
An exceptional diverse trilobite ichnofauna from the uppermost Devonian of Anti-Atlas, Morocco
WAHIBA BEL HAOUZ, ABDELOUAHED LAGNAOUI, CHRISTIAN KLUG, MEHDI MAANAN, and LAHSSEN BAIDDER
Bel Haouz, W., Lagnaoui, A., Klug, C., Maanan, M., and Baidder, L. 2026. An exceptional diverse trilobite ichnofauna from the uppermost Devonian of Anti-Atlas, Morocco. Acta Palaeontologica Polonica 71 (2): 273–295.
We present a highly diverse ichnoassemblage of latest Devonian (Famennian) age from the eastern Anti-Atlas of Morocco, dominated by trilobite and other arthropod trace fossils. It consists of invertebrate and vertebrate ichnotaxa, including Cruziana lobosa, Diplichnites gouldi, Rusophycus antiatlasensis isp. nov., Rusophycus cf. carleyi, associated with Arenicolites isp., Fustiglyphus isp., Lockeia isp., Mammillichnis isp., Planolites isp., Paracanthorhaphe isp., Treptichnus isp., and Undichna isp. These ichnoassemblages are ascribed to the archetypal Cruziana ichnofacies, indicating a shallow-marine environment within low hydrodynamic-energy settings, stable substrate conditions and regular organic matter sources. The results of this ichnological study are remarkable in the following aspects: (i) the report of the youngest occurrence of C. lobosa, (ii) introduction of R. antiatlasensis isp. nov., as a new ichnospecies and (iii) the evidence of collective behaviour and predation as well as protection strategies of trilobites. Judging from the diversity of the invertebrate and vertebrate trace fossils and previously described fish and cephalopod remains, a shallow marine ecosystem was present during the latest Devonian in the eastern Anti-Atlas.
Key words: Trilobita, invertebrate ichnoassemblages, ichnodiversity, palaeoethology, new ichnotaxa, Famennian, Devonian, Morocco.
Wahiba Bel Haouz [wahiba.belhaouz@gmail.com; ORCID: https://orcid.org/0000-0002-4799-9697 ], Mehdi Maanan [mehdi.maanan@gmail.com; ORCID: https://orcid.org/0000-0003-3019-6860 ], and Lahssen Baidder [lbaidder@gmail.com; ORCID: https://orcid.org/0000-0001-5318-7102 ], Géosciences, Géomatique, Planétologie and Environnement Laboratory, Department of Geology, Faculty of Sciences Ain Chock, Hassan II University, km 8 Route d’El Jadida, 20100, Casablanca, Morocco.
Abdelouahed Lagnaoui [abdelouaed.lagnaoui@uhp.ac.ma; ORCID: https://orcid.org/0000-0002-4176-1987 ], Laboratory of Education, Sciences and Techniques, Higher School of Education and Training Berrechid, Hassan First University, Route de Casablanca km 3.5, BP 539, 26100, Berrechid, Casablanca-Settat, Morocco.
Christian Klug [chklug@pim.uzh.ch; ORCID: https://orcid.org/0000-0002-4099-7453 ], Paläontologisches Institut, Universität Zürich, Karl-Schmid-Strasse 4, 8006, Zürich, Switzerland.
Received 30 November 2025, accepted 12 February 2026, published online 17 June 2026.
Copyright © 2026 W. Bel Haouz et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Devonian sections of the eastern Anti-Atlas of Morocco are of great interest to vertebrate and invertebrate palaeontology for almost a century (Clariond 1934a, b; Roch 1934; Frey et al. 2013, 2019). Although it is rich in invertebrate and vertebrate ichnofossils (Klug et al. 2018), ichnological publications focusing on the Devonian are rather rare in comparison to the ones on body fossils. Very few papers on vertebrate and invertebrate ichnofauna have been published, all in recent years (e.g., Klug and Hoffmann 2018; Lagnaoui et al. 2019; Klug et al. 2021; Bel Haouz et al. 2026). Apart from its importance for palaeobiological (Klug et al. 2021), as well as palaeoenvironmental and palaeoecological interpretation (Lagnaoui et al. 2019; Bel Haouz et al. 2026), the Devonian trace fossils significantly contribute to our understanding of benthic activity, ecosystem evolution, particularly where traditional palaeontological approach is limited by complex sedimentary environments and episodic preservation.
The Devonian of the eastern Anti-Atlas of Morocco is well-known for its rich, diverse and well-preserved faunal associations including trilobites, cephalopods, bivalves, brachiopods, crinoids, and fishes (Klug and Pohle 2018; Frey et al. 2019). All of these could leave traces in and on sediments within their habitats. The arthropod fossil record is especially well documented through body fossils of trilobites and a few other groups (e.g., Klug et al. 2008; De Baets et al. 2010; Jobbins et al. 2020), with a unique report of xiphosuran trace fossils from the Middle Devonian strata (Bel Haouz et al. 2026). Here, we provide a first comprehensive analysis of trilobite ichnoassemblages associated with vertebrate and invertebrate trace fossils from the uppermost Devonian of the eastern Anti-Atlas of Morocco and their ethological, palaeoenvironmental and palaeoecological implications.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank https://zoobank.org/NomenclaturalActs/90EABD51-E986-4566-8F36-3A44E50B992E
Institutional abbreviation.—ESEFB-LTM, Higher School of Education and Training of Berrechid, Life Traces Museum, Berrechid, Morocco.
Geological setting
The Moroccan Anti-Atlas is a large, NE-SW-oriented anticlinorium, mainly composed of well-exposed Precambrian basement rocks (exposed as inliers) covered by Palaeozoic Cambrian to Carboniferous sediments. The Eastern Anti-Atlas is considered as having the world’s most complete sedimentary record from the Precambrian to the Carboniferous (Wendt et al. 1984; Wendt 2021a, b). The outcrops extend over a 1000 km belt along the western and northern margins of the Tindouf Basin (Fig. 1), providing an excellent biostratigraphic illustration of the Devonian exposed mostly in isolated E-W oriented synforms and antiforms (Wendt et al. 1984; Döring and Kazmierczak 2001; Frey et al. 2013), comprising siliciclastics and carbonates (e.g., Michard 1976; Wendt 1985, 1988; Baidder et al. 2016; Letsch et al. 2019).
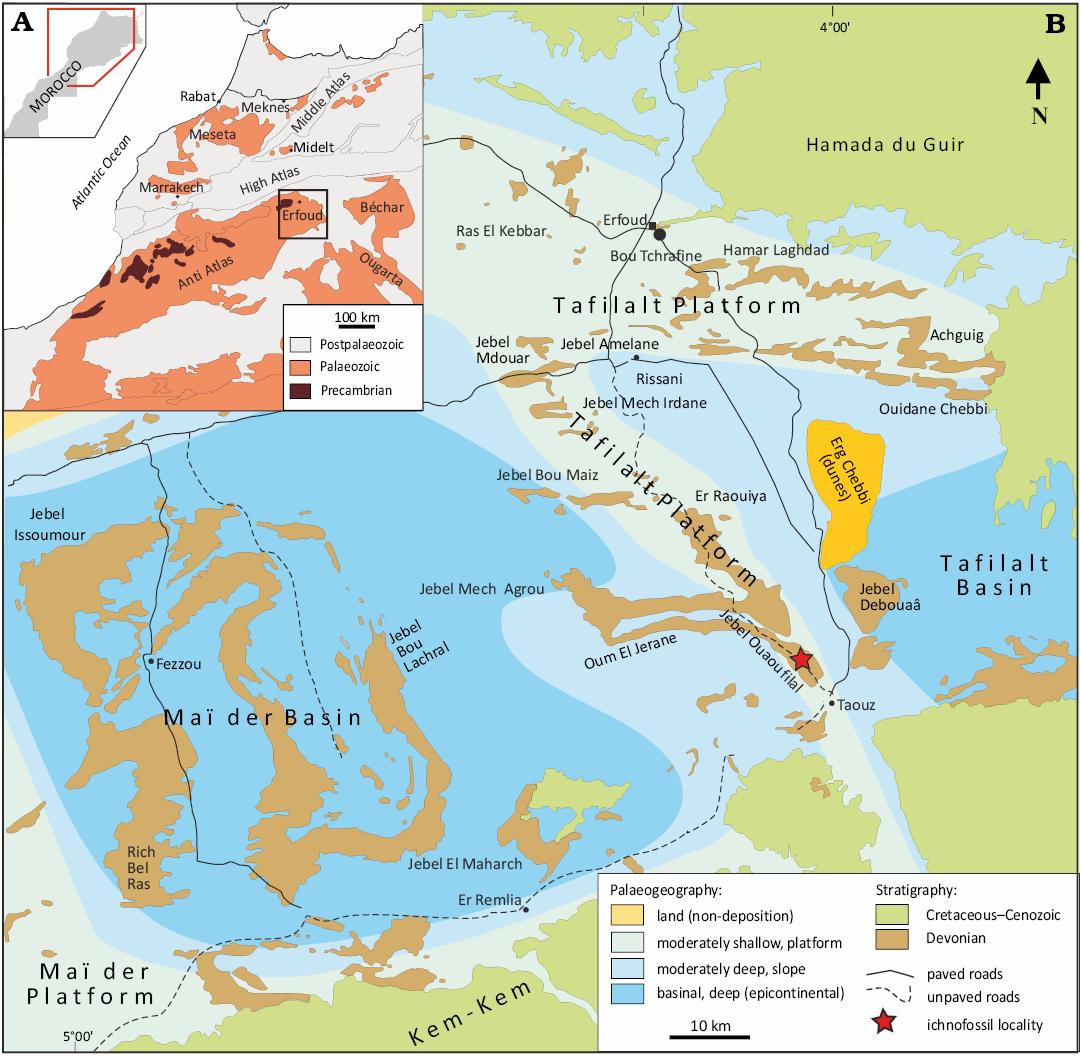
Fig. 1. Geographic location and geological setting of the study area. A. The location of the Tafilalt region in the Anti-Atlas of Morocco. B. A superimposed geographic, palaeogeographic, and geological map showing the approximate outlines of the Maïder and Tafilalt basins, as well as the Tafilalt platform, and the Late Devonian palaeogeography of the eastern Anti-Atlas (modified after Wendt and Belka 1991; Frey et al. 2018).
During the Palaeozoic, the eastern Anti-Atlas was part of the broad passive continental margin of northwestern Gondwana (Scotese 2014; Hüneke et al. 2023). This palaeogeographic position, combined with the moderate southern latitude, enabled diverse ecosystems to be established repeatedly (e.g., Töneböhn 1991; Frey et al. 2013; Klug et al. 2018). The platforms and basins were established along the northern margin of the West African Craton during an extensive early Hercynian phase (Wendt et al. 1984).
Due to differential subsidence, the eastern Anti-Atlas area experienced increasing differences in sediment accumulation rates and depositional environments during the Devonian. A homoclinal carbonate ramp dipping with some topographic variation (Lubeseder et al. 2010) was formed. During the Middle Devonian, the region began to differentiate into the central, approximately N-S running pelagic, somewhat shallower Tafilalt Platform, that was a pivot connecting the two flanking Maïder and Tafilalt basins to the west and east, respectively (Fig. 1), as well as the Maïder Platform to the southwest, surrounded by uplifted areas to the northwest and south (Wendt et al. 1984; Wendt 1985, 1988a, b; Wendt and Belka 1991; Baidder et al. 2016; Ait Daoud et al. 2019). Two different fold directions dominate the ductile deformation structures in the eastern Anti-Atlas (Tafilalt and Maïder), which resulted from late Variscan compression in a NE-SW direction (Ait Daoud et al. 2019), an E-W axis fold dominates in the western part, and a NW-SE axis in the eastern part (Kaufmann 1998; Tawadros 2018; Ait Daoud et al. 2019).
The materials of the newly studied ichnofauna presented herein were collected from the Aoufilal Formation between the villages of El Khraouia and Taouz as well as from the Filon 12 mine, located on the Tafilalt platform (Fig. 2). The Tafilalt Platform, also known as Tafilalt pelagic ridge, is a palaeogeographic element, extending across the eastern Ougnat-Ouzina Axis and the Ougnat-Erfoud High (Wendt 1985, 1989; Wendt and Belka 1991; Baidder et al. 2008, 2016; Lubeseder et al. 2010; Hüneke et al. 2023); its biostratigraphy and palaeontology are well documented (Wendt et al. 1984; Klug et al. 2018; Wendt 2021a, b), consisting mainly of carbonatic sediments showing lateral variations (Wendt and Belka 1991; Lubeseder et al. 2010). They are characterised by a break in sedimentation above the Frasnian–Famennian boundary; thus, the lower Famennian is marked by cephalopod-rich limestone (Wendt et al. 1984; Wendt 1988; Hüneke et al. 2023). During the Eifelian, Givetian, and mid-Frasnian, the sedimentary facies of Tafilalt platform underwent significant changes on a platform surface that sloped gently towards the north (Wendt et al. 1984; Wendt 1985, 1988), transitioning laterally from two facies:
(i) nodular cephalopod limestones of Eifelian, Givetian, and mid-Frasnian age (Lubeseder et al. 2010); these transition in the upper Frasnian to the Famennian into more massive, fossiliferous limestones (Kellwasser Limestone) contain various skeletal debris including tentaculitids, cephalopods, trilobites, brachiopods, bryozoans, corals, crinoids, and gastropods (Hüneke et al. 2023), deposited in previously shallow or even emergent areas, where sea level fell during the late Famennian (Wendt 1988). These limestones are weakly bioturbated, with open-spaced burrows that are often filled with internal sediment and/or calcite cement. They are associated with a hardground on the crest of the ridge. Thus, the moderately to highly diverse Famennian marine assemblages indicate normal salinity and occasionally well-oxygenated bottom conditions, as well as repeated anoxic phases (Wendt et al. 1984; Klug et al. 2008, 2018).
(ii) The much thicker sequences of basinal facies (upper and lower slopes) are dominated by alternating greyish to dark, well-bedded limestones, marly limestones, and marls (including the lower Eifelian to middle Givetian limestones, Givetian–Frasnian marls, and Kellwasser limestones). The carbonate facies show a continuum ranging from laminated mudstones to coarser-grained, cross-bedded limestones (Hüneke et al. 2023). The depositional slopes had an average inclination of 2°, which resulted in slumping and debris flows. The water was probably much shallower than 500 m, locally and temporally reaching the photic zone during the late Famennian (Wendt 1988).
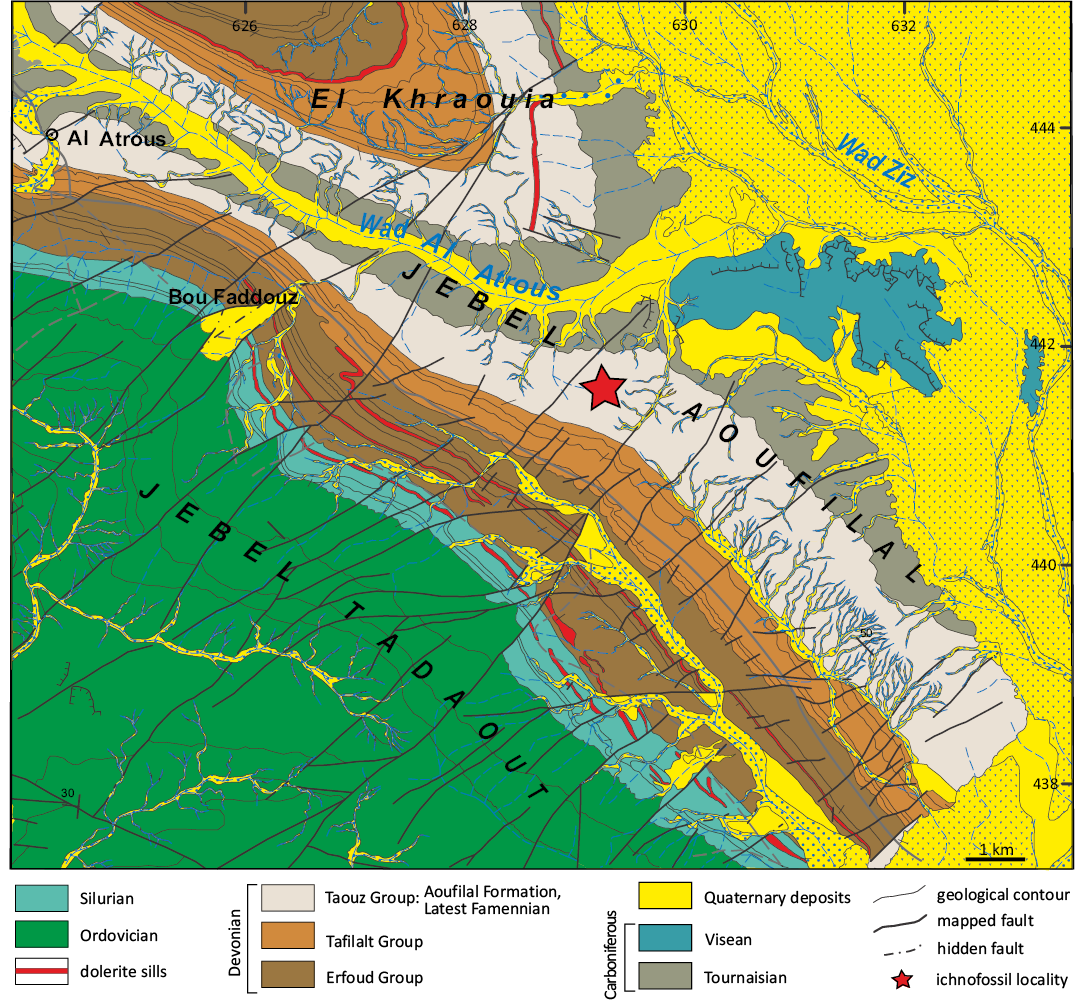
Fig. 2. Detailed geological setting of the study area. A. Geological map of the Al Atrous region showing the locations of trace fossils (after Álvaro et al. 2014).
The Palaeozoic deposits in the South Tafilalt region consists of graptolitic argillaceous limestones and limestones with Orthoceras of the Ordovician–Silurian age (Fig. 2A, Álvaro et al. 2014). The Devonian successions are subdivided into three formal group, which are: Erfoud, Tafilalet and Taouz Groups mounds (Fig. 2A, Álvaro et al. 2014, and reference therein). During the Early Devonian the strata consist mainly of deep-marine shales and terrigenous mudstones intercalated with fossil-rich limestones (Lochkovian–Praguian), as well as shallow-marine fossil-rich limestones (Late Praguian–Early Emsian). These strata are overly by the stratigraphic marker terrigenous mudstones, trilobite-rich marly limestones and argillaceous pelites (Late Emsian–Lowermost Eifelian). These successions are overlain by nodular, lumachellic and turbiditic limestones (Late Eifelian–Middle Givetian), which are followed by Cephalopod-rich limestones, bituminous limestones and the Kellwasser black shales (Frasnian–Early Famennian). The Devonian deposits capped by the terrigenous mudstones and the Hangenberg sandstone equivalent that is ended by microconglomerate layers, corresponding to Aoufilal Formation (Late Famennian). Finally, the Carboniferous is composed of deltaic deposits consisting mainly of claystones, conglomerates, sandstones with brachiopods, argillaceous pelites and chaotic breccia (Tournaisian–Visean) (Fig. 2A, Álvaro et al. 2014; Najih et al. 2019 and reference therein). The invertebrate trace fossils described herein were collected from the Aoufilal Formation (Fig. 3), which consists of a series of clastic mudstones and sandstones 400 m thick (Grès d’Aoufilal in the Taouz region) with cross stratification, wave ripple marks, and ferruginous crusts. This formation is capped by a microconglomerate layer containing highly bioturbated sandstones, referred to the Famennian and marking the transition to the Tournaisian (Strunian) (Wendt et al. 1984). The Famennian deposits on the Tafilalt Platform range in thickness from 50 m to just a few condensed metres (especially in cephalopod limestone), with some or hiati. In contrast, the successions of the marginal and southern Tafilalt Platform display relatively continuous sedimentation, suggesting subtidal conditions (Wendt et al. 1984). Clastic input was almost non existent in the central and northern parts of the platform, but increased towards the south.
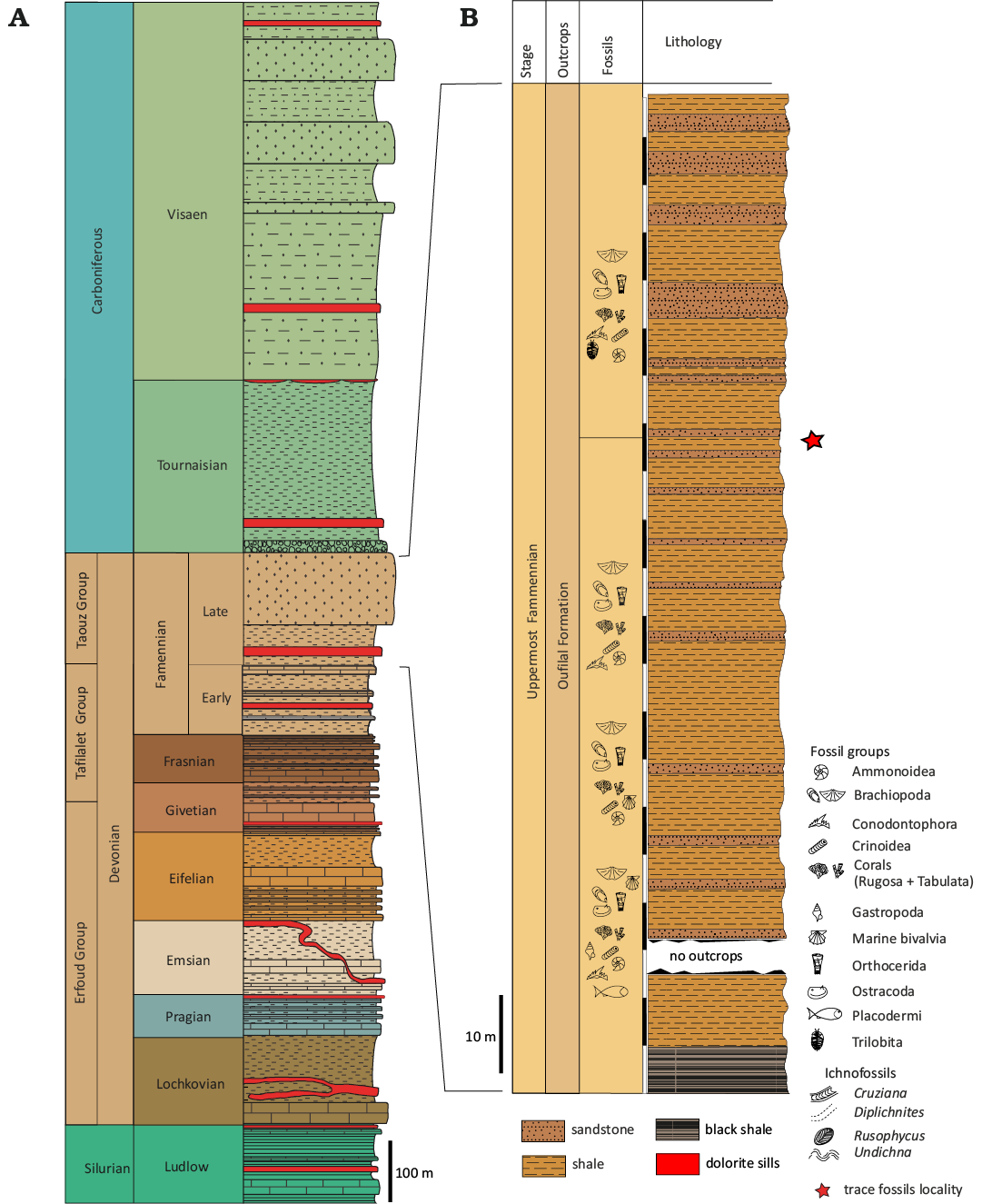
Fig. 3. Stratigraphic subdivision of the study area. A. Middle to upper Palaeozoic stratigraphy subdivision of the South Tafilalt region, modified (after Álvaro et al. 2014; Najih et al. 2019). B. Lithological section of the upper Famennian stage of the Aoufilal Formation, showing the stratigraphic position of the ichnofossiliferous strata.
Material and methods
The invertebrate trace fossils described herein come from the Aoufilal Formation in El Khraouia, Taouz and Filon 12, located on the Tafilalt platform, in the eastern Anti-Atlas (Fig. 1). The material was collected during field trips to various sites in the Tafilalt region (see Fig. 2) between 2018 and 2025. More than 60 specimens were photographed, analysed and measured. The described traces formed at the sediment-water interface through the activities of animals on the seafloor. These trails were preserved by the subsequent deposition of coarse sediment that is typically more marly, and filled moulds on the base of the overlying bed (hypichnial casts). The ichnofossil-bearing sequences contain disarticulated trilobites, which are often abundant. All these invertebrate burrows are preserved in convex hyporelief within fine-grained sandstones.
Specimens were photographed under natural white light and a spotlight. Measurements were taken for several parameters of trace morphology and pattern of the best-preserved specimens according to standards of description and identification of arthropod trackways suggested by Trewin (1995). The well-preserved specimens are housed at ESEFb-LTM.
Systematic palaeoichnology
Ichnogenus Cruziana d’Orbigny, 1842
Type ichnospecies: Cruziana rugosa d’Orbigny, 1842, by subsequent designation Miller (1889); Liriuni, Bolivia, Ordovician.
Cruziana lobosa Seilacher, 1970
Figs. 4–6.
Material.—Four slabs with many traces preserved as convex hyporelief in fine-grained sandstones from the uppermost Devonian (Famennian) of the Jebel Aoufilal area, El Khraouia, eastern Anti-Atlas, Morocco. ESEFB-LTM-101, slab with three curved to winding, well-preserved trails (Fig. 4); ESEFB- LTM-102, slab with curved to winding trail overlapped by imbricated burrows and series of parallel striae (Fig. 5); ESEFB-LTM-103, slab with curved to winding invertebrate and overcrossing burrow/trails with other trace fossils (Fig. 6A1); ESEFB-LTM-104: slab with curved to winding and overcrossing trails (Fig. 6B1).
Original diagnosis.—Deep furrow with rounded, blunt-ended transverse leg markings rather than distinct claw marks, and smooth pleural lobes (Seilacher 1970).
Emended diagnosis.—Horizontal, tapered, bilobate, ribbon-like trail with rounded, blunt-ended transverse scratches separated by a median furrow and surrounded by two lateral grooves and ridges.
Description.—Unbranched, horizontal, trace fossils preserved in curved, winding to looping paths in convex hyporelief (Figs. 4–6). The trails consist of long, shallow to moderately deep, tapered, bilobate, ribbon-like structures separated by a median furrow with fine, slightly oblique, blunt-ended transverse scratch produced by the endopods of a trilobite or similar arthropod in the central region, which are enveloped by two exopodal lateral continuous grooves and ridges (Figs. 4A1, 5A1, 6A1, B1). The regular scratches are arranged in a chevron pattern, converging at an angle of 50–70° with respect to the median axis, and may vary by several tens of degrees along the length of a single specimen (Figs. 4A1, A2, 5A1–A4, 6A2–A5, B2, B3). The burrows are frequently criss-crossed with about 90° preferred angle (Figs. 4, 6A1, B1).

Fig. 4. Trace fossil Cruziana lobosa Seilacher, 1970 (slab ESEFB-LTM-101), from the Famennian (uppermost Devonian) of the Jebel Aoufilal, eastern Anti-Atlas, Morocco. Three curved, winding invertebrate burrow/trails preserved in convex hyporelief: general view showing the bilobate morphology and fine transverse striation (A1), close-up highlighting the detailed scratches and lobe structure (A2), line drawing emphasizing the course and morphology of the three trails (1–3) (A3). Scale bars 20 mm.
ESEFB-LTM-101 exhibits three curved to winding ploughing trails, which are approximately 10 mm wide and over 160 mm long (Fig. 4). It bears three trails on one slab: (i) trail T3 is an entirely eroded, 10 mm wide and over 160 mm long, without ornamentation and scratches; (ii) trail T2 consists of 10 mm wide and more than 160 mm long, mostly eroded burrow, but preserves faint to locally well-preserved ornamentation and scratch traces, and (iii) a well-preserved trail T1 of a 10 mm wide and more than 130 mm long burrow with very well-preserved ornamentation and scratch traces. The endopodal lobes are well developed with continuous and a densely packed chain of small uniserial pustule-like elevations, each being approximately 2 mm wide (Fig. 4A3).
ESEFB-LTM-102 contains a curved to winding trail measuring approximately 13 mm in width and over 200 mm in length (Fig. 5). The grooves/ridges are characterised by small, closely spaced pustule-like elevations and measure about 2 mm in width.

Fig. 5. Trace fossil Cruziana lobosa Seilacher, 1970 (slab ESEFB- LTM-102), from the Famennian (uppermost Devonian) of the Jebel Aoufilal, eastern Anti-Atlas, Morocco. A curved, winding invertebrate burrow/trail preserved in convex hyporelief: general view of the specimen (A1), detailed views of successive segments showing C. lobosa overlapped by imbricated burrows and surrounded by parallel scratches (A2–A4), line drawing (A5), emphasizing the main morphological features, including C. lobosa. (Cr), Monomorphichnus lineatus Crimes et al., 1977 (Mo) and Planolites isp. (Pal). Scale bars 20 mm.
ESEFB-LTM-103 and ESEFB-LTM-104 exhibit a combination of the two kinds of trace fossils described above, consisting of meandering, ploughing structures; the traces transform into shallow trails with faint scratches and sometimes smooth surfaces (Fig. 6). These traces criss-cross each other, and are 5–8 mm wide and over 200 mm long. The endopodal lobes are well developed and rarely flanked by a low, sharp ridge, each approximately 0.5–1 mm wide. The ornamentation and scratches are well-preserved, locally faintly preserved or missing.

Fig. 6. Trace fossils Cruziana lobosa Seilacher, 1970, from the Famennian (uppermost Devonian) of the Jebel Aoufilal, eastern Anti-Atlas, Morocco. A. ESEFB-LTM-103, slab showing multiple overlapping and intersecting bilobate trails preserved in convex hyporelief: general view (A1), detailed views with clearly defined transverse scratches (A2–A5). B. ESEFB- LTM-104, slab from the same layer as A, displaying several parallel and intersecting trails: general view (B1), close-up views of individual specimens (B2–B5), displaying variation in width, curvature and preservation of transverse ridges C. lobosa (Cr), Undichna isp. (Un), and enigmatic trackways (Et). Scale bars 20 mm.
Remarks.—Cruziana was first defined by d’Orbigny (1842) as a ribbon-shaped fucoid. Subsequently, all ribbon-like, bilobate repichnial traces were included to Cruziana, while shorter, suboval, buckle-like cubichnial trace fossils were distinguished as Rusophycus (Miller 1889; Seilacher 1955; Stanley and Pickerill 1998). The ichnogenus Cruziana comprises numerous species that are distributed across the Palaeozoic and lower Mesozoic. In the Devonian, Cruziana is less widespread than in other systems (lower Cambrian to Silurian). All species display transverse scratches, but no recognisable claw formula is identified in our material. Seilacher (2007) reported three ichnospecies of Cruziana from the Devonian: Cruziana rhenana Seilacher, 1970, which displays pleural impressions bounding short rusophyciform lobes; C. uniloba Seilacher, 1970, which consists of heavy endopodal scratches on either side that meet along the midline; and C. lobosa, which has less pronounced endopodal scratches. Morphologically, the species that most closely resembles the invertebrate trace fossils described here is Cruziana quadrata (Seilacher, 1970), which has never been recorded from the Devonian before. Cruziana quadrata has thinner, oblique scratches and it has clearly rounded, blunt to bundle-shaped endopodial striellae. Generally, C. quadrata has a large width ranging from 40–65 mm (Seilacher 1970). The features of C. quadrata are not recorded in the Famennian material studied herein, which have a maximum width of 13 mm. The tapered, bilobate, ribbon-like burrow with rounded, blunt-ended transverse scratch traces separated by a median furrow and surrounded by two lateral grooves and ridges are the main diagnostic features of the ichnospecies C. lobosa. The latter differs from C. quadrata in size and the endopodal scratches, which are sharper in C. quadrata and less sharp in C. lobosa (Seilacher 2007). Therefore, we assign our material to C. lobosa.
Stratigraphic and geographic range.—Upper Ordovician Georgian Bay Formation of Southern Ontario in Eastern Canada (reported with incertitude, Stanley and Pickerill 1998). Middle Devonian Aouinet Quenine Formation in Libya (Seilacher 1970). Uppermost Devonian (Famennian) of the Jebel Aoufilal area (locality?}, Anti-Atlas, Morocco, youngest occurrence of C. lobosa.
Ichnogenus Diplichnites Dawson, 1873
Type ichnospecies: Diplichnites aenigma Dawson, 1873; Pennsylvanian strata of the Coal Mine Point, Joggins, Nova Scotia, Canada. However, Dawson’s type specimen has not been located and no replicas are known.
Diplichnites gouldi Gevers in Gevers et al., 1971
Fig. 7.
Material.— Nine slabs with many traces preserved as convex hyporelief in fine-grained sandstones from the uppermost Devonian (Famennian) of the Jebel Aoufilal area (El Khraouia and Filon 12), eastern Anti-Atlas, Morocco. ESEFB-LTM-105, slab with long trackway consisting of two parallel rows of tracks interrupted by a short bilobate burrow (Fig. 7A); ESEFB-LTM-106, slab with a short trackway consisting of two parallel rows of tracks (Fig. 7B); ESEFB-LTM-107, slab with a short trackway consisting of two parallel rows of dense and closely spaced tracks (Fig. 7C); ESEFB-LTM-108–113, six slabs with variably long trackways consisting of two parallel rows of tracks (Fig. 7D–H).

Fig. 7. Trace fossil Diplichnites gouldi Gevers in Gevers et al., 1971, and associated trace fossils, Arenicolites isp. (Ar), Fustiglyphus isp. (Fu), Lockeia isp. (Lo), Mammillichnis isp. (Ma) Treptichnus isp. (Tr), from the Famennian (uppermost Devonian) of the Jebel Aoufilal, eastern Anti-Atlas, Morocco. Slabs displaying invertebrate trackways preserved as convex hyporeliefs in fine-grained sandstone. A. ESEFB-LTM-105, a slab with a long trackway consisting of two parallel rows of tracks, interrupted by a short bilobate burrow. B–H. Slabs displaying shorter trackways, consisting of two parallel rows of closely spaced impressions. The trackways vary in curvature and track density, yet they all display the characteristic paired arrangement typical for Diplichnites. ESEFB-LTM-106 (B), ESEFB-LTM-107 (C), ESEFB-LTM-108 (D), in situ slabs photographed in the outcrop during 2023 fieldtrip (E–G). Close up view of the diplichnites in E1, photographed in the outcrop during 2023 fieldtrip (E2, E3). H. Lose slab photographed in the outcrop during March 2022 fieldtrip. Scale bars 20 mm.
Description.—Simple, straight to gently curved trackways consisting of two parallel rows of closely spaced and well-organised to unorganised series of tracks in bilateral symmetry (Fig. 7). The trackways are preserved as positive hyporelief or negative epirelief, with lengths between 100 mm and more than 600 mm. The internal width varies between 14 and 16 mm, and the external width between 16 and 24 mm with an external/internal width ratio of less than 1.5. The minimum width of the tracks ranges from 11 to 18 mm, and the maximum width from 14 to 22 mm, with internal widths ranging from 8 to 14 mm. The tracks show diverse extramorphological variations of imprint, from comma internal (Fig. 7), internally tapered (Fig. 7C), elongate (Fig. 7H), gently curved to crescentic dimorphic-shaped (Fig. 7D, A3) to ellipsoidal (Fig. 7E1, E3, F, G) of multilegged arthropods. The track rows are oriented either obliquely (angle of 72° to midline), or perpendicularly to the midline of the trackways. The imprints are not connected; the sets of tracks are slightly oblique to the trackway axis, a series of separated metachronal waves that sometimes intersect. Locally, raised rims envelop the imprints (Fig. 7E1, E2). No grooves or ridges are present.
Remarks.—Ichnospecies of Diplichnites are generally characterized by bilaterally symmetric tracks composed of two rows of small imprints, consisting of unorganised series of sigmoidal scratches (Seilacher 2007). Since its initial description by Dawson (1873), Diplichnites has been the subject of extensive discussion and diagnostic revisions. Indeed, its ichnotaxonomy encompasses several species including Diplichnites aenigma Dawson, 1873, Diplichnites triassicus Linck, 1943, Diplichnites cuithensis Briggs et al., 1979, Diplichnites gouldi Gevers in Gevers et al., 1971, and Diplichnites minimus Walter & Gaitzsch, 1988. Diplichnites aenigma and D. cuithensis are considerably broader than the trackways under discussion (Briggs et al. 1979, Chaney et al. 2013; Buckman et al. 2024). The latter is distinguished by an imprint series of at least 23 that correspond to the number of walking appendages of an Arthropleura (Trewin and McNamara 1994). Diplichnites triassicus and D. gouldi are quite similar and sometime regarded as synonyms (Melchor and Cardonatto 2014). However, D. triassicus is applied mostly to relatively narrow trackways having an imprint series of 7 to 9 arranged en echelon (Briggs et al. 1979, 1984; Pollard 1985; Goldring et al. 2005; Pollard et al. 2008). Diplichnites minimus tends to form asymmetric trackways, and it is characterized by the small size of the track rows consisting of highly variable, composite, and poorly defined tracks that differ from those previously described; thus, its inclusion under Diplichnites is dubious (sensu Melchor and Cardonatto 2014). Thus, asymmetric trackways with oblique to chevron like sets of imprints, assigned to Diplichnites since Seilacher (1955), are regarded later as Petalichnus (Osgood 1970; Osgood and Drennen 1975; Rindsberg 1994), Based on morphological features, the specimens described here can be confidently identified as Diplichnites gouldi, which consists of two parallel series of tracks with an approximate spacing of 1 mm.
Stratigraphic and geographic range.—Cambrian–Permian, the North and South Americas, Europe, Asia and Africa.
Ichnogenus Rusophycus Hall, 1852
Type ichnospecies: Fucoides biloba Vanuxem 1842; Silurian, New York, USA (see Fillion and Pickerill 1990; Gibb and Pemberton 2017).
Emended diagnosis.—Short, bilobate and rarely multilobate trace fossils with an oblong, ovate or coffee-bean-shaped outline. Bilaterally symmetric lobes, preserved as a convex hypichnia with a distinct deep or shallow median furrow, or as a concave epichnia with a distinct deep or shallow median ridge. Lobes are parallel or sometimes merged near the posterior, ornamented with oblique to transverse striation or almost smooth surface (after Gibb and Pemberton 2017).
Rusophycus antiatlasensis isp. nov.
Fig. 8.
Zoobank LSID: urn:lsid:zoobank.org:pub:6CE4EBF6-C959-462B-88 E5-8B88E18226B7
Etymology: Derived from the name of the region where it is collected, the eastern Anti-Atlas Mountains of Morocco.
Holotype: ESEFB-LTM-114, series of 11 short bilobate invertebrate trace fossils having an oblong to ovate-shaped outline, preserved in fine-grained sandstones (Fig. 8A1).
Type locality: Jbel Oufilal, Taouz region, Tafilalet Platform, Eastern Anti-Atlas, Morocco.
Type horizon: Uppermost part of Oufilal Formation, Taouz Group, Late Famennian, Devonian.
Material.—Holotype and ESEFB-LTM-115, slab with isolated short bilobate trace fossils with an oblong to ovate outline (Fig. 8B1), from the Late Famennian Oufilal Formation, Taouz Group in Jbel Oufilal, Taouz region, Tafilalet Platform, Eastern Anti-Atlas, Morocco.
Diagnosis.—Short, bilobate trace fossils with an oblong to ovate outline. Bilaterally symmetric lobes, with a distinct cylindrical shallow median ridge/furrow. Lobes are parallel and ornamented with transverse to slightly oblique striae.
Description.—These short, bilobate trace fossils have an oblong to ovate shape; they are well-preserved in convex hyporeliefs and concave epireliefs consisting of two juxtaposed symmetric and parallel lobes (Fig. 8). They occur as individuals or are arranged in population-like assemblages of up to eleven serially repeated traces, with a preferred orientation within the slab. Distances between burrows are not uniform; sometimes the burrows intersect or overlap locally (Fig. 8A1, A4), or occur as traces continuous with Diplichnites (Fig. 8A1, A2, B1, B2). Lobes are characterised by transverse (approximately perpendicular to slightly inclined with an angle of 70–80° to the median ridge/furrow), close-spaced striae, which are deeply to shallowly impressed toward the outer margins (Fig. 8A1, B1). The burrows exhibit carapace imprints within moulds of scratches. The lobes are parallel and well developed, separated entirely by a distinct axial, cylindrical shallow furrow; they extend from the anterior to the posterior end, which are slightly tapered toward both ends in some specimens. The axial cylindrical furrow is smooth and shows no ornamentation or striation on its wall. The overall morphology of these trace fossils is oblong to ovate; on average, they measure 28 mm in length and 15 mm in width. The length/width ratio is 1.86.

Fig. 8. Trace fossil Rusophycus antiatlasensis isp. nov. and associated trace fossils, Paracanthorhaphe isp. (Pa), preserved as convex hyporeliefs in fine-grained sandstone from the Famennian (uppermost Devonian) of the Jebel Aoufilal, eastern Anti-Atlas, Morocco. A. ESEFB-LTM-114. A1, slab displaying a series of eleven short bilobate trace fossils with oblong to ovate outlines; A2, three R. antiatlasensis isp. nov., two preserved in continuity with Diplichnites (Di) trackways; A3, couple R. antiatlasensis isp. nov. juxtaposed; A4, two R. antiatlasensis isp. nov., one extended (left), second in continuuity with Diplichnites trackway (right); A5, interpretive drawing showing the morphology and orientation of the individual traces, as well as their inferred arrangement on the bedding surface. B. ESEFB-LTM-115, short bilobate trace. Rusophycus antiatlasensis isp. nov. in continuity with Diplichnites trackway (B1), with close-up view (B2). Scale bars 20 mm.
Remarks.—The material described herein consists of bilobate ovate, elliptical to coffee-beanshaped trace fossils with the two lobes are parallel and separated by a median furrow or ridge and covered by transverse striae or rugae, occuring as isolated traces or in groups. Thus, all thses features are the main diagnostic features of the ichnogenus Rusophycus (Osgood 1970; Häntzschel 1975; Osgood and Drennen 1975; Alpert 1976; Fillion and Pickerill 1990). Rusophycus is considered as a continuous or an extension trace of Cruziana, and the difference between both ichnotaxa is their length/width ratio, which is greater than 2 for Cruziana and less than 2 for Rusophycus (Fillion and Pickerill 1990). The length/width ratio of the Moroccan specimens is less than 2, which is matching the characteristics of the ichnogenus Rusophycus. The genus was first diagnosed by Hall (1852) in botanical terms as a branched or simple plant fossil before being considered as an animal trace fossil produced by the burrowing activities of trilobites (Dawson 1864; Fillion and Pickerill 1990). It has a widespread geographical distribution as a common Palaeozoic ichnotaxon, recorded from the lower Cambrian to the Triassic, and exhibits great morphological variation.
Therefore, we sort numerous ichnospecies of Rusophycus into three groups: (i) the “R. biloba group”, including Rusophycus biloba Vanuxem, 1842, Rusophycus pudicum Hall, 1852, Rusophycus cryptolithi Osgood, 1970, Rusophycus unilobus Seilacher, 1970, Rusophycus latus Webby, 1983, Rusophycus lavonensis Crimes & Anderson, 1985, Rusophycus crebrus Orłowski, 1992, Rusophycus magnus Orłowski, 1992, Rusophycus versans Schlirf, 2001, and Rusophycus exsilius Stachacz, 2012; this group is characterised by an oval to elliptical outline and a bilobed excavation; the lobes are either entirely or partially separated by a shallow furrow covered in fine or delicate scratches and striations; (ii) the “R. carbonarius group” comprises Rusophycus carbonarius Dawson, 1864, Rusophycus dispar Linnarsson, 1869, and Rusophycus subnotous Gibb et al., 2017, which are characterised by small, oval- to heart-shaped bilobed traces with two symmetric posteriorly tapering lobes and an anterior V-shaped gap; (iii) the “R. carleyi group”, as defined by Gibb et al. (2017) in an attempt to summarise and reduce the number of ichnospecies, includes the following: Rusophycus didymus Salter, 1856, Rusophycus eutendorfensis Linck, 1942, Rusophycus polonicus Orłowski et al., 1970, Rusophycus morgati Baldwin, 1977, Rusophycus carleyi James, 1885, and Rusophycus moyensis Mángano, 2002. The main features of this group include an oval outline and convex hyporelief consisting of two juxtaposed lobes separated by a median furrow opening mesially at the anterior end with oblique, V-shaped ridges; it exhibits trilobite-specific features such as coxal traces, genal spines, pleural impressions, endopodite and exopodite traces.
Other ichnospecies of Rusophycus are characterised by striate or rugose, transversely parallel lobes, which are: Rusophycus pudicus, R. subangulatus, R. clavatus, and R. bilobatus. The latter is defined as being composed of two distinct and symmetric lobes separated by a deep and short longitudinal groove. Rusophycus grenvillensis Billings, 1862, is characterised by a median furrow that does not extend over the entire length of the burrow (Hall 1852). Our specimens differ from all previously reported ichnospecies of Rusophycus by having a median cylindrical ridge/furrow. Unlike the first group, they are covered with transverse to slightly oblique, closely spaced striae, which are deeply to shallowly impressed toward the margins of the trace fossils. It also differs from the group 2 in that the outline is not heart-shaped, but instead has an oblong to elongately elliptical shape, as well as in the shape of the ridge/furrow. Moreover, it differs from the group 3 by the absence of a mesial opening displaying the traces and impressions of the different morphological features of trilobites. Based on its morphological features; bilobate oblong to ovate outline with distinct cylindrical shallow median ridge/furrow, we assign our material to Rusophycus antiatlasensis isp. nov. Thus, R. antiatlasensis isp. nov. has been regarded here as belonging to the “R. carleyi group”.
Stratigraphic and geographic range.—The only occurrence of Rusophycus antiatlasensis isp. nov. is the Late Famennian (Devonian) Oufilal Formation (Taouz Group) from Jbel Oufilal in the Taouz region (Eastern Anti-Atlas, Morocco).
Rusophycus cf. carleyi James, 1885
Fig. 9C, D, E.
Material.—Three slabs with continuous trackways consisting of two parallel rows preserved as convex hyporelief in fine-grained sandstones from the Famennian (uppermost Devonian) of the Jebel Aoufilal area (El Khraouia and Filon 12), eastern Anti-Atlas, Morocco. ESEFB-LTM-116, 117, elliptical bilobate trace fossils (Fig. 9C, D); ESEFB-LTM-118, isolated elliptical bilobate trace fossils (Fig. 9E).
Description.—Bilobate, elliptical to ovate trace fossils preserved as convex hyporelief and concave epirelief, consisting of two lateral convex lobes separated from the anterior of the lobe traces by a depression or median mesial opening and extending posteriorly (Fig. 9C, D, E). The bilobate trace fossils are sometimes recorded at the end or interrupt Diplichnites trackways having two parallel rows of tracks, occur isolated. The median mesial opening is shallowly impressed. The specimens described herein do not exhibit any exopodal, spinal, cephalic or pygidial traces. The only remaining morphological feature is the coxal impressions. The specimens measure approximately 42 mm in average length and 22 mm in average width with a length/width ratio of 1.9.

Fig. 9. Trace fossil Rusophycus cf. carleyi James, 1885, and associated trace fossils, Paracanthorhaphe isp., preserved as convex hyporeliefs in fine-grained sandstone from the Famennian (uppermost Devonian) of the Jebel Aoufilal, eastern Anti-Atlas, Morocco. A. ESEFB-LTM-116, slab displaying continuous trackways, consisting of two parallel rows of impressions ending in an elliptical, bilobate trace fossil (A1), close up (A2). These occur alongside other invertebrate traces. B. In situ slab displaying continuous trackways that also end in elliptical bilobate traces, associated with further invertebrate traces,. C. ESEFB-LTM-117, slab displaying continuous trackways comprising two parallel rows interspersed with elliptical bilobate trace fossils (C1), close up (C2). D. ESEFB-LTM-118, slab displaying continuous trackways consisting of two parallel rows that end in an elliptical bilobate trace fossil (D1), close up (D2). E–G. In situ slabs, isolated elliptical bilobate trace fossil (E), trackways consisting of two parallel rows (F, G). Scale bars 20 mm. Abbreviations: Pa, Planolites isp. Tr, Treptichnus isp.
Remarks.—Following the synonymies suggested by Gibb et al. (2017) of the four ichnospecies of Rusophycus having an ovoid outline, lateral lobes with coxal impressions in the mesial opening occur in Rusophycus polonicus Seilacher, 1970, Rusophycus morgati Baldwin, 1977, Rusophycus carleyi James, 1885, and Rusophycus moyensis Mángano et al., 2002. Rusophycus carleyi has been diagnosed as an “Elliptical shaped trace with two distinct convex hyporelief lateral lobes from anterior to posterior. Mesial opening with subcircular to subrectangular impressions” (Gibb et al. 2010: 277). The trace fossils described herein exhibit characteristics that are similar to those of R. carleyi such as an ovate to elliptical outline, the bilobate depression (comprising two lateral lobes separated by a mesial opening) and two longitudinal series of slightly eroded ridges or coxal traces, which may have been imprinted by the ventral soft body, including the digestive system and appendages (Gutiérrez-Marco et al. 2017). This distinguishes it from other Rusophycus, as it presents coxal traces or imprints of other morphological features that are not observable in our samples because of external factors (Osgood 1970, Seilacher 1970, 1991). These characteristics place it within the “R. carleyi group”. Therefore, we assign the material described herein to Rusophycus cf. carleyi.
Rusophycus isp.
Figs. 7A1, A3, 9A, B.
Material.—Three slabs with long trackways preserved as convex hyporelief in fine-grained sandstones from the Famennian (uppermost Devonian) of the Jebel Aoufilal area (El Khraouia and Filon 12), eastern Anti-Atlas, Morocco. ESEFB-LTM-105, slab with long trackway consisting of the double row of tracks interrupted with a short bilobate burrow (Fig. 7A1, A3); ESEFB-LTM-119 and 120, slabs with continuous trackways with two parallel rows ending with elliptical bilobate trace fossils cooccurring with other invertebrate traces (Fig. 9A, B).
Description.—Short bilobate trace fossils preserved as convex hyporelief depressions with oblong to ovate outlines (Figs. 7A1, A3, 9A, B). Lobes are not well separated and the median furrow is either missing or only faintly preserved. ESEFB-LTM-120 shows an elongate posterior end, which looks like a telson trace at the end of the circular trace (Fig. 9B). ESEFB-LTM-119 bears an oblong trace fossil with scratches at its outer margins (Fig. 9A). The traces measure 28–50 mm in length and 15–22 mm in width, with a length/width ratio of less than 2. Such trace fossils are usually associated mostly as continuous traces alongside a Diplichnites gouldi trackway, even at its end or interrupting it (see Figs. 7A, 9A, B).
Remarks.—The specimens described herein are referred to Rusophycus isp. based on their short oval shape and bilobate character. The quality of the preservation does not allow an assignment at the level of ichnospecies. Seilacher (1970) cautioned ichnologists that narrow Cruziana and Rusophycus has fewer details and is thus more difficult to characterize. We prefer to keep it on open nomenclature.
Associated trace fossils
Arenicolites isp.
Trace fossils recorded (Fig. 7A1, H) as simple pairs of circles (apertures) preserved as convex hyporelief on bedding planes (horizontal section of upper surfaces). Each opening measures about 1–4 mm in diameter. The circles of each pair shows either approximately the same or a slightly differing diameter.
The invertebrate burrows described herein show the characteristic features of the ichnogenus Arenicolites Salter, 1857. The latter consists of simple, vertical, U-shaped burrows with distinct walls, ending in two apertures at the bedding plane or surface (Rindsberg and Kopaska-Merkel 2005). Arenicolites is commonly referred to domichnial behaviour (dwelling burrows) of suspension feeders, e.g., polychaete worms and amphipod crustaceans (Rindsberg and Kopaska-Merkel 2005).
Fustiglyphus isp.
Invertebrate trace fossils (Fig. 7D) characterised by horizontal, straight strings, preserved in convex hyporelief. They appear as narrow cylinders of varying length encircled at irregular or varying intervals by ring-like knots or swellings, which do show neither a bifurcation nor an invagination. Here, we refer tentatively these burrows to Fustiglyphus isp. due to the presence of unbranched swellings in the form of knots (Häntzschel 1975; Stanley and Pickerill 1993). Fustiglyphus Vialov, 1971, has been regarded as being produced by peristaltic movements of organisms such as amphipods, gastropods or holothurians (Osgood 1970; Häntzschel 1975); thus, the swellings have been suggested to represent brood chambers belonging to the recently introduced ethological category sequestrichnia (Stanley and Pickerill 1993; Vallon and Rindsberg 2022).
Lockeia isp.
Horizontal to subhorizontal, symmetric to asymmetric, smooth-walled, unornamented, almond-shaped, elongate, and elliptical burrows (Figs. 5A1, 7A–C, E1, H), which are in some specimens tapered at both ends. The burrows are mainly preserved in convex hyporelief. The trace fossil described herein fits the diagnosis of the ichnogenus Lockeia James, 1879, whose tracemakers are commonly referred to bivalves.
Mammillichnis isp.
Conical to subconical, elevated to flat-topped mound (Fig. 7H) with a convex or concave, semicircular apex and a tent-like tubercle at the centre of the apex. The mound may be encircled by a shallow furrow on the outer circuit. Such mounds been regarded as cubichnia, domichnia, calichnia, fodinichnia or digestichnia, most probably produced by worm-like organisms (Chamberlain 1971), or even praedichnial structures used as traps (Uchman 1998).
Monomorphichnus lineatus Crimes et al., 1977
A group of eight unpaired ridges or grooves, bearing straight striae (Fig. 5A1–A3). These are preserved in convex hyporelief, measuring approximately 10–40 mm in length, 0.2–1 mm in width. The spacing between successive ridges is 3–6 mm. These trace fossils match the morphological features of Monomorphichnus Crimes, 1970, which are a series of parallel, straight and sigmoidal ridges or grooves. Monomorphichnus comprises five ichnospecies: Monomorphichnus lineatus Crimes et al., 1977, which has single ridges; Monomorphichnus bilinearis Crimes, 1970, with a paired associated ridge, Monomorphichnus multilineatus Alpert, 1976, which has deep central ridges; Monomorphichnus intersectus Fillion & Pickerill, 1990, is characterised by intersecting dig marks; and Monomorphichnus pectenensis Legg, 1985, with striae between ridges. The specimens described herein matchs the diagnosis of M. lineatus, which can be distinguished from M. bilinearis the most similar ichnotaxon by the absence of paired ridges. Monomorphichnus was originally described by Crimes (1970) and attributed to trilobites that got caught up in a current and raked the sediment surface with their endopodite claws. Therefore, the fact that the ridges are repeated laterally reveals that these traces may have been formed by the swimming and grazing movements of the trilobite, where each groove probably resulted from contact with the surface by a single- or multi-clawed limb, or even just one claw (Crimes et al. 1977). By measuring the ridges, we can estimate that the animal was approximately 40 mm long on average and not much larger.
Planolites isp.
Planolites consists of simple, lined, unbranched, cylindrical, horizontal to gently oblique simple burrows (Figs. 5A1, 7A1, H) that have a smooth and structureless wall, preserved in endichnial to hypichnial full relief. The burrow width measures approximately 5–10 mm. Their fillings are structureless, suggesting that they were actively filled. Most described burrows show a distinctively lined burrow; these burrows are assigned in open nomenclature to Planolites isp. The record of Planolites Nicholson, 1873, extends through the whole Phanerozoic, which could imply that a wide range of groups may have been potential producers. It is interpreted as exhibiting both feeding (fodinichnial) and dwelling (domichnial) behaviour (Pemberton and Frey 1982).
Paracanthorhaphe isp.
These thin, branched, smooth burrows (Fig. 8A1–A3) are preserved as positive hyporelief, with the fill resembling the surrounding rock. The traces are consistently 1–2 mm in diameter and extend for several meters. These burrows display a well-organized branching pattern with primary branches that then branch once or twice. Paracanthorhaphe Wu, 1985, has been interpreted as complex feeding structures (Buatois et al. 1996).
Treptichnus isp.
These horizontals to subhorizontal, straight to slightly curved systems of rows of short, twig-like burrows are preserved in positive hyporelief (Figs. 7A1, A2, F, H, 9A1). They are arranged as segments branching off in regular, locally irregular intervals, at low angles resembling zigzag projections. These burrows measure 10–22 mm in length and approx. 1–2 mm in width. They are referred to Treptichnus isp. in open nomenclature. The producer has been interpreted as a surface detritus feeder (Jensen 1997), deposit feeder (Seilacher 1955), predator (Vannier et al. 2010) and undermat miner (Seilacher 2007). Cambrian and Devonian examples were probably made by priapulid worms (Vannier et al. 2010).
Undichna isp.
Two nonparallel pairs of horizontally undulating, sinusoidal vertebrate trace fossil is preserved in convex hyporelief (Fig. 6A). These characteristics represent the diagnosis features of the ichnogenus Undichna Anderson, 1976. This ichnotaxon has been already described from the same stratigraphic unit of the same locality (Klug et al. 2021).
Unidentified trackways
Series of bilaterally symmetric, lunate trace fossils are preserved in convex hyporelief (Fig. 6A). Each trace measures up to 60 mm in length and 15 mm in width. The length of the whole trackway is 200 mm. The spacing between two successive imprints is consistent, approximately 4 cm. The lunate structure reminds of the ichnogenus Crescentichnus, but the trackway pattern and the well-organised series of traces has never been observed in Crescentichnus to our knowledge. A similar trace fossil from the lower Cambrian of Sweden is preserved in concave epirelief and has been referred to cf. Protovirgularia isp. (McLoughlin et al. 2021: fig. 11c). The rarity of specimens and the quality of the preservation do not yet allow the introduction of a new ichnotaxon. These specimens are subject of separate detailed ichnotaxonomic analysis.
Discussion
Tracemakers and behaviour.—Cruziana is one of the most studied ichnotaxa due to its importance, abundance and usefulness in biostratigraphy, together with its relevance to tracemaker behaviour (see Seilacher 2007). It is recorded from the early Cambrian to Mesozoic, including Triassic sediments after the extinction of trilobites (e.g., Bromley and Asgaard 1979; Hminna et al. 2020). Thus, the Palaeozoic Cruziana is usually referred to trilobites as possible tracemakers, but other arthropods (and even brachiopods) may well have produced it (Seilacher 1985, 2007). Cruziana has been regarded as documenting a locomotion activity (repichnia) as an animal moved across the substrate surface, with grazing traces (pascichnia) or ploughing horizontally within the sediments of detritus-feeding organisms (Goldring 1985; Seilacher 1985, 2007; Neto de Carvalho 2006). Increasingly, authors interpret Cruziana as the result of feeding in addition to locomotion (Kesidis et al. 2018). Such behaviour occurs while deposit- or detritus-feeders with appendages move across and through sediments searching for food (Goldring 1985; Seilacher 1985, 2007; Neto de Carvalho 2006). Cruziana is a kind of bilobed furrow characterised by distinctive striae, which are the main ichnotaxonomic features used to distinguish its ichnospecies.
The slightly oblique, blunt-ended transverse scratches in Cruziana lobosa could be more related to anatomically controlled or behavioural features than mode of preservation. The central area of the furrow is characterised by V-shaped, blunt-ended scratch traces, which were presumably made by the endopodite walking legs digging down, inwards and backwards (Seilacher 1955; Crimes 1970; Birkenmajer and Bruton 1971). The V-shaped traces can extend to the outer edges of the whole trace on each side. The lateral ridges are mud grooves that correspond to series of wisp-like lineation, bounded on both lobes, and probably excavated by the pleurae (Seilacher 1970). However, the lateral groove in some specimens is formed at the extreme outer edge of the trilobite’s carapace beyond the limit of the pleurae (Crimes 1975). These lateral ridges may have been caused by trilobites dragging their genal spines excavating through the sediment (cf. Crimes 1968, 1970, 1975). Thus, Caster (1938) documented lateral traces made by genal spines by modern and ancient horseshoe crab trackways. Alternatively, they were caused by the endopodites dragging non-functional legs outside the area where the functional endopodites were digging. The ~90° preferred angle of the burrows crisscrossing might indicate that the tracemakers could detect previous burrows at a distance, and preferred to minimize the amount of rebioturbated sediment (Rindsberg and Martin 2003).
The diversity of trilobites during the Late Devonian was much reduced compared to that of the Early Devonian, which is related to the several global events such as Kellwasser, Condroz-Enkeberg, Annulata, Dasberg, and Hangenberg events (Walliser 1996; Lerosey-Aubril and Feist 2012; Bault et al. 2021). Only two trilobite orders survived into the latest Devonian (Famennian) through the Kellwasser Event: Phacopida are represented by eight genera and the Proetida by eleven (Bault et al. 2021 and references therein). Phacopids were represented by taxa about 10 mm wide and proetids were slightly larger. Accordingly, we suggest phacopids as potential tracemakers of studied C. lobosa based on morphometric measurements of latest Devonian trilobite groups.
The ichnogenus Diplichnites has been regarded as having recorded a locomotion behaviour of trilobites or other many-legged arthropods (e.g., millipedes, centipedes, pill bugs among others), through metachronal waves of motion passing along the body, with a smaller distance than the body length between successive waves for continued support. This ichnogenus has been reported from Cambrian to Triassic sediments from in both marine and nonmarine settings (Seilacher 1955; Bromley and Asgaard 1979; Pickerill 1992; Seilacher 2007). The constant spacing of imprints reflects this origin from multiple, oppositely paired, very small walking limbs (Gevers et al. 1971); however, estimating the number of legs is difficult to impossible due to overlap and the identical size and shape of imprints. Diplichnites gouldi has been reported from marine and nonmarine deposits from Ordovician to Permian rocks (Braddy 1995; Johnson et al. 1994). Among all the arthropods previously listed, the trilobites were likely the most able to sidle, i.e., to move at an angle to the body axis, resulting in separated metachronal waves that produced a set of footprints on the two parallel rows (Seilacher 2007). Furthermore, the connection of Diplichnites and Rusophycus in the same trace provides further evidence that trilobites were primarily responsible for producing these traces from the uppermost Devonian of Morocco (Figs. 6A, B1, B4, 7A1, A3, F). No morphological changes were observed in the tracks within the same samples resulting from homopodous producers; therefore, any differences in trackway shape observed between the samples could be due to substrate moisture, cohesiveness or animal behaviour (Seilacher 2007; Trewin 1995). The Diplichnites described herein exhibited variable ranges of size, thus the morphometric measurements suggest proetids or phacopids as the potential tracemakers of Diplichnites gouldi from the Anti-Atlas.
The Palaeozoic records of Rusophycus are commonly referred to trilobites as the most likely tracemakers. This is because, in exceptional cases, the preserved body fossil of the tracemaker has been found in situ, or clear traces of trilobite-specific morphological features such as size, shape, cephalic border and genal spines have been preserved (Osgood 1970; Campbell 1975). Therefore, Rusophycus is interpreted as a resting trace (cubichnion), revealing the complete outline of the producing organism body (Osgood 1975). These traces may exhibit morphological characteristics, such as traces made by coxal and genal spines, as well as impressions of the cephalic and pygidial margins, while burrowing and digging within the sediment (Brandt et al. 1995; Osgood 1975). Thus, these trilobite traces received their bilobate shape through their filter-feeding mechanism, which formed a ventral feeding chamber that brushed the sediment towards the median line, where it was washed away by some current. Consequently, the transverse striation and ornamentation along its length are consistent with the proposition of a feeding chamber (Seilacher 1985). Moreover, Rusophycus might also have served as an incubation site (egg depository) for trilobites. Trilobites may have carried spawn attached to their swimming feet and used shallow waters for incubation (Fenton and Fenton 1937). Alternatively, to avoid predators or attempt to remain hydrated, the trilobite dug shallow burrows quickly where it covered its body with a thin layer of sediment (Dawson 1864; Fenton and Fenton 1937; Osgood 1975). According to Osgood (1970), this behaviour involved a combination of vertical and slight horizontal and/or lateral movements. It occurred when the organism rested briefly while moving horizontally across the substrate.
Rusophycus antiatlasensis isp. nov. is a bilobate trace fossil with an oblong to ovate outline, ornamented with transverse to slightly oblique striations, separated by a cylindrical shallow median furrow or ridge (Fig. 10D–B). The latter could be an impression of the digestive tract, which generally had a tubular structure, which started with the slit-like mouth leading to a short, narrow oesophagus connecting to a J-shaped crop and a long, narrow intestine that extends to the posterior most part of the trunk (El Albani et al. 2024: fig. 2C, D). The transverse to slightly oblique striation in both lobes could have resulted from burrowing and digging activities by the legs; thus, while resting, it could have formed impressions of the exopodites or even the endopodites. Proetids and/ or phacopids have nearly the same dorsal shield outline, which is mostly oblong and thus, both groups could be referred as the most likely tracemakers of R. antiatlasensis isp. nov. They are arranged in population assemblages of up to eleven serially repeated traces, with an evident preferential orientation within the slab, which is regarded as reflecting gregarious behaviour of trilobites. The latter has previously been recorded from the Lower Ordovician of Morocco; it is regarded as mechanical stimulation linked to motion and touch sensors or seasonal mating behaviour linked to migration of adults to spawning grounds (Vannier et al. 2019). Knowing that most of trilobites were highly visual creatures that probably also had a well-developed chemical sense, the gregarious behaviour could be guided by one of these senses. Rusophycus carleyi has been regarded as a biotic interaction rather than a simple resting trace (Brandt et al. 1995). Its range to the Upper Devonian is questionable. Gibb et al. (2010) suggested asaphid trilobites as the most likely tracemakers of R. carleyi, which cannot be the case in the latest Devonian specimens because asaphids were already extinct by that time.

Fig. 10. The Diplichnites–Rusophycus association. A. The association of Diplichnites–Rusophycus is preserved on the surfaces of bedding planes. PMO235.006, housed at Natural History Museum of University of Oslo, Credit photograph to Hans Arne Nakrem) (A1). Close-up (A2) showing the morphological details of the trackways and resting traces (Rusophycus), which exhibit bilobed impressions and paired tracks (Diplichnites). B. A continuous trackway showing the transition from a Diplichnites locomotion trace to a Rusophycus resting trace. This suggests a behavioural link between the resting and walking phases of the organism that made the tracks. C, D. Diverse Rusophycus antiatlasensis nov. isp. specimens displaying the extramorphological variations with the median tube-like structure. C. ESEFBLTM-121 (C1). Close-up (C2). D. ESEFBLTM-122.
Most of the Diplichnites gouldi trackways described herein are associated with Rusophycus antiatlasensis isp. nov. or Rusophycus isp., at the beginning, middle or end of the trackways (Fig. 10), indicating that they were produced by the same animal. In this case, the makers of the combined Diplichnites–Rusophycus moved forward, assuming a resting position before moving forward again; this operation was repeated in the same trackway and recorded at least in three specimens (Fig. 10). This behaviour can be interpreted as the producer being on the lookout and stalking for its prey; it took a resting position so that its prey does not sense its presence. We note that most of the surfaces with combined Diplichnites–Rusophycus bear other invertebrate trace fossils as well, such as Arenicolites isp., cf. Fustiglyphus isp., Lockeia isp., Mammillichnis isp., Planolites isp., Paracanthorhaphe isp., and Treptichnus isp. We speculate that the tracemakers of Diplichnites–Rusphycus were hunting for the makers of other ichnotaxa. Based on the association of diverse trilobite trace fossils cooccurring with other invertebrate burrows, Martinsson (1965), Bergström (1973), Jensen (1990), and Brandt et al. (1995) discussed the predatory behaviour of trilobites. These cooccurrences have been interpreted as a result of the trilobite locating the prey from above and behind, active pursuit, and digging to a depth considerably above or greater than the lowest part of the prey burrow (Bergström 1973; Jensen 1990). Alternatively, this trace may have formed when the Diplichnites–Rusophycus tracemaker sensed or observed a predator while moving on the substrate; then took up a resting or hiding position and covered its body with sediment to hide from predators. Indeed, the same stratigraphic unit yielded several surfaces with cooccurrences of Diplichnites, Cruziana, and Rusophycus alongside Undichna presumably made by predatory chondrichthyan fish in the same area (Figs. 6A1, 10C) (Klug et al. 2021: figs. 4, 5).
Palaeoenvironment and palaeoecology.—The highly diverse ichnoassemblage reported herein consists mainly of trace fossils of trilobites and associated animals; the assemblage includes pascichnia (Cruziana lobosa), repichnia (Diplichnites gouldi, Mammillichnis isp., Undichna isp.), cubichnia (Rusophycus antiatlasensis isp. nov., Rusophycus cf. carleyi, Lockeia isp.), fodinichnia (Fustiglyphus isp., Paracanthorhaphe isp., Treptichnus isp.), and domichnia (Arenicolites isp., Planolites isp.) of various animal groups. These ichnotaxa are ascribed to the archetypal Cruziana ichnofacies, which is typically referred to shallow-marine environments from the lower to upper shoreface transition to upper offshore in a continental shelf setting (e.g., Pemberton et al. 2001; Seilacher 2007). The predominance of horizontal invertebrate trace fossils commonly indicates low hydrodynamic energy settings in moderately warm and shallow open-marine environments with stable substrate conditions and regular organic matter sources, particularly in subtidal settings below fair-weather wave base but above storm wave base (i.e., MacEachern et al. 2012). The occurrence of the vertical domichnial burrows (Arenicolites isp.) points to short phases of moderate to high hydrodynamic energy (Ekdale 1985). Fine-grained sandstones with wave-induced current ripples indicate a very shallow environment. The excellent preservation of this ichnofauna may be linked with episodic low-oxygen conditions linked in turn to the environmental perturbations of the Hangenberg Event, which inhibited bioturbation and thereby hindered the destruction of the ichnofossils.
Conclusions
The uppermost Devonian sedimentary succession of the eastern Anti-Atlas of Morocco contains a diverse but still poorly documented invertebrate and vertebrate ichnoassemblage representing diverse behaviours, which are: pascichnia (Cruziana lobosa), repichnia (Diplichnites gouldi, Mammillichnis isp., Undichna isp.), cubichnia (Rusophycus antiatlasensis isp. nov., Rusophycus cf. carleyi, Lockeia isp.), fodinichnia (Fustiglyphus isp., Paracanthorhaphe isp., Treptichnus isp.), and domichnia (Arenicolites isp., Planolites isp.). Several of these ichnogenera (Cruziana, Diplichnites, Monomorphichnus, Rusophycus) are referred here to proetid and phacopid trilobites, while others were probably made by polychaete worms, amphipods, crustaceans, gastropods or holothurians, bivalves, and fishes. They are referred to the archetypal Cruziana ichnofacies, indicating a shallow-marine environment with low hydrodynamic-energy settings, stable substrate conditions and regular organic matter sources. The structure of Cruziana lobosa could be related more to anatomically controlled features or behavioural than to preservational conditions. Diplichnites gouldi was produced by homopodous trilobites. Rusophycus antiatlasensis isp. nov. is introduced here for oblong to ovate Rusophycus having a cylindrical median furrow/ridge. Serially repeated assemblages of multiple Rusophycus may reflect a gregarious behaviour of trilobites, most probably related to migration of adults for mating and spawning, or alternatively to locally abundant prey. The association of Diplichnites gouldi, R. antiatlasensis isp. nov., and Rusophycus isp. indicates a predation strategy through stalking prey, as interrupted by hiding in the sediment from predators such as fishes (documented by abundant Undichna isp.).
Given its potentially pivotal role for the understanding of the evolution of Late Devonian ecosystems, just after a series of events and tightly linked to the global mass extinctions of the Hangenberg Event that influenced the marine life on earth profoundly, fossil exploration for both trace and body fossils in eastern Anti-Atlas should be intensified.
Acknowledgements
We warmly thank Moha Mezzan and Ali Karaoui (both El Khraouia, Merzouga, Morocco) for providing some specimens and assisting in the fieldtrips. We thank also Hans A. Nakrem (Natural History Museum, University of Oslo, Oslo, Norway) and Maximo A. Rojo (Oslo, Norway) for providing photographs of Diplichnites–Rusophycus. The reviewers Andrew K. Rindsberg (University of West Alabama, Livingston, USA) and anonymous reviewer are acknowledged for their constructive comments and suggestions that improved the manuscript greatly. CK acknowledges the financial support of the Swiss National Science Foundation (SNSF) for funding our work (grants nr.953 200021_169627, 200020_184894, 205320_215642).
Editor: Krzysztof Hryniewicz
References
Ait Daoud, M., Essalhi, A., Essalhi, M., and Toummite, A. 2019. The role of Variscan shortening in the control of mineralization deposition in Tadaout-Tizi N’rsas mining district (eastern Anti-Atlas, Morocco). Bulletin of The Mineral Research and Exploration 161: 13–32. Crossref
Alpert, S.P. 1976. Planolites and Skolithos from the upper Precambrian–Lower Cambrian White-Inyo Mountains, California. Journal of Paleontology 49: 508–521.
Álvaro, J.J., Aretz, M., Benharref, M., Hibti, M., Pouclet, A., El Hadi, H., Koukaya, A., Ettachfini, M., and Boudad, L. 2014. Carte Géologique du Maroc au 1/50 000, feuille de Tawz. Mémoire explicatif. Notes et Mémoires du Service géologique du Maroc 551bis: 1–128.
Anderson, A. 1976. Fish trail from the Early Permian of South Africa. Palaeontology 19: 397–409.
Baidder, L., Michard, A., Soulaimani, A., Fekkak, A., Eddebbi, A., Rjimati, E.-C., and Raddi, Y. 2016. Fold interference pattern in thick-skinned tectonics; a case study from the external Variscan belt of eastern Anti-Atlas, Morocco. Journal of African Earth Sciences 119: 204–225. Crossref
Baidder, L., Raddi, Y., Tahiri, M., and Michard, A. 2008. Devonian extension of the Pan-African crust north of the West African Craton and its bearing on the Variscan foreland deformation: evidence from eastern Anti-Atlas (Morocco). In: N. Ennih and J.P. Liégeois (eds.), The Boundaries of the West African Craton. Geological Society, London, Special Publications 297: 453–465. Crossref
Baldwin, C. T. 1977. Rusophycus morgati: an asaphid produced trace fossil from the Cambro-Ordovician of Brittany and northwest Spain. Journal of Palaeontology 51: 411–425.
Bault, V., Crônier, C., Allaire, N., and Monnet, C. 2021. Trilobite biodiversity trends in the Devonian of North Africa. Palaeogeography, Palaeoclimatology, Palaeoecology 565: 110208. Crossref
Bel Haouz, W., Lagnaoui, A., Klug, C., Baidder, L., and Maanan, M. 2026. Middle Devonian ichnofossils from Hamar Laghdad (eastern Anti-Atlas, Morocco). Swiss Journal of Palaeontology 145: 403–431. Crossref
Bergström, J. 1973. Organization, life, and systematics of trilobites. Fossils and Strata 2: 1–69. Crossref
Billings, E. 1862. New species of fossils from different parts of the Lower, Middle, and Upper Silurian rocks of Canada. Geological Survey of Canada, Paleozoic Fossils 1 (4): 96–168.
Birkenmajer, K. and Bruton, D.L. 1971. Some trilobite resting and crawling traces. Lethaia 4: 303–319. Crossref
Braddy, S.J. 1995. Coconino Sandstone (Lower Permian), northern Arizona. Early Permian Footprints and Facies. New Mexico Museum of Natural History and Science Bulletin 6: 1–219.
Brandt, D.S., Meyer, D.L., and Lask, P.B. 1995. Isotelus (Trilobita) “hunting burrow” from Upper Ordovician strata, Ohio. Journal of Palaeontology 69:1079–1083. Crossref
Briggs, D.E.G., Plint, A.G., and Pickerill, R.K. 1984. Arthropleura trails from the Westphalian of eastern Canada. Palaeontology 27: 843–55.
Briggs, D.E.G., Rolfe, W.D.I., and Brannan, J. 1979. A giant myriapod trail from the Namurian of Arran, Scotland. Palaeontology 22: 273–291.
Bromley, R. and Asgaard, U. 1979. Triassic freshwater ichnocoenoses from Carlsberg Fjord, east Greenland. Palaeogeography, Palaeoclimatology, Palaeoecology 28: 39–80. Crossref
Buatois, L.A, Mángano,
M.G., Wu, X., and Zhang, G. 1996. Trace fossils from Jurassic
lacustrine turbidites of the Anyao Formation (central China) and their
environmental and evolutionary significance. Ichnos
4: 287–303. Crossref
Buckman, J.O., Cuthbert S.J., and Polson, P.G. 2024. Arthropleura trackway (Diplichnites cuithensis) from the Carboniferous, Serpukhovian, Limestone Coal Formation, Clackmannan Group, Linn Park, Glasgow. Scottish Journal of Geology 60 (1): sjg2021-019. Crossref
Campbell, K.S.W. 1975. The functional morphology of Cryptolithus. Fossils and Strata 4: 65–86. Crossref
Caster, K.E. 1938. A restudy of the tracks of Paramphibius. Journal of Paleontology 12: 3–60.
Chamberlain, C.K. 1971. Morphology and ethology of trace fossils from the Ouachita Mountains, southeast Oklahoma. Journal of Paleontology 45: 212–246.
Chaney, D.S., Lucas, S.G., and Elrick, S. 2013. New occurrence of an arthropleurid trackway from the Lower Permian of Utah. In: S.G. Lucas, W.A. DiMichele, J.E. Barrick, J.W. Schneider, and J.A. Spielmann (eds.), The Carboniferous–Permian Transition. New Mexico Museum of Natural History and Science, Bulletin 60: 64–65.
Clariond, L. 1934a. Sur le Dévonian du Tafilalt et du Maroc. Compte rendu sommaire des séances de la Société géologique de France 1934: 5–11.
Clariond, L. 1934b. apropos d’une coupe de la région d’Erfoud. Compte rendu sommaire des séances de la Société géologique de France 1934: 223–224.
Crimes, T.P. 1968. Cruziana: a stratigraphically useful trace fossil. Geological Magazine 105: 360–364. Crossref
Crimes, T.P. 1970. Trilobite tracks and other trace fossils from the Upper Cambrian of North Wales. Geological Journal 7: 47–68. Crossref
Crimes, T.P. 1975. The production and preservation of trilobite resting and furrowing traces. Lethaia 8: 35–48. Crossref
Crimes, T.P. and Anderson, M.M. 1985. Trace fossils from the late Precambrian–Early Cambrian strata of southeastern Newfoundland (Canada): temporal and environmental implications. Journal of Paleontology 59: 310–343.
Crimes, T.P., Legg, I., Marcos, A., and Arboleya, M. 1977. ?Late Precambrian–low Lower Cambrian trace fossils from Spain. In: T.P. Crimes and J.C. Harper (eds.), Trace Fossils 2. Geological Journal, Special Issue 9: 91–138.
Dawson, J.W. 1864. On the fossils of the genus Rusophycus. The Canadian Naturalist and Geologist 1: 363367.
Dawson, J.W. 1873. Impressions and footprints of aquatic animals and imitative markings on Carboniferous rocks. American Journal of Science 105: 16–24. Crossref
De Baets, K., Klug, C., and Plusquellec, Y. 2010. Zlíchovian faunas with early ammonoids from Morocco and their use for the correlation between the eastern Anti-Atlas and the western Dra Valley. Bulletin of Geosciences 85: 317–352. Crossref
D’Orbigny, A.C.V. 1842. Voyage dans l’Amérique Méridionale, Tome Troisieme, 4. Partie, Paléontologie. 188 pp. Pitois-Levrault et Levrault, Paris.
Döring, S. and Kazmierczak, M. 2001. Stratigraphy, geometry, and facies of a Middle Devonian ramp-to-basin transect (Eastern Anti-Atlas, SE Morocco). Facies 44: 137–150. Crossref
Ekdale, A.A. 1985. Trace fossils and mid-Cretaceous anoxic events in the Atlantic Ocean. SEPM Special Publication 35: 333–342. Crossref
El Albani, A., Mazurier, A., Edgecombe, G.D., Azizi, A., El Bakhouch, A., Berks, H.O., Bouougri, E.H., Chraiki, I., Donoghue, P.C.J., Fontaine, C., Gaines, R.R., Ghnahalla, M., Meunier, A., Trentesaux, A., and Paterson, J.R. 2024. Rapid volcanic ash entombment reveals the 3D anatomy of Cambrian trilobites. Science 384: 1429–1435. Crossref
Fenton, C.L. and Fenton, M.A. 1937. Trilobite “nests” and feeding burrows. The American Midland Naturalist 18: 446–451. Crossref
Fillion, D. and Pickerill, R.K. 1990. Ichnology of the Upper Cambrian? to Lower Ordovician Bell Island and Wabana Group so eastern Newfoundland, Canada. Palaeontographica Canadiana 7: 1–119.
Frey, L., Naglik, C., Hofmann, R., Schemm-Gregory, M., Frýda, J., Kröger, B., Taylor, P.D., Wilson, M.A., and Klug, C. 2013. Diversity and palaeoecology of Early Devonian invertebrate associations in the Tafilalt (Anti-Atlas, Morocco). Bulletin of Geosciences 89: 75–112. Crossref
Frey, L., Pohle, A., Rücklin, M., and Klug, C. 2019: Fossil-Lagerstätten and preservation of vertebrates and invertebrates from the Devonian of Morocco (eastern Anti-Atlas). Lethaia 53: 242–266. Crossref
Gevers, T.W., Frakes, L.A., Edwards, L.N., and Marzolf, J. E. 1971. Trace fossils in the lower Beacon sediments (Devonian), Darwin Mountains, Southern Victoria Land, Antarctica. Journal of Palaeontology 45: 81–94.
Gibb, S. and Pemberton, S.G. 2017. Rusophycus by any other name is still Rusophycus carleyi. Ichnos 24: 19–26. Crossref
Gibb, S., Chatterton, B.D.E., and Gingras, M.K. 2010. Rusophycus carleyi (James, 1885), trace fossils from the Lower Ordovician of Southern Morocco, and the trilobites that made them. Ichnos 17: 271–283. Crossref
Gibb, S., Pemberton S.G., and Chatterton, B.D.E. 2017. Arthropod trace fossils of the upper Lower Cambrian Gog Group, southern Rocky Mountains of Canada. Ichnos 24: 91–123. Crossref
Goldring, R. 1985. The formation of the trace fossil Cruziana. Geological Magazine 122: 65–72. Crossref
Goldring, R., Pollard, J.E., and Radley, J.D. 2005. Trace fossils and pseudofossils from the Wealden strata (non-marine Lower Cretaceous) of southern England. Cretaceous Research 26: 665–685. Crossref
Gutiérrez-Marco, J.C., García-Bellido, D.C., Rábano, I., and Sá, A.A. 2017. Digestive and appendicular soft-parts, with behavioural implications, in a large Ordovician trilobite from the Fezouata Lagerstätte, Morocco. Scientific Reports 7 (1): 39728. Crossref
Hall, J. 1852. Palaeontology of New York, Vol. 2. 362 pp. Printed by C. Van Benthuysen, Albany.
Häntzschel, W. 1975. Trace fossils and problematica. In: R.C. Moore (ed.), Treatise on Invertebrate Palaeontology, Part W. 269 pp. Geological Society of America, New York and University of Kansas Press, Lawrence.
Hminna, A., Lagnaoui, A., Zouheir, T., Saber, H., and Schneider, J.W. 2020. Late Triassic ichnoassemblage from a playa-lake system of the Coastal Meseta, Morocco: Palaeoenvironmental and palaeoecological implications. Journal of African Earth Sciences 172: 103995. Crossref
Hüneke, H., Gibb, M.A., Mayer, O., Kniest, J.F., Mehlhorn, P., Gibb, L.M., and Baidder, L. 2023. Bioclastic bottom‐current deposits of a Devonian contourite terrace: facies variability and depositional architecture (Tafilalt Platform, Morocco). Sedimentology 70: 1413–1471. Crossref
James, J.F. 1885. The fucoids of the Cincinnati Group, Part. 2. Journal of the Cincinnati Society of Natural History 7: 151–166.
James, U.P. 1879. Descriptions of newly discovered species of fossils and remarks on others, from the lower and upper Silurian rocks of Ohio. Paleontologist, Cincinnati 3: 17–24.
Jensen, S. 1990. Predation by early Cambrian trilobites on infaunal worms —evidence from the Swedish Mickwitzia Sandstone. Lethaia 23: 29–42. Crossref
Jensen, S. 1997. Trace fossils from the Lower Cambrian Mickwitzia sandstone, south-central Sweden. Fossils and Strata 42: 1–110. Crossref
Jobbins, M., Haug, C., and Klug, C. 2020. Famennian thylacocephalans from Morocco and their role in Late Devonian foodwebs. Scientific Reports 10 (5129): 1–9. Crossref
Johnson, E.W., Briggs, D.E.G., Suthren, R.J., Wright, J.L., and Tunnicliffe, S.P. 1994. Non-marine arthropod traces from the subaerial Ordovician Borrowdale Volcanic Group, English Lake District. Geological Magazine 131: 395–406. Crossref
Kaufmann, B. 1998. Facies, stratigraphy and diagenesis of Middle Devonian reef- and mud-mounds in the Mader (eastern Anti-Atlas, Morocco). Acta Geologica Polonica 48: 43–106.
Kesidis, G., Budd, G.E., and Jensen, S. 2019. An intermittent mode of formation for the trace fossil Cruziana as a serial repetition of Rusophycus: the case of Cruziana tenella (Linnarsson 1871). Lethaia 52: 133–148. Crossref
Klug, C. and Hoffmann, R. 2018. Trace fossils of Actiniaria (Conichnus) from the Early Devonian of Morocco. In: C. Klug and D. Korn, (eds.), Palaeontology of the Devonian of Hamar Laghdad (Tafilalt, Morocco). Special Volume Honouring Jobst Wendt. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 290: 65–74. Crossref
Klug, C. and Pohle, A. 2018. The eastern Amessoui Syncline—a hotspot for Silurian to Carboniferous cephalopod research. Münstersche Forschungen zur Geologie und Paläontologie 110: 244–260.
Klug, C., Kröger, B., Rücklin, M., Korn, D., Schemm-Gregory, M., De Baets, K., and Mapes, R.H. 2008. Ecological change during the early Emsian (Devonian) in the Tafilalt (Morocco), the origin of the Ammonoidea, and the first African pyrgocystid edrioasteroids, machaerids and phyllocarids. Palaeontographica Abteilung A 283 (4–6): 83–176. Crossref
Klug, C., Samankassou, E., Pohle, A., De Baets, K., Franchi, F., and Korn, D. 2018. Oases of biodiversity: Early Devonian palaeoecology at Hamar Laghdad, Morocco. In: C. Klug, and D. Korn (eds.), Palaeontology of the Devonian of Hamar Laghdad (Tafilalt, Morocco). Special Volume Honouring Jobst Wendt. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 290: 9–48. Crossref
Klug, C., Lagnaoui, A., Jobbins, M., Bel Haouz, W., and Najih, A. 2021. The swimming trace Undichna from the latest Devonian Hangenberg Sandstone equivalent of Morocco. Swiss Journal of Palaeontology 140: 19. Crossref
Lagnaoui, A., Bel Haouz, W., and Najih, A. 2019. Trackway and resting traces of trilobites with associated invertebrate ichnofossils from the Early Carboniferous of the Al Atrous region (Eastern Anti-Atlas, Morocco). In: D.K. Nurgaliev (ed.), Kazan Golovkinsky Stratigraphic Meeting, 2019—Late Paleozoic Sedimentary Earth Systems: Stratigraphy, Geochronology, Petroleum Resources—Fifth All-Russian Conference “Upper Paleozoic of Russia”. September 24–28, 2019, Kazan, Russia. Abstract Volume, 150–151. KFU Publishing House, Kazan
Legg, I.C. 1985. Trace fossils from a middle Cambrian deltaic sequence, northern Spain. In: H.A. Curran (ed.), Biogenic Structures: Their Use in Interpreting Depositional Environments. Society of Economic Paleontologists and Mineralogists Special Publications 35: 151–165. Crossref
Lerosey-Aubril, R. and Feist, R. 2012. Quantitative approach to diversity and decline in late Palaeozoic trilobites. In: J.A. Talent (ed.), Global Biodiversity, Extinction Intervals and Biogeographic Perturbations through Time, 535–555. Springer Science, Berlin. Crossref
Letsch, D., Large, S.J.E., Bernasconi, S.M., Klug, C., Blattmann, T.M., Winkler, W., and von Quadt, A. 2019. Northwest Africa’s Ediacaran to early Cambrian fossil record, its oldest metazoans and age constraints for the basal Taroudant Group (Morocco). Precambrian Research 320: 438–453. Crossref
Linck, O. 1942. Die Spur Isopodichnus. Senckenbergiana 25: 232–255.
Linck, O. 1943. Die Buntsandstein-Kleinfahrten von Nagold (Limulidichnulus nagoldensis n.g. n.s., Merostomichnites triassicus n. sp.). Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Monatshefte, Abteilung B 1943: 9–27.
Linnarsson J.G.O. 1869. On some fos sils found in the Eophyton sandstone at Lugnas in Sweden. Geological Magazine 6: 393–406.
Lubeseder, S., Rath, J.,
Rücklin, M., and Messbacher, R. 2010. Controls on Devonian
hemi-pelagic limestone deposition analysed on cephalopod ridge to
slope sections, Eastern Anti-Atlas, Morocco. Facies
56: 295–315. Crossref
MacEachern, J.A., Bann, K.L., Gingras, M.K., Zonneveld, J.P., Dashtgard, S.E., and Pemberton, S.G. 2012. The ichnofacies paradigm. In: D. Knaust and R.G. Bromley (eds.), Trace Fossils as Indicators of Sedimentary Environments. Developments in Sedimentology 64: 103–138. Crossref
Mángano, M.G., Buatois, L.A., and Muñiz-Guinea, F. 2002. Rusophycus moyensis n. isp. en la transición Cámbrica–Tremadociana del noroeste Argentino: implicancias palaeoambientales y bioestratigráficas. Revista Brasileira de Palaeontologia 4: 35–44.
Martinsson, A. 1965. Aspects of a Middle Cambrian thanatotope on Öland. Geologiska Föreningen i Stockholm Förhandlingar 87: 181–230. Crossref
McLoughlin, S., Vajda, V., Topper, T.P., Crowley, J.L., Liu, F., Johansson, O., and Skovsted, C.B. 2021. Trace fossils, algae, invertebrate remains and new U-Pb detrital zircon geochronology from the lower Cambrian Torneträsk Formation, northern Sweden. GFF 143: 103–133. Crossref
Melchor, R.N. and Cardonatto M.C. 2014. Insights on the behavior of late Paleozoic aquatic Crustaceans (Pygocephalomorpha?): compound trace fossils from western Argentina. Ichnos: An International Journal for Plant and Animal Traces 21: 76–99. Crossref
Michard, A. 1976. Eléments de Géologie Marocaine. 408 pp. Ministère de l’Energie et des Mines, Service Géologique du Maroc, Rabat.
Miller, S.A. 1889. North American Geology and Palaeontology for the Use of Amateurs, Students, and Scientists. 718 pp. Press of Western Methodist Book Concern, Cincinnati. Crossref
Najih, A., Montero, P., Verati, C., Chabou, M.C., Fekkak, A., Baidder, L., Ezzouhairi, H., Bea, F., and Michard, A. 2019. Initial Pangean rifting north of the West African Craton: Insights from late Permian U-Pb and 40Ar/39Ar dating of alkaline magmatism from the Eastern Anti-Atlas (Morocco). Journal of Geodynamics 132: 101670. Crossref
Neto de Carvalho, C. 2006. Roller coaster behavior in the Cruziana rugosa group from Penha Garcia (Portugal): implications for the feeding program of trilobites. Ichnos: An International Journal for Plant and Animal Traces 13: 255–265. Crossref
Nicholson, H.A. 1873. Contributions to the study of the errant annelides of the older Palaeozoic rocks. Royal Society of London Proceedings 21: 288–290. [= Geological Magazine 10: 309–310]. Crossref
Orłowski, S. 1992. Trilobite trace fossils and their stratigraphical significance in the Cambrian sequence of the Holy Cross Mountains, Poland. Geological Journal 27: 15–34. Crossref
Orłowski, S., Radwański, A., and Roniewicz, P. 1970. The trilobite ichnocoenosis in the Cambrian sequence of the Holy Cross Mountains. In: T.P. Crimes and J.C. Tarper (eds.), Trace Fossils. Geological Journal Special Issue 3: 345–360.
Osgood, R.G., Jr. 1970. Trace fossils of the Cincinnati area. Palaeontographica Americana 41: 281–444.
Osgood, R.G., Jr. 1975. The history of invertebrate ichnology. In: R.W. Frey (ed.), The Study of Trace Fossils: A Synthesis of Principles, Problems, and Procedures in Ichnology, 3–12. Springer, Berlin. Crossref
Osgood, R.G., Jr. and Drennen, W.T. III. 1975. Trilobite trace fossils from the Clinton Group (Silurian) of east-central New York State. Bulletins of American Palaeontology 67: 299–349.
Pemberton, S.G. and Frey, R.W. 1982. Trace fossil nomenclature and the Planolites—Palaeophycus Dilemma. Journal of Paleontology 56 (4): 843–881.
Pemberton, S.G., Spila, M., Pulham, A.J., Saunders, T., MacEachern, J.A., Robbins, D., and Sinclair, I.K. 2001. Ichnology and sedimentology of shallow to marginal marine systems: Ben Nevis and Avalon Reservoirs, Jeanne D’Arc Basin. Geological Association of Canada, Short Course Notes 15: 1–343.
Pickerill, R.K. 1992. Carboniferous nonmarine invertebrate ichnocoenoces from southern New Brunswick, eastern Canada. Ichnos 2: 21–35. Crossref
Pollard, J.E. 1985. Isopodichnus, related arthropod trace fossils and notostracans from Triassic fluvial sediments. Transactions of Royal Society of Edinburgh: Earth Sciences 76: 273–285. Crossref
Pollard, J.E., Selden, P., and Watts, S. 2008. Trace fossils of the arthropod Camptophyllia from the Westphalian (Carboniferous) rocks of Lancashire, UK and their palaeoenvironmental context. Palaeogeography, Palaeoclimatology, Palaeoecology 270: 399–406. Crossref
Rindsberg, A.K. 1994. Ichnology of the Upper Mississippian Hartselle Sandstone of Alabama, with notes on other Carboniferous formations. Bulletin of the Geological Survey of Alabama 158: 1–107.
Rindsberg, A.K. and Martin, A.J. 2003. Arthrophycus in the Silurian of Alabama (USA) and the problem of compound trace fossils. Palaeogeography, Palaeoclimatology, Palaeoecology 192: 187–219. Crossref
Rindsberg, A.K. and Kopaska-Merkel, D.C. 2005. Treptichnus and Arenicolites from the Steven C. Minkin Paleozoic Footprint Site (Langsettian, Alabama, USA). In: R.J. Buta, A.K. Rindsberg, and D.C. Kopaska-Merkel (eds.), Pennsylvanian Footprints in the Black Warrior Basin of Alabama. Alabama Paleontological Society Monograph 1: 121–141.
Roch, E. 1934. Sur des phénomènes remarquebles observés dans la région d’Erfoud. Association pour l’Etude Géologique de la Méditerranée Occidentale 5 (11): 15–40.
Salter, J.W. 1856. On fossil remains in the Cambrian rocks of the Longmynd and north Wales. Quarterly Journal of the Geological Society of London 12: 246–252. Crossref
Salter, J.W. 1857. On annelide-burrows and surface-markings from the Cambrian rocks of the Longmynd, No. 2. Quarterly Journal of the Geological Society of London 13: 199–206. Crossref
Schlirf, M., Uchman, A., and Kümmel, M. 2001. Upper Triassic (Keuper) nonmarine trace fossils from the Haßberge area (Franconia, south-eastern Germany). Paläontologische Zeitschrift 75: 71–96. Crossref
Scotese, C.R. 2014. Atlas of Devonian Paleogeographic Maps, PALEOMAP Atlas for ArcGIS, Vol. 4, The Late Paleozoic, maps 65–72, Mollweide Projection, Paleomap Project, Evanston, IL.
Seilacher, A. 1955. Spuren und Lebenweise der Trilobiten. In Beiträge zur Kenntnis des Kambriums in der Salt Range (Pakistan) (O.H. Schindewolfe and A. Seilacher). Akademie der Wissenschaften und der Literatur zu Mainz, Mathematisch-Naturwissen schaftliche Klasse, Abhandlungen 10: 342–372.
Seilacher, A. 1970. Cruziana stratigraphy of “nonfossiliferous” Palaeozoic sandstones. In: T.P. Crimes and J.C. Harper (eds.), Trace Fossils. Geological Journal Special Issue 3: 447–476.
Seilacher, A. 1985. Trilobite palaeobiology and substrate relationships. Transactions of the Royal Society of Edinburgh: Earth Sciences 76: 231–237. Crossref
Seilacher, A. 1991. An updated Cruziana stratigraphy of Gondwanan Palaeozoic sandstones. In: M.J. Salem, O.S., Hammuda, and B.A. Eliagoubi (eds.), The Geology of Libya, 1565–1580. Elsevier, Amsterdam.
Seilacher, A. 2007. Trace Fossil Analysis. 226 pp. Springer, Berlin.
Silva, R.C., Dominato, V.H., and Fernandes, A.C.S. 2012. Novos registros e aspectos palaeoambientais dos icnofósseis da Formação Pimenteira, Devoniano da Baciado Parnaíba, Piauí. Brasil. Gaea Journal of Geoscience 8: 33–41. Crossref
Stachacz, M. 2012. New finds of Rusophycus from the lower Cambrian Ociesęki Sandstone Formation (Holy Cross Mountains, Poland). Geological Quarterly 56: 237–248. Crossref
Stanley, D.C.A. and Pickerill, R.K. 1998. Systematic Ichnology of the Late Ordovician Georgian Bay Formation of Southern Ontario, Eastern Canada. 162 pp. Royal Ontario Museum, Ontario. Crossref
Tawadros, E.E. 2018. Geology of North Africa. 930 pp. CRC Press, Boca Raton.
Töneböhn, R. 1991. Bildungsbedingungen epikontinentaler Cephalopodenkalke (Devon, SE-Marokko). Göttinger Arbeiten zur Geologie und Paläontologie 47: 1–114.
Trewin, N.H. 1995. A draft system for the identification and description of arthropod trackways. Palaeontology 37: 811–823.
Trewin, N.H. and McNamara, K.J. 1994. Arthropods invade the land: trace fossils and paleoenvironments of the Tumblagooda Sandstone (late Silurian) of Kalbarri, western Australia. Transactions of the Royal Society of Edinburgh: Earth Sciences 85: 177–210. Crossref
Uchman, A. 1998. Taxonomy and ethology of flysch trace fossils: a revision of the Marian Książkiewicz collection and studies of complementary material. Annales Societatis Geologorum Poloniae 68: 105–218.
Vallon, L.H. and Rindsberg, A.K. 2022. Cutting through sponge and time—a new record of Koptichnus rasmussenae (trace fossil) from the Kerteminde Marl (middle Paleocene), Denmark. Bulletin of the Geological Society of Denmark 70: 131–137. Crossref
Vannier, J., Calandra, I., Gaillard, C., and Żylińska, A. 2010. Priapulid worms: Pioneer horizontal burrowers at the Precambrian–Cambrian boundary. Geology 38 (8): 711714. Crossref
Vannier, J., Vidal, M., Marchant, R., El Hariri, K., Kouraiss, K., Pittet, B., El Albani, A., Mazurier, A., and Martin, E. 2019. Collective behaviour in 480-million-year-old trilobite arthropods from Morocco. Scientific Reports 9: 14941. Crossref
Vanuxem, L. 1842. Natural History of New York. Geology, Vol. 3. 653 pp. W. & A. White & J. Visscher, Albany.
Vialov, O.S. 1971. The rare Mesozoic problematica from Pamir and Caucasus [in Russian, with English summary]. Paleontologičeskij sbornik 7 (2): 85–93.
Walliser, O.H. 1996. Patterns and causes of global events. In: O.H. Walliser (ed.), Global Events and Event Stratigraphy in the Phanerozoic, 7–19. Springer, Berlin. Crossref
Walter, H. and Gaitzsch, B. 1988. Beiträge zur Ichnologie limnisch–terrestrischer Sedimentation sräume Teil II: Diplichnites minimus n. ichnosp. aus dem Permosiles des Flechtinger Höhenzuges. Freiberger Forschungshefte C 427: 73–84.
Webby, B.D. 1983. Lower Ordovician arthropod trace fossils from western New South Wales. Proceedings of the Linnean Society of New South Wales 107: 59–74.
Wendt, J. 1985. Disintegration of the continental margin of north western Gondwana, Late Devonian of the eastern Anti-Atlas (Morocco). Geology 13: 815–818. Crossref
Wendt, J. 1988a. Condensed carbonate sedimentation in the Late Devonian of the eastern Anti-Atlas (Morocco). Eclogae Geologicae Helvetiae 81: 155–173.
Wendt, J. 1988b. Facies pattern and paleogeography of the Middle and Late Devonian in the eastern Anti-Atlas (Morocco). In: N.J. McMillan, A.F. Embry, and D.E. Glass (eds.), Devonian of the World, Proceedings of the Second International Symposium on the Devonian System, Alberta. Canadian Society of Petroleum Geologists: Geologic Notes 14: 531–539.
Wendt, J. 2021a. Middle and Late Devonian paleogeography of the eastern Anti-Atlas (Morocco). International Journal of Earth Sciences 110: 1531–1544. Crossref
Wendt, J. 2021b. Middle and Late Devonian sea-level changes and synsedimentary tectonics in the eastern Anti-Atlas (Morocco). Journal of African Earth Sciences 182: 104247. Crossref
Wendt, J. and Belka, Z. 1991. Age and depositional environment of Upper Devonian (early Frasnian to early Famennian) black shales and limestones (Kellwasser facies) in the eastern Anti-Atlas, Morocco. Facies 25: 51–90. Crossref
Wendt, J., Aigner, T., and Neugebauer, J. 1984. Cephalopod limestone deposition on a shallow pelagic ridge: the Tafilalt Platform (Upper Devonian, eastern Anti-Atlas, Morocco). Sedimentology 31: 601–625. Crossref
Wu, X. 1985. Trace fossils and their significance in non-marine turbidite deposits of Mesozoic coal and oil-bearing sequences from Yima-Jiyuan basin, western Henan, China [in Chinese with English summary]. Acta Sedimentologica Sinica 3: 23–31.
Acta Palaeontol. Pol. 71 (2): 273–295, 2026
https://doi.org/10.4202/app.01312.2025