
Reassessing diagnostic postcranial traits in Pleistocene elephants: evidence from Palaeoloxodon antiquus and Mammuthus in Italy
DARIO STEFANELLI, MARIA MARINO, BENIAMINO MECOZZI, RAFFAELE SARDELLA, ANDREA ZAZZERA, and MARCO P. FERRETTI
Stefanelli, D., Marino, M., Mecozzi, B., Sardella, R., Zazzera, A., and Ferretti, M.P. 2026. Reassessing diagnostic postcranial traits in Pleistocene elephants: evidence from Palaeoloxodon antiquus and Mammuthus in Italy. Acta Palaeontologica Polonica 71 (1): 155–171.
The fossil elephant remains from the lower Middle Pleistocene site of Contrada Monticelli (Apulia, southern Italy) are described here in detail for the first time. The material, consisting of dental and postcranial bones from a single individual, was analyzed both morphologically and morphometrically. Estimates of shoulder height and body mass were calculated for the specimen. Based on dental characters, the Contrada Monticelli elephant is assigned to Palaeoloxodon antiquus. A comparative analysis with Mammuthus and Palaeoloxodon specimens from the Lower and Middle Pleistocene of Italy indicates the Contrada Monticelli individual represents a medium-sized, old male straight-tusked elephant. The comparative study also demonstrated that several morphological traits of the humerus, ulna, femur, and tibia, previously proposed as diagnostic in distinguishing between Mammuthus and Palaeoloxodon, are not consistent when intraspecific variability is taken into account. Furthermore, the morphometric analyses revealed no significant differences in size and proportions of the studied limb bones between Mammuthus meridionalis and Palaeoloxodon antiquus that would allow for a reliable discrimination between the two taxa. The Contrada Monticelli elephant represents one of the few known European straight-tusked elephant skeletons dated to the interval between 0.8 and 0.5 million years ago, corresponding to the dispersal of the genus into Eurasia, and contributes to a better understanding of the biochronological, paleoecological, and paleobiogeographical context of the early evolution of Palaeoloxodon in Europe.
Key words: Elephantidae, Palaeoloxodon, morphology, anatomy, Middle Pleistocene, Mediterranean Europe, Italy, Contrada Monticelli.
Dario Stefanelli [dario.stefanelli@uniba.it; ORCID: https://orcid.org/0000-0002-6601-1875], Maria Marino [maria.marino@uniba.it; ORCID: https://orcid.org/0000-0001-6239-0786] and Andrea Zazzera [andrea.zazzera94@gmail.com; ORCID: https://orcid.org/0000-0001-7989-5449], Dipartimento di Scienze della Terra e Geoambientali, Università degli Studi di Bari Aldo Moro, Campus universitario, via Orabona 4, 70126, Bari, Italy.
Beniamino Mecozzi [beniamino.mecozzi@uniroma1.it; ORCID: https://orcid.org/0000-0002-7097-3217], Dipartimento di Biologia Ambientale, Sapienza Università di Roma, Piazzale Aldo Moro 5, I-00185, Rome, Italy.
Raffaele Sardella [raffaele.sardella@uniroma1.it; ORCID: https://orcid.org/0000-0002-9752-6281], Dipartimento di Scienze della Terra, laboratorio PaleoFactory, Sapienza Università di Roma, Piazzale Aldo Moro 5, 00185, Roma, Italy.
Marco P. Ferretti [marcopeter.ferretti@unicam.it; ORCID: https://orcid.org/0000-0002-9660-628X], Sezione di Geologia, Università degli Studi di Camerino, Via Gentile III da Varano 7, I-62032, Camerino (Macerata), Italy.
Received 31 October 2025, accepted 15 January 2026, published online 26 March 2026.
Copyright © 2026 D. Stefanelli et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Proboscideans are one of the most commonly found groups of Quaternary large mammals. Although more frequently found at open-air palaeontological sites, their remains are also not uncommon in cave or karstic settings (Schreve and Jones 2005). In Europe, after the extinction of the tetralophodontid gomphothere Anancus arvernensis at around 2.0 Ma, proboscideans were represented exclusively by members of the family Elephantidae. Five elephantid species are currently recognized in the Pleistocene of continental Europe: the mammoths Mammuthus rumanus (latest Pliocene–early Early Pleistocene), Mammuthus meridionalis (Early Pleistocene), Mammuthus trogontherii (Middle Pleistocene), Mammuthus primigenius (Late Pleistocene), and the straight tusked elephant Palaeoloxodon antiquus (Middle–Late Pleistocene). All these species are known from the Quaternary of Italy, in many cases represented by complete skeletons (Palombo and Ferretti 2005). The paleoclimatic and paleoenvironment significance of elephantids is well established and elephants are widely recognized as ecologically key-stone species (Haynes 1991). In addition, during the Quaternary humans interacted with elephantids in complex ways, ranging from scavenging and hunting to potential cultural and symbolic association (Konidaris and Tourloukis 2021; Palombo and Cerilli 2021). Identification of elephant fossil remains at the generic or species level is essential in investigating all these aspects.
The straight tusked elephant coexisted in western Europe with M. trogontherii and M. primigenius during much of the Middle and Late Pleistocene, while the earliest Palaeoloxodon record in Europe may be contemporaneous with the latest M. meridionalis populations. A major issue in the study of Pleistocene elephants in Europe is that in absence of dental (especially molars) and cranial remains, it is often a difficult task to discriminate between the two genera. As a matter of fact, postcranial elements of different elephant genera, both extant and extinct, are often difficult or even impossible to distinguish, due to their similar size and morphology (Lister and Stuart 2010; Athanassiou 2011). Overall, the role of postcranial bones in the taxonomic attribution of proboscidean fossil remains is still controversial and has not received sufficient attention. As a result, taxonomic attributions are often tentative and based on the biochronological, paleogeographical, and paleoenvironmental context.
In this paper we describe for the first time in detail the elephantid material from the lower Middle Pleistocene site (Galerian; Isernia FU) of Contrada Monticelli (CM) (Apulia, southern Italy), comparing it to several skeletons of P. antiquus and M. meridionalis, from various Italian sites. Aims of this study is to: (i) provide a definitive taxonomic identification of the CM sample; (ii) investigate in depth the morphological and morphometric characters of the limb bones, in order to verify the validity of the morphological differences between European Palaeoloxodon and Mammuthus suggested so far by previous authors, and, on the other hand, evaluate the possible identification of new distinctive characters; (iii) determine whether the CM elephantid sample represents a single individual and provide an estimate of its shoulder height (SH), body mass (BM), and sex; (iv) discuss the significance of the CM elephantid sample within the context of the Italian Middle Pleistocene mammal biochronology, biogeography, and paleoecology.
Institutional abbreviations.—IGF, Museum of Geology and Paleontology, University of Florence, Italy; MSTB-CM, Museum of Earth Sciences, University of Bari, Italy; MUST, University Museum of Earth Sciences, University of Rome, Italy.
Other abbreviations.—BM,
body mass; CM, Contrada Monticelli; FU, Faunal Unit; GL, greatest
length; MIS, Marine Isotope Stages; POC,
parieto-occipital crest;
SH, shoulder height. We follow standard convention in abbreviating tooth
families as I, C, P, and M, with upper and lower case letters referring
to upper and lower teeth, respectively.
Geological setting
Contrada Monticelli (Fig. 1; 40°49’50’’N, 17°10’27’’E) is located, near Putignano town in the Apulian region, in the southeastern part of the Italian peninsula. Unfortunately, the fossil site is no longer accessible, as it is now buried beneath a private residence, preventing any further stratigraphic or taphonomic analyses.
Fig. 1. Geographic location of the Contrada Monticelli fossiliferous site (Apulia, Southern Italy).
The fossiliferous deposit consisted of the infill of a karst cavity formed within the Calcare di Altamura Formation (Azzaroli 1967; Azzaroli et al. 1968), which is regionally dated to the late Turonian–Maastrichtian, Upper Cretaceous (Ciaranfi et al. 1988). The cavity, locally known as a grave (sinkhole), had an opening of approximately 4.5 × 5 m and was about 11 m deep. It was filled with red soil interbedded with several cinerite levels (Luperto Sinni and Colucci 1985).
The walls and bottom of the sinkhole were covered with reddish sands, mixed with fossilized bone remains. The sinkhole likely formed as a result of intensified karst processes along a fracture (or fault?) running NW-SE, parallel to the state road on which the sinkhole was located (Luperto Sinni and Colucci 1985).
Excavations of the deposit began in spring 1978, thanks to the involvement of the Superintendency of Antiquities of Bari and the Institute of Geology and Paleontology of the University of Bari, under the direction of Elena Luperto Sinni. Fossil remains were recovered from depths of approximately 3–11 m below the ground surface. However, a detailed stratigraphic framework of the infill deposits during the fossil recovery is lacking.
Following a pause during the winter of 1978, excavations resumed in the summers of 1979 and 1980 but were subsequently halted due to lack of funding (Luperto Sinni and Colucci 1985).
Over the decades, Contrada Monticelli (CM) has yielded abundant faunal remains of terrestrial mammals such as Bison cf. schoetensacki, Canis mosbachensis, Dama cf. roberti, Stephanorhinus hundsheimensis, furthermore, additional remains of equids, lagomorphs, and rodents were also found, though they are fragmentary and only identifiable at the family or order level (Luperto Sinni and Colucci 1985; Mazza and Varola 1999; Mecozzi et al. 2017; Stefanelli et al. 2021, 2024a, b). In addition to the taxa mentioned, an elephant referred to Elephas (=Palaeoloxodon) antiquus has been reported by Luperto Sinni and Colucci (1985). This Galerian faunal assemblage is biochronologically homogeneous and has been attributed to the Isernia Faunal Unit (lower Middle Pleistocene) by several recent studies (Stefanelli et al. 2021, 2024a, b).
In the Apulian hinterland, calcareous rocks experienced intense karst activity during the Middle to Late Pleistocene, forming a complex network of fissures (Selleri et al. 2003; Selleri 2007). These fissures were later infilled with sediments and occasionally preserved vertebrate fossils. Notable fossiliferous karst sites in Apulia include Pirro Nord, one of the most significant Early Pleistocene sites in Europe (e.g., Pavia et al. 2010; Arzarello et al. 2012; Berto et al. 2024; Duval et al. 2024), and the Maglie area, located in the Salentine Peninsula, which has yielded thousands of remains dated from the late Middle Pleistocene to the Early Holocene (Mecozzi et al. 2021 and references therein).
However, apart from Pirro Nord, most Apulian fossil deposits are attributed to the late Middle Pleistocene or more recent periods. Therefore, CM represents the only known deposit of early Middle Pleistocene age in the region and one of the few in southern Italy, alongside Isernia La Pineta, Loreto, and Notarchirico.
Material and methods
Material.—The elephantid material from Contrada Monticelli is represented by 32 specimens. These include three incomplete molars, four tusk fragments, and 25 skeletal bones (SOM 1: table 1, Supplementary Online Material available at http://app.pan.pl/SOM/app71-Stefanelli_etal_SOM.pdf). Further elephant skeletal material is embedded in a block of bone remains cemented with reddish sand (see 3D model in SOM 3). All the CM material is currently preserved at the Museum of Earth Sciences of the University of Bari Aldo Moro, Italy. The catalogued specimens are labelled with progressive numbers preceded by the acronym MSTB-CM (Museo di Scienze della Terra Bari-Contrada Monticelli).
Table 1. List of preserved elements Palaeoloxodon antiquus from Contrada Monticelli. L, left; R, right; ? uncertain assignment.
|
Anatomical portion |
Specimen |
|
tusk |
MSTB-CM 30559, 30560, 30668, QF59a |
|
tooth |
MSTB-CM 30531, 30709, QF59b, 30812(fragm.) |
|
vertebra |
MSTB-CM 30582, 30583 |
|
rib |
MSTB-CM 30564, 30569, 30572–30577, 30676, 30812 |
|
scapula |
MSTB-CM 30581, 30585(R) |
|
humerus |
MSTB-CM 30562(?), 30567(?), 30568(?), 30588 |
|
ulna |
MSTB-CM 30571(R) |
|
radius |
MSTB-CM 30566(R), 30587(?) |
|
femur |
MSTB-CM 30570, 30586(L) |
|
tibia |
MSTB-CM 30563(R) |
|
metapodial |
MSTB-CM 30584 |
Morphological and morphometric analysis.—Dental measurements were conducted following the protocols of Maglio (1973), whereas skeletal measurements were taken according to Göhlich (1998). All measurements were taken with a tape measure or sliding digital caliper. The original abbreviations used in Göhlich (1998) have been replaced by a number whose correspondence with the original abbreviations is shown in SOM 1: table 2. In the comparative analysis of the CM elephant material we used both published data and direct observation of osteological material, including mounted skeletons of Mammuthus meridionalis from Upper Valdarno, housed at the Museum of Geology and Paleontology of the University of Florence, and of Palaeoloxodon antiquus from Fonte Campanile, Viterbo, exhibited at the University Museum of Earth Sciences (MUST). Measurements taken on the CM specimens, and on the comparative material are reported in SOM 1: table 2. With the aim to verify the actual reliability of certain morphological traits in distinguishing between European Palaeoloxodon antiquus (the only mainland Europe full-sized representative of the genus in the Middle to Late Pleistocene) and mammoths (specifically the Quaternary species occurring in Europe), we compared these diagnostic characteristics, cited in the literature (see below), with skeletal material directly observed on specimens exhibited in the museums of Florence, Rome and Bari, as well as with material illustrated in published sources. To support this analysis, comparative tables were constructed, selecting the main morphological features of the humerus, ulna, femur, and tibia identified by Andrews and Cooper (1928), Maccagno (1962), Melentis (1963), Kroll (1991) and Davies (2002) as taxonomically significant at the generic level: these characters were compared and checked. The most important measurements have been plotted in scatter diagrams and analyzed through Principal Component Analysis (PCA) with PAST v. 4.13 (Hammer et al. 2001).
Table 2. Selected anatomical characters of the humerus, ulna, femur and tibia considered diagnostic for distinguishing between Palaeoloxodon and Mammuthus species, as described by previous authors and in this study. *observed in M. primigenius (Ferretti 2008)
|
Character number |
Reference |
Character |
P.
antiquus |
M. meridionalis M. primigenius (from references) |
P.
antiquus |
M.
meridionalis |
|
Humerus |
||||||
|
1 |
shape of articular head |
less spherical |
more spherical |
spherical or |
spherical or |
|
|
2 |
lateral groove, midway along the diaphysis |
present |
absent |
variably present |
absent* |
|
|
3 |
angle of the lateral epicondyle |
110–115° |
140–145° |
117–140° |
125–147° |
|
|
4 |
distal epiphysis |
relatively broad |
relatively narrow |
relatively broad |
relatively narrow |
|
|
5 |
trochlear furrow |
deep |
shallow |
shallow to deep |
shallow to deep |
|
|
6 |
trochlea width |
relatively broad |
relatively narrow |
relatively broad |
relatively narrow |
|
|
7 |
medial and lateral |
sharp |
rounded |
sharp |
rounded |
|
|
8 |
shape of trochlea |
hourglass-shaped |
cylindrical |
variable degree |
variable degree |
|
|
Ulna |
||||||
|
1 |
shape of trochlear notch (incisura semilunaris) |
saddle-shaped |
flat |
saddle-shaped |
flat |
|
|
2 |
olecranon |
larger and |
smaller and |
smaller and more |
more robust and dorso- caudally projected |
|
|
3 |
coronoid processes |
extended |
extended antero- |
variably extended transversely |
variably extended |
|
|
Femur |
||||||
|
1 |
head shape |
less spherical and more medially projected |
more spherical and less medially projected |
distinctly spherical and
moderately |
distinctly spherical and
moderately |
|
|
2 |
height of head relative to greater trochanter |
about the same height |
head of the femur rises more above the greater trochanter |
head of the femur rises more above the greater trochanter |
head of the femur rises more above the greater trochanter |
|
|
3 |
shape of |
straight diaphysis |
diaphysis with curved medial and lateral edges |
straight diaphysis |
straight diaphysis |
|
|
4 |
proportion of distal epiphysis with respect to bone length |
relatively wide |
relatively narrow |
relatively wide |
relatively narrow |
|
|
5 |
angle of trochlear groove |
ca. 145° |
ca. 130° |
ca. 130–145° |
ca. 130–145° |
|
|
6 |
intercondylar spacing |
distal condyles very close to each other |
distal condyles |
rather close |
rather close |
|
|
7 |
width of distal |
relatively narrow |
relatively broad |
relative breadth |
relative breadth |
|
|
8 |
width of caudal part |
relatively broader |
relatively narrower |
relative breadth |
relative breadth |
|
|
Tibia |
||||||
|
1 |
lateral proximal |
relatively wider antero-posteriorly |
relatively narrower antero-posteriorly |
shape of the lateral
articular surface does not differ between |
shape of the lateral
articular surface does not differ between |
|
Tusk dentine Schreger pattern.—A cross-section of a proximal tusk fragment from CM was performed with a circular saw in order to measure the so-called “Schreger angles”. The latter were measured using the ImageJ software. Comparisons were made with a tusk polished section of Mammuthus meridionalis (IGF 305) preserved at the Museum of Geology and Paleontology of the University of Florence (SOM 2: fig. 3) and with data taken from the literature.
Body size and sex determination.—To estimate shoulder height of the CM elephant, from single long bone, in our case the femur, the linear regression equation proposed by Larramendi (2016) was used in this work:
SH = X × 2.56,
where SH = skeletal shoulder height, X = femur length from caput (mm). Shoulder height, however, must also consider the part of flesh and integument that covers the skeleton of a living animal. Osborn (1942) added 6.67% for flesh, Roth (1990) and Christiansen (2004) added around 5%, and Larramendi (2016) added 5.5%. Following previous authors, 6% was added to skeletal shoulder heights to account for flesh, providing estimates of live shoulder height. In Lister and Stuart (2010) live shoulder heights are estimated at 5%, 6% and 7% above skeletal height. In this work, the live shoulder heights are estimated at 6% as in Erkek and Lister (2021). The shoulder height (both skeletal and with flesh) estimations were calculated based on the greatest length from caput (measurement 1) of the femur.
In this work, the formula recently proposed by Larramendi (2016) was used to estimate the body mass of P. antiquus from shoulder height:
BM = 3.63 × 10-4 × SH2.903,
where BM = body mass in kg and SH = shoulder height (with flesh) in cm.
Differences in body size between males and females are commonly found among many extant and extinct elephants. In this work, to determine the sex, the SH and BM of the CM specimen were compared with the analogous ones calculated for the specimens of P. antiquus described in the literature, for which the sex had been established.
For archiving and research purposes, the skeletal elements of the CM elephant were digitalized using a structured-light EinScan-Pro HD 3D scanner equipped with the Color Pack accessory to acquire both geometry and texture. Data acquisition was performed with the dedicated EXScan-Pro software in handheld mode, using a spatial resolution of 0.3 mm with a nominal accuracy of ±0.045 mm and a volumetric accuracy of 0.3 mm/m. With the same software the obtained point cloud was then fixed and the mesh with texture was built.
Systematic palaeontology
Class Mammalia Linnaeus, 1758
Order Proboscidea Illiger, 1811
Family Elephantidae Gray, 1821
Genus Palaeoloxodon Matsumoto, 1924
Type species: Palaeoloxodon naumanni (Makiyama, 1924), Sahama Formation (Late Pleistocene), Shizuoka (Japan)
Palaeoloxodon antiquus (Falconer & Cautely, 1847)
Figs. 2–4.
1823 Elephas priscus; Goldfuss 1823: 489, pl. 57: 1.
1847 Elephas antiquus; Falconer and Cautley 1847: 25–80, pl. 12d: 4, 5, pl. 14a: 1–13, pl. 14b.
1875 Elephas ausonius Forsyth Major, 1875; Verri 1886: 453.
1897 Elephas platyrhynchus; Graells 1897: 569, pl. 18: 1–11.
Material.—A partial skeleton (detailed list of preserved elements Table 1, see also SOM 1: table 1) from Contrada Monticelli, Italy, Pleistocene.
Description.—Preservation and minimum number of individuals: Both the isolated dental and skeletal elements and those still embedded into the cemented block are in a variable state of preservation. Most of the remains are fractured and/or incomplete and covered with a reddish patina. Some of the bones, however, are complete and in a relatively good state of preservation, allowing for a thorough anatomical description. For those elements embedded in the block only general observations could be done.
There is no evidence that suggest the presence of more than one individual in the CM elephantid material. Among the isolated remains, there are no duplicated elements and the molars and the size and developmental stage of the bones support their attribution to a single individual.
Tusks: Four tusk fragments are present in the CM sample; one is from near the proximal end (MSTB-CM QF59a; Fig. 2B), as indicated by a small pulp cavity visible at the center; two represent intermediate sections of the tusk (MSTB-CM 30559, 30668) and one derives from near the distal end of the tusk (MSTB-CM 30560; Fig. 2C). Despite the poor state of preservation and the partial coverage by the encrusting sediment (which did not allow for a reliable measurement of the circumference), only a weak curvature is noted, especially in the distal fragment MSTB-CM 30560. The proximal portion (MSTB-CM QF59a), shows a slightly elliptical cross-section, with diameters of approximately 138 and 120 mm. Overall, being fragments of different parts of the tusk (proximal, intermediate, distal), these can belong to one or a maximum of two tusks, and this is consistent with the presence at CM of a single individual.

Fig. 2. Dental remains of Palaeoloxodon antiquus (Falconer & Cautely, 1847) from Contrada Monticelli, southern Italy, lower Middle Pleistocene. A. MSTB-CM 30812, tooth fragment in occlusal view. B. MSTB-CM QF59a and b, proximal tusk fragment and M3 cemented together in lateral view. C. MSTB-CM 30560, distal tusk fragment in lateral view. Scale bars 50 mm.
Molars: Cheek tooth material is in poor condition. MSTB-CM QF59b (Fig. 2B) is interpreted as rather worn upper third molar (M3) with only a small posterior part missing. The occlusal surface of the molar is cemented with one of the tusk fragments (MSTB-CM QF59a), preventing the observation of the occlusal morphology. However, observing its lateral profile, towards the base of the crown, a lamellar frequency of 5 and a total of 14–15 lamellae can be counted. MSTB-CM 30531 consists of incomplete and poorly preserved and interpreted as rather worn lower third molar (m3) based on the roots that are noticeably curved backwards (see SOM 2: fig. 2). MSTB-CM 30812 (Fig. 2A), consists of a small tooth fragment that, however, has a clean occlusal surface. This molar fragment reveals the initial wear pattern of the lamellae, characterized by a distinctive dot-dash-dot configuration.
Vertebrae: MSTB-CM 30582, 30583, are two fragmentary vertebrae, MSTB-CM 30582 consists of the very large body of a thoracic vertebra.
Ribs: MSTB-CM 30564, 30569, 30572–30577, 30676 and 30812, represent either intermediate or proximal rib fragments, which are the most abundant bone element in the elephantid sample from CM. Their state of preservation prevents the identification of their position along the rib cage. In some cases, the rib fragments are attached to other unidentified bone fragments. Further fragmentary ribs were recognized on the surface of the cemented bone block (see SOM 2: fig. 1: 4, 5).
Scapulae: MSTB-CM 30581, highly abraded fragment and MSTB-CM 30585, the articular portion of the right scapula, characterized by a large elliptical glenoid cavity and the supraglenoid tubercle that extends ventrally lower than the articular surface. The articular portion of the left scapula MSTB-CM 30581 is embedded within the cemented bone block (see SOM 2: fig. 1: 2).
Humerus: The humerus is represented by the portion of the diaphysis (MSTB-CM 30562), two very fragmentary remains (MSTB-CM 30567, 30568) and a left distal fragment preserving part of the medial side of the diaphysis and the entire epiphysis (MSTB-CM 30588; Fig. 3A). It is not possible to determine with certainty whether the previous fragments belong to the same bone. In cranial view, the diaphysis of MSTB-CM 30588 appears rather broad and flattened, with a slightly arched medial profile. The humeral crest is pronounced and ends at approximately the same level as the epicondylar crest. The latter is slightly damaged both in its ascending portion, where it then joins the diaphysis, and in its descending lateral branch, where a substantial portion is missing. Above the trochlea is a large radial fossa that is not very deep and approximately elliptical in shape. The trochlea has a medial lip that is approximately the same height as the medial one, but wider. The trochlear groove is wide and shallow. The medial margin of the trochlea is slightly convex and vertical, while the lateral margin is flat and slightly inclined in a medial direction. The trochlea has a moderately cylindrical shape, with quite sharp edges. In caudal view, the olecranon fossa is high, wide, but shallow. On either side of it, the epicondyles are very different from each other, with the medial one being clearly more massive than the lateral one.
Ulna: MSTB-CM 30571 (Fig. 3B) is a complete ulna, although it is extensively fractured. Part of the olecranon tuberosity is missing, while a bone fragment (probably a rib) is attached to its distal part. In cranial view, the trochlear notch (incisura semilunaris) has a saddle-shaped basis and is also quite wide in a medio-lateral direction. The cranial surface of the diaphysis is slightly concave. The distal epiphysis does not expand much transversely, though it is heavily fractured. In medial view, the notable antero-posterior elongation of the proximal epiphysis is evident. The anconeal process is moderately curved forward and a deep fossa for articulation with the radius is evident.

Fig. 3. Palaeoloxodon antiquus (Falconer & Cautely, 1847) from Contrada Monticelli, southern Italy, lower Middle Pleistocene. A. MSTB-CM 30588, left humerus in cranial (A1) and caudal (A2) views. B. MSTB-CM 30571, right ulna and unnumbered rib fragment in cranial (B1) and caudal (B2) views. Scale bars 20 mm.
Radius: MSTB-CM 30566 consists of the proximal two thirds of the right radius, exhibiting a significant degree of fracturing and abrasion. The proximal epiphysis presents an articular surface for the humerus that is approximately triangular in proximal view, with rounded corners. In cranial view, the radial tuberosity and the coronoid process are quite prominent. Immediately distal to the proximal epiphysis, the diaphysis exhibits a strong torsion around its axis and has a distinctly curved profile.
Femora: Two femora are present in the CM elephantid sample. MSTB-CM 30570 consists of an articular head; MSTB-CM 30586 (Fig. 4) is a nearly complete left femur. In cranial view, the articular head is large, spherical and facing upwards, with a short neck. In both bones there are no traces of a fovea capitis. The portion of the greater trochanter is the only slightly damaged part of the bone. Proceeding distally, the diaphysis narrows and, in its central part, has a straight columnar shape, then widens again near the distal epiphysis. The patellar surface is rather small and hourglass shaped in distal view. It is oriented towards the medial side and its lateral lip is slightly more pronounced than the medial one. These two parts are separated by a rather deep groove. The medial epicondyle is more prominent than the lateral one, which is flatter. In caudal view, the trochanteric fossa is elliptical in shape, rather narrow and elongated downwards, but not very deep. The intertrochanteric crest is poorly marked. The condyles, although partially covered by a large fragment of another strongly cemented bone, are large and rounded, close to each other and with the lateral one noticeably more voluminous than the medial one. The intercondylar fossa is narrow and shallow. The right femur is embedded within the cemented bone block from which only the side of the proximal epiphysis emerges.

Fig. 4. Palaeoloxodon antiquus (Falconer & Cautely, 1847) from Contrada Monticelli, southern Italy, lower Middle Pleistocene. MSTB-CM 30586, left femur and MSTB-CM 30587, left radius fragment in cranial (A1) and caudal (A2) views. Scale bar 20 mm.
Tibia: MSTB-CM 30563 is a right tibia missing the proximal end. In cranial view, there is a rather pronounced tibial crest and the medial malleolus is prominent. In caudal view, a broad distal articular surface (tibial cochlea) with a pronounced concavity occupies the entire distal end.
Stratigraphic and geographic range.—Lower Middle Pleistocene–Upper Pleistocene; Europe and Western Asia.
Results
Tusks and molars.—Although fragmentary, the tusk remains from CM lack the pronounced, often twisted curvature characteristic of mammoths. Instead, they display a pattern characteristic of P. antiquus. Consistently, the molar fragment MSTB-CM 30812 shows the typical dot-dash-dot wear pattern of Palaeoloxodon, while Mammuthus molars are characterized by either a dash-dot-dash wear pattern or a row of uniformly sized enamel rings (Albayrak and Lister 2012).
Appendicular skeleton.—In this section we examine the various morphological characters of the limb bones considered in the literature to distinguish between Mammuthus (European Pleistocene species) and Palaeoloxodon antiquus. For each character (Table 2) we compare descriptions provided by previous authors with our own direct observation of several elephant skeletons (see Material and methods). We then assess how each character appears in the CM specimens.
Humerus (Table
2): (i) Melentis (1963) reported that Palae-oloxodon
has a less spherical articular head than Mammuthus.
According to our direct observations, however, there are
no evident differences in the degree of curvature of the articular head
between Palaeoloxodon and Mammuthus.
The shape of the humeral articular head is, in fact, much more flattened
than that of the femur, so much so that osteological measurement
protocols (e.g., Göhlich 1998) only include
measurements of the diameters of the femoral articular head. In the
humerus, the anteroposterior and transverse diameters of the entire
proximal epiphysis are measured (see measurements
4 and 5 of the humerus in SOM 1: table 4). In the CM specimen the articular
head is not observable because the proximal part of the humerus is
missing.
(ii) According to Kroll (1991) and Davies (2002), Palaeoloxodon has a triangular-shaped groove along the lateral side of the diaphysis (deltoid fossa), while this feature is absent in Mammuthus. Our comparative study demonstrates that a deltoid fossa may be either present or absent in both Palaeoloxodon and Mammuthus, as previously noted by Ferretti (2008). In the CM specimen the presence of groove midway along the diaphysis is not observable because part of the diaphysis is missing.
(iii) Melentis (1963) noted that the angle of the lateral epicondyle could vary between 110–115° for Palaeoloxodon and 140–145° for Mammuthus. According to our direct observations, the amplitude of the angle formed by the lateral epicondyle is not particularly different between Palaeoloxodon and Mammuthus (see Table 2: humerus character 3). In the CM specimen, the part relating to the lateral epicondyle was slightly damaged, therefore it was not possible to make an adequate estimate of the angle.
(iv) According to Maccagno (1962) there are differences in the proportion of the distal epiphysis between the two genera, with Palaeoloxodon having a broader epiphysis than Mammuthus. Although the number of M. meridionalis specimens directly examined by us is rather low, the data seem to support Maccagno’s (1962) observation: the width of the distal epiphysis, relative to the length of the humerus, tends to be greater in Palaeoloxodon antiquus. In the CM individual the distal epiphysis is notably wide, with a transversal width among the highest recorded (see SOM 1: table 4).
(v) Melentis (1963) reports that the trochlear articular surface (distal articular surface) is characterized by a deeper central groove in Palaeoloxodon than in Mammuthus. According to our direct observations, the depth of the trochlear groove is variable in both Palaeoloxodon and Mammuthus with no consistent difference between the two genera (SOM 2: fig. 4). Regarding the depth, the CM individual has a shallow trochlear groove (Fig. 3A1).
(vi) According to Melentis (1963), the trochlear articular surface is relatively wider in Palaeoloxodon and narrower in Mammuthus. Based on our direct observation, the trochlea of Palaeoloxodon is wider than that of Mammuthus (see measurement 10 of the humerus in SOM 1: table 4). The CM individual has a particularly wide trochlea (see measurement 10 in SOM 1: tables 2, 4).
(vii) Melentis (1963) observed that the medial and lateral edges of the trochlear articular surface are sharp in Palaeoloxodon, while in Mammuthus are blunter. From observations made directly on the comparative material, the edges of the trochlea tend to be indeed sharper in Palaeoloxodon than in Mammuthus (see SOM 2: fig. 4). In the CM specimen, the edges of the trochlea are quite sharp (see Fig. 3A1).
(viii) Andrews and Cooper (1928) noted that Mammuthus has a more cylindrical trochlea than Palaeoloxodon, which, according to them, possesses medially and laterally enlarged condyles, producing an hourglass-like shape. In our comparative sample, there do not seem to be particular differences between Palaeoloxodon and Mammuthus regarding the degree of cylindricity of the trochlea in cranial view (e.g., for P. antiquus see Maccagno 1962: pl. 12: 2b). In the CM specimen, the trochlea is cylindrical (see Fig. 3A1).
Ulna (Table 2): (i) In Davies (2002), Palaeoloxodon is described as possessing a saddle-shaped trochlear notch, while this is flat in Mammuthus. According to our direct observations, the trochlear notch is more pronounced in Palaeoloxodon and, in general, the articular surface with the trochlea of the humerus has a wavy saddle-like shape. This surface in Mammuthus is more flattened. In the CM specimen the trochlear notch is saddle-shaped (see Fig. 3B1).
(ii) According to Davies (2002), the olecranon in Palaeoloxodon is large and overhanging, while in Mammuthus it is smaller and more upright. Direct observations have shown that in Palaeoloxodon the olecranon is smaller and more elongated backwards, while in Mammuthus it is more robust and projected dorso-caudally. Unfortunately, the olecranon part is damaged in the CM specimen, therefore it is not possible to evaluate this character.
(iii) Maccagno (1962) reported that the two articular processes of the ulna for the humerus (processus coronoideus lateralis and processus coronoideus medialis) were more extended transversely in Palaeoloxodon and more extended antero-posteriorly in Mammuthus. According to our direct observation, however, no significant differences between Mammuthus and Palaeoloxodon were found in the transverse and antero-posterior development of the articular surface for the humerus. In the ulna of the CM specimen, the proximal articular processes for the humerus are relatively broad and antero-posteriorly short (see Fig. 3B).
Femur (Table 2): (i) According to Melentis (1963), the articular head of the femur is less spherical and more medially projected in Palaeoloxodon than in Mammuthus. In directly observed specimens, there does not appear to be any particular differences in the degree of sphericity and medial projection of the articular head between Mammuthus and Palaeoloxodon (see SOM 2: fig. 5). The CM specimen has a spherical and moderately medially projected femoral head (Fig. 4).
(ii) Andrews and Cooper (1928) observed that the head of the femur and the greater trochanter are at about the same height in Palaeoloxodon, while the head of the femur rises more above the greater trochanter in Mammuthus. We observed in our comparative sample that this trait is quite variable both in Mammuthus and Palaeoloxodon, with the femoral head ranging from slightly to markedly higher than the greater trochanter (see SOM 2: fig. 5). In the CM specimen, this trait cannot be assessed as a large portion of the greater trochanter is missing. However, the articular head appears significantly elevated.
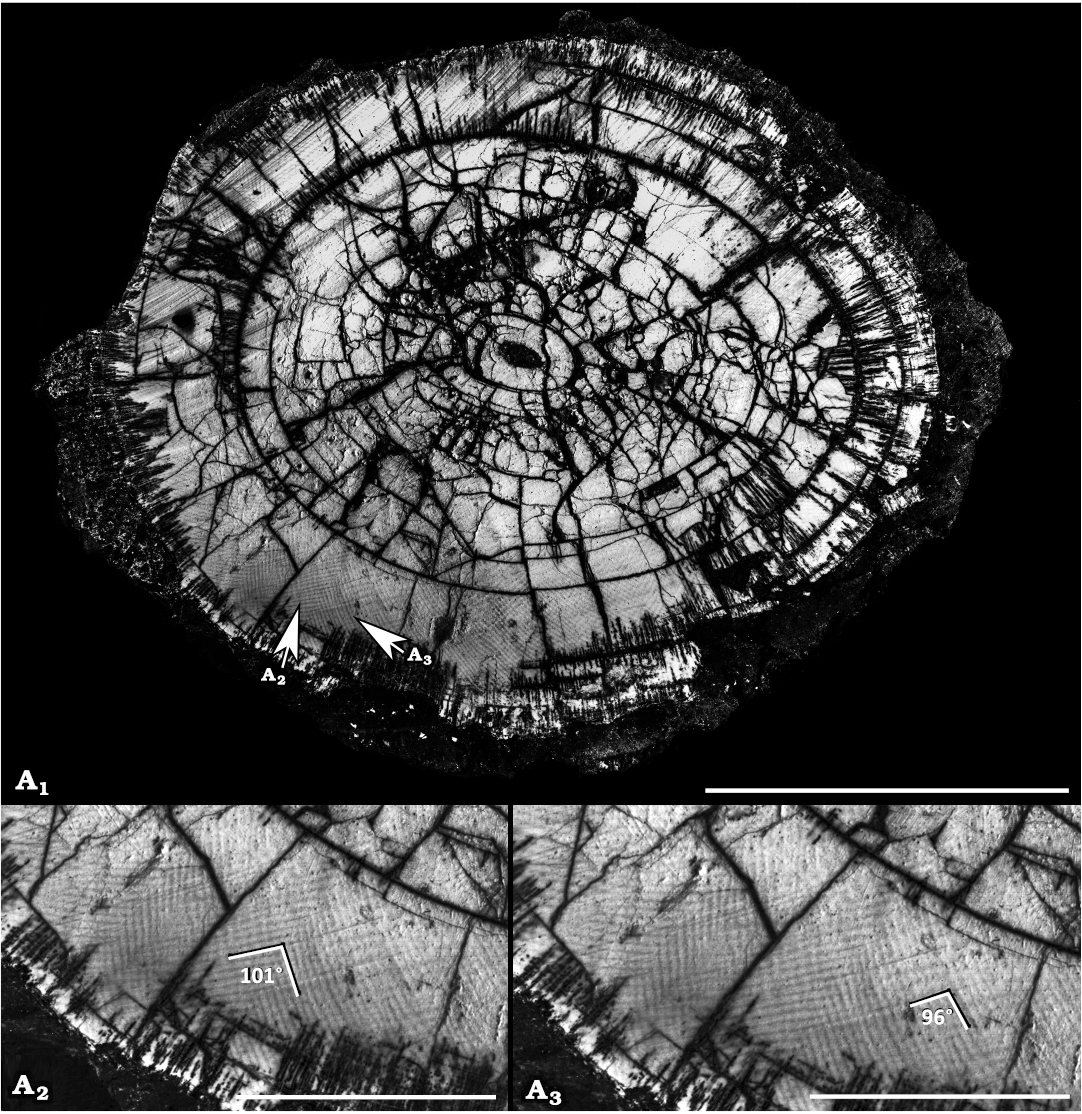
Fig. 5. Transverse section of the proximal portion of the tusk of Palaeoloxodon antiquus (Falconer & Cautely, 1847) from Contrada Monticelli, southern Italy, lower Middle Pleistocene. MSTB-CM QF59a, photo with enhanced contrast (A1), Schreger angles in the outer portion of the tusk section vary from about 101° (A2) to about 96° (A3). Scale bar A1 50 mm, A2, A3 10 mm.
(iii) Melentis (1963) and Davies (2002) describe the diaphysis of the femur of Palaeoloxodon in frontal view as possessing a straight lateral profile, while that of Mammuthus would be characterized by curved medial and lateral edges. From our observation, there does not appear to be significant differences in the shape of the diaphysis and the degree of curvature of its medial and lateral edges between Mammuthus and Palaeoloxodon (see SOM 2: fig. 5). The diaphysis of the CM specimen is straight in anterior view (Fig. 4).
(iv) According to Maccagno (1962), the distal epiphysis of the femur is relatively broader (transversally) in Palaeoloxodon and narrower in Mammuthus. Although our M. meridionalis sample is limited, it supports Maccagno’s (1962) conclusion that the femur distal width, relative to greatest length of the bone, is generally greater in P. antiquus than in Mammuthus. In the CM specimen the distal epiphysis is rather broad (see SOM 1: table 4).
(v) According to Melentis (1963), the angle of the trochlear groove is larger in Palaeoloxodon (ca. 145°) than in Mammuthus (ca. 130°). However, according to our direct observation, the amplitude of the trochlear groove angles shows a certain variability, with values for Palaeoloxodon and Mammuthus overlapping in the range of ca. 130–145° (see Table 2: femur character 3). The CM specimen displays a trochlear groove with an angle of approximately 130°.
(vi) Davies (2002) reported that the distal condyles, in posterior view, are very close together in Palaeoloxodon, whereas they are more widely spaced in Mammuthus. Among the directly observed specimens, there do not appear to be significant differences in the degree of separation of the condyles between the two genera (see SOM 2: fig. 5). In the CM specimen the distal condyles are positioned very close to each other (Fig. 4A2).
(vii) Melentis (1963)
reported that the distal articular surface for the tibia (seen in distal
view) is relatively narrow transversally in Palaeoloxodon
and broad in Mammuthus. According to
our direct observation, there do not appear to
be any significant differences in the shape of the articular
surface for the tibia between Mammuthus and
Palaeoloxodon (see measurement 12 of the
femur in SOM 1: table 3).
In the CM specimen, the breadth of the articular surface for the tibia
falls between the minimum and maximum values observed in the comparative
sample.
(viii) Melentis (1963) states that the caudal part of the lateral condyle is relatively broader (transversally) in Palaeoloxodon and narrower in Mammuthus. In directly observed specimens, the caudal part of the lateral condyle does not show particular differences in width between Palaeoloxodon and Mammuthus (see SOM 2: fig. 5). Like the other Elephantidae examined, the CM individual also appears to have a rather broad caudal part of the lateral condyle.
In addition to the characters discussed above, we observed that the studied femora of Mammuthus have a deeper trochanteric fossa than those of Palaeoloxodon. Indeed, this appear to be the only character that distinguishes the observed individuals of M. meridionalis from P. antiquus. The CM specimen exhibits a shallow but rather wide trochanteric fossa (see SOM 2: fig. 5).
Tibia (Table 2): (i) According to Andrews and Cooper (1928), the tibia of Palaeoloxodon differs from that of Mammuthus in the relative position of the lateral proximal articular surface for the femur (condylus lateralis), which is much lower than the medial one and in being wider antero-posteriorly. In the specimens we examined, however, Palaeoloxodon and Mammuthus seem not to differ in these traits. In the CM individual, this character is not observable as the entire proximal extremity is missing.
Schreger pattern analyses of tusk dentine.—In MSTB-CM QF59a the Schreger angles ranges 95–110° (Fig. 5). These values are higher than that typically observed in Mammuthus, while are consistent with the observed range in Palaeoloxodon (Espinoza and Mann 1993; Palombo and Villa 2001; Trapani and Fisher 2003). Schreger angles measured on the tusk section of M. meridionalis (IGF 305) ranged from 75° to 90°, consistent with the reported range in Mammuthus (Espinoza and Mann 1993; Palombo and Villa 2001; Trapani and Fisher 2003).
Morphometric analysis.—The
state of preservation of the molars from CM and their advanced stage of
wear does not allow determining the original total number of plates and
crown heigh, two important parameters in species identification. The
most complete molar, MSTB-CM QF59b, presents however a rather
narrow crown and a lamellar frequency which are consistent
with and attribution to P. antiquus.
Regarding the postcranial skeleton, our morphometric analysis did not
reveal significant differences in size and robustness between the
studied M. meridionalis and
P. antiquus specimens. As the femur is the most complete long
bone in the CM elephantid sample, we conducted a PCA
on our femur sample, based on five measurements (measurements 1, 6, 7,
10, and 11;
see SOM
1: table 4). PC1 accounts for most of the sample total variance (96.32%)
and is highly correlated with variable 1 (GL) and therefore with overall
size. Accordingly, a scatterplot of the first two
principal components (Fig, 6) shows a clear separation of
male and female specimens along PC1. In contrast, the ranges of
variation of the two species largely overlap as the analysis revealed no
clear distinction between the femora of P.
antiquus and M. meridionalis (Fig.
6). None of the other PCs improved species discrimination.
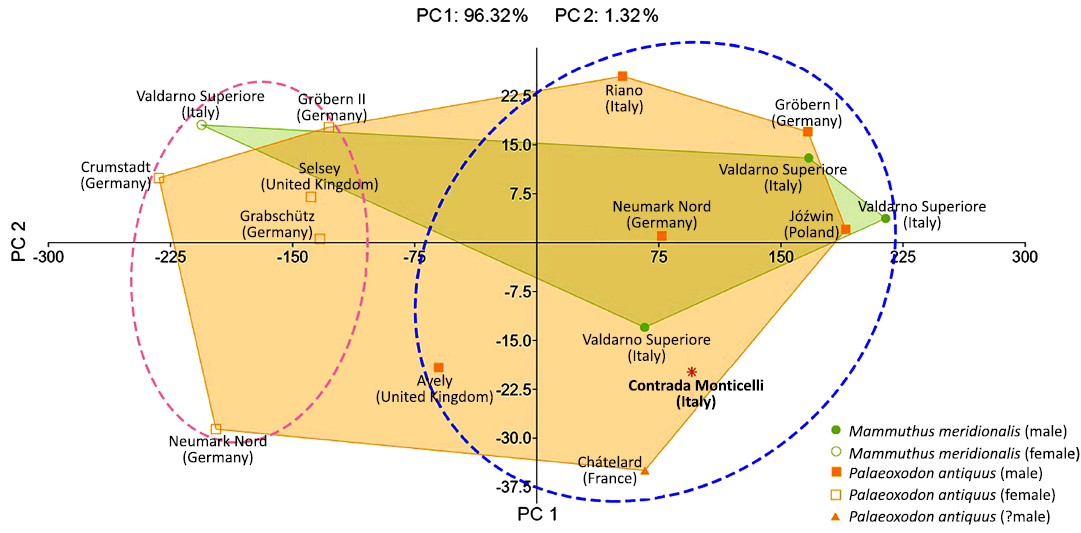
Fig. 6. 2D scatterplot of PC1 and PC2 from the Principal Component Analysis of femur variables 1, 6, 7, 10, and 11 in the studied sample of Palaeoloxodon antiquus and Mammuthus meridionalis. The analysis did not separate the two taxa. PC1, which primarily reflects size variation, distinguishes females (pink dashed oval) from males (blue dashed oval) individuals. Original measurements are reported in SOM 1: table 2.
Shoulder height, body mass, and sex.—To calculate the SH, the equation of Larramendi (2016) was applied both to the Contrada Monticelli individual and to the other comparison specimens of P. antiquus (see SOM 1: table 5). The results obtained showed a flesh SH for the CM individual of 3636 mm, a height comparable to that of medium-high sized male specimens (Fig. 7).
BM was estimated using the equation of Larramendi (2016) applied both to the individual from CM and to the other specimens of P. antiquus and M. meridionalis in the comparative sample (see SOM 1: table 5). The results obtained showed a BM for the CM individual of 9881 kg, and a shoulder height with flesh of 3.6 m, comparable to that of medium-large male P. antiquus specimens (Fig. 7).
The Upnor specimen was not included in the comparison, as only the proximal part of the femur is preserved, the rest is restored and its total length (1545 mm, according to Andrews and Cooper 1928) is most likely exaggerated (Asier Larramendi personal communication, 2025).
A reliable method for estimating the sex of an elephant skeleton is based on pelvis proportions (Lister 1996). In the absence of this skeletal element, as is the case for the CM elephant, body size, bone robustness, and tusk dimensions can be used as a proxy to estimate the sex of the animal, as elephants display a marked sexual dimorphism in these traits. Comparing the SH and BM estimates obtained for the CM elephant, based on linear measurements of the femur (MSTB-CM 30586), the CM elephant appears to belong to a large adult male, although its tusks are relatively slender. A principal component analysis (PCA) based on five femoral measurements did not clearly separate P. antiquus and M. meridionalis, which formed two largely overlapping clusters (Fig. 6). On the other hand, male and female individuals are separated along the PC1 axis, which correlates primarily with measurement 1 (femur GL), and thus reflects overall size. The CM specimen fell well within the male P. antiquus cluster.
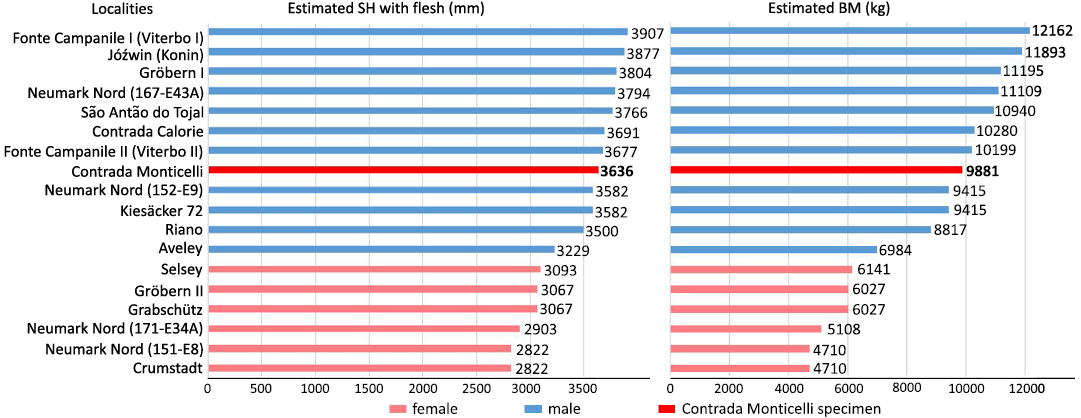
Fig. 7. Bar-charts comparing the estimated shoulder height with flesh (SH, in mm, following Larramendi 2016) and the estimated body mass (BM, in kg, following Larramendi 2016) of the Palaeoloxodon antiquus specimen from Contrada Monticelli with that of 17 other skeletons of P. antiquus. Data and relative references are reported in SOM 1: table 5.
Discussion
Taxonomy, sex, and age of the Contrada Monticelli elephant.—The CM elephantid was referred to as Elephas (= Palaeoloxodon) antiquus by Luperto Sinni and Colucci (1985), the most common elephant species in the Middle Pleistocene of central and southern Italy, although they did not provide supporting evidence for their taxonomic attribution. Our morphological and morphometric analysis of the entire CM sample allows us to confirm this preliminary attribution to Palaeoloxodon antiquus, based on dental morphology. In particular, the occlusal surface of the molar MSTB-CM 30812 (Fig. 2A), shows the dot-dash-dot initial wear pattern typical of Palaeoloxodon, and different from those usually observed in Mammuthus. The M3 (MSTB-CM QF59b) possesses a relatively narrow crown (SOM 1: table 3), compatible with the known range of variability observed in Italian samples of P. antiquus (Palombo and Ferretti 2005).
Based on the preserved fragments, the tusks of the CM elephant appear to have been only slightly curved, resembling the tusk morphology of Palaeoloxodon antiquus and contrasting with the spirally curved tusks of mammoths. Consistently, the dentine of the CM tusk shows Schreger angles ranging from 95° to 110° (Fig. 5), which fall within the observed range of Palaeoloxodon, and of extant Loxodonta and Elephas. In contrast, Mammuthus possesses distinctly narrower angles (Palombo and Villa 2001), further confirming the utility of this trait in discriminating among elephantid taxa (Athanassious 2011; Katona et al. 2012; Virág 2014; Kevrekidis and Mol 2016).
The arrival of elephants of the genus Palaeoloxodon from Africa into Eurasia occurred at the end of the Early Pleistocene (approximately 0.9–0.8 Ma, MIS 21–19; Palombo and Ferretti 2005; Lister 2016; Mecozzi et al. 2025b). From a dental perspective, these forms are similar to the African species P. recki. In particular, the total number of lamellae and the degree of hypsodonty show substantial stasis throughout the time span in which the genus existed in Eurasia. In contrast, the skull shows morphological evolution in the Eurasian forms, especially evident in the development of a peculiar structure of this genus known as the parieto-occipital crest (POC; Larramendi et al. 2020).
European remains of Palaeoloxodon are commonly attributed to a single species, P. antiquus. The earliest finds of this species come from northern Italy (Slivia, MIS 21) and England (Pakefield, MIS 17), which attest to its wide latitudinal distribution in Europe during the 0.9–0.7 Ma time interval (Stuart and Lister 2001; Palombo and Ferretti 2005). It is worth noting that the skull of P. antiquus is only known starting from MIS 12 (e.g., Marathousa 1, Greece; Konidaris et al. 2018; Athanassiou 2022), at which point the development of the POC is already well pronounced.
In the Middle East (a likely dispersal route between Africa and Eurasia followed by Palaeoloxodon) and Central Asia, elephant skulls attributable to this genus have been found, dated between 0.8 and 0.7 Ma, which, however, do not show development of a POC, and have been assigned to different species (P. recki and P. turkmenicus, respectively; Dubrovo 1960; Saegusa and Gilbert 2008). In the absence of cranial remains showing the dorsal portion, it is currently not possible to determine the degree of development of the POC in pre-MIS 12 European Palaeoloxodon populations, which includes the Italian material from CM, Isernia La Pineta, and Notarchirico. As a matter of fact, although these earlier specimens are indistinguishable from late Middle and Late Pleistocene P. antiquus based on dental and postcranial features, they may nonetheless represent a distinct taxon. For this reason, sites dated between 0.9 and 0.5 Ma, such as CM, are of particular importance for our understanding of the diversity of early European palaeoloxodont elephants.
Our estimates of shoulder height (SH) and body mass (BM) for the CM elephant are close to the averages calculated for adult male Palaeoloxodon antiquus from various European localities (Erkek and Lister 2021), suggesting it was a medium-sized male individual (M3 in use), with relatively slender tusks. The specimens studied by Erkek and Lister (2021) originate from sites younger than the Isernia FU. No size estimates currently exist for P. antiquus specimens older than MIS 12, except for the nearly complete skeletons from Contrada Calorie (Palombo et al. 2024) and CM. Interestingly, the SH and BM estimates reported for the Contrada Calorie elephant are very similar to those proposed here for the CM elephant (see Fig. 7). At present, due to the scarcity of well-preserved early P. antiquus skeletal material, it is not possible to determine whether the extreme body sizes observed at younger sites, such as Neumark Nord 1 (MIS 7/5; Larramendi et al. 2017), already characterized the earliest European populations of P. antiquus.
On the other hand, the available evidence suggests that P. antiquus did not exhibit a latitudinal gradient in body size across its European range (Larramendi et al. 2017; Erkek and Lister 2021; Palombo et al. 2024).
Diagnostic value of the post-cranial skeleton.—All Elephantimorpha (the clade that includes derived proboscideans with horizontal tooth progression) exhibit a rather stereotypical skeletal structure, likely linked to their graviportal adaptations for supporting their very large body size. Only a few minor postcranial characters have been identified that appear to differentiate Mammutidae, Gomphotheriidae, and Elephantidae (Tassy 1985; Ferretti 2010), while differences within families are more elusive and mostly concern bone proportions and robusticity (Reggiani 2001; Larramendi 2016). These, however, are difficult to assess with small samples, considering that postcranial bones change in proportion during growth, and that males are generally more robust than females. In fact, the only consistently marked difference between elephantid genera is the well-known variation in the dorsal silhouette of the vertebral column, which is primarily due to differences in the length and inclination of the vertebral spinous processes (Shoshani 1996). However, this complex feature can only be assessed in sufficiently complete and well-preserved vertebral columns, a rather rare occurrence.
Our comparative anatomical analysis of four long bones (humerus, ulna, femur, and tibia) previously considered by several authors to possess diagnostic traits distinguishing Palaeoloxodon from Mammuthus, revealed that most of these traits are subject to extensive individual variation, or that traits regarded as diagnostic for one genus were also observed in the other. These therefore cannot be used to discriminate between the two genera.
The humerus, ulna, and femur do appear to show some consistent differences between the two genera; however, these need to be tested on a larger sample to confirm their diagnostic value. As first observed by Maccagno (1962) and Melentis (1963), Palaeoloxodon is characterized by a proportionally wider distal humerus compared with Mammuthus, as indicated by the greater widths of both the distal epiphysis and the distal articular surface, relative to the bone length. Consistently, the CM humerus is characterized by a particularly broad distal epiphysis. We were also able to confirm Melentis’ (1963) observation that the distal articular surface of the humerus of Palaeoloxodon possesses distinctly sharper edges with respect to Mammuthus.
Regarding the ulna, we confirmed Davies’ (2002) observation that in Palaeoloxodon, the trochlear notch (incisura semilunaris) is clearly saddle-shaped, whereas in Mammuthus it is relatively flat. Davies (2002) also highlighted differences in the robusticity and orientation of the olecranon between the two genera. We observed consistent differences in the development of the olecranon as well, although they differ from the traits described by Davies (2002). Specifically, in Mammuthus, the olecranon is more robust and projects in a dorso-caudal direction, while in Palaeoloxodon it appears less pronounced and positioned lower. Additionally, the distance from the anconeal process to the posterior edge of the olecranon is greater in Palaeoloxodon than in Mammuthus.
Regarding the femur, Maccagno’s (1962) observation on the proportions of the distal epiphysis is supported: Palaeoloxodon exhibits a larger relative distal width than Mammuthus, consistent with pattern observed in the hume-rus. Furthermore, another feature that seems to distinguish M. meridionalis from P. antiquus, is the different depth of the trochanteric fossa. The Mammuthus femora examined have a deeper trochanteric fossa than those of Palaeoloxodon.
The morphometric analyses did not reveal significant size differences between the Mammuthus and Palaeoloxodon specimens studied.
Despite this, significant individual size differences were observed among individuals of the same genus, but these are attributable to sexual dimorphism. As is well known, both extant and extinct elephants exhibit pronounced sexual dimorphism, particularly in body size, with males being considerably larger than females (Haynes 1991; Shoshani 1996; Sukumar 2003). Healthy, fully grown males of E. maximus are approximately 15% taller and weigh 48% more than females, whereas males of L. africana are 23% taller and weigh twice as much (100%) as females (Larramendi 2016). Larramendi et al. (2017) found that sex differences in size were even greater in P. antiquus from Neumark Nord 1, with fully grown males being approximately 33% taller and weighing approximately 136% more than females, suggesting greater sexual dimorphism than in extant elephants. Consistently, several authors have used the size of the postcranial elements to distinguish sexes of extinct elephantids (e.g., Kroll 1991; Averianov 1996; Tsoukala and Lister 1998; Palombo and Villa 2003).
Biochronological, paleobiogeographical, and palaeoecological remarks.—Faunal assemblages from southern Italy, such as those from Isernia La Pineta (MIS 15) and the upper levels of Notarchirico (MIS 16), dated to approximately 0.58 and between 0.66 and 0.61 Ma, respectively, and referred to the Isernia FU, exhibit a taxonomic composition similar to that of CM (Sala 2006; Breda et al. 2015; Peretto et al. 2015; Mecozzi et al. 2024b). These assemblages are characterized by the occurrence of Bison cf. schoetensacki and Stephanorhinus hundsheimensis, and the primitive fallow deer Dama roberti, whose known fossil record spans from 0.7 to 0.5 Ma (Breda and Lister 2013; Mecozzi et al. 2024a). During this interval, Palaeoloxodon antiquus had already achieved a wide geographical distribution across Europe, ranging from the British Isles to southern Italy, and from the Iberian Peninsula to Greece (Palombo et al. 2010). Despite this wide range, complete or partial skeletons of this species are extremely rare in Europe prior to MIS 12–11, and, as stated above, no complete skulls, preserving the dorsal part, are known. The CM specimen therefore represents one of the few straight-tusked elephant partial skeletons known from this time interval in Europe, along with those from Contrada Calorie (Palombo et al. 2024) and Notarchirico (Mecozzi et al. 2024b).
In the Italian Peninsula, the straight-tusked elephant reached as far south as southern Calabria during the Middle Pleistocene and eventually colonized Sicily (Palombo and Ferretti 2005).
P. antiquus represents the most frequently recorded elephant species in the Middle Pleistocene of Italy, whereas findings of Mammuthus are extremely rare. No Italian sites are currently known to feature both genera in clear stratigraphic association, with the sole exception of the Ponte Galeria Formation near Rome. The classic “Ponte Galeria fauna” (Ponte Galeria 2) originates from the sand and gravel beach deposits underlying the Venerupis senescens clays, dated to approximately 0.7 Ma (Milli 1997). The faunal list includes, among others, Mammuthus trogontherii and Palaeoloxodon antiquus (Milli and Palombo 2005). Due to the scarcity of remains and the impossibility of verifying the exact stratigraphic position of specimens belonging to historical collections, it cannot be excluded that, actually, the two species may originate from different depositional layers (Petronio and Sardella 1999). This would be consistent with the hypothesis that P. antiquus and M. trogontherii had distinct ecological requirements and thus occupied different niches.
In Italy, Mammuthus trogontherii is considered exceptionally rare and is only reported as far south as Lazio and Abruzzo (Palombo and Ferretti 2005). During the coldest phases of the Late Pleistocene, M. primigenius expanded into the Italian Peninsula, reaching its southernmost range at Cardamone, located at the southern tip of Apulia (Rustioni et al. 2003).
Mesowear and stable isotope analyses from other European contexts indicate that M. trogontherii maintained a predominantly grazing diet, while P. antiquus displayed a broader range of dietary strategies, often classified as a mixed-feeder with a browsing tendency (Saarinen and Lister 2016; Tsakalidis et al. 2025). This degree of dietary plasticity may have allowed P. antiquus to exploit a wider array of environments and vegetational structures. Isotope data obtained from the dental enamel of P. antiquus from Europe suggest in fact that these animals lived in different environmental and climatic conditions (e.g., Grube et al. 2010; Briatico and Bocherens 2023; Roditi et al. 2024). For example, the straight-tusked elephant sample from Steinheim an der Murr and Mauer (Germany) indicated cooler and more humid conditions compared to that of other European samples (Grube et al. 2010). As a general trend, stable isotopic data indicate a transition from a wooded landscape and humid climate in Europe during the mid-Middle Pleistocene (MIS 15–11), to more open and arid environments at the end of Middle Pleistocene (MIS 7) (Mecozzi et al. 2025a). Due to the poor preservation of dental remains from the CM locality, mesowear analysis could not be performed. Nevertheless, the overall composition of the faunal assemblage from CM suggests the presence of mild climatic conditions and a heterogeneous landscape, characterized by open habitats interspersed with areas of tree cover. This environmental setting would have supported mixed-feeding strategies and reduced potential trophic competition among large herbivores.
Conclusions
The complete early Middle Pleistocene elephant sample from Contrada Monticelli has been studied here for the first time. The material consists of 32 dental and skeletal elements belonging to a single adult (M3 in use) male individual. Molar traits and Schreger pattern of the tusk dentine allowed us to confidently refer the specimen to Palaeoloxodon antiquus, the straight-tusked elephant. Diagnostic features of the humerus, ulna, and femur of the CM elephant are consistent with such an attribution. The CM elephant had an estimated shoulder height with flesh of approximately 3.6 m and a body mass of nearly 10 t, corresponding to a medium-sized male P. antiquus individual (Larramendi 2016; Erkek and Lister 2021), comparable to the specimen from Contrada Calorie (Palombo et al. 2024).
Our morphological comparison of the limb bones of late Early Pleistocene Mammuthus meridionalis from Upper Valdarno (Tasso FU) and various Middle Pleistocene Palaeoloxodon antiquus from Italian sites indicates that most of the morphological characters previously reported as diagnostic between Mammuthus and Palaeoloxodon are not reliable, when intraspecific variability is taken into consideration. However, the two genera appear to differ in specific traits of the distal articular surface of the humerus and femur, and the proximal epiphysis of the ulna. These differences merit further investigation by using a larger comparative sample that should also include other Mammuthus species. The morphometric analysis we conducted did not reveal any differences between Mammuthus and Palaeoloxodon in terms of the size and proportions of the long limb bones. On the other hand, the size of the long bones analyzed (humerus, ulna, femur, and tibia), particularly their greatest length, clearly distinguishes adult male and female individuals in both genera. The elephant from Contrada Monticelli represents one of the few European skeletons of straight-tusked elephant known from the 0.8–0.5 Ma interval, corresponding to the early dispersal of the genus in Eurasia. Although it provides no information on the cranial morphology of these early European palaeoloxodontine populations, particularly regarding the degree of development of the POC, the CM specimen nonetheless contributes to our understanding of the biochronological, paleoecological, and paleobiogeographical context of Palaeoloxodon early evolution in Europe. The presence of the straight-tusked at CM, along with other mixed-feeder herbivores such as Bison cf. schoetensacki, Dama cf. roberti, and Stephanorhinus hundsheimensis, suggests a temperate climate and a landscape with substantial woodland components as well as more open areas, as indicates the occurrence of Equus sp. in the CM assemblage. In this varied environment, different herbivores likely took advantage of the diverse vegetation, using their dietary flexibility to minimize competition.
Acknowledgements
The authors thank the Museo di Scienze della Terra and Dipartimento di Scienze della Terra e Geoambinetali (University of Bari Aldo Moro, Italy) and Nicola Venisti for the facilitation to access the Museum and to study the fossil sample from Contrada Monticelli, Giuseppe Di Prizio (University of Bari Aldo Moro, Italy) for his help in obtaining the tusk section and Nicola Antonio Morano (University of Bari, Aldo Moro, Italy) for his assistance in photographing the fossils and taking measurements. The authors also thank Stefano Dominici and Andrea Savorelli for facilitating access to the Museum of Geology and Paleontology of the University of Florence, the University Museum of Earth Sciences of Sapienza the University of Rome (MUST) for their willingness to allow the collection of photographic and morphometric data on their mammoth and elephant specimens. The authors also thank Asier Larramendi (Eofauna Scientific Research, Donostia, Basque Country, Spain) and Athanassios Athanassiou (Hellenic Ministry of Culture, Superintendency of Palaeoanthropology-Speleology, Athens, Greece) for the constructive comments and suggestions that improved the manuscript. This study was financially supported by Geoscience PhD scholarship to DS (University of Bari-Italy), and by fund “FrancescangeltCTCAVAPONT2022” to MM (University of Bari-Italy). RS acknowledges the Italian Ministry of University and Research, PRIN 2022 scorrimento Prot. 2022JWFEKE “The evolution of mammals in changing ecosystems of the Middle Pleistocene of Italy”.
Editor: Eli Amson
References
Albayrak, E. and Lister, A.M. 2012. Dental remains of fossil elephants from Turkey. Quaternary International 276: 198–211. Crossref
Andrews, C.W. and Cooper, C.F. 1928. On a Specimen of Elephas antiquus From Upnor. 25 pp. British Museum (Natural History), London. Crossref
Arzarello, M., Pavia, G., Peretto, C., Petronio, C., and Sardella, R. 2012. Evidence of an early Pleistocene hominin presence at Pirro Nord (Apricena, Foggia, southern Italy): P13 site. Quaternary International 267: 56–61. Crossref
Athanassiou, A. 2011. A skeleton of Mammuthus trogontherii (Proboscidea, Elephantidae) from NW Peloponnese, Greece. Quaternary International 255 (2012): 9–28. Crossref
Athanassiou, A. 2022. The Fossil Record of Continental Elephants and Mammoths (Mammalia: Proboscidea: Elephantidae) in Greece. In: E. Vlachos (ed.), Fossil Vertebrates of Greece Vol. 1, 345–391. Springer Nature, Cham. Crossref
Averianov, A.O. 1996. Sexual dimorphism in the mammoth skull, teeth, and long bones. In: J. Shoshani and P. Tassy (eds.), The Proboscidea: Evolution and Paleoecology of Elephants and Their Relatives, 260–267. Oxford University Press, New York. Crossref
Azzaroli, A.1967. Carta Geologica d’Italia. Schede per formazioni sedimentarie: Formazione del Calcare di Altamura. Bollettino del Servizio Geologico d’Italia 88: 151–156.
Azzaroli, A., Radina, B., Ricchetti, G., and Valduga, A. 1968. Note Illustrative della Carta Geologica d’Italia, Altamura. Servizio Geologico Italiano 189: 1–21.
Berto, C., Arnaud, J., Lopez-Garcia, J.M., Luzi, E., and Arzarello, M. 2024. Analysis of the Early Pleistocene small mammals from Pirro Nord 13 (Apricena, southern Italy) and their implications for reconstructing the palaeoenvironment of the early human occupation in Europe. Palaeogeography, Palaeoclimatology, Palaeoecology 647: 112251. Crossref
Breda, M. and Lister, A.M. 2013. Dama roberti, a new species of deer from the early Middle Pleistocene of Europe, and the origins of modern fallow deer. Quaternary Science Reviews 69: 155–167. Crossref
Breda, M., Peretto, C., and Thun-Hohenstein, U. 2015. The deer from the early Middle Pleistocene site of Isernia La Pineta (Molise, Italy): revised identifications and new remains from the last 10 years of excavation. Geological Journal 50: 290–305. Crossref
Briatico, G. and Bocherens, H. 2023. Middle Pleistocene ecology in central Italy. New isotopic insights from fauna tooth enamel of Casal de’ Pazzi. Journal of Mediterranean Earth Sciences 15: 141–52.
Christiansen, P. 2004. Body size in proboscideans, with notes on elephant metabolism. Zoological Journal of the Linnean Society 140: 523–549. Crossref
Ciaranfri, N., Pieri, P., and Ricchetti, G. 1988. Note alla carta geologica delle Murge e del Salento (Puglia centromeridionale). Memorie Società Geologica Italiana 41: 499–460.
Davies, P. 2002. The Straight-Tusked Elephant (Palaeoloxodon antiquus) in Pleistocene Europe. 524 pp. Unpublished Ph.D. thesis, University of London, London.
Dubrovo, I.A. 1960. Ancient elephants of the USSR. Trudy Paleontologičeskogo Instituta Akademii Nauk USSR 85: 1–78. [in Russian]
Duval, M., Arnold, L.J., Bahain, J.-J., Pares, J.M., Demuro, M., Falgueres, C., Shao, Q., Voinchet, P., Arnaud, J., Berto, C., Berruti, G.L.F., Daffara, S., Sala, B., and Arzarello, M. 2024. Re-examining the earliest evidence of human presence in western Europe: new dating results from Pirro Nord (Italy). Quaternary Geochronology 82: 101519. Crossref
Erkek, E.E. and Lister, A.M. 2021. The skeleton of a straight‐tusked elephant, Palaeoloxodon antiquus (Falconer and Cautley, 1847) from Selsey, England, and growth and variation in Palaeoloxodon of the European Pleistocene. Journal of Quaternary Science 36: 211–223. Crossref
Espinoza, E.O. and Mann, M.-J. 1993. The history and significance of the Schreger pattern in proboscidean ivory characterization. Journal of the American Institute for Conservation 32: 241–248. Crossref
Falconer, H. and Cautley, P.T. 1847. Fauna Antiqua Sivalensis, Being the Fossil Zoology of the Siwalik Hills, in the North India, Atlas. 80 pp. Smith, Lender and Co, London. Crossref
Ferretti, M. 2008. The dwarf elephant Palaeoloxodon mnaidriensis from Puntali Cave, Carini (Sicily; late Middle Pleistocene): anatomy, systematics and phylogenetic relationships. Quaternary International 182: 90–108. Crossref
Ferretti, M.P. 2010. Anatomy of Haplomastodon chimborazi (Mammalia, Proboscidea) from the late Pleistocene of Ecuador and its bearing on the phylogeny and systematics of South American gomphotheres. Geodiversitas 32: 663–721. Crossref
Göhlich, U.B. 1998. Elephantoidea (Proboscidea, Mammalia) aus dem Mittel-und Obermiozän der Oberen Süßwassermolasse Süddeutschlands: Odontologie und Osteologie. Münchner Geowissenschaftliche A 36: 1–245.
Goldfuss, G. A. 1823. Osteologische Beitrage zur Kenntness Verschiedener Saugetiere der Vorwelt. Nova Acta Physico-Medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum 11: 451–490.
Graells, M. de la P. 1897. Fauna Mastodológica Ibérica. Memorias de la Real Academia de Ciencias Exactas, Físicas y Naturales de Madrid 17: 1–806.
Grube, R., Palombo, M.R., and Iacumin, P. 2010. What did elephants from Neumark Nord eat? In: H. Meller (ed.), Elefantenreich-Eine Fossilwelt in Europa, 252–274. State Museum of Prehistory, Halle. [Complementary volume to the special exhibition].
Hammer, Ø., Harper, D.A., and Ryan, P.D. 2001. PAST: paleontological statistics software package for education and data analysis. Paleontologia Electronica 4: 1–9.
Haynes, G. 1991. Mammoths, Mastodonts and Elephants: Biology, Behavior and the Fossil Record. 413 pp. Cambridge University Press, New York.
Katona, L., Kovács, J., Kordos, L., Szappanos, B., and Linkai, I. 2012. The Csajág mammoths (Mammuthus primigenius): Late Pleniglacial finds from Hungary and their chronological significance. Quaternary International 255: 130–138. Crossref
Kevrekidis, C. and Mol, D. 2016. A new partial skeleton of Elephas (Palaeoloxodon) antiquus Falconer & Cautley, 1847 (Proboscidea, Elephantidae) from Amyntaio, Macedonia, Greece. Quaternary International 406 (B): 35–56. Crossref
Konidaris, G.E. and Tourloukis, V. 2021. Proboscidea-Homo Interactions in Open-air Localities During the Early and Middle Pleistocene of Western Eurasia: A Palaeontological and Archaeolocigal Perspective. 104 pp. Tübingen University Press, Tübingen.
Konidaris, G.E., Athanassiou, A., Tourloukis, V., Thompson, N., Giusti, D., Panagopoulou, E., and Harvati, K. 2018. The skeleton of a straight-tusked elephant (Palaeoloxodon antiquus) and other large mammals from the Middle Pleistocene butchering locality Marathousa 1 (Megalopolis Basin, Greece): preliminary results. Quaternary International 497: 65–84. Crossref
Kroll, W. 1991. Der Waldelefant von Crumstadt. Ein Beitrag zur Osteologie des Waldelefanten, Elephas (Palaeoloxodon) antiquus Falconer & Cautley, 1847. 104 pp. Ph.D. thesis, Ludwig-Maximilians-Universität, Munich.
Larramendi, A. 2016. Shoulder height, body mass, and shape of proboscideans. Acta Palaeontologica Polonica 61: 537–574. Crossref
Larramendi, A., Palombo, M.R., and Marano, F. 2017. Reconstructing the life appearance of a Pleistocene giant: Size, shape, sexual dimorphism and ontogeny of Palaeoloxodon antiquus (Proboscidea: Elephantidae) from Neumark‐Nord 1 (Germany). Bollettino della Società Paleontologica Italiana 56: 299–317.
Larramendi, A., Zhang, H., Palombo, M.R., and Ferretti, M.P. 2020. The evolution of Palaeoloxodon skull structure: Disentangling phylogenetic, sexually dimorphic, ontogenetic, and allometric morphological signals. Quaternary Science Reviews 229: 106090. Crossref
Lister, A.M. 1996. Sexual dimorphism in the mammoth pelvis: an aid to gender determination. In: H. Shoshani and P. Tassy (eds.), The Proboscidea, Evolution and Palaeoecology of Elephants and Their Relatives, 254–259. Oxford University Press, Oxford. Crossref
Lister, A.M. 2016. Dating the arrival of straight‐tusked elephant (Palaeoloxodon spp.) in Eurasia. Bulletin du Musée d’Anthropologie préhistorique de Monaco 6: 123–128.
Lister, A.M. and Stuart, A.J. 2010. The West Runton mammoth (Mammuthus trogontherii) and its evolutionary significance. Quaternary International 228: 180–209. Crossref
Luperto Sinni, E. and Colucci, M. 1985. Il giacimento fossilifero di Contrada Monticelli. In: Gruppo Puglia Grotte (eds.), Atti del 1° Convegno Regionale di Speleologia, Castellana-Grotte, 207–219. Amministrazione comunale, Castellana-Grotte.
Maccagno, A.M. 1962. Gli elefanti fossili di Riano. Geologica Romana 1: 33–131.
Maglio, V.J. 1973. Origin and evolution of the Elephantidae. Transactions of the American Philosophical Society 63: 1–149. Crossref
Mazza, P. and Varola, A. 1999. Revision of the Middle Pleistocene Rhinoceros remains from Contrada Monticelli (Castellana, Bari, Southern Italy). Il Quaternario 12: 99–104.
Mecozzi, B., Bellucci, L., Giustini, F., Iannucci, A., Iurino, D.A., Mazzini, I., Strani, F., and Sardella, R. 2021. A reappraisal of the Pleistocene mammals from the karst infilling deposits of the Maglie area (Lecce, Apulia, southern Italy). Rivista Italiana di Paleontologia e Stratigrafia 127 (2): 355–382.
Mecozzi, B., Fiore, I., Giaccio, B., Giustini, F., Mercurio, S., Monaco, L., Argento, A., Bucci Casari, F., Caricola, I., Lemorini, C., Lucchini, F., Mazzini, I., Palombo, M.R., Sardella, R., Sposato, A., Spinapolice, E.E. and Alhaique, F. 2025a. From meat to raw material: the Middle Pleistocene elephant butchery site of Casal Lumbroso (Rome, central Italy). PLoS One 20(10): e0328840. Crossref
Mecozzi, B., Iannucci, A., Carpentieri, M., Pineda, A., Rabinovich, R., Sardella, R. and Moncel, M.H. 2024a. Climatic and environmental changes of ~100 thousand years: the mammals from the early Middle Pleistocene sequence of Notarchirico (southern Italy). PLoS One 19: e0311623. Crossref
Mecozzi, B., Iurino, D.A., Bertè, D., and Sardella, R. 2017. Canis mosbachensis (Canidae, Mammalia), from the Middle Pleistocene of Contrada Monticelli (Putignano, Apulia, southern Italy). Bollettino della Società Paleontologica Italiana 56: 71–78.
Mecozzi, B., Sardella, R., and Breda, M. 2024b. Late Early to late Middle Pleistocene medium-sized deer from the Italian Peninsula: implications for taxonomy and biochronology. Palaeobiodiversity and Palaeoenvironments 104: 191–215. Crossref
Mecozzi, B., Zorzin, R. and Tomelleri, I. 2025b. A review of the latest Early Pleistocene mammal record from Selva Vecchia (Verona, northern Italy) and its implications for “the 0.9 Ma event” in European faunal dispersal during MIS 21. Quaternary Science Reviews 368: 109575. Crossref
Melentis, J.K. 1963. Studien über fossile Vertebraten Griechenlands. 3. Die Osteologie der pleistozänen Proboscidier des Beckens von Megalopolis in Peloponnes (Griechenland). Annales Géologiques des Pays Helléniques 14: 1–107.
Milli, S. 1997. Depositional setting and high-frequency sequence stratigraphy of the Middle–Upper Pleistocene to Holocene deposits of the Roma Basin. Geologica Romana 33: 99–136.
Milli, S. and Palombo, M.R. 2005. The high-resolution sequence stratigraphy and the mammal fossil record: a test in the Middle–Upper Pleistocene deposits of the Roman Basin (Latium, Italy). Quaternary International 126–128: 251–270. Crossref
Osborn, H.F. 1942. Proboscidea, Vol. 2. 960 pp. American Museum of Natural History, New York.
Palombo, M.R. and Cerilli, E. 2021. Human-elephant interactions during the Lower Palaeolithic: Scrutinizing the role of environmental factors. In: G.E. Konidaris, R. Barkai, V. Tourloukis, and K. Harvati (eds.), Human-Elephant Interactions: From Past to Present, 105–143. Tübingen University Press, Tübingen.
Palombo, M.R. and Ferretti, M.P. 2005. Elephant fossil record from Italy: knowledge, problems, and perspectives. Quaternary International 126–128: 107–136. Crossref
Palombo, M.R. and Villa, P. 2001. Schreger lines as support in the Elephantinae identification. In: G. Carravetta, P. Gioia, M. Mussi, and M.R. Palombo (eds.), La Terra degli Elefanti. The Worlds of Elephant, 656. Consiglio Nazionale delle Ricerche, Roma.
Palombo, M.R. and Villa, P. 2003. Sexually dimorphic characters of Elephas (Palaeoloxodon) antiquus from Grotte Santo Stefano (Viterbo, Central Italy). In: J.W.F. Reumer, J. de Vos, and D. Mol (eds.), Advances in Mammoth Research. Proceedings of the Second International Mammoth Conference, Rotterdam, May 16–20, 1999. Denisea 9: 293–315.
Palombo, M.R., Albayrak, E., and Marano, F. 2010. The straight-tusked Elephants from Neumark-Nord, a glance into a lost world. In: H. Meller (ed.), Elefantenreich. Eine Fossilwelt in Europa, 218–251. Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt Landesmuseum für Vorgeschichte, Halle.
Palombo, M.R., Sardella, R., and Bellucci, L. 2024. The straight-tusked elephant from Contrada Calorie (Basilicata, Southern Italy). Preliminary notes. Journal of Mediterranean Earth Sciences 16: 1–26.
Pavia, G., Bertok, C., Ciampo, G., Di Donato, V., Martire, L., Masini, F., Santangelo, N., Taddei, R.E., and Zunino, M. 2010. Depositional events in the Pliocene to Lower Pleistocene succession of the Apricena-Lesina-Poggio Imperiale minig district (western Gargano, Foggia, southern Italy). Bollettino della Società Geologica Italiana 129: 132–155.
Peretto, C., Arnaud, J., Moggi-Cecchi, J., Manzi, G., Nomade, S., Pereira, A., Falguéres, C., Bahain, J.J., Grimaud-Hervé, D., Berto, C., Sala, B., Lembo, G., Muttillo, B., Gallotti, R., Thun Hohenstein, U., Vaccaro, C., Coltorti, M., and Arzarello, M. 2015. A human deciduous tooth and new 40Ar/39Ar dating results from the Middle Pleistocene archaeological site of Isernia La Pineta, Southern Italy. PLoS ONE 10: e0140091. Crossref
Petronio, C. and Sardella, R. 1999. Biochronology of the Pleistocene mammal fauna from Ponte Galeria (Rome) and remarks on the Middle Galerian faunas. Rivista Italiana di Paleontologia e Stratigrafia 105: 155–164.
Reggiani, P. 2001. Morphological differences in Mammuthus meridionalis and Palaeoloxodon antiquus carpal bones. In: G. Carravetta, P. Gioia, M. Mussi, and M.R. Palombo (eds.), La Terra degli Elefanti. The Worlds of Elephant, 661–664. Consiglio Nazionale delle Ricerche, Roma.
Roditi, E., Bocherens, H., Konidaris, G.E., Athanassiou, A., Tourloukis, V., Karkanas, P., Vangelis Tourloukis, V., Karkanas, P., Panagopoulou, E., and Harvatiet, K. 2024. Life-history of Palaeoloxodon antiquus reveals Middle Pleistocene glacial refugium in the Megalopolis basin, Greece. Scientific Report 14: 1390. Crossref
Roth, V.L. 1990. Insular dwarf elephants: a case study in body mass estimation and ecological inference. In: J. Damuth and B. MacFadden (eds.), Body Size in Mammalian Paleobiology: Estimation and Biological Implications, 151–179. Cambridge University Press, Cambridge.
Rustioni, M., Ferretti, M.P., Mazza, P., Pavia, M., and Varola, A. 2003. The vertebrate fauna from Cardamone (Apulia, southern Italy): an example of Mediterranean mammoth fauna. Deinsea 9: 395–404.
Saarinen, J. and Lister, A.M. 2016. Dental mesowear reflects local vegetation and niche separation in Pleistocene proboscideans from Britain. Journal of Quaternary Science 31: 799–808. Crossref
Saegusa, H. and Gilbert, W.H. 2008. Elephantidae. In: W.H. Gilbert and B. Asfaw (eds.), Homo erectus. Pleistocene Evidence from the Middle Awash, Ethiopia, 193–226. University of California Press, Berkeley. Crossref
Sala, B. 2006. Le nuove specie rinvenute a Isernia La Pineta. In: C. Peretto (ed.), Preistoria in Molise, gli Insediamenti del Territorio di Isernia, 36–38. Centro Europeo di Ricerche Preistoriche, Collana Ricerche 3, Pomezia.
Schreve, D. and Jones, A.K. 2005. Vertebrate overview. In: S. Elias (ed.), Encyclopedia of Quaternary Science (Third edition), 272–285. Elsevier, Amsterdam. Crossref
Selleri, G. 2007. Karstic landscape evolution of southern Apulia fore-land during the Pleistocene. Geografia Fisica e Dinamica Quaternaria 30: 77–86.
Selleri, G., Sanso, P., and Walsh, N. 2003. The karst of Salento region (Apulia, southern Italy): constraints for management. Acta Carsologica 32: 19–28. Crossref
Shoshani J. (1996). Skeletal and other basic anatomical features of elephants. In: J. Shoshani and P. Tassy (eds.), The Proboscidea—Evolution and Palaeoecology of Elephants and Their Relatives, 9–20. Oxford University Press, Oxford. Crossref
Stefanelli, D., Mecozzi, B., Francescangeli, R., Girone, A., Marino, M., Montenegro, V., Sardella, R., and Breda, M. 2021. The Fallow deer from Contrada Monticelli (Bari, Southern Italy): the southernmost occurrence of a large Dama of the early Middle Pleistocene. Bollettino della Società Paleontologica Italiana 60: 55–60.
Stefanelli, D., Mecozzi, B., Marino, M., Sardella, R. and Breda, M. 2024a. The postcranial variability of Quaternary European rhinoceroses: the case study of Stephanorhinus hundsheimensis from the Middle Pleistocene site of Contrada Monticelli (Apulia, southern Italy). Historical Biology 37: 758–777. Crossref
Stefanelli, D., Sorbelli, L., Mecozzi, B., Maiorano, P., Marino, M. and Sardella, R. 2024b. The Bison sample from the early Middle Pleistocene site of Contrada Monticelli (Apulia, southern Italy). Rivista Italiana di Paleontologia e Stratigrafia 130: 525–544. Crossref
Stuart, A.J. and Lister, A.M. 2001. The mammalian faunas of Pakefield/Kessingland and Corton, Suffolk, UK: evidence for a new temperate episode in the British early Middle Pleistocene. Quaternary Science Reviews 20: 1643–1656. Crossref
Sukumar, R. 2003. The Living Elephants: Evolutionary Ecology, Behaviour, and Conservation. 494 pp. Oxford University Press, Oxford. Crossref
Tassy, P. 1985. La place des mastodontes Miocènes de l’ancien monde dans la phylogénie des Proboscidea (Mammalia): hypothèses et conjectures. 861 pp. Ph.D. Thesis, Université Pierre et Marie Curie, Paris.
Trapani, J. and Fisher, D.C. 2003. Discriminating proboscidean taxa using features of the Schreger pattern in tusk dentin. Journal of Archaeological Science 30: 429–438. Crossref
Tsakalidis, C., Konidaris, G.E., Tsoukala, E., and Kostopoulos, D.S. 2025. Dietary Reconstruction of Pliocene–Pleistocene Mammoths and Elephants (Proboscidea) from Northern Greece Based on Dental Mesowear Analysis. Quaternary 8: 19. Crossref
Tsoukala, E. and Lister, A.M. 1998. Remains of straight-tusked elephant, Elephas (Palaeoloxodon) antiquus Falc. & Caut., 1847 ESR-dated to oxygen isotope stage 6 from Grevena (W. Macedonia, Greece). Bollettino della Società Paleontologica Italiana 37: 117–139.
Verri, A. 1886. Relazione sulle escursioni nei dintorni di Terni. Bollettino della Società Geologica Italiana 5: 507–514
Virág, A. 2014. Dental remains of Elephas antiquus Falconer & Cautley 1847 (Mammalia, Proboscidea, Elephantidae) in Hungary. Hantkeniana 9: 117—131.
Acta Palaeontol. Pol. 71 (1): 155–171, 2026
http://doi.org/10.4202/app.01304.2025