
Ichnotaxonomy and paleobiology of a bird track assemblage from the Miocene Vinchina Formation of La Rioja Province, Argentina
MARTIN E. FARINA, VERÓNICA KRAPOVICKAS, and CLAUDIA A. MARSICANO
Farina, M.E., Krapovickas, V., and Marsicano, C.A. 2026. Ichnotaxonomy and paleobiology of a bird track assemblage from the Miocene Vinchina Formation of La Rioja Province, Argentina. Acta Palaeontologica Polonica 71 (1): 29–50.
This study examines the bird ichnoassemblage of the Miocene Vinchina Formation (La Rioja Province, Argentina), recorded in fluvio-lacustrine environments. Both unpublished material and previously collected specimens were analysed across three transverse sections cropping out from north to south in the Quebrada Pedregal Negro, the Quebrada de la Troya, and the Quebrada del Yeso. The Pedregal Negro creek has yielded only Aviadactyla vialovi, recently synonymised with Ornithotarnocia lambrechti. The low ichnodiversity observed is likely due more to limited exploration efforts and poor outcrop exposure in the creek than to palaeoenvironmental constraints. Exposures at Quebrada de la Troya yield six ichnotaxa preserved in fluvial and lacustrine settings: Aramayoichnus rheae (a large rheid and the oldest rheid track in southern South America), Phoenicopterichnus rector (Phoenicopteridae), Ardeipeda isp. (Ciconiiformes), cf. Jindonornipes, cf. Aquatilavipes (small aquatic birds, possibly Recurvirostridae or Charadriidae), and Rionegrina isp. (a functional didactyl phorusrhacid). At Quebrada del Yeso creek, five ichnospecies of birds were recognised and preserved in a playa-lake environment: Phoenicopterichnus rector, Gragliavipes gavenskii (Threskiornithidae), Avipeda isp. (likely Scolopacidae or Charadriidae), together with a ralliform morphotype and a Cariamidae morphotype. Overall, the trackmakers represented in the three ichnoassemblages correspond well with the globally known Miocene avian body fossil record. While the Pedregal Negro section warrants further exploration, differences in ichnodiversity between Quebrada del Yeso and Quebrada de la Troya likely reflect ecological factors, as the track-bearing levels in the latter represent environments with greater freshwater availability, which may have supported more diverse bird communities.
Key words: Aves, trace fossil, palaeoecology, continental environments, ichnodiversity, fluvial environments, playa-lake, Miocene, Argentina.
Martin E. Farina [martin@gl.fcen.uba.ar; https://orcid.org/0000-0002-1068-6275], Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales; Instituto de Estudios Andinos “Don Pablo Groeber”, Estudios Paleobiológicos en Ambientes Continentales (IDEAN-CONICET), Universidad de Buenos Aires, Ciudad Universitaria, Intendente Güiraldes 2160, C1428EGA, Buenos Aires, Argentina.
Verónica Krapovickas [vkrapovickas@gl.fcen.uba.ar; https://orcid.org/0009-0003-9500-8901], Instituto de Geociencias, UNAM, Blvd. Juriquilla No. 3001, Juriquilla, CP: 76230, Querétaro, México. Instituto de Estudios Andinos “Don Pablo Groeber”, Estudios Paleobiológicos en Ambientes Continentales (IDEAN-CONICET); Laboratorio de Paleontología de Vertebrados, Facultad de Ciencias Exactas y Naturales, Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Ciudad Universitaria, Intendente Güiraldes 2160, C1428EGA, Buenos Aires, Argentina. Machine Learning and Data Analytics Lab, Department of Artificial Intelligence in Biomedical Engineering (AIBE), Friedrich-Alexander University Erlangen-Nürnberg (FAU), Erlangen, Germany.
Claudia A. Marsicano [claumar@gl.fcen.uba.ar; https://orcid.org/0000-0002-0121-6730], Instituto de Estudios Andinos “Don Pablo Groeber”, Estudios Paleobiológicos en Ambientes Continentales (IDEAN-CONICET); Laboratorio de Paleontología de Vertebrados, Facultad de Ciencias Exactas y Naturales, Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Ciudad Universitaria, Intendente Güiraldes 2160, C1428EGA, Buenos Aires, Argentina.
Received 29 August 2025, accepted 17 November 2025, published online 23 February 2026.
Copyright © 2026 M.E. Farina et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Vinchina Formation, located in north-western Argentina, has attracted ichnological interest since the mid-20th century, following the pioneering works of Frenguelli (1950) and Bonaparte (1965). These studies focused on footprints found at Quebrada del Yeso creek and interpreted the age of the unit based on ichnofossils and imprecise K-Ar radiometric ages, alternatively as Permian (Frenguelli 1950), Oligocene (Krapovickas and Nasif 2011), and Pliocene (Bonaparte 1965).
Nearly fifty years later, new ichnological research on the Vinchina Formation has expanded to include both vertebrate (Krapovickas 2010; Krapovickas and Nasif 2011; Melchor et al. 2012; Farina et al. 2021; Vera and Krapovickas 2022; Krapovickas et al. 2017, 2023; De la Fuente et al. 2024) and invertebrate traces (Krapovickas 2010; Melchor et al. 2010; Valencia et al. 2024). The age of the unit has now been constrained using high-precision CA-TIMS U-Pb zircon ages to the early–late Miocene (Ciccioli et al. 2014).
The exceptional diversity and distinct foot morphology observed in the tetrapod footprints make the Vinchina Formation one of the foremost sites for studying Neogene tetrapod faunas in the region, offering valuable palaeontological insights despite the scarcity of body fossils. Reconstructing ancient communities from the ichnological record requires accurately identifying both tracks and their trackmakers. This process involves linking footprints to the broader taxonomic groups best reflecting the observed anatomical features, together with a detailed analysis of their preservation (e.g., Mancuso et al. 2022).
However, establishing a direct correspondence between ichnotaxonomy and traditional taxonomy is not always possible or advisable. To study past communities using footprints, it is useful to move beyond strict taxonomic classification. Approaches such as analysing species richness, size distribution, and aspects of functional morphology provide valuable insights into past ecological structures (e.g., Lockley et al. 2021; Farina et al. 2021).
The Miocene was a period of rapid evolutionary radiation among birds (e.g., Nagy 2020). The diversification of passerines coincided with the spread of temperate habitats, mountain uplift, and glacial retreat (Jetz et al. 2012). Rheids, which inhabited forested environments in Patagonia during the early Miocene, had expanded into open habitats across central, north-western, and north-eastern Argentina by the late Miocene–Pliocene (Noriega et al. 2017). Notably, ecological niches during this period were already comparable to those observed today.
Therefore, the bird track record of the Vinchina Formation is expected to reflect this increasing diversity. This study focuses on the bird footprint record of the Vinchina Formation with the following objectives: (i) to review the morphology and ichnotaxonomy of avian tracks from three different sites; (ii) to identify potential trackmakers; (iii) to compare the ichnological and body fossil records; and (iv) to infer bird communities within their palaeoenvironmental contexts.
Institutional abbreviations.—CPBA, Colección de Paleovertebrados del Repositorio de Colecciones Paleontológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Autónoma de Buenos Aires, Argentina; CRILAR, Centro Regional de Investigaciones Científicas y Transferencia Tecnológica de La Rioja, La Rioja Province, Argentina; PVL, Instituto Miguel Lillo, Colección de Paleozoología de Vertebrados, Tucumán Province, Argentina.
Other abbreviations.—II–III, III–IV, I–IV, interdigital angles; DivTotal, total divarication angle); DI–IV, digit impression number; FL, track length; FW, track width; LI–IV, digit length.
Geological setting
The Vinchina Formation forms part of the Vinchina Basin, which crops out in the Sierra de los Colorados and extends in a north-east to south-west direction. It is bounded to the north by the Toro Negro Range, to the south by the Umango-Espinal Range, to the east by the Valle del Río Bermejo, and to the west by the Bolsón de Jagüel (Ciccioli et al. 2010). The Vinchina Formation was formally defined by Turner (1964), and Ramos (1970) later recognised two members, upper and lower, separated by a low-angle erosive surface (Ramos 1970; Marenssi et al. 2000). The Vinchina succession overlies Palaeogene rocks assigned to the eolian Vallecito Formation and has an average thickness of approximately 3500 m, reaching its maximum (6451 m) at Quebrada del Yeso Creek (Schencman 2016).
Geographically, the Vinchina outcrops are divided into three sectors: southern (Yeso), central (La Troya, Pedregal Negro, Piedras Moras), and northern (Pozuelos, Larga, El Cardón) (Schencman 2016). In this study, we analysed track-bearing levels corresponding to three creeks that cut the succession in an east-west direction: the Quebrada Pedregal Negro, the Quebrada de la Troya, and, further south, the Quebrada del Yeso (Fig. 1).
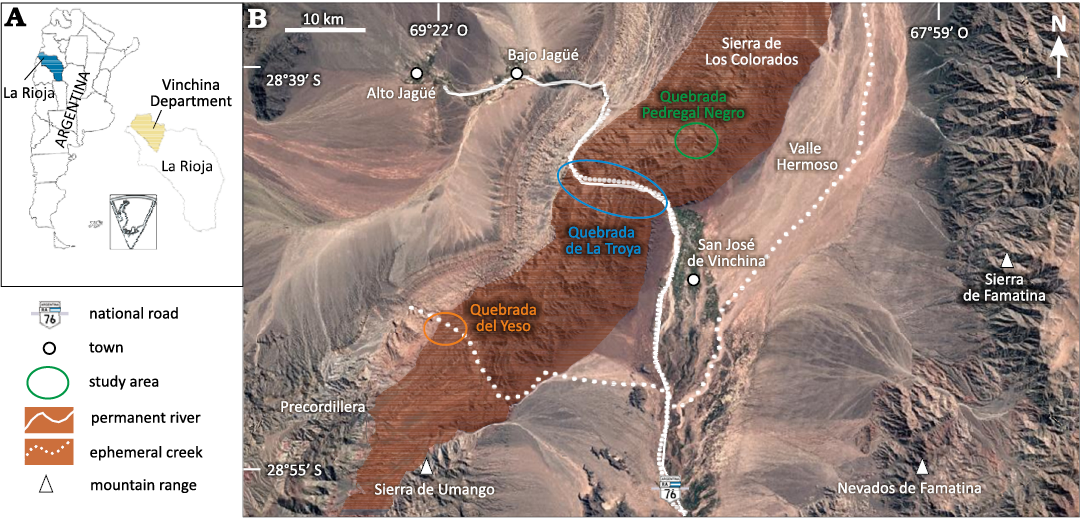
Fig. 1. A. Location of the study area (asterisk) within Argentina. B. Overview of the Vinchina Formation (brown shadow), with study areas (map source: Google Earth 2025).
The Quebrada Pedregal Negro Creek lies within the central sector of the Vinchina Formation outcrops and remains the least studied area, both palaeontologically and geologically. In this sector, the Vinchina Formation reaches a thickness of 1996 m and is composed of reddish sandstones and siltstones with scarce conglomerates and abundant desiccation cracks (Fig. 2) (Schencman 2016). Although Schencman (2016) undertook a detailed facies analysis of this area, difficult access prevented a complete examination of the upper member. The more extensively studied lower member represents a multichannel fluvial system with well-developed interchannel areas where fluvial-aeolian interaction is evident (Schencman 2016).
Here, the analysed bird footprints are preserved in beds of medium-grained sandstones and siltstones interbedded with conglomerates (Fig. 2; “Facies 5” of Schencman 2016). These footprints occur alongside mammal tracks such as Tacheria troyana and Carnivoripeda sudamericana, the latter representing the oldest footprint record of a metatherian carnivore in South America, as well as invertebrate traces typical of the Scoyenia ichnofacies (Krapovickas et al. 2023). Below these track-bearing beds, footprints of a small freshwater turtle attributed to Chelidae or Podocnemidae (cf. Chelonipus torquatus) were also identified (Krapovickas et al. 2023).
The Quebrada de la Troya section represents one of the most thoroughly studied parts of the Vinchina succession and corresponds to the central part of the outcrops. The sedimentary succession reaches a thickness of about 6500 m, with both lower and upper members well developed, and is predominantly composed of fluvial deposits (Schencman et al. 2018). The numerous track-bearing levels in this portion of the Vinchina infilling have been extensively studied in recent years (Vera and Krapovickas 2022; Krapovickas et al. 2023; De la Fuente et al. 2025).
The lower member consists of medium- to coarse-grained sandstone packages, occasionally including thin beds of intraformational and extraformational conglomerates interbedded with mudstones (Tripaldi et al. 2001). The track-bearing intervals in the lower member have been interpreted as the deposits of anastomosed rivers with multi-episodic channels and sandy-muddy floodplains (Tripaldi et al. 2001). Invertebrate trace fossils indicate a range of bioturbation, including Capayanichnus vinchinensis, attributed to crab burrows (Melchor et al. 2010). Tetrapod tracks such as Pichipoda vizcainoi, possibly produced by a dasypodid mammal, have also been recorded (De la Fuente et al. 2025).
The upper member is composed of muddy sandstones intercalated with tuffaceous sandstones and conglomerates containing both fresh and altered volcanic clasts (Limarino et al. 2001; Schencman et al. 2018). Four bird track-bearing intervals have been identified within this member (Fig. 2). The first, from bottom to top, corresponds to multi-episodic channels with moderately developed sandy-muddy floodplain deposits (Tripaldi et al. 2001; Schencman et al. 2018), preserving bird tracks attributed to cf. Aquatilavipes (Fig. 2).
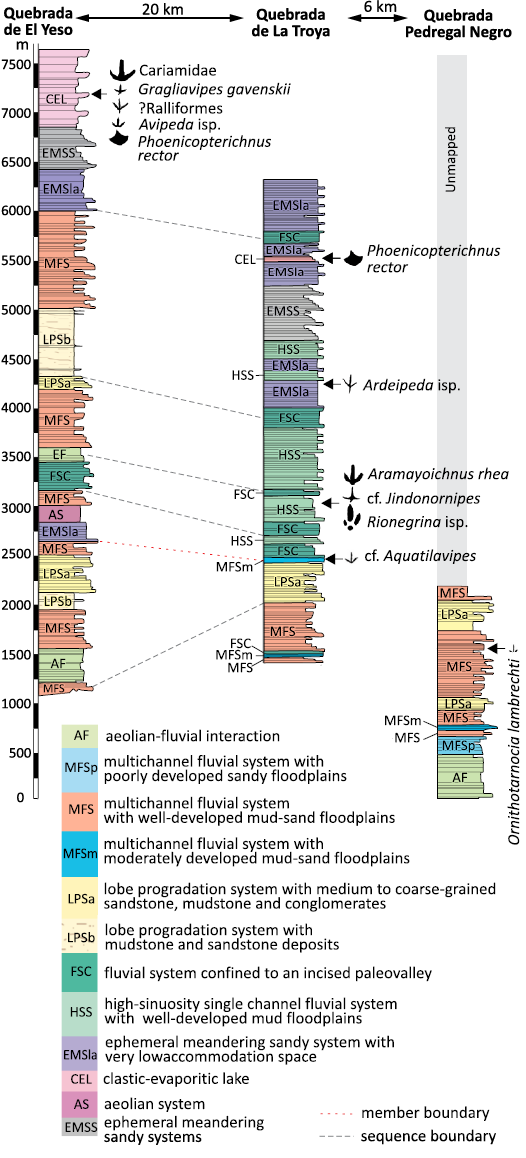
Fig. 2. Lithostratigraphic sections, bird track-bearing levels, and paleoenvironments of the Vinchina Formation in the three studied creeks. Based on Schencman (2016).
The second track-bearing interval has been interpreted as deposited by a meandering fluvial system (Schencman et al. 2018). Invertebrate traces include Skolithos isp., Capayanichnus vinchinensis, horizontal tunnels of Palaeophycus tubularis, and meniscate horizontal traces such as Taenidium barretti, all characteristic of the Scoyenia ichnofacies (Valencia et al. 2024). The tetrapod footprints are found in crevasse-splay deposits (Valencia et al. 2024) and include mammal trackmakers such as Tacheria troyana (a dinomyid rodent), Macrauchenichnus troyana (a macraucheniid litoptern), Llastaya yesera (a toxodontid), and Tridactylonicha marsicanae (a typothere notoungulate) (Krapovickas and Nasif 2011; Vera and Krapovickas 2022; De la Fuente et al. 2025), along with bird tracks belonging to Aramayoichnus rheae isp. nov., Rionegrina isp., and cf. Jindongornipes isp. (Fig. 2).
The third track-bearing interval corresponds to beds deposited in an ephemeral meandering sandy fluvial environment (Schencman et al. 2018), particularly in abandoned fluvial channels (Valencia et al. 2024). Towards the top of this member, the fourth track-bearing interval comprises a 30 m thick clastic-evaporitic lacustrine deposit (Schencman et al. 2018). Melchor et al. (2012) reported the presence of Phoenicopterichnus rector, interpreted as having been produced by flamingo trackmakers.
The southernmost of the studied localities, Quebrada del Yeso Creek, is well known for the ichnological content of its succession and was the first to be studied ichnologically (Frenguelli 1950). The section begins with prominent eolian deposits showing fluvial-aeolian interactions, unique to this area, and is overlain by ephemeral lake deposits, possibly reflecting a progressive decrease in aridity during deposition (Schencman et al. 2018). Towards the top of the succession, a 768 m thick clastic-evaporitic lacustrine system is developed, including abundant track-bearing levels (Facies 11 of Schencman 2016). These correspond to material herein assigned to Phoenicopterichnus rector, Gragliavipes gavenskii, Avipeda isp., a ?ralliform morphotype, and a Cariamidae morphotype.
The Vinchina Formation has been dated using U-Pb zircon ages from three tuffaceous levels. Near the base, Ciccioli et al. (2014) reported an age of 15.6±0.4 Ma, and from the upper third of the succession, an age of 9.24±0.034 Ma. Additionally, Collo et al. (2017) obtained a LA-ICP-MS U-Pb age of 12.62±0.4 Ma from a tuffaceous level observable across all localities in the lower member. Detrital and volcanic zircon U-Pb ages from the overlying Toro Negro Formation range from 6.9 to 2.3 Ma (Amidon et al. 2016), indicating that the Vinchina Formation succession was deposited during the middle to late Miocene.
Material and methods
This study analyses both slabs collected in the field and material examined in situ at the study sites. Measurements of track and trackway parameters were taken from photographs using the open-source software ImageJ (version 1.54d) and directly from the original material with a calliper. Measurements of bird footprints follow McCrea and Sarjeant (2001) (Fig. 3).
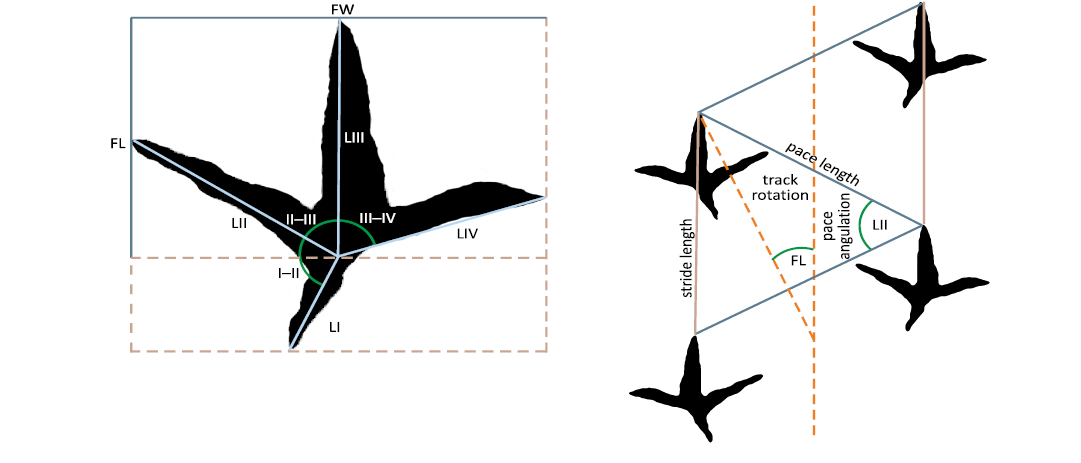
Fig. 3. Measurements on bird footprints following McCrea and Sarjeant (2001).
Terminology and quantitative parameters used to describe footprints were based on Leonardi (1987) and Lallensack et al. (2025): track length (FL), track width (FW), interdigital angles (II–III; III–IV; I–IV), and total divarication angle (DivTotal). We also include digit impression number (Dº) and digit length (Lº). A trackway is defined as a sequence of at least three consecutive tracks produced by the same individual. Parameters used to describe trackways follow Lallensack et al. (2025): pace length, pace angulation, stride length, and track rotation. Descriptive terminology, such as footprint length, relative digit size, and divarication, follows Herrero et al. (2023); however, we prefer to use giant rather than enormous, which we consider grammatically more appropriate.
The ichnotaxobases adopted follow De Valais and Melchor (2008). For ichnogeneric-level diagnosis, these include: number, morphology, relative length, and orientation of digit imprints, especially that of digit I; average FL/FW ratio of footprints; number of phalangeal pad impressions; divarication angles between the impressions of digits II–III, III–IV, and II–IV; presence or absence of webbing; rotation of the footprint relative to the midline; pace; pace angulation; and stride length. For ichnospecific-level diagnosis, the taxobases include: range (maximum and minimum) of FL/FW ratios; proportion of digit lengths I:II:III:IV; range (maximum and minimum) of the angles between all digit impressions; morphology of the sole (metatarsal pad); and range (maximum and minimum) of pace angulation and stride length. We follow Abbassi et al. (2025) for ichnogeneric and ichnospecific validity, and Farina et al. (2025) for ichnofamily classification.
For ichnotaphonomic classification, we use the Taphonomic Tetrapod-Track Modes (TTTM) proposed by Mancuso et al. (2022): High Fidelity Mode (HFM), Moderate Fidelity Mode (MFM), and Low Fidelity Mode (LFM).
Slab reconstructions followed the standard photogrammetric methodology outlined by Falkingham et al. (2018). Sets of photographs were taken with a Canon PowerShot G16 digital camera, and mesh models were generated using Agisoft Metashape Professional (version 1.8.5). Depth-colour maps were produced using the open-source software ParaView 5.11.0 (www.paraview.org). The resulting models are available on FigShare (https://doi.org/10.6084/m9.figshare.29825477). Measurements of the studied material provided in SOM (Supplementary Online Material available at http://app.pan.pl/SOM/app71-Farina_etal_SOM.pdf).
Systematic palaeontology
Ichnofamily Avipedidae Sarjeant & Langston, 1994
Ichnogenus Aramayoichnus Aramayo et al., 2015
Type ichnospecies: Aramayoichnus rheae Aramayo et al., 2015; Late Pleistocene, Pehuen Có, Buenos Aires, Argentina.
Aramayoichnus rheae Aramayo et al., 2015
Fig. 4.
Material.—Four tracks arranged in a trackway in situ preserved at the base of the upper member of the Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. Three tracks were measured digitally, while the remaining one was inaccessible.
Description.—Giant mesaxonic tridactyl tracks without interdigital webbing. Digit III extends beyond digits II and IV. The average footprint length/width ratio (FL/FW) is 1.37, ranging 1.23–1.51. The digits are robust, with distinguishable digital pads, particularly on digit III. The metatarsal pad exhibits a circular shape with an average diameter of 32 mm. The ratio of digits II and IV to digit III is approximately 0.75, with an average length of 126 mm for digit III. The angles are subequal, averaging 27° for II–III and 29° for III–IV, with a total divarication of 54°. Digits II and IV are equal, averaging 96 mm each, while the digit III length is 126 mm. The ratio of digits II and IV to digit III is approximately 0.75. The digits show claw impressions. The metatarsal pad is well preserved at the divergence of all digits, with a circular morphology and an average diameter of 32 mm. The posterior margin of the track forms a V-shape with a smooth apex. The average pace length is 322 mm, ranging from 290 to 355 mm. The average stride length is 646 mm, with a trackway angle of 179°. Track rotation is 4°.

Fig. 4. Rheiform bird track Aramayoichnus rheae Aramayo et al., 2015, trackway in situ preserved at the base of the upper member of the Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. Trackway (A1), detail (A2).
Remarks.—The ichnofamily Avipedidae was originally proposed by Sarjeant and Langston (1994) and later emended by McCrea et al. (2014), who proposed its division into two ichnofamilies: Avipedidae and Limiavipedidae. This reclassification was based on significant differences in track patterns associated with long-legged birds, using the anatomy of the likely trackmaker as a key criterion. However, in the present study, and following the simplified morphological criteria proposed and discussed by Farina et al. (2025), we choose to retain the original classification by Sarjeant and Langston (1994), avoiding divisions that may not necessarily reflect meaningful taxonomic differences.
The ichnogenus was described by Aramayo et al. (2015) based on Pleistocene tracks attributed to rheiform birds showing a strong similarity to the tracks in this study. The most notable difference is the greater width of the digits in the Vinchina specimens. Two other primary differences lie in the length of the lateral digits, which is relatively greater in the Vinchina material, and the interdigital angles, which are more open in the Pehuen Có material. The material from Vinchina locality has a well-defined morphology of the autopodium, corresponding to a High Fidelity Mode (TTTM HFM) compared to that from Pehuen Có, suggesting that these differences may be attributed to taphonomic factors.
Geographic and stratgraphic range.—Base of the upper member of the Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Ichnogenus Avipeda Vyalov, 1965
Type ichnospecies: Avipeda phoenix Vyalov, 1965; Burdigalian (Miocene), Ukraine.
Avipeda isp.
Fig. 5A.
Material.—17 tracks on four slabs (PVL-2680, 2681, 2683, and 2689) preserved at the top of the upper member of the Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Small tridactyl footprints with forward-oriented digits, occasionally connected. The average length is 24 mm and the width is 31 mm. The digits are slender and tubular in shape with well-defined claw impressions. The lateral digits are subequal and 25% shorter than digit III. The interdigital angle between digits II–III measures 65°, between digits III–IV measures 46°, with a total divarication angle (II–IV) of 111° that generally exceeds 100°. The tracks are preserved as concave epirelief.

Fig. 5. Scolopacidae or Charadriidae birds trackways preserved at the top of the upper member of the Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. A. Avipeda isp. (PVL2689), general view of the slab (A1), details (A2–A4), A3 inverted to improve visualization. B. cf. Aquatilavipes isp. (CRILAR-Ic 45), photo (B1), and depth-color map (B2).
Remarks.—The material was originally referred by Bonaparte (1965) to the order Charadriiformes. Slabs PVL-2682 and 2688, each containing a single poorly preserved track, were excluded from the study. Similarly, slabs PVL-2686 and 2687 were not included due to their poor preservation. Slab PVL-2683 contains three tracks, one of which is incomplete. Slab PVL-2680 displays two tridactyl tracks, one right and one left, which likely belong to the same individual. Slab PVL-2689 may represent a different ichnogenus, but the preservation is not optimal. These tracks are characterised by the proximal union of the digits, a feature that distinguishes them from other tracks. This particularity may be due to differences in preservation processes. Additionally, there may be two distinct trackways, although they are not easily distinguishable. These tracks correspond to the Moderate Fidelity Mode (TTTM MFM).
Ichnogenus Rionegrina Melchor et al., 2023
Type ichnospecies: Rionegrina pozosaladensis Melchor et al., 2023; Late Miocene, Pozo Salado locality, Río Negro Province, Argentina.
Rionegrina isp.
Fig. 6.
Material.—Four footprints in situ arranged in a single trackway preserved at the base of the upper member of the Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. Due to their distance and height, it was not possible to place a scale. The description is exclusively morphological and proportional.
Description.—Mesaxonic tridactyl footprints without interdigital webbing. Digit III extends beyond digits II and IV. Digit II is significantly reduced, measuring 65% shorter than digit III, while digit IV is 40% shorter than the same digit. The average FW/FL is 1.46. The digits are thick, with digital pads distinguishable only on digit II, which displays two pads. Interdigital angles are 27° between digits II–III and 37° between digits III–IV. The total divarication angle is 65°. Digits III and IV show claw marks. No metatarsal pad is observed. The digits do not appear to be joined proximally. Step length is 3:1 relative to digit III, and stride is 6:1. The average step angle is 170°. Track rotation is 10°.
Remarks.—Due to the impossibility of including a scale, proportions were used. For this purpose, the value 1 was arbitrarily assigned to digit III of the third track in the trackway (Fig. 6A2). For this reason, the values expressed, with the exception of the angles, do not include units, as they indicate proportions and ratios. Additionally, the digits are heavily eroded, particularly digits II and III, where little depth is observable. Digit II, when preserved, does not appear to be significantly affected by erosion. These tracks correspond to the Moderate Fidelity Mode (TTTM MFM).

Fig. 6. Trackway of Phorusrhacidae bird Rionegrina isp. preserved at the base of the upper member of the Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. Due to the track height, a scale could not be placed but the rough estimation is 37 × 24 cm. General view (A1), details (A2, A3), arbitrarily assigned FL = 1 for comparative purposes.
Ichnogenus Aquatilavipes Currie, 1981
Type ichnospecies: Aquatilavipes swiboldae Currie, 1981; Aptian (Cretaceous), British Columbia, Canada.
cf. Aquatilavipes isp.
Fig. 5B.
Material.—A single isolated track in a slab CRILAR-Ic 45 preserved at the top of the lower member of Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Small
tridactyl footprints with no interdigital webbing. The average length is
28 mm and the width is 35 mm.
The average FL/FW is 0.80. The
total divarication angle
(II–IV) is 105°. The interdigital
angles are 55° between digits
II–III and 50° between digits III–IV.
Digit IV is slightly longer than digit II,
measuring 23 mm and 19
mm, respectively. Both digits show slight claw impressions. Digit
III is 30% longer than the lateral
digits, with a length of 28 mm. The
metatarsal pad is well preserved at the divergence of all digits, but
phalangeal pads are not distinguishable.
Remarks.—The original diagnosis of Aquatilavipes was emended by McCrea and Sarjeant (2001), who proposed characteristics that are clearly observed in the specimens presented in this study, such as digits joined proximally, a well-marked heel, a ratio between digit III and the lateral digits greater than 25%, and an interdigital angle exceeding 95°. However, McCrea et al. (2014) invalidated this amendment by grouping the tracks under the ichnogenus Limiavipes, using size as a criterion along with a multivariate analysis. Nevertheless, size is not a valid ichnotaxobase, and the new diagnosis does not specify the features derived from the multivariate analysis. Therefore, we continue to use the original diagnosis amended by McCrea and Sarjeant (2001). This track corresponds to the High Fidelity Mode (TTTM HFM).
Cariamidae morphotype
Fig. 7.
Material.—Three slabs with 14 footprints identified as PVL-2690, PVL-2691 and PVL-2692 preserved at the top of the upper member of the Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Large mesaxonic tridactyl footprints, unwebbing or semi-palmate. In well-preserved specimens, digit III projects prominently beyond digits II and IV. The average footprint length is 115 mm, with a width of 109 mm, yielding an FL/FW ratio of 1.06. The digits display well-defined digital pads in most cases. The interdigital angle between digits II–III measures 34°, while that between digits III–IV is 42°, resulting in a total divarication angle of 76°. Digit II averages 92 mm in length, contrasting with digit IV at 84 mm, whereas digit III extends to 112 mm. Digit II is approximately 20% shorter than digit III, and digit IV reaches about 0.75 the length of digit III. Claw marks are occasionally present on the digits. A well-preserved metatarsal pad is visible at the divergence of all digits. The posterior margin of the footprint forms a V-shape with a smoothly rounded apex.

Fig. 7. Track of Cariamidae bird Cariamidae indet. (slab PVL-2690) preserved at the top of the upper member of the Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. General view: photograph (A1) and depth-color map (A2); detail: photograph (A3) and depth-color map (A4).
Remarks.—Bonaparte (1965) reported the presence of 18 tracks. At least two of the tracks on slab PVL 2691 appear to exhibit interdigital webbing, which has also been noted by Tineo et al. (2017). Despite some morphological similarity, these specimens differ from A. rheae described. Compared to the specimens from Quebrada de La Troya, the tracks from Quebrada del Yeso exhibit higher anatomical fidelity. The digital pads are more defined, and the digits are thinner. The same is observed with the metatarsal pad, which is well defined. Quantitatively, they are slightly shorter and wider, resulting in a lower FW/FL (1.06 compared to 1.37). Additionally, the interdigital angles and total divarication are significantly larger than in A. rheae. The digits are not subequal. These data suggest that the tracks were produced by a distinct trackmaker (likely a Cariamidae) rather than by rheids (see Discussion below). These tracks correspond to the High Fidelity Mode (TTTM HFM).
Ichnofamily Culcitapedidae Sarjeant & Reynolds, 2001
Ichnogenus Phoenicopterichnus Aramayo et al., 2015
Type ichnospecies: Phoenicopterichnus rector Aramayo & Manera de
Blanco, 1987; Late Pleistocene, Pehien Co, Buenos Aires, Argentina.
Phoenicopterichnus rector Aramayo & Manera de Blanco, 1987
Fig. 8A, B, Table 1, SOM: table 6.
2021 Phoenicopterichnus lucioi; Farina et al. 2021: 9, 11, figs. 9C, 10B–D.
2021 Phoenicopterichnus vinchinaensis; Farina et al. 2021: 9, 11, figs. 9D, 10G–I.
2021 Anatipeda isp.; Farina et al. 2021: 9, 11, figs. 9A, 10A.
Material.—One trackway composed of 4 tracks preserved in situ (digitally reconstructed in 3D using photogrammetry and depth and false-color maps) from the playa lake at the top of the upper member at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. 35 isolated tracks on 11 slabs (CRILAR-Ic 25–35) preserved at the top of the upper member of Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Medium-sized webbed tridactyl bird tracks with thick digits and claw marks. FL/FW approximately 0.84. Total dimensions are 73 mm in length and 89 mm in width, while the divarication between digits II–III and III–IV is subequal, averaging around 50°. The total divarication angle is 109°. The posterior margin is moderately convex. Digit III averages 71 mm in length, while digit IV averages 59 mm in length. The smallest digit is digit II, measuring 57 mm in length. The metatarsal pad impression is well preserved at the convergence of all digits. The webbing inserts at the tips of digits II and IV, while the insertion on digit III ranges from 9 to 13 mm from the tip. A concavity is present, with its base located 45 mm from the posterior margin between digits II and III, and it is less pronounced between digits III and IV, located 51 mm from the posterior margin. The step measures 120 mm, the pace angulation is 120°, and the stride is 232 mm. The track rotation is −11°. The tracks are preserved as convex hyporelief.
Table 1. Summary of descriptive measurements (in mm, except angles). Abbreviations: II–III, III–IV, I–IV, interdigital angles; DivTotal, total divarication angle; FL, track length; FW, track width; LI–IV, digit length.
|
Ichnotaxa |
FL |
FW |
FL/FW |
LI |
LII |
LIII |
LIV |
I–II |
II–III |
III–IV |
II–IV |
DivTotal |
Pace |
Pace |
Stride |
Track rotation |
|
cf. Aquatilavipes isp. |
28 |
35 |
0.80 |
– |
19 |
28 |
23 |
– |
55 |
50 |
– |
105 |
– |
– |
– |
– |
|
Aramayoichnus rheae |
126 |
93 |
1.37 |
– |
96 |
126 |
96 |
– |
28 |
27 |
– |
55 |
322 |
179 |
646 |
5 |
|
Ardeipeda isp. |
96 |
115 |
0.83 |
39 |
72 |
96 |
74 |
117 |
56 |
52 |
108 |
225 |
– |
– |
– |
– |
|
Avipeda isp. |
24 |
32 |
0.81 |
– |
19 |
24 |
20 |
– |
47 |
65 |
– |
112 |
– |
– |
– |
– |
|
Cariamidae |
113 |
108 |
1.02 |
– |
92 |
110 |
82 |
– |
35 |
42 |
78 |
– |
– |
– |
– |
– |
|
cf. Jindonornipes isp. |
39 |
55 |
0.80 |
– |
31 |
37 |
34 |
96 |
64 |
66 |
97 |
221 |
– |
– |
– |
– |
|
Gragliavipes gavenskii |
32 |
46 |
0.71 |
16 |
28 |
32 |
28 |
96 |
58 |
63 |
122 |
214 |
47 |
151 |
96 |
19 |
|
Ornithotarnocia lambrechti |
20 |
21 |
0.95 |
10 |
15 |
20 |
15 |
50 |
44 |
37 |
82 |
90 |
103 |
175 |
195 |
– |
|
Phoenicopterichnus rector |
72 |
88 |
0.84 |
– |
56 |
70 |
59 |
– |
50 |
49 |
100 |
– |
121 |
121 |
233 |
–12 |
|
?Ralliformes indet. |
33 |
36 |
0.92 |
10 |
23 |
33 |
29 |
126 |
50 |
47 |
97 |
223 |
– |
– |
– |
– |
Remarks.—Phoenicopterichnus was established by Aramayo and Manera de Bianco (1987) and emended by Melchor et al. (2012). The ichnogenus was divided into three ichnospecies by Farina et al. (2021) based on statistical clustering (predictive clustering) and the Ward method. The observed characteristics did not have a direct correlation with the ichnotaxobases followed, but the groups appeared sufficiently robust to propose new ichnospecies. Abbassi et al. (2025) considered the ichnospecies P. lucioi and P. vinchinaensis as nomen dubium, as the study was based on isolated tracks and the analysed characters appeared to overlap. We agree with these observations. On the other hand, the same study includes tetradactyl tracks identified as Anatipeda isp. A new review of the material indicates that these are actually overlapping tracks, and they have now been analysed as tridactyl tracks within this group. All tracks correspond to the High Fidelity Mode (TTTM HFM), with the exception of slabs CRILAR-Ic 26, 27, 29, which correspond to the Moderate Fidelity Mode (TTTM MFM).
Stratigraphic and geographic range.— Top of the upper member at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina and top of the upper member of Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Ichnofamily Gruipedidae Sarjeant & Reynolds, 2001
Ichnogenus Jindongornipes Lockley et al., 1992
Type ichnospecies: Jindongornipes kimi Lockley et al., 1992; Jindong Formation, Cenomanian (Cretaceous), Dukemyeon, Koseong-gun, Kyongsangnam Province, Korea.
cf. Jindongornipes isp.
Fig. 8C, Table 1, SOM: table 7.
Material.—Four isolated tracks preserved in situ at the base of the upper member of Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Small semipalmate tetradactyl footprints with poorly developed interdigital webbing. The average length is 38 mm and the width is 54 mm, with an average FL/FW of 0.80. Digits II, III and IV are oriented forward, while digit I is oriented posteromedially. The total interdigital angle (I–IV) is 220°. The lateral interdigital angles show little variation, averaging 63° between digits II–III, while greater variation is observed between digits III–IV, ranging from 47° to 80°, with an average of 66°. The angle between digits II–IV averages 97°, and between digits I–II is 96°, with no webbing observed between them. Digits II and IV are subequal and occasionally preserve claw impressions, as does the well-marked digit III. Digit I is not completely preserved in any track. Digit II measures 30 mm with pronounced curvature in its final third. Digit III is visibly straight and measures 36 mm, while digit IV is also straight with a length of 33 mm. The metatarsal pad is generally preserved at the divergence of all digits. The interdigital webbing is not present in all tracks, being poorly defined and mainly located in the proximal third of the digits, particularly between digits III and IV. The posterior margin of the footprint is slightly convex. No trackways were identified.

Fig. 8. Bird trackways from the upper member of Vinchina Formation at Quebrada de La Troya (A, C) and Quebrada del Yeso (B, D), nearby San José de Vinchina town, La Rioja Province Miocene of Argentina. A, B. Phoenicopterichnus rector Aramayo et al., 2015. A. Trackway (in situ), photograph (A1) and depth-color map (A2). B. Footprint (slab CRILAR-Ic 34), previously assigned to Anatipeda isp. by Farina et al. (2021). C. cf. Jindongornipes isp.trackway (in situ). D. ?Ralliformes footprint (slab PVL-2684).
Remarks.—Not all tracks are well preserved or exhibit complete characteristics. Two tracks are overlapping, one is visibly eroded, and the last one appears covered at the end of digit III. The tracks are preserved as positive hyporelief, except for one that is eroded and shows negative hyporelief. Preservation varies from moderate to poor. Although sharp contours and relatively deep impressions are observed, the digital impressions are not always continuous and exhibit overlapping. These tracks correspond to the Moderate Fidelity Mode (TTTM MFM). Due to these characteristics, it is challenging to assign the tracks to a specific ichnospecies.
?Ralliformes morphotype
Fig. 8D.
Material.—A slab with a single footprint identified as PVL-2684, preserved at the top of the upper member of Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Small
semipalmate tetradactyl footprint with interdigital webbing preserved
between digits III and IV. The
average length is 33 mm and the width is 36 mm,
with an average FL/FW of 0.92. Digits II, III,
and IV are forward-oriented, while
digit I is oriented almost in
opposition to digit III. The total
interdigital angle (I–IV) is 223°.
The interdigital angle between digits I–II measures
126°, digits
II–III 50°, and digits III–IV
47°. Digit I measures 10
mm in length and is slightly fusiform in shape. The
remaining digits are straight except for digit III,
which shows slight curvature. Digit II measures 23 mm, being shorter
than digit IV (29 mm), while digit III measures 33 mm. All
digits except digit I preserve claw impressions, with the most
conspicuous marks on digit IV. No metatarsal or digital pads are
observed. The connection between digits is not well defined, though
interdigital webbing is clearly preserved between digits III and IV.
Remarks.—In the original description, Bonaparte (1965)mentions two tracks located on the same slab. However, only one track is visible, as the other is sectioned and only the tips of the digits are barely distinguishable. Since the original work does not include illustrations, it is difficult to determine whether the slab was later dissected or if it does not match the author’s description. The measurements presented in that work and in this study show minimal variations, possibly due to differences in the methodology used to measure the tracks, which is not detailed in the original work. However, these differences are not significant. These tracks correspond to the High Fidelity Mode (TTTM HFM).
Ichnofamily Ignotornidae Kim et al., 2006
Ichnogenus Ardeipeda Panin & Avram, 1962
Type ichnospecies: Ardeipeda egretta Panin and Avram, 1962, emend. Sarjeant and Langston 1994; Miocene, Romania.
Ardeipeda isp.
Fig. 9A.
Material.—Eight isolated in situ tracks on a fallen slab preserved over an approximately 42 m² surface, slabs identified as CRILAR-Ic 6, 44; showing intense trampling by sloths and Ardeipeda isp., preserved at the first occurrence of ephemeral meandering sandy channels with very low accommodation space on the upper member of Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Giant, semipalmate tetradactyl footprints with elongated, subtriangular digits bearing claw marks. The footprints average 96 mm in length and 115 mm in width, resulting in an FL/FW ratio of 0.83. The total divarication angle (I–IV) is 225°. The interdigital angle I–II measures 116°, digits II–III measures 56°, and between digits III–IV measures 52°. Digit I averages 39 mm in length, digit II 72 mm, digit III 95 mm, and digit IV 74 mm. No metatarsal or digital pads are preserved. The tracks occur as isolated impressions in negative epirelief, and no associated trackways were identified.

Fig. 9. Trackways from the upper member of Vinchina Formation at Quebrada de La Troya (A) and Quebrada del Yeso (B) and near the top of the lower member of Vinchina Formation at Quebrada Pedregal Negro (C), nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. A. Ardeipeda isp. (Ciconiidae) (slab CRILAR-Ic 44), general view (A1), detail (A2 photograph, A3 depth-color map). B. Gragliavipes gavenskii Farina et al., 2025 (slab CRILAR-Ic 42), photograph (B1) and depth-color map (B2). C. Ornithotarnocia lambrechti Kordos, 1985 (slab CPBA-V 14324), photograph (C1), depth-color map (C2), and contour map (C3).
Remarks.—The ichnogenus Ardeipeda was proposed by Panin and Avram (1962) and later amended by Sarjeant and Langston (1994), who pointed out multiple violations of the International Code of Zoological Nomenclature. The revised description by Sarjeant and Langston (1994) characterizes Ardeipeda as tetradactyl tracks, with three forward-oriented digits and a fourth directed backward, lacking interdigital webbing. However, Ardeipeda incerta (junior synonym of A. egretta follow Abbassi et al. 2025), mentioned in the original work by Panin and Avram (1962), shows what appears to be a small interdigital web (Panin 2022). This observation, along with the inclusion of the ichnogenus in the morphofamily Gruipedidae (Sarjeant and Langston 1994; Mustoe 2002), which groups tracks either lacking webbing or with webbing limited to the proximal portion of the digits, suggests that a new emendation of the ichnogenus could be reconsidered in future works. This would avoid the unnecessary creation of new ichnofamilies, as the presence and morphology of interdigital webbing are typically ichnotaxobases at the ichnofamily level. These tracks correspond to the High Fidelity Mode (TTTM HFM).
Ichnogenus Gragliavipes Farina et al., 2025
Type ichnospecies: Gragliavipes gavenskii Farina et al., 2025; Miocene, Quebrada del Yeso, Vinchina Formaton, Argentina.
Gragliavipes gavenskii Farina et al., 2025
Fig. 9B.
Material.—A single slab with 45 tracks, 27 of which are isolated and 18 arranged in two trackways. The collected slab was identified as CRILAR-Ic 42, and its silicone mold was identified as CPBA-V14422. Preserved at the top of the upper member of Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Small semipalmate tetradactyl tracks with interdigital webbing occasionally preserved, most prominently between digits III and IV. The average footprint length is 31 mm and the width is 45 mm, yielding an FL/FW ratio of 0.70. Digits II, III and IV are forward-oriented, while digit I projects posteromedially. The total interdigital angle (I–IV) measures 215°. The interdigital angle between digits I–II is 94°, with no webbing observed between them. The lateral angles range from 41° to 79° between digits II–III (averaging 60°) and from 49° to 72° between digits III–IV (averaging 62°), while the angle between digits II–IV averages 122°. Digit I is elongated and straight, lacking claw impressions, and measures 16 mm in length, approximately 50% of digit III and 2/3 the length of digits II and IV. Digit III is the longest (32 mm) and distinctly acuminate, while digits II and IV are subequal in length, averaging 27–28 mm, with digit IV displaying slight curvature and digit II often more pronounced in this feature. The metatarsal pad is well preserved at the divergence of all digits. Interdigital webbing is inconsistently present across specimens, appearing faint and primarily restricted to the proximal third of the digits, particularly between digits III and IV. The posterior margin of the track is convex with a slightly angular outline. The step length averages 48 mm (ranging from 27 to 66 mm), while the stride length averages 96 mm (varying between 64 and 129 mm). The trackway angle averages 149°, with a broad range from 102° to 178°. Track rotation is moderate, averaging 20°, but exhibiting considerable variation (−50° to 53°).
Remarks.—The tracks are preserved on a slab that was naturally split and later reconstructed in the laboratory. They are preserved as positive epirelief, except for two that appear as negative epirelief. This apparent contradiction in stratinomic preservation can be explained by the detachment of the infill during the natural splitting of the slabs in the field. Preservation varies from very good to poor. The tracks within the trackways correspond to the High Fidelity Mode (TTTM HFM) due to their sharp contours, continuous digital impressions along the trackway, and occasional claw marks. However, isolated tracks on the same surface exhibit overlapping footprints, thicker digital impressions, and a rough texture, resulting in poorly defined shapes and the loss of some morphological features. These isolated tracks would correspond to the Low Fidelity Mode (TTTM LFM). For a detailed ethological and ecological analysis of the trackways, see Farina et al. (2025).
Stratigraphic and geographic range.—Top of the upper member of Vinchina Formation at Quebrada del Yeso, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Ichnofamily Koreanaornipodidae Lockley et al., 2006
Ichnogenus Ornithotarnocia Kordos, 1985
Type ichnospecies: Ornithotarnocia lambrechti Kordos, 1985; Lower Miocene, Ipolytarnóc, Hungary.
Ornithotarnocia lambrechti Kordos, 1985
Fig. 9C.
Material.—A trackway in a slab preserved in situ with 11 footprints. 7 footprints were recovered as silicone molds identified as CPBA-V 14324. Preserved near the top of the lower member of Vinchina Formation at Quebrada Pedregal Negro, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Description.—Small tridactyl bird footprints with no interdigital webbing, occasionally preserving the hallux. The average length is 20 mm and the width is 21 mm. The digits are forward-facing and slender with a tubular shape, showing no fusion between them and no visible claw impressions. Digit III is 25% longer than the lateral digits. The average interdigital angle between digits II–III measures 44°, digits III–IV 37°, and digits II–IV averages 81°, typically below 90°. The total divarication angle averages 90°. The trackway shows an average stride length of 206 mm, ranging from 187 mm to 276 mm, with an average pace angle of 175°, showing little variation (minimum 167°, maximum 179°). The average step length is 116.7 mm, ranging between 92 mm and 193 mm. The footprints form a single trackway with a total length of 1,250 mm. All tracks are preserved as concave hyporelief.
Remarks.—This material was originally assigned to Avidactyla vialovi by Krapovickas et al. (2023). The ichnogenus was later invalidated and synonymized with Ornithotarnocia (Abbassi et al. 2025), as the symmetry of the track and the total divarication angle overlap. We follow this suggestion. These tracks correspond to the High Fidelity Mode (TTTM HFM).
Stratigraphic and geographic range.—Lower member of Vinchina Formation at Quebrada Pedregal Negro, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina.
Discussion
Ichnotaxonomy.—The assignment to Aquatilavipes is problematic due to the limited information available from the trackway. The original ichnogenus and its ichnospecies (including those now grouped under the ichnogenus Limiavipes by McCrea et al. 2014) are considerably larger than the tracks presented here, with the latter being 20–40% smaller than the previously described ichnospecies. However, since size is not considered a valid ichnotaxobase, and its use is recommended to be limited (e.g., De Valais and Melchor 2008; Bertling et al. 2022), this difference alone is not taxonomically decisive. A comparable small tridactyl ichnogenus is Ornithotarnocia. In this case, the main difference lies in the lack of proximal union between digits, and the length of digit III never exceeds 25% more than that of the lateral digits. These features differentiate Ornithotarnocia from the tracks described here. In Aquatilavipes, by contrast, the digits are joined proximally and the track width exceeds its length, unlike Ornithotarnocia. The interdigital angle II–IV is less than 92°, whereas in Aquatilavipes it is over 100°. In Ornithotarnocia, digits II and IV are subequal, whereas in Aquatilavipes, digit IV is longer than digit II. Similarly, the tracks differ from Avipeda, which exhibits a total divarication angle that does not exceed 95°.
The ichnogenus Aramayoichnus was proposed by Aramayo et al. (2015) based on Pleistocene tracks attributed to rheiform birds, which shows notable morphological similarity to the tracks described in this study. The specimens from Pehuen Có exhibit larger pace and stride ratios (approximately 5:1 vs. 5:2 and 10:1 vs. 5:1, respectively). The pace angulation is similar in both ichnotaxa. The interdigital angles are smaller in the Vinchina specimens. In Aramayoichnus, digit III is 50% longer than the lateral digits, whereas in the Vinchina tracks digit III is 40% longer than digit IV.
The ichnogenus was also reported by Tineo et al. (2017) in the upper Miocene–Pliocene Guandacay Formation in San Ramón de la Nueva Orán, Salta Province, Argentina. Until now, this was the oldest known record of rheid tracks in north-western Argentina. However, the Vinchina tracks extend the temporal range of Aramayoichnus rheae to the middle Miocene. The tracks from Guandacay are notably similar to those from Vinchina, particularly in their interdigital angles, although in the Guandacay tracks, the II–III interdigital angle is the narrowest. Additionally, there is a slight difference in the FL/FW ratios. The differences between the Guandacay and Vinchina tracks are based on the same set of characteristics as those distinguishing the Vinchina and Pehuen Có tracks: in the Guandacay tracks, digit II is not reduced, the pace ratio is 4:1 (vs. 5:2), the stride ratio is 8:1 (vs. 5:1), and the pace angulation is similar. Digit III is 40% longer than the lateral digits, exactly matching the specimens in this study but differing from the Pehuen Có specimens (50% longer). These minor morphological and behavioural differences (evidenced in pace and stride ratios) do not warrant the erection of a new ichnospecies.
Aramayoichnus is comparable to tridactyl ichnotaxa included in the ichnofamily Grallatoridae. The main difference is the impression of a metatarsal pad in Aramayoichnus, absent in Grallatoridae. For instance, Anchisauripus, which shows a low FL/FW, displays a wider divarication (Aramayo et al. 2015) and more differentiated digital pads. Grallator, another comparable tridactyl ichnogenus, has a much higher FL/FW (around 2.2) and a more robust digit II compared to the Vinchina specimens, where they are subequal. These differences apply to the entire ichnofamily. The ichnogenus Eubrontes, on the other hand, has a larger FL/FW (between 1.4 and 1.5), and the projection of digits II and IV aligns with that of digit III. Additionally, it displays well-distinguished claw marks, morphologically different from those in the Vinchina specimens, which are not clear. Finally, although not an ichnotaxobase, the size is more than double.
Ardeipeda
differs from other ichnogenera such as Gruipeda in
the size of digit I, which is longer and aligned with digit III. In
contrast, in Gruipeda digit I is shorter,
spur-like, and not oriented with digit III. Additionally, Gruipeda
lacks a defined interdigital webbing. Similarly, Ignotornis
has digit I oriented posteromedially, with a spur-like shape and a
length representing 25% of digit III, whereas in Ardeipeda
it represents 50%. Furthermore, Ardeipeda
does not exhibit the curvature in digits II and IV that is observed in Ignotornis. Finally, the separation (hypex)
between digits II–III and
III–IV shows no difference in Ardeipeda,
unlike Ignotornis, where the hypex III–IV
is positioned further forward than that between II–III. Persiavipes
shows general morphological similarity, but digit I is long and slender,
with no connection to the rest of the track. Additionally, it inserts
obliquely relative to the rest of the track, unlike Ardeipeda,
where digit I is elongated and aligned with digit III. The
interdigital angles II–III and III–IV are different, whereas in Ardeipeda
they are subequal. On the other hand, Iranipeda
is a large tetradactyl track with a similar arrangement to Ardeipeda.
However, the digits are not joined together, and it exhibits prominent
pads, with no traces of interdigital webbing.
Avipeda is a small tridactyl track, with digits occasionally joined, a total divarication greater than 95°, and digit III being 25% wider than the lateral digits. It differs from Koreanaornis in that the latter is often tetradactyl. Abbassi et al. (2025) question whether non-tetradactyl ichnospecies should be included in Koreanaornis. In this case, since no traces of a hallux are observed, Avipeda is inferred. The possible affinities with Ornithotarnocia are not well-defined for Avipeda (Abbassi et al. 2025), but it is an ichnogenus that is occasionally tetradactyl, asymmetrical, and often exhibits proximally joined digits. Another ichnogenus, Aquatilavipes, features proximally joined digits, a digit IV longer than digit II, and an asymmetrical shape.
Jindonornipes was proposed by Lockley et al. (1992) without providing a specific diagnosis, instead offering a description of the specimens. Later, Lockley et al. (2006) included it in the ichnofamily Jindongornipodidae. It differs from other ichnofamilies, such as Gruipedidae, in the orientation of digit I, which is aligned backward with digit III, whereas in Jindongornipodidae, digit I is angled. Additionally, digit II is shorter than digit IV, and they are not subequal. It differs from the ichnofamily Ignotornidae by the more developed interdigital webbing between digits III and IV, which is absent in the Vinchina specimens. Furthermore, Ignotornidae exhibits a hypex that tends to be asymmetrical, while in Vinchina specimens, it is symmetrical.
There are two ichnospecies within the ichnogenus: Jindonornipes kimi and Jindonornipes falkbuckleyi. The latter was the first record for the Cenozoic (Lockley et al. 2021), making the Vinchina record the second for this era. Jindonornipes kimi and J. falkbuckleyi differ in that the latter appears to have a small webbing in the space between digits III and IV, but it is less pronounced or absent between digits II and III. This characteristic is not represented in the Vinchina taxa, as they occasionally show webbing between digits II and III and between III and IV, being more evident in the former case, when preserved. Therefore, it cannot be assigned to an existing ichnospecies or proposed as a new one.
The taxonomic discussion of Gragliavipes gavenskii has been extensively addressed in Farina et al. (2025) when the new ichnogenus was erected (see “Discussion” in Farina et al. 2025).
In Ornithotarnocia, we can observe a morphology comparable to Koreanaornis and Aquatilavipes. Although the affinities between Ornithotarnocia and Koreanaornis do not seem well-defined (Abbassi et al. 2025), Koreanaornis is 30% larger in both length and width. The interdigital angles are much larger between II–III, though smaller between III–IV. The total divarication is up to 40% wider. The average step length is greater in Ornithotarnocia but shows more variation between the maximum and minimum (92 mm and 193 mm compared to 51 mm and 130 mm). The stride is slightly longer in Ornithotarnocia (average 206 mm, minimum 187 mm, maximum 295 mm) compared to Koreanaornis (average 150 mm, minimum 91 mm, maximum 211 mm). Ornithotarnocia is similar to Aquatilavipes in that both are slightly symmetrical; however, differences in size, total divarication, the ratio between digits II and IV, and the proximal union of the digits indicate that they belong to separate taxa.
Phoenicopterichnus has traditionally been compared with Culcitapeda. Melchor et al. (2012) mentioned that Culcitapeda was established based on scarce material of webbed tridactyl tracks with moderate to poor preservation. According to these authors, none of the characteristics used to differentiate it from Phoenicopterichnus are considered diagnostic of Culcitapeda, and until a detailed description of the ichnogenus is provided, it is regarded as a junior synonym of Phoenicopterichnus (Abbasi et al. 2025).
A similar morphology is observed in Uhangrichnus, with the notable difference that Uhangrichnus may occasionally exhibit a hallux (Lockley et al. 2012). Additionally, the ratios of Pace/LIII and Stride/LIII differ. In this study, the ratios are 1.4 and 3.8, respectively, while in Uhangrichnus, they are significantly higher, at 2.1 and 4.2, respectively. The total divarication angle (II–IV) is also higher in Uhangrichnus compared to the tracks presented here (averaging around 111° in contrast to 94° in this work).
The tracks described at Quebrada de La Troya and Quebrada del Yeso show some differences in size. Specifically, the tracks from Quebrada del Yeso are smaller in both length (71 mm vs. 80 mm) and width (86 mm vs. 97 mm). Despite this, the FL/FW ratio remains at 0.84 in both locations. The proportions between the digits are maintained, with only a slight variation in digit II, which shows a slightly smaller ratio relative to digit III. The size difference does not affect the ichnotaxonomic assignment, and therefore, the tracks are also assigned to Phoenicopterichnus rector. However, this size difference holds paleobiological significance and will be analyzed in detail in the corresponding section.
Melchor et al. (2023) described Rionegrina pozosaladensis, didactyl tracks from the Río Negro Formation in Río Negro Province. These are giant tracks with a length exceeding 300 mm, markedly mesaxonic, and with a reduced digit II. Except for size, which cannot be measured numerically, Rionegrina isp. aligns with the tracks in this study. It shows a reduced digit II, an FL/FW of ~1.46, and fusiform impressions in digit III, while digit IV is elongated but shorter. The main difference is the presence of conspicuous claws in the Río Negro specimens, which are not as visible in the Vinchina tracks. The ratio between the digits is exactly the same (digit II is approximately 60% shorter than digit III, and digit IV is 40% shorter than digit III). Based on these proportions and considering that size is not an ichnotaxobase, these tracks are included within the ichnogenus Rionegrina.
Compared to the rheids previously presented in this study, these specimens exhibit a greater pace/LIII (3:1 vs. 5:2) and a larger stride/LIII (6:1 vs. 5:1). This difference may be attributed to a behavioural change. The interdigital angles are identical for II–III but larger for III–IV, resulting in a greater total divarication. The FL/FW ratio is higher (1.46 vs. 1.37), likely due to the reduced width caused by the diminished size of digit II.
Trackmakers.—Aramayoichnus was erected by Aramayo et al. (2015), and its nomenclatural act included referred material comparable to rheids tracks (e.g., Bonaparte 1965; Ramos 1970; Aramayo and Manera de Bianco 1996, 2009; Aramayo 2009). Morphologically, it corresponds to a large-sized bird, clearly cursorial due to the pronounced development of plantar pad structures. The three robust digits, with well-defined digital and plantar pads, are characteristic of extant rheas (e.g., Pteronecmia pennata). It is notable that in the Toro Negro succession (late Miocene to early Pleistocene), which overlies the Vinchina Formation, rhea body fossils have been recorded (Tambussi et al. 2021).
Aramayo et al. (2015) identified differences between the tracks of the Lesser Rhea (Pterocnemia pennata) and the Greater Rhea (Rhea americana), indicating that the former are smaller, exhibit a lower FL/FW ratio, and show a narrower digit divergence angle. Except for the digit divergence angle, the same differences are observed between the tracks from Quebrada La Troya (larger, higher FL/FW) and those from Quebrada del Yeso (smaller, lower FL/FW). Although a direct ichnotaxonomic assignment between fossil tracks and extant species cannot be made, these patterns support the hypothesis of two different trackmakers exhibiting similar ecological habits. These tracks can be distinguished from those of other cursorial birds, such as the Emu (Dromaius novaehollandiae), as the latter is significantly larger (Milàn 2006) and has a reduced digit II. Other extant cursorial birds with similar morphology, such as ostriches (Struthio camelus), differ by being didactyl. Additionally, cassowaries (Casuarius casuarius) are considerably larger than these tracks and display a prominent claw mark on digits II and III (Moore 2003). Moas (Dinornithiformes) also show a much greater size and a significantly wider digit divarication. The morphology of straight toes with well-defined plantar pads further differentiates the Vinchina tracks from these taxa.
The fossil record of rheas is well documented in South America. The oldest known occurrence of the family comes from the middle Paleocene in Chubut Province (Tambussi 1995). In the Neogene, records are of the early Miocene age with the genus Opisthodactylus, including the type species Opisthodactylus patagonicus Ameghino, 1891, from lower Miocene of Patagonia. Tambussi et al. (2021) reported the presence of Opisthodactylus cf. kirchneri Noriega et al., 2017, in the Toro Negro Formation, adjacent to the Vinchina Formation.
According to Noriega et al. (2017), Opisthodactylus–Pteronemia is the sister group of Rhea americana and reflects a biogeographic shift from closed to open habitats. These environmental changes occurred during the Neogene and Pleistocene. These paleoenvironmental and temporal inferences are consistent with the presence of rhea tracks in the Vinchina Formation. Additionally, Opisthodactylus cf. kirchneri extends the biostratigraphic range of the genus from the early to late Miocene and expands its geographic distribution from Patagonia to northwestern Argentina.
As its name suggests, Ardeipeda was originally assigned to a large heron species (Lockley et al. 2007; Anfinson et al. 2009). The tetradactyl foot morphology, with digit I directed posteriorly, is characteristic of both herons and other members of the Order Ciconiiformes. The foot morphology of the Goliath Heron (Ardea goliath), the largest known heron, corresponds well with the Ardeipeda tracks and this species represents a plausible trackmaker. Another possible candidate is the Shoebill (Balaeniceps rex), which has similar size and morphology. These species are sympatric and share low-oxygen wetland habitats. However, in the Shoebill, digit I is oriented posteromedially, not directly backward. The Threskiornithidae family (e.g., ibises and spoonbills) could also be considered within potential trackmakers, but their digit I is also posteromedially oriented. On the other hand, the Ciconiidae family shows a better morphological match. This group includes tetradactyl species with three forward-facing toes and one directed backward, partially webbed feet, and clawed digits. Ciconiidae is a diverse group that inhabits aquatic environments, which aligns with the studied bearing-facies. Additionally, these birds have a gregarious behaviour, breeding in large colonies (Coulter et al. 2020), which is consistent with the trampling surface where the tracks were found (Fig. 10). Considering that Ciconiidae includes more species within the expected range for the Ardeipeda trackmaker, it is more parsimonious to assign the tracks to Ciconiidae rather than Ardeidae, where only the Goliath Heron matches.

Fig. 10. Trampling surface (in situ) with Ciconiidae bird Ardeipeda isp. and sloths tracks, preserved at the upper member of Vinchina Formation at Quebrada de La Troya, nearby San José de Vinchina town, La Rioja Province, Miocene of Argentina. General view (A1), enlargement (A2).
The assignment of cf. Aquatilavipes is complex. This ichnogenus exhibits a morphology widely distributed among aquatic birds. It consists of small tridactyl tracks oriented forward, wider than long, with varying interdigital angles, clearly indicating aquatic or similar birds (Anfison et al. 2009). Even when excluding webbing aquatic birds, it remains difficult to narrow down the classification to specific families. Anfison et al. (2009) compared the size of Aquatilavipes found in North America and identified similarities with the genera Himantopus (Family Recurvirostridae) and Pluvialis (Family Charadriidae). The hypothesis proposed by Anfison et al. (2009) appears to be the most consistent with both morphological and environmental data. Additionally, the Charadriidae family was already present in South America during the Miocene (Tambussi and Degrange 2013).
Avipeda shows a morphology widely distributed among extant birds. The forward-oriented, slender tridactyl footprints ending in claws are found in many families outside of Charadriiformes, the group originally proposed by Bonaparte (1965) as the potential trackmaker. Referring to the opinion of ornithologist C.C. Olrog, Bonaparte suggested that the trackmaker might belong to the family Thinocoridae (seedsnipes), although he did not elaborate much on the hypothesis (Bonaparte 1965). Although there is fossil evidence of this family dating back to the late Miocene (Agnolin et al. 2016), Bonaparte’s suggestion can be considered, at best, speculative. Members of the Thinocoridae family are tetradactyl, with a small digit I, a condition absent in the Vinchina tracks. In support of Bonaparte’s original proposal, this family is commonly found in arid environments (Winkler et al. 2020) and dry lakes (Toner 2020), indicating an ecological match. However, ecological similarity alone is not sufficient to support a confident assignment. Anfison et al. (2009) suggest that, based on size and proportions, the ichnogenus Avipeda could correspond to a trackmaker related to Caladris (e.g., Caladris alba, Family Scolopacidae) or Charadrius (e.g., Charadrius melodus, Family Charadriidae). Anfison et al. (2009) proposal is consistent with most of the available evidence. Although the wide distribution of this morphology across different orders makes it difficult to undoubtedly identify the trackmaker, it most likely belonged to the Scolopacidae or Charadriidae.
In the case of cf. Jindongornipes, a good analysis of the trackmaker affinities cannot be done due to the poor preservation and the limited number of specimens. However, based on some morphological traits (wider-than-long tracks, semipalmate, and posteromedial orientation of the hallux), and the depositional environment where the tracks were found it is reasonable to infer that the trackmaker was an aquatic bird.
The trackmaker of Ornithotarnocia produced small tridactyl tracks without proximal connection, a morphology that is typical of shorebirds and has been previously discussed in Krapovickas et al. (2023). Weidmann and Reichel (1979) and Sarjeant and Reynolds (2001) agree that the morphology and habits are very similar to those of sandpipers, particularly within the family Scolopacidae. However, within the Scolopacidae family, is frequently present semiwebbing between digits III and IV, which are not observed in the trackmaker of Ornithotarnocia. A similar situation is seen in Charadriiformes, which likewise exhibit partial webbing or hallux traces under certain conditions. These discrepancies allow us to rule out representatives of these families as likely trackmakers. Based on the current evidence, we maintain the hypothesis that the trackmaker belongs to the Scolopacidae.
Regarding the trackmaker of Phoenicopterichnus, the morphology quantitatively and qualitatively matches that of modern flamingos. The tridactyl tracks, with well-preserved webbing, curved digits II and IV, and a prominent digital pad, strongly suggest a member of Phoenicopteridae. The proportions of the fossil tracks align with those of modern flamingo tracks. This interpretation is consistent with a previous proposal by Melchor et al. (2012). Although the trackway does not allow for inferences about leg length, the short step might suggest relatively short legs. In the case of Phoenicopterichnus from Quebrada del Yeso, only isolated tracks are available, as no trackway was identified. Farina et al. (2021) conducted a multivariate analysis on these tracks, identifying three groups based on quantitative and qualitative morphological characteristics. While these groups were assigned to three different ichnospecies, it was not proposed that they correspond exactly to modern flamingo species. However, they clearly indicated an increase in diversity within the group, which is supported by the body fossil record (McDonald and Steadman 2022). In South America, the fossil record of flamingos includes members of the family Palaelodidae in Brazil of late Oligocene–early Miocene age (Alvarenga 1990) and in Argentina of the late Miocene–Pliocene age (Agnolín 2009). The modern family Phoenicopteridae is recorded in Brazil from the rocks of late Oligocene, as well as in Argentina and Uruguay of the late Miocene–Pliocene age, with records extending into the Quaternary (Cuello 1988; Alvarenga 1990; Ubilla et al. 1990; Tambussi and Noriega 1998; Noriega and Agnolín 2008).
The tetradactyl and semipalmate Gragliavipes footprints match that of different potential trackmakers. The tracks of Gragliavipes, measuring under 40 mm in length and displaying a short membrane between digits III and IV, suggest a possible link to small ibises (Threskiornithidae), small herons (Ciconiiformes), or even shorebirds (Charadriiformes). Their preservation in lacustrine and fluvial deposits within the Vinchina succession is consistent with the current habitats of all proposed trackmakers. The small size of the tracks resemble those of Charadriiformes, where digit I either fails to leave a trace or is much shorter than in Gragliavipes. The fossil tracks are smaller than those of current Ciconiiformes. The Threskiornithidae family has a larger size and the first digit is oriented inward, which differentiates it from Ciconiiformes, where the first digit aligns with the third. For this reason, a trackmaker from the Threskiornithidae family is inferred. A more extensive discussion on the trackmaker and the fossil record of the potential trackmaker can be found in Farina et al. (2025). Based on the current morphological evidence, Gragliavipes trackmaker is considered a member of the Threskiornithidae, possibly an ibis.
The morphotype assigned to ?Ralliformes was originally proposed by Bonaparte (1965) without going into much detail in the discussion. The author argued that due to the variety of bird groups exhibiting this morphology, it cannot be definitively assigned to any of them. The orientation of the digits, a tetradactyl pes with digit I pointing backward, spans a diversity of birds. Even when limiting potential trackmakers to aquatic birds, the assignment remains difficult. Therefore, the original assignment by Bonaparte is maintained, although it is not definitive.
For Cariamidae, their tracks are approximately 10% shorter and 15% wider than those of Aramayoichnus. The FL/FW varies between the specimens recorded at Quebrada de la Troya and del Yeso. We can infer different but related trackmakers, with similar life habits. Based on the integument present in the plantar pads, it can be assumed that the trackmaker was a bird with cursorial habits, similar to those of the current Rheidae family, although not as developed as in the case of Aramayoichnus. The morphology of the digits and digital pads suggests a distinct trackmaker with similar cursorial habits. The thinner pads (compared to rheids), a defined metatarsal pad joined to the digits, and rounded posterior margin align with a large cursorial bird. Seriemas (Cariamidae) are well-documented in the fossil record for this period and are plausible candidates. Particularly Noriegavis santacrucensis, an early Miocene basal seriema from the Santa Cruz Formation (Noriega et al. 2009). Though inferred to be less cursorial than extant seriemas and capable of sustained flight (Gonzaga 1996; Mayr and Noriega 2013), its phylogenetic affinity supports this hypothesis. The digits are non-subequal, a significant feature as Mayr (2002) identified the short digit II and curved claw as apomorphies of the Cariamidae+Phorusrhacidae clade. In this case, digit IV appears shortest, though this may represent measurement error due to the lack of a clear trackway making it difficult to determine which foot was measured. Additionally, seriemas possess a digital webbing that was also observed in these footprints. These morphological characteristics suggest a trackmaker belonging to Cariamidae, possibly Noriegavis or a closely related taxon.
The footprints attributed to Rionegrina isp. were extensively studied by Melchor et al. (2023). These authors observed that these late Miocene (~8 Ma) well-preserved tracks show a didactyl posture with cursorial adaptations. They also noted a strong mesaxony and that digit II had an elevated claw, which appeared to be used for immobilizing prey. Although the Rionegrina tracks examined during the present work could not be measured precisely because they were inaccessible, it is reasonable to infer a moderately-sized phorusrhacid as a trackmaker as suggested by Melchor et al. (2023).
Geographically, the fossil bird community and ichnological record of the Toro Negro Formation (overlying the Vinchina Formation, late Miocene–early Pleistocene in age) provide a direct comparison. Its base includes passerine tracks and a single incumbent bird footprint (Krapovickas et al. 2009), alongside skeletal remains of eagles (Vinchinavis paka), a probable scavenging condor, and rheids (Tambussi et al. 2021). Despite sharing the same basin, geographic setting, and similar age, the track and body fossil records show little correlation, except for rheids.
Also geographically proximate but in a separate basin, the Las Flores Formation (late Miocene) shares age and paleoenvironmental conditions with the Vinchina Formation. Agnolin et al. (2025) reported a bird-bone assemblage in Las Flores Formation (Iglesia Basin), preserved in fluvial floodplain and ephemeral lake deposits, alongside three Vinchina ichnoassemblages. Both sites are comparable (Table 2), but key differences exist, notably, the absence of cursorial birds in Las Flores Formation, likely reflecting an ecological bias toward aquatic taxa. For example, tadornines exhibit tetradactyl webbed feet, though their elevated hallux may not register in tracks, potentially leading to misidentification as tridactyl (e.g., Phoenicopterichnus-like traces).
Table 2. Comparisons between the bird skeletal record of the Las Flores Formation and the ichnological record from the Vinchina Formation.
|
Skeletal record |
Ichnological record |
|
Palaelodids |
Phoenicopterichnus |
|
Tadornines |
|
|
Heron |
Ardeipeda |
|
Recurvirostrid |
Aquatilavipes |
|
Avipeda |
|
|
Fulica |
?Ralliform |
Ichnodiversity.—The ichnodiversity of the Vinchina Formation varies across the different sections, which seems to correlate with the proximal-distal fluvial gradient observed in the outcrops from north to south (Ciccioli et al. 2010; Fig. 11). The Pedregal Negro corresponds to a multi-channel fluvial system with well-developed inter-channel areas (Schencman 2016). This setting is favourable for track preservation, as indeed, footprints from various vertebrate groups have been reported (Krapovickas et al. 2023). However, the ichnospecies Ornithotarnocia lambrechti is the only avian track recorded to date in this succession. This low ichnodiversity likely reflects limited paleontological sampling rather than environmental constraints, as the site remains sparsely explored. The Quebrada de La Troya section is predominantly composed of fluvial deposits. The first track-bearing level with cf. Aquatilavipes is include anastomosing fluvial succession with multi-episodic channels and sandy-muddy floodplain deposits. The second track-bearing level is the most diverse preserving cf. Jindonornipes, Aramayoichnus rheae, and Rionegrina isp., and it is included in deposits representing a high-sinuosity single-channel fluvial system with well-developed muddy floodplains, The third track-bearing level containing Ardeipeda isp. is included in an ephemeral meandering sandy system with very low accommodation space (Schencman 2016). Finally, near the top of the creek, a clastic-evaporitic lake deposit contains the Phoenicopterichnus tracks. In the Quebrada del Yeso section, five ichnotaxa were identified: Avipeda isp., Gragliavipes gavenskii, Phoenicopterichnus rector, a morphotype attributed to ?Ralliformes, and a morphotype attributed to Cariamidae. Most of the tracks are concentrated in beds near the top of the upper member, close to its contact with the Toro Negro Formation levels. The bearing levels correspond to facies identified as a playa-lake setting (Schencman 2016). In general, the higher diversity of bird tracks is concentrated on the clastic-evaporitic lake beds within the Vinchina succession.
The low ichnodiversity in Quebrada Pedregal Negro relative to the other sites likely indicates limited sampling effort. The differences in ichnotaxa distribution appear to reflect ecological preferences of the trackmakers. The assemblage from Quebrada de La Troya is not only more diverse than that from Quebrada del Yeso, but also shows larger sizes, suggesting a greater niche occupation (Woodward and Hildrew 2002). This pattern aligned with the paleoenvironmental proximal-distal fluvial gradient previously described for the Vinchina Formation. In Quebrada de La Troya, a wide range of fluvial environments with abundant freshwater has recorded bird tracks. These environments likely supported a greater diversity of trackmakers, while the ephemeral lacustrine system in the Quebrada del Yeso section is more restrictive. More stable ecosystems, with low salinity and high nutrient input, are generally less stressful and allow for a more diverse fauna (Scott et al. 2012), possibly explaining the differences between both sections. At the community level, the presence of shared ichnospecies (e.g. Phoenicopterichnus) and others that are different (e.g. Aramayoichnus, Gragliavipes, Rionegrina) may represent the replacement of species between sites, typical of species turnover (Baselga 2010).

Fig. 11. Reconstruction of the inferred ichnofauna from the Vinchina Formation. Trackmakers from Quebrada Pedregal Negro are shown in green, those from Quebrada de La Troya in dark blue, and those from Quebrada del Yeso in light blue. Species with uncertain assignment are indicated in brown. Not to scale.
Conclusions
The Vinchina Formation exhibits remarkable ichnological diversity, forming a rich ichnoassemblage across the three sites analysed. At Quebrada Pedregal Negro only one ichnotaxon, assigned to Ornithotarnocia lambrechti, has been identified. However, given the site’s preservational potential, and the limited exploration, the low ichnodiversity is most likely the result of a sampling bias rather than ecological or sedimentary constraints.
The Quebrada de la Troya section preserves footprints of different types and morphologies along nearly its entire extent. In total, six ichnotaxa were identified, suggesting a diverse avifauna, including Phoenicopterichnus rector, attributed to flamingos that were likely foraging along saline lakes; Ardeipeda isp., represented by numerous superimposed footprints probably produced by members of the family Ciconiiformes; Aramayoichnus rheae, assigned to large rheids and it represents the oldest record of rheiform tracks in Argentina. Additionally, cf. Jindongornipes isp. and cf. Aquatilavipes isp. represent small aquatic birds of uncertain affinities. In the case of Aquatilavipes, the most parsimonious proposal suggests a producer within the families Recurvirostridae or Charadriidae. Finally, Rionegrina isp. was analysed based on morphological proportions, without absolute data, and corresponds to a functionally didactyl footprint attributable to phorusrhacid. All identified taxa are consistent with the known bird fossil record of South America.
Five ichnospecies were recognised in the Quebrada del Yeso, many of which were reassigned from their original descriptions. Phoenicopterichnus rector, attributed to a flamingo, is represented here by numerous individual tracks, and are smaller in size compared to those described in Quebrada de La Troya. Gragliavipes gavenskii is likely a member of the family Threskiornithidae. The morphotype of ?Ralliformes lacks sufficient information to confidently assign a trackmaker, while for Avipeda isp., the trackmaker is likely within the families Scolopacidae or Charadriidae. Finally, a morphotype attributable to Cariamidae was identified, potentially representing a basal seriema form comparable to Noriegavis santacrucensis. The inferred trackmaker from Cariamidae in both sites shows similar ecology and morphology, but based on morphometric characters, we can infer that they are different.
The avian ichnoassemblage from the Vinchina Formation show a strong taxonomic, environmental, and temporal correspondence with the fossil assemblage of the Las Flores Formation (late Miocene), with the exception of the absence of cursorial birds. The ichnodiversity across the Vinchina Formation varies in relation to a proximal-distal fluvial gradient observed along the north-south axis of its outcrops. In Quebrada de La Troya, the dominant freshwater fluvial systems support a higher diversity of bird species. In contrast, the dominant ephemeral and saline lacustrine system of Quebrada del Yeso was ecologically more restrictive. The presence of both shared and distinct ichnospecies at the community level reflects a pattern characteristic of species turnover. Meanwhile, further field studies and collections are needed in Pedregal Negro to make more accurate interpretations and comparissons with the other two sections of the unit.
The Vinchina Formation shows an ichnoassemblage with a high taxonomic and functional diversity, coinciding with the period of expansion of modern bird lineages, which reinforces the idea of ecological enhancement of birds during the late Miocene.
Acknowledgements
Pedro “Peter” González and director Ayelén Carrizo (both Museo Quyllur Ñan, San José de Vinchina, Argentina), the Subsecretaría de Patrimonio of the Province of La Rioja, and the community of Vinchina are acknowledged for their support during the fieldwork. We are also grateful to Pablo Ortiz (PVL) for allowing us to access the collections under his care. Field assistance by Rocío Vera (IDEAN-CONICET, Universidad de Buenos Aires, Argentina), Lucas Fernández Piana (Universidad de San Andrés, Argentina), and Anne Koelewijn (Friedrich-Alexander University Erlangen-Nürnberg, Erlangen, Germany) in Quebrada Pedregal Negro is deeply appreciated. We thank Patricia Ciccioli (IGEBA-CONICET, Universidad de Buenos Aires, Argentina) and Oscar Limarino (Universidad de Buenos Aires, Argentina) for sharing information on new material and providing coordinates to identify classic localities. We also acknowledge the administrative staff of the Instituto de Estudios Andinos “Don Pablo Groeber” for their prompt management and logistical help. We are indebted to the reviewers, Peter Falkingham (Liverpool John Moores University, UK) and Anthony Martin (Emory University, Atlanta, USA) for their valuable comments that greatly improved this manuscript. Finally, we thank Agostina Padovan (Buenos Aires, Argentina) for her help with the English grammar revision and the editor, Eudald Mujal, for his careful and meticulous editorial work. IDEAN Contribution Nº R-533. This research was supported by the Fondo para la Investigación Científica y Tecnológica, Argentina PICT-2020-2635 and Proyecto de cooperación Bilateral CONICET-BAYLAT 2021–2022.
Editor: Eudald Mujal
References
Abbassi, N., Melchor, R.N., and Díaz-Martínez, I. 2025. Ichnotaxonomy of Paleogene and Neogene tetrapod tracks. Vertebrate Ichnology 1: 749–892. Crossref
Agnolín, F.L. 2009. Una nueva especie del género Megapaloelodus (Aves: Phoenicopteridae: Palaelodinae) del Mioceno Superior del noroeste de Argentina. Revista del Museo Argentino de Ciencias Naturales 11: 23–32. Crossref
Agnolín, F.L., Tomassini, R.L., and Contreras, V.H. 2016. Oldest record of thinocoridae (Aves, Charadriiformes) from south America. Annales de paleontologie 102: 1–6. Crossref
Alvarenga, H.M.F. 1990. Flamingos fósseis da bacia de Taubaté, Estado de São Paulo, Brasil: descrição de nova espécie. Anais da Academia brasileira de Ciências 62: 335–345.
Ameghino, F. 1891. Enumeración de las aves fósiles de la República Argentina. Revista Argentina de Historia Natural 1: 255–259.
Amidon, W.H., Ciccioli, P.L., Marenssi, S.A., Limarino, C.O., Fisher, G.B., Burbank, D.W., and Kylander-Clark, A. 2016. U-Pb ages of detrital and volcanic zircons of the Toro Negro Formation, northwestern Argentina: Age, provenance and sedimentation rates. Journal of South American Earth Sciences 70: 237–250. Crossref
Anfinson, O.A., Lockley, M.G., Kim, S.H., Kim, K.S., and Kim, J.Y. 2009. First report of the small bird track Koreanaornis from the Cretaceous of North America: implications for avian ichnotaxonomy and paleoecology. Cretaceous Research 30: 885–894. Crossref
Aramayo, S.A. 2009. A brief sketch of the Monte Hermoso human footprint site, South Coast of Buenos Aires Province, Argentina. Ichnos 16: 49–54. Crossref
Aramayo, S.A. and Manera de Bianco, T. 1987. Hallazgo de una icnofauna continental (Pleistoceno tardío) en la localidad Pehuen-Có (partido de Coronel Rosales), provincia de Buenos Aires, Argentina. Parte II: Carnivora, Artiodactyla y Aves. 4º Congreso Latinoamericano de Paleontología (La Paz), Resúmenes 1: 532–547.
Aramayo, S.A. and Manera de Bianco, T. 1996. Edad y nuevos hallazgos de icnitas de mamíferos y aves en el yacimiento paleoicnológico de Pehuen-Có (Pleistoceno Tardío), Provincia de Buenos Aires, Argentina. Asociación Paleontológica Argentina. Publicación Especial 4 (1): 47–57.
Aramayo, S.A. and Manera de Bianco, T. 2009. Late Quaternary palaeoichnological sites from the southern Atlantic coast of Buenos Aires Province, Argentina: mammal, bird and hominid evidence. Ichnos 16: 25–32. Crossref
Aramayo, S.A., Manera de Bianco, T., Bastianelli, N.V., and Melchor, R.N. 2015. Pehuen Có: Updated taxonomic review of a late Pleistocene ichnological site in Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 439: 144–165. Crossref
Baselga, A. 2010. Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography 19: 134–143. Crossref
Bertling, M., Buatois, L.A., Knaust, D., Laing, B., Mángano, M.G., Meyer, N., Mikulás, R., Minter, N.J., Neumann, C., Rindsberg, A.K., Uchman, A. and Wisshak, M. 2022. Names for trace fossils 2.0: theory and practice in ichnotaxonomy. Lethaia 55 (3): 1–19. Crossref
Bonaparte, J.F. 1965. Nuevas icnitas de la Quebrada del Yeso (La Rioja) y reconsideración de la edad de los afloramientos. Acta Geológica Lilloana 7: 5–16.
Ciccioli, P.L., Limarino, C.O., Friedman, R., and Marenssi, S.A. 2014.New high precision U-Pb ages for the Vinchina Formation: Implications for the stratigraphy of the Bermejo Andean foreland basin (La Rioja Province, western Argentina). Journal of South American Earth Sciences 56: 200–213. Crossref
Cioccili, P.L., Limarino, C.O., Marenssi, S.A., Tedesco, A.M., and Tripaldi, A. 2010. Estratigrafía de la cuenca de vinchina (terciario), sierras pampeanas, provincia de la Rioja. Revista de la Asociación Geológica Argentina 66: 146–155.
Collo, G., Dávila, F.M., Teixeira, W., Nóbile, J.C., Sant, L.G., and Carter, A. 2017. Isotopic and thermochronologic evidence of extremely cold lithosphere associated with a slab flattening in the Central Andes of Argentina. Basin Research 29: 16–40. Crossref
Coulter, M.C., Rodgers, J.A., Jr., Ogden, J.C., and Depkin, F.C. 2020. Wood Stork (Mycteria americana), versión 1.0. In: A.F. Poole and F.B. Gill (eds.), Birds of the World. Laboratorio de Ornitología de Cornell, Ithaca. https://doi.org/10.2173/bow.woosto.01 Crossref
Cuello, J.P. 1988. Lista de las aves fósiles de la región neotropical y de las islas antillanas. Paula-Coutiana 2: 3–79.
Currie, P.J. 1981. Bird footprints from the Gething Formation (Aptian, Lower Cretaceous) of northeastern British Columbia, Canada. Journal of Vertebrate Paleontology 1 (3–4): 257–264. Crossref
De la Fuente, J.M., Vera, R.B., Farina, M.E., and Krapovickas, V. 2025. Small mammal trackways from the Neogene Vinchina Basin, Argentina: Insights into Xenarthra (Dasypodidae), Notoungulata (Typotheria), and Rodentia (Caviomorpha). Journal of South American Earth Sciences 152: 105297. Crossref
De Valais, S. and Melchor, R.N. 2008. Ichnotaxonomy of bird-like footprints: an example from the Late Triassic–Early Jurassic of northwest Argentina. Journal of Vertebrate Paleontology 28: 145–159. Crossref
Falkingham, P.L., Bates, K.T., Avanzini, M., [and 19 more] 2018. A standard protocol for documenting modern and fossil ichnological data. Palaeontology 61: 469–480. Crossref
Farina, M.E., Krapovickas, V., Fernández Piana, L., Vera, R.B., and De Los Ángeles Ordoñez, M. 2021. Flamingo-like footprints and the problem of addressing biological diversity in the past. Historical Biology 33: 912–926. Crossref
Farina, M.E., Krapovickas, V., and Marsicano, C.A. 2025. A new avian footprint taxon (Gragliavipes gavenskii, Ignotornidae) from the Cenozoic of South America and a reappraisal of avian ichnofamilies from the Cretaceous and Cenozoic. Historical Biology [published online https://doi.org/10.1080/08912963.2025.2481654]. Crossref
Frenguelli, J. 1950. Ichnites en el Paleozoico Superior del oeste argentino. Revista de la Asociación Geológica Argentina 5 (3): 136–149.
Gonzaga, L.P. 1996. Family Cariamidae (Seriemas). In: J. del Hoyo, A. Elliot, and J. Sargatal (eds.), Handbook of the Birds of the World. Vol. 3: Hoatzin to Auks. 821 pp. Lynx Editions, Barcelona.
Herrero, C., Herrero, E., Martín-Chivelet, J., and Pérez-Lorente, F. 2022. Vertebrate ichnofauna from Sierra de las Cabras tracksite (late Miocene, Jumilla, SE Spain). Mammalian ichnofauna. Journal of Iberian Geology 48: 241–279. Crossref
Jetz, W., Thomas, G.H., Joy, J.B., Hartmann, K., and Mooers, A.O. 2012. The global diversity of birds in space and time. Nature 491: 444–448. Crossref
Kim, J.Y., Kim, S.H., Kim, K.S., and Lockley, M. 2006. The oldest record of webbed bird and pterosaur tracks from South Korea (Cretaceous Haman Formation, Changseon and Sinsu Islands): more evidence of high avian diversity in East Asia. Cretaceous Research 27: 56–69. Crossref
Kordos, L. 1985. Lábnyomok Az Ipolytarnóci Alsó-Miocén Korú Homokköben. Geologica Hungarica 46: 259–415.
Krapovickas, V. 2010. El rol de las trazas fósiles de tetrápodos en los modelos de icnofacies continentales en ambientes de climas áridos-semiáridos. 358 pp. Doctoral dissertation, Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales, Buenos Aires.
Krapovickas, V. and Nasif, N. 2011. Large caviomorph rodent footprints of the late Oligocene Vinchina Formation, Argentina. Palaeontologia Electronica 14 (2): 12A.
Krapovickas, V., Ciccioli, P.L., Mángano, M.G., Marsicano, C.A., and Limarino, C.O. 2009. Paleobiology and paleoecology of an arid–semiarid Miocene South American ichnofauna in anastomosed fluvial deposits. Palaeogeography, Palaeoclimatology, Palaeoecology 284: 129–152. Crossref
Krapovickas, V., Munoz, N.A., Alonso-Muruaga, P.J., Ciccioli, P.L., Marenssi, S., and Vizcaíno, S.F. 2018. Tracheria troyana (Krapovickas and Nasif, 2011): redefinition, environmental distribution, and heritage conservation. Ichnos 25: 221–231. Crossref
Krapovickas, V., Vera, R.B., Farina, M.E., Piana, L.R.F., and Koelewijn, A. 2023. A new vertebrate ichnological association sheds light on the small metatherian record of the middle Miocene in South America. Journal of South American Earth Sciences 129: 104529. Crossref
Lallensack, J.N., Leonardi, G., and Falkingham, P.L. 2025. Glossary of fossil tetrapod tracks. Palaeontologia Electronica 28 (1): a8. Crossref
Leonardi, G. (ed.) 1987. Glossary and Manual of Tetrapod Footprint Palaeoichnology. 75 pp., 20 plates. Departamento Nacional da Produçao Mineral, Brasil.
Limarino, C., Tripaldi, A., Marenssi, S., Net, L., Re, G., and Caselli, A. 2001. Tectonic control on the evolution of the fluvial systems of the Vinchina Formation (Miocene), northwestern Argentina. Journal of South American Earth Sciences 14: 751–762. Crossref
Lockley, M.G., Houck, K., Yang, S.Y., Matsukawa, M., and Lim, S.K. 2006. Dinosaur-dominated footprint assemblages from the Cretaceous Jindong Formation, Hallyo Haesang National Park area, Goseong County, South Korea: evidence and implications. Cretaceous Research 27: 70–101.
Lockley, M., Kim, K.S., Lim, J.D., and Romilio, A. 2021. Bird tracks from the Green River Formation (Eocene) of Utah: ichnotaxonomy, diversity, community structure and convergence. Historical Biology 33: 2085–2102. Crossref
Lockley, M.G., Lim, J.D., Kim, J.Y., Kim, K.S., Huh, M., and Hwang, K.G. 2012. Tracking Korea’s early birds: a review of Cretaceous avian ichnology and its implications for evolution and behaviour. Ichnos 19: 17–27. Crossref
Lockley, M.G., Reynolds, R., Milner, A.C., and Varhalmi, G. 2007. Preliminary overview of mammal and bird tracks from the White Narrows Formation, southern Nevada. In: S.G. Lucas, J.A. Spielmann, and M.G. Lockley (eds.), Cenozoic Vertebrate Tracks and Traces. New Mexico Museum History and Science Bulletin 42: 91–96.
Lockley, M.G., Yang, S.Y., Matsukawa, M., Fleming, F., and Lim, S.K. 1992. The track record of Mesozoic birds: evidence and implications. Philosophical Transactions of the Royal Society of London B 336: 113–134. Crossref
Mancuso, A.C., Marsicano, C.A., and Krapovickas, V. 2022. Tetrapod-track taphonomic modes (TTTM) applied to Triassic lacustrine systems from southern South America. Journal of African Earth Sciences 196: 104735. Crossref
Marenssi, S.A., Net, L., Caselli, A., Tripaldi, A., and Limarino, C.O. 2000. Hallazgo de discordancias intraformacionales en la Formación Vinchina (Neógeno), quebrada de La Troya, La Rioja, Argentina. Revista de la Asociación Geológica Argentina 55: 414–418.
Mayr, G. 2002. A new specimen of Salmila robusta (Aves: Gruiformes: Salmilidae n. fam.) from the Middle Eocene of Messel. Paläontologische Zeitschrift 76: 305–316. Crossref
Mayr, G. and Noriega, J.I. 2013. A well-preserved partial skeleton of the poorly known early Miocene seriema Noriegavis santacrucensis. Acta Palaeontologica Polonica 60: 589–598. Crossref
McCrea, R.T. and Sarjeant, W.A.S. 2001. New ichnotaxa of bird and mammal footprints from the Lower Cretaceous (Albian) Gates Formation of Alberta. In: D.H. Tanke and M.W. Skrepnick (eds.): Mesozoic Vertebrate Life, 453–478. Indiana University Press, Bloomington.
McCrea, R.T., Buckley, L.G., Plint, A.G., Currie, P.J., Haggart, J.W., Helm, C.W., and Pemberton, S.G. 2014. A review of vertebrate track-bearing formations from the Mesozoic and earliest Cenozoic of western Canada with a description of a new theropod ichnospecies and reassignment of an avian ichnogenus. New Mexico Museum of Natural History and Science Bulletin 62: 5–93.
McDonald, H.G. and Steadman, D.W. 2023. Fossil Flamingo (Phoenicopteriformes) from the Miocene (Hemingfordian) of Southern California, USA. Historical Biology 35: 1574–1582. Crossref
Melchor, R.N., Cardonatto, M.C., and Visconti, G. 2012. Palaeonvironmental and palaeoecological significance of flamingo-like footprints in shallow-lacustrine rocks: an example from the Oligocene–Miocene Vinchina Formation, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 315: 181–198. Crossref
Melchor, R.N., Feola, S.F., Cardonatto, M.C., Espinoza, N., Rojas-Manriquez, M.A., and Herazo, L. 2023. First terror bird footprints reveal functionally didactyl posture. Scientific Reports 13 (1): 16474. Crossref
Melchor, R.N., Genise, J.F., Farina, J.L., Sánchez, M.V., Sarzetti, L., and Visconti, G. 2010. Large striated burrows from fluvial deposits of the Neogene Vinchina Formation, La Rioja, Argentina: a crab origin suggested by neoichnology and sedimentology. Palaeogeography, Palaeoclimatology, Palaeoecology 291: 400–418. Crossref
Milàn, J. 2006. Variations in the morphology of emu (Dromaius novaehollandiae) tracks reflecting differences in walking pattern and substrate consistency: ichnotaxonomic implications. Palaeontology 49: 405–420. Crossref
Moore, L. 2003. Ecology and Population Viability Analysis of the Southern Cassowary Casuarius casuarius johnsonii Mission Beach, North Queensland. 222 pp. Doctoral dissertation, James Cook University, Townsville.
Mustoe, G.E. 2002. Eocene bird, reptile, and mammal tracks from the Chuckanut Formation, northwest Washington. Palaios 17 (4): 403–413. Crossref
Nagy, J. 2020. Biologia Futura: rapid diversification and behavioural adaptation of birds in response to Oligocene–Miocene climatic conditions. Biologia Futura 71 (1): 109–121. Crossref
Noriega, J.I. and Agnolin, F.L. 2008. El registro paleontológico de las Aves del “Mesopotamiense” (Formación Ituzaingó; Mioceno tardío-Plioceno) de la provincia de Entre Ríos, Argentina. In: F.G. Aceñolaza (ed.), Temas de la biodiversidad del Litoral Fluvial Argentino III. Instituto Superior de Correlacion Geologica, Miscelanea 17 (2): 271–290.
Noriega, J.I., Vizcaíno, S.F., and Bargo, M.S. 2009. First record and a new species of seriema (Aves: Ralliformes: Cariamidae) from Santacrucian (early–middle Miocene) beds of Patagonia. Journal of Vertebrate Paleontology 29: 620–626. Crossref
Noriega, J.I., Jordan, E.A., Vezzosi, R.I., and Areta, J.I. 2017. A new species of Opisthodactylus Ameghino, 1891 (Aves, Rheidae), from the late Miocene of northwestern Argentina, with implications for the paleobiogeography and phylogeny of rheas. Journal of Vertebrate Paleontology 37 (1): e1278005. Crossref
Panin, N. 2022. The palaeo-ichnology of the Miocene deposits in the Romanian Carpathians. Geo-Eco-Marina 28: 73–87.
Panin, N. and Avram, E. 1962. Noe urme de pas de vertebrate in Miocenul Subcarpatilor Ruminestkya. Studie si Cercetari de Gélogie, Géophyzica, si Géografie, Serie de Géologie 7: 455–484.
Ramos, V. 1970. Estratigrafıa y estructura del Terciario en la sierra de los Colorados (Provincia de La Rioja), Republica Argentina. Revista de la Asociación Geológica Argentina 25: 359–382.
Sarjeant, W.A.S. and Langston, W. 1994.Vertebrate footprints and invertebrate traces from the Chadronian (Late Eocene) of Trans-Pecos Texas. Bulletin (Texas Memorial Museum) 36: 1–86.
Sarjeant, W.A. and Reynolds, R.E. 2001. Bird footprints from the Miocene of California. In: R. Reynolds (ed.): The Changing Face of the East Mojave Desert. Abstracts from the 2001 Desert Symposium, 21–40. California State University, Desert Studies Consortium, Riverside.
Schencman, L.J. 2016. Arquitectura de sistemas fluviales, modelos depositacionales y estratigrafía secuencial en cuencas de antepaís: la Formación Vinchina, un caso de estudio. 269 pp. Doctoral dissertation, Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales, Buenos Aires.
Schencman, L.J., Marenssi, S.A., and Díaz, M. 2018. Evolución paleoambiental de la Formación Vinchina (Mioceno) en la sierra de Los Colorados, La Rioja, Argentina. Revista de la Asociación Geológica Argentina 75 (1): 17–38.
Scott, J.J., Buatois, L.A., and Mangano, M.G. 2012. Lacustrine environments. Developments in Sedimentology 64: 379–417. Crossref
Tambussi, C.P. 1995. The fossil Rheiformes from Argentina. Courier Forschunginstitut Senckenberg 181: 121–129.
Tambussi, C.P. and Degrange, F.J. 2013.The paleogene birds of South America. In: C.P. Tambussi and F.J. Degrange (eds.): South American and Antarctic Continental Cenozoic Birds: Paleobiogeographic Affinities and Disparities, 29–47. Springer Netherlands, Dordrecht. Crossref
Tambussi, C.P. and Noriega, J.I. 1998. Registro de presbiornítidos (Aves, Anseriformes) en sedimentitas de la Formación Vaca Mahuida, La Pampa, Argentina. Publicación Electrónica de la Asociación Paleontológica Argentina 5: 51–54.
Tambussi, C.P., Degrange, F.J., Ciccioli, P.L., and Prevosti, F. 2021. Avian remains from the Toro Negro Formation (Neogene), Central Andes of Argentina. Journal of South American Earth Sciences 105: 102988. Crossref
Tineo, D.E., Pérez, L.M., Bona, P., Comerio, M.A., Noriega, J.I., and Poiré, D.G. 2017. The oldest record of Aramayoichnus rheae from the Neogene of northwestern Argentina. Ameghiniana 55: 109–116. Crossref
Toner, S. 2020. Least Seedsnipe (Thinocorus rumicivorus), version 2.0. In: T.S. Schulenberg, S.M. Billerman, and B.K. Keeney (eds.), Birds of the World. Laboratorio de Ornitología de Cornell, Ithaca. https://doi.org/10.2173/bow.thinoc1.01 Crossref
Tripaldi, A., Net, L., Limarino C., Marenssi, S., and Caselli, A. 2001. Paleoambientes sedimentarios y procedencia de la Formación Vinchina, Mioceno, noroeste de la provincia de La Rioja. Revista de la Asociación Geológica Argentina 56: 443–465.
Turner, J.C.M. 1964. Descripción Geológica de la Hoja 15 c, Vinchina. Provincia de la Rioja. Escala 1:200 000. Carta Geológico-Económica de la República. 81 pp. Dirección Nacional de Geología y Minería, Buenos Aires.
Ubilla, M., Perea, D., Tambussi, C., and Tonni, E.P. 1990. Primer registro fósil de Phoenicopteridae (Aves: Charadriiformes) para el Uruguay (Mio-Plioceno). Anais da Academia Brasileira de Ciências 62: 61–68.
Valencia, G.L., Buatois, L.A., Mángano, M.G., Farina, M.E., and Krapovickas, V. 2024.Evaluating trace fossils, fluvial architecture, and colonization patterns in channel and overbank deposits from the Miocene Vinchina Formation, western Argentina. Palaios 39: 363–383. Crossref
Vera, R.B. and Krapovickas, V. 2022. Paleobiology and paleoecology of ungulates from the Miocene of South America based on an ichnological analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 606: 111247. Crossref
Vialov, O.S. 1965. Stratigrafiâ neogenovyh molass Predkarpatskogo progiba. 194 pp. Naukova Dumka, Kiev.
Weidmann, M. and Reichel, M. 1979. Traces de pattes d’oiseaux dans la Molasse suisse. Eclogae Geologicae Helvetiae 72 (3): 953–971.
Winkler, D.W., Billerman, S.M., and Lovette, I.J. 2020. Seedsnipes (Thinocoridae), versión 1.0. In: S.M. Billerman, B.K. Keeney, P.G. Rodewald, and T.S. Schulenberg (eds.), Birds of the World. Laboratorio de Ornitología de Cornell, Ithaca. https://doi.org/10.2173/bow.thinoc1.01 Crossref
Woodward, G.U.Y. and Hildrew, A.G. 2002. Body‐size determinants of niche overlap and intraguild predation within a complex food web. Journal of Animal Ecology 71: 1063–1074. Crossref
Acta Palaeontol. Pol. 71 (1): 29–50, 2026
https://doi.org/10.4202/app.01284.2025