
Overlooked diversity of the predatory fungus gnat Macrocera in Baltic amber
ALICJA PEŁCZYŃSKA, WIESŁAW KRZEMIŃSKI, and AGNIESZKA SOSZYŃSKA
Pełczyńska, A., Krzemiński, W., and Soszyńska, A. 2026. Overlooked diversity of the predatory fungus gnat Macrocera in Baltic amber. Acta Palaeontologica Polonica 71 (2): 219–245.
The genus Macrocera (Diptera: Keroplatidae) is highly diverse, comprising over 200 species distributed worldwide, yet it remains poorly documented in the extensive sciaroid fossil record. This study presents the results of our investigation of Macrocera in Baltic amber, which led to the discovery of five new species (Macrocera barteki Pełczyńska sp. nov., Macrocera hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov., Macrocera poseidonis Pełczyńska, Krzemiński, & Soszyńska sp. nov., Macrocera stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov., and Macrocera tymoni Pełczyńska sp. nov.) and to the re-examination of the only previously described valid species, Macrocera electricornis. A diagnostic key for species identification is provided. Our results indicate that Macrocera had already attained its modern morphological body plan by the Eocene, highlighting Baltic amber as a largely unexplored source of information on the evolutionary history of certain insect groups. We further discuss the delimitation of the genus and emphasise the need for its comprehensive revision.
Key words: Diptera, Keroplatidae, fossil insects, paleodiversity, Baltic amber, Eocene.
Alicja Pełczyńska [alicja.pelczynska@edu.uni.lodz.pl; ORCID: https://orcid.org/0000-0002-7000-3552 ], University of Lodz, Faculty of Biology and Environmental Protection, Department of Invertebrate Zoology and Hydrobiology, 90-237 Łódź, Poland; University of Copenhagen, Faculty of SCIENCE, Natural History Museum of Denmark, 2100 Copenhagen, Denmark.
Wiesław Krzemiński [wieslawk4@gmail.com; ORCID: https://orcid.org/0000-0001-5685-891X ], Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, 31-016 Kraków, Poland.
Agnieszka Soszyńska [agnieszka.soszynska@biol.uni.lodz.pl; ORCID: https://orcid.org/0000-0002-2661-6685 ], University of Lodz, Faculty of Biology and Environmental Protection, Department of Invertebrate Zoology and Hydrobiology, 90-237 Łódź, Poland.
Received 12 December 2025, accepted 3 February 2026, published online 19 May 2026.
Copyright © 2026 A. Pełczyńska et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The genus Macrocera Meigen, 1803 (subfamily Macrocerinae, tribe Macrocerini) is arguably one of the most remarkable taxa within the predatory fungus gnat family Keroplatidae (Diptera: Sciaroidea). It is the most speciose taxon, comprising more than 200 extant species, and is distributed worldwide, from the low Arctic of Greenland to Subantarctic regions, with records even from the Crozet Islands (Matile 1990; Evenhuis 2006). Moreover, new extant and fossil species continue to be described each year (Mantič and Ševčík 2017; Pełczyńska et al. 2024, 2025; Ševčík 2024; Lim et al. 2025).
In particular, the larvae of those species whose biology has been studied are predatory, spinning sticky webs coated with an acidic fluid to capture and kill small invertebrate prey, or possibly combining carnivory with fungivory (Matile 1990). However, the genus remains poorly studied, and available observations suggest that trophic strategies within Macrocera may be more diverse. Larvae have been recorded from natural caves as well as artificial underground habitats, such as cellars and basements, but also frequently occur in narrow crevices under tree bark, where the availability of potential prey may be limited, and have been reared from grass and sedge tussocks (Laurence 1982; Matile 1990; Ševčík and Roháček 2008). Therefore, further studies are required to better understand the biology and ecology of this genus.
In adults, Macrocera is perhaps best known for its long antennae, which can be up to five times the length of the body (Matile 1990). This characteristic is shared by most species, and in reference to it the genus name is derived from the Greek words makros (μακρός), meaning “long”, and keras (κέρας), meaning “horn” (Mantič and Ševčík 2017). Given their distinctive anatomy, species richness, and relatively large size, it is somewhat surprising that this genus is poorly reported and documented among the numerous Sciaroidea inclusions known from Baltic amber. This prompted a closer examination of this fossil resin.
The fauna of Baltic amber might be perceived as relatively well studied, given that ca. 3500 species of arthropods have been described over more than 300 years of research (Penney and Preziosi 2013). At the same time, Diptera constitute the dominant group among the numerous insect inclusions found in this resin, accounting for more than 70% of all inclusions. Most of these are nematoceran flies, including many sciaroids (Perkovsky et al. 2007). However, only eight nominal species of Macrocera have been proposed from Baltic amber, and only one of them remains valid within this genus.
Representatives of the genus Macrocera in Baltic amber were first reported by Loew (1850), although he did not provide formal species descriptions. The first species were described much later by Meunier (1899) as Macrocera grandis Meunier, 1899, Macrocera minuta Meunier, 1899, and Macrocera socatta Meunier, 1899. However, the first two should be considered nomina nuda, as no descriptions or illustrations were provided. The type specimens of the third species are most likely lost, and the original description is insufficient to allow a reliable diagnosis; therefore, it should be treated as a nomen dubium (Evenhuis 2006; Blagoderov et al. 2019).
Among the five species described later by Meunier et al. (1904), four were subsequently reassigned: Macrocera abundare (Meunier, 1904), Macrocera ciliata (Meunier, 1904), and Macrocera filiformis (Meunier, 1904) were transferred to Kelneria Matile, 1979, while Macrocera elegantissima was redescribed as Micrepimera Matile, 1990 (Blagoderov et al. 2019). As a result, only Macrocera electricornis Evenhuis, 2006 (originally described as Macrocera longicornis by Meunier, a junior homonym of Macrocera longicornis Fabricius, 1781) should be regarded as a valid species of Macrocera known from Baltic amber.
Macrocera electricornis was described from a single specimen, and because the original description included only a very limited set of morphological characters, we re-examined the holotype and provide a redescription of this species. This enabled accurate comparisons with the newly discovered species from Baltic amber.
The main aim of this study was to analyse the actual richness of Macrocera in Eocene Baltic amber. This diversity had previously remained unexplored, as initially hypothesised, and our findings confirm this. Consequently, we describe five new species and provide a diagnostic key for their identification. The delimitation of the genus is also discussed.
Institutional abbreviations.—CCHH, the collection of Christel and Hans Werner Hoffeins, Hamburg, Germany deposited in SDEI; GZG, Geowissenschaftliches Zentrum, Universität Göttingen, Göttingen, Germany; ISEA PAS, Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland; SDEI, Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany.
Other abbreviations.—A₁/A₂, first/second anal vein; anepm, anepimeron; anepst, anepisternum; C, costal vein; cer scl, cerebral sclerite; Cu, cubital vein; flgm, flagellomere; frm, radio-medial fusion; h, humeral crossvein; kepst, katepisternum; ltg, laterotergite; med, mediotergite; M₁/M₂, first/second branch of media; M₃₊₄, fourth branch of media; Mb, mediobasal vein; m-cu, medio-cubital crossvein; oc, ocellus; pcelli, posterior ocelli; ped, pedicel; plp, palpomere; plv, pulvillus; R₁, anterior branch of radius; R₂₊₃/R₂₊₃₊₄₊₅, second branch of radius; R₄₊₅, third branch of radius; Rb, radiobasal vein; Rs, radial sector; Sc, subcostal vein; scp, scape; sc-r, subcostal-radial crossvein; sct, scutum; sctl, scutellum.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:ADC93CCC-B187-4210-B161-D2F0DB5479BF
Material and methods
The investigated holotype of Macrocera electricornis, GZG.BST.3064 (former Königsberg number Z8194), and the holotypes of the newly described species (CCHH 582-1, MP/3247, 4422, 5331, 5332) originate from Eocene Baltic amber. This resin was presumably produced by plant assemblages that covered large parts of Fennoscandia. However, it is not deposited at its place of origin, as it was redistributed across the Northern European Plain through fluvial transport, marine transgressions, and glacial processes (Szwedo and Sontag 2013; Chang et al. 2023). Consequently, Baltic amber lacks an original stratigraphic context, which poses a serious challenge for precise dating. The broadly accepted age range of Baltic amber extends from the Lutetian to the Priabonian (approx. 47.8–33.9 Ma; Grimaldi and Ross 2018). While it is generally impossible to date specimens collected from beaches and shallow nearshore waters more precisely, more refined age estimates can be obtained for specimens recovered from the “Blue Earth” layer of amber-bearing sediments (see the Discussion for further details; Ross et al. 2026). Unfortunately, the majority of amber specimens in collections lack precise provenance data, with labels typically providing no more information than an attribution to Baltic amber. This is also the case for the specimens investigated in this study.
The Baltic amber origin of the investigated specimens was verified using Fourier transform infrared spectroscopy (FTIR) with a Nicolet iS5 FTIR spectrometer equipped with a diamond crystal attenuated total reflectance (ATR) accessory. The obtained spectra have been archived in the database of the ISEA PAS, in accordance with the guidelines proposed by Zakrzewska et al. (2020).
The amber pieces were manually ground using wet sandpaper with grits ranging from 220 to 2500 and subsequently polished with a fine abrasive paste on a felt polishing wheel. Morphological analyses were then performed using transmitted light microscopy with a Leica M205 C stereomicroscope.
Photographs of Macrocera electricornis were taken at the University of Lodz (Poland) using a Leica M205 C stereomicroscope equipped with a Leica DMC5400 camera. Image stacking and subsequent processing were performed using Leica Application Suite X software (LAS X; Leica Microsystems). Photographic documentation of the newly described species was carried out at the Natural History Museum of Denmark using a Canon EOS R7 camera equipped with an Infinity K2/SC lens and Nikon 5× or 10× microscope objectives. Focus-stacked image series were captured using a Cognisys StackShot 3X Macro Rail, and image stacking was performed with Zerene Stacker software (Zerene Systems). Line drawings were subsequently prepared in CorelDRAW 2018 based on the photographs.
The holotype of Macrocera electricornis (GZG.BST. 3064, old Königsberg number Z8194) is currently housed at the GZG. The holotypes of Macrocera barteki Pełczyńska sp. nov. (MP/3247), Macrocera poseidonis Pełczyńska, Krzemiński, & Soszyńska sp. nov. (MP/5332), Macrocera stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov. (MP/5331), and Macrocera tymoni Pełczyńska sp. nov. (MP/ 4422) are deposited at the ISEA PAS. The holotype of Macrocera hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov. (CCHH 582-1) is a part of the collection of C. & H.W. Hoffeins and is deposited in the collection of SDEI.
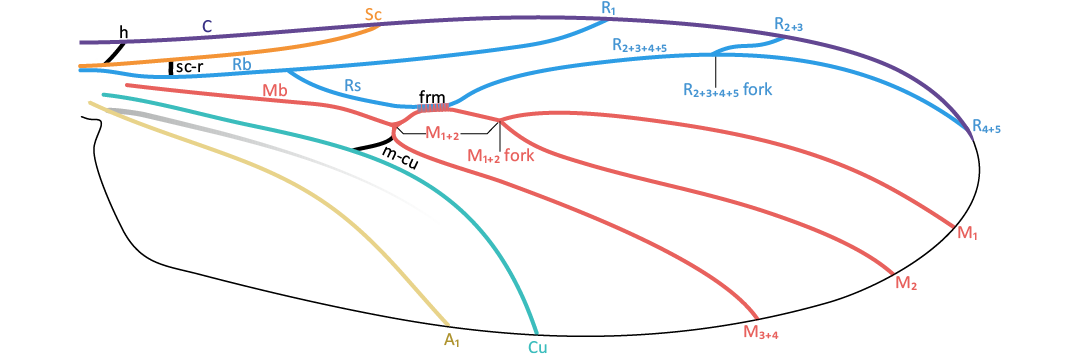
Fig. 1. Hypothetical ground plan of the Keroplatidae wing.
The morphological terminology used in this publication mainly follows the nomenclature outlined in the Manual of Afrotropical Diptera (Volume 1) by Cumming and Wood (2017). The wing vein nomenclature (Fig. 1) follows that used in our previous works (Pełczyńska et al. 2024, 2025), based on the nomenclature proposed by Ševčík et al. (2020, 2022). The following terminology is applied: basal part of M1+2 = M1+2 section from its origin to the start of frm; M1+2 fork stem = M1+2 section from the end of the radio-medial fusion to the M1+2 fork; R2+3+4+5 fork stem = R2+3+4+5 section from the end of the radio-medial fusion to the R2+3+4+5 fork. Additionally, in the case of the Mb, which atrophies to various degrees in different species (see the Discussion for further details), its course has been marked on wing reconstructions even in cases where only its trace remains, and the degree of its preservation has been indicated in the text.
For male genitalia color coding was used as follows: yellow, cercus; violet, epandrium; pink, gonocoxites; blue, gonostylus; green, hypoproct; grey, membranous structures.
Systematic palaeontology
Class Insecta Linnaeus, 1758
Order Diptera Linnaeus, 1758
Infraorder Bibionomorpha Hennig, 1948
Superfamily Sciaroidea Billberg, 1820
Family Keroplatidae Rondani, 1856
Subfamily Macrocerinae Rondani, 1856
Tribe Macrocerini Rondani, 1856
Genus Macrocera Meigen, 1803
Type species: Macrocera lutea Meigen, 1804, by original designation; Germany, Recent.
Diagnosis.—See Meigen (1803).
Macrocera electricornis Evenhuis, 2006
Figs. 2, 3.
1904 Macrocera longicornis Meunier, 1904 nec Fabricius, 1781; Meunier 1904a: 91; 1904b: 102.
2006 Macrocera electricornis Evenhuis, 2006, nomen novum pro Macrocera longicornis Meunier, 1904; Evenhuis 2006: fig. 36.
Holotype: GZG.BST.3064, male in 15×10×4 mm piece of Baltic amber (Fig. 2A1, A2; IR spectrum Fig. 20A).
Type locality: Baltic region.
Type horizon: Baltic amber, mid-Eocene (ca. 47.8–33.9 Ma according
Diagnosis.—Antennae more than 2× wing length; vein R1 terminating in C before the R2+3+4+5 fork; R2+3+4+5 fork situated distal to the level of the M1+2 fork; R2+3 approx. 0.5× length of the R2+3+4+5 fork stem; apical part of M1+2 approx. 2.3× frm length; fore tibial spur longer than apical width of tibia; male genitalia wider than apex of abdomen; gonocoxites forming two widely separated arms, largely divided dorsally; gonostyli broad, laterally flattened, shorter than gonocoxites, ending with two small, subequal teeth.
Description.—Body (Fig. 2A1): Approx. 3.9 mm long; wing more than 3.0 mm; antennae incomplete, exceeding 8.3 mm in length.
Head (Fig. 2A3, A4): Subspherical, wider than long, inserted just below anterior margin of scutum; eyes large, well separated, occupying most of lateral part of head capsule, inner margins slightly notched at level of antennal insertion, without interommatidial setae; large, distinct cerebral sclerite present, angular at antero-external angles, narrowing posteriorly, bare; three ocelli present, closely grouped, located on anterior margin of cerebral sclerite, forming a triangle, median ocellus not reduced.

Fig. 2. Predatory fungus gnat Macrocera electricornis Evenhuis, 2006 (male, holotype, GZG.BST.3064), Eocene Baltic amber, Baltic region. General view of the specimen (A1), amber piece containing specimen (A2), head in frontal (A3) and dorsal (A4) views; apical spur of the fore tibia (A5), mid tibia (A6), and hind tibia (A7), tip of the tarsus of fore leg (A8).
Antennae (Fig. 2A1): Incomplete; preserved segments indicate length exceeding 2× wing length; scapus as long as broad, globular, approx. 2.5× wider than first flagellomere; pedicel short, annular, approx. 2.1× wider than long, narrower than scape, 1.7× wider than first flagellomere; flagellum with 12 segments preserved; flagellomeres distinctly elongated, first flagellomere approx. 7.2× as long as broad, subsequent flagellomeres elongate, cylindrical, bearing sparse row of setae. Palpi (Fig. 2A3): four maxillary palpomeres visible, all longer than broad; apical palpomere distinctly elongated, approx. 7.4× as long as broad, 1.8× longer than proximal palpomere.
Wing (Fig. 3A1, A3): Membrane hyaline, without microtrichia or visible markings; C bearing microtrichia throughout length, remaining veins apparently bare; C terminating at wing tip, beyond end of R4+5, at approx. one-third distance between terminations of R4+5 and M1; Sc ending in C at level of tip of Rb cell, well before level at which A1 reaches wing margin; R1 extending beyond half wing length, terminating in C distal to R2+3+4+5 fork, at approx. half distance between terminations of Cu and M3+4; R2+3 short, approx. 0.2× length of R2+3+4+5 fork stem; frm long, ending before level at which A1 reaches wing margin; Mb absent; M1+2 fork stem approx. 2.3× longer than frm, ending just before level of Cu termination; M1 approx. 5× longer than M1+2 fork stem; opening of cell M2 approx. 1.3× wider than opening of cell M1; base of M3+4 not atrophied; opening of cell M3+4 approx. 1.7× wider than opening of cell M2; basal portion of M1+2 and m-cu distinct; Cu reaching wing margin; A1 ending at wing margin; A2 absent.
Thorax (Fig. 3A2): Higher than long; scutum arched, almost semicircular in lateral view, covered with sparse setation arranged in three rows (one dorsocentral and two lateral); scutellum scale-shaped; anepimeron bare, not reduced, reaching ventral margin of pleura; anepisternum, katepisternum, laterotergite and mediotergite bare. Legs (Fig. 2A5–A8): fore coxa with sparse, long setae on anterior surface; mid and hind coxae with few setae; femora irregularly covered with thin setae; tibiae bearing ventral row of short, robust setae; fore tibia with single long spur (approx. 1.25× apical width of tibia), sensory pit and row of apical bristles; mid and hind tibiae with long spurs of unequal length, ventral spur slightly shorter (longer spurs approx. 1.4× and 2.8× apical width of tibiae, respectively); tarsal claws small; pulvilli small.
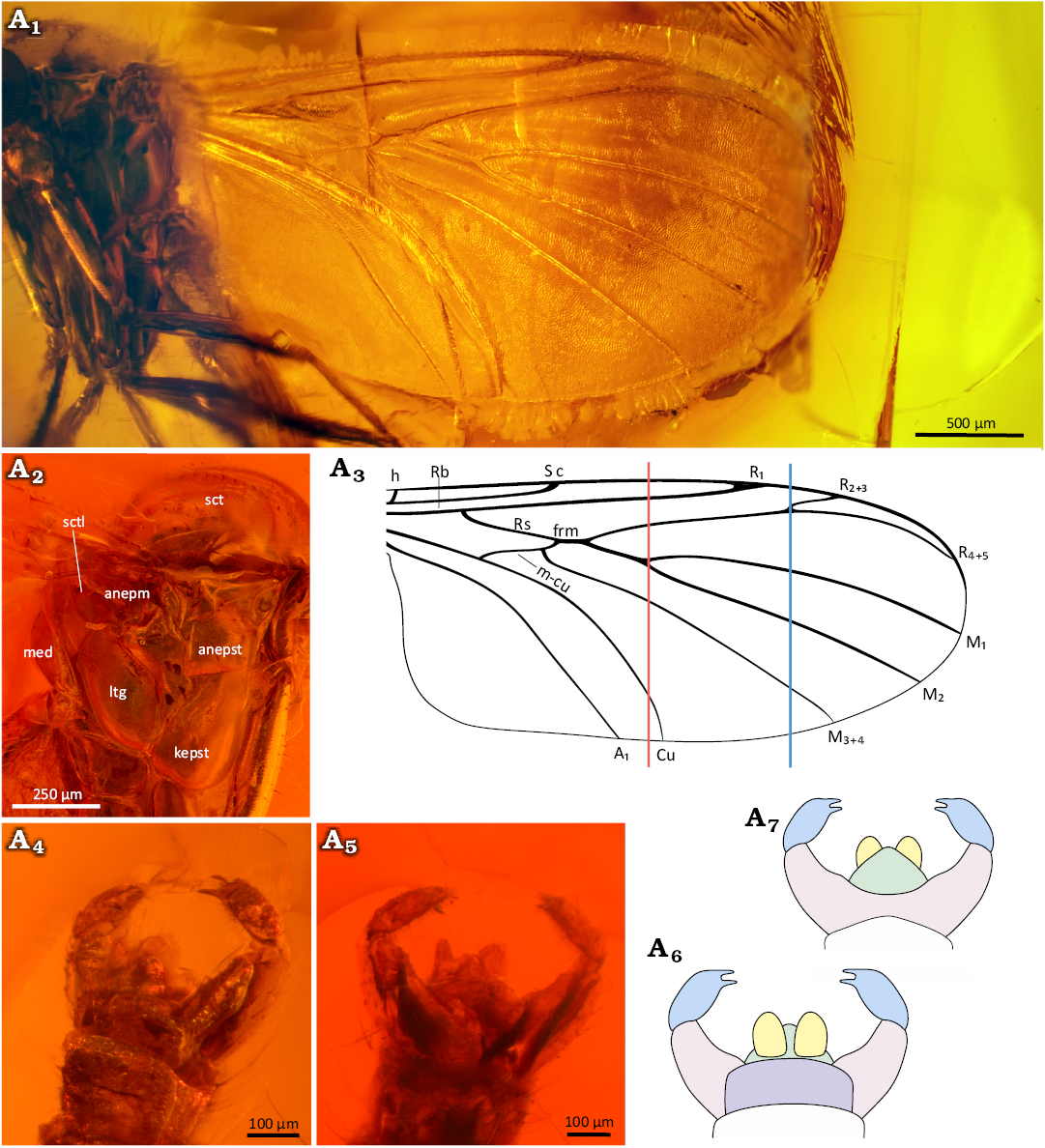
Fig. 3. Predatory fungus gnat Macrocera electricornis Evenhuis, 2006 (male, holotype GZG.BST.3064), Eocene Baltic amber, Baltic region. Wing in ventral view (A1, A3), thorax in lateral view (A2), genitalia in dorso-lateral (A4, A6) and ventral (A5, A7) views. Photographs (A1, A2, A4, A5), explanatory drawings (A3, A6, A7, not to scale). Abbreviations: anepm, anepimeron; anepst, anepisternum; kepst; kepst, katepisternum; ltg, laterotergite; med, mediotergite; sct, scutum; sctl, scutellum. Colour coding: yellow, cercus; violet, epandrium; pink, gonocoxites; blue, gonostylus; green, hypoproct. Lines: red, level of the M1+2 fork; blue, level of the R2+3+4+5 fork.
Abdomen (Fig. 3A4–A7): Sparsely setose; eight segments visible; segment I short; segments II–IV approx. equal in length; subsequent segments gradually decreasing in length; male terminalia (Fig. 3A4–A7) wider than apex of abdomen; gonocoxites forming two widely separated arms, largely divided dorsally beneath epandrium; pair of cerci present; gonostyli broad, laterally flattened, shorter than gonocoxites, ending with two subequal teeth pointed ventrally; aedeagus not visible.
Remarks.—The original description of this species was based on a single specimen (GZG.BST.3064; Fig. 2A1, A2) and included insufficient number of morphological characters; it stated that vein C extends beyond termination of vein Cu; however, present observations indicate that it is considerably shorter, terminating shortly after end of R4+5, at approx. one-third distance between terminations of R4+5 and M1; wing on which original reconstruction was primarily based appears optically shortened, therefore length-to-width ratio was not provided; wing was digitally stretched in present reconstruction to recreate its natural proportions; wing apex reconstructed based on second wing.
Stratigraphic and geographic range.—Baltic amber, mid-Eocene (ca. 47.8–33.9 Ma according to Seyfullah et al. 2018). Kaliningrad Region, Russian Federation.
Macrocera barteki Pełczyńska sp. nov.
Figs. 4, 5.
Zoobank LCID: urn:lsid:zoobank.org:act:E8A5C7D2-1EC9-4B45-8319-61361D67EE93
Etymology: A patronym barteki dedicated to the first author’s brother, Bartosz (“Bartek”) Pełczyński.
Holotype: MP/3247, male in 26×11×4 mm piece of Baltic amber (Fig. 4A1, A2; IR spectrum Fig. 20B).
Type locality: Baltic region.
Type horizon: Baltic amber, mid-Eocene (ca. 47.8–33.9 Ma according to Seyfullah et al. 2018).
Material.—Holotype only.
Diagnosis.—Antennae more than 1.7× wing length; vein R1 terminating in C at approx. the level of the R2+3+4+5 fork; R2+3+4+5 fork situated just proximal to the level of the M1+2 fork; R2+3 approx. 0.5× the length of the R2+3+4+5 fork stem; apical part of M1+2 approx. 5.6× frm length; fore tibial spur shorter than apical width of tibia; male genitalia as wide as apex of abdomen; gonocoxites small, positioned parallel to each other; gonostyli short and broad, laterally flattened, approx. as long as gonocoxites, ending with two very small, subequal teeth pointed ventrally; aedeagus not visible.
Description.—Body (Fig. 4A1): Approx. 3.9 mm long; wing 4.3 mm; antennae incomplete, exceeding 12 mm in length.
Head (Fig. 5A1): Subspherical, wider than long, inserted just below anterior margin of scutum; eyes large, well separated, occupying most of lateral part of head capsule, inner margins slightly notched at level of antennal insertion, interommatidial setae present; distinct cerebral sclerite present, angular at antero-external angles, narrowing posteriorly, with few short setae along anterior margin; three ocelli present, closely grouped, located on anterior margin of cerebral sclerite, forming a triangle, median ocellus not reduced.
Antennae (Figs. 4A1, 5A1): Incomplete, preserved portion exceeding 2× wing length; scapus as long as broad, globular, approx. 3.9× wider than first flagellomere; pedicel short, annular, approx. 1.4× wider than long, distinctly smaller than scape, 2.4× wider than first flagellomere; flagellum with 12 segments preserved; flagellomeres distinctly elongated, first flagellomere approx. 11.4× as long as broad, subsequent flagellomeres elongate, cylindrical, bearing a sparse row of longer setae on anterior surface.
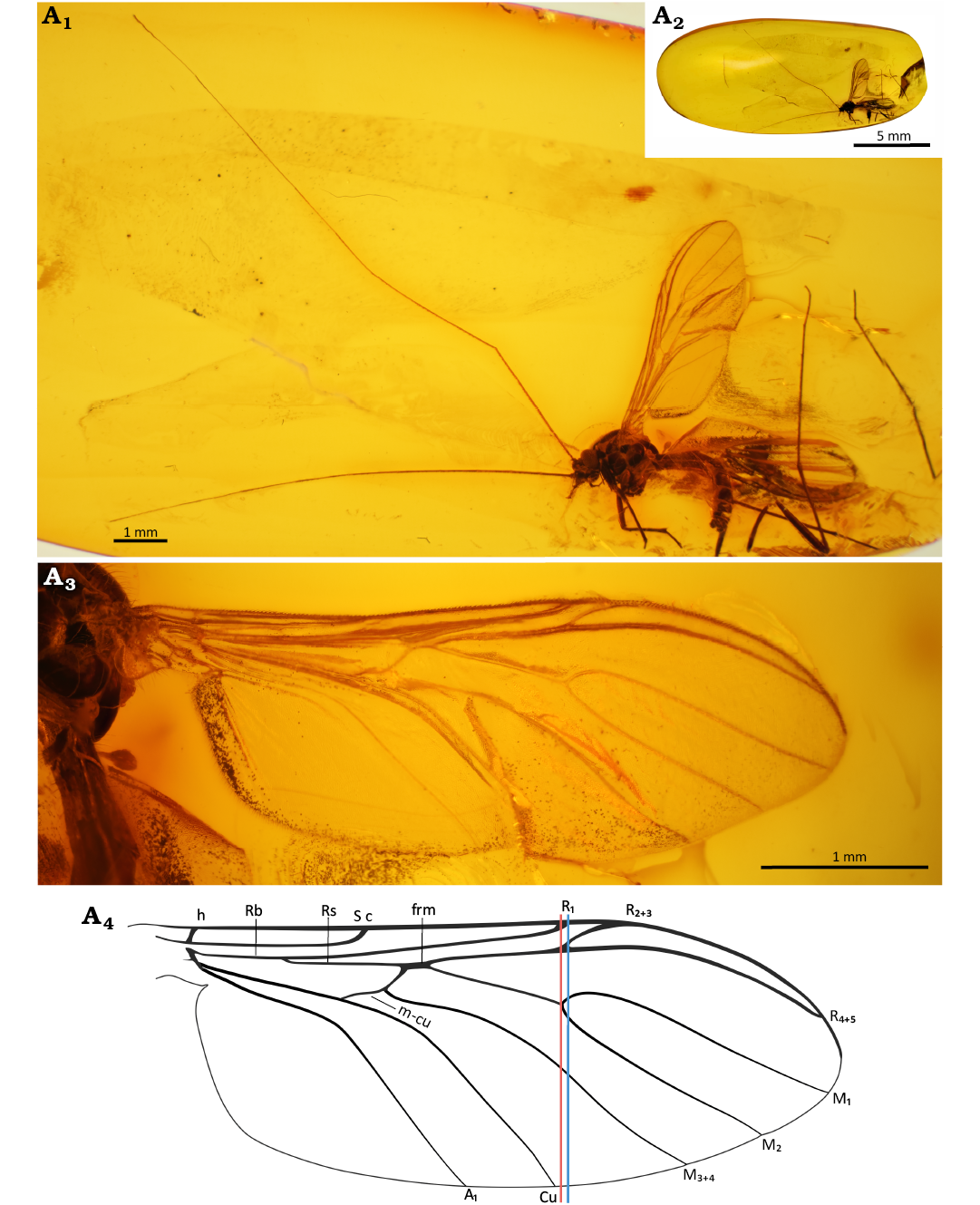
Fig. 4. Predatory fungus gnat Macrocera barteki Pełczyńska sp. nov. (male, holotype MP/3247), Eocene Baltic amber, Baltic region. General view of the specimen (A1), amber piece containing specimen (A2), wing in ventral view (A3, A4). Photographs (A1–A3), explanatory drawing (A4, not to scale). Lines: red, level of the M1+2 fork; blue, level of the R2+3+4+5 fork.
Palpi (Fig. 5A1): Four maxillary palpomeres visible, all longer than broad; apical palpomere distinctly elongated, approx. 18× as long as broad, 2.3× longer than proximal palpomere. Wing (Fig. 4A3, A4): 2.4× longer than wide; membrane hyaline, without microtrichia or visible markings; C bearing microtrichia throughout its length; microtrichia present on dorsal surface of Rb, R2+3+4+5 and R4+5; C terminating at wing tip, beyond end of R4+5, at approx. half the distance between terminations of R4+5 and M1; Sc ending in C well before level of tip of Rb cell, at approx. half the length of m-cu; R1 extending beyond half wing length, terminating in C at approx. the level of the R2+3+4+5 fork, just distal to Cu termination; R2+3+4+5 fork situated just proximal to level of M1+2 fork; R2+3 approx. 0.5× the length of R2+3+4+5 fork stem; frm ending before level at which A1 reaches wing margin; Mb absent; M1+2 fork stem approx. 5.6× longer than frm, ending just before level of Cu termination; M1 approx. 2.1× longer than M1+2 fork stem; opening of cell M2 approx. as wide as that of cell M1; base of M3+4 not atrophied; opening of cell M3+4 approx. 1.6× wider than opening of cell M2; basal part of M1+2 and m-cu distinct; Cu reaching wing margin; A1 ending at wing margin; A2 absent. Thorax (Fig. 5A5): higher than long; scutum arched, almost semicircular in lateral view, covered with sparse, short setation on lateral edges and dorsal surface, arranged in three distinct rows (one dorsocentral and two lateral); scutellum bearing a row of trichia along its margin; anepimeron bare, not reaching ventral margin of pleura; anepisternum, katepisternum, laterotergite and mediotergite bare.
Legs (Fig. 5A2–A4): Fore coxa with sparse, long setae on anterior surface; mid and hind coxae without visible setation; femora irregularly covered with very small, sparse setae; fore tibia bearing a ventral row of short, robust setae and a single relatively short spur (approx. 0.55× apical width of tibia), a sensory pit, and a row of apical bristles; mid and hind legs detached.
Abdomen (Fig. 5A6–A9): Sparsely setose, bearing few thin, long setae; eight segments visible; segment I short; segments II–III approx. equal in length; subsequent segments gradually decreasing in length; male terminalia (Fig. 5A6–A9) as wide as apex of abdomen; gonocoxites small, positioned parallel to each other; pair of cerci present; gonostyli short and broad, laterally flattened, approx. as long as gonocoxites, ending with two very small, subequal teeth pointed ventrally; aedeagus not visible.

Fig. 5. Predatory fungus gnat Macrocera barteki Pełczyńska sp. nov. (male, holotype MP/3247), Eocene Baltic amber, Baltic region. Head in lateral view (A1), spur of the fore tibia (A2) and detached leg (A3), tip of the tarsus of a detached leg (A4), thorax in lateral view (A5), genitalia in dorsalo-apical (A6), dorsal (A8), and ventral (A7, A9) views. Photographs (A1–A7), explanatory drawings (A8, A9, not to scale). Abbreviations: anepm, anepimeron; anepst, anepisternum; cer scl, cerebral sclerite; flgm, flagellomere; kepst, katepisternum; ltg, laterotergite; med, mediotergite; oc, ocelli; ped, pedicel; plp, palpomere; scp, scape; sct, scutum. Colour coding: yellow, cercus; violet, epandrium; pink, gonocoxites; blue, gonostylus; green, hypoproct.
Remarks.—Mid and hind legs detached; however, one loose leg (Fig. 5A3, A4), located near the body, is assumed to originate from the specimen; it bears a single tibial spur and a preserved tarsus with distinct claws, whereas pulvilli are not observable; shape of cerci indicated in the figure with a dashed line, as its interpretation remains uncertain.
Stratigraphic and geographic range.—Type locality and horizon only.
Macrocera hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov.
Figs. 6–8.
Zoobank LCID: urn:lsid:zoobank.org:act:A3248BC4-BAED-4ECA-B3A0-BE5256068861
Etymology: A patronym hoffeinsorum dedicated to Christel and Hans Werner Hoffeins, from whose collection the specimen originates.
Holotype: CCHH 582-1, male in 17×11×5 mm piece of Baltic amber embedded in synthetic resin to prevent oxidation (Fig. 6A1, A2; IR spectrum Fig. 20C).
Type locality: Baltic region.
Type horizon: Baltic amber, mid-Eocene (ca. 47.8–33.9 Ma according to Seyfullah et al. 2018).
Material.—Holotype only.
Diagnosis.—Antennae more than 1.8× wing length; vein R1 terminating in C just after the R2+3+4+5 fork; R2+3+4+5 fork situated at approx. the level of the M1+2 fork; R2+3 approx. 0.4× the length of the R2+3+4+5 fork stem; M1+2 fork stem approx. 7× frm length; fore tibial spur shorter than apical width of tibia; male genitalia wider than apex of abdomen; gonocoxites forming two widely separated arms; gonostyli flattened, shorter than gonocoxites, ending with two teeth of equal length, pointed ventrally.
Description.—Body (Fig. 6A2): 4.2 mm long; wing 4.6 mm long; antennae incomplete, more than 8.1 mm long.
Head (Fig. 7A1): Subspherical, wider than long, inserted just below anterior margin of scutum; eyes large, well separated, occupying most of lateral part of head capsule, inner margins notched at level of antennal insertion, without interommatidial setae visible; cerebral sclerite distinct but poorly visible due to positioning of specimen.

Fig. 6. Predatory fungus gnat Macrocera hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov. (male, holotype CCHH 582-1), Eocene Baltic amber, Baltic region. Amber piece containing specimen (A1), general view of the specimen (A2).
Antennae (Figs. 6A2, 7A1): Incomplete, remaining part exceeding wing length; scapus as long as broad, globular in shape, significantly broader than first flagellomere; pedicel poorly visible, short, annular in shape, narrower than scape; flagellum incomplete; flagellomeres distinctly elongated with first flagellomere approx. 9.2× long as broad, subsequent flagellomeres elongate, cylindrical and with row of sparse setae.
Palpi (Fig. 7A1): Four maxillary palpomeres visible; palpomeres II–IV longer than broad; apical palpomere distinctly elongated, approx. 10.1× as long as broad, 2.2× longer than proximal palpomere.
Wing (Fig. 7A3, A4): 2.5× longer than wide; membrane hyaline, without microtrichia or visible markings; C bearing microtrichia throughout its length; microtrichia present on dorsal surface of Rb, R2+3+4+5 and R4+5; C terminating at wing tip, beyond end of R4+5, just after half the distance between terminations of R4+5 and M1; Sc ending in C before level of tip of Rb cell, at approx. the level of m-cu base; R1 extending beyond half wing length, terminating in C just distal to level of R2+3+4+5 fork, at approx. one-third of the distance between terminations of Cu and M3+4; R2+3+4+5 fork situated at approx. the level of the M1+2 fork; R2+3 approx. 0.4× the length of the R2+3+4+5 fork stem; frm ending before level at which A1 reaches wing margin; Mb represented by a faint trace only, not reaching tip of Rb cell; M1+2 fork stem approx. 7× longer than frm, ending at approx. the level of the R2+3+4+5 fork; M1 approx. 2× longer than M1+2 fork stem; opening of cell M2 approx. as wide as that of cell M1; base of M3+4 present but weakened; opening of cell M3+4 approx. 2.6× wider than opening of cell M2; basal part of M1+2 and m-cu distinct; Cu reaching wing margin; A1 ending at wing margin, close to Cu; A2 absent.
Thorax (Fig. 7A2): Higher than long; scutum arched, almost semicircular in lateral view, covered with sparse, short setation along lateral edges, dorsal surface poorly visible; anepimeron bare, not reaching ventral margin of pleura; anepisternum bearing a few short setae near dorsoanterior margin; katepisternum, laterotergite and mediotergite bare.

Fig. 7. Predatory fungus gnat Macrocera hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov. (male, holotype CCHH 582-1), Eocene Baltic amber, Baltic region. Head in frontal view (A1), thorax in lateral view (A2), wing in lateral view? (A3, A4). Photographs (A1–A3), explanatory drawing (A4, not to scale). Abbreviations: anepm, anepimeron; anepst, anepisternum; kepst, katepisternum; ltg, laterotergite; med, mediotergite; plp, palpomere; scp, scape; sct, scutum; sctl, scutellum. Lines: red, level of the M1+2 fork; blue, level of the R2+3+4+5 fork.
Legs (Fig. 8A1–A4): Fore coxa with dense, long setae on anterior surface; mid and hind coxae appearing bare; femora bearing thin setae along ventral margin; tibiae covered with very minute setae, slightly longer setae present only on hind legs; fore tibia with a single relatively short spur (approx. 0.8× apical width of tibia), a sensory pit, and a row of apical bristles; mid and hind tibiae with short spurs of unequal length, ventral spur slightly shorter (longer spurs measuring approx. 0.9× apical width of tibiae); tarsal claws distinct; pulvilli not observable.
Abdomen (Fig. 8A5, A6): Partially concealed beneath wings, densely setose; segment I short; segments II–III approx. equal in length; subsequent segments gradually decreasing in length; male terminalia (Fig. 8A5, A6) visible only ventrally, wider than apex of abdomen; gonocoxites forming two widely separated arms, not fully fused ventrally; gonostyli flattened, shorter than gonocoxites, ending with two teeth of equal length, pointed ventrally; aedeagus and associated membranous structures visible.
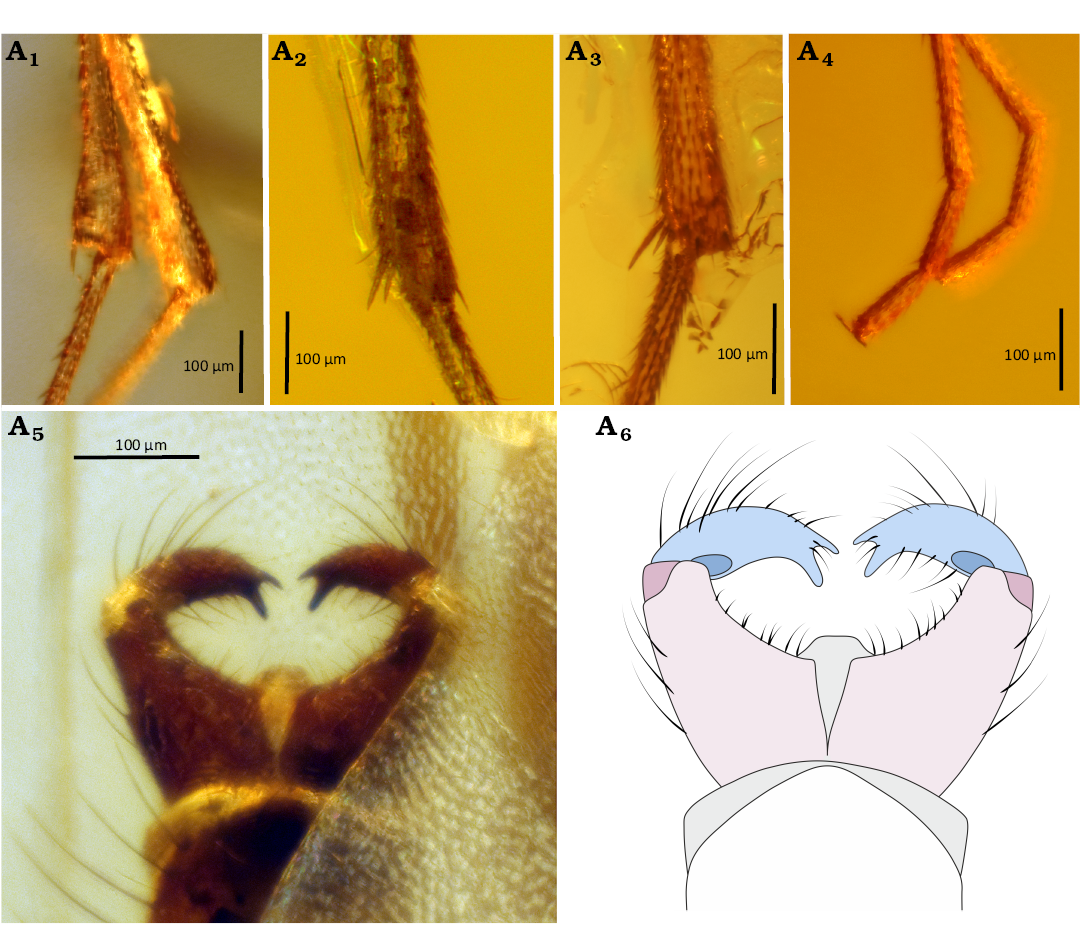
Fig. 8. Predatory fungus gnat Macrocera hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov. (male, holotype CCHH 582-1), Eocene Baltic amber, Baltic region. Spur of the fore tibia (A1), mid tibia (A2), and hind tibia (A3), tip of the tarsus of fore leg (A4), genitalia in ventral view (A5, A6). Photographs (A1–A5), explanatory drawing (A6, not to scale). Color coding: pink, gonocoxites; blue, gonostylus; grey, membranous structures.
Remarks.—Exact number of preserved flagellomeres not specified, as divisions between segments are difficult to observe.
Stratigraphic and geographic range.—Type locality and horizon only.
Macrocera poseidonis Pełczyńska, Krzemiński, & Soszyńska sp. nov.
Figs. 9, 10.
Zoobank LCID: urn:lsid:zoobank.org:act:21B46F16-1D11-48DB-9E 3E-07CBE5C95799
Etymology: Named after Poseidon, the ancient Greek god of the sea, traditionally depicted with a trident. The name refers both to the common association of Baltic amber with the sea and to the distinctive three-toothed gonostylus of this species.
Holotype: MP/5332, male in 10×9×4 mm piece of Baltic amber (Fig. 9A1, A2; IR spectrum Fig. 20D).
Type locality: Baltic region.
Type horizon: Baltic amber, mid-Eocene (ca. 47.8–33.9 Ma according to Seyfullah et al. 2018).
Material.—Holotype only.
Diagnosis.—Antennae more than 1.7× wing length; vein R1 terminating in C just before the R2+3+4+5 fork; R2+3+4+5 fork situated far distal to the level of the M1+2 fork; R2+3 approx. 0.4× the length of the R2+3+4+5 fork stem; M1+2 fork stem part approx. 1.9× frm length; fore tibial spur approx. as long as apical width of tibia; male genitalia distinctly wider than apex of abdomen; gonocoxites long, forming two widely separated arms, largely divided dorsally; gonostyli laterally flattened, slightly shorter than gonocoxites, ending with three sharp teeth of unequal length, median tooth slightly shorter than outer ones, all pointed ventrally.
Description.—Body (Fig. 9A1): Approx. 3.8 mm long; wing 3.9 mm; antennae incomplete, more than 6.6 mm long.
Head (Fig. 9A4, A6): Subspherical, wider than long, inserted just below anterior margin of scutum; eyes large, well separated, occupying most of lateral part of head capsule, interommatidial setae present; distinct cerebral sclerite present; three ocelli present, closely grouped, located on anterior margin of cerebral sclerite, forming a triangle, median ocellus not reduced.
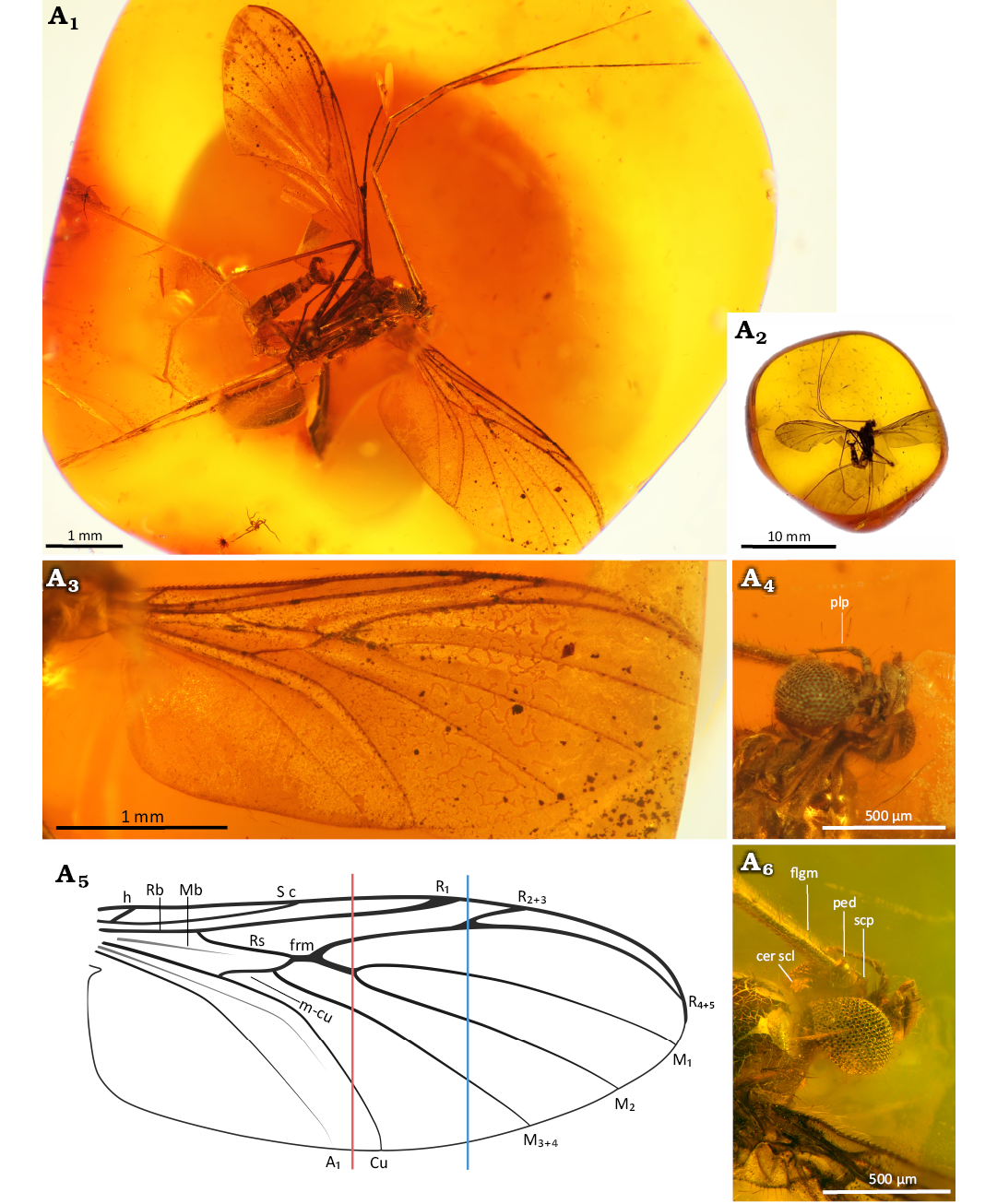
Fig. 9. Predatory fungus gnat Macrocera poseidonis Pełczyńska, Krzemiński, & Soszyńska sp. nov. (male, holotype MP/5332), Eocene Baltic amber, Baltic region. General view of the specimen (A1), amber piece containing specimen (A2), wing in ventral view (A3, A5), head in ventral (A4) and dorso-lateral (A6) views. Photographs (A1–A4, A6), explanatory drawing (A5, not to scale). Abbreviations: cer scl, cerebral sclerite; flgm, flagellomere; ped, pedicel; plp, palpomere; scp, scape. Lines: red, level of the M1+2 fork; blue, level of the R2+3+4+5 fork.
Antennae (Fig. 9A1, A6): Incomplete; preserved segments indicate a length exceeding 2× wing length; scapus as long as broad, globular, approx. 2.3× wider than first flagellomere; pedicel short, annular, approx. 1.8× wider than long, distinctly smaller than scape, 1.6× wider than first flagellomere; flagellum with 8 segments preserved; flagellomeres distinctly elongated, first flagellomere approx. 8.4× as long as broad, subsequent flagellomeres elongate, cylindrical, densely covered with short setae and bearing a row of longer setae along anterior surface.
Palpi (Fig. 9A4): Four maxillary palpomeres visible, all longer than broad; apical palpomere distinctly elongated, approx. 10.4× as long as broad, 2.3× longer than proximal palpomere.
Wing (Fig. 9A3, A5): 2.3× longer than wide; membrane hyaline, without microtrichia or visible markings; C bearing microtrichia throughout its length; microtrichia present on dorsal surface of Sc, Rb, R2+3+4+5, R2+3 and R4+5; C terminating at wing tip, beyond end of R4+5, at approx. half the distance between terminations of R4+5 and M1; Sc ending in C just distal to level of tip of Rb cell; R1 extending beyond half wing length, terminating in C just proximal to level of R2+3+4+5 fork, at approx. half the distance between terminations of Cu and M3+4; R2+3+4+5 fork situated far distal to level of M1+2 fork; R2+3 approx. 0.4× the length of the R2+3+4+5 fork stem; frm ending before level at which A1 reaches wing margin; Mb represented only by a faint trace; M1+2 fork stem approx. 1.9× longer than frm, ending just before approx. half the distance between terminations of A and Cu; M1 approx. 7.6× longer than M1+2 fork stem; opening of cell M2 approx. 1.3× wider than opening of cell M1; base of M3+4 not atrophied; opening of cell M3+4 approx. 1.6× wider than opening of cell M2; basal part of M1+2 and m-cu distinct; Cu reaching wing margin; A1 ending at wing margin; A2 absent.
Thorax (Fig. 10A1): Higher than long; scutum arched, nearly semicircular in lateral view, covered with setae along lateral edges and dorsal surface, arranged in three distinct rows (one dorsocentral and two lateral); scutellum bearing a row of trichia along dorsal margin; anepimeron bare, not reduced, extending to ventral margin of pleura; anepisternum, katepisternum and mediotergite bare; laterotergite with a few setae along dorsal margin.
Legs (Fig. 10A2, A3): Fore coxa bearing sparse, long setae on anterior surface; mid and hind coxae without visible setation; femora irregularly covered with sparse, thin setae; tibiae covered with short setae; fore tibia bearing a ventral row of more robust setae, a single spur (approx. as long as apical width of tibia), a sensory pit, and a row of apical bristles; mid tibiae with a pair of long spurs of unequal length, ventral spur slightly shorter (longer spur approx. 1.7× apical width of tibia); tarsal claws small; pulvilli small.
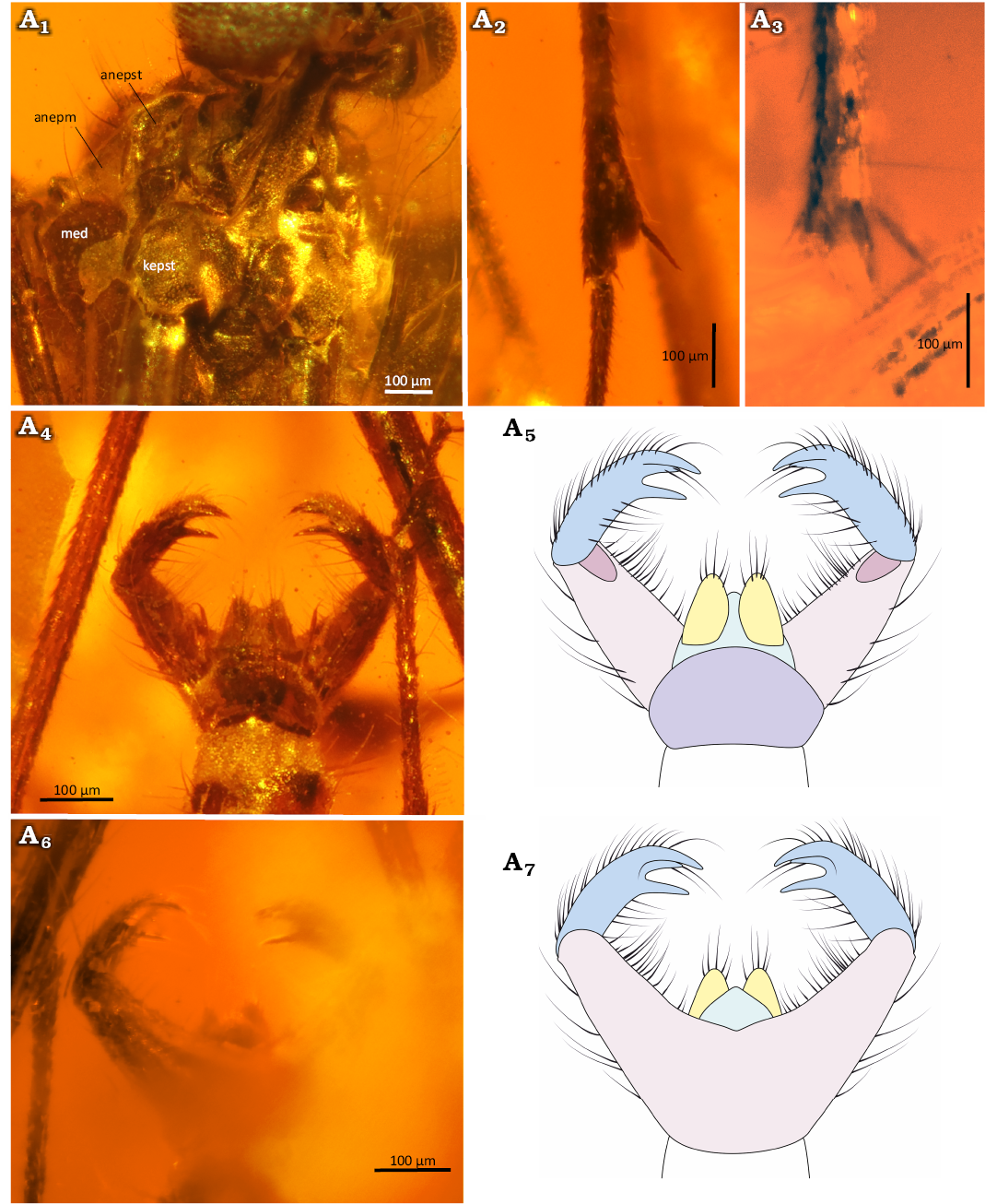
Fig. 10. Predatory fungus gnat Macrocera poseidonis Pełczyńska, Krzemiński, & Soszyńska sp. nov. (male, holotype MP/5332), Eocene Baltic amber, Baltic region. Thorax in anterolateral view (A1), spur of the fore tibia (A2) and mid tibia (A3), genitalia in dorsal (A4, A5) and ventral (A6, A7) views. Photographs (A1–A4, A6), explanatory drawings (A5, A7, not to scale). Abbreviations: anepm, anepimeron; anepst, anepisternum; kepst, katepisternum. Colour coding: yellow, cercus; violet, epandrium; pink, gonocoxites; blue, gonostylus; green, hypoproct.
Abdomen (Fig. 10A4–A7): Densely covered with thin, long setae; eight segments visible; segment I short; segments II–III approx. equal in length; subsequent segments gradually decreasing in length; segment VIII partially retracted into VII; male terminalia (Fig. 10A4–A7) distinctly wider than apex of abdomen; gonocoxites long, forming two widely separated arms, largely divided dorsally beneath epandrium; pair of cerci present; gonostyli laterally flattened, slightly shorter than gonocoxites, ending with three sharp teeth of unequal length, median tooth slightly shorter than outer ones, all pointed ventrally; aedeagus not visible.
Remarks.—Spurs on hind tibiae not observed but most likely present.
Stratigraphic and geographic range.—Type locality and horizon only.
Macrocera stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov.
Figs. 11, 12.
Zoobank LCID: urn:lsid:zoobank.org:act:0A8B06DD-8BBF-4AB5-A158-68A63680F636
Etymology: A playful combination of English words “stuck in amber”, referring to the fact that the holotype is preserved, entrapped within fossil resin.
Holotype: MP/5332, male in 10×9×4 mm piece of Baltic amber (Fig. 11A1, A2).
Type locality: Baltic region.
Type horizon: Baltic amber, mid-Eocene (ca. 47.8–33.9 Ma according to Seyfullah et al. 2018).
Material.—Holotype only.
Diagnosis.—Antennae more than 2.4× wing length; vein R1 terminating in C after the R2+3+4+5 fork; R2+3+4+5 fork situated far distal to the level of the M1+2 fork; R2+3 very short, approx. 0.2× the length of the R2+3+4+5 fork stem; M1+2 fork stem approx. 4.3× frm length; fore tibial spur much longer than apical width of tibia; male genitalia wider than apex of abdomen; gonocoxites short and robust, forming two widely separated arms; gonostyli short and broad, laterally flattened, approx. as wide as gonocoxites, ending with two large, subequal teeth pointed ventrally; aedeagus not visible.
Description.—Body (Fig. 11A1): Approx. 4.4 mm long; wing 4.3 mm; antennae approx. 10.5 mm long.
Head (Fig. 12A1): Subspherical, wider than long, inserted just below anterior margin of scutum; eyes large, well separated, occupying most of lateral part of head capsule; very few interommatidial setae present.
Antennae (Figs. 11A1, 12A1): Length exceeding 2× wing length; scapus approx. as long as broad, annular, approx. 2.1× wider than first flagellomere; pedicel short and broad, annular, approx. 1.6× wider than long, distinctly smaller than scape, approx. 1.4× wider than first flagellomere; flagellomeres distinctly elongated, first flagellomere approx. 9.2× as long as broad, subsequent flagellomeres elongate, cylindrical, bearing sparse, longer, more robust setae on anterior surface.
Palpi (Fig. 12A1): Four maxillary palpomeres visible; two distal palpomeres longer than broad; apical palpomere distinctly elongated, approx. 6.7× as long as broad, 1.7× longer than proximal palpomere.
Wing (Fig. 11A3, A4): Membrane hyaline, without microtrichia or visible markings; C bearing microtrichia throughout its length; microtrichia present on all veins except Cu and A1; C terminating at wing tip, beyond end of R4+5, at approx. half the distance between terminations of R4+5 and M1; Sc ending in C distal to the level of the Rb cell tip; R1 extending beyond half wing length, terminating in C distal to the level of the R2+3+4+5 fork; R2+3+4+5 fork situated far distal to the level of the M1+2 fork; R2+3 very short, approx. 0.2× the length of the R2+3+4+5 fork stem; frm very short; Mb absent; M1+2 fork stem approx. 4.3× longer than frm; M1 approx. 6.7× longer than M1+2 fork stem; base of M3+4, basal part of M1+2 and m-cu not atrophied; A2 absent.
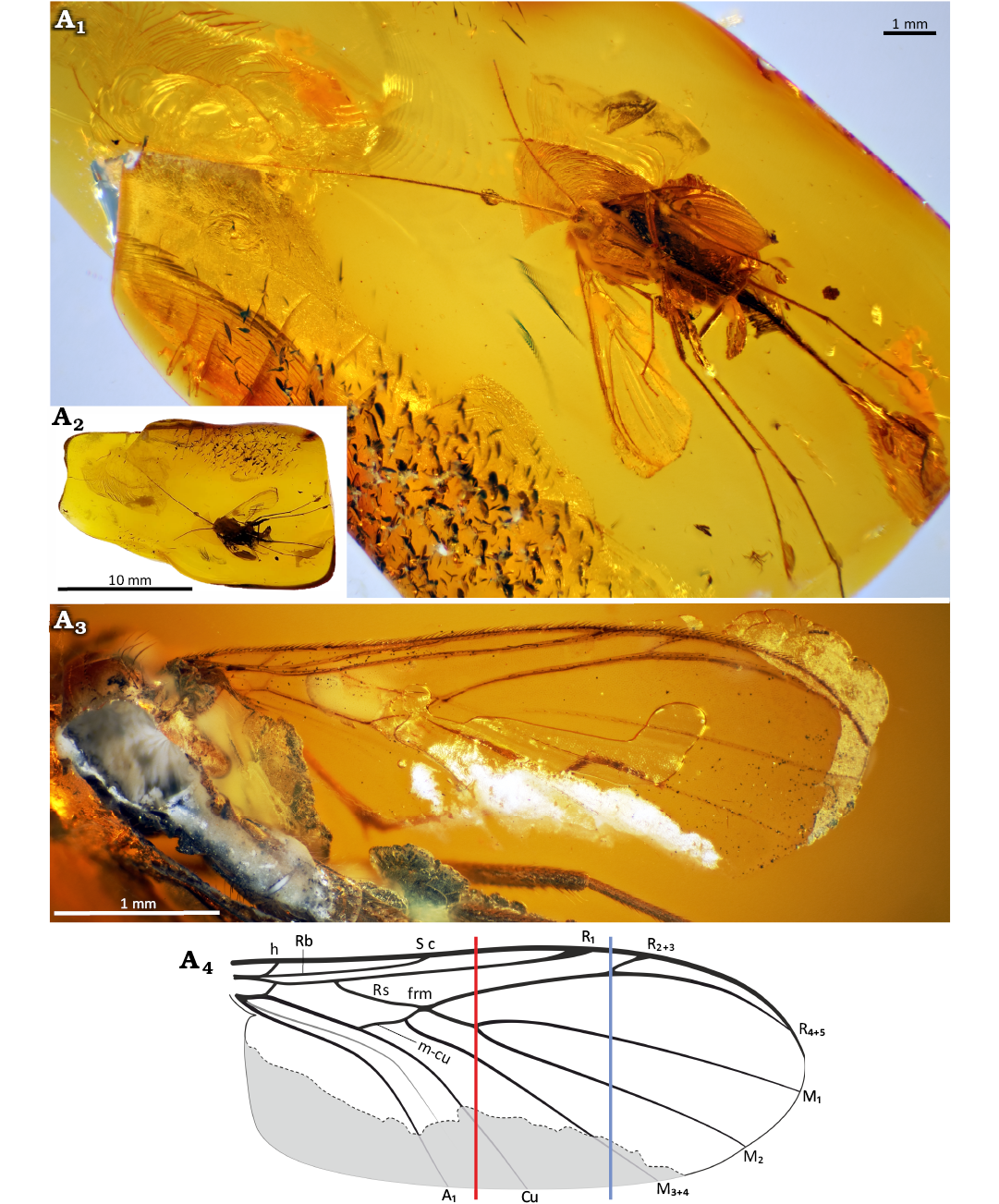
Fig. 11. Predatory fungus gnat Macrocera stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov. (male, holotype MP/5331), Eocene Baltic amber, Baltic region. General view of the specimen (A1), amber piece containing specimen (A2), wing in ventral view (A3, A4). Photographs (A1–A3), explanatory drawing with the hypothesized reconstruction of the missing part of the wing indicated in grey (A4, not to scale). Lines: red, level of the M1+2 fork; blue, level of the R2+3+4+5 fork.
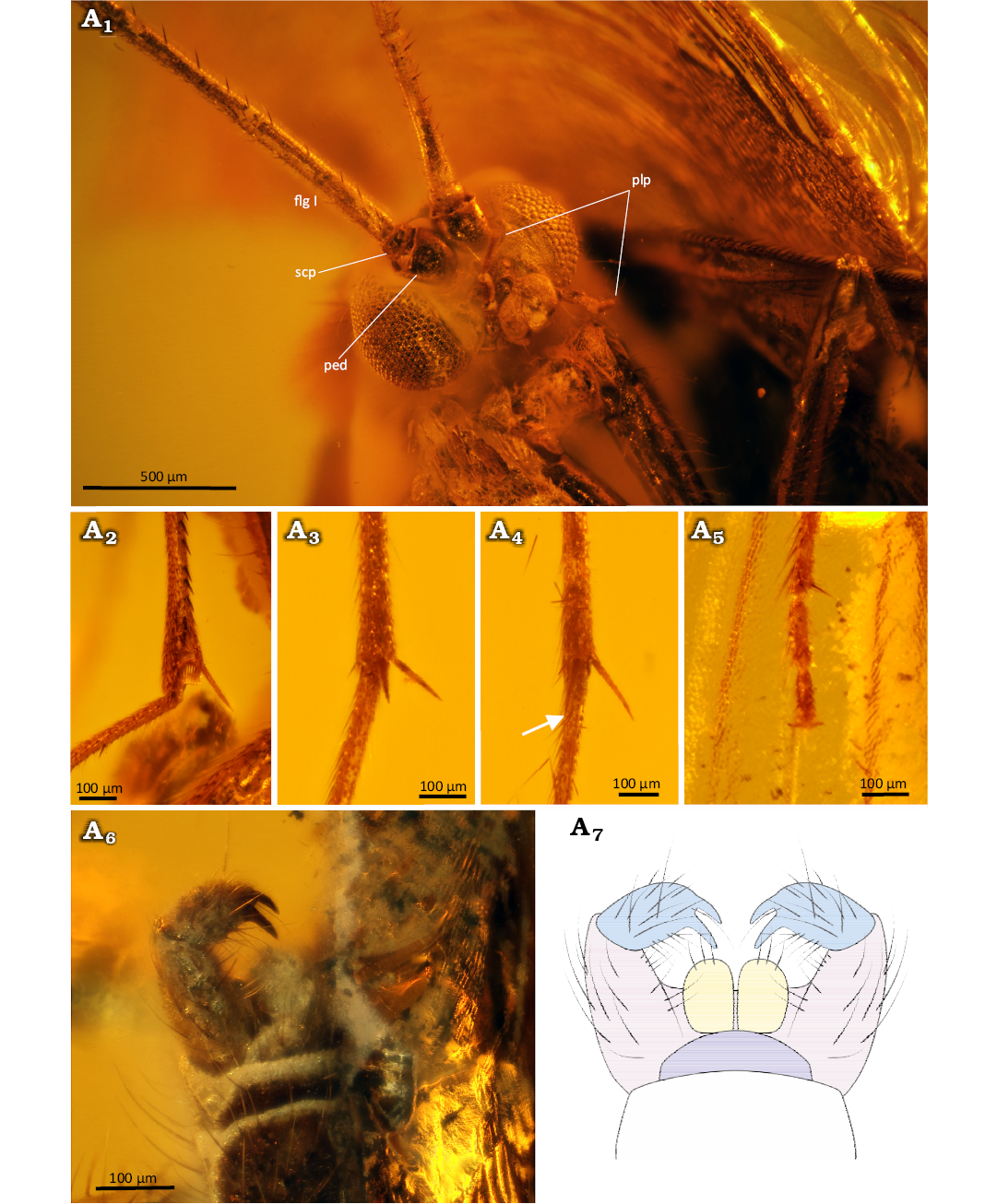
Fig. 12. Predatory fungus gnat Macrocera stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov. (male, holotype MP/5331), Eocene Baltic amber, Baltic region. Head in frontal view (A1), spur of the fore tibia (A2), mid tibia (A3), and hind tibia, tip of the fore tarsus (A4, shorter spur indicated by arrow), genitalia in dorsal view (A5, A6). Photographs (A1–A6), explanatory drawing (A7, not to scale). Abbreviations: flgm, flagellomere; ped, pedicel; plp, palpomere; scp, scape. Colour coding: yellow, cercus; violet, epandrium; pink, gonocoxites; blue, gonostylus; green, hypoproct.
Thorax: Pleural sclerites not visible; scutum partially damaged during specimen preparation, with setae along lateral edges and dorsal surface arranged in three distinct rows (one dorsocentral and two lateral) preserved only on remaining parts; scutellum abraded during amber preparation.
Legs (Fig. 12A2–A5): Fore coxa covered with sparse, long setae; femora and tibiae irregularly covered with thin, dense setae; fore tibia bearing a ventral row of robust setae, a single long spur (approx. 1.9× apical width of tibia), a sensory pit, and a row of apical bristles; mid and hind tibiae with long spurs of unequal length, ventral spur slightly shorter (longer spurs approx. 1.7× and 1.9× apical width of tibiae, respectively); tarsal claws small; pulvilli small.
Abdomen (Fig. 12A6, A7): Densely covered with thin, long setae; segment VIII partially retracted into VII; male terminalia (Fig. 12A6, A7) wider than apex of abdomen; gonocoxites short and robust, forming two widely separated arms; pair of small cerci present; gonostyli short and broad, laterally flattened, approx. as wide as gonocoxites, ending with two large, subequal teeth pointed ventrally; aedeagus not visible.
Remarks.—Posterior part of head and lateral side of thorax not visible due to body orientation in amber; exact number of preserved flagellomeres could not be determined because divisions between segments are indistinct; descriptions of wing, thorax and abdomen incomplete due to specimen damage.
Stratigraphic and geographic range.—Type locality and horizon only.
Macrocera tymoni Pełczyńska sp. nov.
Figs. 13, 14.
Zoobank LCID: urn:lsid:zoobank.org:act:35ECFE0C-80EE-4B1C-BEAD-AA04FB7BB997
Etymology: A patronym tymoni dedicated to Tymon Pabis (Łódź, Poland), in gratitude for creating a pleasant and supportive atmosphere during the preparation of this work, and for the many board games we played together.
Holotype: MP/4422, male in 27×10×6 mm piece of Baltic amber (Fig. 13A1, A2; IR spectrum Fig. 20E).
Type locality: Baltic region.
Type horizon: Baltic amber, mid-Eocene (ca. 47.8–33.9 Ma according to Seyfullah et al. 2018).
Material.—Holotype only.
Diagnosis.—Antennae more than 2.5× wing length; vein R1 terminating in C after the R2+3+4+5 fork; R2+3+4+5 fork situated far distal to the level of the M1+2 fork; R2+3 approx. 0.4× the length of the R2+3+4+5 fork stem; basal part of Mb present; M1+2 fork stem approx. 5× frm length; fore tibial spur much longer than apical width of tibia; male genitalia densely covered with long setae, wider than apex of abdomen; gonocoxites short and robust, forming two widely separated arms; gonostyli broad, laterally flattened, approx. as long as gonocoxites, ending with two very robust, subequal teeth pointed ventrally.
Description.—Body (Fig. 13A2): Approx. 4.6 mm long; wing 4.5 mm; antennae incomplete, more than 11.2 mm long.
Head (Fig. 14A1): Subspherical, wider than long, inserted just below anterior margin of scutum; eyes large, well separated, occupying most of lateral part of head capsule, interommatidial setae present; distinct cerebral sclerite present, angular, sparsely covered with small setae; three ocelli present, closely grouped, located on anterior margin of cerebral sclerite, forming a triangle, median ocellus not reduced.
Antennae (Figs. 13A2, 14A1): Incomplete; preserved segments indicate a length exceeding 2× wing length; scapus broader than long, annular, approx. 2.5× wider than first flagellomere; pedicel short and broad, annular, approx. 1.6× wider than long, distinctly smaller than scape, 1.6× wider than first flagellomere; flagellum with 12 segments preserved; flagellomeres distinctly elongated, first flagellomere approx. 10.6× as long as broad, subsequent flagellomeres elongate, cylindrical, bearing a row of longer, more robust setae on anterior surface.

Fig. 13. Predatory fungus gnat Macrocera tymoni Pełczyńska sp. nov. (male, holotype MP/4422), Eocene Baltic amber, Baltic region. Amber piece containing specimen (A1), general view of the specimen (A2), wing in dorsal view (A3, A4). Photographs (A1–A3), explanatory drawing (A4, not to scale). Lines: red, level of the M1+2 fork; blue, level of the R2+3+4+5 fork.

Fig. 14. Predatory fungus gnat Macrocera tymoni Pełczyńska sp. nov. (male, holotype MP/4422), Eocene Baltic amber, Baltic region. Head and thorax in lateral view (A1), spur of the fore tibia (A2), mid tibia (A3), and hind tibia (A4), tip of the tarsus (A5, claws indicated by arrow), genitalia in dorso-apical (A6), dorsal (A7) and ventral (A8, A9). view. Photographs (A1–A6, A9), explanatory drawings (A7, A8, not to scale). Abbreviations: anepm, anepimeron; anepst, anepisternum; cer scl, cerebral sclerite; flgm, flagellomere; kepst, katepisternum; ltg, laterotergite; med, mediotergite; oc, ocelli; ped, pedicel; plp, palpomere; plv, pulvillus; scp, scape; sct, scutum; sctl, scutellum. Colour coding: yellow, cercus; violet, epandrium; pink, gonocoxites; blue, gonostylus.
Palpi (Fig. 14A1): Four maxillary palpomeres visible; two distal palpomeres longer than broad; apical palpomere distinctly elongated, approx. 5.5× as long as broad, 1.8× longer than proximal palpomere.
Wing (Fig. 13A3, A4): 2.3× longer than wide; membrane hyaline, without microtrichia or visible markings; C bearing microtrichia throughout its length; microtrichia present on dorsal surface of all veins; C terminating at wing tip, beyond end of R4+5, at approx. one-third of the distance between terminations of R4+5 and M1; Sc ending in C far distal to level of tip of Rb cell; R1 extending beyond half wing length, terminating in C distal to the level of the R2+3+4+5 fork, at approx. half the distance between terminations of Cu and M3+4; R2+3 approx. 0.4× the length of the R2+3+4+5 fork stem; R2+3+4+5 fork situated far distal to the level of the M1+2 fork; frm short, ending before level at which A1 reaches wing margin; basal part of Mb present; M1+2 fork stem approx. 5× longer than frm, ending at the level of Cu termination; M1 approx. 2.1× longer than M1+2 fork stem; opening of cell M2 approx. as wide as that of cell M1; base of M3+4 partially atrophied; opening of cell M3+4 approx. 1.6× wider than opening of cell M2; basal part of M1+2 and m-cu partially atrophied; Cu reaching wing margin; A1 ending at wing margin; A2 absent.
Thorax (Fig. 14A1): Higher than long; scutum arched, almost semicircular in lateral view, covered with setation along lateral edges and dorsal surface, arranged in three distinct rows (one dorsocentral and two lateral), setae gradually increasing in length posteriorly; scutellum bearing a row of trichia along its margin; anepimeron bare, not reduced, reaching ventral margin of pleura; anepisternum, katepisternum and laterotergite with very few setae; mediotergite bare.
Legs (Fig. 14A2–A5): Coxae bearing sparse, long setae; femora irregularly covered with thin, dense setae; tibiae irregularly covered with thin, dense setae; fore tibia bearing a ventral row of short, robust setae and a single spur (approx. 1.2× apical width of tibia), a sensory pit, and a row of apical bristles; mid and hind tibiae with long spurs of unequal length, ventral spur slightly shorter (longer spurs approx. 1.8× and 1.6× apical width of tibiae, respectively); tarsal claws small; pulvilli small.
Abdomen (Fig. 14A6–A9): Densely covered with thin, long setae; eight segments visible; segment I short; segment II longest; subsequent segments gradually decreasing in length; segment VIII partially retracted into VII; male terminalia (Fig. 14A6–A9) densely covered with long setae, wider than apex of abdomen; gonocoxites short and robust, forming two widely separated arms; pair of small cerci present; gonostyli broad, laterally flattened, approx. as long as gonocoxites, ending with two very robust, subequal teeth pointed ventrally; aedeagus not visible.
Stratigraphic and geographic range.—Type locality and horizon only.
Identification key to species of Macrocera Meigen, 1803, known from Baltic amber
1. – R2+3+4+5 fork before or at the level of M1+2 fork (Fig. 15A) ..................................................................................................2
– R2+3+4+5 fork beyond the level of M1+2 fork (Fig. 15B) .. 3
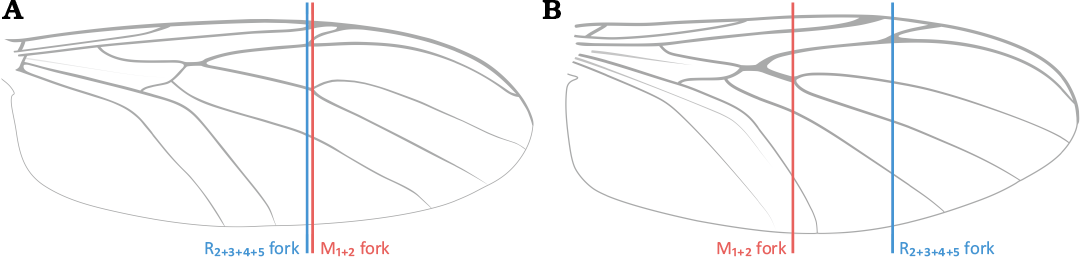
Fig. 15. Relative position of R2+3+4+5 fork to M1+2 fork: R2+3+4+5 fork before or at the level of M1+2 fork (A); R2+3+4+5 fork beyond the level of M1+2 fork (B).
2. – Genitalia as wide as apex of the abdomen; gonocoxites small, positioned parallel to each other; gonostyli approx. as long as gonocoxites (Fig. 16A) .......... M. barteki Pełczyńska sp. nov.
– Genitalia wider than apex of the abdomen; gonocoxites forming two widely separated arms, not fully fused ventrally; gonostyli flattened, shorter than gonocoxites (Fig. 16B) .......................................... M. hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov.
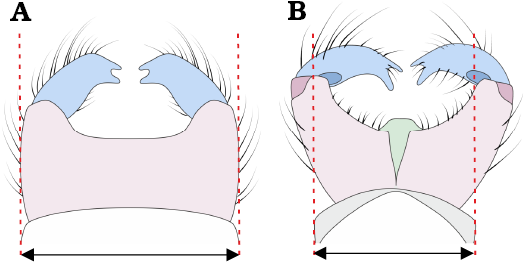
Fig. 16. Width of male genitalia relative to width of apical tip of the abdomen: genitalia as wide as apex of the abdomen (A); genitalia wider than apex of the abdomen (B).
3. – Gonostyli ending with three teeth (Fig. 17A) ....... M. poseidonis Pełczyńska, Krzemiński, & Soszyńska sp. nov.
– Gonostyli ending with two teeth (Fig. 17B) .................... 4
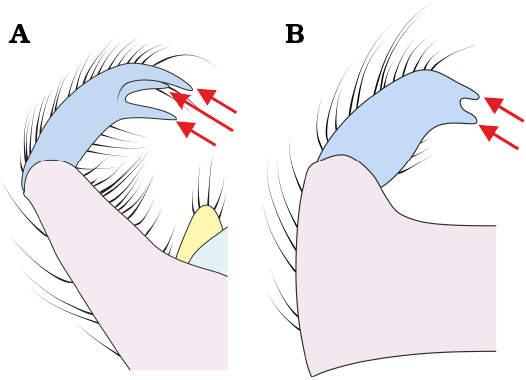
Fig. 17. Apical projections of gonostyli: gonostyli ending with three teeth (A); gonostyli ending with two teeth (B). Arrows indicate the apical tips of the gonostylar teeth.
4. – Frm relatively long, with M1+2 fork stem about 2× its length (Fig. 18A) .......... M. electricornis Evenhuis, 2006
– Frm short, with M1+2 fork stem more than 4× its length (Fig. 18B) .............................................................................. 5
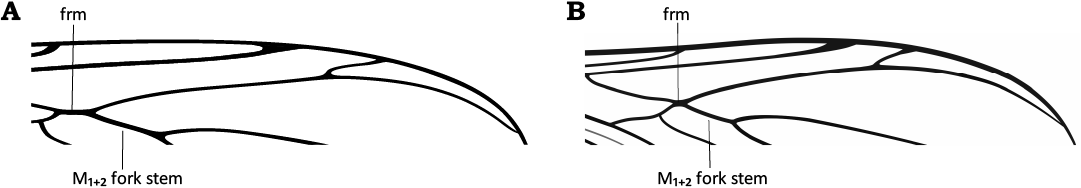
Fig. 18. Length of M1+2 fork stem relative to frm: M1+2 fork stem more than 2× longer than frm (A); M1+2 fork stem more than 4× longer than frm (B).
5. – R2+3 short, approx. 0.2× the length of the R2+3+4+5 fork stem (Fig. 19A) ......................... M. stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov.
– R2+3 long, approx. 0.4 × the length of the R2+3+4+5 fork stem (Fig. 19B) ................. M. tymoni Pełczyńska sp. nov.

Fig. 19. Length of R2+3 relative to R2+3+4+5 fork stem: R2+3 short, approx. 0.2× the length of the R2+3+4+5 fork stem (A); R2+3 long, approx. 0.4 × the length of the R2+3+4+5 fork stem (B).
Results
The Baltic origin of the investigated specimens was confirmed using Fourier transform infrared (FTIR) spectroscopy with attenuated total reflectance (ATR). The spectra of GZG.BST.3064, MP/3247, 4422, 5331, show a characteristic doublet of peaks in the range of 1260–1160 cm⁻¹, with a maximum at approx. 1150 cm⁻¹ (Fig. 20). This diagnostic feature, known as the Baltic shoulder, results from the presence of succinic acid components that characterize Baltic amber (Wolfe et al. 2016; Drąg et al. 2020). However, the spectrum of CCHH 582-1 (Fig. 20C) is distorted due to its embedding in epoxy resin, and FTIR analysis was not performed for MP/5332 because of technical limitations related to its very small size (10×9×4 mm) and irregular shape, which prevented the acquisition of reliable spectra.
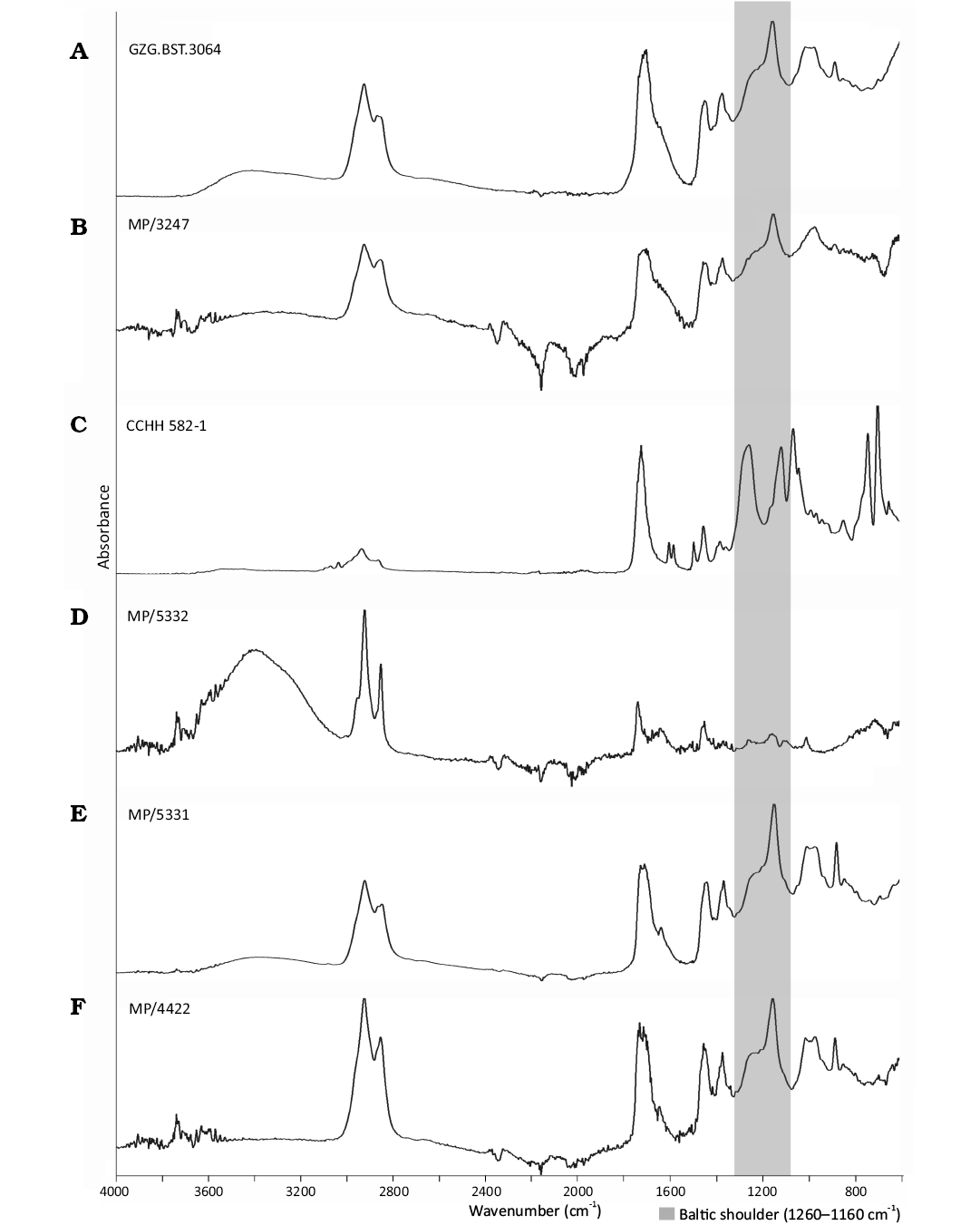
Fig. 20. Fourier transform infrared spectroscopy (attenuated total reflectance) spectra obtained from investigated Baltic amber specimens; the Baltic shoulder wavelength of occurrence marked. GZG.BST.3064 (A); MP/3247 (B); CCHH 582-1 (C); MP/5332 (D); MP/4422 (E).
Discussion
Age of the material.—It is estimated that hundreds of tons of Baltic amber are extracted annually from the upper part of the Blue Earth layer of the Prussian Formation (Sambian Peninsula). The age of such specimens can potentially be correlated with the age of this layer, which has been dated to the early Priabonian (approx. 36–35 Ma) (Penney and Preziosi 2013; Drohojowska et al. 2024; Ross et al. 2026). In contrast, specimens collected, for example, along the Baltic or North Sea coasts have an unknown provenance and cannot be dated more precisely than within the broadly accepted age range of Baltic amber (Lutetian to Priabonian, 47.8–33.9 Ma) (Seyfullah et al. 2018). Therefore, we recommend preserving the most detailed possible provenance data for Baltic amber specimens, as discrepancies in their geographical origin and the potentially different ages of their source deposits may be highly relevant for future research.
Delimitation of the Macrocera.—Within the genus Macrocera, most species conform to a relatively conservative morphological bauplan. However, some taxa exhibit more pronounced departures from this pattern. This is exemplified, among others, by the Mediterranean species group, which is characterised by more complex male terminalia bearing modifications that exceed simple allometric variation (Matile 1990, Mantič and Ševčík 2017).
Historically, many species have been assigned to Macrocera largely on the basis of markedly elongated and filiform male antennae, a character that, although distinctive, may be prone to homoplasy. Consequently, the inclusion of morphologically divergent species complicates generic delimitation and suggests the possible paraphyly of the genus.
This issue was already recognised by Matile (1990), who emphasised the need for a revision of Macrocera. Such a revision should, however, be supported by molecular data, which would allow for a more robust reconstruction of relationships among species and species groups within the genus. Therefore, for the purposes of the present study, we retain the traditional, broad concept of Macrocera sensu lato.
Morphological comparison.—Wing venation is rather uniform among Macrocerinae and is prone to parallel evolution within Keroplatidae (Matile 1990; Mantič et al. 2020; Ševčík 2024). Nevertheless, differences can be observed among the species described here.
In particular, the length of the M₁₊₂ fork stem varies between species. Two clearly distinct states can be recognised. The M₁₊₂ vein may fork distinctly before the R₂₊₃₊₄₊₅ forking, or approx. at the same level, as observed in M. barteki Pełczyńska sp. nov. (Fig. 4A3, A4) and M. hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov. (Fig. 7A3, A4. The long M₁₊₂ fork stem observed in these two species can be regarded as a plesiomorphic feature and also distinguishes them from M. electricornis, previously described from Baltic amber (Matile 1990). No apparent intermediate states were observed among the species described herein.
Another character that can be interpreted as apomorphic is the length of frm, as it is generally assumed that the radial and medial sectors have become increasingly fused during the evolution of Keroplatidae (Matile 1990). The most plesiomorphic condition of this character is observed in M. stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov. (Fig. 11A3, A4), in which frm is very short and several times shorter than in M. electricornis.
Variation is also evident in the degree of preservation of the Mb. Its presence should be regarded as plesiomorphic, as this vein undergoes partial or complete atrophy in most Sciaroidea, including Keroplatidae (Hennig 1954; Matile 1990). Within Macrocerinae, a fully preserved Mb vein reaching the apex of the basal cell occurs only in the genera Vockerothia Matile, 1990, where it is complete, and Hesperodes Coquillett, 1900, where it is present but weakened apically. Partial preservation of Mb, excluding its apical part, may also occur in some species of Macrocera. Among the new species described here, clear variation in the degree of sclerotization of this vein is observed, ranging from the presence of a distinct basal part in M. tymoni Pełczyńska sp. nov. (Fig. 13A3, A4) to complete atrophy in M. stuckinamberi Pełczyńska, Krzemiński, & Soszyńska sp. nov. (Fig. 11A3, A4), a condition shared with M. electricornis. It should be noted, however, that the apparent condition of this vein may be affected by taphonomic processes, which can obscure weakly sclerotised veins in amber inclusions
With respect to male genitalia, the most typical condition in Macrocerinae involves gonostyli bearing two apical teeth, as observed in M. electricornis and in most of the species described here. Only a single species, M. poseidonis Pełczyńska, Krzemiński, & Soszyńska sp. nov., exhibits a third tooth (Fig. 10A4–A7). Although this condition is less common, it is not unique and has also been known in species such as Macrocera caudata Matile, 1977, and Macrocera annuliventris Matile, 1972.
At present, it is not possible to place these species along a plesiomorphic-apomorphic axis or to infer their relationship to a potential stem lineage without proper phylogenetic analyses. Consequently, any such interpretation would be premature.
Fossil record and morphological conservatism of the Macrocera.—The analysis of the newly described Baltic amber species requires placing them within a broader evolutionary context and comparing them with taxa known from other fossil materials. Macrocera is additionally represented by seven fossil species recorded from other resins as well as from compression fossils (Evenhuis 2006; Pełczyńska et al. 2024, 2025)
The presence of this taxon in the fossil record has been documented in the Lower Cretaceous, in Álava amber (Burgos, Spain), dated to the late Albian (approx. 105 Ma) (Barrón et al. 2015; Pérez-de la Fuente et al. 2020; Pełczyńska et al. 2025). From this fossil resin, two of the oldest representatives of Macrocerinae were described, both originally placed in Hegalari Blagoderov & Arillo, 2002 (H. antzinako Blagoderov & Arillo, 2002, and H. minor Blagoderov & Arillo, 2002) (Barrón et al. 2015; Pérez-de la Fuente et al. 2020; Pełczyńska et al. 2025). Subsequently, H. minor was reassigned to Macrocera (Pełczyńska et al. 2025).
This indicates that Macrocera, together with Hegalari, represents one of the oldest lineages within the family Keroplatidae, as only a single older record is known, Lebanognoriste prima Blagoderov & Grimaldi, 2004 (Keroplatidae: Lygistorrhininae), described from Lebanese amber dated to the late Barremian (approx. 125 Ma) (Pełczyńska et al. 2024).
Furthermore, the Cretaceous M. minor closely resembles extant representatives of Macrocera, exhibiting elongate flagellomeres (although less pronounced than in the species described herein), a typical genital structure with a gonostylus bearing two apical teeth, short tibial spurs, and elongate apical palpomeres. This combination of characters suggests a high degree of morphological conservatism and implies that the evolutionary origins of Macrocera may extend even further back into the Mesozoic (Pełczyńska et al. 2025).
Three slightly younger species (Macrocera vonneguti Pełczyńska & Blagoderov, 2025, Macrocera pawli Pełczyńska, 2025, and Macrocera sevciki Pełczyńska & Krzemiński, 2025) originate from Early Cretaceous Burmese amber (Myanmar), dated to the earliest Cenomanian (approx. 99 Ma) (Shi et al. 2012). Together with M. minor, all Mesozoic representatives of the genus differ from the younger Eocene fauna by, among other features, a much more slender shape of the gonocoxites and more prominent tibial setation. In contrast, the shortening of the gonocoxites and the reduction of tibial setation observed in Baltic amber species can be regarded as apomorphic (Matile 1990).
Other Eocene deposits contain Macrocera apithanos Kerr & Greenwalt, 2022, described from a compression fossil from the Kishenehn Formation (Montana, USA), dated to the Lutetian (45.8–46.6 Ma). Two younger compression fossils are also known: Macrocera archaica Armbruster, 1938, from the Randeck Maar Formation of Germany, dated to the Miocene (15.98–13.82 Ma), and Macrocera umbonata Statz, 1944, from the Rott Formation of Germany, dated to the Oligocene (approx. 23–24 Ma) (Mörs 1995; Rasser et al. 2013; Greenwalt et al. 2022). All these species possess a short M1+2 fork stem, which distinguishes them from M. hoffeinsorum Pełczyńska, Krzemiński, & Soszyńska sp. nov. and M. barteki Pełczyńska sp. nov.
In addition, M. apithanos is characterised by a very short Sc vein that terminates in C distinctly before the apex of the basal cell, approx. at the origin of M₃₊₄, which separates it from the Baltic amber species. Macrocera archaica, in contrast, appears to have a strongly shortened R₁ vein reaching the wing margin at the level of the M₁₊₂ fork, accompanied by a short M₁₊₂ fork stem. In M. umbonata, vein R₄₊₅ reaches the wing margin very close to the tip of M₁, as R₄₊₅ is very long and M₁ terminates high, near the wing apex. None of these character states are observed in the species described here.
The newly described species, together with M. electricornis, previously described from Baltic amber, do not differ markedly from extant representatives of the genus. This is not unexpected, given that Macrocera appears to have been well established already in considerably older fossil deposits.
Conclusions
We posit that Macrocera remains poorly documented in Baltic amber inclusions. This pattern is consistent with observations made in other taxa and further supports the view that the palaeodiversity preserved in this fossil resin is still far from fully explored (Penney and Preziosi 2013).
Consequently, research on the Baltic amber fauna continues to hold substantial potential, as numerous arthropod species appear to remain undescribed. Improved knowledge of this palaeobiota will provide important insights into the evolutionary history and biogeography of insect lineages, as well as into the palaeoenvironment of Eocene Europe.
Acknowledgements
The authors would like to thank Katarzyna Kopeć (ISEA PAS) for her assistance with the spectroscopy analyses, Lars Vilhelmsen (Natural History Museum of Denmark, University of Copenhagen, Denmark) for proofreading the manuscript, Anders Alexander Illum (Natural History Museum of Denmark, University of Copenhagen, Denmark) for technical support during the preparation of photographic documentation. The authors express our sincere gratitude to Christel and Hans Werner Hoffeins (Hamburg, Germany) for kindly providing access to specimens from their private collection and subsequently agreeing to deposit the studied material in the SDEI collection. Finally, the authors would like to thank the reviewers for their insightful comments and valuable suggestions. This research was funded by the National Science Center, Poland (grant no. 2020/37/B/NZ8/03042).
Authors’ contributions
AP took the lead in writing the manuscript and was responsible for material preparation, photography, and graphic illustrations. AP and WK made taxonomic decisions. AS and WK participated in establishing the research plan and concept, provided access to the material, and supervised the project and secured financial support. All authors provided critical feedback, contributed to refining the manuscript, and approved its final version.
Conflict of interest: The authors declare that they have no conflicts of interest in relation to this work.
Data availability statement: The data underlying this article will be shared on reasonable request to the corresponding author.
Editor: Andrzej Kaim
References
Armbruster, L. 1938. Versteinerte Honigbienen aus dem obermiocänen Randecker Maar. Archiv für Bienenkunde 19: 97–133.
Barrón, E., Peyrot, D., Rodríguez-López, J.P., Meléndez, N., López del Valle, R., Najarro, M., Rosales, I., and Comas-Rengifo, M.J. 2015. Palynology of Aptian and upper Albian (Lower Cretaceous) amber-bearing outcrops of the southern margin of the Basque-Cantabrian basin (northern Spain). Cretaceous Research 52: 292–312. Crossref
Billberg, G. J. 1820. Enumeratio insectorum in Museo. 138 pp. Typis Gadelianis. Stockholm.
Blagoderov, V. and Arillo, A. 2002. New Sciaroidea (Insecta : Diptera) in Lower Cretaceous amber from Spain. Studia Dipterologica 9: 3–12.
Blagoderov, V. and Grimaldi, D. 2004. Fossil Sciaroidea (Diptera) in Cretaceous Ambers, Exclusive of Cecidomyiidae, Sciaridae, and Keroplatidae. American Museum Novitates 2004: 1–76. Crossref
Blagoderov, V., Krzemiński, W., and Skibińska, K. 2019. A new species of Micrepimera Matile (Diptera: Keroplatidae) from Baltic Amber. Palaeoentomology 2: 643–649. Crossref
Chang, S.-C., Li, Y., and Zheng, D. 2023. Dating Amber: Review and Perspective. Minerals 13: 948. Crossref
Coquillett, D.W. 1900. Papers from the Harriman Alaska Expedition. IX. Entomological Results (3): Diptera. Proceedings of the Washington Academy of Sciences 2: 389–464.
Cumming, J.M. and Wood, D.M. 2017. Adult morphology and terminology. In: A.H. Kirk-Spriggs and B.J. Sinclair (eds.), Manual of Afrotropical Diptera, Vol. 1., 89–133. SANBI, Pretoria.
Drąg, K., Matuszewska-Skoczek, M., Dumała-Słowińska, M., and Żyła, G. 2020. Identification of treated Baltic amber by FTIR and FT-Raman spectroscopy. Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy 279: 121404. Crossref
Drohojowska, J., Śladowska, J., and Szwedo, J. 2024. First whiteflies from the Eocene amber of Denmark. Acta Palaeontologica Polonica 69: 649–676. Crossref
Evenhuis, N.L. 2006. Catalog of the Keroplatidae of the World (Insecta: Diptera). 178 pp. Bishop Museum Press, Honolulu.
Fabricius, J.C. and Fabricius, J.C. 1781. Species insectorum, exhibentes eorum differentias specificas, synonyma auctorum, loca natalia, metamorphosin, adjectis observationibus, descriptionibus. 517 pp. C.E. Bohnii, Hamburg. Crossref
Greenwalt, D.E., Amorim, D.D.S., Hauser, M., Kerr, P.H., Fitzgerald, S.J., Winterton, S.L., Cumming, J.M., Evenhuis, N.L., and Sinclair, B.J. 2022. Diptera of the Middle Eocene Kishenehn Formation II. Palaeontologia Electronica 25 (2): 1–52. Crossref
Grimaldi, D.A. and Ross, A.J. 2018. Extraordinary Lagerstätten in amber, with particular reference to the Cretaceous of Burma. In: D. Penney (ed.), Terrestrial Conservation Lagerstätten: Windows into the Evolution of Life on Land, 287–342. Cambridge University Press, Cambridge. Crossref
Hennig, W. 1948. Die Larvenformen der Dipteren. Vol. 1. 184 pp. Akademie Verlag, Berlin.
Hennig, W. 1954. Flügelgeäder und System der Dipteren unter Berücksichtigung der aus dem Mesozoikum beschriebenen Fossilien. Beiträge zur Entomologie Contributions to Entomology 4: 245–388.
Laurence, B.R. 1982. A cave dwelling mycetophilid in central London. Entomologist’s monthly Magazine 117 (1981): 198.
Lim, W., Bang, W.J., and Shin, S. 2025. New records of Macrocera Meigen 1803 (Diptera, Keroplatidae) from the Korean Peninsula with description of a new species. Zootaxa 5665: 239–255. Crossref
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Loew, H. 1850. Ueber den Bernstein u. Die Bernsteinfauna. 48 pp. Mittler u. Sohn. Berlin. Crossref
Mantič, M. and Ševčík, J. 2017. Macrocera rohaceki sp. nov. and other interesting records of Keroplatidae (Diptera) from southern and central Europe, with DNA sequence data. Acta Entomologica Musei Nationalis Pragae 57: 751–764. Crossref
Mantič, M., Sikora, T., Burdíková, N., Blagoderov, V., Kjærandsen, J., Kurina, O., and Ševčík, J. 2020. Hidden in Plain Sight: Comprehensive Molecular Phylogeny of Keroplatidae and Lygistorrhinidae (Diptera) Reveals Parallel Evolution and Leads to a Revised Family Classification. Insects 11: 348. Crossref
Matile, L. 1972. Keroplatinae des îles de l’Océan Indien Occidental (Diptera, Mycetophilidae). Cahiers Orstom 16: 105–123.
Matile, L. 1977. Keroplatinae de Madagascar (Diptera: Mycetophilidae). Annals of the Natal Museum 23 (1): 23–36.
Matile, L. 1979. Un nouveau genre de Keroplatidae de l’ambre oligocene de la Baltique (Diptera: Mycetophilidae). Revue Française d’Entomologie 1: 36–41.
Matile, L. 1990. Recherches sur la systématique et l’évolution des Keroplatidae (Díptera, Mycetophiloidea). 682 pp. Mémoire du Muséum national d’Histoire naturelle, Paris.
Meigen, J. 1803. Versuch einer neuen Gattungseinteilung [sic!] der eurpoäischen zweiflügligen Insekten. Magazin für Insektenkunde 2: 259–281.
Meigen, J.W. 1804. Klassifikazion und beschreibung der europäischen zweiflügligen insekten: (Diptera Linn.). 414 pp. Karl Reichard, Braunschweig.
Meunier, F. 1899. Études sur quelques Diptères de l’ambre tertiaire. Bulletin de la Société entomologique de France 5 (4): 111–112.
Meunier, F.A. 1904. Monographie des Cecidomyidae, des Sciaridae, des Mycetophilidae et des Chironomidae de l’ambre de la Baltique. 275 pp. Société scientifique de Bruxelles, City of Brussels. Crossref
Mörs, T. 1995. Die Sedimentationsgeschichte der Fossillagerstätte Rott und ihre Alterseinstufung anhand neuer Säugetierfunde (Oberoligozän, Rheinland). CFS Courier Forschungsinstitut Senckenberg 187: 1–129.
Pełczyńska, A., Blagoderov, V., Krzemiński, W., and Soszyńska, A. 2025. Long story of long antennae: Mesozoic origins of the subfamily Macrocerinae (Diptera: Keroplatidae). Acta Geologica Sinica (English Edition) 99: 1225–1248. Crossref
Pełczyńska, A., Krzemiński, W., Blagoderov, V., Vilhelmsen, L., and Soszyńska, A. 2024. Eocene amber provides the first fossil record and bridges distributional gap in the rare genus Robsonomyia (Diptera: Keroplatidae). Scientific Reports 14: 9252. Crossref
Penney, D. and Preziosi, R.F. 2013. Estimating fossil ant species richness in Eocene Baltic Amber. Acta Palaeontologica Polonica 59: 927–929. Crossref
Pérez-de la Fuente, R., Engel, M.S., Delclòs, X., and Peñalver, E. 2020. Straight-jawed lacewing larvae (Neuroptera) from Lower Cretaceous Spanish amber, with an account on the known amber diversity of neuropterid immatures. Cretaceous Research 106: 104200. Crossref
Perkovsky, E.E., Rasnitsyn, A.P., Vlaskin, A.P., and Taraschuk, M.V. 2007. A comparative analysis of the Baltic and Rovno amber arthropod faunas : representative samples. African Invertebrates 48: 229–245.
Rasser, M., Bechly, G., Böttcher, R., Ebner, M., Heizmann, E.P.J., Höltke, O., Joachim, C., Kern, A.K., Kovar-Eder, J., Nebelsick, J.H., Roth-Nebelsick, A., Schoch, R.R., Schweigert, G., and Ziegler, R. 2013. The Randeck Maar: Palaeoenvironment and habitat differentiation of a Miocene lacustrine system. Palaeogeography Palaeoclimatology Palaeoecology 392: 426–453. Crossref
Rondani, C. 1856. Dipterologiae Italicae Prodromus. Vol. I: Genera italica ordinis Dipterorum ordinatim disposita et distincta et in familias et stirpes aggregata. 226 pp. A. Stocchi. Parma. Crossref
Ross, A.J., Bojarski, B., and Szwedo, J. 2026. A critical review of the age of Baltic amber from the Samland Peninsula, Russia. Earth and Environmental Science Transactions of The Royal Society of Edinburgh, 1–14. Crossref
Ševčík, J. 2024. Davimacrocera, a new extant genus of Keroplatidae, with notes on the fossil genus Burmacrocera (Diptera). Insects 15: 1018. Crossref
Ševčík, J. and Roháček, J. 2008. Fungus gnats (Diptera: Mycetophilidae and Keroplatidae) reared from grass and sedge tussocks in the Czech Republic. Časopis Slezského Muzea v Opavě (A) 57: 175–178.
Ševčík, J., Krzemiński, W., and Skibińska, K. 2020. Intriguing and beautiful: Adamacrocera adami gen. et sp. nov. from the Upper Cretaceous amber of Myanmar represents a new subfamily of Keroplatidae (Diptera: Bibionomorpha). Insects 11: 552. Crossref
Ševčík, J., Krzemiński, W., and Skibińska, K. 2022. Extant genus in the Mesozoic: Paleoplatyura Meunier (Diptera: Keroplatidae) found in the Cretaceous amber of Myanmar. Insects 13 (1): 24. Crossref
Seyfullah, L.J., Beimforde, C., Dal Corso, J., Perrichot, V., Rikkinen, J., and Schmidt, A.R. 2018. Production and preservation of resins—past and present. Biological Reviews 93: 1684–1714. Crossref
Shi, G., Grimaldi, D., Harlow, G., Wang, J., Wang, J., Yang, M., Lei, W., Li, Q.-L., and Li, X. 2012. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretaceous Research 37: 155–163. Crossref
Statz, G. 1944. Neue Dipteren (Nematocera) aus dem Oberoligocän von Rott. III. Familie Limnobiidae (Stelzmücken). IV. Familie: Tipulidae (Schnaken). V. Familie: Culicidae (Stechmücken). Palaeontographica A 95: 93–120.
Szwedo, J. and Sontag, E. 2013. The flies (Diptera) say that amber from the Gulf of Gdańsk, Bitterfeld and Rovno is the same Baltic amber. Polish Journal of Entomology 82: 379–388. Crossref
Wolfe, A.P., McKellar, R.C., Tappert, R., Sodhi, R.N.S., and Muehlenbachs, K. 2016. Bitterfeld amber is not Baltic amber: Three geochemical tests and further constraints on the botanical affinities of succinite. Review of Palaeobotany and Palynology 225: 21–32. Crossref
Zakrzewska, M., Singh, H., Wagner-Wysiecka, E., and Giłka, W. 2020. Minute and diverse in fossil sticky stuff: Tanytarsini (Diptera: Chironomidae) from early Eocene Indian Cambay amber. Zoological Journal of the Linnean Society 189: 1398–1425. Crossref
Acta Palaeontol. Pol. 71 ( 2): 219–245, 2026
https://doi.org/10.4202/app.01318.2025