotypic fenestrate genus Juanopora elegans gen. et sp. nov. is described from the Wietrznia Beds (upper Givetian) of Wietrznia-I quarry at Kielce. The described bryozoans show palaeobiogeographical connections to the Middle–Upper Devonian of Germany, Spain, and Armenia. The bryozoan faunas of the Holy Cross Mountains are in accordance with the worldwide acme of the sessile filter-feeding benthos during the Givetian age, followed by a two-stage decline in diversity due to the Taghanic and Frasnian crises.
Key words: Bryozoa, taxonomy, evolution, palaeobiogeography, Devonian, Poland.
Andrej Ernst [Andrej.Ernst@uni-hamburg.de; ORCID: https://orcid.org/0000-0002-1299-2559 ], Institut für Geologie, Universität Hamburg, D-20146 Hamburg, Germany.
Grzegorz Racki [grzegorz.racki@us.edu.pl; ORCID: https://orcid.org/0000-0003-4609-8341 ], Institute of Earth Sciences, Department of Natural Sciences, University of Silesia in Katowice, Będzińska str. 58, PL 41-200 Sosnowiec, Poland.
Patrick N. Wyse Jackson [wysjcknp@tcd.ie; ORCID: https://orcid.org/0000-0001-5605-0670 ], Department of Geology, Trinity College, Dublin 2, Ireland.
Received 11 August 2025, accepted 18 November 2025, published online 19 February 2026.
Copyright © 2026 A. Ernst et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
In the Middle to Upper Devonian carbonate succession of the Holy Cross Mountains in south-central Poland (Fig. 1A), bryozoans are mostly a minor component in numerous benthic assemblages, which are usually dominated by brachiopods. Although reported in several publications (e.g., Pajchlowa 1957; Racki et al. 1985; Racki 2024), since the monographic work of Gürich (1896) this fossil group has been taxonomically studied exclusively from open shelf facies of the northern Łysogóry Region by Kiepura (1965, 1973). Research was later significantly extended to the southern (Kielce) carbonate platform, and included many localities representing lithology variations from clayey-marly strata to stromatoporoid-coral reef-type limestones (Morozova et al. 2002).
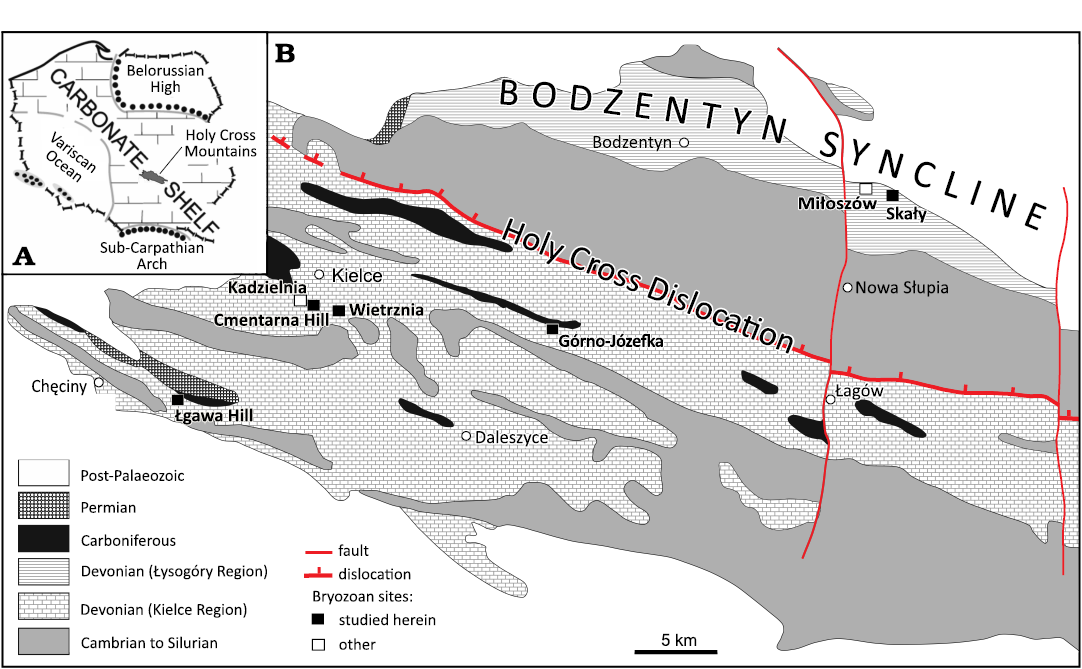
Fig. 1. Location of bryozoan sites in Poland (A) and the Holy Cross Mountains (B, based on Racki 1993: fig. 2) against paleogeography and regional geology; see Fig. 2 for refined stratigraphic-facies setting.
The present contribution is a continuation of the work of Morozova et al. (2002), based on new specimens located in museum collections and on samples more recently collected during fieldwork undertaken by the second author and students of the University of Silesia. Its main objective is to provide a taxonomic analysis of bryozoan material from five Middle Devonian and Frasnian sites located in the north-western part of the Holy Cross Mountains (Figs. 1B and 2), including providing important data from the famous Eifelian “brachiopod shale” of the Skały Formation (Pajchlowa 1957; Racki et al. 2022). The nine taxa described here include a new monotypic genus of fenestellids. The migration history of the bryozoan faunas during gradual drowning of the Kielce carbonate platform (Fig. 2) is updated, as is information pertaining to their regional succession and zoogeographical affinities with Western Europe, especially the Eifel Mountains.
Institutional abbreviations.—GIUS, Institute of Earth Sciences of the University of Silesia in Katowice, Sosnowiec, Poland.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in Zoobank: urn:lsid:zoobank.org:pub:80C55D6F-6D65-40A6-AAE6-D26510271F92
Material and methods
The studied material comprises five samples containing loose fragments of bryozoan colonies. The friable marly shale and compact limestone samples come from following localities in Holy Cross Mts, Poland (Fig. 1B): Skały; Górno, Józefka Hill; Kielce, Wietrznia-I quarry; Bolechowice, Łgawa Hil; and Kielce, Cmentarna Hill (for details, see the section on the geological setting).
Thin sections were made from the studied samples. The separate fragments were imbedded in epoxide resin (SpeciFix-20), and then cut and polished. In total, 46 thin sections were made (24 × 48 mm). The studied material is deposited at the Institute of Earth Sciences of the University of Silesia in Katowice, Sosnowiec, Poland.
Bryozoan morphology has been studied using a binocular microscope. Morphologic character terminology is partly adopted from Boardman (1960) and Anstey and Perry (1970) for trepostomes, from Hageman (1991a, b) and Snyder (1991) for fenestrates. The following morphologic characters were measured and used for statistics in the studied material: branch width, exo- (endo-) zone width, axial ratio (ratio of endozone width to the branch width), autozooecial aperture width, aperture spacing (along branch, diagonally), acanthostyle diameter, exilazooecia or metazooecia width, wall thickness in exozone, vesicle diameter, number of vesicles per aperture, dissepiment width, fenestrule width (length), distance between branch (dissepiment) centres, number of apertures per fenestrule length, maximal chamber width, width of superstructure openings. The spacing of structures is measured as a distance between their centres. Statistics were summarized using arithmetic mean, sample standard deviation, coefficient of variation, and minimum and maximum values.
Geological setting
The Middle to Upper Devonian epicontinental succession in the Holy Cross Mountains records a continuous stepwise drowning of an increasingly differentiated, vast bank-to-reef platform (Racki 1993; Szulczewski 1995; Narkiewicz et al. 2006; Racki et al. 2022), in general agreement with the transgressive-regressive cyclicity of the Euramerican eustatic curve of Johnson et al. (1985; Fig. 2).
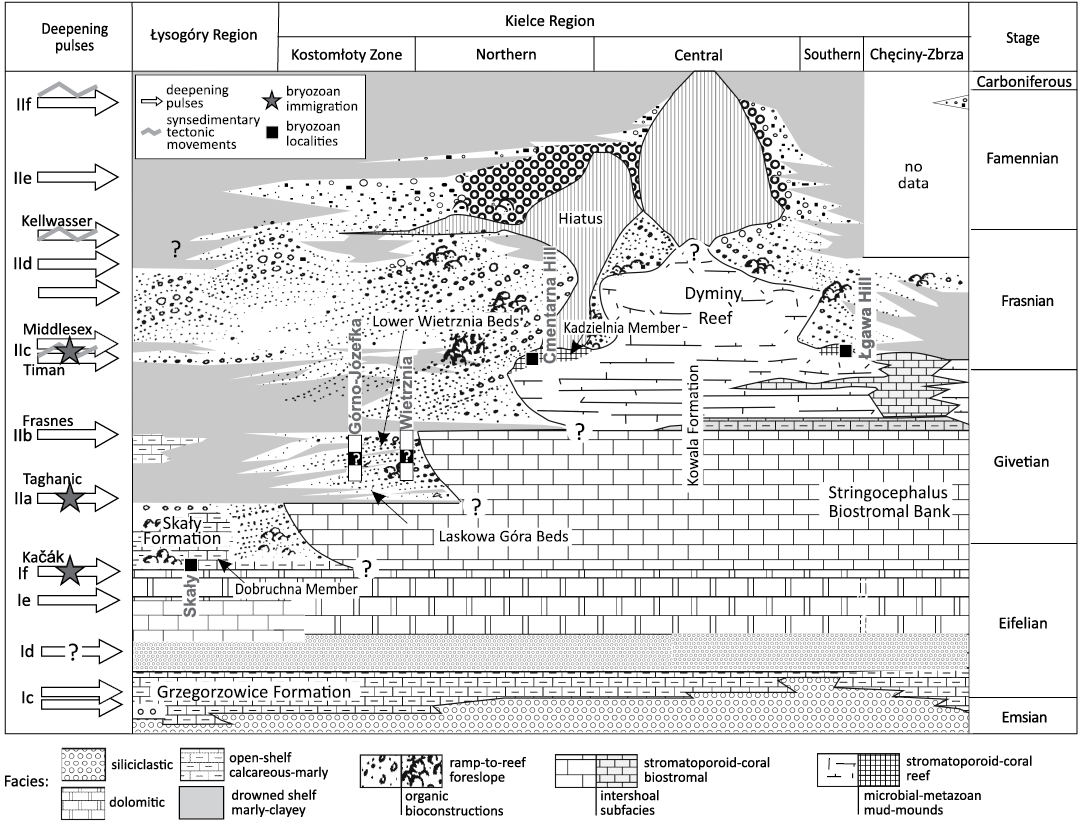
Fig. 2. Position of the bryozoan-bearing sections under this study (see Fig. 1B) relative to the developmental stages of the Middle–Late Devonian bank-to-reef complex of the Holy Cross Mountains; idealized stratigraphic-facies cross section according to Racki (1993: fig. 3, modified) to emphasize eustatic control of the rhythmic depositional pattern and major bryozoan immigration events; transgressive-regressive cycles Ic–IId after Johnson et al. (1985, modified; see Halamski et al. 2022: fig. 7); selected global biotic events following Racki (1993) and Becker (2025).
The Holy Cross Mountains area is divided into two distinct paleogeographic-tectonic domains: the Kielce Paleohigh and the Łysogóry Paleolow, corresponding to a carbonate platform and a hemipelagic basin, respectively (Fig. 1B). The major facies regions are coupled with the transitional Kostomłoty Facies Zone (Racki 1993), which provides an opportunity to compare the bryozoan record in different sedimentary regimes (Fig. 2). However, the subsymmetric facies plan in the Late Devonian is shown by the central location of the Dyminy Reef, surrounded by intrashelf basins: localized Chęciny-Zbrza (southern) and larger Łysogóry-Kostomłoty (northern), as summarized in Racki (1993), Szulczewski (1995) and Racki et al. (2022).
Three of the studied outcrops have already been investigated by Morozova et al. (2002, 2006). The most important data are provided for the uppermost Eifelian (Polygnathus ensensis Conodont Zone) “brachiopod shales” in the Łysogóry Region (Figs. 1B and 2), recently formalized as the Dobruchna Member in the basal part of the shaly-calcareous Skały Formation (Racki et al. 2022). Two studied samples of fossiliferous marls were collected by graduate students of the University of Silesia from trench A dug in 1989 (Czaplikowska 1990; Woźniak 1992; see Woźniak et al. 2022).
Some species come from two upper Givetian localities in the northern part of the Kielce Region: the Górno-Józefka hill (road cut, set A) and the Wietrznia-I quarry (section Wi-IA, basal set A). They represent the bryozoan-enriched calcareous-marly succession of the Laskowa Góra Beds and their lateral equivalents in the basal Wietrznia Beds, roughly dated as an undivided interval of the Schmidtognathus hermanni to Mesotaxis falsiovalis conodont zones (Makowski in Racki et al. 1993; Racki and Bultynck 1993; Baliński et al. 2016).
The relatively frequently encountered Frasnian fenestellids in the northern foreslope successions of the Dyminy Reef, especially in the Kadzielnia Limestone Member of the Kowala Formation, have been recorded since the 19th century (see Racki 2024). They are described here for the first time from two localities of microbial-metazoan mud-mounds, Cmentarna Hill (set B1; Gawlik in Racki et al. 1993) in Kielce and the Łgawa Hill (set J; Racki 1993) near Bolechowice. The fossiliferous mid-slope bioconstructions are uncertainly dated as Palmatolepis transitions to Palmatolepis punctata conodont zones (Racki 2024).
Systematic palaeontology
Phylum Bryozoa Ehrenberg, 1831
Class Stenolaemata Borg, 1926
Superorder Palaeostomata Ma et al., 2014
Order Cystoporata Astrova, 1964
Suborder Fistuliporina Astrova, 1964
Family Fistuliporidae Ulrich, 1882
Genus Cyclotrypa Ulrich, 1896
Type species: Fistulipora communis Ulrich, 1890; Middle Devonian; Iowa, USA.
Emended diagnosis.—Encrusting colonies, often with multiple overgrowths. Cylindrical autozooecia with thin walls and complete diaphragms. Autozooecial apertures rounded. Lunaria absent. Autozooecia separated by extrazooidal vesicular skeleton. Acanthostyles occurring in roofs of vesicles. Low maculae often developed.
Remarks.—Cyclotrypa Ulrich, 1896, differs from Fistulipora M’Coy, 1849, and Eridopora Ulrich, 1882, in the absence of lunaria.
Stratigraphic and geographic range.—Silurian to Permian; Europe, North America, Asia.
Cyclotrypa sp.
Fig. 3A, B; Table 1.
Material.—Single colony, two thin sections (GIUS 4-2239 Sk/Bry-XIV-5N-1, -5N-2) from Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone); Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Description.—Encrusting colony, 1.75 mm in thickness. Autozooecia growing from thin epitheca, bending sharply at their bases towards colony surface. Autozooecial apertures circular to oval. Basal diaphragms absent. Vesicles large, separating autozooecia in 1–2 rows, 6–9 surrounding each autozooecial aperture, polygonal in tangential section, box-like to hemispheric, with plane or concave roofs, consisting of granular skeleton. Autozooecial walls thick, laminated.
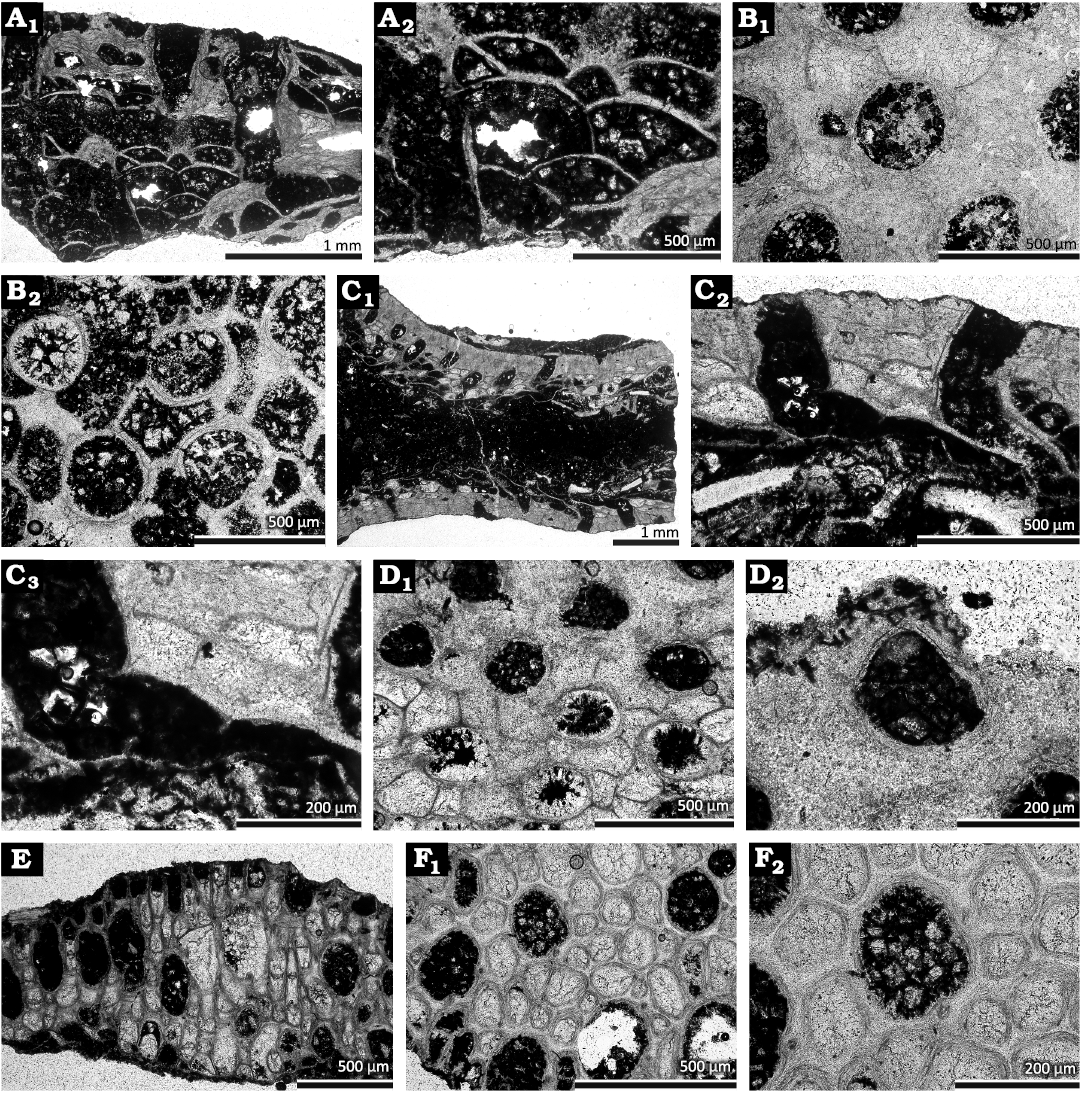
Fig. 3. Cystoporate and trepostome bryozoans from the Devonian of Poland, Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone), Skały. A, B. Cyclotrypa sp. A. Longitudinal section showing autozooecia and vesicles (GIUS 4-2239 Sk/Bry-XIV-5N-1) (A1, A2). B. Tangential section showing autozooecial apertures and vesicles (GIUS 4-2239 Sk/Bry-XIV-5N-2) (B1, B2). C, D. Fistuliphragma gracilis Ernst, 2008. C. Longitudinal section of a tubular colony showing autozooecial chamber, vesicles and hemiphragms (GIUS 4-2239 Sk/Bry-XIV-5A-2) (C1–C3). D. Tangential section showing autozooecial apertures with lunaria and vesicles (GIUS 4-2239 Sk/Bry-XIV-5E-1) (D1, D2). E, F. Leioclema passitabulatum Duncan, 1939. E. Longitudinal section showing autozooecial chambers and mesozooecia (GIUS 4-2239 Sk/Bry-XIV-5L-1). F. Tangential section showing autozooecial apertures, mesozooecia and acanthostyles (GIUS 4-2239 Sk/Bry-XIV-5H-1) (F1, F2).
Table 1. Summary of descriptive statistics for Cyclotrypa sp. (single colony measured). Abbreviations: CV, coefficient of variation; MAX, maximal value; MIN, minimal value; N, number of measurements; SD, sample standard deviation; X, mean.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Aperture width (mm) |
20 |
0.30 |
0.27 |
0.34 |
0.021 |
7.14 |
|
Aperture spacing (mm) |
20 |
0.49 |
0.43 |
0.55 |
0.030 |
6.07 |
|
Vesicle width (mm) |
20 |
0.16 |
0.11 |
0.25 |
0.038 |
23.36 |
|
Vesicles per aperture |
7 |
7.0 |
6.0 |
9.0 |
1.155 |
16.50 |
Remarks.—Cyclotrypa sp. is similar to Cyclotrypa communis (Ulrich, 1890), from the Lower to Middle Devonian (Emsian–Givetian) of USA and Europe, but differs in having larger autozooecial apertures (average aperture width 0.30 mm vs. 0.20 mm in C. communis). Cyclotrypa sp. is similar to C. cyclostoma (Schlütter, 1889) [= C. nekhoroshevi Kiepura, 1973] from the Middle Devonian (Eifelian–Givetian) of Europe and North Africa, but differs in the absence of apertural nodes.
Genus Fistuliphragma Bassler, 1934
Type species: Fistulipora spinulifera Rominger, 1866; Traverse Group, Middle Devonian; Michigan, USA.
Emended diagnosis.—Solid ramose and encrusting colonies, the latter usually in form of hollow tubes. Secondary overgrowth occurring. Autozooecia growing from a thin epitheca, bending sharply at their bases towards colony surface, with circular apertures and large, prominent lunaria. Hemiphragms present, positioned in alternating pattern in autozooecia, originating in laminated skeleton of autozooecia, rare in endozones, rare to abundant in exozones. Basal diaphragms rare to abundant. Vesicular skeleton well-developed. In the centre of each vesicle roof a single acanthostyle is present. Monticules present.
Remarks.—Fistuliphragma Bassler, 1934, is similar to Cliotrypa Ulrich & Bassler in Bassler, 1929, and Strotopora Ulrich in Miller, 1889, in the presence of hemiphragms, but differs from both genera in the absence of gonozooecia.
The species Fistuliphragma crustiformis Yang & Lu, 1983 (originally “crustiforme”, changed herewith to “crustiformis”, according to the Article 11.9 of the Code of Zoological Nomenclature), reported from the Pennsylvanian (Kasimovian–Gzhelian) of China (Xinjiang) belongs to the genus Xiapora Ernst & Gorgij, 2013. This species possesses incomplete vesicles which were interpreted as hemiphragms (Yang and Lu 1983: 271, pl. 5: 8–10).
Stratigraphic and geographic range.—Lower–Middle Devonian (Emsian–Givetian); Europe, China, North Africa, North America.
Fistuliphragma gracilis Ernst, 2008
Fig. 3C, D; Table 2.
2008 Fistuliphragma gracilis sp. nov.; Ernst 2008: 329–332, fig. 11A–G.
2011 Fistuliphragma gracilis Ernst, 2008; Ernst et al. 2011: 305–307, fig. 3e–g.
2012 Fistuliphragma gracilis Ernst, 2008; Ernst et al. 2012: 741–742, fig. 7a–d.
Material.—Five colonies, nine thin sections (GIUS 4-2239 Sk/Bry-XIV-5A-1, -5A-2, -5D, -5E-1, -5E-2, -5G-1, -5G-2, -5O-1, 5O-2) from Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone); Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Description.—Encrusting or hollow ramose colonies formed by encrusting overgrowths of ephemeral cylindrical objects. Encrusting sheets 0.6–1.0 mm thick. Autozooecia moderately long, narrow in endozones. Diaphragms absent. Hemiphragms abundant, long, proximally curved; alternating in outer endozone and in exozone. Vesicles in outer endozone and exozone, 7–9 surrounding each autozooecial aperture, polygonal in tangential section, box-like to hemispheric, with plane or concave roofs, consisting of granular skeleton. Vesicular roofs containing acanthostyles, 0.018–0.025 mm in diameter. Autozooecial walls granular, 0.005–0.010 mm thick in endozones; laminated, 0.010–0.015 mm thick in exozones. Lunaria in outer exozone and endozone, horse-shoe shaped, large, with thick skeletal deposits. Maculae not observed.
Table 2. Summary of descriptive statistics for Fistuliphragma gracilis Ernst, 2008 (five colonies measured). Abbreviations: CV, coefficient of variation; MAX, maximal value; MIN, minimal value; N, number of measurements; SD, sample standard deviation; X, mean.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Aperture width (mm) |
25 |
0.17 |
0.13 |
0.21 |
0.023 |
13.77 |
|
Aperture spacing (mm) |
25 |
0.39 |
0.32 |
0.50 |
0.042 |
10.92 |
|
Vesicle width (mm) |
25 |
0.09 |
0.06 |
0.14 |
0.023 |
24.00 |
Remarks.—Fistuliphragma gracilis Ernst, 2008, differs from Fistuliphragma eifelensis Ernst, 2008, in having smaller colonies, and smaller autozooecia (average autozooecial aperture width 0.17 mm vs. 0.28 mm in F. eifelensis).
Stratigraphic and geographic range.—Middle Devonian (lower Eifelian), Kierspe, Germany. Middle Devonian (upper Eifelian), western Rhenish Massif, Germany. Santa Lucia Formation, Lower–Middle Devonian (upper Emsian–lower Eifelian), Abelgas, Paradilla, Cantabrian Mountains, NW Spain. Dobruchna Member in the bottom part of the Skały Formation, Middle Devonian (uppermost Eifelian, Polygnathus ensensis Conodont Zone), Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Order Trepostomata Ulrich, 1882
Suborder Halloporina Astrova, 1965
Family Heterotrypidae Ulrich, 1890
Genus Leioclema Ulrich,
1882
[= Lioclema Ulrich, 1882]
Type species: Callopora punctata Hall, 1858; Mississippian; Iowa, USA.
Emended diagnosis.—Encrusting, branched, less commonly massive colonies. Autozooecia with polygonal to rounded-polygonal, sometimes petaloid apertures. Autozooecial diaphragms rare. Mesozooecia abundant, with abundant diaphragms, often beaded. Acanthostyles abundant, commonly large. Autozooecial walls thin in endozone; laminated, regularly thickened in exozones (modified after Astrova 1978).
Remarks.—Leioclema Ulrich, 1882, differs from Heterotrypa Nicholson, 1879, in having rare autozooecial diaphragms and abundant acanthostyles and mesozooecia, and from Stigmatella Ulrich & Bassler, 1904, in having abundant mesozooecia.
Stratigraphic and geographic range.—Lower Silurian to Pennsylvanian (Carboniferous); worldwide.
Leioclema passitabulatum Duncan, 1939
Fig. 3E–F; Table 3.
1939 Lioclema passitabulatum sp. nov.; Duncan 1939: 251, pl. 16: 8–10.
1964 Lioclema passitabulatum Duncan, 1939; Astrova 1964: 33, pl. 10: 2a, b.
2007 Leioclema passitabulatum Duncan, 1939; Ernst and Schroeder 2007: 210, figs. 2I–K, 3A–C.
2009 Leioclema passitabulatum Duncan, 1939; Ernst and May 2009: 779, figs. 7.1–7.4.
2012 Leioclema passitabulatum Duncan, 1939; Ernst et al. 2012: 743, fig. 8a–d.
Material.—Three colonies, three thin sections (GIUS 4-2239 Sk/Bry-XIV-5H-1, -5J-1, -5L-1) from Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone); Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Description.—Encrusting colonies, 0.90–1.08 mm in thickness. Autozooecia budding from a thin epitheca, briefly oriented parallel to the substrate, then bending sharply and intersecting the colony surface at right angles. Epitheca 0.013–0.018 mm thick. Autozooecial apertures rounded-polygonal to petaloid due to indenting acanthostyles. Autozooecial diaphragms few to absent, thin, straight or slightly deflected proximally. Mesozooecia abundant, 6–11 surrounding each aperture, polygonal in cross section, slightly beaded, containing planar diaphragms. Acanthostyles small, abundant, 1–5 surrounding each aperture, originating from the base of exozone, often indenting autozooecia, having distinct calcite cores and dark laminated sheaths. Autozooecial walls granular, in endozone 0.010–0.015 mm thick; in exozone 0.025–0.030 mm thick, distinctly laminated. Maculae not observed.
Table 3. Summary of descriptive statistics for Leioclema passitabulatum Duncan, 1939 (three colonies measured). Abbreviations: CV, coefficient of variation; MAX, maximal value; MIN, minimal value; N, number of measurements; SD, sample standard deviation; X, mean.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Aperture width (mm) |
35 |
0.18 |
0.13 |
0.23 |
0.024 |
13.31 |
|
Aperture spacing (mm) |
34 |
0.34 |
0.25 |
0.43 |
0.050 |
14.66 |
|
Mesozooecium width (mm) |
34 |
0.09 |
0.04 |
0.14 |
0.022 |
25.62 |
|
Acanthostyle diameter (mm) |
17 |
0.031 |
0.025 |
0.040 |
0.005 |
14.68 |
|
Mesozooecia per aperture |
16 |
8.6 |
6.0 |
11.0 |
1.147 |
13.30 |
Remarks.—Leioclema passitabulatum Duncan, 1939, is similar to Leioclema alpenense Duncan, 1939, but differs from it in containing more abundant acanthostyles (1–5 vs. 2 acanthostyles around each aperture, respectively).
Stratigraphic and geographic range.—Lower Devonian, Lochkovian, Bortschovski Horizon, Podolia, Ukraine. Lower Devonian, Pragian, Konӗprusy Limestone, Czech Republic. Middle Devonian, Eifelian, Traverse Group, Michigan, USA. Middle Devonian, lower Eifelian, Kierspe, Germany. Dobruchna Member in the bottom part of the Skały Formation, Middle Devonian (uppermost Eifelian (Polygnathus ensensis Conodont Zone), Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland. Cürten Formation, Middle Devonian, lower Givetian, Dollendorf Syncline, Rhenish Slate Massif, Germany.
Order Cryptostomata Vine, 1884
Suborder Rhabdomesina Astrova & Morozova, 1956
Family Lenaporidae Ernst & Königshof, 2010
Genus Bigeyella Morozova & Weiss in Morozova et al., 2006
Type species: Bigeyella sparsa Morozova & Weiss in Morozova et al., 2006; Middle Wietrznia Series (Member C), Upper Devonian (Frasnian); central Poland.
Emended diagnosis.—Branched colonies. Autozooecia tubular, with inflated bases, initially polygonal, irregular in transverse section becoming rhombic, bending abruptly in exozones, growing in spiral arrangement from the median axis. Autozooecial diaphragms occurring; hemisepta absent. Autozooecial apertures circular to oval, arranged in regular alternating longitudinal rows. Metazooecia occasionally present. Acanthostyles surrounding autozooecial apertures present, varying in number and size. Autozooecial walls laminated, thick, with dividing hyaline layer in endozone; laminated, merged, containing abundant tubules in exozones (emended after Morozova & Weiss in Morozova et al. 2006).
Remarks.—Bigeyella differs from Lenapora Ernst & Königshof, 2010, in having acanthostyles and metazooecia.
Stratigraphic and geographic range.—Lower–Upper Devonian (Emsian–Frasnian), Europe, Kazakhstan, Canada.
Bigeyella indigena (Morozova & Weiss in Morozova et al., 2002)
Fig. 4A–C; Table 4.
2002 Primorella indigena sp. nov.; Morozova & Weiss in Morozova et al. 2002: 314–315, fig. 6H–J.
2024 Bigeyella indigena (Morozova & Weiss in Morozova et al., 2002); Ernst et al. 2024: 15, fig. 3e, f.
Material.—Ten colonies, eighteen thin sections (GIUS 4-292 Gó/Bry A-1a, A-1b, A-1c, A-2a, A-2b, A-2c, A-3a, A-3b, A-4a, A-4b, A-5a, A-6a, A-6b, A-7a, A-8a, A-8b, A-9a, A-10a) from Laskowa Góra Beds, Middle Devonian (upper Givetian) (?Polygnathus ansatus–Schmidtognathus hermanni, and Klapperina disparilis conodont zones); Górno, Józefka Hill (road cut, set A), Poland.
Description.—Branched colonies, 0.92–1.90 mm in diameter. Endozone 0.38–0.70 mm wide, exozone 0.24–0.60 mm wid. Axial ratio 0.32–0.50. Autozooecia tubular, budding from distinct branch axis, growing in spiral arrangement from the median axis and bending abruptly in exozones. Diaphragms not observed. Acanthostyles small, with distinct hyaline cores and laminated sheaths. Autozooecial apertures oval, arranged in regular alternating longitudinal rows. Autozooecial walls laminated, 0.02–0.04 mm thick in endozone; laminated, containing tubules in exozone.
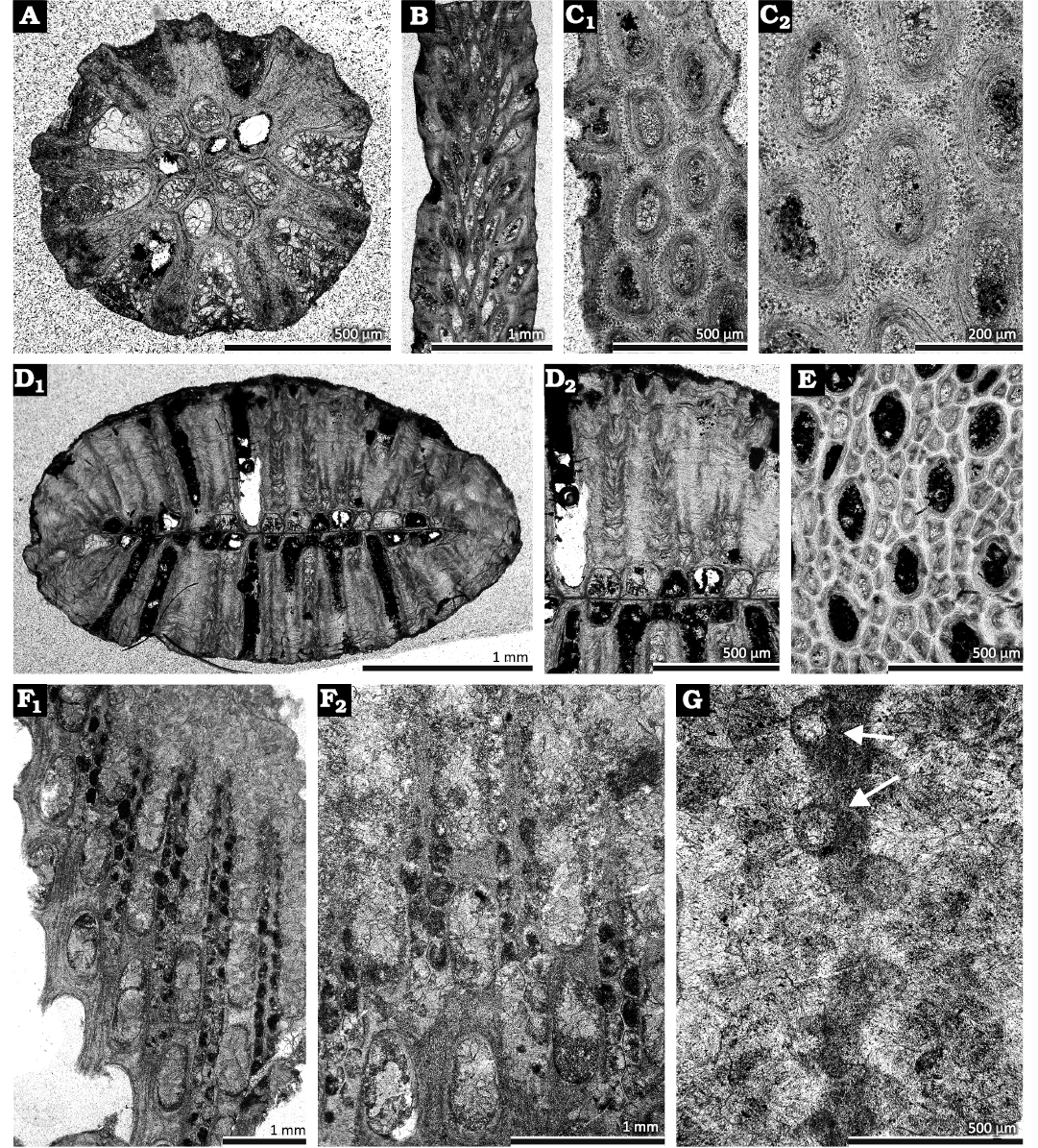
Fig. 4. Cryptostome and fenestrate bryozoans from the Devonian of Poland. A–C. Bigeyella indigena (Morozova & Weiss in Morozova et al., 2002), from the Laskowa Góra Beds, Middle Devonian (upper Givetian) (?Polygnathus ansatus–Schmidtognathus hermanni, and Klapperina disparilis conodont zones) of Górno, Józefka Hill. A. Transverse section (GIUS 4-292 Gó/Bry A-8a). B. Longitudinal section (GIUS 4-292 Gó/Bry A-4b). C. Tangential section (GIUS 4-292 Gó/Bry A-3a) (C1, C2). D, E. Intrapora leunisseni Ernst, 2008, from the Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone) of Skały. D. Transverse section showing autozooecia, metazooecia, and mesotheca{?} (GIUS 4-2239 Sk/Bry-XIV-5C-2) (D1, D2). E. Tangential section showing autozooecia, metazooecia, and acanthostyles (GIUS 4-2239 Sk/Bry-XIV-5I-1). F, G. Rectifenestella sp. from the Kadzielnia Limestone Member of the Kowala Formation, Lower Frasnian (Palmatolepis transitans Conodont Zone). F. Tangential section showing autozooecial chambers (GIUS 4-259 GC/Bry-1a), Bolechowice, Łgawa Hill (F1, F2). G. Apertures with stellate structures (arrows) (GIUS 4-256 GŁ/Bry-1b), Kielce, Cmentarna Hill.
Table 4. Summary of descriptive statistics for Bigeyella indigena (Morozova & Weiss in Morozova et al., 2002) (ten colonies measured). Abbreviations: CV, coefficient of variation; MAX, maximal value; MIN, minimal value; N, number of measurements; SD, sample standard deviation; X, mean.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Branch width (mm) |
7 |
1.19 |
0.92 |
1.90 |
0.355 |
29.92 |
|
Exozone width (mm) |
7 |
0.35 |
0.24 |
0.60 |
0.135 |
38.21 |
|
Endozone width (mm) |
7 |
0.48 |
0.38 |
0.70 |
0.108 |
22.53 |
|
Axial ratio |
7 |
0.41 |
0.32 |
0.50 |
0.069 |
16.82 |
|
Aperture width (mm) |
40 |
0.10 |
0.07 |
0.13 |
0.019 |
19.13 |
|
Aperture spacing along branch (mm) |
20 |
0.40 |
0.30 |
0.48 |
0.057 |
14.15 |
|
Aperture spacing diagonally (mm) |
20 |
0.25 |
0.20 |
0.30 |
0.025 |
9.85 |
|
Acanthostyle diameter (mm) |
20 |
0.029 |
0.020 |
0.040 |
0.005 |
17.84 |
Remarks.—Bigeyella indigena (Morozova & Weiss in Morozova et al., 2002) differs from Bigeyella sparsa Morozova & Weiss in Morozova et al., 2006, in having larger autozooecial apertures (aperture width 0.07–0.13 mm vs. 0.07–0.08 mm in B. sparsa).
Stratigraphic and geographic range.—Middle Devonian, upper Givetian of Poland (?Polygnathus ansatus–Schmidtognathus hermanni, Klapperina disparilis conodont zones), Górno, Józefka Hill (road cut, set A); Upper Devonian, middle Frasnian of Poland: Laskowa Góra Beds. Upper Devonian, ?lower–middle Frasnian, Noravank section, Central Armenia.
Suborder Ptilodictyina Astrova & Morozova, 1956
Family Intraporidae Simpson, 1897
Genus Intrapora Hall, 1883
Type species: Intrapora puteolata Hall, 1883; Middle Jeffersonville Limestone, Middle Devonian; Eastern USA.
Emended diagnosis.—Bifoliate colonies consisting of dichotomous branches, leaf-like, frondose. Mesotheca straight or slightly undulating. Autozooecia short, abruptly bending in exozones, Superior hemisepta present or absent. Diaphragms occasionally occurring. Metazooecia abundant, often separating autozooecia. Acanthostyles present or absent.
Remarks.—Intrapora Hall, 1883, differs from Ensiphragma Astrova in Astrova & Yaroshinskaya, 1968, in arrangement of metazooecia. Metazooecia in Intrapora are arranged more or less irregularly, whereas the metazooecia of Ensiphragma are arranged in pairs between apertures.
Stratigraphic and geographic range.—Lower Devonian–Mississippian (Emsian–Tournaisian), Europe, North America, and Asia.
Intrapora leunisseni Ernst, 2008
Fig. 4D, E; Table 5.
2008 Intrapora leunisseni sp. nov.; Ernst 2008: 357, figs. 25H, 26A–F.
Material.—Four colonies, six thin sections (GIUS 4-2239 Sk/Bry-XIV-5C-1, -5C-2, -5C-3, -5F-1, -5I-1, -5M-1) from Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone); Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Description.—Bifoliate, branched colonies. Branch cross section lens-shaped. Branches 2.68–2.95 mm wide and 1.63–1.70 mm thick. Mesotheca 0.010–0.015 mm thick, three-layered, with central dark layer and two outer bright layers. Autozooecia long, growing from a mesotheca, semicircular to trapezoid at the base in cross section, becoming oval to petaloid in the exozone, arranged in 9–15 rows on branches. Autozooecial diaphragms rare, hemisepta absent. Metazooecia small, abundant, separating autozooecia in 1–3 rows, 8–11 surrounding each autozooecial aperture, polygonal to narrowly oval in cross section, often sealed by calcitic skeleton at colony surface. Metazooecial diaphragms abundant, thick. Acanthostyles abundant, 2–5 constantly surrounding each autozooecial aperture and occurring randomly between autozooecia, having narrow hyaline cores and wide laminated sheaths.
Table 5. Summary of descriptive statistics for Intrapora leunisseni Ernst, 2008 (four colonies measured). Abbreviations: CV, coefficient of variation; MAX, maximal value; MIN, minimal value; N, number of measurements; SD, sample standard deviation; X, mean.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Aperture width (mm) |
30 |
0.11 |
0.09 |
0.15 |
0.015 |
13.58 |
|
Aperture spacing along branch (mm) |
13 |
0.49 |
0.42 |
0.57 |
0.048 |
9.86 |
|
Aperture spacing diagonally (mm) |
13 |
0.31 |
0.27 |
0.35 |
0.024 |
7.53 |
|
Metazooecia width (mm) |
30 |
0.05 |
0.03 |
0.10 |
0.016 |
29.51 |
|
Acanthostyle diameter (mm) |
30 |
0.032 |
0.020 |
0.043 |
0.006 |
18.47 |
Remarks.—Intrapora leunisseni Ernst, 2008, is similar to Intrapora texera Troizkaya, 1975, from the Mississippian (Tournaisian) of central Kazakhstan. However, I. texera has abundant autozooecial diaphragms and wider branches (branch width about 5 mm vs. 2.68–2.95 mm in I. leunisseni.
Stratigraphic and geographic range.—Ahbach-Formation, Middle Devonian, lowermost lower Givetian, Üxheim-Ahütte, Müllertchen quarry, Hillersheim syncline, Eifel, western Rhenish Massif, Germany. Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone), Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Order Fenestellida Elias & Condra, 1957
Family Fenestellidae King, 1849
Genus Rectifenestella Morozova, 1974
Type species: Fenestella medvedkensis Schulga-Nesterenko, 1951; Pennsylvanian (Kasimovian); Russian Platform.
Emended diagnosis.—Reticulate colonies consisting of fine to intermediately robust branches and straight dissepiments. Autozooecia triangular to pentagonal in mid tangential section. Superior hemisepta present; inferior hemisepta absent. Low keel carrying one row of intermediate nodes (modified after Morozova 2001: 45).
Remarks.—Rectifenestella Morozova, 1974, differs from Laxifenestella Morozova, 1974, in having a pentagonal shape of autozooecia and in the absence of inferior hemisepta, from Minilya Crockford, 1944, in having a single row of nodes on the keel instead of two alternating rows in Minilya.
Stratigraphic and geographic range.—Devonian–Permian; worldwide.
Rectifenestella sp.
Fig. 4F, G; Table 6.
Material.—Two colonies, four thin sections (GIUS 4-256 GŁ/Bry-1a, -1b, GIUS 4-259 GC/Bry-1a, -1b) from Kadzielnia Limestone Member of the Kowala Formation; Lower Frasnian; Palmatolepis transitans Conodont Zone at Bolechowice, Łgawa Hill (set J) and ?Palmatolepis transitans Conodont Zone at Kielce, Cmentarna Hill (set B1), Poland.
Description.—Exterior: Reticulate colonies with straight branches, bifurcating, joined by moderately wide dissepiments. Autozooecia arranged in two alternating rows on branches. Autozooecial apertures circular, surrounded by 8 apertural nodes (stellate structure); 3–5 apertures spaced per fenestrule length. Apertural nodes 0.012–0.020 mm in diameter. Fenestrules oval to rectangular, moderately long and narrow. Median keel low. Keel nodes small, intermediately spaced, granular core stellate in shape. Reverse side longitudinally striated.
Table 6. Summary of descriptive statistics for Rectifenestella sp. (two colonies measured). Abbreviations: N, number of measurements; X, mean; MIN, minimal value; MAX, maximal value; SD, sample standard deviation; CV, coefficient of variation.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Branch width (mm) |
32 |
0.28 |
0.22 |
0.38 |
0.037 |
13.28 |
|
Dissepiment width (mm) |
30 |
0.23 |
0.15 |
0.36 |
0.058 |
24.99 |
|
Fenestrule width (mm) |
33 |
0.31 |
0.22 |
0.42 |
0.044 |
14.15 |
|
Fenestrule length (mm) |
36 |
0.74 |
0.45 |
1.00 |
0.104 |
14.02 |
|
Distance between branch centres (mm) |
36 |
0.60 |
0.47 |
0.75 |
0.064 |
10.62 |
|
Distance between dissepiment centres (mm) |
39 |
0.98 |
0.73 |
1.20 |
0.099 |
10.10 |
|
Aperture width (mm) |
32 |
0.08 |
0.07 |
0.09 |
0.008 |
10.34 |
|
Aperture spacing along branch (mm) |
32 |
0.26 |
0.22 |
0.32 |
0.029 |
10.95 |
|
Aperture spacing diagonally (mm) |
32 |
0.23 |
0.18 |
0.27 |
0.022 |
9.57 |
|
Apertures per fenestrule length |
40 |
3.6 |
3.0 |
5.0 |
0.552 |
15.56 |
|
Maximal chamber width (mm) |
40 |
0.13 |
0.10 |
0.15 |
0.010 |
8.10 |
Interior: Autozooecia triangular to pentagonal in mid-tangential section; with well-developed long vestibule; axial wall zigzag; aperture positioned at distal end of chamber. Hemisepta absent. Internal granular skeleton continuous with obverse keel, nodes, peristome and across dissepiments. External laminated skeleton well developed, traversed by abundant microstyles. Microstyles regularly arranged in longitudinal rows on colony reverse surface, 0.02–0.03 mm in diameter. Heterozooecia not observed.
Remarks.—Rectifenestella sp. is similar to Rectifenestella localis Morozova & Weiss in Morozova et al., 2006, from the Middle Devonian (Eifelian) of Poland, but differs from it in wider branches (branch width 0.22–0.38 mm vs. 0.20–0.25 mm in R. localis).
Genus Laxifenestella Morozova, 1974
Type species: Fenestella sarytshevae Schulga-Nesterenko, 1951, subsequently designated by Morozova (1974); Mississippian (Serpukhovian), Moscow Syncline, Russia.
Emended diagnosis.—Reticulate colonies consisting of relatively wide and thick branches and moderately wide dissepiments. Autozooecia arranged in two rows on the branches. Autozooecial chambers rectangular to pentagonal in mid-tangential section. Axial wall between autozooecial rows weakly undulating. Both superior and inferior hemisepta present. Narrow keel with single row of nodes developed (modified after Morozova 2001: 44).
Remarks.—Laxifenestella Morozova, 1974, differs from Fenestella Lonsdale, 1839, in the rectangular to pentagonal shape of autozooecia in mid-tangential section and the presence of well-developed hemisepta.
Stratigraphic and geographic range.—Lower Devonian–upper Permian; worldwide.
Laxifenestella sp.
Fig. 5A.
Material.—Single specimen (GIUS 4-2239 Sk/Bry-XIV- 5K-1) from Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone); Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Description.—Exterior: Reticulate colonies formed by straight branches joined by relatively wide dissepiments. Branches 0.34–0.37 mm in width, dissepiments 0.28–0.34 mm in width. Fenestrules oval to rectangular, 0.21–0.36 mm in width and 0.47–0.58 mm in length. Autozooecia arranged in two rows on branches. Autozooecial apertures circular, with low peristome; 2 to 3 apertures spaced per fenestrule length. Keel high, nodes inconspicuous. Reverse side not observed.
Interior: Autozooecia relatively long, rectangular in mid-tangential section; with short to moderately long vestibule in longitudinal section. Axial wall between autozooecial rows straight; aperture positioned at distal end of chamber. Superior hemisepta long, proximally curved; inferior hemisepta short or indistinct. External laminated skeleton well-developed on both obverse and reverse side. Heterozooecia not observed.
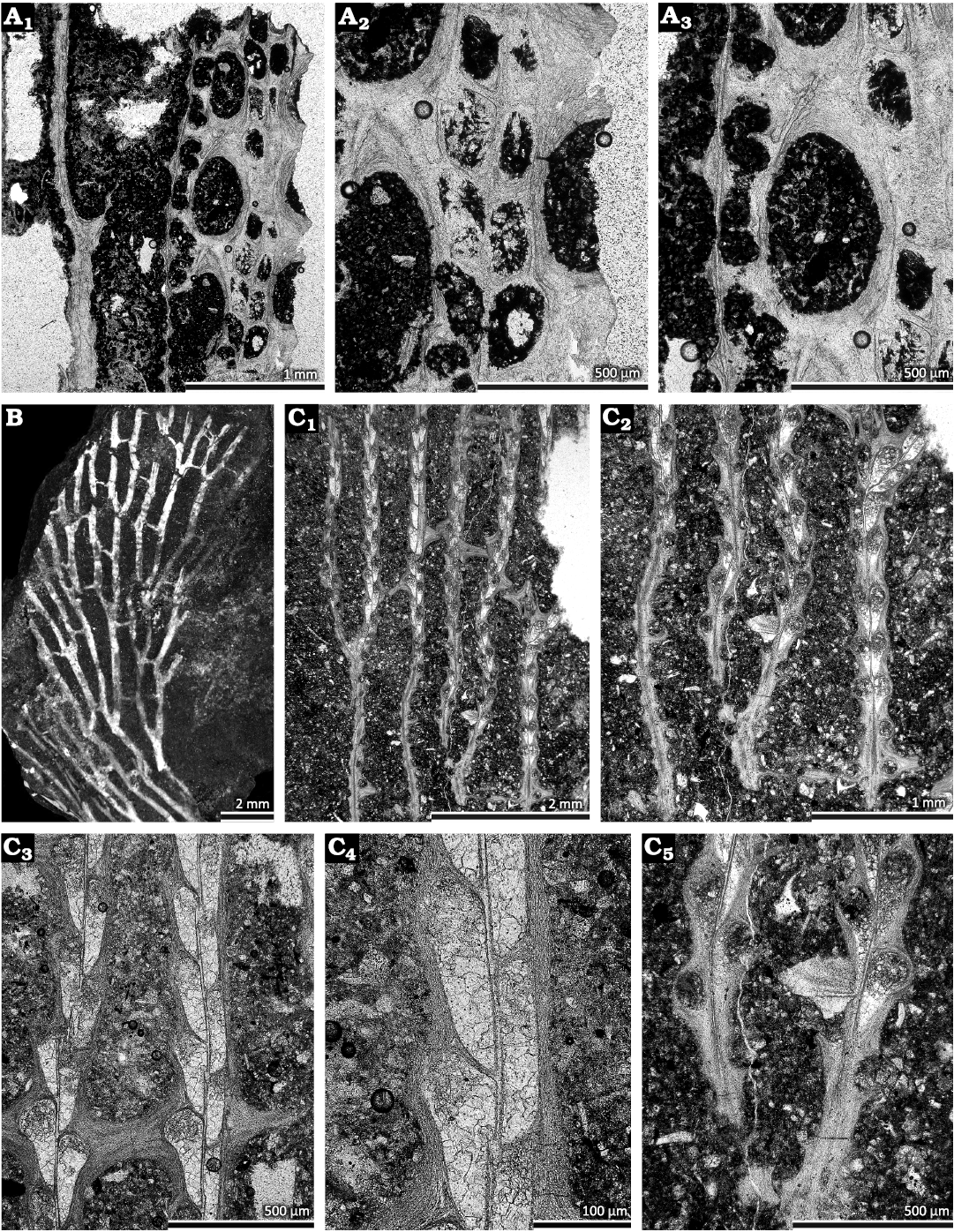
Fig. 5. Fenestrate bryozoans from the Devonian of Poland. A. Laxifenestella sp., tangential section (GIUS 4-2239 Sk/Bry-XIV-5K-1), from the Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone) of Skały (A1–A3). B, C. Juanopora elegans gen. et sp. nov. from the Wietrznia Beds, upper Givetian (?Schmidtognathus hermanni – Mesotaxis falsiovalis conodont zones) of Wietrznia-I quarry at Kielce. B. Colony fragment, paratype (GIUS 4-260b Wt/Bry-2). C. Tangential section showing autozooecial chambers and apertures, holotype (GIUS 4-260b Wt/Br-1a) (C1–C5).
Remarks.—Laxifenestella sp. is superficially similar to Laxifenestella vera (Ulrich, 1890) from the Middle Devonian (Givetian) of USA. However, the latter species possesses closely spaced nodes on the keel. This character could not be observed on the present specimen. Material identified as Laxifenestella vera has been also described from the Middle Devonian of Mongolia and Altai (Nekhoroshev 1926, 1948; Krasnopeeva 1935).
Genus Juanopora nov.
ZooBank LSID: urn:lsid:zoobank.org:act:BB472E06-D35D-447C-9C 9E-9189F08745F7
Type species: Juanopora elegans gen. et sp. nov., by monotypy; see below.
Etymology: Named in honour of Juan Luis Suárez Andrés (SONINGEO, Maliaño, Spain) for his contribution to the knowledge of Palaeozoic bryozoans.
Remarks.—The new genus is mainly characterized by the shape of autozooecia which are long and narrow with rectangular outlines in the middle tangential section. These are unlike those developed in previously documented fenestellid genera. Similar autozooecia are known in the acanthocladiid genus Diploporaria Nickles & Bassler, 1900 (cf. Ernst et al. 2015: 192, fig. 26F–H; Diploporaria tenella Wyse Jackson, 1988). However, this genus possesses hemisepta and a pinnate growth form.
Stratigraphic and geographic range.—As for the type species.
Juanopora elegans gen. et sp. nov.
Fig. 5B, C; Table 7.
ZooBank LSID: urn:lsid:zoobank.org:act:B37F7FB6-E7F1-4EDA-AFCC-5FEB581AE822.
Etymology: From Latin elegans, fine; in reference to its elegant appearance and thin morphological elements.
Type material: Holotype, colony fragment (thin section GIUS 4-260b Wt/Br-1a). Paratype, colony fragment (thin section GIUS 4-260b Wt/Bry-2) from the type locality and horizon.
Type locality: Wietrznia-I quarry at Kielce, section Wi-IA, Poland.
Type horizon: Wietrznia Beds (basal set A), upper Givetian (?Schmidtognathus hermanni–Mesotaxis falsiovalis conodont zones)
Diagnosis.—Reticulate colonies consisting of relatively narrow branches and narrow dissepiments. Fenestrules long and narrow. Autozooecia arranged in two rows on the branches, weakly alternating. Autozooecial chambers rectangular in mid-tangential section, long and narrow. Axial wall between autozooecial rows straight. Hemisepta absent. Keel low; nodes absent.
Description.—Exterior: Reticulate colonies formed by straight narrow branches joined by narrow dissepiments. Fenestrules long and narrow, rectangular. Autozooecia arranged in two rows on branches, weakly alternating, often arranged side-by-side (Fig. 5C3, C4). Autozooecial apertures circular, with low peristome; 4–6 apertures spaced per fenestrule length. Keel low, nodes absent. Reverse side not observed.
Table 7. Summary of descriptive statistics for Juanopora elegans gen. et sp. nov. (single colony measured). Abbreviations: CV, coefficient of variation; MAX, maximal value; MIN, minimal value; N, number of measurements; SD, sample standard deviation; X, mean.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Branch width (mm) |
5 |
0.22 |
0.19 |
0.25 |
0.022 |
9.94 |
|
Dissepiment width (mm) |
6 |
0.12 |
0.09 |
0.14 |
0.020 |
16.67 |
|
Fenestrule width (mm) |
5 |
0.30 |
0.24 |
0.35 |
0.040 |
13.33 |
|
Fenestrule length (mm) |
3 |
1.66 |
1.45 |
1.88 |
0.215 |
12.96 |
|
Distance between branch centres (mm) |
5 |
0.49 |
0.40 |
0.60 |
0.078 |
15.78 |
|
Distance between dissepiment centres (mm) |
3 |
1.70 |
1.60 |
1.85 |
0.132 |
7.78 |
|
Aperture width (mm) |
5 |
0.10 |
0.09 |
0.11 |
0.009 |
9.32 |
|
Aperture spacing along branch (mm) |
5 |
0.28 |
0.23 |
0.32 |
0.033 |
11.60 |
Interior: Autozooecia relatively long, rectangular in mid-tangential section; with short to moderately long vestibule in longitudinal section. Axial wall between autozooecial rows straight; aperture positioned at distal end of chamber. Hemisepta absent. External laminated skeleton well-developed on both obverse and reverse side. Heterozooecia not observed.
Stratigraphic and geographic range.—Middle Devonian (upper Givetian), Holy Cross Mountains, Poland.
Genus Hemitrypella
Nekhoroshev, 1948
[= Neohemitrypa Schastlivceva in Morozova,
2001]
Type species: Hemitrypella tubulosa Nekhoroshev, 1948, by original designation; Middle Devonian; Altai, Russia.
Diagnosis.—Reticulate colonies, conical and conical-tubular shaped, frontal surface exterior. Branches intermediate in width, linear to moderately sinuous, closely or intermediately spaced, dichotomously divided. Two rows of autozooecia per branch, increasing to three rows proximal of branch bifurcations; low straight to sinuous central keel on obverse side of branch with high nodes, composed of core of granular skeleton and sheath of laminar skeleton. Laminar wall extensions of keel nodes fused together forming a fine meshwork of rounded to angular openings, each opening centred over a zooecial aperture in the branch below. Axial wall between autozooecial rows zigzag in tangential sections; autozooecia not strongly inflated laterally, commonly triangular to trapezoid in tangential section; maximum diameter of zooecia corresponds with either length or height; transverse walls at intermediate or high angle to reverse wall; superior hemisepta absent or weakly developed, other interior structures absent. Small- to large-diameter distal tube typically short, opening frontally or slightly inclined laterally and perhaps distally; apertural peristome present or absent. Heterozooecia not observed. Autozooecial walls of granular material that may be absent on obverse side near apertures; laminar extrazooecial skeleton traversed by small to moderate microstyles (after Ernst et al. 2025).
Remarks.—Hemitrypella Nekhoroshev, 1948, differs from Hemitrypa Phillips, 1841, in its conical and conical-tubular colony shape and in the triangular to trapezoid shape of autozooecia in tangential section, which are quadrangular or pentagonal in Hemitrypa.
Stratigraphic and geographic range.—Lower Devonian (Lochkovian)–Mississippian (Tournaisian); Europe, Russia, Kazakhstan, Mongolia, and China.
Hemitrypella nodulosa Ernst et al., 2025
Fig. 6A–C; Table 8.
2025 Hemitrypella nodulosa Ernst et al. 2025: 501–503, fig. 6a–h.
Material.—Three colonies, three thin sections (GIUS 4-2239 Sk/Bry-XIV-5P, -5Q, -5B-1) from the Dobruchna Member in the bottom part of the Skały Formation, Middle Devonian, uppermost Eifelian, Polygnathus ensensis Conodont Zone), Skały, Poland.
Description.—Exterior: Reticulate colonies with straight branches joined by dissepiments. Autozooecia arranged in two alternating rows on branches, having circular apertures with low peristomes, 1–2 spaced per length of a fenestrule. Peristomes smooth. Fenestrules oval to slightly rectangular. Openings in the superstructure irregularly shaped, rounded to petaloid, corresponding to positions of apertures, 0.09–0.13 mm in diameter. Reverse colony surface not observed in the present material.
Interior: Autozooecia triangular to trapezoidal in mid-tangential section; low and elongated, with short vestibule in longitudinal section. Axial wall between autozooecial rows zigzag in tangential sections; aperture positioned at distal end of chamber. Hemisepta absent. Internal granular skeleton continuous with obverse keel, nodes, peristome and across dissepiments. External laminated skeleton well developed. Heterozooecia not observed.
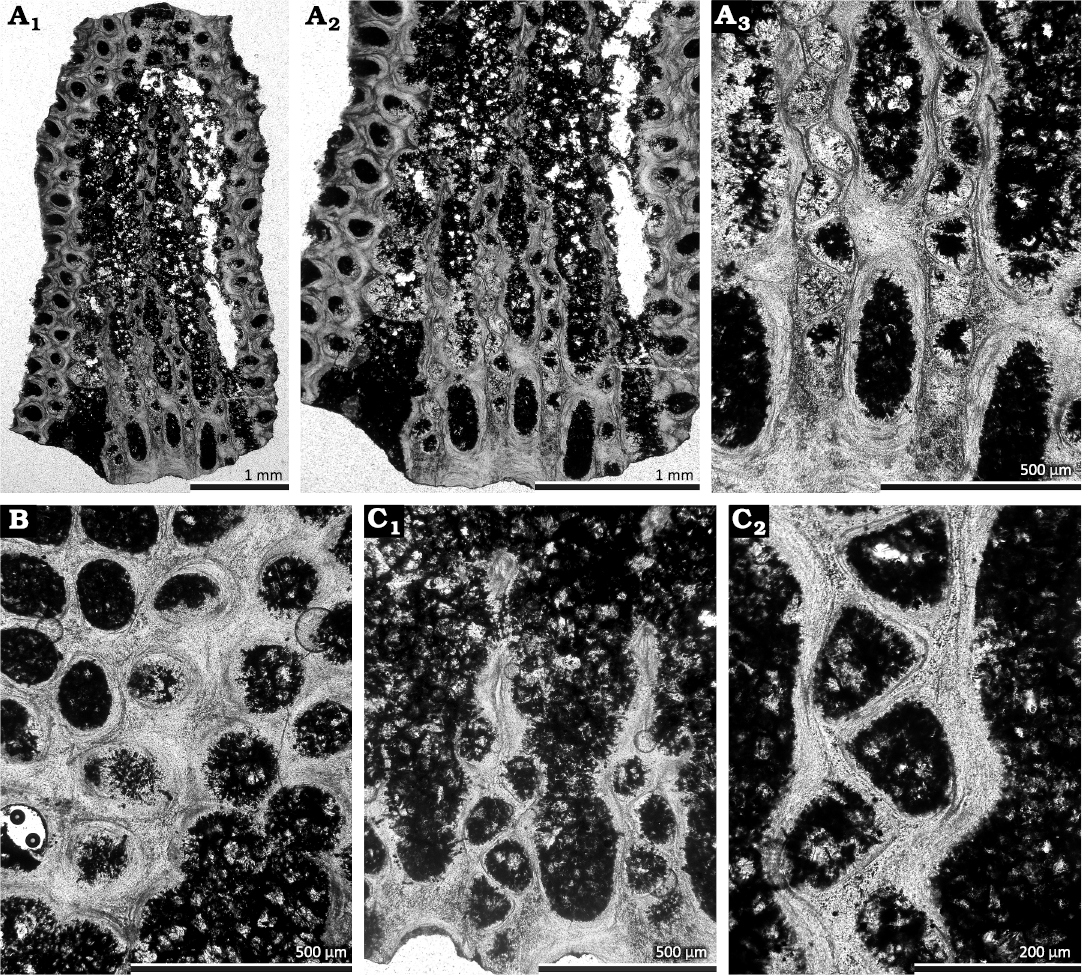
Fig. 6. Fenestrate bryozoan Hemitrypella nodulosa Ernst et al., 2025, from the Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone) of Skały, Poland. A. Tangential section showing autozooecial chambers and protective superstructure (GIUS 4-2239 Sk/Bry-XIV-5B-1) (A1–A3). B. Tangential section showing openings of the superstructure (GIUS 4-2239 Sk/Bry-XIV-5P). C. Tangential section showing autozooecial apertures and chambers (GIUS 4-2239 Sk/Bry-XIV-5Q) (C1, C2).
Table 8. Summary of descriptive statistics for Hemitrypella nodulosa Ernst et al., 2025 (three colonies measured). Abbreviations: CV, coefficient of variation; MAX, maximal value; MIN, minimal value; N, number of measurements; SD, sample standard deviation; X, mean.
| |
N |
X |
MIN |
MAX |
SD |
CV |
|
Branch width (mm) |
7 |
0.23 |
0.19 |
0.30 |
0.042 |
18.45 |
|
Dissepiment width (mm) |
8 |
0.11 |
0.08 |
0.16 |
0.025 |
22.01 |
|
Fenestrule width (mm) |
7 |
0.17 |
0.14 |
0.20 |
0.022 |
13.15 |
|
Fenestrule length (mm) |
4 |
0.40 |
0.37 |
0.47 |
0.047 |
11.72 |
|
Distance between branch centres (mm) |
12 |
0.43 |
0.35 |
0.52 |
0.058 |
13.74 |
|
Distance between dissepiment centres (mm) |
4 |
0.58 |
0.49 |
0.63 |
0.064 |
11.08 |
|
Aperture width (mm) |
7 |
0.07 |
0.07 |
0.08 |
0.005 |
7.20 |
|
Aperture spacing along branch (mm) |
8 |
0.23 |
0.22 |
0.26 |
0.015 |
6.44 |
|
Maximal chamber width (mm) |
12 |
0.12 |
0.10 |
0.13 |
0.010 |
8.70 |
|
Superstructure opening diameter (mm) |
13 |
0.11 |
0.09 |
0.13 |
0.015 |
13.19 |
|
Distance between node centres (mm) |
10 |
0.32 |
0.26 |
0.35 |
0.023 |
7.22 |
Remarks.—Hemitrypella nodulosa Ernst et al., 2025, differs from Hemitrypella tubulosa Nekhoroshev, 1948, in the wider spacing of branches and dissepiments (average distance between branch centres 0.43 mm vs. 0.33 mm in H. tubulosa; average distance between dissepiment centres 0.58 mm vs. 0.39 mm in H. tubulosa), and in wider spacing of autozooecial apertures (average distance between centres of adjacent apertures 0.23 mm vs. 0.19 mm in H. tubulosa).
Hemitrypella nodulosa Ernst et al., 2025, differs from Hemitrypella nekhoroshevi Ariunchimeg in Ariunchimeg and Morozova, 1992, from the Middle Devonian (Eifelian) of Mongolia in having shorter fenestrules (fenestrule length 0.37–0.47 mm vs. 0.47–0.60 mm in H. nekhoroshevi), and in having smaller autozooecial apertures (aperture width 0.07–0.08 mm vs. 0.14 mm in H. nekhoroshevi).
Stratigraphic and geographic range.—Upper Nims Member of the Junkerberg Formation, Middle Devonian (Eifelian), Brühlborn near Rommersheim, Prüm Syncline in western Rhenish Massif, Germany. Lower Baarley Member of the middle Loogh Formation, Middle Devonian (lower Givetian), “Mühlenwäldchen”, Gerolstein Syncline, Eifel, western Rhenish Massif, Germany. Dobruchna Member in the bottom part of the Skały Formation, uppermost Eifelian (Polygnathus ensensis Conodont Zone), Skały (trench A dug in 1989; see Woźniak et al. 2022), Poland.
Discussion
As emphasized by Racki et al. (1985) and Morozova et al. (2002), bryozoan distribution in the studied successions was mainly influenced by environmental changes corresponding to deepening pulses (Johnson et al. 1985) and correlative global biotic events (Becker 2025). The partial submergence of the vast Stringocephalus Biostromal Bank in the late Givetian during the transgressive cycle IIa was a key ecosystem turnover in the Łysogóry Region and Kostomłoty Zone (Racki et al. 1985, 2022; Fig. 2). As documented by bryozoan-productive sites in the Laskowa Góra Beds (Górno, Laskowa; see Morozova et al. 2002, 2006), the wave of southward expansion of the diverse shelf benthos occurred during the multiphase biotic recovery from the Taghanic global crisis (see also the record of Canutrypa francqana in Skompski et al. 2018). This extra-regional immigration is confirmed in this study by the occurrence of Bigeyella indigena also in the older Górno fauna (and the occurrence of this species in the Frasnian of Armenia; Ernst et al. 2024), as well as by the new monotypic fenestellid genus described from the Wietrznia site. This finding increases the total number of species in the Wietrznia bryozoan association to seven. Four of these species were new (see Morozova et al. 2006), and, therefore, it can be alternatively assumed that the specific Wietrznia biota is endemic.
The early–middle Frasnian transgressive events were also clearly manifested in the subsequent bryozoan fauna occurring in the fossiliferous Górno succession (7 taxa; Morozova et al. 2002, 2006), associated with very diverse brachiopod fauna (Baliński et al. 2016). Notably, this diversification is not recognized in the coeval shallow-water reef-type habitats, especially in the rich benthic biota of Kadzielnia-type organic buildups. Besides Rectifenestella, only tiny problematic lophophorates, the hederellids, are found at the Kadzielnia site.
The present study provides new data concerning the oldest migration to the northern periphery of carbonate platform during the latest Eifelian Kačák (If) event (Fig. 2), recorded in the bryozoan association of the Dobruchna Member. However, taxa lists from both Kiepura (1965, 1973) for the Skały site and Wyse Jackson in Halamski et al. (2022) for the Miłoszów section suggest that the successive early and middle Givetian (pre-Taghanic) faunas were quite different in composition. This strongly suggests that the pulsed Kačák transgression was associated with a significant, possibly long-term, colonization of the Łysogóry carbonate ramp habitats by this benthic group (Halamski et al. 2022). In particular, the very diverse bryozoan fauna from Miłoszów, comprising at least 20 taxa (Wyse Jackson in Halamski et al. 2022), still awaits a systematic study to provide a basis for estimating the regional effects of the major Taghanic extinction (Ernst 2013).
In the present paper, six species are described from the Dobruchna Member: Cyclotrypa sp., Fistuliphragma gracilis Ernst, 2008, Leioclema passitabulatum Duncan, 1939, Intrapora leunisseni Ernst, 2008, Laxifenestella sp., Hemitrypella nodulosa Ernst et al., 2025. From them, Leioclema passitabulatum, Intrapora leunisseni, and Hemitrypella nodulosa are known from the lower Givetian of the Rhenish Massif, whereas Fistuliphragma gracilis has originally been described from the Eifelian of the Rhenish Massif. Moreover, Leioclema passitabulatum is known from the Lower Devonian of Ukraine (Lochkovian) and Czech Republic (Pragian), as well as from the Middle Devonian (Eifelian) of Michigan, USA. Fistuliphragma gracilis was also identified from the Lower to Middle Devonian (Emsian–Eifelian) of Spain (Ernst et al. 2011). Noteworthy, the number of reported bryozoan taxa in the well-known Skały site is currently extended to 15 (adding 7 encrusting taxa of Ctenostomata after Kiepura 1965: table 1).
In summary, during the IIa and IIc transgressions (Fig. 2), drowning steps of the northern part of the Kielce platform were clearly associated with successive colonization stages of the ramp-type environments. Clearly, the gently inclined slope of the platform, which was subject to dynamic change due to organic growth and synsedimentary tectonic movements, offered several attractive, low-energy niches for lophophorates and echinoderms. This is evident from the thriving populations of diverse, sessile, filter-feeding, benthic organisms, primarily brachiopods and crinoids (Racki et al. 1985; Baliński et al. 2016; Skompski et al. 2018; Halamski et al. 2022). Conversely, a major (in regional perspective; Racki et al. 1985; Racki 1993) latest Givetian IIb deepening episode apparently terminated the development of bryozoan-rich benthic biotas in the Kostomłoty Zone, even though impoverished associations then appeared in the southern Kielce Region (Morozova et al. 2002). The observed replacement of locally abundant and diverse Givetian bank faunas by generally impoverished Frasnian reef-related associations was a remarkable regional trend. It largely corresponds to the worldwide apex of Givetian bryozoans, followed by a major diversity decline in the lineage’s evolutionary history (Horowitz et al. 1996; Ernst 2013). However, this turnover probably occurred in two phases in the Holy Cross shelf area (i.e., as a result of the Taghanic and Frasnian crises).
Acknowledgements
AE was supported by the Deutsche Forschungsgemeinschaft (ER 278/4-1 and 2). Catherine Reid (University of Canterbury, Christchurch, UK) and Hans Arne Nakrem (Natural History Museum, Oslo, Norway) are appreciated for their helpful and constructive reviews.
Editor: Andrzej Kaim
References
Anstey, R.L. and Perry, T.G. 1970. Biometric procedures in taxonomic studies of Paleozoic bryozoans. Journal of Paleontology 44: 383–398.
Ariunchimeg, Y. and Morozova, I.P. 1992. New Paleozoic bryozoans of Mongolia. Sovmestnaâ Sovetsko-Mongolskaya Paleontologičeskaâ Ekspeditsiâ, Trudy 4: 75–84.
Astrova, G.G. 1964. New order of paleozoic Bryozoa [in Russian]. Paleontologičeskij žurnal 1964 (2): 22–31.
Astrova, G.G. 1965. Morphology, history of development and system of the Ordovician and Silurian Bryozoa [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 106: 1–432.
Astrova, G.G. 1978. The history of development, system, and phylogeny of the Bryozoa: order Trepostomata [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 169: 1–240.
Astrova, G.G. and Morozova, I.P. 1956. About systematics of the order Cryptostomata [in Russian]. Doklady Akademii Nauk SSSR 110 (4): 661–664.
Astrova, G.G. and Yaroshinskaya, A.M. 1968. Early Devonian and Eifelian Bryozoa of Salair and Gorno-Altai [in Russian]. Trudy Tomskogo Gosudarstvennogo Universiteta 202: 47–62.
Baliński, A., Racki, G., and Halamski, A. T. 2016. Brachiopods and stratigraphy of the Upper Devonian (Frasnian) succession of the Radlin Syncline (Holy Cross Mountains, Poland). Acta Geologica Polonica 66: 125–174. Crossref
Bassler, R.S. 1929. The Permian Bryozoa of Timor. Paläontologie von Timor 16: 37–90.
Bassler, R.S. 1934. Notes on fossil and Recent Bryozoa. Journal of the Washington Academy of Sciences 24: 404–408.
Becker, R.T. 2025. Devonian and lower Carboniferous global events in the central Variscan Orogen. In: U. Linnemann (ed.), The Variscan Orogen of Central Europe. Regional Geology Reviews, 889–978. Springer, Cham. Crossref
Boardman, R.S. 1960. Trepostomatous Bryozoa of the Hamilton Group of New York State. U.S. Geological Survey Professional Papers 340: 1–87. Crossref
Borg, F. 1926. Studies on Recent cyclostomatous Bryozoa. Zoologiska Bidrag från Uppsala 10: 181–507.
Crockford, J. 1944. Bryozoa from the Permian of Western Australia. Part I. Cyclostomata and Cryptostomata from the north-west basin and Kimberly district. Proceedings of the New South Wales Linnean Society 69: 139–175.
Czaplikowska, I. 1990. Opracowanie mikropaleontologiczne środkowodewońskich łupków brachiopodowych z Gór Świętokrzyskich. 79 pp. Unpublished MSc. Thesis, Faculty of Earth Sciences, University of Silesia, Sosnowiec.
Duncan, H. 1939. Trepostomatous Bryozoa from the Traverse Group of Michigan. Contributions from the Museum of Paleontology, University of Michigan 5 (10): 171–270.
Ehrenberg, C.G. 1831. Symbolae Physicae, seu Icones et descptiones Corporum Naturalium novorum aut minus cognitorum, quae ex itineribus per Libyam, Aegiptum, Nubiam, Dongalaam, Syriam, Arabiam et Habessiniam, studia annis 1820–1825, redirent. Pars Zoologica, 4, Animalia Evertebrata exclusis Insectis. 10 pls. Officina Academica Berolini, Berolini.
Elias, M.K. and Condra, G.E. 1957. Fenestella from the Permian of West Texas. Geological Society of America Memoir 70: 1–158. Crossref
Ernst, A. 2008. Non-fenestrate bryozoans from the Middle Devonian of the Eifel (western Rhenish Massif, Germany). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 250: 313–379. Crossref
Ernst, A. 2013. Diversity dynamics and evolutionary patterns of Devonian Bryozoa. Palaeobiodiversity and Palaeoenvironments 93: 45–63. Crossref
Ernst, A. and Gorgij, M.N. 2013. Lower Permian bryozoan faunas from Kalmard area, central Iran. Neues Jahrbuch für Geologie und Paläontologie, Abhandlngen 268: 275–324. Crossref
Ernst, A. and Königshof, P. 2010. Bryozoan fauna and microfacies from a Middle Devonian reef complex (Western Sahara, Morocco). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 568: 1–91.
Ernst, A. and May, A. 2009. Bryozoan fauna from the Konĕprusy Limestone (Pragian, Lower Devonian) of Zlatý Kůň near Konĕprusy (Czech Republic). Journal of Paleontology 83 (5): 767–782. Crossref
Ernst, A. and Schröder, S. 2007. Stenolaemate bryozoans from the Middle Devonian of the Rhenish Slate Massif (Eifel, Germany). Neues Jahrbuch für Geologie und Paläontologie 246: 205–233. Crossref
Ernst, A., Dorsch, T., and Keller, M. 2011. A bryozoan fauna from the Santa Lucia Formation (Lower–Middle Devonian) of Abelgas, Cantabrian Mountains, NW-Spain. Facies 57: 301–329. Crossref
Ernst, A., May, A., and Marks, S. 2012. Bryozoans, corals and microfacies of lower Eifelian (Middle Devonian) limestones at Kierspe, Germany. Facies 58: 727–758. Crossref
Ernst, A., Wyse Jackson, P. N., and Aretz, M. 2015. Bryozoan fauna from the Mississippian (Viséan) of Roque Redonde (Montagne Noire, southern France). Geodiversitas 37: 151–213. Crossref
Ernst, A., Serobyan, V., and Danelian, T. 2024. Biostratigraphic, palaeoenvironmental and palaeobiogeographic implications of bryozoan fauna from the Upper Devonian sequences of Armenia. Geobios 85: 10–18. Crossref
Ernst, A., Suárez Andrés, J.L., and Wyse Jackson, P.N. 2025. Hemitrypa (Fenestrata, Bryozoa) from the Middle Devonian of the Eifel (western Rhenish Massif) and taxonomic re-evaluation of some allied hemitrypid genera. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften (Journal of Applied and Regional Geology) 176: 493–508. Crossref
Gürich, G. 1896. Das Palaeozoicum im Polnischen Mittelgebirge. Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft zu St. Petersbourg (series 2) 32: 1–539.
Hageman, S.J. 1991a. Approaches to systematic and evolutionary studies of perplexing groups: An example using fenestrate Bryozoa. Journal of Paleontology 65: 630–647. Crossref
Hageman, S.J. 1991b. Discrete morphotaxa from a Mississippian fenestrate faunule: presence and implications. In: F.P. Bigey and J.-L. d’Hondt (eds.), Bryozoaires actuels et fossiles: Bryozoa living and fossil. Bulletin de la Société des Sciences Naturelles de l’Ouest de la France, Mémoire HS 1: 147–150.
Halamski, A.T., Baliński, A., Racki, G., Amler, M.R.W., Basse, M., Denayer, J., Dubicka, Z., Filipiak, P., Kondas, M., Krawczyński, W., Mieszkowski, R., Narkiewicz, K., Olempska, E., Wrzołek, T., Wyse Jackson, P.N., Zapalski, M.K., Zatoń, M., and Kozłowski, W. 2022. The pre-Taghanic (Givetian, Middle Devonian) ecosystems of Miłoszów (Holy Cross Mts, Poland). Annales Societatis Geologorum Poloniae 92: 323–379. Crossref
Hall, J. 1858. Report on the Geological Survey of Iowa, embracing the results of investigations made during portions of the years 1855, 1856, 1857. Geological Survey of Iowa, Paleontology 1 (2): 1–724.
Hall, J. 1883. Bryozoans of the Upper Helderberg and Hamilton groups. Transactions of the Albany Institute 10: 145–197.
Horowitz, A.S., Pachut, J.F., and Anstey, R.L. 1996. Devonian bryozoan diversity, extinctions, and originations. Journal of Paleontology 70: 373–380. Crossref
Johnson, J.G., Klapper, G. and Sandberg, C.A. 1985. Devonian eustatic fluctuations in Euramerica. Geological Society of America Bulletin 96: 567–587. Crossref
Kiepura, M. 1965. Devonian bryozoans of the Holy Cross Mountains, Poland; Part I, Ctenostomata. Acta Palaeontologica Polonica 10: 11–48.
Kiepura, M. 1973. Devonian bryozoans of the Holy Cross Mountains, Poland. Part 2. Cyclostomata and Cystoporata. Acta Palaeontologica Polonica 18: 323–400.
King, W. 1849. On some families and genera of corals. Annals and Magazine of the Natural History 2: 388–390. Crossref
Krasnopeeva, P.S. 1935. Bryozoans of the Middle and Upper Devonian of Altai [in Russian]. Materialy po geologii Zapadno-Sibirskogo Kraya 20: 43–84.
Lonsdale, W. 1839. Corals. In: R.I. Murchison (ed.): Silurian System, 675–694. John Murray, London.
M’Coy, F. 1849. On some new genera and species of Palaeozoic corals and foraminifera. Annals and Magazine of Natural History 3: 119–136. Crossref
Ma, J.-Y., Buttler, C.J., and Taylor, P.D. 2014. Cladistic analysis of the “trepostome” suborder Esthonioporina and the systematics of Palaeozoic bryozoans. In: A. Rosso, P.N. Wyse Jackson, and J.S. Porter (eds.), Bryozoan Studies 2013. Studi Trentini di Scienze Naturali 94: 153–161.
Miller, S.A. 1889. North American Geology and Paleontology. 664 pp. Western Methodist Book Concern, Cincinnati.
Morozova, I.P. 1974. Revision of the genus Fenestella [in Russian]. Paleontologičeskij žurnal 1974 (2): 54–67.
Morozova, I.P. 2001. Bryozoans of the order Fenestellida [in Russian]. Trudy Paleontologičeskogo Instituta Rossijskoj Akademii Nauk 277: 1–176.
Morozova, I.P., Weis, O.B., and Racki, G. 2002. Emergence and extinction of the Givetian to Frasnian bryozoan faunas in the Kostomɫoty facies zone, Holy Cross Mountains, Poland. Acta Palaeontologica Polonica 47: 307–317.
Morozova, I.P., Weis, O.B., and Racki, G. 2006. New Devonian and Carboniferous bryozoans of the Holy Cross Mountains (Central Poland). Paleontologičeskij žurnal 2006 (5): 58–67. Crossref
Narkiewicz, M., Racki, G., Skompski, S., and Szulczewski, M. 2006. Procesy i zdarzenia w historii geologicznej Gór Świętokrzyskich. In: S. Skompski and A. Żylińska (eds.), Materiały konferencyjne, LXXVII Zjazd Naukowy Polskiego Towarzystwa Geologicznego, Ameliówka k. Kielc 28–30 czerwca 2006 r., 51–77. Państwowy Instytut Geologiczny, Warszawa.
Nekhoroshev, V.P. 1926. Middle Devonian bryozoans of northwest Mongolia with a description of the microscopic method for the determination of fenestellids [in Russian]. Trudy Geologicheskogo muzeya AN SSSR 1: 1–28.
Nekhoroshev, V.P. 1948. Devonian bryozoa of the Altai [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 3: 1–172.
Nicholson, H.A. 1879. On the Structure and Affinities of the “Tabulate Corals” of the Paleozoic Period, with Critical Descriptions of Illustrative Species. 312 pp. William Blackwood and Sons, Edinburgh, Crossref
Nickles, J.M. and Bassler, R.S. 1900. A synopsis of American fossil Bryozoa including bibliography and synonymy. Bulletin of the United States Geological Survey 173: 1–663. Crossref
Pajchlowa, M. 1957. The Devonian in the Grzegorzowice-Skały section [in Polish, with English summary]. Biuletyn Instytutu Geologicznego 122: 145–254.
Phillips, J. 1841. Figures and Descriptions of the Paleozoic Fossils of Cornwall, Devon and West Sommerset. 231 pp. Longman, Brown, Green, Longmans, London.
Racki, G. 1993. Evolution of the bank to reef complex in the Devonian of the Holy Cross Mountains. Acta Palaeontologica Polonica 37 [for 1992]: 87–182.
Racki, G. 2024. An overlooked contribution to Devonian studies in the Holy Cross Mts: rediscovering Alexei Doronin’s 1893 article on the Kadzielnia Limestone. Acta Geologica Polonica 74: e25. Crossref
Racki, G. and Bultynck, P. 1993. Conodont biostratigraphy of the Middle to Upper Devonian boundary beds in the Kielce area of the Holy Cross Mountains. Acta Geologica Polonica 43: 1–33.
Racki, G., Głuchowski, E., and Malec, J. 1985. The Givetian to Frasnian succession at Kostomloty in the Holy Cross Mts, and its regional significance. Bulletin of Polish Academy of Sciences. Earth Sciences 33: 159–171.
Racki, G., Makowski, I., Mikłas, J., and Gawlik, S. 1993. Brachiopod biofacies in the Frasnian reef-complexes: an ex ample from the Holy Cross Mts, Poland. Prace Naukowe Uniwersytetu Śląskiego 1331, Geologia 12–13: 64–109.
Racki, G., Wójcik, K., Halamski, A.T., and Narkiewicz, M. 2022. Middle Devonian Skały Formation in the Holy Cross Mountains (Poland)—formal description and subdivision based on new field data. Annales Societatis Geologorum Poloniae 92: 425–444. Crossref
Rominger, C. 1866. Observations on Chaetetes and some related genera, in regard to their systematic position; with an appended description of some new species. Proceedings of the Academy of Natural Sciences of Philadelphia 18: 113–123.
Schlüter, C. 1889. Anthozoen des rheinischen Mitteldevon. Abhandlungen zur geologischen Special-Karte von Preussen und den thüringischen Staaten 8 (4): 259–465.
Schulga-Nesterenko, M.I. 1951. Carboniferous Fenestellida of the Russian Platform [in Russian]. Trudy Paleontologičeskogo Instituta 32: 1–157.
Simpson, G.B. 1897. A handbook of the genera of the North American Paleozoic Bryozoa: with an introduction upon the structure of living species. In: 14th Annual Report of the State Geologist (of New York) for the Year 1894, 407–608. James B. Lyon, Albany.
Skompski, S., Baliński, A., Szulczewski, M., and Zawadzka, I. 2018. Middle/Upper Devonian brachiopod shell concentrations from the intra-shelf basinal carbonates of the Holy Cross Mountains (central Poland). Acta Geologica Polonica 68: 607–633.
Snyder, E.M. 1991. Revised taxonomic procedures and paleoecological applications for some North American Mississippian Fenestellidae and Polyporidae (Bryozoa). Palaeontographica Americana 57: 1–275.
Szulczewski, M. 1995. Depositional evolution of the Holy Cross Mts. (Poland) in the Devonian and Carboniferous—a review. Geological Quarterly 3: 471–488.
Troizkaya, T.D. 1975. Main features of development of bryozoans at the boundary between the Devonian and the Carboniferous in Central Kazakhstan [in Russian]. Paleontologičeskii žurnal 1975 (3): 54–69.
Ulrich, E.O. 1882. American Palaeozoic bryozoa. Journal of the Cincinnati Society of Natural History 5: 121–175.
Ulrich, E.O. 1890. Palaeozoic bryozoa: III. Geological Survey 8: 283–688.
Ulrich, E.O. 1896. Bryozoa. In: C. Eastman (ed.), Zittel’s Textbook of Palaeontology, Vol. 1, 257–291. Macmillan, London.
Ulrich, E.O. and Bassler, R.S. 1904. A revision of Palaeozoic Bryozoa. Part II: On genera and species of Trepostomata. Smithsonian miscellaneous collections 47: 15–55.
Vine, G.R. 1884. Fourth report of the Committee consisting of Dr. H.R. Sorby and Mr. G.R. Vine, appointed for the purpose of reporting on fossil Polyzoa. In: Reports of the 53rd Meeting of the British Association for the Advancement in Sciences, 161–209. John Murray, London.
Woźniak, P. 1992. Środkowodewońskie zespoły ramienionogowe z warstw skalskich w Górach Świętokrzyskich. 38 pp. Unpublished M.Sc. Thesis, Faculty of Earth Sciences, University of Silesia, Sosnowiec.
Woźniak, P., Halamski, A.T., and Racki, G. 2022. Cyclic ecological replacement of brachiopod assemblages in the top-Eifelian Dobruchna Brachiopod Shale Member (Skały Formation) of the Holy Cross Mountains (Poland). Annales Societatis Geologorum Poloniae 92: 445–463. Crossref
Wyse Jackson, P.N. 1988. New fenestrate Bryozoa from the Carboniferous of County Fermanagh. Irish Journal of Earth Sciences 9: 197–208.
Yang, J. and Lu, L. 1983. Upper Carboniferous and Lower Permian bryozoans from Kalpin of western Xinjiang. Paleontologia Cathayana 1: 259–317.
Acta Palaeontol. Pol. 71 (1): 1–17, 2026
https://doi.org/10.4202/app.01282.2025